 ,2)同济大学航空航天与力学学院,上海 200092
,2)同济大学航空航天与力学学院,上海 200092THE COMPLEX DYNAMICS OF ABNORMAL PHENOMENON OF NEURAL ELECTRONIC OSCILLATIONS INDUCED BY NEGATIVE FEEDBACK 1)
Lan Yuqun, Guan Linan, Gu Huaguang,2)School of Aerospace and Applied Mechanics, Tongji University, Shanghai 200092, China通讯作者: 2) 古华光,教授,主要研究方向:神经动力学,非线性动力学.E-mail:guhuaguang@tongji.edu.cn
收稿日期:2019-01-30接受日期:2019-04-1网络出版日期:2019-07-18
| 基金资助: |
Received:2019-01-30Accepted:2019-04-1Online:2019-07-18
作者简介 About authors

摘要
传统观念认为,负反馈容易使系统达到稳定平衡点而正反馈容易引起振荡.本研究基于神经元理论模型,提出了负反馈可以诱发稳定平衡点、也就是静息、变为振荡、也就是放电的新观点.在Hopf分岔点附近,作用在静息上的一次足够大的负向脉冲电流的抑制性刺激,能够引起一个动作电位及随后的衰减振荡的后电位;而能够在后电位上诱发出动作电位的负脉冲电流强度阈值也是衰减振荡的.在模型中,引入具有时滞($\tau$)的负反馈来模拟抑制性自突触,一个动作电位诱发的负反馈自突触电流会作用到比动作电位延迟$\tau$的后电位上.随时滞增加,能够诱发出放电的负反馈增益强度阈值呈现出具有衰减振荡特点的类似多重相干共振的特性,衰减振荡的周期与电流阈值曲线的周期以及分岔点附近的放电周期相关.另外,负反馈还能诱发出放电与静息共存的复杂行为.本研究的结果不仅揭示了负反馈的新的反常调控作用,还有助于理解在现实神经系统中存在的慢抑制性自突触的潜在功能.
关键词:
Abstract
In traditional viewpoint, it is easy to achieve stable equilibrium for a system with negative feedback and stable oscillations for a system with positive feedback. In the present investigation of the nonlinear neural system, a novel viewpoint that negative feedback can induce stable equilibrium, i.e. the resting state, changed to oscillations, i.e. firing, is proposed. In a theoretical neuron model, inhibitory stimulation mediated by the negative impulsive current with enough strength can induce an action potential from the resting state near a Hopf bifurcation point and the after-potential with damping oscillations following the action potential. The strength threshold of the second negative impulsive current applied within the after-potential to evoke the second action potential exhibits damping oscillations as well with respect to the application phase of the current. After introducing negative feedback with time delay ($\tau $) into the theoretical model to simulate the inhibitory autapse, the negative feedback current induced by an action potential is applied at the phase $\tau $ of the after-potential following the action potential. The negative feedback gain threshold to induce firing from the resting state exhibits characteristics of damping oscillations with increasing time delay corresponding to application phase of the current, which resembles the time delay induced-multiple coherence resonances. The oscillation period is associated with the period of strength threshold curve of the second impulsive current to evoke the second action potential from after-potential and the period of the firing near the Hopf bifurcation. Furthermore, the negative feedback can also induce complex dynamics such as the coexistence of the firing and resting state. The results of the present paper not only present a novel modulation effect of the negative feedback, which is a contrast to the tradition viewpoint, but also are helpful for understanding the potential functions of slow inhibitory autapse that can induce negative impulsive current with time delay in the real neural system.
Keywords:
PDF (3283KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
蓝昱群, 关利南, 古华光. 负反馈诱发神经电振荡的反常现象的复杂动力学 1). 力学学报[J], 2019, 51(4): 1122-1133 DOI:10.6052/0459-1879-19-038
Lan Yuqun, Guan Linan, Gu Huaguang.
引言
正反馈或负反馈是调控动力系统行为的重要手段.一般而言,采用负反馈主要目的是使得系统变为稳定、达到平衡点,而采用正反馈是为了引起振荡.但在非线性动力系统中往往存在违反常规概念的行为.比如神经系统,其非线性的电活动会表现出十分复杂的乃至反常规概念的动力学行为[1-13].比如目前已经获得广泛认可的随机共振和相干共振现象,揭示了噪声有助于而不是扰乱神经系统的信息处理[1].神经电活动包括静息和放电,分别对应稳定平衡点和振荡;随着去极化电流的增加,低电位水平的极化静息通过分岔变为放电,再经分岔变为高电位水平的去极化静息[2-3].极化静息是通常的对应正常生理状态的静息.神经系统通过电活动进行信息处理,进而实现其所负载的感觉、运动控制和脑高级功能等[4].因此,神经电活动的分岔在信息处理中起到重要的作用.一般而言,神经系统的兴奋性作用会引起静息变为放电或引起电活动的增强,抑制性作用会引起放电变为静息或是引起放电活动降低.但是,一些神经系统的理论研究发现了与常规概念不同的现象[5,7-9].例如,在靠近Hopf分岔点附近的极化静息,不仅是兴奋性作用,抑制性刺激也能引起放电或增强电活动:包括一次阈上的抑制性脉冲刺激可以诱发一次动作电位,两次时间间隔合适的阈下抑制性脉冲刺激也能够诱发一次动作电位[4];在兴奋性作用能引起的放电的频率较低时,随机施加一些抑制性刺激,可以增加放电频率[7];具有合适相位差的周期性抑制性刺激(在前)和周期性兴奋性刺激(在后)的联合作用,比单纯的周期性兴奋性刺激更能引起放电增强[8].更为重要的是,最后一个理论预期已经被实验证实[8-9],这也说明了理论预期的重要性.但是,这些研究只关注了极化静息和来自于神经系统外部的作用或调控,而抑制性的自我反馈对于神经系统的极化静息行为有什么样的调控作用,比如能否诱发出稳定、持续的放电行为,目前还不清晰.
在现实神经系统,一个神经元的抑制性自突触(autapse)就能够实现负反馈,将动作电位转变为抑制性自突触电流作用到该神经元自己[14-16].因此,揭示负反馈电流对电活动的影响,不仅是揭示负反馈的调控机制的作用,还有助于从理论上来预期抑制性自突触的潜在功能.自突触是一个起始和结束于同一个神经元的特殊突触,已经被实验证实在中枢神经系统(脑)的神经元中普遍存在[14-21].实验揭示兴奋性自突触能够诱发持续性的放电[18],众多理论研究揭示了自突触可以引起神经元的兴奋性转迁、随机共振和节律模式的转迁以及神经元网络的螺旋波、随机共振、同步及同步转迁等[22-33].特别是,兴奋性自突触能诱发单神经元及网络产生相干共振的现象得到了关注,这其中也包括时滞诱发的多重相干共振[31].但是,对于抑制性自突触的研究还有待开展.实验证实了抑制性自突触是慢突触(动作电位与自突触电流有相位差),单次抑制性脉冲刺激可以诱发具有抑制性自突触的神经元产生一次动作电位,及抑制性自突触可以引起放电精确性增强等[16-18],但是,关于抑制性自突触对极化静息的影响的理论研究尚不多见.
基于此,本文进行抑制性自突触介导的负反馈电流对极化静息的调控作用的理论模型研究.选用Rulkov神经元模型[34-35],考虑自突触介导的负反馈,考虑时滞来对应于慢突触.结果发现,合适的时滞和自突触增益能够诱发稳定平衡点,也就是静息转变为振荡,也就是放电.该结果不仅揭示了负反馈不同于常规概念的新调控作用,提供了神经电活动的反常规概念的复杂非线性行为,实际上也揭示了抑制性自突触的潜在功能.
1 模型
1.1 无自突触的单神经元模型、超临界Rulkov模型
超临界Rulkov模型是二维离散映射神经元模型,已经被广泛用于研究神经元的动力学行为[31-32]. 本研究选取这一模型,其动力学方程如下 \begin{equation} \label{eq1} \left. {{\begin{array}{l} {x_{n + 1} = f(x_n ,y_n + \beta ) + I} \\ {y_{n + 1} = y_n-\mu (x_n + 1-\sigma )} \\ \end{array} }} \right\} (1) \end{equation} 其中,$x_{n}$代表神经元膜电位,$y_{n}$是神经元细胞膜上离子通道的门控离子浓度. $n=0,1, 2, 3, \cdots $, 代表迭代次数.设定为不同数值可用于模拟特定神经元行为,$\beta $和$\sigma $代表外部对系统的影响,$I$为背景电流,方程中$f\left( {x,y + \beta } \right)$ 的函数形式如下其中,参数$\alpha =1$和$\sigma $为系统参数,$\mu =0.004$, $\beta =1$. 无自突触的神经元模型的研究中,{$\sigma $}和$I$为控制参数. Rulkov模型是无量纲的.
1.2 具有自突触介导的反馈电流的Rulkov神经元模型
在超临界Rulkov模型引入抑制性自突触介导的负反馈电流,模型的表达式如下式中,$I_{{aut}} =-g\left( {x_n-x_{{re}} }\right)\dfrac{1}{1 + {e}^{-\lambda \left( {x_{n-\tau }-\varTheta _{s} } \right)}}$为自突触电流.
其中,$g$为突触耦合强度,决定着耦合电流的强度;$\tau $为突触的时滞; $x_{{re}}$为突触的反转电位,决定突触是兴奋性还是抑制性的;$\lambda $决定了突触电流的响应速度, 是确保放电脉冲转换为非零的耦合电流的膜电位阈值. 当耦合强度$g=0$时,耦合电流$I_{{aut}} = 0$,自突触模型退化为无自突触的Rulkov模型.
本研究中$x_{re} =-1.6$,确保自突触是抑制性的和耦合电流脉冲是负的;$\varTheta _{ s} =-0.7$,膜电位$x_{n-\tau } $高于该阈值的时段对应的抑制性自突触电流为负、也就是抑制性的,低于该阈值的时段耦合电流为0,$\lambda = 30$,$I=0$.
2 结果与讨论
2.1 无自突触神经元模型的分岔和电活动
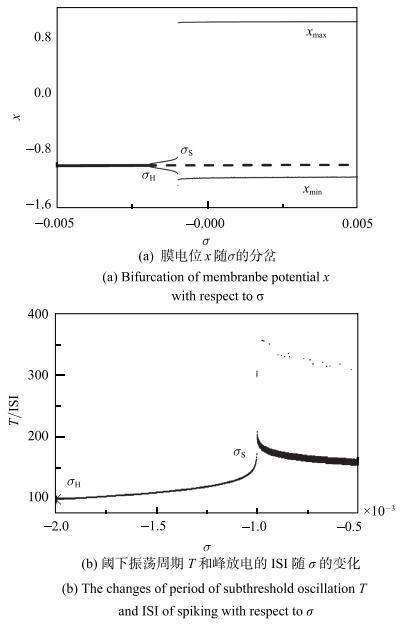
超临界Rulkov模型的一个重要特征就是静息能通过超临界Hopf分岔产生阈下振荡和放电,如图1所示. 当$I=0$时,超临界Rulkov神经元模型随参数$\sigma $的分岔如图1(a)所示. 对应稳定平衡点的静息(粗实线,$\sigma < \sigma _{H})$经过Hopf分岔($\sigma =\sigma _{ H})$变为不稳定平衡点(虚线,$\sigma > \sigma _{ H})$,同时产生对应小幅值极限环的阈下振荡($\sigma _{H }< \sigma < \sigma _{ S})$,再变为对应大幅值的放电($\sigma > \sigma _{ S})$,$x_{\min}$ (上细实线)和$x_{\max}$ (下细实线)对应振荡的极小值和极大值. 图1(b)展现了峰峰间期(interspike interval, ISI)随$\sigma $变化的情况.图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1无自突触Rulkov模型的电活动随$\sigma $变化
Fig. 1The changes of eletronic behaviors of Rulkov model without autapse with respect to $\sigma $
$\sigma =\sigma _{H}=-0.002$为超临界Hopf分岔点,如图1(a)所示. 当$\sigma < \sigma _{ H}$时,系统的动力学行为为稳定平衡点 (粗实线),对应于静息状态. 例如,当$\sigma =-0.003$时,神经元的动力学行为为稳定焦点,系统稳定后表现出静息行为,膜电位为$-1.003$,如图2(a)所示,相图上对应于一个点 ($-9.0\times 10^{-6}$,-1.003),如图2(e)所示,其中静息(圆点)、阈下振荡(细线)和峰放电(粗线),箭头表示运行方向.
图2
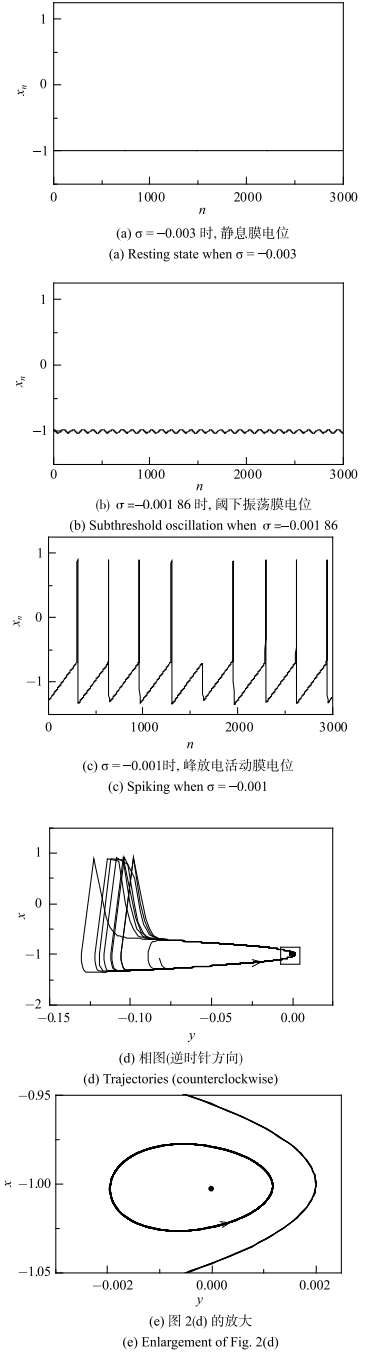 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2无自突触Rulkov神经元模型不同电活动模式
Fig. 2Different electronic behaviors of Rulkov model
当$\sigma $增加越过分岔点$\sigma_{H}$,稳定焦点变为不稳定平衡点(虚线), 同时产生一小幅值振荡的稳定极限环,对应于阈下振荡,振荡的极小值和极大值分别对应$x_{min}$和$x_{max}$的细实线, 如图1(a)所示. 例如,当$\sigma = -0.001~86$时,阈下振荡行为如图2(b)所示,振荡周期$T=102$. 阈下振荡存在的参数区间为$\sigma _{H} < \sigma < \sigma _{S} \approx -0.001$,随着{$\sigma $}增大,阈下振荡的振荡幅值增加,如图1(a)所示,振荡周期也增大,如图1(b)所示. 当$\sigma = -0.001~003$时,振荡周期$T=162$. 阈下振荡的周期$T$的范围约在 102$\sim $172,如图1(b)所示. 图2(d)对应图2(a)~图2(c)的静息、阈下振荡和峰放电活动的相图,图2(e)是它的局部放大图,箭头表示运行方向为逆时针方向.
当$\sigma $增加而越过$\sigma_{S} \approx-0.001$时,振荡幅值突然增大,动力学行为也从阈下振荡变为放电. 例如,当$\sigma =-0.001$时,神经元表现出放电行为,膜电位如图2(c)所示,最小的放电ISI为296. 进一步增大$\sigma $,ISI以极快的速度衰减至200左右.
2.2 无自突触神经元模型电活动的相图及推测
图2(a)~图2(c)所示的静息、阈下振荡和放电的电活动的相图如图2(d)所示,图2(e)是图2(d)的放大图.图2(c)的放电相图为一逆时针方向的大幅振荡的闭环;图2(b)的阈下振荡的相图为一逆时针方向的小幅值的闭环,位于放电的闭环的内部的膜电位$x$较小但$y$较大处;图2(a)的静息的相图为一点,位于阈下振荡的闭环内部,其膜电位$x$的中部.根据图2(d)和图2(e)所示相图的位置关系,可以进行如下推测:如果静息状态的电活动轨迹也就是图2(d)和图2(e)所示的圆点受到较大的扰动,使得电活动轨迹能穿越阈下振荡的闭环达放电活动轨迹附近,就会诱发一个动作电位或一次放电产生,然后再回到静息状态;但是,如果扰动较小,不能达到放电轨迹附近,则不能诱发动作电位或放电.由于圆点在$x$方向位于阈下振荡闭环的中部,向上或向下离放电的轨线的距离是相当的,那么,不论正向或兴奋性的,或者是负向或抑制性的膜电位扰动,只要扰动足够大,就都能引起放电或动作电位.这是Hopf分岔点附近的焦点所对应的静息所具备的特点,而鞍结分岔点附近的结点所对应的静息,不具备负向扰动能够产生放电或动作电位的特性[5].
2.3 兴奋性脉冲和抑制性脉冲刺激静息诱发动作电位
我们将通过对静息状态分别施加一个兴奋脉冲和一个抑制脉冲来验证我们的推测.将无自突触的Rulkov模型中的$I$选为一脉冲电流$I_{n}$,其脉冲宽度$\Delta T=11$,脉冲强度为$\Delta A$,如图3(a)和图3(b)虚线所示.图3
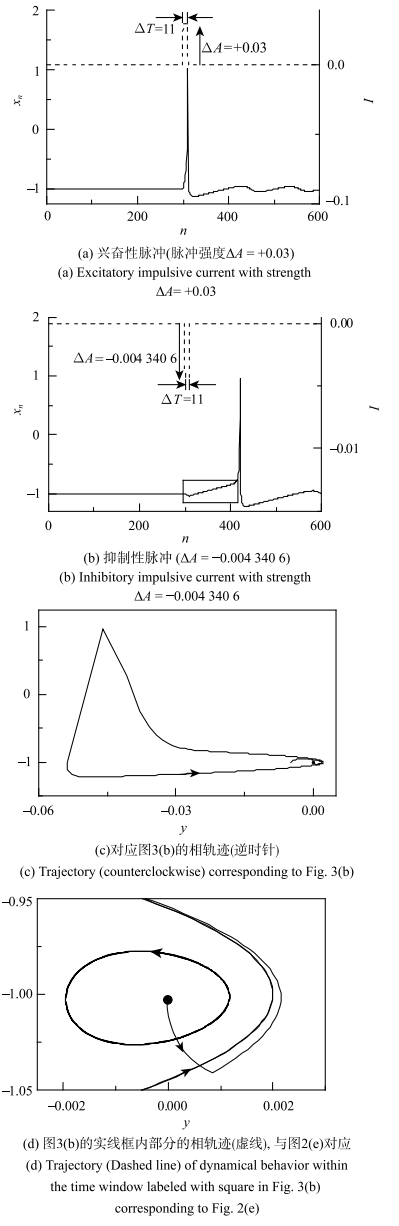 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3受到兴奋性质不同的脉冲电流,静息($\sigma =-0.003$)可以变为动作电位(脉冲宽度$\Delta T=11$)
Fig. 3An action potential can be evoked when the resting state ($\sigma =-0.003$) is stimulated by an impulsive current (impulsive width $\Delta T=11$) with different excitable characteristics
当$\sigma \!=\!-0.003$,无自突触神经元的电活动为静息.引入$\Delta A\!=\!+0.03$的正向或兴奋性脉冲电流(图3(a)的虚线), 会诱发一次放电或动作电位,然后系统衰减振荡到静息,如图3(a)实线所示. 这验证了上述的推测,也符合传统神经生理学的观点:兴奋性刺激诱发放电. 但是当$\Delta A$较小时,不能诱发动作电位.
而引入$\Delta A=-0.004~340~6$的负向或抑制性电流(图3(b)的虚线)的刺激,也会诱发一次放电或动作电位,然后系统衰减振荡到静息, 如图3(b)实线所示,与上述的推测结果一致. 这是不同于传统神经生理学的新观点:抑制性刺激诱发放电,然而它确实是符合非线性动力学的理论观点. 但较小的$\left| {\Delta A} \right|$不能诱发动作电位.能诱发动作电位的抑制性脉冲的强度阈值$\Delta A \approx -0.004~340~6$. 当脉冲强度$-0.004~340~6 <\Delta A<0$时, 抑制性脉冲($\Delta T=11$)不能诱发动作电位;当脉冲强度$\Delta A \le-0.004~340~6$时, 抑制性脉冲($\Delta T=11$)能诱发动作电位.
图3(b)所示的电活动的整个过程的相图如图3(c)所示,逆时针方向. 图3(b)的方框所示的时段, 也就是从静息膜电位水平到受到抑制性脉冲作用而放电前的时段,对应的相图如图3(d)所示.
从图3(c)和图3(d)可以看到大幅值的动作电位和小幅值的阈下振荡,受脉冲刺激前运动轨迹停留在静息(圆点)态上, 受到抑制性脉冲刺激后,相轨迹(虚线)离开静息然后逆时针往右下方运动、 也就是$x$降低,进而穿越对应图2(e)阈下振荡的极限环(细线)达到对应图2(e)的放电的运动轨迹(粗线)附近时, 膜电位开始上升然后产生动作电位. 该结果验证了2.2节的推测.
2.4 第二次抑制性脉冲刺激诱发动作电位的强度阈值
一次抑制性脉冲诱发的动作电位后的膜电位表现出衰减振荡,在衰减振荡的不同相位上,再次施加合适的抑制性脉冲刺激(虚线), 还会引起第二次动作电位(实线);但刺激相位不同时能再次诱发动作电位的抑制性脉冲的强度阈值不同, 如图4所示. 记第二次脉冲刺激相位与第一次动作电位峰值的峰值前的$x_{n }=\varTheta _{S}=-$0.7处(图中空心圆点)的时间差(也对应相位差)为$\Delta t$. 例如,当$\Delta t=86$时,$\Delta A=-0.003~0$的抑制性脉冲(虚线)不能诱发出第二个动作电位(实线),如图4(a)所示,而$\Delta A=-0.003~1$的脉冲(虚线),则能诱发出第二个动作电位(实线),如图4(b)所示. 类似地,当$\Delta t=207$时,$\Delta A=-0.001~6$的抑制性脉冲(虚线)不能诱发动作电位(实线), 如图4(c)所示,而$\Delta A=-0.001~7$脉冲(虚线),则能诱发出第二个动作电位(实线), 如图4(d)所示. 这说明$\Delta t=86$时,诱发第二个动作电位的抑制性脉冲强度阈值为$-0.003~1$,而$\Delta t=207$时,该阈值变为$-0.001~7$.图4
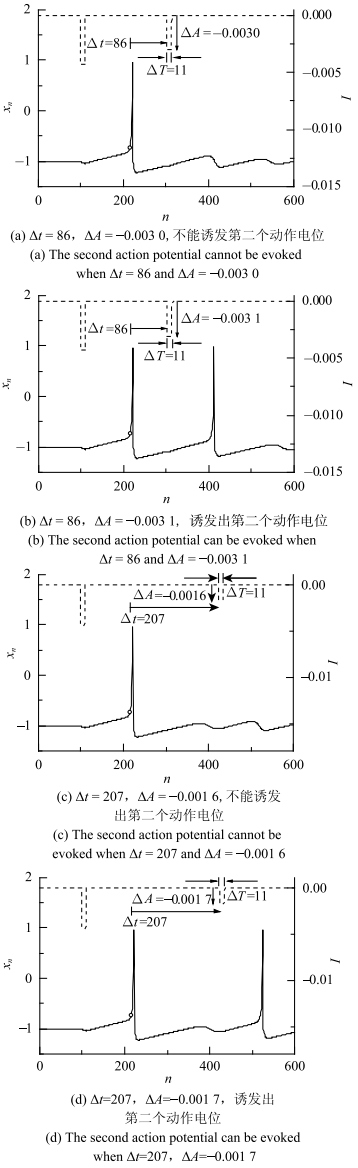 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4模型受二次刺激时序图,脉冲宽度$\Delta T=11$
Fig. 4Time series diagram of Rulkov model with two inhibitory impulsive currents, the impulsive width $\Delta T=11$
图4结果显示,在动作电位后的膜电位的合适相位$\Delta t$处给予合适强度$\Delta A$的抑制性脉冲作用,能够诱发出第二动作电位. 诱发第二次动作电位的抑制性脉冲($\Delta T$=11)的强度阈值$\Delta A$的绝对值$\left| {\Delta A} \right|$随着$\Delta t$的变化如图5所示. 可以发现,诱发第二个动作电位的脉冲阈值强度的绝对值$\left| {\Delta A} \right|$随着$\Delta t$的增大呈现出衰减振荡,有一定的周期性,振荡周期大约为100, 类似时滞诱发的多重共振现象.
图5
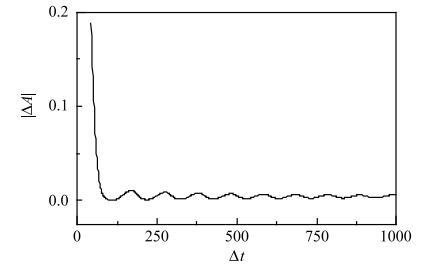 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5在膜电位的不同相位($\Delta t)$再次施加抑制性脉冲诱发第二个动作电位的脉冲强度$\Delta A$的绝对值$|\Delta A|$的阈值曲线
Fig. 5The strength threshold curve of absolute value $\left| {\Delta A} \right|$ of the second inhibitory impulsive current applied at different phase ($\Delta t)$ to evoke the second action potential
2.5 抑制性自突触诱发静息变为放电
当具有自突触的Rulkov模型的$g=0$和$\sigma =-0.003$时,在极大初值范围内其动力学行为为静息. 耦合强度$g$和时滞$\tau $取合适的值时,原先的静息会变为放电,说明抑制性自突触电流可使静息变为放电行为. 例如,图6(a)和图6(b)与图7(a)与图7(b)所示为系统稳定后膜电位与耦合电流时序图,图6与图7的模型初值分别相等. 当$\tau =51$时,$g=0.107$的自突触耦合电流为负向或抑制性脉冲串,如图6(a)虚线所示, 能够诱发静息变为放电,如图6(a)实线所示;而$g$略降低为0.106时,不能诱发放电, 耦合电流为零,如图6(b). 再例如,$\tau =197$时,$g=0.070$的抑制性耦合电流能够诱发放电,如图7(a);而$g=0.069$时不能诱发放电,如图7(b).图6
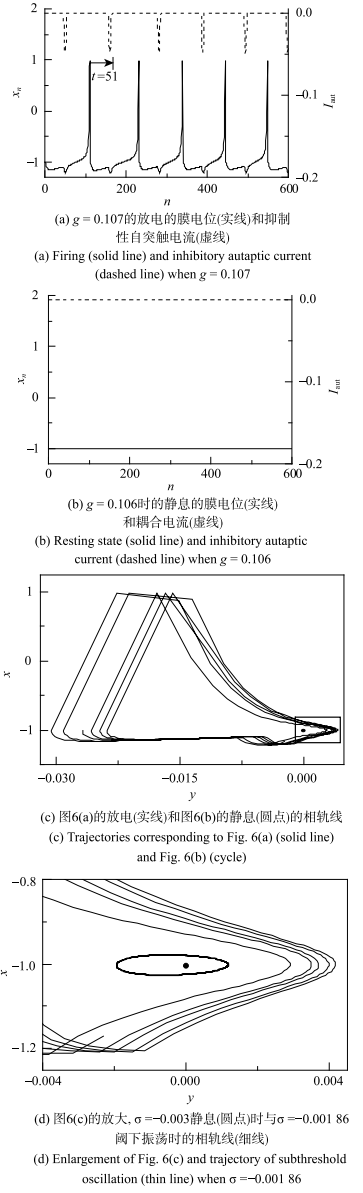 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6抑制性自突触($\tau =51$)诱发Rulkov模型($\sigma =-0.003$) 的电活动
Fig. 6Electronic behaviors induced by inhibitory autapse with $\tau =51$ in the Rulkov model ($\sigma =-0.003$)
图7
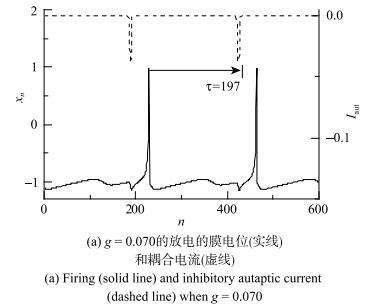 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7抑制性自突触($\tau =197$)诱发Rulkov模型($\sigma =-0.003$) 产生电活动
Fig. 7Electronic behaviors induced by inhibitory autapse with $\tau =197$ in the Rulkov model ($\sigma =-0.003$)
图7
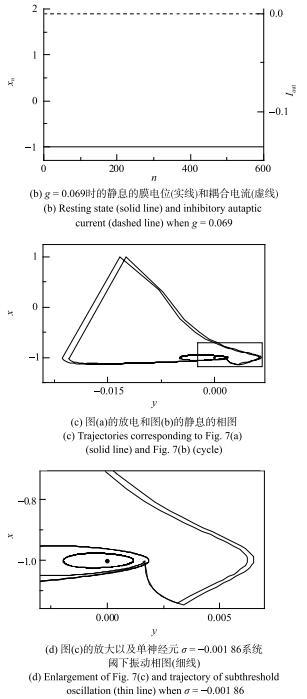 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7抑制性自突触($\tau =197$)诱发Rulkov模型($\sigma =-0.003$) 产生电活动(续)
Fig. 7Electronic behaviors induced by inhibitory autapse with $\tau =197$ in the Rulkov model ($\sigma =-0.003$ (continued))
在($y$, $x)$相平面上,图6(a)的放电($g=0.107$)和图6(b)的静息($g=0.106$)的轨迹如图6(c)所示, 图6(d)是图6(c)放大图以及单神经元$\sigma =-0.001~86$系统阈下振动相图(细线),与图2(c)和图2(d)的静息、阈下振荡和放电的相图类似, 静息对应的点位于放电的相轨迹的内部且靠近$x_{n}$较小、$y_{n}$较大处. 图7(c)和图7(d)的相图展示了图7(a)的放电($g=0.070$)和图7(b)的静息($g=0.069$)的轨迹, 表现出与图6(c)和图6(d)相类似的结果,表明较大的抑制耦合电流诱发静息变为放电.
与2.3节中的抑制性刺激脉冲相比较,抑制性自突触的耦合电流$I_{ aut}$的极大值由耦合强度$g$决定,与2.3节中的抑制性脉冲的强度$\Delta A$有关,而自突触的时滞$\tau $决定着抑制性耦合电流的作用相位,对应于2.3节中的抑制性脉冲的$\Delta t$. 在2.3节中发现合适的$\Delta t$和$\Delta A$组合能够诱发静息变为放电,而具有自突触的Rulkov模型中,抑制耦合电流诱发静息变为放电的动力学机制与2.3节相类似:不同$\tau $下的引起放电的自突触的耦合强度$g$一般是不同的,如图6和图7所示. 例如$\tau =51$时,诱发放电的自突触电导阈值为$g=0.107$,而$\tau =197$时的阈值为$g=0.070$. 结果说明,不同$\tau $下的诱发放电的抑制性自突触的阈值$g$不同.
2.6 抑制性自突触诱发的放电和静息的共存行为
抑制性自突触电流能诱发模型产生放电和静息的共存行为:不同的初值,有可能诱发模型产生放电,也可能还是静息.以时滞$\tau =214$,耦合强度$g=0.027$的自突触神经元模型为例(图8),选取不同初值,得到不同的动力学行为. 如图8(a)所示,当初值$x_{0}=1.25$, $y_{0}=0.1$时,模型产生放电行为;图8(b)中初值$x_{0}=1.25$,$y_{0}=-0.1$,模型的动力学行为为静息.图8
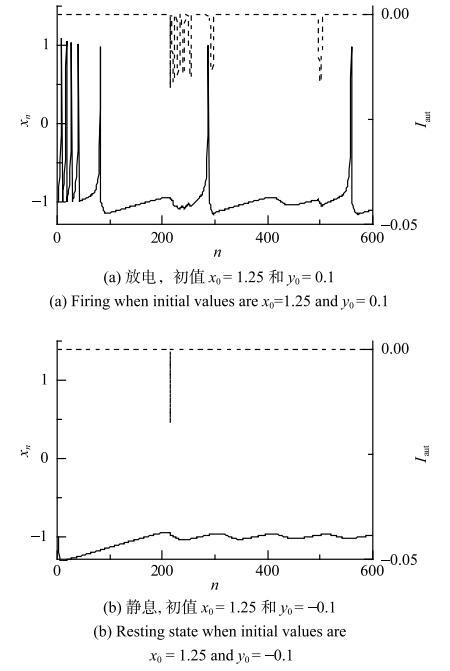 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8具有抑制性自突触的Rulkov模型在$\tau =214$和$g=0.027$的放电和静息行为的共存
Fig. 8Co-existence of firing and resting state of the Rulkov model with inhibitory autapse when $\tau =214$ and $g=0.027$
对应初值$x_{0}=-$1.25, $ y_{0}=0.1$的模型在$(\tau,g)$平面的动力学行为如图9(a)所示,对应初值$x_{0} =1.25$, $y_{0}=-0.1$的模型在($\tau $,$g)$平面的动力学行为如图9(b)所示,灰色区域代表放电行为,黑色区域代表静息行为.模型在($\tau $,$g)$双参数平面内的动力学行为的分布具有以下特征:
(1)诱发模型放电的$\tau -g$阈值曲线(即黑灰区域分隔线)随着$\tau $的增大呈现出衰减振荡现象,阈值曲线$g$随着$\tau $增大重复出现极大值、极小值,类似于时滞诱发的多重共振现象;
(2) 两图对应初值的放电阈值曲线有明显区别,图9(b)中静息区域明显大于图9(a). 这说明初值对模型的动力学行为有影响,模型存在放电和静息的共存行为.
图9
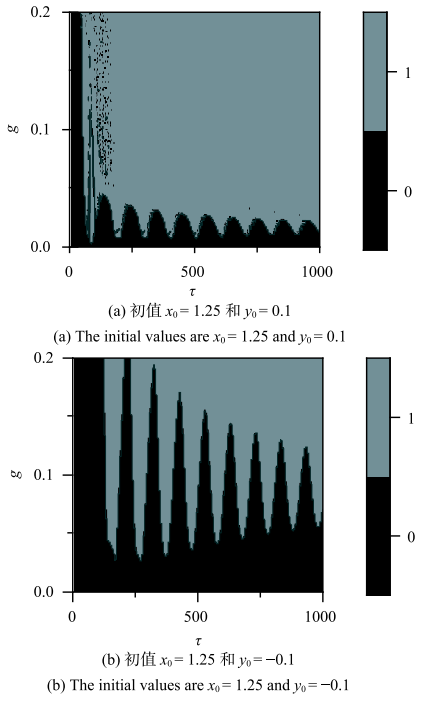 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9有抑制性自突触的Rulkov模型在($\tau ,g$)平面内的放电(灰)和静息(黑)的分布
Fig. 9Distribution of firing (gray) and resting state (black) in plane ($\tau ,g$)
随机选取100个初值, 分布如图10(c),在$\tau $从0至1000、$g$从0至4范围内,计算每个($\tau $,$g)$在100个不同初值下放电行为的概率,结果如图10(a)所示.
图10
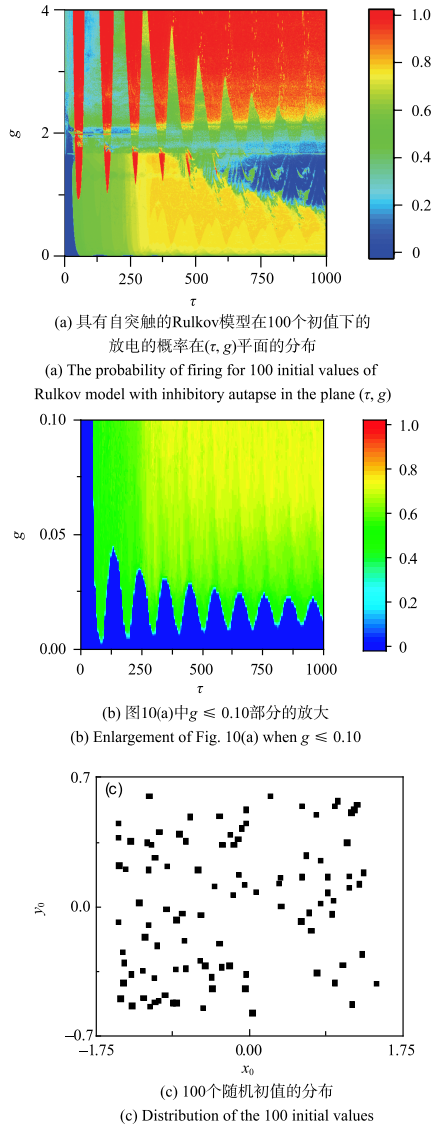 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10随机初值引起放电概率
Fig. 10Firing probability corresponding to 100 random initial values
图10(b)是图10(a) 中$g$较小时($g \le 0.10$)的放大图,蓝色部分代表放电行为概率为零,也就是静 息行为为100%的区域,可以将其看作是单稳的静息区域. 因此,该蓝色区域的边界线也可以看作是静息变为放电的概率阈值曲线, 与图10所示的电导阈值曲线有类似的特征:随着$\tau $的增大是衰减振荡的.
此外,$g$较大时且$\tau $较长时,放电概率接近或等于1 (红色),就是放电是主要的动力学行为,如图10(a)右上部分所示.当$\tau $较短时且$g$中等到较大时,概率分布随$\tau $变化大致具有一定的周期性,如图10(a)的左上部分所示. 此外,$g$中等且$\tau $较长时,概率分布的结果无明显规律,如图10(a)右中部 所示.
2.7 概率阈值曲线的特征
图10(b)中蓝色部分的边界也就是概率阈值曲线表现出衰减振荡现象,并且有一定的周期性. 阈值曲线的极大值的$\tau $分别为136, 242, 347, 449, 551, 655, 756, 856, 957,记为$\tau_{\max}(i), i=1, 2, 3, 4, 5, 6, 7, 8, 9$. 利用最小二乘法可以获得线性回归方程:$\tau_{\max}(i)=102.42 i+37.25$,相关系数为0.999~9. 阈值曲线的极小值的$\tau $分别为87, 197, 302, 404, 508, 609, 709, 811, 911,记为$\tau_{\min}(i), i=1, 2, 3, 4, 5, 6, 7, 8, 9$. 利用最小二乘法可以获得线性回归方程:$\tau_{\min}(i)=102.62 i-8.86$,相关系数为0.999~8.振荡的周期约为102~103,与图5所示的抑制性脉冲电流阈值曲线的振荡周期一致.3 结论
本研究提出了非线性系统的负反馈可以将稳定变为振荡、对应非线性神经系统的抑制性自突触电流可以诱发静息变为放电、的新观点. 该观点具有以下意义:对于不具有自突触的神经元,给出了一种新的调控放电的措施;对于具有自突触的神经元,揭示了自突触的可能的潜在作用,当然,还要等待实验的验证. 不同时滞下诱发放电的反馈增益,也就是自突触的耦合强度的阈值曲线表现类似多重相干共振的衰减振荡特征,振荡周期与靠近Hopf分岔附近的电活动的周期相关,这就为通过调控反馈增益和时滞诱发出放电提供能了具体的理论依据. 而神经元的电活动是神经系统信息处理的基本环节,因此,负反馈或抑制性自突触也会在神经信息处理中发挥重要的作用.抑制性自突触电流可以诱发静息变为放电的现象确认,又为反常规概念的非线性动力学行为的添加了新的示例, 进一步丰富了反常规概念的非线性动力学行为的内容:以前更多关注了外界的抑制性刺激[4,7-9],而本研究则关注了内在的抑制性反馈. 除了抑制性作用在极化静息附近的研究,也有抑制性自突触作用到去极化静息附近的研究: 理论模型的结果揭示了抑制性自突触电流能够增大而不是缩小靠近去极化静息的放电的参数区间[36]. 此外,还有研究发现抑制性自突触能增强簇放电的电活动[37],兴奋性作用不是提高而是降低神经簇放电的簇内放电个数[38]. 除了这些神经元层次的结果,在网络层次的研究还发现,合适的抑制性耦合能够促进同相同步而不仅仅是引起反相同步, 还会引起网络内神经元放电频率的增加[39-46],并能通过分岔进行理论解释.本文结果和这些研究结果就较为全面和深入的揭示了神经系统的反常规概念的复杂非线性动力学, 丰富了非线性动力学的内涵.
此外,时滞是引起本文研究的反常现象产生的重要因素,今后应该加强对于时滞引起的复杂动力学[47-48]的研究.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI [本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
DOI [本文引用: 3]
DOIURL [本文引用: 2]
DOIURL [本文引用: 3]
DOIURL [本文引用: 2]
DOI [本文引用: 3]
Magsci

利用随机的相变动力学理论研究了一个具有不同相位的神经振子群模型,并考察神经振子群对刺激信息的处理及神经编码的动态演化. 通过对动力学模型的数值分析,在二维相空间上描述了神经元集群内不同振子簇发放动作电位时,数密度随时间演化的图像. 数值分析的结果表明该模型能够用来描述注意和记忆的神经动力学机制,并且证明了只有高维的神经动力学模型才能更深刻地描述神经元集群的动力学特性,而以往的编码模型丢失了大量有用的神经信息.
Magsci
利用随机的相变动力学理论研究了一个具有不同相位的神经振子群模型,并考察神经振子群对刺激信息的处理及神经编码的动态演化. 通过对动力学模型的数值分析,在二维相空间上描述了神经元集群内不同振子簇发放动作电位时,数密度随时间演化的图像. 数值分析的结果表明该模型能够用来描述注意和记忆的神经动力学机制,并且证明了只有高维的神经动力学模型才能更深刻地描述神经元集群的动力学特性,而以往的编码模型丢失了大量有用的神经信息.
DOIMagsci
<p>生物神经元系统中存在着丰富的同步模式, 不同同步模式的实现条件已经被广泛地研究. 然而, 不同同步模式之间的转迁是神经动力学研究领域的难点问题, 近年来在此方面开展了许多相应的研究工作. 本文主要阐述近年来在神经元系统同步转迁动力学方面的研究进展, 揭示神经元系统在耦合、时滞和网络拓扑等不同参数作用下呈现的复杂的同步转迁动力学行为及其可能的动力学机制. 最后总结研究进展的内容并提出对同步动力学今后研究的展望.</p>
DOIMagsci
<p>生物神经元系统中存在着丰富的同步模式, 不同同步模式的实现条件已经被广泛地研究. 然而, 不同同步模式之间的转迁是神经动力学研究领域的难点问题, 近年来在此方面开展了许多相应的研究工作. 本文主要阐述近年来在神经元系统同步转迁动力学方面的研究进展, 揭示神经元系统在耦合、时滞和网络拓扑等不同参数作用下呈现的复杂的同步转迁动力学行为及其可能的动力学机制. 最后总结研究进展的内容并提出对同步动力学今后研究的展望.</p>
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
DOIURL
DOIURL [本文引用: 2]
DOIURL
DOIURL [本文引用: 2]
DOIURL
DOIURL
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL
DOIURL
DOIURL
DOIURL
DOI
DOIURL
DOIURL
DOI [本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIMagsci [本文引用: 1]
<p>神经放电节律在神经系统功能实现中起着重要的作用.具有自突触(起始和结束于同一细胞的突触)的神经元普遍存在于神经系统,本文研究了单神经元模型在抑制性自突触作用下的放电节律.结果发现,随着时滞和/或耦合强度的增加,可以诱发Rulkov神经元模型放电节律的加周期分岔.随着放电节律的周期数的增加,平均放电频率增大,当时滞和/或耦合强度大于某一阈值时,频率大于没有自突触时的放电频率.用快慢变量分离方法可以获得没有自突触的神经放电节律的分岔结构,可用于认识外界负向脉冲诱发的新节律.这些新的节律模式与加周期分岔中的节律模式一致.研究结果不仅揭示了抑制性自突触可以诱发典型的非线性现象——加周期分岔,还给出了抑制性自突触可以提高放电频率的新现象,与以前的自突触压制放电的观点不同,进一步丰富了对抑制性自突触诱发的非线性现象的认识.</p>
DOIMagsci [本文引用: 1]
<p>神经放电节律在神经系统功能实现中起着重要的作用.具有自突触(起始和结束于同一细胞的突触)的神经元普遍存在于神经系统,本文研究了单神经元模型在抑制性自突触作用下的放电节律.结果发现,随着时滞和/或耦合强度的增加,可以诱发Rulkov神经元模型放电节律的加周期分岔.随着放电节律的周期数的增加,平均放电频率增大,当时滞和/或耦合强度大于某一阈值时,频率大于没有自突触时的放电频率.用快慢变量分离方法可以获得没有自突触的神经放电节律的分岔结构,可用于认识外界负向脉冲诱发的新节律.这些新的节律模式与加周期分岔中的节律模式一致.研究结果不仅揭示了抑制性自突触可以诱发典型的非线性现象——加周期分岔,还给出了抑制性自突触可以提高放电频率的新现象,与以前的自突触压制放电的观点不同,进一步丰富了对抑制性自突触诱发的非线性现象的认识.</p>
DOIMagsci [本文引用: 1]
非线性动力学在识别神经放电的复杂现象、机制和功能方面发挥了重要作用.不同于传统观念,本文提出了兴奋性作用可以降低而不是增加簇内放电个数的新观点.在簇放电模式休止期的适合相位施加强度合适的脉冲或自突触电流,能诱发簇内放电个数降低;电流的施加相位越早,所需的强度阈值越大,簇内放电个数越少.进一步,利用快慢变量分离获得的簇放电的动力学性质进行了理论解释.簇放电模式表现出低电位的休止期和高电位的放电的交替,存在于快子系统的鞍结分岔点和同宿轨分岔点之间;放电起始于鞍结分岔、结束于同宿轨分岔;越靠近同宿轨分岔从休止期跨越到放电所需的电流强度越大.因此,电流在休止期上的作用相位越早,就越靠近同宿轨分岔,因而从休止期跨越到放电需要的电流强度阈值越大,放电起始相位到同宿轨分岔之间的区间变小导致放电个数变少.研究结果丰富了非线性现象及机制,对兴奋性作用提出了新看法,给出了调控簇放电模式的新途径.
DOIMagsci [本文引用: 1]
非线性动力学在识别神经放电的复杂现象、机制和功能方面发挥了重要作用.不同于传统观念,本文提出了兴奋性作用可以降低而不是增加簇内放电个数的新观点.在簇放电模式休止期的适合相位施加强度合适的脉冲或自突触电流,能诱发簇内放电个数降低;电流的施加相位越早,所需的强度阈值越大,簇内放电个数越少.进一步,利用快慢变量分离获得的簇放电的动力学性质进行了理论解释.簇放电模式表现出低电位的休止期和高电位的放电的交替,存在于快子系统的鞍结分岔点和同宿轨分岔点之间;放电起始于鞍结分岔、结束于同宿轨分岔;越靠近同宿轨分岔从休止期跨越到放电所需的电流强度越大.因此,电流在休止期上的作用相位越早,就越靠近同宿轨分岔,因而从休止期跨越到放电需要的电流强度阈值越大,放电起始相位到同宿轨分岔之间的区间变小导致放电个数变少.研究结果丰富了非线性现象及机制,对兴奋性作用提出了新看法,给出了调控簇放电模式的新途径.
DOIURL [本文引用: 1]
DOIURL
DOIURL
DOIURL
DOIURL
DOIURL
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}