 ,1, 何燕华1, 何静怡1, 王艳1, 舒鼎铭,1, 罗成龙,11
,1, 何燕华1, 何静怡1, 王艳1, 舒鼎铭,1, 罗成龙,11 Optimization of transfection conditions of chicken primordial germ cells
Zou Xian,1, He Yanhua1, He Jingyi1, Wang Yan1, Shu Dingming,1, Luo Chenglong,11 第一联系人:
罗成龙,博士,研究员,研究方向:家禽种质资源保护、遗传育种与健康生产。E-mail:
收稿日期:2020-07-7
| 基金资助: |
Received:2020-07-7
| Fund supported: |

摘要
为获得鸡原始生殖细胞(primordial germ cells, PGCs)的最佳转染效率,本研究比较不同质粒用量和不同细胞数在3种转染试剂(Lipofectamine 2000、3000和LTX & Plus Reagent)中PGCs的转染效率,利用荧光激活细胞分选技术(fluorescence activated cell sorting technology, FACS)辅助优化Lipofectamine 3000转染试剂,经FACS进一步分选获得带绿色荧光蛋白(GFP)的PGCs,继续培养3周后,移植回注到受体鸡胚中,移植3.5 d后分离性腺拍照观察。结果显示,转染试剂Lipofectamine 3000的转染效率最高,质粒、Lipofectamine 3000转染试剂和PGCs细胞数的配比为3 μg: 4 μL: 0.5×10 4个,转染5 h转染效率最高,达到23.4%,与现有的研究结果相比提高了2倍以上。移植回注PGCs到受体鸡胚中,荧光显微镜观察到鸡胚性腺中有GFP阳性细胞。本研究综合考虑转染试剂、质粒用量和细胞数量的影响因素以优化PGCs的转染条件,为高效制备转基因鸡及基因编辑鸡的研究奠定基础。
关键词:
Abstract
Keywords:
PDF (845KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
邹娴, 何燕华, 何静怡, 王艳, 舒鼎铭, 罗成龙. 鸡原始生殖细胞转染条件优化. 遗传[J], 2021, 43(3): 280-288 doi:10.16288/j.yczz.20-212
Zou Xian.
鸡(Gallus gallus)已被广泛用于早期胚胎发生、毒理学和干细胞等研究,特别是鸟类转基因和基因组编辑研究[1,2]。原始生殖细胞(primordial germ cells,PGCs)是精原细胞和卵原细胞的祖先细胞,能将遗传信息传递给下一代。鸡PGCs经过体外培养及基因修饰后,仍保持其生物学特性,移植回注到鸡胚血脉系统后可迁移到性腺并发育成功能性配子,产生转基因后代[3]。近年来,CRISPR/Cas9技术的发展及应用,使基因编辑鸟类模型成为可能,包括将LOXP位点导入鸡IgH位点、靶向卵球蛋白基因、标记基因插入Z染色体和抗J亚型禽白血病鸡模型[4⇓⇓~7]。但是,在利用鸡PGCs为载体进行基因操作过程中,需经历基因克隆、细胞转染、PGCs移植等步骤,而细胞转染技术则是此过程中的重要环节。目前鸡PGCs转染技术主要有脂质体转染、腺病毒或慢病毒转染和电转染,其中脂质体转染技术因其安全、操作简便、成本低而应用较多。但鸡PGCs脂质体转染效率较低(约5%~10%)[8],限制了其应用。本研究以鸡PGCs为模型,用3种不同脂质体转染试剂(Lipofectamine 2000、3000和 LTX & Plus Reagent)进行转染,摸索质粒浓度、转染试剂复合物配比以及细胞数量等,分析转染效率,旨在优化脂质体转染试剂介导的PGCs转染条件。
1 材料与方法
1.1 材料
种蛋为惠阳胡须鸡种蛋,来源于广东省农业科学院动物科学研究所原种鸡场。在Rcom PRO 50孵化箱(Rutoex)中38.5℃、相对湿度60%孵化至所需日龄。PiggyBac转座子和mPB转座酶质粒由华南农业大学提供;pPB-GFP转座子质粒由本实验室构建,内含由pGK启动子启动双基因puro和GFP表达的片段(图1)。图1
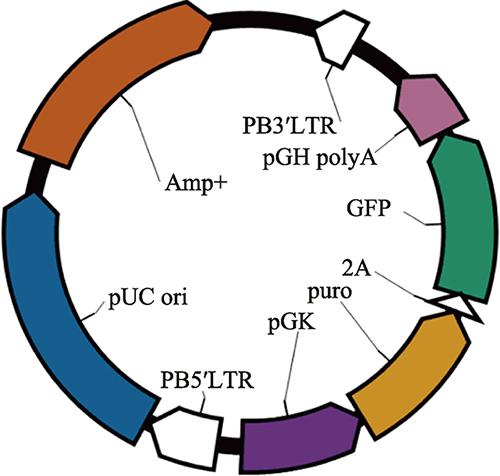 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1pPB-GFP质粒图谱
Fig. 1The plasmid map of pPB-GFP
1.2 细胞分离及培养
从孵育至5.5 d的惠阳胡须鸡胚性腺分离获得PGCs,然后接种至经钴源处理的BRL饲养层上培养,培养箱条件37℃、5% CO2。PGCs培养液中含56% KO-DMEM、30% BRL培养液、7.5%胎牛血清(FBS)、2.5%鸡血清、1×GS非必需氨基酸、1×GlutaMAXTM、1×双抗(青霉素和链霉素)、0.1 mmol/L β-巯基乙醇、重组人碱性成纤维细胞生长因子(rhFGF)、人干细胞因子(human SCF)。细胞每2 d进行半量换液,每3~4 d传代1次。从孵育至5.5 d的惠阳胡须鸡胚分离获得胎儿成纤维细胞(chicken embryo fibroblasts,CEF),培养箱条件为39℃、5% CO2,培养液中含89% DMEM、10%胎牛血清(FBS)和1%双抗(青霉素和链霉素)。1.3 qRT-PCR验证分析
利用qRT-PCR验证多能性基因POU5F1、PRDM14、NANOG和DDX4在PGCs和CEF中的表达量。PCR扩增体系为10 μL,包括5 μL SYBR® Green Master Mix,上下游引物各0.5 μL,1 μL cDNA 和3 μL ddH 2O。扩增条件:95℃ 1 min;95℃ 30 s,54~58℃ 30 s,72℃ 1 min,34个循环。参考NCBI中鸡相关基因序列,利用Primer 5.0软件设计扩增引物,GAPDH作为内参基因,由Sangon Biotech公司合成,引物信息见表1。Table 1
表1
表1用于qRT-PCR检测的基因引物
Table 1
| 基因 | 引物序列(5′→3′) | 扩增产物 长度(bp) |
|---|---|---|
| POU5F1 | CCACCCTCCCCACCTCTAC | 181 |
| TGTGCCAGCCTAACCTCCT | ||
| PRDM14 | TGTTCGCCTACCGCTACTACCG | 106 |
| AGTGCTGGCGGAGTGTGTGTG | ||
| NANOG | AGACCTCTCCTTGACCACA | 171 |
| CCACTCTCACCTTTATCCT | ||
| DDX4 | TCCCAGAGCCCACACAGATG | 124 |
| AAGTGATGCGCCCTCCTCTC | ||
| GAPDH | GGTGGTGCTAAGCGTGTTAT | 151 |
| ACCTCTGCCATCTCTCCACA |
新窗口打开|下载CSV
1.4 免疫荧光检测PGCs特征
PGCs铺到有盖玻片的24孔细胞培养板中制作细胞爬片,用4%多聚甲醛室温固定10~15 min,然后Tris base缓冲液(TBS)洗净;加入适量0.1% TritonX-100,室温破膜10 min后TBS缓冲液洗净;10%驴血清室温孵育20 min;甩去血清,滴加一抗工作液,4℃过夜;次日从4℃冰箱拿出,复温15 min,TBS缓冲液洗净,每张切片滴加50 μL荧光二抗(稀释比均为1∶400),37℃孵育30 min,TBS缓冲液洗净(此步骤开始进行避光);甩去TBS,每张切片滴加50 μL新鲜配置的DAPI (稀释比为1∶500),进行荧光染核,室温孵育10 min,TBS冲洗3次,每次5 min;甩去TBS,用抗荧光淬灭封片剂进行封片,避光保存于4℃,在激光共聚焦显微镜下拍照。1.5 实验分组与带绿色荧光蛋白(GFP)质粒的转染
前期我们在DF1细胞(鸡胚成纤维细胞系)中研究发现当转座子质粒pPB-GFP与转座酶质粒mPB的比例为3~4∶1时,转染效率最高。因此,本研究选择二者比例为3∶1,在PGCs中进行转染试验。转染前将PGCs转移到24孔板中,每孔接种103个PGCs。将PGCs随机分组,即Lipofectamine 2000 (添加质粒和Lipofectamine 2000,用A表示)、Lipofectamine 3000 (添加质粒和Lipofectamine 3000,用B表示)和Lipofectamine LTX & Plus Reagent(添加质粒和Lipofectamine LTX & Plus Reagent,用C表示)。根据转染试剂说明书做如下分组:A组依质粒剂量及转染试剂Lipofectamine 2000剂量分为A1 (1 μg: 2 μL)、A2 (3 μg: 2 μL)、A3 (5 μg: 3 μL)和A4 (6 μg: 4 μL);B组依质粒剂量及转染试剂Lipofectamine 3000剂量分为B1(1 μg: 2 μL)、B2(1.5 μg: 3 μL)、B3 (3 μg: 4 μL)和B4(5 μg: 6 μL);C组依质粒剂量及转染试剂Lipofectamine LTX剂量分为C1 (1 μg: 1 μL)、C2 (2 μg: 2 μL)、C3 (3 μg: 2 μL)和C4 (5 μg: 3 μL)。每小组3个重复。然后按照转染试剂盒说明进行转染,转染6 h后收集细胞,离心,重悬后接种到新的饲养层上,加入新鲜PGCs培养液,置于37℃、5% CO 2培养箱中继续培养48 h后用倒置显微镜照相并分析。
筛选出较高转染效率的组别后,提高细胞密度至0.3×104个/孔或0.5×104个/孔,减少转染时间至5 h,并用含有血清培养基转染,进一步优化转染条件。
1.6 流式细胞术分选稳定表达GFP蛋白的PGCs
将细胞接种到T25培养皿中,脂质体方法转染GFP蛋白质粒。同时以转染GFP蛋白质粒的DF1细胞作为阳性对照,以未转染任何质粒的DF1细胞作为阴性对照进行流式细胞分选。上机分选前,制备无菌操作液清洗流式细胞仪(BD FACSAriaII)。分别收集PGCs和DF1细胞到15 mL离心管中,并用PBS悬浮细胞,细胞数要大于106个/mL。分别用阳性和阴性对照DF1细胞设定流式细胞仪的取样参数和十字门的范围,再进行PGCs分选。1.7 阳性PGCs移植受体鸡及性腺分离
将PGCs培养液收集到15 mL离心管中,常温离心,用PBS洗一次,常温离心,之后加入适量的PBS悬浮细胞,细胞计数。取所需细胞数,加PBS至总体积的90%,然后加入10%台盼蓝,混匀。取2~2.5日龄鸡胚,鸡蛋尖端处开壳0.5~1 cm,在体视显微镜下找到鸡胚背主动脉,针斜面向下,沿着血流方向轻轻吹入1~3 µL含有PGCs的溶液,溶液中PGCs总量约为2000个。在开口周围涂上蛋清,用膜封好开口,待蛋清干了之后,放入孵化箱继续孵化。移植注射3.5 d后取出活胚,在体视显微镜下用镊子分离鸡胚性腺,置于滴有80 μL PBS的载玻片上,盖上盖玻片,倒置荧光显微镜下观察并拍照。
1.8 数据处理
荧光显微镜下统计每孔绿色荧光PGCs数量,数据用JMP 8.0 (SAS Institute, Cary, NC)统计软件检验,进行差异显著性分析(P<0.05为差异显著)。2 结果与分析
2.1 PGCs分离培养
本研究共分离40只鸡胚,其中有10个雄性PGCs在第3周结束之前细胞数达到5.0×104,被认为已成功建立了品系。这10个PGCs品系均来自单个胚胎。这些细胞呈圆形、边缘光亮、体积较大、细胞核大等形态特征,通常能看到细胞两个连在一起或成串,符合PGCs的形态特征(图2A)。PGCs培养60 d后,qRT-PCR检测生殖细胞标记基因POU5F1、PRDM14、NANOG和DDX4,结果表明,它们在PGCs中表达量高,在CEF中表达量极低(图2B),表明本研究分离的PGCs具有多能性。同时,通过免疫荧光法检测到几乎所有PGCs均表达了生殖细胞表面标志蛋白SSEA-1和DAZL (图2:C~F)。2.2 PGCs转染条件的优化
用3种不同转染试剂转染携带GFP蛋白的piggyBac转座子质粒。结果表明,Lipofectamine 3000转染试剂中B3阳性细胞数最多,高于所有组别;其次是Lipofectamine 3000转染试剂B2,第三是Lipofectamine LTX & Plus Reagent转染试剂C3 ( 表2)。实验过程中还观察到Lipofectamine 2000转染试剂组以及其他两种试剂的高浓度组B4和C4对PGCs伤害较大,48 h后观察到大量PGCs死亡。在B3转染条件基础上,提高每孔中PGCs细胞数量到0.5×104,并减少转染时间至5 h,同时加入含有血清培养基,48 h后,在荧光显微镜下计数,发现阳性细胞数更多,每孔阳性细胞数超过了1000个。初步判断此条件下PGCs转染效率更佳(命名为D1),即质粒3 μg、Lipofectamine 3000试剂4 μL和PGCs细胞数0.5×10 4的配比。2.3 稳定表达GFP蛋白PGCs细胞株的获得
为了确定最佳转染条件,根据B3转染条件和D1转染条件分别转染PGCs细胞,72 h后,利用GFP基因表达绿色荧光蛋白,用流式细胞仪进行阳性细胞分选。以未转染任何质粒的DF1细胞作为阴性对照、转染GFP蛋白质粒的DF1细胞作为阳性对照。图3可知,DF1阴性对照组中阳性细胞比例为0% (图3A),DF1阳性对照组中阳性细胞比例为46.2% (图3B),B3转染条件下阳性PGCs比例为12.7% (图3C),D1转染条件下阳性PGCs比例为23.4% (图3D)。图2
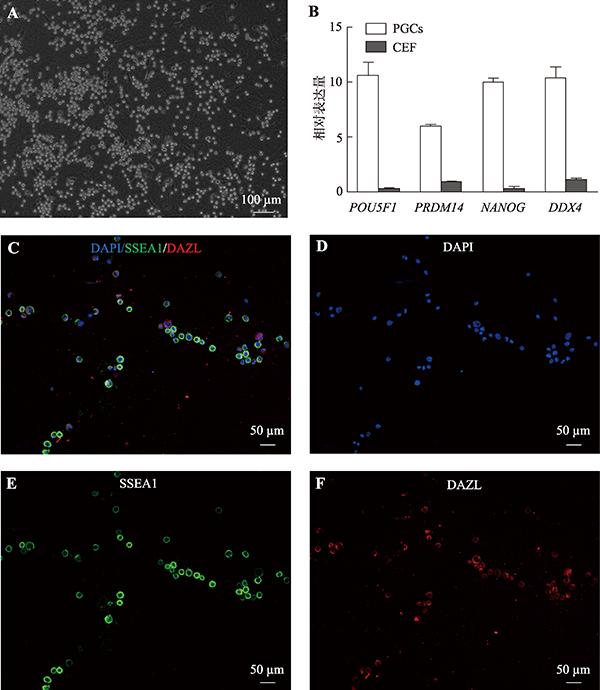 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2PGCs的分离培养
A:PGCs体外培养3周后的形态特征;B:qRT-PCR检测POU5F1、PRDM14、NANOG和DDX4基因在PGCs和CEF中表达情况;C:PGCs中生殖细胞表面标志蛋白SSEA-1(绿色荧光)、DAZL蛋白(红色荧光)的表达;D:PGCs核染色;E :PGCs中生殖细胞表面标志蛋白SSEA-1蛋白;F:PGCs中生殖细胞表面标志蛋白DAZL蛋白。
Fig. 2Isolation and culture of PGCs
Table 2
表2
表2不同转染试剂和不同剂量GFP质粒转染PGCs后48 h的阳性细胞数(个/孔)
Table 2
| Lipofectamine 2000 | Lipofectamine 3000 | Lipofectamine LTX & Plus Reagent |
|---|---|---|
| A1 (20.30±1.70)b | B1 (50.40±7.10)c | C1 (30.00±5.50)c |
| A2 (40.50±3.50)a | B2 (102.10±3.10)b | C2 (55.90±4.40)b |
| A3 (46.50±5.10)a | B3 (278.50±10.90)a | C3 (80.90±8.10)a |
| A4 (40.70±10.80)a | B4 (75.20±8.60)b | C4 (63.30±9.20)b |
新窗口打开|下载CSV
图3
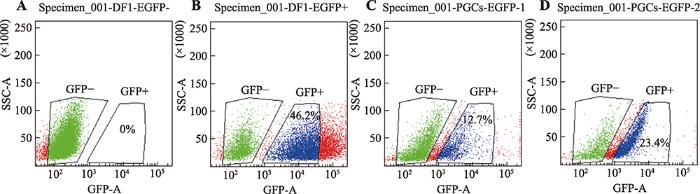 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3流式细胞仪分选阳性PGCs
A:未转染任何质粒的阴性DF1细胞分选结果;B:转染了GFP蛋白质粒的DF1细胞分选结果;C:B3转染条件下阳性PGCs分选结果;D:D1转染条件下阳性PGCs分选结果。
Fig. 3Screening of positive PGCs by flow cytometry
将流式细胞仪分选后的PGCs接种到24孔板继续培养,状态良好(图4)。继续培养72 h后进行传代。
图4
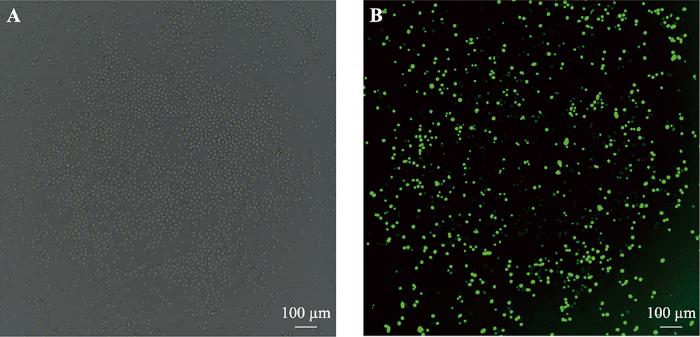 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4稳定表达绿色荧光蛋白的PGCs
A:白光视野;B:绿色荧光视野。由于鸡PGCs是悬浮细胞,无法固定,会漂来漂去,导致PGCs并不总在同一平面或同一位置,因此绿色荧光视野下拍摄的PGCs荧光有强有弱,A和B图并不能完全匹配上。
Fig. 4PGCs were stably expressing green fluorescent protein
2.4 鸡早期胚胎注射技术生产转基因鸡
阳性PGCs培养3周后,大量细胞继续表达GFP蛋白,表明GFP基因已稳定整合到PGCs中。把它们注射到2~2.5胚龄的100只受体鸡胚中,继续孵化至5.5~6胚龄,分离性腺,在荧光显微镜下观察到有2只鸡胚性腺表达绿色荧光蛋白(图5),表明本研究分离、筛选的PGCs能够在受体鸡中继续发育。考虑到此次注射的PGCs在体外培养时间较长(约200 d),本文重新分离PGCs并转染pPB-GFP质粒,流式细胞术分选阳性PGCs,继续培养3周后注射到200只鸡胚中,随机取了15枚,荧光显微镜下观察到5枚鸡胚性腺表达绿色荧光蛋白。经PCR鉴定性腺中GFP基因表达情况,结果表明阳性性腺中可以检测到GFP基因的表达,而阴性性腺则无法检测到(图5E)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5含有表达绿色荧光蛋白PGCs的鸡胚性腺
A:阴性鸡胚性腺(白光视野);B:阴性鸡胚性腺(绿色荧光视野);C:阳性鸡胚性腺(白光视野);D:阳性鸡胚性腺(绿色荧光视野);E:PCR鉴定阳性性腺。泳道1和2为表达GFP蛋白的性腺,3和4为注射了阳性PGCs但不表达GFP蛋白的性腺,5和6为未注射阳性PGCs的性腺,M为DL2000 Marker。
Fig. 5Chicken embryo gonads containing PGCs that were stably expressing green fluorescent protein
3 讨论
鸡肉是人类最重要的食物来源之一,也是用于生产药用蛋白的生物反应器[9]。随着基因编辑技术的发展,鸡胚胎作为发育和细胞生物学的模型有了更多的机遇,但鸟类在基因组编辑技术方面一直落后于哺乳动物[1,10,11]。本研究旨在探究提高鸡PGCs转染效率的方法,为家禽基因组编辑技术的应用奠定基础。本研究利用piggyBac转座子,比较了3种不同转染试剂的转染效率,优化了转染条件,经流式细胞仪分析转染效率最高达到了23.4%,PGCs回注到鸡胚血液中,获得了表达GFP蛋白的鸡胚。Macdonald等[12]利用piggyBac及Tol2转座子有效转染PGCs后回注到鸡胚血脉系统,获得表达GFP蛋白的转基因鸡,piggyBac的转染效率为10.5%。陈美娟等[8]利用piggyBac转座子转染PGCs后回注到鸡胚血脉系统,获得表达GFP蛋白的鸡胚,转染效率最高为6.16%。由此可知,本研究中PGCs的转染效率高于上述研究结果,筛选出的阳性PGCs可以在受体鸡胚中继续发育,为后续开展基因编辑鸡的研究奠定了良好基础。
本研究用脂质体转染方法转染PGCs,比较转染试剂的转染效率。结果发现,质粒3 μg、Lipofectamine 3000试剂4 μL和PGCs细胞数10 4的配比时转染效率最高。据报道,在一定范围内, 质粒用量和转染效率成正比,但当DNA加入过量时,转染效率反而下降,与本研究结果一致[13,14]。本研究还发现,质粒浓度和转染试剂剂量相同情况下,提高细胞密度,可以提高转染效率。这可能是由于细胞密度大,细胞间空隙较小,质粒DNA与细胞接触面多,进入细胞的阻力变小,从而提高转染水平。Lipofectamine 3000试剂在操作过程中不需要使用无血清、无双抗培养基,对PGCs后续生长影响较小。另外,在PGCs传代后第2 d、3 d、4 d时用最优转染条件进行转染,发现转染效率差异不显著(数据未呈现),表明PGCs传代后第2~4 d均可进行转染试验。
目前获得PGCs的方法主要有两种:一是从孵化2.5 d左右鸡胚循环血液中获得;二是从孵化5 d左右鸡胚性腺中获得。血液和性腺PGCs的细胞生长和特性基本没有差异,均表现出FGF依赖性的生长速率,且两种方法获得的PGCs均能在体外培养并进行基因修饰后产生转基因鸡[3,7,15⇓⇓⇓⇓⇓~21]。本研究从孵化5.5 d鸡胚性腺中分离培养PGCs。与循环血液中PGCs相比,使用性腺PGCs的优点是可以从一个胚胎中获得更多的PGCs,且每个胚胎都更容易发育出许多PGCs,更有效地筛选出具有生殖细胞多能性的PGCs系,以产生胚系嵌合体和转基因鸡。
相比脂质体转染试剂Lipofectamine 2000和Lipofectamine LTX & Plus Reagent,Lipofectamine 3000转染试剂更适用于转染PGCs,最优转染条件为:质粒3μg、Lipofectamine3000试剂4μL和PGCs细胞数0.5×104的配比, 且可以用有血清有双抗的培养基。本研究综合考虑转染试剂、质粒用量和细胞数量的影响因素以优化PGCs的转染条件, 为高效制备转基因鸡及基因编辑鸡的研究奠定基础。
(责任编委: 姜雨)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}