 ,1,2, аьЭЈДя,1,2
,1,2, аьЭЈДя,1,2The molecular mechanism of apical hook development in dicot plant
Min Cao,1,2, Tongda Xu,1,2ЭЈбЖзїеп: аьЭЈДя,НЬЪк,ВЉЪПЩњЕМЪІ,баОПЗНЯђЃКЩњГЄЫиаХКХзЊЕМЭООЖЕФбаОПМАЦфдкХЉзїЮяжаЕФгІгУЁЃE-mail:tdxu@sibs.ac.cn
БрЮЏ: аэВй
ЪеИхШеЦк:2021-03-22аоЛиШеЦк:2021-06-8ЭјТчГіАцШеЦк:2021-08-20
| ЛљН№зЪжњ: |
Received:2021-03-22Revised:2021-06-8Online:2021-08-20
| Fund supported: |
зїепМђНщ About authors

Вмчы,ВЉЪП,баОПЗНЯђЃКЩњГЄЫиаХКХзЊЕМЭООЖЕФЗжзгЛњжЦЁЃE-mail:
2014—2019ФъОЭЖСгкжаЙњПЦбЇдКЗжзгжВЮяПЦбЇзПдНДДаТжааФ,дкаьЭЈДяПЮЬтзщЙЅЖСВЉЪПбЇЮЛ,ФПЧАдкУРЙњМгжнSalkInstituteforBiologicalStudiesНјааВЉЪПКѓбЕСЗЁЃВЉЪПЦкМфЕФбаОПЗНЯђЮЊЩњГЄЫиаХКХзЊЕМЕФЗжзгЛњжЦЁЃЭЈЙ§баОПЩњГЄЫи-TMK1етвЛЗЧОЕфЩњГЄЫиаХКХзЊЕМЭООЖЕїПиЖЅЖЫЭфЙГЗЂг§ЕФЗжзгЛњжЦ,НвЪОСЫЖЅЖЫЭфЙГЗЂг§Й§ГЬжаФкВрЯИАћжаИпХЈЖШЩњГЄЫивжжЦЯИАћЩьГЄЕФдвђ,ВћУїСЫЩњГЄЫи-TMK1-IAA32/34аХКХЭЈТЗдкЖЅЖЫЭфЙГЗЂг§Й§ГЬжагыОЕфЕФTIR1НщЕМЕФЩњГЄЫиаХКХЭЈТЗЕФВювьадЕїПиЛњжЦ,ЮЊжВЮяЩњГЄЫиаХКХзЊЕМЬсЙЉСЫаТЕФбаОПЗНЯђЁЃВЉЪПТлЮФЁЖЩњГЄЫиЭЈЙ§РрЪмЬхМЄУИTMK1ЕїПижВЮяВювьадЩњГЄЕФЗжзгЛњжЦЁЗЛёЕУ2020ФъжаЙњПЦбЇдКгХауВЉЪПЩњТлЮФЁЃ

еЊвЊ
ЫЋзгвЖжВЮяжжзгдкЭСШРжаУШЗЂКѓ,ЦфЯТХпжсЖЅЖЫЛсаЮГЩЭфЙГЕФЬиЛЏНсЙЙ,БЃЛЄзгвЖКЭЖЅЖЫЗжЩњзщжЏдкЦЦЭСЙ§ГЬжаВЛЪмЭСШРЛњаЕСІЕФЦЦЛЕ,БЃжЄгзУчЫГРћЦЦЭСЁЃЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬЗжЮЊЭфЙГаЮГЩЁЂЮЌГжМАДђПЊ3ИіНзЖЮ,ЦфКЫаФдкгкФкЭтСНВрЯИАћЕФВювьадЩњГЄЕМжТЭфЙГНсЙЙЁЃНќФъРДбаОПБэУї,жВЮяМЄЫиМАЛЗОГаХКХЖдЖЅЖЫЭфЙГЗЂг§ИїИіЙ§ГЬЦ№зХжСЙиживЊЕФЕїПизїгУЁЃШЛЖј,ЖЅЖЫЭфЙГСНВрЯИАћВЛЖдГЦЩњГЄШчКЮБЛОЋзМЕїПиЕФЗжзгЛњжЦФПЧАШдВЛЪЎЗжЧхГўЁЃБОЮФзлЪіСЫНќФъРДЖЅЖЫЭфЙГЗЂг§ЕїПиЛњжЦЕФбаОПНјеЙ,ВЂзХжиВћЪіСЫжВЮяМЄЫиЩњГЄЫидкЖЅЖЫЭфЙГЗЂг§жаЕФЙиМќзїгУМАЦфЗжзгЛњжЦ,ВЂЖдИУСьгђЮДРДЕФбаОПЗНЯђНјааСЫеЙЭћ,вдЦкЮЊЯрЙиСьгђЕФПЦбаШЫдБШЋУцСЫНтжВЮяМЄЫиаХКХЯрЛЅзїгУЕФФЃЪНЬсЙЉВЮПМЁЃ
ЙиМќДЪЃК
Abstract
After the seeds of the dicot model plant Arabidopsis germinate in the soil, the tip of the hypocotyl will form a specialized structure called apical hooks to protect the cotyledons and shoot apical meristems from the mechanical damage during the soil emerging process. The development process of the apical hook is divided into three stages: the apical hook formation, maintenance, and opening. In recent decades, studies have shown that different kinds of plant hormones and environmental signals play a vital role in the development of the apical hook. As the downstream of a variety of signals, the asymmetric distribution of auxin and the signal transduction pathways play a decisive role in the development of the apical hook. However, the detailed mechanism of the asymmetric signal transduction pathway of the cells on both sides of the apical hook is still unclear. In this review, we summarize the molecular mechanisms of the development of apical hook and further refine the role of auxin in the development of apical hook, and prospect for future research directions in this field.
KeywordsЃК
PDF (780KB)дЊЪ§ОнЖрЮЌЖШЦРМлЯрЙиЮФеТЕМГіEndNote|Ris|BibtexЪеВиБОЮФ
БОЮФв§гУИёЪН
Вмчы, аьЭЈДя. ЫЋзгвЖжВЮяЖЅЖЫЭфЙГЗЂг§ЕФЕїПиЛњжЦ. вХДЋ[J], 2021, 43(8): 723-736 doi:10.16288/j.yczz.21-105
Min Cao.
жжзгЕФУШЗЂЪЧжВЮяЩњУќжмЦкжажСЙиживЊЕФвЛВНЁЃдкЮќЪеЭСШРжаЫЎЗжжЎКѓжжзгПЊЪМУШЗЂ,ЦфЕквЛЯюЬєеНОЭЪЧЦЦЭСЖјГіНгЪеЙтКЭПеЦјРДНјааЙтКЯзїгУДгЖјЮЌГжжВЮяздЩэЕФе§ГЃЩњГЄЗЂг§ЁЃжВЮядкЭСШРжавдЛЦЛЏУчЕФзДЬЌбИЫйНЋЯТХпжсЩьГЄ[1,2],ВЂЧвВЩгУвЛЖЈЕФЗНЪНРДБЃЛЄЖЅЖЫЗжЩњзщжЏКЭзгвЖБмУтЫќУЧдкЦЦЭСЖјГіЕФЙ§ГЬжаЪмЕНЫ№КІЁЃЕЅзгвЖжВЮяКЭЫЋзгвЖжВЮяВЩШЁСЫСНжжНиШЛВЛЭЌЕФЗНЪНРДБЃЛЄЦфЖЅЖЫЗжЩњзщжЏКЭзгвЖЃКЕЅзгвЖжВЮяЖЅЖЫаЮГЩМсгВЕФХпбПЧЪ(coleoptile)зщжЏ,НЋЖЅЖЫЗжЩњзщжЏАќдкЦфжа[3];ЖјЫЋзгвЖжВЮядкЦЦЭСЙ§ГЬжа,згвЖКЭЖЅЖЫЗжЩњзщжЏМАвЛВПЗжЯТХпжсзщжЏЯђЯТЭфЧњ,аЮГЩЭфЙГзДНсЙЙ,гЩЭфЙГДІЕФЯТХпжсгХЯШНгДЅЭСШР,ШЫУЧНЋетИіОжВПЬиЛЏЕФзщжЏГЦжЎЮЊЖЅЖЫЭфЙГ(apical hook)[4]ЁЃжЎЧАЕФбаОПвбОБэУї,ОпгаЖЅЖЫЭфЙГШБЯнБэаЭЕФЭЛБфЬхЦфЦЦЭСЖјГіЕФФмСІЯджјадНЕЕЭ,етЫЕУїЖЅЖЫЭфЙГНсЙЙЖдгкЫЋзгвЖжВЮяЫГРћЦЦЭСЖјГіЗЧГЃживЊ[5,6,7]ЁЃ
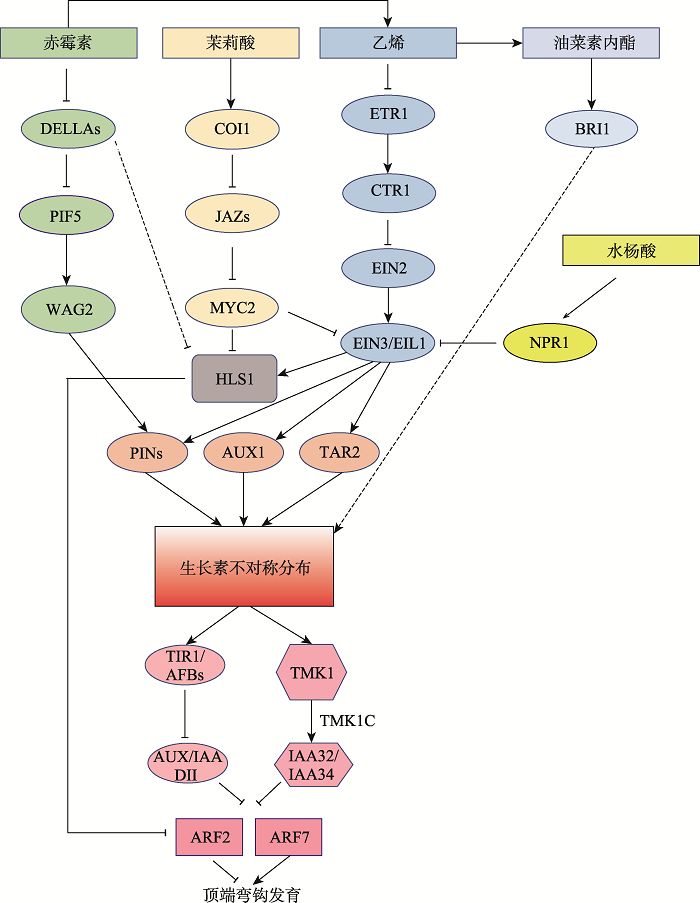
ЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬПЩвдЗжЮЊ3ИіНзЖЮЃКЖЅЖЫЭфЙГаЮГЩНзЖЮ,ЮЌГжНзЖЮКЭДђПЊНзЖЮ[8]ЁЃФПЧАбаОПШЫдБвбОПЩвдЪЕЪБЙлВьЖЅЖЫЭфЙГЕФећИіЗЂг§ЖЏЬЌЙ§ГЬ[9]ЁЃвдФтФЯНц(Arabidopsis thaliana)ЮЊР§,ЖЅЖЫЭфЙГЕФаЮГЩДггзУчЭЛЦЦжжЦЄПЊЪМ,гЩвЛВПЗжЯТХпжсНсЙЙГжајЯђЯТЭфЧњгыЩњГЄЗНЯђаЮГЩ180°;аЮГЩ180°жЎКѓЕФЖЅЖЫЭфЙГПЊЪМНјШыЮЌГжНзЖЮ,дкИУНзЖЮЖЅЖЫЭфЙГГжајЯђЯТЭфЧњБЃГж180°ВЂАщЫцЯТХпжсЕФПьЫйЩњГЄ;етИіЙ§ГЬГжајвЛЖЮЪБМфКѓЖЅЖЫЭфЙГжиаТДђПЊ,жБЕНзгвЖЭъШЋжБСЂГЩ0°,АщЫцзгвЖЭъШЋеЙПЊ[9,10,11,12]ЁЃећИіЖЅЖЫЭфЙГЗЂг§Й§ГЬЪмЕНЖржжМЄЫиМАЛЗОГаХКХЕФЕїПи,ДгЖјОЋзМПижЦЖЅЖЫЭфЙГЗЂг§ЕФВЛЭЌНзЖЮ,ЭъГЩЦЦЭСУШЗЂЙ§ГЬ(ЭМ1)ЁЃ
ЭМ1
 аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPT
аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPTЭМ1ЖржжМЄЫиЕїПиЫЋзгвЖжВЮяЖЅЖЫЭфЙГЗЂг§
ГрУЙЫиЁЂввЯЉЁЂмдРђЫсЁЂгЭВЫЫиФкѕЅМАЫЎбюЫсЕФаХКХЯрЛЅзїгУВЂзюжезїгУгкЩњГЄЫиЕФКЯГЩМАдЫЪф,ЕїПиЖЅЖЫЭфЙГДІЩњГЄЫиЕФВЛЖдГЦЗжВМ,гЩЯТгЮTIR1КЭTMK1НщЕМЕФЩњГЄЫиаХКХзЊЕМЭООЖзюжеЕїПиЖЅЖЫЭфЙГЕФЗЂг§ЁЃМ§ЭЗБэЪОе§ЕїПизїгУ,TаЭМ§ЭЗБэЪОИКЕїПизїгУ,ащЯпЕФМ§ЭЗБэЪООпЬхзїгУЛњжЦЮДжЊЁЃ
Fig. 1Plant hormone signaling network involved in the regulation of apical hook development
1 ввЯЉаХКХЭЈТЗДйНјВЂбгГЄЖЅЖЫЭфЙГЮЌГжЙ§ГЬ
1.1 ввЯЉКЯГЩЕїПиЖЅЖЫЭфЙГЕФЗЂг§
дкЖЅЖЫЭфЙГЗЂг§Й§ГЬжаЪЉМгЭтдДввЯЉ(ethylene)ДІРэ,ЛсЕМжТЖЅЖЫЭфЙГГЪЯж270°ЕФЭфЧњ,ВЂбгГЄЖЅЖЫЭфЙГЕФЮЌГжЙ§ГЬ[11,13],БЛГЦЮЊОЕфЕФввЯЉШ§жиЗДгІжЎвЛЁЃбаОПЗЂЯж,ЖдвАЩњаЭЭтдДЪЉМгввЯЉКЯГЩЧАЬх1-aminocyclopropane-1-carboxylic acid (ACC)ПЩвдДйНјЖЅЖЫЭфЙГМгОчЭфЧњ[14],ВЂЧвввЯЉЕФЙ§СПКЯГЩЭЛБфЬхeto1КЭeto2ОљБэЯжГіЖЅЖЫЭфЙГГЪЯж270°ЭфЧњЕФБэаЭ[15,16,17],етЫЕУїввЯЉЕФКЯГЩЖдЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬжСЙиживЊЁЃ
1.2 ввЯЉаХКХЭЈТЗЕїПиЖЅЖЫЭфЙГЕФЗЂг§
дкввЯЉЪмЬхETR1 (ethylene receptor 1)ЭЛБфЬхetr1жа,ЪЉМгЭтдДввЯЉЮоЗЈДйНјЖЅЖЫЭфЙГНјвЛВНЭфЧњ[18,19,20]ЁЃETR1ЯТгЮЕФМЄУИCTR1ЮЊввЯЉаХКХЭЈТЗжаЕФИКЕїПивђзг,ЦфШБЪЇЭЛБфЬхctr1-1дкВЛЪЉМгЭтдДввЯЉЕФЧщПіЯТвВБэЯжГіЖЅЖЫЭфЙГМгОчжС270°ЕФБэаЭ[21,22,23]ЁЃCTR1ЯТгЮЕзЮяEIN2ЮЊввЯЉаХКХЭЈТЗжаЕФе§ЕїПивђзг,ЦфЭЛБфЬхein2дкЪЉМгввЯЉЕФЧщПіЯТБэЯжГіЖдввЯЉВЛУєИаЕФБэаЭ[22,23,24,25]ЁЃввЯЉаХКХЭЈТЗжаживЊЕФзЊТМвђзгEIN3/EIL1вВВЮгыввЯЉЕїПиЕФЖЅЖЫЭфЙГЗЂг§Й§ГЬ,ЦфЭЛБфЬхein3eil1гыetr1КЭein2вЛбљ,дкЪЉМгввЯЉЕФЧщПіЯТБэЯжГіЖдввЯЉВЛУєИаЕФБэаЭ[26,27,28,29]ЁЃетаЉбаОПНсЙћБэУї,ввЯЉМАЦфаХКХЭЈТЗзЊЕМЭООЖЖдгкЕїПиЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬжСЙиживЊЁЃЧАЦкгабаОПБэУї,ввЯЉПЩФмЭЈЙ§ДйНјЩњГЄЫиКЯГЩЛљвђTAR2ЕФБэДяРДЬсИпЖЅЖЫЭфЙГДІЩњГЄЫи(auxin)ЕФКЌСП,НјЖјгАЯьЖЅЖЫЭфЙГСНВрЩњГЄЫиЗжВМ[30,31];Г§ДЫжЎЭт,гаЯрЙиБЈЕРжЄУїЭтдДЪЉМгЩњГЄЫидЫЪфвжжЦМСNPA (1-naphthylphthalamic acid),ПЩвдвжжЦctr1МАEIN3oxЕФЖЅЖЫЭфЙГМгОчЕФБэаЭ[32];ЭЌЪБ,ввЯЉЛЙПЩвдЭЈЙ§діЧПPIN3ЁЂPIN4КЭPIN7дкЖЅЖЫЭфЙГДІБэЦЄЯИАћЕФЖЈЮЛРДЕїПиЩњГЄЫидЫЪфвдМАЭфЙГСНВрЩњГЄЫиЕФВювьЛЏЗжВМ[12];ввЯЉПЩвддіЧПЖЅЖЫЭфЙГФкВрЯИАћжаAUX1ЕФбЛЗРДЪЙФкВрЯИАћЛ§РлИќЖрЕФЩњГЄЫи,НјЖјЕїПиЖЅЖЫЭфЙГЕФЭфЧњГЬЖШ[11]ЁЃзлЩЯЫљЪі,ввЯЉЭЈЙ§аЭЌЕїПиЩњГЄЫиЕФОжВПКЯГЩКЭМЋаддЫЪфРДНщЕМЖЅЖЫЭфЙГСНВрЩњГЄЫиВЛЖдГЦЗжВМ,зюжегАЯьЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬЁЃ
ДЫЭт,ввЯЉПЩвдДйНјHOOKLESS1ЕФБэДя[32,33],ВЂЧвДйНјHOOKLESS1ЕААзЕФЛ§Рл,НјЖјвжжЦЩњГЄЫиаХКХЭЈТЗжаARF2 (auxin response factor 2)ЕААзЕФЛ§Рл[33],ВЮгыЖдЖЅЖЫЭфЙГЗЂг§ЕФЕїПи[33]ЁЃ
дкжВЮяЦЦЭСЙ§ГЬжа,ЭСШРФІВСВњЩњЕФЛњаЕСІЛсгеЕМЯТХпжсМАЖЅЖЫЭфЙГВњЩњввЯЉ,ДгЖјЮЌГжЖЅЖЫЭфЙГ,ШЗБЃжВЮяГЩЙІЦЦЭСЖјГі[6,7,34]ЁЃввЯЉаХКХЭЈТЗЯрЙидЊМўЕФвХДЋбЇЗжЮівВжЄЪЕСЫетвЛЙлЕу,дкввЯЉаХКХЭЈТЗГжајМЄЛюЭЛБфЬхctr1ЛђЙ§СПКЯГЩЭЛБфЬхeto1жа,СНжжЭЛБфЬхОљПЩвдГЩЙІЦЦЭСЖјГі,ЖјЖдввЯЉВЛУєИаЕФЭЛБфЬх,Р§Шчein2КЭein3eil1,дђЦфЦЦЭСЕФИХТЪЯрБШвАЩњаЭвЊЕЭ[6,7,34]ЁЃетаЉЪ§ОнБэУїввЯЉаХКХЭЈТЗЪЧжВЮяаЭЌЭСШРЛЗОГМАжВЮяЗЂг§,ЧЩУюРћгУЖЅЖЫЭфЙГАяжњгзУчЦЦЭСУШЗЂЕФЙиМќЛњжЦЁЃ
2 ГрУЙЫиаХКХЭЈТЗЕїПиЖЅЖЫЭфЙГЗЂг§
2.1 ГрУЙЫиКЯГЩЕїПиЖЅЖЫЭфЙГЗЂг§
ЫцзХЖдЖЅЖЫЭфЙГбаОПЕФВЛЖЯЩюШы,ДѓМвЗЂЯжГрУЙЫи(gibberellins, GA)вВВЮгыжВЮяЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬЁЃЖдАЕЯТЩњГЄЕФФтФЯНцгзУчЪЉМгГрУЙЫиПЩвдДйНјЖЅЖЫЭфЙГМгОчЭфЧњ[32,35,36]ЁЃЖјЭтдДЪЉМгГрУЙЫиКЯГЩвжжЦМСPAC (paclobutrazol)Кѓ,гзУчЮоЗЈаЮГЩЖЅЖЫЭфЙГ,ЧвИУБэаЭПЩвдБЛдйДЮЪЉМгГрУЙЫиЫљЛиИД[36]ЁЃНјвЛВНбаОПБэУї,ФтФЯНцжаГрУЙЫиКЯГЩУИЭЛБфЬхga1-3БэЯжГіЖЅЖЫЭфЙГШБЪЇЕФБэаЭ[36],етЫЕУїГрУЙЫиЕФКЯГЩЕїПиСЫЖЅЖЫЭфЙГЕФЗЂг§ЁЃ2.2 ГрУЙЫиаХКХЭЈТЗЕїПиЖЅЖЫЭфЙГЗЂг§МАЛњжЦ
габаОПБэУї,ГрУЙЫиаХКХЭЈТЗжаЕФИКЕїПивђзгDELLAМвзхЕААзШБЪЇЭЛБфЬхвВБэЯжГіЖЅЖЫЭфЙГМгОчЭфЧњЕФБэаЭ,ЖјЕБЙ§СПЛ§РлDELLAЕААзЪБ,ЖЅЖЫЭфЙГЮоЗЈаЮГЩЭфЧњЕФаЮзД,жБНгНјШыЖЅЖЫЭфЙГЕФДђПЊНзЖЮ[32,35,36]ЁЃЖдгкГрУЙЫиЕїПиЖЅЖЫЭфЙГЗЂг§ЕФЗжзгЛњжЦ,габаОПБэУїГрУЙЫиПЩвдЭЈЙ§МЄЛюPIN3КЭPIN7ЛљвђЕФзЊТМРДЕїПиЩњГЄЫиЕФдЫЪф[37]ЁЃгыжЎЯрЖдгІ,pin3pin7ЫЋЭЛБфЬхБэЯжГіЖЅЖЫЭфЙГЖдГрУЙЫиВЛУєИаЕФБэаЭ[37]ЁЃетаЉНсЙћЫЕУїГрУЙЫиЮЛгкPIN3КЭPIN7ЕФЩЯгЮ,ЭЈЙ§ЕїПиЩњГЄЫидЫЪф,зюжеЕМжТЖЅЖЫЭфЙГСНВрЯИАћЩњГЄЫиХЈЖШЬнЖШЕФБфЛЏРДгАЯьЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬЁЃГ§ДЫжЎЭт,вВгажЄОнБэУїГрУЙЫиПЩвдЕїПиWAG2ЛљвђЕФБэДя[38]ЁЃWAG2ЪєгкAGCаЭМЄУИ(ЖЏЮяжаЭЌдДЕФЕААзМЄУИA,G,CМвзхЕФЭГГЦ),ПЩвдСзЫсЛЏЩњГЄЫизЊдЫЕААзPINЕААз[39,40]ЁЃгаШЄЕФЪЧ,WAG2ЕААздкЖЅЖЫЭфЙГДІГЪЯжВЛЖдГЦЕФБэДя,дкЖЅЖЫЭфЙГДІФкВрБэДяНЯИп,ЖјЭтВрБэДяНЯЕЭ[38]ЁЃWAG2дкЖЅЖЫЭфЙГФкВрЕФЛ§РлПЩФмЭЈЙ§СзЫсЛЏPINЕААзРДЕїПиPINЕААзЕФЩњГЄЫидЫЪфЛюад,ДгЖјЮЌГжЖЅЖЫЭфЙГФкВрЕФЩњГЄЫиЗДгІРДДйНјЖЅЖЫЭфЙГЕФЭфЧњзДЬЌ[39,40]ЁЃвВгабаОПБэУї,WAG2дкГрУЙЫигеЕМЯТЕФБэДяЪЧЪмзЊТМвђзгPIF5ЕїПи[41]ЁЃГрУЙЫиЭЈЙ§НЕНтЯТгЮЕФзЊТМвжжЦзгDELLAЕААз,ЪЙЕУPIF5МЄЛюWAG2ЕФзЊТМ,ЖјWAG2ЕААзПЩвдСзЫсЛЏPINЕААзРДЕїПиЩњГЄЫидкЖЅЖЫЭфЙГФкВрЕФЛ§Рл,зюжеЕїПиЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬ[37,38]ЁЃШЛЖјWAG2дкЖЅЖЫЭфЙГДІЕФОЋзМБэДяЕїПиЛњжЦШдВЛЧхГў,гаД§гкНјвЛВННтЮіЁЃ
ГрУЙЫиГ§СЫЭЈЙ§PINЕААзЕїПиЩњГЄЫидЫЪф,ЭЌЪБвВЭЈЙ§ввЯЉаХКХЭЈТЗРДЮЌГжЖЅЖЫЭфЙГБмУтЦфЙ§дчДђПЊЁЃЭЈЙ§вХДЋбЇКЭЩњЮяЛЏбЇбаОПЪжЖЮЗЂЯж,дкФтФЯНцdellaЭЛБфЬхжа,ЦфввЯЉКЌСПИпгквАЩњаЭжВЮя,етЫЕУїГрУЙЫиаХКХЭЈТЗПЩФмЕїПиввЯЉЕФКЯГЩЭООЖ[35,36,41]ЁЃНјвЛВНбаОПжЄУї,ввЯЉКЯГЩЯрЙиЕФУИACS5/ETO2КЭACS8ЛљвђЕФзЊТМЪмЕНГрУЙЫиЕФе§ЕїПи[35,36,41],ЦфЕїПиЛњжЦвВЪЧЭЈЙ§DELLAЕААзНЕНтКѓPIF5жБНгНсКЯACS8ЕФЦєЖЏзгЧјгђЧ§ЖЏзЊТМЁЃ
ГрУЙЫиГ§СЫПЩвдЕїПиввЯЉЕФКЯГЩ,ЛЙПЩвджБНгЕїПиЖЅЖЫЭфЙГЗЂг§Й§ГЬжаЕФживЊдЊМўHOOKLESS1ЕФБэДя[32,37]ЁЃбаОПБэУї,EIN3ПЩвджБНгНсКЯHOOKLESS1ЕФЦєЖЏзгЧјРДЕїПиHOOKLESS1ЕФБэДя,ЖјDELLAЕААзПЩвджБНгКЭEIN3ЕААзЯрЛЅзїгУ,РДвжжЦEIN3ЕФзЊТМЛюад[32,37],ДгЖјЕїПиЖЅЖЫЭфЙГЕФЗЂг§ЁЃ
3 мдРђЫсаХКХЭЈТЗдкЖЅЖЫЭфЙГЗЂг§жаЕФЕїПизїгУ
мдРђЫс(jasmonic acid,JA)ЕїПиЖЅЖЫЭфЙГЕФЮЌГжЙ§ГЬжївЊгыввЯЉаХКХЭЈТЗКЭЙтаХКХЭЈТЗгаЙиЁЃгабаОПБэУї,ЕБЖдАЕЯТЩњГЄЕФФтФЯНцгзУчЪЉМгЭтдДмдРђЫсДІРэКѓ,ЖЅЖЫЭфЙГЛсЬсЧАДђПЊ[42,43]ЁЃЭЌЪБ,мдРђЫсДІРэПЩвдвжжЦввЯЉЙ§СПКЯГЩЭЛБфЬхeto1МАаХКХМЄЛюЭЛБфЬхctr1ЕФЖЅЖЫЭфЙГЕФБэаЭ[42,43],ВЂЧвИУЙ§ГЬЪмЕНCOI1-JAZаХКХЭЈТЗЕїПи[42,43]ЁЃввЯЉДІРэПЩвдДйНјHOOKLESS1ЛљвђЕФБэДя,ЖјмдРђЫсДІРэПЩвдвжжЦввЯЉгеЕМHOOKLESS1БэДя[42,43]ЁЃНјвЛВНбаОПБэУї,мдРђЫсаХКХЭЈТЗжаЕФзЊТМвђзгMYC2ЁЂMYC3ЁЂMYC4гыввЯЉаХКХЭЈТЗжаЕФзЊТМвђзгEIN3КЭEIL1ЯрЛЅзїгУ,НјЖјвжжЦEIN3КЭEIL1ЕФзЊТМЛюадДгЖјвжжЦввЯЉаХКХЭЈТЗ[42,43]ЁЃДЫЭт,вВгабаОПБэУїмдРђЫсПЩвдЭЈЙ§вжжЦЙтаХКХЭЈТЗжазЊТМвђзгPIF4ЕФзЊТМЛюадРДвжжЦHOOKLESS1ЕФБэДя[44],ВЂЧвИУЙ§ГЬЪЧЭЈЙ§MYC2гыPIF4жБНгЯрЛЅзїгУНщЕМЕФ[44]ЁЃ4 ЫЎбюЫсаХКХЭЈТЗдкЖЅЖЫЭфЙГЗЂг§жаЕФЕїПизїгУ
ЫЎбюЫс(salicylic acid, SA)ЪЧживЊЕФУтвпЗРгљЯрЙиЕФжВЮяМЄЫи,жївЊВЮгыжВЮяПЙВЁЁЂвЖЦЌЫЅРЯЕШЩњЮябЇЙ§ГЬ[45,46,47]ЁЃШЛЖјЫЎбюЫсЖддчЦкжВЮяЗЂг§ЕФзїгУбаОПНЯЩйЁЃзюНќбаОПБэУї,ЫЎбюЫсаХКХЭЈТЗвВВЮгыЖЅЖЫЭфЙГЕФЗЂг§ЕїПиЁЃЭтдДЪЉМгЫЎбюЫсДІРэПЩвдДйНјЖЅЖЫЭфЙГЕФДђПЊ,ВЂЧввжжЦСЫввЯЉгеЕМЕФЖЅЖЫЭфЙГМгОчЭфЧњЕФБэаЭ,етЫЕУїввЯЉКЭЫЎбюЫсдкЖЅЖЫЭфЙГЗЂг§Й§ГЬжаЪЧЛЅЯрозПЙЕФ[48]ЁЃНјвЛВНбаОПБэУї,ЫЎбюЫсЪмЬхNPR1ЕФNЖЫПЩвджБНгКЭввЯЉаХКХЭЈТЗжаЕФEIN3дкЯИАћКЫФкЛЅзїЁЃNPR1НсКЯEIN3КѓвжжЦEIN3ЕФзЊТМЛюад,ДгЖјвжжЦHLS1МАЦфЫћEIN3/EIL1ЯТгЮЛљвђЕФБэДя,ДгЖјвжжЦЖЅЖЫЭфЙГЕФаЮГЩ[48]ЁЃ5 гЭВЫЫиФкѕЅдкЖЅЖЫЭфЙГЗЂг§жаЕФзїгУ
5.1 гЭВЫЫиФкѕЅЕФКЯГЩдкЖЅЖЫЭфЙГЗЂг§жаЕФзїгУ
дкжВЮяАЕаЮЬЌНЈГЩЗЂг§Й§ГЬжа,гЭВЫЫиФкѕЅ(brassinolides, BR)ЕФКЯГЩЛљвђЭЛБфЬхdet2БэЯжЮЊЖЅЖЫЭфЙГШБЪЇЕФБэаЭ[49]ЁЃДЫЭт,СэвЛИіПижЦгЭВЫЫиФкѕЅКЯГЩЭЛБфЬхcpdвВБэЯжЮЊЖЅЖЫЭфЙГШБЪЇЕФБэаЭ[50]ЁЃВЂЧвЭтдДЪЉМгeBL (24-epibrassinolide)ПЩвдЛжИДdet2ЖЅЖЫЭфЙГЗЂг§ШБЯнЕФБэаЭЁЃНјвЛВНбаОПЯдЪО,ЭтдДЪЉМггЭВЫЫиФкѕЅКЯГЩвжжЦМСвВЛсЕМжТЖЅЖЫЭфЙГЗЂг§ШБЯн[51]ЁЃетБэУїгЭВЫЫиФкѕЅЖдЖЅЖЫЭфЙГЕФЗЂг§ОпгаживЊзїгУЁЃ5.2 гЭВЫЫиФкѕЅЕїПиЖЅЖЫЭфЙГЗЂг§ЕФЛњжЦ
гаБЈЕРжЄУїввЯЉЖдЖЅЖЫЭфЙГЭфЧњЕФДйНјзїгУЪЧвРРЕгкгЭВЫЫиФкѕЅКЯГЩМАЦфЯТгЮаХКХЭЈТЗЕФЁЃввЯЉДІРэКѓЛсДйНјBRКЯГЩБЈИцЛљвђCPD:GUSЕФБэДя[51]ЁЃдкdet2ЭЛБфЬхжа,ЭтдДввЯЉДІРэВЛФмДйНјЖЅЖЫЭфЙГЕФЭфЧњНЧЖШЁЃетЫЕУїввЯЉЛсДйНјжВЮяЬхBRЕФКЯГЩРДЕїПиЖЅЖЫЭфЙГЕФЗЂг§[51]ЁЃКѓајЕФбаОПБэУї,bri1 bzr1-1DЫЋЭЛБфЬхБэЯжГіРрЫЦгквАЩњаЭЖЅЖЫЭфЙГЕФБэаЭ[52]ЁЃетЫЕУїBRаХКХЭЈТЗПЩФмВЮгыЕНЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬЁЃввЯЉДІРэВЛНіНіЕїПиBRЕФКЯГЩ,вВгаПЩФмНјвЛВНЕїПиBRЕФаХКХЭЈТЗНјЖјзюжегАЯьЖЅЖЫЭфЙГЕФЗЂг§ЁЃ6 ЩњГЄЫидкЖЅЖЫЭфЙГЗЂг§Й§ГЬжаЕФзїгУ
ЩњГЄЫизїЮЊжВЮязюживЊЕФМЄЫижЎвЛ,МИКѕВЮгыВЂЕїПиСЫжВЮяЩњГЄЗЂг§ЕФИїИіНзЖЮЁЃЩњГЄЫиЕФХЈЖШВювьдкЖржжзщжЏЭфЧњЩњГЄЙ§ГЬжаЦ№зХживЊЕФзїгУ[53]ЁЃР§ШчдкжВЮяЯђЙтадЩњГЄЙ§ГЬжа,ЩњГЄЫидкБГЙтВрЛ§Рл,ЕМжТСНВрЯИАћВювьадЩњГЄ,зюжеЪЙжВЮяЯђЙтЭфЧњЁЃдкжВЮяЕФИљЯђЕиадЩњГЄЙ§ГЬжа,ЩњГЄЫидкНќЕиВрЛ§Рл,ЕМжТНќЕиВрЕФЯИАћЩьГЄЪмЕНвжжЦ,ЕМжТжВЮяЯђЕиЩњГЄ[54,55,56]ЁЃШчЧАЫљЪі,ЦфЫћМЄЫиШчввЯЉЁЂГрУЙЫиЕШЕїПиЖЅЖЫЭфЙГЩњГЄ,зюжеЖМЛсОлНЙЕНЖдЩњГЄЫиХЈЖШЗжВМвдМАЯТгЮаХКХЭЈТЗЕФЕїПиЁЃдкЖЅЖЫЭфЙГДІ,ЭЈЙ§ЩњГЄЫиБЈИцЛљвђDR5-GUSЕШЗЂЯжЩњГЄЫиФкЭтВрГЪЯжВЛЖдГЦЗжВМ[57]ЁЃдкhookless1ЭЛБфЬхжа,ЖЅЖЫЭфЙГВЛФмаЮГЩ,ВЂЧвDR5-GUSдкСНВрЕФВЛЖдГЦЗжВМдкhookless1ЭЛБфЬхжавВЯћЪЇСЫ,етЫЕУїСНВрЕФЩњГЄЫиХЈЖШВювьЖдЖЅЖЫЭфЙГЕФаЮГЩЗЧГЃживЊ[57]ЁЃетаЉЗЂЯжвтЮЖзХЩњГЄЫидкЖЅЖЫЭфЙГФкЭтВрВЛЖдГЦЗжВМДгЖјгеЕМВЛЭЌЯТгЮаХКХЭЈТЗОіЖЈФкЭтЯИАћВювьадЩњГЄ[58],ЪЧЖЅЖЫЭфЙГЗЂг§ЕФКЫаФЛњжЦжЎвЛЁЃ6.1 ЩњГЄЫиЕФКЯГЩЕїПиМАЦфдкЖЅЖЫЭфЙГЗЂг§жаЕФзїгУ
ЩњГЄЫиКЯГЩзїЮЊжВЮяЬхФкЩњГЄЫиЕФживЊРДдДжЎвЛ,ЖджВЮяЕФЩњГЄЗЂг§Ц№зХЗЧГЃживЊЕФзїгУЁЃдкФтФЯНцжа,пХпс-3-БћЭЊЫс(indole-3-propionic acid, IPA)вРРЕЕФКЯГЩЭЈТЗЦ№зХжїЕМзїгУ[59,60,61]ЁЃжВЮяЬхФкКЌгавЛРрЩЋАБЫсАБЛљзЊвЦУИ,ПЩвдНЋЩЋАБЫс(Trp)зЊЛЏЮЊIPA[31,62]ЁЃвђДЫИУЕААзвВБЛУќУћЮЊTAA1 (TRYPTOPHAN AMINOTRANSFERASE of ARABIDOPSIS)[62]ЁЃКѓајЕФбаОПЗЂЯжTAA1ЕААздкФтФЯНцжаЛЙгаСНИіЭЌдДЕААзTAR1КЭTAR2[63,64]ЁЃИУМвзхЛљвђЭЛБфКѓжВЮяЩњГЄОпгабЯжиШБЯн,ВЂЧвЬхФкЩњГЄЫиКЌСПвВБШвАЩњаЭжВЮяЯджјНЕЕЭ[63,64]ЁЃДЫЭт,YUCМвзхЕААзвВВЮгыЕНЩњГЄЫиЕФКЯГЩЕїПижа[65,66,67]ЁЃYUCМвзхдкФтФЯНцжага11ИіГЩдБ,ЦфЙ§БэДяжВжъОљБэЯжГіЯрЫЦЕФЩњГЄЫиКЌСПЩ§ИпЕФБэаЭЁЃетЫЕУїYUCМвзхжаЕААзЕФЙІФмБШНЯРрЫЦ[65,66]ЁЃбаОПБэУї,TAAМвзхКЭYUCМвзхЪєгкЭЌвЛЬѕЩњГЄЫиКЯГЩЭЈТЗжаЕФСНИіВНжшжаЕФживЊУИЃКЩЋАБЫсОЙ§TAAЕФДпЛЏЩњГЩIPA,IPAОЙ§YUCМвзхЕФДпЛЏзюжеЩњГЩIAA[61,63]ЁЃЧАЦкбаОПЗЂЯжЕБЪЙгУЩњГЄЫиМЋаддЫЪфвжжЦМСNPAЛђеп1-NOA (1-naphthoxyacetic acid)ДІРэжВЮяЪБ,гЩгкЩњГЄЫиЮоЗЈдкЯИАћжЎМфдЫЪфЖјдкЩњГЄЫиКЯГЩЕФВПЮЛДѓСПЛ§Рл,ЕМжТжВЮяЕФЖЅЖЫЭфЙГЯћЪЇ,ВЂЧвзгвЖжаЕФDR5-GUSЛюадЯджјдіИп[11,12,33]ЁЃетЫЕУїзгвЖжаЕФЩњГЄЫиЪЧЖЅЖЫЭфЙГДІаЮГЩЩњГЄЫиВЛЖдГЦЗжВМХЈЖШЬнЖШЕФвЛИіживЊРДдДЁЃШЛЖј,ШЫУЧвВЗЂЯжСЫЩњГЄЫиПЩвддкЖЅЖЫЭфЙГЧјгђЕФЯИАћжаКЯГЩЁЃР§ШчБрТыЩњГЄЫиКЯГЩЭЈТЗжаживЊЕФСНРрДпЛЏУИYUC1ЁЂTAA1/WEI8КЭTAR2ЛљвђБЛжЄУїдкЖЅЖЫЭфЙГДІБэДя[31,68],ВЂЧввХДЋбЇжЄОнжЄУї,wei8-2 tar2-1КЭyuc1/ 2/4/6ЭЛБфЬхЕФгзУчЮоЗЈаЮГЩе§ГЃЕФЖЅЖЫЭфЙГ[31,68]ЁЃЕЋЪЧ,жЎЧАЕФбаОПБэУїдкЖЅЖЫЭфЙГЗЂг§Й§ГЬжа,етаЉЕїПиЩњГЄЫиКЯГЩЕФЛљвђдкЖЅЖЫЭфЙГДІВЂВЛГЪЯжВЛЖдГЦБэДяЁЃжЛгаTAR2ЛљвђдкввЯЉДІРэЕФЬѕМўЯТдкЖЅЖЫЭфЙГЮЌГжНзЖЮЕФФкВрЯИАћжаЕФБэДяЩдЮЂдіЧП[11,31]ЁЃетЫЕУїЩњГЄЫиЕФКЯГЩЫфШЛЪЧЩњГЄЫиЕФРДдД,ЕЋВЂВЛЪЧаЮГЩЖЅЖЫЭфЙГДІФкЭтВрЩњГЄЫиХЈЖШВюЕФжївЊдвђЁЃ
6.2 ЩњГЄЫиЕФдЫЪфЕїПиМАЦфдкЖЅЖЫЭфЙГЗЂг§жаЕФзїгУ
жЎЧАЕФбаОПвбОжЄЪЕ,ЩњГЄЫиЕФМЋаддЫЪфЖдЖЅЖЫЭфЙГЕФЗЂг§жСЙиживЊЁЃЩњГЄЫиЕФМЋаддЫЪфВЛНіНіДгзгвЖжаЯђЯТдЫЪфЩњГЄЫи,ВЂЧвЖЅЖЫЭфЙГФкЭтВрвВДцдкЩњГЄЫиЕФМЋаддЫЪфЁЃЩњГЄЫиМЋаддЫЪфЕААзЗжЮЊФкдЫЕААзКЭЭтдЫЕААзСНДѓРр,ЗжБ№ИКд№НЋЩњГЄЫидЫНјЯИАћЛђепдЫГіЯИАћ[69,70,71,72]ЁЃдкФтФЯНцжа,ЩњГЄЫиФкдЫЕААзжївЊгЩ4ИіЕААззщГЩ——AUX1ЁЂLAX1ЁЂLAX2КЭLAX3[71]ЁЃЦфжа,дкЖЅЖЫЭфЙГДІжївЊЦ№зїгУЕФЪЧAUX1КЭLAX3,ИКд№НЋЩњГЄЫиДгзгвЖДІЯђЯТдЫЪфЕНЖЅЖЫЭфЙГДІ[11,71]ЁЃAUX1жївЊЖЈЮЛгкБэЦЄЯИАћ,ЖјLAX3жївЊЖЈЮЛгкЮЌЙмзщжЏжаЁЃвђДЫ,ЩњГЄЫиФкдЫЕААзМвзхЕФзїгУжївЊЪЧНЋзгвЖКЭЖЅЖЫЭфЙГДІКЯГЩЕФЩњГЄЫиЯђЯТдЫЪф,ЖјЖдЩњГЄЫидкЖЅЖЫЭфЙГФкЭтВрЕФВЛЖдГЦЗжВМЕФНЈСЂзїгУНЯШѕ[11,71]ЁЃдкФтФЯНцжа,ЛЙгаСНДѓРрФЄЕААззїЮЊЩњГЄЫиЭтдЫЕААзЁЃетСНИіМвзхЗжБ№ЪЧPINЛљвђМвзх(Опга8ИіГЩдБ)вдМАСНИіBаЭATPНсКЯЕФзЊдЫЕААзABCB1КЭABCB19[69,72,73]ЁЃвХДЋбЇЗжЮіжЄУїabcb1,abcb19ЫЋЭЛБфЬхдкЖЅЖЫЭфЙГаЮГЩКЭДђПЊЙ§ГЬжагаШБЯн,ВЂЧвдкРћгУЩњГЄЫиБЈИцдЊМўDR5-GUSРДЙлВьЫЋЭЛБфЬхжаЩњГЄЫиЕФаХКХМЄЛюЧщПіЪБЗЂЯждкabcb1,abcb19ЫЋЭЛБфЬхжаDR5-GUSаХКХЯджјНЕЕЭ[74,75]ЁЃгаШЄЕФЪЧ,ABCB19БЛЗЂЯждкЖЅЖЫЭфЙГДІгаВЛЖдГЦБэДяЕФЯжЯѓЁЃбаОПБэУї,ABCB19ЖЈЮЛгкЖЅЖЫЭфЙГФкВрДІЕФБэЦЄЯИАћФЄЩЯ[75]ЁЃ
ЕБЪЙгУPINЕААзМвзхвжжЦМСNPAРДвжжЦЩњГЄЫиЯИАћЭтдЫЭООЖЪБ,ЖЅЖЫЭфЙГдкаЮГЩНзЖЮОЭЛсдьГЩжиДѓШБЯн,ЦфЖЅЖЫЭфЙГжБНгДђПЊГжајБЃГжжБСЂзДЬЌ[12,69],ВЂЧвЩњГЄЫидкЖЅЖЫЭфЙГФкВрДІЕФЛ§РлвВЛсЪмЕНвжжЦЁЃPINЕААзМвзхдкФтФЯНцжаЪЧживЊЕФвЛРрЩњГЄЫиЭтдЫЕААз[12,69,72,76,77]ЁЃдкPINЕААзМвзхжа,PIN3ЁЂPIN4КЭPIN7дкЖЅЖЫЭфЙГДІОљгаБэДя,ВЂЧвЦфБэДяВПЮЛгаВПЗжжиЕў[12,58]ЁЃгаШЄЕФЪЧ,PIN3ЁЂPIN4КЭPIN7вВБЛЗЂЯждкЖЅЖЫЭфЙГДІгаВЛЖдГЦБэДяЕФЯжЯѓ,ЫќУЧдкЖЅЖЫЭфЙГЭтВрДІЯИАћБэДяСПИќИп[12,58]ЁЃдкpin3ЙІФмШБЪЇЭЛБфЬхжа,гЩгкЩњГЄЫидЫЪфЕФШБЯнЕМжТжВЮяЮоЗЈдкЖЅЖЫЭфЙГФкВрЛ§РлзуЙЛКЌСПЕФЩњГЄЫи,ЕМжТpin3ЭЛБфЬхжаЕФЖЅЖЫЭфЙГВЛФмЭъШЋБеКЯ[12,58]ЁЃpin4КЭpin7ЕЅЭЛБфЬхЕФБэаЭНЯШѕ,ЕЋpin3pin4Лђpin3pin7ЕФЫЋЭЛБфЬхБэЯжГіНЯЧПЕФЖЅЖЫЭфЙГаЮГЩШБЯнЕФБэаЭ,ЫЕУїPIN3ЁЂPIN4КЭPIN7дкЕїПиЖЅЖЫЭфЙГаЮГЩЙ§ГЬжаДцдкзХЙІФмШпгр[12,58]ЁЃ
6.3 ЩњГЄЫиаХКХзЊЕМЭООЖМАЦфдкЖЅЖЫЭфЙГЗЂг§жаЕФзїгУ
ЩњГЄЫидкЖЅЖЫЭфЙГДІНЈСЂФкВрХЈЖШИп,ЭтВрХЈЖШЕЭЕФВЛЖдГЦХЈЖШВюКѓ,БиаывЊЭЈЙ§аХКХзЊЕМЭООЖРДЪЙЯИАћИажЊВЛЭЌХЈЖШЕФЩњГЄЫиаХКХ,зюжеЕїПиЯргІЕФЩњЮябЇЙ§ГЬ[78]ЁЃЩњГЄЫиКЫФкЪмЬхTRANSPORT INHIBITOR RESPONSE1/AUXIN SIGNALING F- BOX PROTEIN1-3 (TIR1/AFB1-3)дкЩњГЄЫиаХКХзЊЕМжаЦ№зХживЊЕФзїгУ[79,80,81]ЁЃЩњГЄЫигы TIR1 ЪмЬхНсКЯКѓ,ПЩвдДйНјTIR1ЕААзгыAUXIN/INDOLE-3- ACETIC ACID (Aux/IAA)ЕААзЕФЛЅзї[79,81]ЁЃAux/IAAПЩвдКЭзЊТМвђзгAUXIN RESPONSE FACTOR (ARF),дкЩњГЄЫиХЈЖШНЯЕЭЕФЧщПіЯТаЮГЩЖўОлЬх[82,83,84]ЁЃЕБЩњГЄЫиХЈЖШЩ§ИпМЄЛюTIR1аХКХЭЈТЗЪБ,TIR1ПЩвдЪЙ Aux/IAAЕААзЖрЗКЫиЛЏЪЙЦфЭЈЙ§26SЕААзУИЬхЭООЖНЕНтЁЃAux/IAAЕААзЕФНЕНтЪЙЕУЦфЖдARFЕФвжжЦзїгУНтГ§,РДМЄЛюARFЕФзЊТМЛюад,ЕїПиЛљвђБэДя[82,83,84]ЁЃбаОПБэУї,ЖЅЖЫЭфЙГЮЌГжНзЖЮЕФФкЭтВрЩњГЄЫиХЈЖШВю,ЕМжТФкЭтВрЯИАћЕФВЛЖдГЦЩьГЄ,НјЖјЮЌГжЖЅЖЫЭфЙГЭфЧњЕФзДЬЌ[58,85]ЁЃФПЧАЕФбаОПвбОЗЂЯжвЛаЉВЮгыЩњГЄЫиаХКХзЊЕМЭООЖжаЕФзщЗжвВВЮгыЕНЖЅЖЫЭфЙГЕФЕїПи,Р§ШчЩњГЄЫиКЫФкЪмЬхTIR1/AFBМвзхЕААзВЮгыЕїПиЖЅЖЫЭфЙГЕФЮЌГжЙ§ГЬ,ВЂЧвtir1afb1afb2afb3ЖрЭЛБфЬхБэЯжГіЖЅЖЫЭфЙГШБЪЇЕФБэаЭ[79,80,81]ЁЃГ§ДЫжЎЭт,Aux/IAAЕААзМвзхвВВЮгыЕНЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬжаЁЃбаОПжЄУї,ЕБжВЮяБэДяВЛЪмTIR1МвзхНЕНтЕФЭЛБфаЮЪНЕФAux/IAAЕААзЪБ(Р§ШчSHY2/IAA3ЁЂBDL/ IAA12ЕШ),ЖЅЖЫЭфЙГвВЛсГіЯжЗЂг§ШБЯнЕФБэаЭ[86,87,88]ЁЃетаЉжЄОнЫЕУїдкФкВрЩњГЄЫиЛ§РлКѓЦфЖдгІЕФаХКХзЊЕМЭООЖЖдгкЖЅЖЫЭфЙГЕФе§ГЃЗЂг§Й§ГЬжСЙиживЊЁЃГ§ДЫжЎЭт,ЩњГЄЫиаХКХЭЈТЗжаЕФживЊзЊТМвђзгМвзхARFМвзхЕААзвВВЮгыЕНетвЛЩњЮябЇЙ§ГЬжаЁЃзЊТММЄЛюзгШчNPH4/ARF7КЭARF19ЕФЙІФмШБЪЇЭЛБфЬхЛсБэЯжГіЖЅЖЫЭфЙГЗЂг§ШБЯнЕФБэаЭ,ЦфБэаЭгыЯдадЪЇЛюЕФAux/IAAЕААзЭЛБфЬхБэаЭРрЫЦ[89,90,91,92]ЁЃГ§ДЫжЎЭт,ARFМвзхжаЕФзЊТМвжжЦзгвВВЮгыЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬЁЃШчзЊТМвжжЦзгARF1КЭARF2ЪЧЖЅЖЫЭфЙГЗЂг§ЕФИКЕїПивђзг,arf1arf2ЫЋЭЛБфЬхБэЯжГідіЧПЭфЧњЕФЖЅЖЫЭфЙГБэаЭ[90]ЁЃгаШЄЕФЪЧ,ФтФЯНцжаетСНИібЧМвзхЕФзЊТМвђзгОљЭЈЙ§МЄЛюЛђепвжжЦЛљвђзЊТМРДДйНјЛђепвжжЦЯИАћЕФВЛЖдГЦЩњГЄ,НјЖјЕїПиЖЅЖЫЭфЙГЕФЗЂг§[90,91,93]ЁЃШЛЖј,ФПЧАЛЙУЛгажЄОнжЄУїARFМвзхзЊТМвђзгдкЖЅЖЫЭфЙГДІгаВЛЖдГЦБэДяЕФЧщПіЁЃвђДЫ,гЩЩњГЄЫиХЈЖШЬнЖШЕМжТЕФЬивьЪБПеМфМЄЛюЯТгЮзЊТММЄЛюаЭЛђепзЊТМвжжЦаЭARFЕААзПЩФмзюжеЕМжТЯТгЮЕФаЇгІЕААзЕФВЛЖдГЦБэДя,зюжеЕМжТЖЅЖЫЭфЙГЕФЭфЧњЙ§ГЬЁЃ
зюаТЕФбаОПЗЂЯж,дкЖЅЖЫЭфЙГЮЌГжНзЖЮ,ЦфФкВрЯИАћЕФИпХЈЖШЩњГЄЫиФмДйНјЯИАћФЄЩЯЕФРрЪмЬхМЄУИЕААзTMK1МєЧааЮГЩTMK1CФЉЖЫЦЌЖЮВЂДгЯИАћФЄзЊдЫЕНЯИАћжЪКЭЯИАћКЫФк,НјЖјЕїПиЯТгЮЭЈТЗ[94]ЁЃИУбаОПНјвЛВНЗЂЯж,МєЧаКѓЕФTMK1CФмЬивьКЭСНИіЗЧОЕфAux/IAAМвзхзЊТМвжжЦзгIAA32КЭIAA34ЛЅзїВЂСзЫсЛЏIAAЕААз[94]ЁЃШЛЖјгыTMK1CЛЅзїЕФIAA32/34ВЂВЛОпгагыTIR1ЛЅзїЕФНсЙЙЧјгђ,вђДЫВЛФмБЛTIR1ЫљНЕНт,етвтЮЖзХTIR1-НщЕМЕФЩњГЄЫиаХКХЭООЖКЭTMK1-НщЕМЕФЩњГЄЫиЭООЖЭЈЙ§бЁдёВЛЭЌIAAЕААзРДЧјЗжЯТгЮаХКХЭООЖ[94]ЁЃИУбаОПЛЙвтЭтЗЂЯж,гыжЎЧАБЈЕРЕФTIR1/AFBНщЕМЕФЩњГЄЫиЖдгкAux/IAAЕААзЗКЫиЛЏНЕНтЙ§ГЬЯрЗД,ЩњГЄЫиЭЈЙ§TMK1МєЧаКѓаЮГЩЕФTMK1CРДЮШЖЈIAA32КЭIAA34ЕААз,зюжевРШЛЭЈЙ§ARFзЊТМвђзгРДЕїПиЛљвђБэДя,дкЩњГЄЫиОлМЏЕФЕиЗНвжжЦЯИАћЩњГЄ,ДгЖјЕМжТЖЅЖЫЭфЙГФкЭтВрЕФВювьадЩњГЄ[94]ЁЃ
7 НсгягыеЙЭћ
жВЮязїЮЊвЛжжЙЬзХдкЭСШРжаЩњГЄЕФЩњЮя,ЦфжжзгЦЦЭСЖјГіЕФЙ§ГЬЖдгкжВЮяКѓајЭъГЩЙтКЯзїгУЛ§РлгЊбј,вдМАКѓЦкЩњГЄЗЂг§жСЙиживЊЁЃЫЋзгвЖжВЮяВЩгУЖЅЖЫЭфЙГетвЛЬиЛЏНсЙЙРДБЃЛЄзгвЖКЭЖЅЖЫЗжЩњзщжЏдкЦЦЭСЙ§ГЬжаВЛЪмЭСШРЛњаЕФІВСЕФЫ№ЩЫЁЃЖЅЖЫЭфЙГЕФЗЂг§Й§ГЬАщЫцзХЖржжжВЮяМЄЫиЕФаЭЌзїгУ,ЖјжВЮяЕФЗЂг§гыЖдЛЗОГЕФЯьгІЪЧЖржжМЄЫиаЭЌзїгУЕФНсЙћ,дкЮДРДвВШдЪЧжВЮяМЄЫиСьгђЕФбаОПШШЕуЁЃ7.1 ЦфЫћМЄЫидкЖЅЖЫЭфЙГЗЂг§Й§ГЬжаЕФзїгУ
ФПЧАЖЅЖЫЭфЙГЗЂг§Й§ГЬжаЕФжВЮяМЄЫибаОПжївЊМЏжагкввЯЉЁЂГрУЙЫиЁЂмдРђЫсЁЂЫЎбюЫсЁЂгЭВЫЫиФкѕЅКЭЩњГЄЫиЁЃШЛЖј,ФтФЯНцжаЦфЫћЕФживЊжВЮяМЄЫи,ШчЯИАћЗжСбЫи(cytokinin, CK)ЁЂЭбТфЫс(abscisic acid, ABA)КЭЖРНЧН№ФкжЌ(strigolactones, SLs)дкЖЅЖЫЭфЙГЗЂг§Й§ГЬжаЕФзїгУЛЙВЛЧхГўЁЃетаЉМЄЫиЪЧЗёВЮгыЖЅЖЫЭфЙГЕФЗЂг§ЕїПиЧвгыЦфЫћМЄЫиаХКХЭЈТЗЕФЛЅзїЛЙгаД§гкНјвЛВНбаОПЁЃЦфжа,ЯИАћЗжСбЫигыЩњГЄЫиЙВЭЌВЮгыИљЕФЗЂг§[95]ЁЃЖРНХН№ФкжЌВЮгыЕїПивЖЦЌаЮЬЌКЭЕиЩЯВПЗжЗжжЇЗЂг§[96]ЁЃЭбТфЫсдкЕїПижжзгУШЗЂЙ§ГЬЦ№зХжСЙиживЊЕФзїгУ[97],ЕЋЖдгкЭбТфЫсЪЧЗёВЮгыжжзгУШЗЂКѓЖЅЖЫЭфЙГЕФЗЂг§ЕїПиШДЯЪгаБЈЕРЁЃзюаТЕФбаОПЗЂЯжPP2CМвзхВЮгыСЫжжзгУШЗЂКѓЖЅЖЫЭфЙГЕФаЮГЩЙ§ГЬ[98],етПЩФмАЕЪОЭбТфЫсвВВЮгыСЫЖЅЖЫЭфЙГЕФЗЂг§ЕїПиЁЃвђДЫ,ЯИАћЗжСбЫиЁЂЖРНЧН№ФкжЌКЭЭбТфЫсЖдЖЅЖЫЭфЙГЗЂг§ЕФЕїПиМАЦфЗжзгЛњжЦЛсЪЧЖЅЖЫЭфЙГЗЂг§СьгђжЕЕУЙизЂЕФЮЪЬтЁЃ7.2 НјвЛВННтЮіжВЮяВЛЖдГЦЩњГЄЕФЗжзгЛњжЦ
ЖЅЖЫЭфЙГЕФЗЂг§ЪЧбаОПжВЮяВювьадЩњГЄЕФОЕфФЃаЭ,ЮЊНвЪОжВЮяВЛЭЌЗЂг§НзЖЮЕФВювьадЩњГЄЕїПиЛњжЦЬсЙЉживЊЯпЫїЁЃвђДЫ,ЖЅЖЫЭфЙГФкЭтВрВювьадЩњГЄЕФЕїПиЛњжЦЪЧЗёЖдЦфЫћВювьадЩњГЄ(ШчжВЮяЯђжиСІадЗДгІ)ЭЌбљЪЪгУгаД§гкНјвЛВНбаОПЁЃДЫЭт,ЩњГЄЫидкжВЮяЬхФкВЛЭЌзщжЏжаЕїПиЯИАћЩьГЄЕФЗжзгЛњжЦвВВЛОЁЯрЭЌЁЃР§Шч,ЩњГЄЫиВЛЖдГЦЗжВМЕїПиЯИАћВЛЖдГЦЩьГЄ,дкЖЅЖЫЭфЙГКЭИљЕФЯђЕиадЩњГЄЕФЩњЮябЇЙ§ГЬжа,ЩњГЄЫиЛ§РлЕФвЛВрЛсвжжЦЯИАћЕФЩьГЄ;ЖјдкЯТХпжсЯђЙтадЩњГЄетвЛЩњЮябЇЙ§ГЬжа,ЩњГЄЫиЛ§РлЕФвЛВрШДДйНјЯИАћЕФЩьГЄЁЃетаЉНсЙћБэУїЩњГЄЫидкВЛЭЌзщжЏжаЕїПиЯИАћЩьГЄЕФЙ§ГЬЪЧЖрбљЧвИДдгЕФЁЃжВЮяЩњГЄЫиаХКХЭЈТЗАќРЈTIR1НщЕМЕФОЕфзЊТМЕїПиЭЈТЗКЭTMK1НщЕМЕФЗЧОЕфаХКХЭЈТЗ,зїЮЊЖЅЖЫЭфЙГЕФКЫаФЛњжЦжЎвЛ,гаД§гкНјвЛВННтЮіЦфОЋзМЕїПиЛњжЦЁЃБШШч,зюГѕЖЅЖЫЭфЙГаЮГЩЪЧШчКЮЦ№ЪМЕФ?ЩњГЄЫиФкЭтВрВЛЖдГЦЗжВМШчКЮОЋзМБЛЩњГЄЫиКЯГЩЁЂдЫЪфКЭаХКХзЊЕМШ§епаЭЌЕїПиЕФ?TIR1КЭTMK1НщЕМЕФаХКХЭЈТЗЪЧШчКЮаЕїЕФ?зюНќЕФбаОПЗЂЯж,ЛњаЕСІФмЕїПижВЮяЖЅЖЫЭфЙГаЮГЩ,ЕЋИУЙ§ГЬвРРЕгкTMKМвзхНщЕМЕФЗЧОЕфаХКХЭЈТЗ,ЖјВЛвРРЕгкTIR1НщЕМЕФаХКХЭЈТЗ[99],ЦфОпЬхЛњжЦвВгаД§гкНјвЛВННтЮіЁЃ7.3 МЄЫиаХКХЭЈТЗЛЅзїЕФзщжЏКЭЯИАћЬивьад
жВЮяМЄЫиЕФаЭЌзїгУВЮгыСЫЖржжжВЮяЗЂг§ЕФЩњЮябЇЙ§ГЬ,ШЛЖјдкВЛЭЌЕФЩњЮябЇЙ§ГЬжажВЮяМЄЫиЕФаЭЌзїгУвВВЛОЁЯрЭЌЁЃР§ШчдкЖЅЖЫЭфЙГЗЂг§Й§ГЬжа,ввЯЉКЭмдРђЫсЁЂЫЎбюЫсЭЈЙ§ЛЅЯрозПЙРДЕїПиЖЅЖЫЭфЙГЕФЗЂг§,ЖјдкжВЮяПЙВЁЙ§ГЬжажВЮяЛсЭЌЪБМЄЛюввЯЉЁЂмдРђЫсКЭЫЎбюЫсЭЈТЗРДЕїПижВЮяУтвпЗДгІ[100]ЁЃжВЮяШчКЮдкУцЖдВЛЭЌЗЂг§ЛЗОГЯТВЩШЁВЛЭЌЩѕжСЯрЗДЕФМЄЫиЛЅзїЕїПиЗНЪНвВЪЧНёКѓЕФбаОПШШЕужЎвЛЁЃДЫЭт,IAA32/34жЛЬивьдкЖЅЖЫЭфЙГДІБэДя,етЫЕУїЬиЖЈзщжЏЛђЯИАћРраЭжаДцдкзХЬивьЕФЕААзДЋЕнаХКХРДЕїПиИУзщжЏЕФЩњЮябЇЙІФмЁЃвђДЫ,ЭЈЙ§ВЛЭЌзщжЏЕФзЊТМзщбЇЗжЮіРДбАевВЛЭЌзщжЏдкИаЪмЛЗОГаХКХЪБЕФЬивьадзщЗжПЩФмЪЧЮДРДМЄЫиаХКХзЊЕМЗНЯђЕФбаОПШШЕуЁЃИЃНЈХЉСжДѓбЇаьЭЈДяПЮЬтзщМђНщ
аьЭЈДяНЬЪкгк2014ФъШЋжАЛиЙњ,дкжаЙњПЦбЇдКЩЯКЃФцОГжааФГЩСЂПЮЬтзщ,2017ФъПЮЬтзщзЊЕНИЃНЈХЉСжДѓбЇ,жївЊЮЇШЦжВЮяЩњГЄЫиЕФЗЧзЊТМЕїПиЛњжЦЁЂжВЮяЩњГЄЫиХЈЖШаЇгІВњЩњЕФЗжзгЛњжЦМАЩњГЄЫиОжВПХЈЖШШчКЮБЛЩЯгЮЗЂг§КЭЛЗОГаХКХОЋзМЕїПиЕФЗжзгЛњжЦЕШСьгђНјаабаОПЁЃдкжВЮяМЄЫиЗЂг§КЭЛЗОГЯьгІжаШЁЕУСЫвЛЯЕСабаОПГЩЙћ,дкNatureЁЂScienceЁЂCellЁЂNature CommunicationsКЭPNASЕШЙњМЪжЊУћЦкПЏЗЂБэТлЮФ20РДЦЊЁЃгыДЫЭЌЪБ,ПЮЬтзщЛЙГаЕЃСЫПЦММВПЁЂЙњМвздШЛПЦбЇЛљН№ЮЏЁЂЩЯКЃЪаПЦбЇЛљН№ЮЏКЭИЃНЈЪЁПЦбЇаЛсЕШПЮЬтЁЃПЮЬтзщЭјеОЃК

(д№ШЮБрЮЏ: аэВй)
ВЮПМЮФЯз дЮФЫГађ
ЮФЯзФъЖШЕЙађ
ЮФжав§гУДЮЪ§ЕЙађ
БЛв§ЦкПЏгАЯьвђзг
DOI:10.1016/j.cub.2008.10.034URL [БОЮФв§гУ: 1]
DOI:10.1016/j.cub.2015.03.020URL [БОЮФв§гУ: 1]
DOI:10.1007/s00709-016-1023-6URL [БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
DOI:10.1080/15592324.2017.1330239URL [БОЮФв§гУ: 1]
[БОЮФв§гУ: 3]
DOI:10.1016/j.cub.2015.11.053URL [БОЮФв§гУ: 3]
PMID:10409511 [БОЮФв§гУ: 1]

Arabidopsis seedlings develop a hook-like structure at the apical part of the hypocotyl when grown in darkness. Differential cell growth processes result in the curved hypocotyl hook. Time-dependent analyses of the hypocotyl showed that the apical hook is formed during an early phase of seedling growth and is maintained in a sequential phase by a distinct process. Based on developmental genetic analyses of hook-affected mutants, we show that the hookless mutants (hls1, cop2) are involved in an early aspect of hook development. From time-dependent analyses of ethylene-insensitive mutants, later steps in hook maintenance were found to be ethylene sensitive. Regulation of differential growth was further studied through examination of the spatial pattern of expression of two hormone-regulated genes: an ethylene biosynthetic enzyme and the ethylene receptor ETR1. Accumulation of mRNA for AtACO2, a novel ACC (1-aminocyclopropane-1-carboxylic acid) oxidase gene, occurred within cells predominantly located on the outer-side of the hook and was tightly correlated with ethylene-induced exaggeration in the curvature of the hook. ETR1 expression in the apical hook, however, was reduced by ethylene treatment. Based on the expression pattern of ETR1 and AtACO2 in the hook-affected mutants, a model for hook development and maintenance is proposed.
[БОЮФв§гУ: 2]
DOI:10.1111/nph.2014.202.issue-4URL [БОЮФв§гУ: 1]
DOI:10.1242/dev.040790PMID:20110325 [БОЮФв§гУ: 7]
Dark-grown dicotyledonous seedlings form a hook-like structure at the top of the hypocotyl, which is controlled by the hormones auxin and ethylene. Hook formation is dependent on an auxin signal gradient, whereas hook exaggeration is part of the triple response provoked by ethylene in dark-grown Arabidopsis seedlings. Several other hormones and light are also known to be involved in hook development, but the molecular mechanisms that lead to the initial installation of an auxin gradient are still poorly understood. In this study, we aimed to unravel the cross-talk between auxin and ethylene in the apical hook. Auxin measurements, the expression pattern of the auxin reporter DR5::GUS and the localization of auxin biosynthesis enzymes and influx carriers collectively indicate the necessity for auxin biosynthesis and efficient auxin translocation from the cotyledons and meristem into the hypocotyl in order to support proper hook development. Auxin accumulation in the meristem and cotyledons and in the hypocotyl is increased approximately 2-fold upon treatment with ethylene. In addition, a strong ethylene signal leads to enhanced auxin biosynthesis at the inner side of the hook. Finally, mutant analysis demonstrates that the auxin influx carrier LAX3 is indispensable for proper hook formation, whereas the auxin influx carrier AUX1 is involved in the hook exaggeration phenotype induced by ethylene.
DOI:10.1242/dev.041277PMID:20110326 [БОЮФв§гУ: 9]
The apical hook of dark-grown Arabidopsis seedlings is a simple structure that develops soon after germination to protect the meristem tissues during emergence through the soil and that opens upon exposure to light. Differential growth at the apical hook proceeds in three sequential steps that are regulated by multiple hormones, principally auxin and ethylene. We show that the progress of the apical hook through these developmental phases depends on the dynamic, asymmetric distribution of auxin, which is regulated by auxin efflux carriers of the PIN family. Several PIN proteins exhibited specific, partially overlapping spatial and temporal expression patterns, and their subcellular localization suggested auxin fluxes during hook development. Genetic manipulation of individual PIN activities interfered with different stages of hook development, implying that specific combinations of PIN genes are required for progress of the apical hook through the developmental phases. Furthermore, ethylene might modulate apical hook development by prolonging the formation phase and strongly suppressing the maintenance phase. This ethylene effect is in part mediated by regulation of PIN-dependent auxin efflux and auxin signaling.
[БОЮФв§гУ: 1]
DOI:10.1111/nph.2014.202.issue-4URL [БОЮФв§гУ: 1]
DOI:10.1038/nature02516URL [БОЮФв§гУ: 1]
PMID:9952448 [БОЮФв§гУ: 1]
The Arabidopsis mutants eto1 (ethylene overproducer) and eto3 produce elevated levels of ethylene as etiolated seedlings. Ethylene production in these seedlings peaks at 60 to 96 h, and then declines back to almost wild-type levels. Ethylene overproduction in eto1 and eto3 is limited mainly to etiolated seedlings; light-grown seedlings and various adult tissues produce close to wild-type amounts of ethylene. Several compounds that induce ethylene biosynthesis in wild-type, etiolated seedlings through distinct 1-aminocyclopropane-1-carboxylic acid (ACC) synthase (ACS) isoforms were found to act synergistically with eto1 and eto3, as did the ethylene-insensitive mutation etr1 (ethylene resistant), which blocks feedback inhibition of biosynthesis. ACS activity, the rate-limiting step of ethylene biosynthesis, was highly elevated in both eto1 and eto3 mutant seedlings, even though RNA gel-blot analysis demonstrated that the steady-state level of ACS mRNA was not increased, including that of a novel Arabidopsis ACS gene that was identified. Measurements of the conversion of ACC to ethylene by intact seedlings indicated that the mutations did not affect conjugation of ACC or the activity of ACC oxidase, the final step of ethylene biosynthesis. Taken together, these data suggest that the eto1 and eto3 mutations elevate ethylene biosynthesis by affecting the posttranscriptional regulation of ACS.
DOI:10.1105/tpc.006882URL [БОЮФв§гУ: 1]
PMID:8211181 [БОЮФв§гУ: 1]
Ethylene behaves as a hormone in plants, regulating such aspects of growth and development as fruit ripening, flower senescence, and abscission. Ethylene insensitivity is conferred by dominant mutations in the ETR1 gene early in the ethylene signal transduction pathway of Arabidopsis thaliana. The ETR1 gene was cloned by the method of chromosome walking. Each of the four known etr1 mutant alleles contains a missense mutation near the amino terminus of the predicted protein. Although the sequence of the amino-terminal half of the deduced ETR1 protein appears to be novel, the carboxyl-terminal half is similar in sequence to both components of the prokaryotic family of signal transducers known as the two-component systems. Thus, an early step in ethylene signal transduction in plants may involve transfer of phosphate as in prokaryotic two-component systems. The dominant etr1-1 mutant gene conferred ethylene insensitivity to wild-type Arabidopsis plants when introduced by transformation.
DOI:10.1104/pp.011635URL [БОЮФв§гУ: 1]
PMID:9974395 [БОЮФв§гУ: 1]
The ETR1 receptor from Arabidopsis binds the gaseous hormone ethylene. A copper ion associated with the ethylene-binding domain is required for high-affinity ethylene-binding activity. A missense mutation in the domain that renders the plant insensitive to ethylene eliminates both ethylene binding and the interaction of copper with the receptor. A sequence from the genome of the cyanobacterium Synechocystis sp. strain 6803 that shows homology to the ethylene-binding domain of ETR1 encodes a functional ethylene-binding protein. On the basis of sequence conservation between the Arabidopsis and the cyanobacterial ethylene-binding domains and on in vitro mutagenesis of ETR1, a structural model for this copper-based ethylene sensor domain is presented.
PMID:8431946 [БОЮФв§гУ: 1]
We isolated a recessive Arabidopsis mutant, ctr1, that constitutively exhibits seedling and adult phenotypes observed in plants treated with the plant hormone ethylene. The ctr1 adult morphology can be phenocopied by treatment of wild-type plants with exogenous ethylene and is due, at least in part, to inhibition of cell elongation. Seedlings and adult ctr1 plants show constitutive expression of ethylene-regulated genes. The epistasis of ctr1 and other ethylene response mutants has defined the position of CTR1 in the ethylene signal transduction pathway. The CTR1 gene has been cloned, and the DNA sequences of four mutant alleles were determined. The gene encodes a putative serine/threonine protein kinase that is most closely related to the Raf protein kinase family.
DOI:10.1126/science.1225974URL [БОЮФв§гУ: 2]
DOI:10.1073/pnas.1214848109URL [БОЮФв§гУ: 2]
DOI:10.1016/j.cell.2015.09.037URL [БОЮФв§гУ: 1]
PMID:10381874 [БОЮФв§гУ: 1]
Ethylene regulates plant growth, development, and responsiveness to a variety of stresses. Cloning of the Arabidopsis EIN2 gene identifies a central component of the ethylene signaling pathway. The amino-terminal integral membrane domain of EIN2 shows similarity to the disease-related Nramp family of metal-ion transporters. Expression of the EIN2 CEND is sufficient to constitutively activate ethylene responses and restores responsiveness to jasmonic acid and paraquat-induced oxygen radicals to mutant plants. EIN2 is thus recognized as a molecular link between previously distinct hormone response pathways. Plants may use a combinatorial mechanism for assessing various stresses by enlisting a common set of signaling molecules.
DOI:10.1073/pnas.1103959108URL [БОЮФв§гУ: 1]
DOI:10.1105/tpc.18.00018URL [БОЮФв§гУ: 1]
DOI:10.1105/tpc.110.076588URL [БОЮФв§гУ: 1]
PMID:17307926 [БОЮФв§гУ: 1]
Ethylene signaling in Arabidopsis thaliana converges on the ETHYLENE-INSENSITIVE3 (EIN3)/EIN3-Like (EIL) transcription factors to induce various responses. EIN3 BINDING F-BOX1 (EBF1) and EBF2 were recently shown to function in ethylene perception by regulating EIN3/EIL turnover. In the absence of ethylene, EIN3 and possibly other EIL proteins are targeted for ubiquitination and subsequent degradation by Cullin 1-based E3 complexes containing EBF1 and 2. Ethylene appears to block this ubiquitination, allowing EIN3/EIL levels to rise and mediate ethylene signaling. Through analysis of mutant combinations affecting accumulation of EBF1, EBF2, EIN3, and EIL1, we show that EIN3 and EIL1 are the main targets of EBF1/2. Kinetic analyses of hypocotyl growth inhibition in response to ethylene and growth recovery after removal of the hormone revealed that EBF1 and 2 have temporally distinct but overlapping roles in modulating ethylene perception. Whereas EBF1 plays the main role in air and during the initial phase of signaling, EBF2 plays a more prominent role during the latter stages of the response and the resumption of growth following ethylene removal. Through their coordinated control of EIN3/EIL1 levels, EBF1 and EBF2 fine-tune ethylene responses by repressing signaling in the absence of the hormone, dampening signaling at high hormone concentrations, and promoting a more rapid recovery after ethylene levels dissipate.
DOI:10.1038/nchembio.1178PMID:23377040 [БОЮФв§гУ: 1]
We identify an Arabidopsis pyridoxal-phosphate-dependent aminotransferase, VAS1, whose loss-of-function simultaneously increases amounts of the phytohormone auxin and the ethylene precursor 1-aminocyclopropane-1-carboxylate. VAS1 uses the ethylene biosynthetic intermediate methionine as an amino donor and the auxin biosynthetic intermediate indole-3-pyruvic acid as an amino acceptor to produce L-tryptophan and 2-oxo-4-methylthiobutyric acid. Our data indicate that VAS1 serves key roles in coordinating the amounts of these two vital hormones.
DOI:10.1016/j.cell.2008.01.047PMID:18394997 [БОЮФв§гУ: 5]
Plants have evolved a tremendous ability to respond to environmental changes by adapting their growth and development. The interaction between hormonal and developmental signals is a critical mechanism in the generation of this enormous plasticity. A good example is the response to the hormone ethylene that depends on tissue type, developmental stage, and environmental conditions. By characterizing the Arabidopsis wei8 mutant, we have found that a small family of genes mediates tissue-specific responses to ethylene. Biochemical studies revealed that WEI8 encodes a long-anticipated tryptophan aminotransferase, TAA1, in the essential, yet genetically uncharacterized, indole-3-pyruvic acid (IPA) branch of the auxin biosynthetic pathway. Analysis of TAA1 and its paralogues revealed a link between local auxin production, tissue-specific ethylene effects, and organ development. Thus, the IPA route of auxin production is key to generating robust auxin gradients in response to environmental and developmental cues.
DOI:10.1038/cr.2012.29URL [БОЮФв§гУ: 6]
DOI:10.1016/j.devcel.2004.07.002URL [БОЮФв§гУ: 4]
DOI:10.1016/j.cub.2015.12.003URL [БОЮФв§гУ: 2]
PMID:14756759 [БОЮФв§гУ: 4]
Dark-grown Arabidopsis seedlings develop an apical hook by differential elongation and division of hypocotyl cells. This allows the curved hypocotyl to gently drag the apex, which is protected by the cotyledons, upwards through the soil. Several plant hormones are known to be involved in hook development, including ethylene, which causes exaggeration of the hook. We show that gibberellins (GAs) are also involved in this process. Inhibition of GA biosynthesis with paclobutrazol (PAC) prevented hook formation in wild-type (WT) seedlings and in constitutive ethylene response (ctr)1-1, a mutant that exhibits a constitutive ethylene response. In addition, a GA-deficient mutant (ga1-3) did not form an apical hook in the presence of the ethylene precursor 1-aminocyclopropane-1-carboxylate (ACC). Analysis of transgenic Arabidopsis seedlings expressing a green fluorescent protein (GFP)-repressor of ga1-3 (RGA) fusion protein suggested that ACC inhibits cell elongation in the apical hook by inhibition of GA signaling. A decreased feedback of GA possibly causes an induction of GA biosynthesis based upon the expression of genes encoding copalyl diphosphate synthase (CPS; GA1) and GA 2-oxidase (AtGA2ox1). Furthermore, expression of GASA1, a GA-response gene, suggests that differential cell elongation in the apical hook might be a result of differential GA-sensitivity.
PMID:14963246 [БОЮФв§гУ: 6]
Plants undergo two different developmental programs depending on whether they are growing in darkness (skotomorphogenesis) or in the presence of light (photomorphogenesis). It has been proposed that the latter is the default pathway followed by many plants after germination and before the seedling emerges from soil. The transition between the two pathways is tightly regulated. The conserved COP1-based complex is central in the light-dependent repression of photomorphogenesis in darkness. Besides this control, hormones such as brassinosteroids (BRs), cytokinins, auxins, or ethylene also have been shown to regulate, to different extents, this developmental switch. In the present work, we show that the hormone gibberellin (GA) widely participates in this regulation. Studies from Arabidopsis show that both chemical and genetic reductions of endogenous GA levels partially derepress photomorphogenesis in darkness. This is based both on morphological phenotypes, such as hypocotyl elongation and hook and cotyledon opening, and on molecular phenotypes, such as misregulation of the light-controlled genes CAB2 and RbcS. Genetic studies indicate that the GA signaling elements GAI and RGA participate in these responses. Our results also suggest that GA regulation of this response partially depends on BRs. This regulation seems to be conserved across species because lowering endogenous GA levels in pea (Pisum sativum) induces full de-etiolation in darkness, which is not reverted by BR application. Our results, therefore, attribute an important role for GAs in the establishment of etiolated growth and in repression of photomorphogenesis.
DOI:10.1111/tpj.2011.67.issue-4URL [БОЮФв§гУ: 5]
DOI:10.1242/dev.081240PMID:22992959 [БОЮФв§гУ: 3]
When penetrating the soil during germination, dicotyledonous plants protect their shoot apical meristem through the formation of an apical hook. Apical hook formation is a dynamic process that can be subdivided into hook formation, maintenance and opening. It has previously been established that these processes require the transport and signaling of the phytohormone auxin, as well as the biosynthesis and signaling of the phytohormones ethylene and gibberellin (GA). Here, we identify a molecular mechanism for an auxin-GA crosstalk by demonstrating that the auxin transport-regulatory protein kinase WAG2 is a crucial transcription target during apical hook opening downstream from GA signaling. We further show that WAG2 is directly activated by PHYTOCHROME INTERACTING FACTOR 5 (PIF5), a light-labile interactor of the DELLA repressors of the GA pathway. We find that wag2 mutants are impaired in the repression of apical hook opening in dark-grown seedlings and that this phenotype correlates with GA-regulated WAG2 expression in the concave (inner) side of the apical hook. Furthermore, wag2 mutants are also impaired in the maintenance or formation of a local auxin maximum at the site of WAG2 expression in the hook. WAG2 is a regulator of PIN auxin efflux facilitators and, in line with previous data, we show that this kinase can phosphorylate the central intracellular loop of all PIN-FORMED (PIN) proteins regulating apical hook opening. We therefore propose that apical hook opening is controlled by the differential GA-regulated accumulation of WAG2 and subsequent local changes in PIN-mediated auxin transport.
PMID:21852755 [БОЮФв§гУ: 2]
The analysis of cell polarity in plants is fueled by the discovery and analysis of auxin efflux carrier PIN proteins that show polar localizations in various plant cell types in line with their roles in directional cell to cell auxin transport. As this asymmetry in cellular PIN localization drives directional auxin fluxes, abnormalities in PIN localizations modify auxin transport culminating into range of auxin distribution defective phenotypes. Because of this influence of PIN localization on plant development via changes in auxin distribution, mechanisms establishing, maintaining and altering PIN polarity are of intense interest in the plant field during the recent years. Recent findings suggest that two categories of molecules, namely AGC-3 kinase family members PINOID, WAG1, WAG2 and ARF-GEF family member GNOM predominantly influence the polar localization of PINs. The emerging mechanism for AGC-3 kinases and ARF-GEF action suggest that AGC-3 kinases predominantly phosphorylate PINs at the plasma membrane for eventual PIN internalization and PIN sorting into ARF-GEF GNOM independent polar recycling pathways. In case of mutant for AGC-3 kinases or mutations in AGC-3 kinase-targeted PIN residues, much less phosphorylated PINs are recruited into ARFGEF GNOM-dependent polar recycling pathway. When ARF-GEF GNOM is inactive, the bias is shifted for rerouting less efficiently phosphorylated PINs into GNOM-independent polar recycling pathways that generally prefer efficiently phosphorylated PINs. Thus, balance shifts between the extent of AGC-3 kinase mediated PIN phosphorylation and the functioning of ARFGEF instruct PIN polarity establishment and/or PIN polarity alterations. Recent studies report utilization of this AGC-3 kinase and ARF-GEF PIN polarity regulation module during diverse developmental and response programs including shoot patterning, root growth, phototropism, gravitropism, organogenesis, leaf epidermal cell indentations and fruit valve margin formation. Based on these findings the same theme of phosphorylated PIN sorting into differential polar recycling pathways for PIN polarity establishment and alteration seems to be employed in a context-dependent manner.
DOI:10.1242/dev.052456PMID:20823065 [БОЮФв§гУ: 2]
Polar membrane cargo delivery is crucial for establishing cell polarity and for directional transport processes. In plants, polar trafficking mediates the dynamic asymmetric distribution of PIN FORMED (PIN) carriers, which drive polar cell-to-cell transport of the hormone auxin, thereby generating auxin maxima and minima that control development. The Arabidopsis PINOID (PID) protein kinase instructs apical PIN localization by phosphorylating PINs. Here, we identified the PID homologs WAG1 and WAG2 as new PIN polarity regulators. We show that the AGC3 kinases PID, WAG1 and WAG2, and not other plant AGC kinases, instruct recruitment of PINs into the apical recycling pathway by phosphorylating the middle serine in three conserved TPRXS(N/S) motifs within the PIN central hydrophilic loop. Our results put forward a model by which apolarly localized PID, WAG1 and WAG2 phosphorylate PINs at the plasma membrane after default non-polar PIN secretion, and trigger endocytosis-dependent apical PIN recycling. This phosphorylation-triggered apical PIN recycling competes with ARF-GEF GNOM-dependent basal recycling to promote apical PIN localization. In planta, expression domains of PID, WAG1 and WAG2 correlate with apical localization of PINs in those cell types, indicating the importance of these kinases for apical PIN localization. Our data show that by directing polar PIN localization and PIN-mediated polar auxin transport, the three AGC3 kinases redundantly regulate cotyledon development, root meristem size and gravitropic response, indicating their involvement in both programmed and adaptive plant development.
DOI:10.1105/tpc.107.051508URL [БОЮФв§гУ: 3]
DOI:10.1105/tpc.113.120394URL [БОЮФв§гУ: 5]
DOI:10.1105/tpc.113.122002URL [БОЮФв§гУ: 5]
DOI:10.1105/tpc.18.00018URL [БОЮФв§гУ: 2]
PMID:10972893 [БОЮФв§гУ: 1]
Leaf senescence is a complex process that is controlled by multiple developmental and environmental signals and is manifested by induced expression of a large number of different genes. In this paper we describe experiments that show, for the first time, that the salicylic acid (SA)-signalling pathway has a role in the control of gene expression during developmental senescence. Arabidopsis plants defective in the SA-signalling pathway (npr1 and pad4 mutants and NahG transgenic plants) were used to investigate senescence-enhanced gene expression, and a number of genes showed altered expression patterns. Senescence-induced expression of the cysteine protease gene SAG12, for example, was conditional on the presence of SA, together with another unidentified senescence-specific factor. Changes in gene expression patterns were accompanied by a delayed yellowing and reduced necrosis in the mutant plants defective in SA-signalling, suggesting a role for SA in the cell death that occurs at the final stage of senescence. We propose the presence of a minimum of three senescence-enhanced signalling factors in senescing leaves, one of which is SA. We also suggest that a combination of signalling factors is required for the optimum expression of many genes during senescence.
PMID:17746926 [БОЮФв§гУ: 1]
In an effort to identify the signal compound that mediates systemic acquired resistance (SAR), changes in the content of phloem sap were monitored in cucumber plants inoculated with either tobacco necrosis virus or the fungal pathogen Colletotrichum lagenarium. The concentration of a fluorescent metabolite was observed to increase transiently after inoculation, with a peak reached before SAR was detected. The compound was purified and identified by gas chromatography-mass spectrometry as salicylic acid, a known exogenous inducer of resistance. The data suggest that salicylic acid could function as the endogenous signal in the transmission of SAR in cucumber.
DOI:10.1105/tpc.17.00438URL [БОЮФв§гУ: 1]
DOI:10.1105/tpc.19.00658URL [БОЮФв§гУ: 2]
DOI:10.2307/3869351URL [БОЮФв§гУ: 1]
PMID:8612270 [БОЮФв§гУ: 1]
The cpd mutation localized by T-DNA tagging on Arabidopsis chromosome 5-14.3 inhibits cell elongation controlled by the ecdysone-like brassinosteroid hormone brassinolide. The cpd mutant displays de-etiolation and derepression of light-induced genes in the dark, as well as dwarfism, male sterility, and activation of stress-regulated genes in the light. The CPD gene encodes a cytochrome P450 (CYP90) sharing homologous domains with steroid hydroxylases. The phenotype of the cpd mutant is restored to wild type both by feeding with C23-hydroxylated brassinolide precursors and by ectopic overexpression of the CPD cDNA. Brassinosteroids also compensate for different cell elongation defects of Arabidopsis det, cop, fus, and axr2 mutants, indicating that these steroids play an essential role in the regulation of plant development.
DOI:10.1093/pcp/pci111URL [БОЮФв§гУ: 3]
DOI:10.1093/mp/ssn005PMID:19825546 [БОЮФв§гУ: 1]
We undertook a chemical genetics screen to identify chemical inhibitors of brassinosteroid (BR) action. From a chemical library of 10,000 small molecules, one compound was found to inhibit hypocotyl length and activate the expression of a BR-repressed reporter gene (CPD::GUS) in Arabidopsis, and it was named brassinopride (BRP). These effects of BRP could be reversed by co-treatment with brassinolide, suggesting that BRP either directly or indirectly inhibits BR biosynthesis. Interestingly, the compound causes exaggerated apical hooks, similar to that caused by ethylene treatment. The BRP-induced apical hook phenotype can be blocked by a chemical inhibitor of ethylene perception or an ethylene-insensitive mutant, suggesting that, in addition to inhibiting BR, BRP activates ethylene response. Analysis of BRP analogs provided clues about structural features important for its effects on two separate targets in the BR and ethylene pathways. Analyses of the responses of various BR and ethylene mutants to BRP, ethylene, and BR treatments revealed modes of cross-talk between ethylene and BR in dark-grown seedlings. Our results suggest that active downstream BR signaling, but not BR synthesis or a BR gradient, is required for ethylene-induced apical hook formation. The BRP-related compounds can be useful tools for manipulating plant growth and studying hormone interactions.
DOI:10.1093/jxb/erx463PMID:29309681 [БОЮФв§гУ: 1]
PMID:16096973 [БОЮФв§гУ: 1]
In an attempt to compensate for their sessile nature, plants have developed growth responses to deal with the copious and rapid changes in their environment. These responses are known as tropisms and they are marked by a directional growth response that is the result of differential cellular growth and development in response to an external stimulation such as light, gravity or touch. While the mechanics of tropic growth and subsequent development have been the topic of debate for more than a hundred years, only recently have researchers been able to make strides in understanding how plants perceive and respond to tropic stimulations, thanks in large part to mutant analysis and recent advances in genomics. This paper focuses on the recent advances in four of the best-understood tropic responses and how each affects plant growth and development: phototropism, gravitropism, thigmotropism and hydrotropism. While progress has been made in deciphering the events between tropic stimulation signal perception and each characteristic growth response, there are many areas that remain unclear, some of which will be discussed herein. As has become evident, each tropic response pathway exhibits distinguishing characteristics. However, these pathways of tropic perception and response also have overlapping components - a fact that is certainly related to the necessity for pathway integration given the ever-changing environment that surrounds every plant.
DOI:10.1016/j.cub.2017.05.085URL [БОЮФв§гУ: 1]
DOI:10.3732/ajb.1200591URL [БОЮФв§гУ: 1]
DOI:10.1016/j.devcel.2004.07.002URL [БОЮФв§гУ: 2]
DOI:10.1105/tpc.15.00569URL [БОЮФв§гУ: 6]
DOI:10.1093/jxb/ert080PMID:23580748 [БОЮФв§гУ: 1]
The plant hormone auxin drives plant growth and morphogenesis. The levels and distribution of the active auxin indole-3-acetic acid (IAA) are tightly controlled through synthesis, inactivation, and transport. Many auxin precursors and modified auxin forms, used to regulate auxin homeostasis, have been identified; however, very little is known about the integration of multiple auxin biosynthesis and inactivation pathways. This review discusses the many ways auxin levels are regulated through biosynthesis, storage forms, and inactivation, and the potential roles modified auxins play in regulating the bioactive pool of auxin to affect plant growth and development.
DOI:10.1073/pnas.1108434108URL [БОЮФв§гУ: 1]
DOI:10.1093/mp/ssr104URL [БОЮФв§гУ: 2]
DOI:10.1016/j.cell.2008.01.049PMID:18394996 [БОЮФв§гУ: 2]
Plants grown at high densities perceive a decrease in the red to far-red (R:FR) ratio of incoming light, resulting from absorption of red light by canopy leaves and reflection of far-red light from neighboring plants. These changes in light quality trigger a series of responses known collectively as the shade avoidance syndrome. During shade avoidance, stems elongate at the expense of leaf and storage organ expansion, branching is inhibited, and flowering is accelerated. We identified several loci in Arabidopsis, mutations in which lead to plants defective in multiple shade avoidance responses. Here we describe TAA1, an aminotransferase, and show that TAA1 catalyzes the formation of indole-3-pyruvic acid (IPA) from L-tryptophan (L-Trp), the first step in a previously proposed, but uncharacterized, auxin biosynthetic pathway. This pathway is rapidly deployed to synthesize auxin at the high levels required to initiate the multiple changes in body plan associated with shade avoidance.
DOI:10.1105/tpc.111.231112 [БОЮФв§гУ: 3]
DOI:10.1105/tpc.114.127993URL [БОЮФв§гУ: 2]
DOI:10.1101/gad.1415106URL [БОЮФв§гУ: 2]
DOI:10.1105/tpc.107.053009URL [БОЮФв§гУ: 2]
PMID:11209081 [БОЮФв§гУ: 1]
Although auxin is known to regulate many processes in plant development and has been studied for over a century, the mechanisms whereby plants produce it have remained elusive. Here we report the characterization of a dominant Arabidopsis mutant, yucca, which contains elevated levels of free auxin. YUCCA encodes a flavin monooxygenase-like enzyme and belongs to a family that includes at least nine other homologous Arabidopsis genes, a subset of which appears to have redundant functions. Results from tryptophan analog feeding experiments and biochemical assays indicate that YUCCA catalyzes hydroxylation of the amino group of tryptamine, a rate-limiting step in tryptophan-dependent auxin biosynthesis.
DOI:10.1105/tpc.111.088047URL [БОЮФв§гУ: 2]
DOI:10.1186/gb-2009-10-12-249PMID:20053306 [БОЮФв§гУ: 4]
The PIN-FORMED (PIN) proteins are secondary transporters acting in the efflux of the plant signal molecule auxin from cells. They are asymmetrically localized within cells and their polarity determines the directionality of intercellular auxin flow. PIN genes are found exclusively in the genomes of multicellular plants and play an important role in regulating asymmetric auxin distribution in multiple developmental processes, including embryogenesis, organogenesis, tissue differentiation and tropic responses. All PIN proteins have a similar structure with amino- and carboxy-terminal hydrophobic, membrane-spanning domains separated by a central hydrophilic domain. The structure of the hydrophobic domains is well conserved. The hydrophilic domain is more divergent and it determines eight groups within the protein family. The activity of PIN proteins is regulated at multiple levels, including transcription, protein stability, subcellular localization and transport activity. Different endogenous and environmental signals can modulate PIN activity and thus modulate auxin-distribution-dependent development. A large group of PIN proteins, including the most ancient members known from mosses, localize to the endoplasmic reticulum and they regulate the subcellular compartmentalization of auxin and thus auxin metabolism. Further work is needed to establish the physiological importance of this unexpected mode of auxin homeostasis regulation. Furthermore, the evolution of PIN-based transport, PIN protein structure and more detailed biochemical characterization of the transport function are important topics for further studies.
DOI:10.1242/dev.021071PMID:18787070 [БОЮФв§гУ: 1]
The signalling molecule auxin controls plant morphogenesis via its activity gradients, which are produced by intercellular auxin transport. Cellular auxin efflux is the rate-limiting step in this process and depends on PIN and phosphoglycoprotein (PGP) auxin transporters. Mutual roles for these proteins in auxin transport are unclear, as is the significance of their interactions for plant development. Here, we have analysed the importance of the functional interaction between PIN- and PGP-dependent auxin transport in development. We show by analysis of inducible overexpression lines that PINs and PGPs define distinct auxin transport mechanisms: both mediate auxin efflux but they play diverse developmental roles. Components of both systems are expressed during embryogenesis, organogenesis and tropisms, and they interact genetically in both synergistic and antagonistic fashions. A concerted action of PIN- and PGP-dependent efflux systems is required for asymmetric auxin distribution during these processes. We propose a model in which PGP-mediated efflux controls auxin levels in auxin channel-forming cells and, thus, auxin availability for PIN-dependent vectorial auxin movement.
DOI:10.1105/tpc.112.097766URL [БОЮФв§гУ: 4]
DOI:10.3390/ijms19092759URL [БОЮФв§гУ: 3]
DOI:10.1111/tpj.2009.59.issue-1URL [БОЮФв§гУ: 1]
PMID:11701880 [БОЮФв§гУ: 1]
Arabidopsis possesses several genes related to the multidrug resistance (MDR) genes of animals, one of which, AtMDR1, was shown to be induced by the hormone auxin. Plants having mutations in AtMDR1 or its closest relative, AtPGP1, were isolated by a reverse genetic strategy. Auxin transport activity was greatly impaired in atmdr1 and atmdr1 atpgp1 double mutant plants. Epinastic cotyledons and reduced apical dominance were mutant phenotypes consistent with the disrupted basipetal flow of auxin. The auxin transport inhibitor 1-naphthylphthalamic acid was shown to bind tightly and specifically to AtMDR1 and AtPGP1 proteins. The results indicate that these two MDR-like genes of Arabidopsis encode 1-naphthylphthalamic acid binding proteins that are required for normal auxin distribution and auxin-mediated development.
DOI:10.1111/j.1365-313X.2010.04137.xURL [БОЮФв§гУ: 2]
DOI:10.1038/415806aURL [БОЮФв§гУ: 1]
PMID:16192309 [БОЮФв§гУ: 1]
Plant development displays an exceptional plasticity and adaptability that involves the dynamic, asymmetric distribution of the phytohormone auxin. Polar auxin flow, which requires polarly localized transport facilitators of the PIN family, largely contributes to the establishment and maintenance of the auxin gradients. Functionally overlapping action of PIN proteins mediates multiple developmental processes, including embryo formation, organ development and tropisms. Here we show that PIN proteins exhibit synergistic interactions, which involve cross-regulation of PIN gene expression in pin mutants or plants with inhibited auxin transport. Auxin itself positively feeds back on PIN gene expression in a tissue-specific manner through an AUX/IAA-dependent signalling pathway. This regulatory switch is indicative of a mechanism by which the loss of a specific PIN protein is compensated for by auxin-dependent ectopic expression of its homologues. The compensatory properties of the PIN-dependent transport network might enable the stabilization of auxin gradients and potentially contribute to the robustness of plant adaptive development.
DOI:10.1146/annurev-genet-102108-134148URL [БОЮФв§гУ: 1]
DOI:10.1038/nature03543URL [БОЮФв§гУ: 3]
PMID:15992545 [БОЮФв§гУ: 2]
The plant hormone auxin has been implicated in virtually every aspect of plant growth and development. Auxin acts by promoting the degradation of transcriptional regulators called Aux/IAA proteins. Aux/IAA degradation requires TIR1, an F box protein that has been shown to function as an auxin receptor. However, loss of TIR1 has a modest effect on auxin response and plant development. Here we show that three additional F box proteins, called AFB1, 2, and 3, also regulate auxin response. Like TIR1, these proteins interact with the Aux/IAA proteins in an auxin-dependent manner. Plants that are deficient in all four proteins are auxin insensitive and exhibit a severe embryonic phenotype similar to the mp/arf5 and bdl/iaa12 mutants. Correspondingly, all TIR1/AFB proteins interact with BDL, and BDL is stabilized in triple mutant plants. Our results indicate that TIR1 and the AFB proteins collectively mediate auxin responses throughout plant development.
DOI:10.1038/nature03542URL [БОЮФв§гУ: 3]
DOI:10.1038/nchembio.926PMID:22466420 [БОЮФв§гУ: 2]
The plant hormone auxin regulates virtually every aspect of plant growth and development. Auxin acts by binding the F-box protein transport inhibitor response 1 (TIR1) and promotes the degradation of the AUXIN/INDOLE-3-ACETIC ACID (Aux/IAA) transcriptional repressors. Here we show that efficient auxin binding requires assembly of an auxin co-receptor complex consisting of TIR1 and an Aux/IAA protein. Heterologous experiments in yeast and quantitative IAA binding assays using purified proteins showed that different combinations of TIR1 and Aux/IAA proteins form co-receptor complexes with a wide range of auxin-binding affinities. Auxin affinity seems to be largely determined by the Aux/IAA. As there are 6 TIR1/AUXIN SIGNALING F-BOX proteins (AFBs) and 29 Aux/IAA proteins in Arabidopsis thaliana, combinatorial interactions may result in many co-receptors with distinct auxin-sensing properties. We also demonstrate that the AFB5-Aux/IAA co-receptor selectively binds the auxinic herbicide picloram. This co-receptor system broadens the effective concentration range of the hormone and may contribute to the complexity of auxin response.
DOI:10.1111/tpj.2009.59.issue-1URL [БОЮФв§гУ: 2]
DOI:10.1038/nature05731URL [БОЮФв§гУ: 2]
DOI:10.1038/nplants.2017.105PMID:28714973 [БОЮФв§гУ: 1]
The phytohormone auxin induces or represses growth depending on its concentration and the underlying tissue type. However, it remains unknown how auxin signalling is modulated to allow tissues transiting between repression and promotion of growth. Here, we used apical hook development as a model for growth transitions in plants. A PIN-FORMED (PIN)-dependent intercellular auxin transport module defines an auxin maximum that is causal for growth repression during the formation of the apical hook. Our data illustrate that growth transition for apical hook opening is largely independent of this PIN module, but requires the PIN-LIKES (PILS) putative auxin carriers at the endoplasmic reticulum. PILS proteins reduce nuclear auxin signalling in the apical hook, leading to the de-repression of growth and the onset of hook opening. We also show that the phytochrome (phy) B-reliant light-signalling pathway directly regulates PILS gene activity, thereby enabling light perception to repress nuclear auxin signalling and to control growth. We propose a novel mechanism, in which PILS proteins allow external signals to alter tissue sensitivity to auxin, defining differential growth rates.
DOI:10.1093/pcp/pci111URL [БОЮФв§гУ: 1]
PMID:8624510 [БОЮФв§гУ: 1]
By screening suppressor mutants of the hy2 mutation of Arabidopsis thaliana, two dominant photomorphogenic mutants, shy1-1D and shy2-1D, for two genetic loci designated as SHY1 and SHY2 (suppressor of hy2 mutation) have been isolated. Both of these non-allelic, extragenic suppressor mutations of hy2 are located on chromosome 1 of the Arabidopsis genome. Both mutations suppress the elongated hypocotyl phenotype of hy2 by light-independent inhibition of hypocotyl growth as well as by increasing the effectiveness of light inhibition of hypocotyl elongation. The shy1-1D mutation is partially photomorphogenic in darkness with apical hook opening and reduced hypocotyl elongation. The shy2-1D mutant displays highly photomorphogenic characteristics in darkness such as true leaf development, cotyledon expansion and extremely reduced hypocotyl growth. In regard to hypocotyl elongation, however, the shy2-1D mutation is still light sensitive. Examination of red-far-red light responses shows that the shy1-1D mutation suppresses the hypocotyl elongation of the hy2 mutation effectively in red light but not effectively in far-red light. The shy2-1D suppresses hypocotyl elongation of the hy2 mutation effectively in both red and far-red light. Both mutations can also suppress the early-flowering phenotype of hy2 and have a distinct pleiotropic effect on leaf development such as upward leaf rolling. The data obtained suggest that SHY1 and SHY2 represent a novel class of components involved in the photomorphogenic pathways of Arabidopsis. This is the first report on the identification of dominant mutations in the light signal transduction pathway of plants.
PMID:9744095 [БОЮФв§гУ: 1]
We previously reported a photomorphogenic mutation of Arabidopsis thaliana, shy2-1D, as a dominant suppressor of a hy2 mutation. Here, we report that shy2-1D confers various photo-responsive phenotypes in darkness and the dark phenotypes of the mutant are affected by phytochrome deficiency. Dark-grown seedlings of the mutant developed several photomorphogenic characteristics such as short hypocotyls, cotyledon expansion and opening, and partial differentiation of plastids. When grown further in darkness, the mutant plant underwent most of the developmental stages of a light-grown wild-type plant, including development of foliar leaves, an inflorescence stem with cauline leaves, and floral organs. In addition, two light-inducible genes, the nuclear-encoded CAB and the plastid-encoded PSBA genes, were highly expressed in the dark-grown mutant seedlings. Furthermore, reduced gravitropism, a phytochrome-modulated response, was observed in the mutant hypocotyl in darkness. Thus, shy2-1D is one of the most pleiotropic photomorphogenic mutations identified so far. The results indicate that SHY2 may be a key component regulating photomorphogenesis in Arabidopsis. Surprisingly, double mutants of the shy2-1D mutant with the phytochrome-deficient mutants hy2, hy3(phyB-1) and fre1-1(phyA-201) showed reduced photomorphogenic response in darkness with a longer hypocotyl, a longer inflorescence stem, and a lower level expression of the CAB gene than the shy2-1D single mutant. These results showed that phytochromes function in darkness in the shy2-1D mutant background. The implications of these results are discussed.
DOI:10.1104/pp.105.070987URL [БОЮФв§гУ: 1]
PMID:15960614 [БОЮФв§гУ: 3]
AUXIN RESPONSE FACTORS (ARFs) regulate auxin-mediated transcriptional activation/repression. They are encoded by a gene family in Arabidopsis, and each member is thought to play a central role in various auxin-mediated developmental processes. We have characterized three arf2 mutant alleles, arf2-6, arf2-7 and arf2-8. The mutants exhibit pleiotropic developmental phenotypes, including large, dark green rosette leaves, delayed flowering, thick and long inflorescence, abnormal flower morphology and sterility in early formed flowers, large organ size and delayed senescence and abscission, compared with wild-type plants. In addition, arf2 mutant seedlings have elongated hypocotyls with enlarged cotyledons under various light conditions. The transcription of ACS2, ACS6 and ACS8 genes is impaired in the developing siliques of arf2-6. The phenotypes of all three alleles are similar to those of the loss-of-function mutants obtained by RNA interference or co-suppression. There is no significant effect of the mutation on global auxin-regulated gene expression in young seedlings, suggesting that ARF2 does not participate in auxin signaling at that particular developmental stage of the plant life cycle. Because ARF2 is thought to function as a transcriptional repressor, the prospect arises that its pleiotropic effects may be mediated by negatively modulating the transcription of downstream genes in signaling pathways that are involved in cell growth and senescence.
PMID:15659631 [БОЮФв§гУ: 2]
The AUXIN RESPONSE FACTOR (ARF) gene family products, together with the AUXIN/INDOLE-3-ACETIC ACID proteins, regulate auxin-mediated transcriptional activation/repression. The biological function(s) of most ARFs is poorly understood. Here, we report the identification and characterization of T-DNA insertion lines for 18 of the 23 ARF gene family members in Arabidopsis thaliana. Most of the lines fail to show an obvious growth phenotype except of the previously identified arf2/hss, arf3/ett, arf5/mp, and arf7/nph4 mutants, suggesting that there are functional redundancies among the ARF proteins. Subsequently, we generated double mutants. arf7 arf19 has a strong auxin-related phenotype not observed in the arf7 and arf19 single mutants, including severely impaired lateral root formation and abnormal gravitropism in both hypocotyl and root. Global gene expression analysis revealed that auxin-induced gene expression is severely impaired in the arf7 single and arf7 arf19 double mutants. For example, the expression of several genes, such as those encoding members of LATERAL ORGAN BOUNDARIES domain proteins and AUXIN-REGULATED GENE INVOLVED IN ORGAN SIZE, are disrupted in the double mutant. The data suggest that the ARF7 and ARF19 proteins play essential roles in auxin-mediated plant development by regulating both unique and partially overlapping sets of target genes. These observations provide molecular insight into the unique and overlapping functions of ARF gene family members in Arabidopsis.
PMID:14729917 [БОЮФв§гУ: 1]
We have isolated a dominant, auxin-insensitive mutant of Arabidopsis thaliana, massugu2 (msg2), that displays neither hypocotyl gravitropism nor phototropism, fails to maintain an apical hook as an etiolated seedling, and is defective in lateral root formation. Yet other aspects of growth and development of msg2 plants are almost normal. These characteristics of msg2 are similar to those of another auxin-insensitive mutant, non-phototropic hypocotyl4 (nph4), which is a loss-of-function mutant of AUXIN RESPONSE FACTOR7 (ARF7) (Harper et al., 2000). Map-based cloning of the MSG2 locus reveals that all four mutant alleles result in amino acid substitutions in the conserved domain II of an Auxin/Indole-3-Acetic Acid protein, IAA19. Interestingly, auxin inducibility of MSG2/IAA19 gene expression is reduced by 65% in nph4/arf7. Moreover, MSG2/IAA19 protein binds to the C-terminal domain of NPH4/ARF7 in a Saccharomyces cerevisiae (yeast) two-hybrid assay and to the whole latter protein in vitro by pull-down assay. These results suggest that MSG2/IAA19 and NPH4/ARF7 may constitute a negative feedback loop to regulate differential growth responses of hypocotyls and lateral root formation.
PMID:17900969 [БОЮФв§гУ: 1]
Auxin signaling is key to many plant growth and developmental processes from embryogenesis to senescence. Most, if not all, of these processes are initiated and/or mediated through auxin-regulated gene expression. Two types of transcription factor families are required for controlling expression of auxin response genes. One of these, the auxin response factor (ARF) family, functions by binding to auxin response elements (AuxREs) on promoters of auxin response genes, activating or repressing the auxin response genes, and recruiting a second family of transcription factors, the Aux/IAA repressors, that confer an auxin response to the genes. Recent advances have provided information on regulation of ARF gene expression, ARF roles in growth and developmental processes, and target genes regulated by ARFs.
DOI:10.1038/s41586-019-1069-7URL [БОЮФв§гУ: 4]
DOI:10.1093/aob/mcl027URL [БОЮФв§гУ: 1]
DOI:10.1105/tpc.15.00605URL [БОЮФв§гУ: 1]
DOI:10.3389/fpls.2018.00838URL [БОЮФв§гУ: 1]
DOI:10.3389/fpls.2021.636098PMID:33767720 [БОЮФв§гУ: 1]
During seedling etiolation after germination in the dark, seedlings have closed cotyledons and form an apical hook to protect the meristem as they break through the soil to reach the surface. Once in contact with light, the hook opens and cotyledons are oriented upward and separate. Hook development in the dark after seedling emergence from the seed follows three distinctly timed and sequential phases: formation, maintenance, and eventual opening. We previously identified () as a phytochrome interacting factor (PIF)-repressed gene in the dark necessary for hook development during etiolated growth. encodes the type 2C phosphatase PP2C.D1, and mutants exhibit open hooks in the dark. Recent evidence has described that PP2C.D1 and other PP2C.D members negatively regulate SMALL AUXIN UP RNA (SAUR)-mediated cell elongation. However, the fundamental question of the timing of PP2C.D1 action (and possibly other members of the PP2C.D family) during hook development remains to be addressed. Here, we show that PP2C.D1 is required immediately after germination to form the hook. shows reduced cell expansion in the outer layer of the hook and, therefore, does not establish the differential cell growth necessary for hook formation, indicating that PP2C.D1 is necessary to promote cell elongation during this early stage. Additionally, genetic analyses of single and high order mutants in PP2C.D1, PP2C.D2, and PP2C.D5 demonstrate that the three PP2C.Ds act collectively and sequentially during etiolation: whereas PP2C.D1 dominates hook formation, PP2C.D2 is necessary during the maintenance phase, and PP2C.D5 acts to prevent opening during the third phase together with PP2C.D1 and PP2C.D2. Finally, we uncover a possible connection of PP2C.D1 levels with ethylene physiology, which could help optimize hook formation during post-germinative growth in the dark.Copyright © 2021 Rovira, Sentandreu, Nagatani, Leivar and Monte.
DOI:10.1016/j.devcel.2020.12.008URL [БОЮФв§гУ: 1]
DOI:10.1371/journal.pgen.1006639URL [БОЮФв§гУ: 1]

{kind=link}
{kind=link}