Effect of extensin-like OsPEX1 on pollen fertility in rice
Dai Hang,1, Li Yan1, Liu Shunchun1, Lin Lei1, Wu Juanyan1, Zhang Zhijie1, Peng Qichun1, Li Nan1, Zhang Xiangqian,11 College of Forestry and Landscape Architecture, South China Agricultural University, Guangzhou 510642, China
A:野生型和转基因RNAi植株的主穗;B:野生型和转基因RNAi植株的单穗结实率;C:野生型和转基因RNAi植株中OsPEX1基因的相对表达量。WT:野生型植株;KD:RNAi转基因植株。 Fig. 5Phenotypes of wild type and transgenic rice plants with RNAi-mediated knock-down of OsPEX1 expression and expression analysis of OsPEX1 in the transgenic plants
WT:野生型植株;KD:RNAi转基因植株。 Fig. 6I2-KI staining of pollen grains in wild type and T2 transgenic plants generated by RNAi-mediated knock-down of OsPEX1 expression
RubinsteinAL, BroadwaterAH, LowreyKB.BedingerPA. Pex1, a pollen-specfic gene with an extensin-like domain Proc Natl Acad Sci USA, 1995,92(8):3086- 3090. [本文引用: 2]
BaumbergerN, RingliC, KellerB. The chimeric leucine- rich repeat/extensin cell wall protein LRX1 is required for root hair morphogenesis in Arabidopsis thaliana. Genes Dev, 2001, 15(9):1128- 1139. [本文引用: 2]
BaumbergerN, DoessegerB, GuyotR, DietA, ParsonsRL, ClarkMA, Simmons, MP, Bedinger P, Goff SA, Ringli C, Keller B Whole-genome comparison of leucine-rich repeat extensins in Arabidopsis and rice. A conserved family of cell wall proteins form a vegetative and a reproductive clad Plant Physiol, 2003,131(3):1313- 1326. [本文引用: 1]
WangXX. Functional characterization of Arabidopsis pollen-specific expressed LRXs in pollen germination and pollen tube growth[Dissertation]. Hebei Normal Univ, 2017. [本文引用: 1]
CampbellL, SR.SR. A Comprehensive analysis of RALF proteins in green plants suggests there are two distinct functional groups Turner, Front Plant Sci, 2017,8:37. [本文引用: 1]
MecchiaMA, Santos-FernandezG, DussNN, SomozaSC, Boisson-DernierA, GagliardiniV, Martinez-BernardiniA, FabriceTN, RingliC, MuschiettiJP, GrossniklausU. RALF4/19 peptides interact with LRX proteins to control pollen tube growth in Arabidopsis Science, 2017, 358( 6370): 1600- 1603. [本文引用: 2]
HergerA, GuptaS, KadlerG, FranckCM, Boisson-Dernier A, RingliC. Overlapping functions and protein- protein interactions of LRR-extensins in Arabidopsis PLoS Genet, 2020,16(6): e1008847. [本文引用: 1]
MarzolE, BorassiC, BringasM, SedeA, RodríguezGarcia DR, CapeceL, EstevezJM. Filling the gaps to solve the extensin puzzle Mol Plant, 2018,11(5):645- 658. [本文引用: 1]
NguyenQN, LeeYS, ChoLH, JeongHJ, AnG, JungKH. Genome-wide identification and analysis of Catharanthus roseus RLK1-like kinases in rice Planta, 2015,241(3):603- 613. [本文引用: 1]
NissenKS, WillatsWGT, MalinovskyFG. Understanding CrRLK1L function: cell walls and growth control Trends Plant Sci, 2016,21(6):516- 527. [本文引用: 1]
ChenCJ, ChenH, ZhangY, ThomasHR, FrankMH, HeY, XiaR. TBtools: An integrative toolkit developed for interactive analyses of big biological data Mol Plant, 2020,13(8):1194- 1202. [本文引用: 1]
ZhangXQ, ZhouJS, ZhuHT, LiX Y, ZengR Z. Genetic analysis and gene mapping of an early flowering and multi-ovary mutant in rice Oryza sativa L. Hereditas (Beijing), 2008, 30(10): 1349- 1355. [本文引用: 1]
MaH. Molecular genetic analyses of microsporogenesis and microgametogenesis in flowering plants Annu Rev Plant Biol, 2005,56:393- 434. [本文引用: 1]
PaciniE, GuarnieriM, NepiM. Pollen carbohydrates and water content during development, presentation, and dispersal: a short review Protoplasma, 2006,228(1- 3):73- 77. [本文引用: 1]
YamagataY, YamamotoE, AyaK, WinK T, DoiK, Sobrizal, Ito T, Kanamori H, Wu J, Matsumoto T, Matsuoka M, Yoshimura A. Mitochondrial gene in the nuclear genome induces reproductive barrier in rice Proc Natl Acad Sci USA, 2010,107(4):1494- 1499. [本文引用: 1]
UedaK, YoshimuraF, MiyaoA, HirochikaH, NonomuraK, WabikoH.Collapsed abnormal pollen1 gene encoding the arabinokinase-like protein is involved in pollen development in rice Plant Physiol, 2013, 162(2): 858- 871. [本文引用: 1]
ZhouSR, WangY, LiWC, ZhaoZG, RenYL, WangY, GuSH, LinQB, WangD, JiangL, SuN, ZhangX, LiuLL, ChengZJ, LeiCL, WangJL, GuoXP, WuFQ, IkehashiH, WangHY, WanJM. Pollen semi-sterility1 encodes a kinesin-1-like protein important for male meiosis, anther dehiscence, and fertility in rice Plant Cell, 2011, 23( 1): 111- 129. [本文引用: 2]
WuLN, GuanYS, WuZG, YangK, LvJ, ConverseR, HuangYX, MaoJX, ZhaoY, WangZW, MinHQ, KanDY, ZhangY.OsABCG15 encodes a membrane protein that plays an important role in anther cuticle and pollen exine formation in rice Plant Cell Rep, 2014, 33(11): 1881- 1899. [本文引用: 1]
TangJY, ChuCC. MicroRNAs in crop improvement: fine-tuners for complex traits Nat Plants, 2017, 3( 7): 17077. [本文引用: 1]
GloverJ, GrelonM, CraigS, ChaudhuryA, DennisE. Cloning and characterization of MS5 from Arabidopsis : a gene critical in male meiosis Plant J, 1998,15(3):345- 356. [本文引用: 1]
ZhangPF, SongYL, ZhangGS, ZhaoXL, BaQS, LiuHZ, ZhuWW, LiZK, WangJW, NiuN. Relationship between abnormal metabolism of tapetum and microspore abortion in male sterile line of wheat Sci Agric Sin. 2014, 47(09): 1670- 1680. [本文引用: 1]
JungKH, HanMJ, LeeDY, LeeYS, SchreiberL, FrankeR, FaustA, YephremovA, SaedlerH, KimYW, HwangI, AnG. Wax-deficient anther1 is involved in cuticle and wax production in rice anther walls and is required for pollen development Plant Cell, 2006, 18( 11): 3015- 3032. [本文引用: 1]
LiN, ZhangDS, LiuHS, YinCS, LiXX, LiangWQ, YuanZ, XuB, ChuHW, WangJ, WenTQ, HuangH, LuoD, MaH, ZhangDB. The rice tapetum degeneration retardation gene is required for tapetum degradation and anther development Plant Cell, 2006, 18(11): 2999- 3014. [本文引用: 1]
LiL, LiB, XieC, ZhangT, BorassiC, EstevezJM, LiXS, LiuX.Arabidopsis RAD23B regulates pollen development by mediating degradation of KRP1 J Exp Bot, 2020,71(14):4010- 4019. [本文引用: 1]
WangB, FangRQ, ZhangJ, HanJL, ChenFM, HeFR, LiuYG, ChenLT. Rice LecRK5 phosphorylates a UGPase to regulate callose biosynthesis during pollen development J Exp Bot, 2020, 71(14):4033- 4041. [本文引用: 1]
MaL, XuW, DouLL, KeXN, LiuMY, GengYF, HuangX, JiaYF, LiuQP. Research progress on pollen fertility- related genes in rice ( Oryza sativa L. Jiangsu Agri Sci, 2019, 47(10): 42- 47. [本文引用: 1]
 ,1, 李延1, 刘树春1, 林磊1, 吴娟燕1, 张志杰1, 彭崎春1, 李楠1, 张向前
,1, 李延1, 刘树春1, 林磊1, 吴娟燕1, 张志杰1, 彭崎春1, 李楠1, 张向前
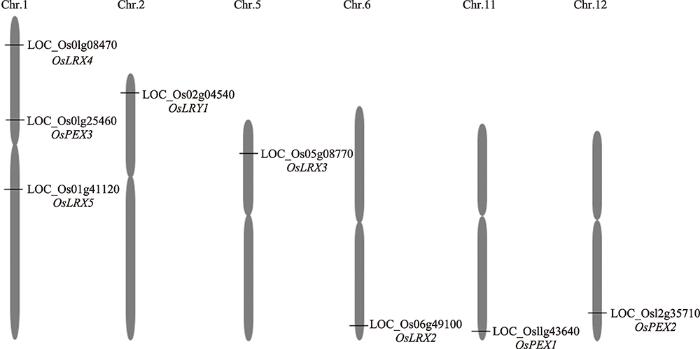 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT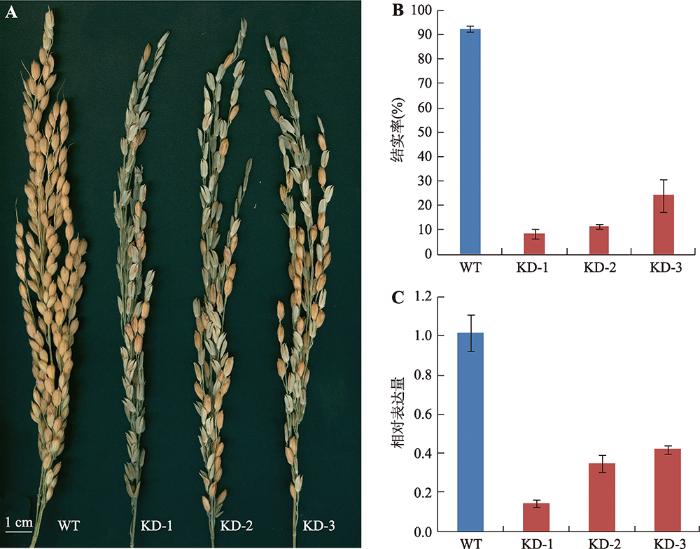 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT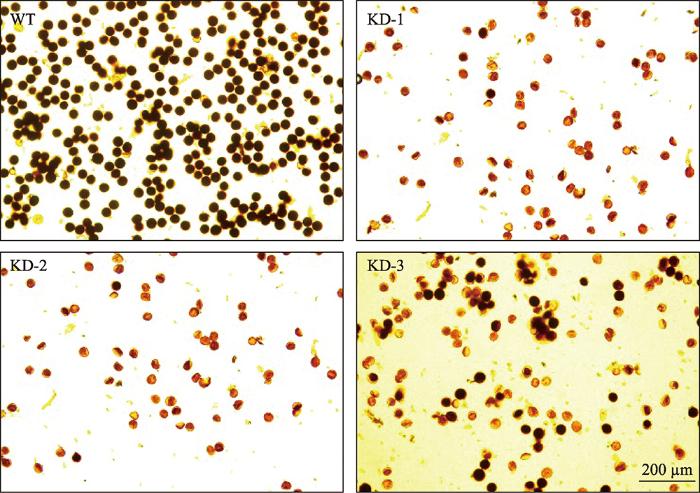 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}