Progress on biosynthesis and function of the natural products of Zi Cao as a traditional Chinese medicinal herb
Hongyan Lin, Xuan Wang, Cong He, Ziling Zhou, Minkai Yang, Zhongling Wen, Hongwei Han, Guihua Lu, Jinliang Qi, Yonghua Yang,State Key Laboratory of Pharmaceutical Biotechnology, Institute for Plant Molecular Biology, School of Life Sciences, Nanjing University, Nanjing 210023, China
Supported by the National Natural Science Foundation of China Nos.U1903201 Supported by the National Natural Science Foundation of China Nos.31670298 Supported by the National Natural Science Foundation of China Nos.31771413 Supported by the National Natural Science Foundation of China Nos.21702100 Supported by the National Natural Science Foundation of China Nos.21907051 the Program for Changjiang Scholars and Innovative Research Team in University from the Ministry of Education of China No.IRT_14R27
作者简介 About authors 林红燕,博士,助理研究员,研究方向:药用植物天然产物化学和分子药理。E-mail:linhy@nju.edu.cn。
Abstract Zi Cao is an important traditional medicinal plant resource in China. Shikonin and its derivatives, as the purple-red naphthoquinones among natural products of its roots, are commonly used clinically in the treatment of sores and skin inflammations. Over the past few decades, due to their highly effective multiple biological activities, pharmacological effects, good clinical efficacy and high utilization value, shikonin and its derivatives have attracted increasing attention of domestic and foreign researchers. For this reason, the wild plant germplasm resources have been suffering a grievous exploitation, leading to a serious threat to the habitat. With the development of the biosynthesis, molecular metabolism and biotechnology, as well as the continuous innovation of research methods on the biological activities and pharmacological effects of plant natural products, significant progress has been made in the research on the biosynthetic pathways and related regulatory genes of shikonin. The pharmacological action and its mechanism of shikonin have also been deeply elucidated, which greatly promoted the basic research and clinical application development of shikonin. In this review, we briefly introduce and analyze the classification of Zi Cao, structure and composition of natural shikonin and its biosynthesis pathway, functional genes related to the regulation of shikonin biosynthesis, and biological activities and pharmacological functions of shikonin. Finally, we address possible prospective for the trend on the future research and development of natural shikonin and its derivatives, hoping to provide a useful reference for the deep mining and development of medicinal natural products from important Chinese medicinal materials, and to promote the modern development of traditional Chinese medicine. Keywords:Zi Cao;shikonin;biosynthesis;gene regulation;pharmacological activity
PDF (914KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 林红燕, 王煊, 何聪, 周紫玲, 杨旻恺, 文钟灵, 韩洪苇, 陆桂华, 戚金亮, 杨永华. 中药植物紫草天然产物的生物合成及其功能研究进展. 遗传[J], 2021, 43(5): 459-472 doi:10.16288/j.yczz.20-341 Hongyan Lin. Progress on biosynthesis and function of the natural products of Zi Cao as a traditional Chinese medicinal herb. Hereditas(Beijing)[J], 2021, 43(5): 459-472 doi:10.16288/j.yczz.20-341
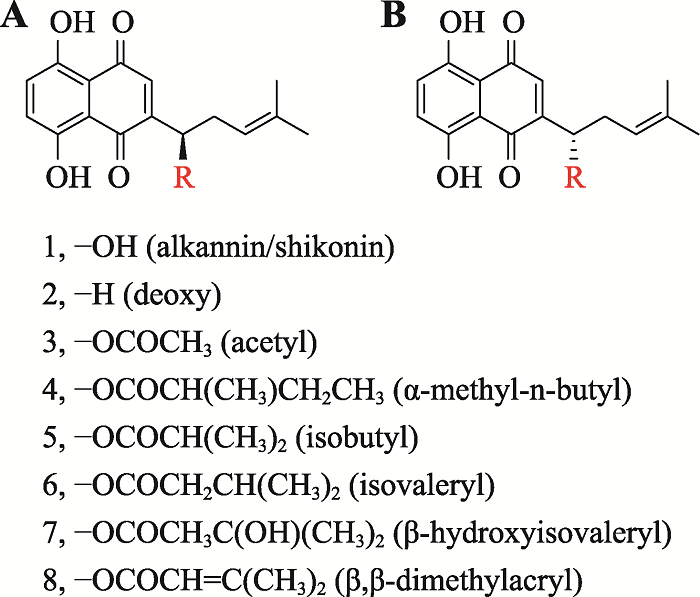
根据参考文献[9]修改绘制。A:紫草宁类;B:阿卡宁类。R为取代基团;1:紫草素;2:去氧紫草素;3:乙酰紫草素;4:ɑ-甲基-正丁酰紫草素;5:异丁酰紫草素;6:异戊酰紫草素;7:β-羟基异戊酰紫草素;8:β,βʹ-二甲基丙烯酰紫草素;此外,4与6又属于同分异构体。 Fig. 1Natural shikonins of Zi Cao
CohenJI, LittA, DavisJI . Comparative floral development in Lithospermum(Boraginaceae) and implications for the evolution and development of heterostyly Am J Bot, 2012,99(5):797-805. [本文引用: 1]
Chinese PharmacopoeiaCommission. 2000. Pharmacopoeia of the People's Republic of China (Part I). Beijing: Chemical Industry Press, 280. [本文引用: 1]
Chinese PharmacopoeiaCommission. 2020. Pharmacopoeia of the People's Republic of China (Part I). Beijing: China Medical Science Press, 355-356. [本文引用: 1]
SagratiniG, CristalliG, GiardinàD, GioventùG, MaggiF, RicciutelliM, VittoriS . Alkannin/shikonin mixture from roots of Onosma echioides(L.) L.: Extraction method study and quantification J Sep Sci, 2008,31(6-7):945-952. [本文引用: 2]
AuberRP, SuttiyutT, McCoyRM, GhasteM, CrookJW, PendletonAL, WidhalmJR, WisecaverJH,. Hybrid de novo genome assembly of red gromwell ( Lithospermum erythrorhizon) reveals evolutionary insight into shikonin biosynthesis Hortic Res, 2020,7:82. [本文引用: 2]
AssimopoulouAN, SturmS, StuppnerH, PapageorgiouVP . Preparative isolation and purification of alkannin/shikonin derivatives from natural products by high-speed counter- current chromatography Biomed Chromatogr, 2009,23(2):182-198. [本文引用: 2]
AlbrehtA, VovkI, SimonovskaB, SrbinoskaM . Identification of shikonin and its ester derivatives from the roots of Echium italicum L J Chromatogr A, 2009,1216(15):3156-3162. [本文引用: 2]
BergmanME, DavisB, PhillipsMA . Medically useful plant terpenoids: biosynthesis, occurrence, and mechanism of action Molecules, 2019,24(21):3961. [本文引用: 1]
EisenreichW, BacherA, ArigoniD, RohdichF . Biosynthesis of isoprenoids via the non-mevalonate pathway Cell Mol Life Sci, 2004,61(12):1401-1426. [本文引用: 1]
MiziorkoHM . Enzymes of the mevalonate pathway of isoprenoid biosynthesis Arch Biochem Biophys, 2011,505(2):131-143. [本文引用: 1]
GaisserS, HeideL . Inhibition and regulation of shikonin biosynthesis in suspension cultures of Lithospermum Phytochemistry, 1996,41(4):1065-1072. [本文引用: 3]
SinghRS, GaraRK, BhardwajPK, KaachraA, MalikS, KumarR, SharmaM, AhujaPS, KumarS . Expression of 3-hydroxy-3-methylglutaryl-CoA reductase, p-hydroxybenzoate-m-geranyltransferase and genes of phenylpropanoid pathway exhibits positive correlation with shikonins content in arnebia [Arnebia euchroma(Royle) Johnston] BMC Mol Biol, 2010,11:88-98. [本文引用: 4]
SzkopinskaA . Ubiquinone. Biosynthesis of quinone ring and its isoprenoid side chain. Intracellular localization Acta Biochim Pol, 2000,47(2):469-480. [本文引用: 1]
PadillaS, JonassenT, Jiménez-HidalgoMA, Fernández- AyalaDJM, López-LluchG, MarboisB, NavasP, ClarkeCF, Santos-Oca?aC . Demethoxy-Q, an intermediate of coenzyme Q biosynthesis, fails to support respiration in Saccharomyces cerevisiae and lacks antioxidant activity J Biol Chem, 2004,279(25):25995-26004. [本文引用: 2]
SommerS, K?hleA, YazakiK, ShimomuraK, BechtholdA, HeideL . Genetic engineering of shikonin biosynthesis hairy root cultures of Lithospermum erythrorhizon transformed with the bacterial ubiC gene Plant Mol Biol, 1999,39(4):683-693. [本文引用: 1]
K?hleA, SommerS, YazakiK, FerrerA, BoronatA, LiSM, HeideL . High level expression of chorismate pyruvate- lyase (UbiC) and HMG-CoA reductase in hairy root cultures of Lithospermum erythrorhizon Plant Cell Physiol, 2002,43(8):894-902. [本文引用: 1]
TakanashiK, NakagawaY, AburayaS, KaminadeK, AokiW, Saida-MunakataY, SugiyamaA, UedaM, YazakiK . Comparative proteomic analysis of Lithospermum erythrorhizon reveals regulation of a variety of metabolic enzymes leading to comprehensive understanding of the shikonin biosynthetic pathway Plant Cell Physiol, 2019,60(1):19-28. [本文引用: 1]
HeideL, NishiokaN, FukuiH, TabataM . Enzymatic regulation of shikonin biosynthesis in Lithospermum erythrorhizon cell cultures Phytochemistry, 1989,28(7):1873-1877. [本文引用: 1]
WangS, WangRS, LiuT, LvCG, LiangJW, KangCZ, ZhouLY, GuoJ, CuiGH, ZhangY, Werck-ReichhartD, GuoLP, HuangLQ . CYP76B74 catalyzes the 3''-hydroxylation of geranylhydroquinone in shikonin biosynthesis Plant Physiol, 2019,179(2):402-414. [本文引用: 1]
OharaK, MuroyaA, FukushimaN, YazakiK . Functional characterization of LePGT1, a membrane-bound prenyltransferase involved in the geranylation of p-hydroxybenzoic acid Biochem J, 2009,421(2):231-241. [本文引用: 1]
SongW, ZhuangYB, LiuT . Potential role of two cytochrome P450s obtained from Lithospermum erythrorhizon in catalyzing the oxidation of geranylhydroquinone during shikonin biosynthesis Phytochemistry, 2020,175:112375. [本文引用: 1]
OshikiriH, WatanabeB, YamamotoH, YazakiK, TakanashiK . Two BAHD acyltransferases catalyze the last step in the shikonin/alkannin biosynthetic pathway Plant Physiol, 2020,184(2):753-761. [本文引用: 1]
LangeBM, SeverinK, BechtholdA, HeideL . Regulatory role of microsomal 3-hydroxy-3-methylglutaryl-coenzyme A reductase for shikonin biosynthesis in Lithospermum erythrorhizon cell suspension cultures Planta, 1998,204(2):234-241. [本文引用: 1]
KatsuyamaY, MatsuzawaM, FunaN, HorinouchiS . Production of curcuminoids by Escherichia coli carrying an artificial biosynthesis pathway Microbiology, 2008,154(Pt 9):2620-2628. [本文引用: 2]
Syk?owska-BaranekK, PietrosiukA, NaliwajskiMR, KawiakA, JeziorekM, WyderskaS, LojkowskaE, ChinouI . Effect of l-phenylalanine on PAL activity and production of naphthoquinone pigments in suspension cultures ofArnebia euchroma(Royle) Johnst In Vitro Cell Dev Biol Plant, 2012,48(5):555-564. [本文引用: 1]
YazakiK, KunihisaM, FujisakiT, SatoF . Geranyl diphosphate: 4-hydroxybenzoate geranyltransferase from Lithospermum erythrorhizon. Cloning and characterization of a ket enzyme in shikonin biosynthesis J Biol Chem, 2002,277(8):6240-6246. [本文引用: 1]
YazakiK, MatsuokaH, ShimomuraK, BechtholdA, SatoF . A novel dark-inducible protein, LeDI-2, and its involvement in root-specific secondary metabolism in Lithospermum erythrorhizon Plant Physiol, 2001,125(4):1831-1841. [本文引用: 1]
PréM, AtallahM, ChampionA, De VosM, PieterseCMJ, MemelinkJ . The AP2/ERF domain transcription factor ORA59 integrates jasmonic acid and ethylene signals in plant defense Plant Physiol, 2008,147(3):1347-1357. [本文引用: 1]
ZhuZQ, AnFY, FengY, LiPP, LiX, MuA, JiangZQ, KimJM, ToTK, LiW, ZhangXY, YuQ, DongZ, ChenWQ, SekiM, ZhouJM, GuoHW . Derepression of ethylene-stabilized transcription factors (EIN3/EIL1) mediates jasmonate and ethylene signaling synergy in Arabidopsis Proc Natl Acad Sci USA, 2011,108(30):12539-12544. [本文引用: 1]
ZhangW, ZouA, MiaoJ, YinY, TianR, PangY, YangR, QiJ, YangY . LeERF-1, a novel AP2/ERF family gene within the B3 subcluster, is down-regulated by light signals in Lithospermum erythrorhizon Plant Biol, 2011,13(2):343-348. [本文引用: 2]
FangRJ, ZouAL, ZhaoH, WuFY, ZhuY, ZhaoH, LiaoYH, TangRJ, PangYJ, YangRW, WangXM, QiJL, LuGH, YangYH . Transgenic studies reveal the positive role of LeEIL-1 in regulating shikonin biosynthesis in Lithospermum erythrorhizon hairy roots BMC Plant Biol, 2016,16(1):121-132. [本文引用: 1]
SauterM, LorbieckeR, OuyangB, PochapskyTC, RzewuskiG . The immediate-early ethylene response gene OsARD1 encodes an acireductone dioxygenase involved in recycling of the ethylene precursor S-adenosylmethionine Plant J, 2005,44(5):718-729. [本文引用: 1]
QiJL, ZhangWJ, LiuSH, WangH, SunDY, XuGH, ShiMW, LiuZ, ZhangMS, ZhangHM, YangYH . Expression analysis of light-regulated genes isolated from a full-length-enriched cDNA library of Onosma paniculatum cell cultures J Plant Physiol, 2008,165(14):1474-1482. [本文引用: 1]
YazakiK, BechtholdA, TabataM . Nucleotide sequence of a cDNA from Lithospermum erythrorhizon homologous to PR-1 of parsley Plant Physiol, 1995,108(3):1331-1332. [本文引用: 1]
YamamuraY, SahinFP, NagatsuA, MizukamiH . Molecular cloning and characterization of a cDNA encoding a novel apoplastic protein preferentially expressed in a shikonin- producing callus strain of Lithospermum erythrorhizon Plant Cell Physiol, 2003,44(4):437-446. [本文引用: 1]
ZhaoH, BalochSK, KongLR, ZhangWJ, ZouAL, WangXM, QiJL, YangYH . Molecular cloning, characterization, and expression analysis of LeMYB1 from Lithospermum erythrorhizon Biol Plantarum, 2014,58(3):436-444. [本文引用: 1]
AndújarI, RecioMC, GinerRM, RíosJL . Traditional Chinese Medicine Remedy to Jury: The Pharmacological basis for the use of shikonin as an anticancer therapy Curr Med Chem, 2013,20(23):2892-2898. [本文引用: 1]
YangF, ChenY, DuanWH, ZhangC, ZhuH, DingJ . SH-7, a new synthesized shikonin derivative, exerting its potent antitumor activities as a topoisomerase inhibitor Int J Cancer, 2006,119(5):1184-1193. [本文引用: 1]
AhnBZ, BaikKU, KweonGR, LimK, HwangBD . Acylshikonin analogues: Synthesis and inhibition of DNA topoisomerase-I J Med Chem, 1995,38(6):1044-1047. [本文引用: 1]
QiuHY, ZhuX, LuoYL, LinHY, TangCY, QiJL, PangYJ, YangRW, LuGH, WangXM, YangYH . Identification of new shikonin derivatives as antitumor agents targeting STAT3 SH2 domain Sci Rep, 2017,7(1):2863-2875. [本文引用: 1]
AcharyaBR, BhattacharyyaS, ChoudhuryD, ChakrabartiG . The microtubule depolymerizing agent naphthazarin induces both apoptosis and autophagy in A549 lung cancer cells Apoptosis, 2011,16(9):924-939. [本文引用: 1]
WangXM, LinHY, KongWY, GuoJ, ShiJ, HuangSC, QiJL, YangRW, GuHW, YangYH . Synthesis and biological evaluation of heterocyclic carboxylic acyl shikonin derivatives Chem Biol Drug Des, 2014,83(3):334-343. [本文引用: 1]
BalochSK, LingLJ, QiuHY, MaL, LinHY, HuangSC, QiJL, WangXM, LuGH, YangYH . Synthesis and biological evaluation of novel shikonin ester derivatives as potential anti-cancer agents RSC Adv, 2014,4(67):35588-35596. [本文引用: 1]
LinHY, HanHW, BaiLF, QiuHY, YinDZ, QiJL, WangXM, GuHW, YangYH . Design, synthesis and biological evaluation of shikonin thio-glycoside derivatives: new anti-tubulin agents RSC Adv, 2014,4(91):49796-49805. [本文引用: 1]
SunWX, HanHW, YangMK, WenZL, WangYS, FuJY, LuYT, WangMY, BaoJX, LuGH, QiJL, WangXM, LinHY, YangYH . Design, synthesis and biological evaluation of benzoylacrylic acid shikonin ester derivatives as irreversible dual inhibitors of tubulin and EGFR Bioorg Med Chem, 2019,27(23):115153-115169. [本文引用: 1]
HuangZS, WuHQ, DuanZF, XieBF, LiuZC, FengGK, GuLQ, ChanASC, LiYM . Synthesis and cytotoxicity study of alkannin derivatives Eur J Med Chem, 2004,39(9):755-764. [本文引用: 1]
DengR, TangJ, XieBF, FengGK, HuangYH, LiuZC, ZhuXF . SYUNZ-16, a newly synthesized alkannin derivative, induces tumor cells apoptosis and suppresses tumor growth through inhibition of PKB/AKT kinase activity and blockade of AKT/FOXO signal pathway Int J Cancer, 2010,127(1):220-229. [本文引用: 1]
ZhangX, CuiJH, ZhouW, LiSS . Design, Synthesis and anticancer activity of shikonin and alkannin derivatives with different substituents on the naphthazarin scaffold Chem Res Chinese U, 2015,31(3):394-400. [本文引用: 1]
WangRB, ZhouW, MengQQ, ZhangX, DingJ, XuY, SongHL, YangK, CuiJH, LiSS . Design, synthesis, and biological evaluation of shikonin and alkannin derivatives as potential anticancer agents via a prodrug approach Chem Med Chem, 2014,9(12):2798-2808. [本文引用: 1]
HuangG, MengQQ, ZhouW, ZhangQJ, DongJY, LiSS . Design and synthesis of biotinylated dimethylation of alkannin oxime derivatives Chinese Chem Lett, 2017,28(2):453-457. [本文引用: 1]
ChangMX, WangHG, NiuJJ, SongY, ZouZH . Alkannin-induced oxidative DNA damage synergizes with PARP inhibition to cause cancer-specific cytotoxicity Front Pharmacol, 2020,11:610205-610218. [本文引用: 1]
YangY, WangJ, YangQ, WuSS, YangZG, ZhuHH, ZhengM, LiuWX, WuW, HeJL, ChenZ . Shikonin inhibits the lipopolysaccharide-induced release of HMGB1 in RAW264.7 cells via IFN and NF-κB signaling pathways Int Immunopharmacol, 2014,19(1):81-87. [本文引用: 1]
KomiY, SuzukiY, ShimamuraM, KajimotoS, NakajoS, MasudaM, ShibuyaM, ItabeH, ShimokadoK, OettgenP, NakayaK, KojimaS . Mechanism of inhibition of tumor angiogenesis by beta-hydroxyisovalerylshikonin Cancer Sci, 2009,100(2):269-277. [本文引用: 1]
WooHJ, JunDY, LeeJY, ParkHS, WooMH, ParkSJ, KimSC, YangCH, KimYH . Anti-inflammatory action of 2-carbomethoxy-2,3-epoxy-3-prenyl-1,4-naphthoquinone (CMEP-NQ) suppresses both the MyD88-dependent and TRIF-dependent pathways of TLR4 signaling in LPS-stimulated RAW264.7 cells J Ethnopharmacol, 2017,205:103-115. [本文引用: 1]
FanXH, ChengL, YanAH . Ameliorative effect of acetylshikonin on ovalbumin (OVA)-induced allergic rhinitis in mice through the inhibition of Th2 cytokine production and mast cell histamine release APMIS, 2019,127(10):688-695. [本文引用: 1]
ZengJC, ZhuBH, SuML . Autophagy is involved in acetylshikonin ameliorating non-alcoholic steatohepatitis through AMPK/mTOR pathway Biochem Biophys Res Commun, 2018,503(3):1645-1650. [本文引用: 1]
CuiLB, YanY, ZhangM, WuJF, TangXX, YangJ, LiLL, YaoK, ZouWG, JiangCH . Acetylshikonin suppresses atherogenesis by attenuating vascular inflammation in apolipoprotein E-deficient mice Int J Clin Exp Med, 2018,11(3):1882-1890. [本文引用: 1]
ZhangZL, FanHY, YangMY, ZhangZK, LiuK . Therapeutic effect of a hydroxynaphthoquinone fraction on dextran sulfate sodium-induced ulcerative colitis World J Gastroentero, 2014,20(41):15310-15318. [本文引用: 1]
AndújarI, RíosJL, GinerRM, CerdáJM, RecioMDC . Beneficial effect of shikonin on experimental colitis induced by dextran sulfate sodium in Balb/C mice Evid Based Complement Alternat Med, 2012,38:271606. [本文引用: 1]
HaghbeenK, PourmolaeiS, MareftjoMJ, MousaviA, NoghabiKA, ShiraziFH, MeshkatA . Detailed investigations on the solid cell culture and antimicrobial activities of the Iranian Arnebia euchroma J Biomed Biotechnol, 2011,165852. [本文引用: 1]
LiHM, TangYL, ZhangZH, LiuCJ, LiHZ, LiRT, XiaXS . Compounds from Arnebia euchroma and their related anti-HCV and antibacterial activities Planta Med, 2012,78(1):39-45. [本文引用: 1]
ShenCC, SyuWJ, LiSY, LinCH, LeeGH, SunCM . Antimicrobial activities of naphthazarins from Arnebia euchroma J Nat Prod, 2002,65(12):1857-1862. [本文引用: 1]
KuoHM, HsiaTC, ChuangYC, LuHF, LinSY, ChungJG . Shikonin inhibits the growth and N-acetylation of 2-aminofluorene in Helicobacter pylori from ulcer patients Anticancer Res, 2004,24(3a):1587-1592. [本文引用: 2]
ChenX, YangL, ZhangN, TurpinJA, BuckheitRW, OsterlingC, OppenheimJJ, HowardOMZ . Shikonin, a component of Chinese herbal medicine, inhibits chemokine receptor function and suppresses human immunodeficiency virus type 1 Antimicrob Agents Chemother, 2003,47(9):2810-2816. [本文引用: 1]
ZhangY, HanH, SunL, QiuH, LinH, YuL, ZhuW, QiJ, YangR, PangY, WangX, LuG, YangY . Antiviral activity of shikonin ester derivative PMM-034 against enterovirus 71 in vitro Braz J Med Biol Res, 2017,50(10):e6586. [本文引用: 2]
ZhangYH, HanHW, QiuHY, LinHY, YuLG, ZhuWZ, QiJL, YangRW, PangYJ, WangXM, LuGH, YangYH . Antiviral activity of a synthesized shikonin ester against influenza A (H1N1) virus and insights into its mechanism Biomed Pharmacother, 2017,93:636-645. [本文引用: 2]
LeeH, BaeS, KimK, KimW, ChungSI, YangY, YoonY . Shikonin inhibits adipogenesis by modulation of the WNT/beta-catenin pathway Life Sci, 2011,88(7-8):294-301. [本文引用: 1]
ShanZL, ZhongL, XiaoCL, GanLG, XuT, SongH, YangR, LiL, LiuBZ . Shikonin suppresses proliferation and induces apoptosis in human leukemia NB4 cells through modulation of MAPKs and c-Myc Mol Med Rep, 2017,16(3):3055-3060. [本文引用: 1]
LanWJ, WanSB, GuWQ, WangHY, ZhouSW . Mechanisms behind the inhibition of lung adenocarcinoma cell by shikonin Cell Biochem Biophys, 2014,70(2):1459-1467. [本文引用: 1]
KimEK, ChoiEJ . Compromised MAPK signaling in human diseases: an update Arch Toxicol, 2015,89(6):867-882. [本文引用: 1]
FuDJ, ShangXF, NiZ, ShiGG . Shikonin inhibits inflammation and chondrocyte apoptosis by regulation of the PI3K/Akt signaling pathway in a rat model of osteoarthritis Exp Ther Med, 2016,12(4):2735-2740. [本文引用: 1]
FanC, ZhangXF, UptonZ . Anti-inflammatory effects of shikonin in human periodontal ligament cells Pharm Biol, 2018,56(1):415-421. [本文引用: 1]
YanY, TanF, MiaoH, WangH, CaoYY . Effect of shikonin against Candida albicans biofilms Front Microbiol, 2019,10:1085-1095. [本文引用: 1]
ShishodiaSK, ShankarJ . Proteomic analysis revealed ROS-mediated growth inhibition of Aspergillus terreus by shikonin J Proteomics, 2020,224:103849-103860. [本文引用: 1]
LiaoZB, ZhuZY, LiL, WangL, WanH, JianYY, CaoYY . Metabonomics on Candida albicans indicate the excessive H3K56ac is involved in the antifungal activity of shikonin . Emerg Microbes Infec, 2019,8(1):1243-1253. [本文引用: 1]
LiaoZB, YanY, DongHH, ZhuZY, JiangYY, CaoYY . Endogenous nitric oxide accumulation is involved in the antifungal activity of shikonin against Candida albicans Emerg Microbes Infec, 2016,5(8):e88. [本文引用: 1]
 ,南京大学医药生物技术国家重点实验室,植物分子生物学研究所,生命科学学院,南京 210023
,南京大学医药生物技术国家重点实验室,植物分子生物学研究所,生命科学学院,南京 210023
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT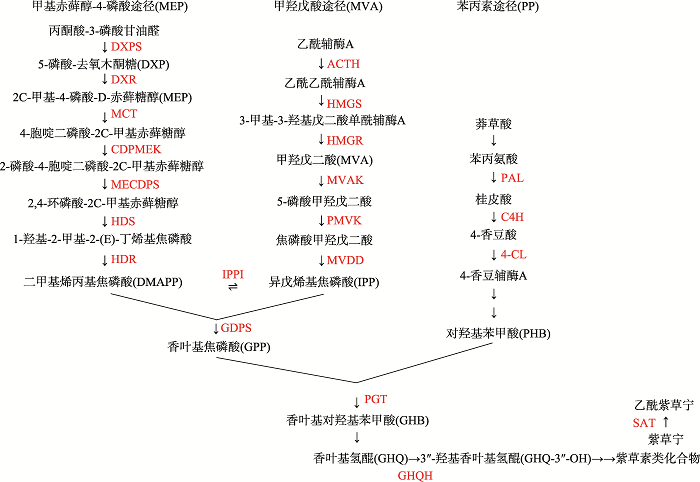 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT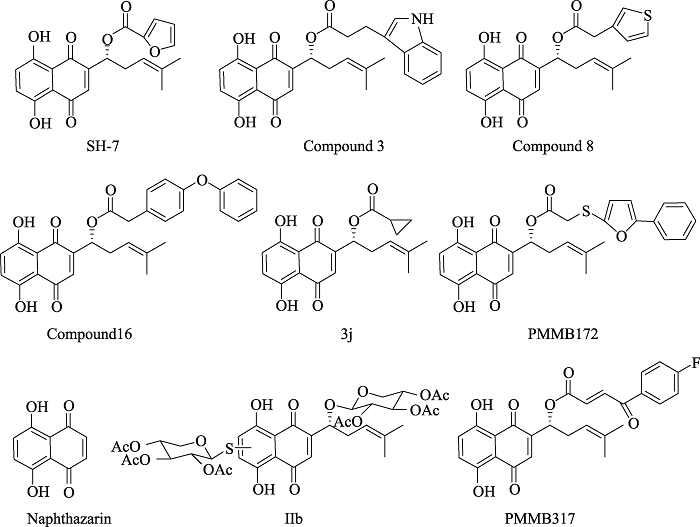 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT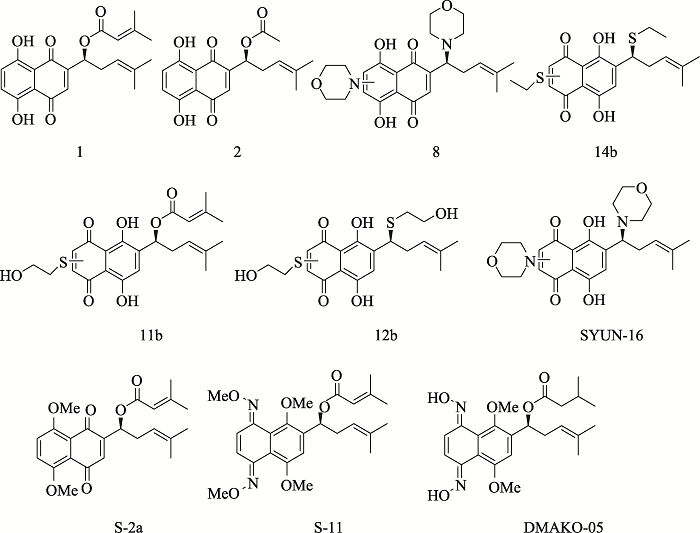 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT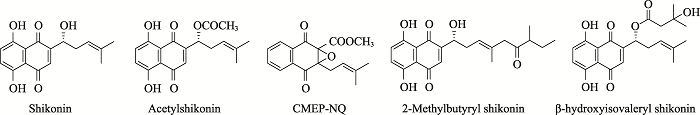 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}