 ,1, 吴宏,1, 于黎,11
,1, 吴宏,1, 于黎,11 Progress on coat color regulation mechanism and its association with the adaptive evolution in mammals
Zhang Yuxing,1, Wu Hong,1, Yu Li,11 通讯作者: 通讯作者: 于黎,博士,研究员,研究方向:动物遗传与进化。E-mail:yuli@ynu.edu.cn吴宏,博士后,研究方向:动物遗传与进化。E-mail:whzxsg@126.com
第一联系人:
收稿日期:2020-11-18
| 基金资助: |
Received:2020-11-18
| Fund supported: |

摘要
哺乳动物类群呈现出的丰富毛色是引人注目的一种生物现象,是研究和理解哺乳动物适应性进化的理想模型之一。哺乳动物的毛色多态在躲避天敌、捕食、求偶及抵御紫外线等方面都具有重要作用。哺乳动物毛发的色素化过程由体内黑色素的数量、质量和分布状况所决定。黑色素的形成过程复杂,包括黑素细胞的分化、成熟,黑素体等细胞器的形态发生及黑色素在黑素细胞中的合成代谢和转运等过程;而在细胞色素化的每个阶段/时相都伴随着一些重要功能基因的参与,并通过基因之间的相互作用形成了黑色素生物代谢的复杂调控网络,进而形成不同的毛色有助于哺乳动物适应不同生存环境。对哺乳动物不同毛色形成机制的探究一直以来都是遗传学及进化生物学的重要研究领域和聚焦热点。本文综述了哺乳动物毛色色素化过程的主要分子机制以及毛色适应性进化的遗传基础,以期为哺乳动物毛色多态及其适应性进化的分子机制研究提供参考。
关键词:
Abstract
Keywords:
PDF (772KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
章誉兴, 吴宏, 于黎. 哺乳动物毛色调控机制及其适应性进化研究进展. 遗传[J], 2021, 43(2): 118-133 doi:10.16288/j.yczz.20-390
Zhang Yuxing.
哺乳动物类群展现出的丰富多彩的毛色特征是一种普遍且备受关注的生物现象。毛色多态性普遍存在于种内和种间,比如:孟加拉虎(Panthera tigris tigris)有4种毛色表型,其中普遍存在的是橙色底黑色条纹,而其变种白虎是白色底黑棕条纹,金虎是金色底红棕色条纹以及雪虎是几乎全身雪白的毛色[1]。马(Equus ferus caballus)的毛色繁多,可大致分为黑色、骝毛、栗毛等[2]。不同品种的家犬(Canis lupus familiaris)毛色也分为多种,例如拉布拉多犬有黑色、巧克力色和黄色[3],德国牧羊犬有黑色、蓝色和肝脏色等[4]。除了种内多态,毛色多态性还体现在种间,例如金丝猴属中有几乎全身为金黄色的川金丝猴(Rhinopithecus roxellana),黑色和白色为主的滇金丝猴(Rhinopithecus bieti)和越南金丝猴(Rhinopithecus avunculus),黑褐色和金黄色为主的黔金丝猴(Rhinopithecus brelichi)以及全身黑色为主的怒江金丝猴(Rhinopithecus strykeri)[5]。在猫科动物中,大部分猫科动物类群间都可以毛色、斑点或条纹的差异而进行区分[6]。
哺乳动物种内或种间的体色多态很大程度上是自然选择作用的结果;特定类群所呈现的特殊毛色分布模式往往反映了其对周围环境的协调和适应,例如躲避天敌、捕食、求偶以及抵御紫外线等[7]。亚利桑那州南部的岩小囊鼠(Chaetodipus intermedius)为了避免被捕食,其毛色类型与其栖息地密切相关,火山熔岩附近区域生活的岩小囊鼠有着深色皮毛,而生活在较远且遍布浅色花岗岩地区的群体其被毛颜色则相对较浅[8]。分布于喀斯特地貌区域的石山叶猴(Trachypithecus francoisi)除了头部、尾部有白色、黄色毛发外,几乎全身被黑色毛发覆盖,该毛色可能是石山叶猴的一种伪装适应策略[9]。大熊猫(Ailuropoda melanoleuca)黑白相间的毛色也与伪装有关,白色的毛发有助于在积雪的栖息地中隐藏,而黑色毛发则有助于在森林中隐藏,这有助于大熊猫在两种不同栖息地中活动[10]。不同猫科动物类群所具有的特殊毛色特征与其捕食行为密切相关,比如鹿和羊,因其仅具有二色视觉而无法有效区分橙色和绿色,因此捕食鹿和羊的老虎(Panthera tigris)其橙色被毛对于鹿和羊来说是一种非常有效的伪装,能够与所处的环境融为一体,使其能不动声色地靠近、捕食猎物[11]。Caro等[12]对鲸类(Cetacea)的着色模式研究发现,吃鱼、虾和乌贼的鲸类往往具有醒目的斑块化体色(例如:白侧腹、白头和不对称的白色下颌骨),这些明显的白色斑块能够迷惑猎物,使其聚集在一起,便于鲸类捕食。此外,性选择作用对于塑造哺乳动物类群间的毛色多态也有着重要影响[13]。如雄狮(Pantheraleo)的鬃毛是性选择的产物,深色的鬃毛对雌狮更有吸引力[14]。Cooper等[15]将10只不同亚种的褐狐猴(Eulemur fulvus)分别放进笼子里,在每个笼子中提供经过处理的不同色彩的雄猴照片,记录每只雌猴看照片的时间,结果发现雌猴更喜欢色彩鲜艳的雄性。在抵御紫外线方面,栖息于高海拔地区的藏猪(Sus scrofa domesticus)通过其黑色的毛发来抵御紫外辐射,能有效避免紫外辐射引起的DNA损伤[16]。
除了自然选择作用外,由于人类文明的兴起和发展,体色作为一种重要的经济性状,不同家养动物类群也受到强烈的人工选择作用而表现出高度的体色多态性。例如驴(Equus asinus)的毛色可分为三粉、黑色、白色等[17];猪也有丰富的毛色表型,如黑色、红色、黑斑等[18];绵羊(Ovis aries)也具有白色、灰色、棕色等多种毛色[19]。
对哺乳动物不同毛色形成机制的探究和揭示一直以来都是遗传学及进化生物学的重要研究领域和聚焦热点。杨广礼[20]、巫小倩等[21]、王磊等[22]先后对动物的毛色形成机制进行了综述。对哺乳动物的毛色长期研究发现,哺乳动物的毛色、眼睛颜色和肤色由体内黑色素的数量、质量和分布状况决定[23]。在哺乳动物中,一般存在真黑素和褐黑素两种黑色素:真黑素(棕黑或深色不溶性聚合物)能使皮肤和毛发表现为黑色或褐色;褐黑素(红黄色可溶性聚合物)能使皮肤和毛发表现为红色和黄色[24]。黑色素在哺乳动物的毛色形成中扮演着关键角色,黑色素的形成有着复杂的调控机制,从黑素细胞的分化成熟到黑色素合成及转运的过程中都有着多个基因的参与调控[25]。此外,越来越多的研究表明,microRNA (miRNAs)也可以调控黑色素沉积从而影响哺乳动物的毛色[21,26]。
本文综述了哺乳动物毛色色素化形成的主要调控机制,进一步阐述了这些重要功能基因产生的不同遗传突变对于哺乳动物适应性进化的重要意义(自然选择与人工选择);同时,也对哺乳动物类群中毛色多态与某些重大遗传疾病之间的关联性进行了探讨,以期为哺乳动物毛色多态、适应性进化以及重要遗传疾病致病机理的分子机制研究提供参考。
1哺乳动物色素化过程调控机制
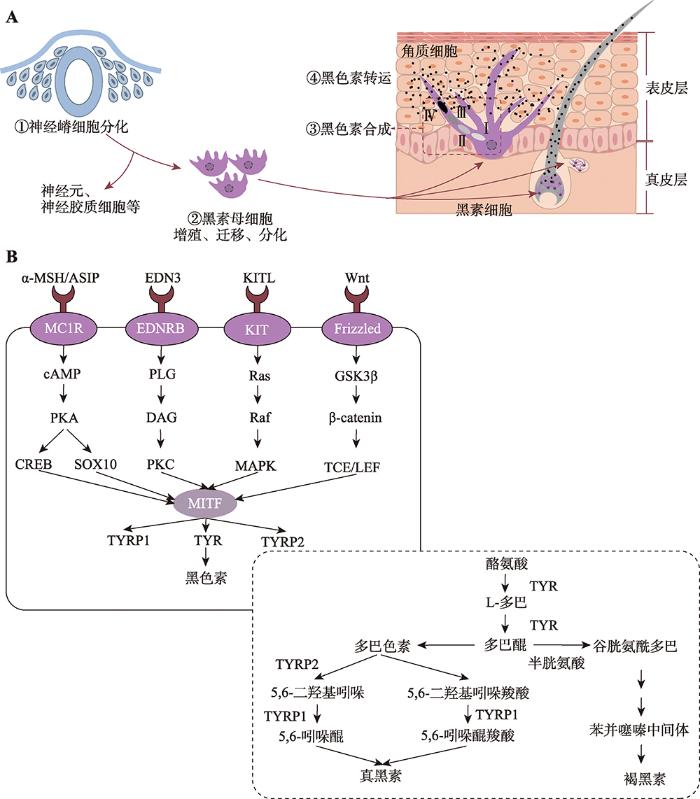
细胞色素化过程包含多个时相和阶段,主要分为:黑素细胞的发育成熟、黑素细胞中重要细胞器的形态发生、黑色素的合成代谢及黑色素的胞内胞外转运和功能行使等[27]。黑素细胞由黑素母细胞分化而来,该细胞起源于胚胎时期躯干神经嵴细胞[28]。神经嵴细胞是一种多能细胞,具有分化为黑素细胞、外周神经系统的神经元、神经胶质细胞和肾上腺髓质的嗜铬细胞等多种细胞的潜能[29]。神经嵴细胞分化为黑素细胞主要有两种途径:第一种通过背侧迁移途径,即沿着体节和外胚层的间隙迁移。前期神经嵴细胞分化为黑素母细胞,黑素母细胞沿着背侧途径增殖并迁移至胚胎各处。随后,黑素母细胞通过基底膜进入表皮层,其中部分黑素母细胞会进一步迁移至毛囊基质中。位于表皮层的黑素母细胞分化为黑素细胞后负责皮肤色素沉着;而迁移到毛囊基质的黑素母细胞,一部分分化为成熟的黑素细胞,还有一部分黑素母细胞则形成黑素细胞干细胞(melanocyte stem cells, MSC),负责维持体内平衡[25]。第二种是腹侧迁移途径,早期的神经嵴细胞沿腹侧迁移分化为神经元和施万细胞前体(schwann cell precursors, SCPs),SCPs进一步分化为施万细胞和黑素细胞,黑素细胞的分化与发育受神经调节蛋白1 (neuregulin-1, NRG1)、胰岛素样生长因子1 (insulin- like growth factor 1, IGF1)和血小板衍生因子(platelet- derived growth factor, PDGF)调控,NRG1抑制黑素细胞的分化而IGF1和PDGF促进SCPs分化为黑素细胞[30,31]。黑素体是黑素细胞中由高尔基体-内质网-溶酶体形成的不连续的膜状细胞器,是黑色素合成的场所[32]。在黑素体中,酪氨酸首先通过酪氨酸酶(tyrosinase, TYR)的催化作用形成多巴,再进一步氧化生成多巴醌(DOPA quinone, DQ),后经一系列酶促反应最终生成真黑素;当半胱氨酸存在时,DQ便与半胱氨酸反应生成半胱氨酸多巴,经过闭环和脱羧反应生成褐黑素[33,22]。此外,真黑素和褐黑素的合成还与黑素体内的pH值有关,低pH值有利于褐黑素合成而高pH值则有利于真黑素合成[34]。黑色素合成后,成熟的黑素体通过各种分子马达运输黑色素到周围的角质细胞中行使色素功能[35,36]。与皮肤黑色素生成不同的是,毛囊中黑色素的生成与毛发周期有关,在毛发生长期,毛囊黑素细胞中黑素体合成黑色素,随后成熟的黑素体转运至周围的皮层角质细胞最终形成有色毛干。与表皮黑色细胞相比,毛囊黑色细胞更大,树突更多,有更大的高尔基体和内质网,能产生更大的黑素体[30](图1)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1哺乳动物细胞色素化过程及其调控机制
A:黑色素合成的细胞过程;B:黑色素合成信号通路。根据参考文献[33]修改绘制。
Fig. 1Pigmentation process and its regulatory mechanism in mammals
1.1黑素细胞发育过程调控
随着第一个毛色基因TYRP1在小鼠(Mus musculus)中被克隆到现在[37],已经发现171个毛色相关基因(378个突变位点)在不同阶段调控毛色/肤色的产生(Wnt蛋白是富含半胱氨酸的分泌型糖蛋白,在诱导神经嵴细胞的迁移、增殖和分化过程中起重要作用[38]。Wnt信号通路能够通过不同方式和途径影响细胞存活、增殖、分化和迁移[39]。Wnt信号通路通过黑素细胞中β-catenin的积累而进入细胞核后与淋巴细胞增强因子(lymphoid enhancer factor 1, LEF1)结合而增强小眼畸形转录因子(microphthalmia-associated transcription factor, MITF)的转录表达,最终促进黑素母细胞的增殖分化,进而影响黑色素合成[40]。此外,Wnt1A和Wnt3A也是Wnt信号通路中重要的调控分子;Wnt1可促进黑素母细胞分化为黑素细胞,Wnt3a促进神经嵴细胞向黑素细胞的分化,维持或上调黑素母细胞中MITF的表达及其下游基因TYR和TYRP1来促进黑色素的生成[41,42]。
内皮素(endothelin 3,EDN3)及其受体EDNRB在黑素细胞的发育中有着不可或缺的作用。这两个基因上发生的遗传突变均会导致黑素细胞前体的数量减少和色素稀释[43]。EDN3在黑素细胞发育的整个过程中是必需的,它在黑素细胞发育早期可以补偿KIT的部分作用,辅助影响黑色素的形成,在KIT基因突变的小鼠中EDN3的过表达能够减少白斑[44]。
KIT/KITL信号通路是黑素母细胞存活所必需。KIT基因编码酪氨酸激酶受体,其表达量变化会影响黑素细胞的迁移过程。黑素母细胞迁移时,KIT与其配体KITL结合后通过MAPK使MITF-M的Ser73磷酸化而导致MITF-M功能上调;与此同时,转录共刺激分子(CBP/p300)也与MITF-M相互作用,进一步激活MITF-M的功能[45]。当KIT基因发生突变不能正常表达时,会影响黑素细胞的正常迁移,导致毛根中缺乏黑素细胞而出现毛色稀释,白色毛发或白色斑点表型[45,46]。KIT和MITF能共同调控黑素细胞的发育,二者之间有着复杂的相互作用,MITF是维持黑素细胞中KIT表达所必需的,而KIT信号传导则可调节黑素细胞中MITF的活性和稳定性。MITF首先在促进黑素母细胞向黑素细胞的分化中起作用,随后通过影响KIT表达来促进黑素细胞的存活和迁移[47]。
MITF属于MiT转录因子家族成员,参与多种细胞类型的分化发育,例如黑素细胞、破骨细胞和肥大细胞等[48]。MITF基因有多个启动子,存在多个具有不同5?外显子的同工型,其中MITF-M几乎只在黑素细胞和黑素细胞瘤中表达。MITF具有螺旋-环-螺旋-亮氨酸拉链结构(bHLHzip),它可以识别并与E-box(5?-CACGTG-3?)、M-box(5?-TCATGTG-3?)序列相互作用,形成专一的二聚体,对胚胎神经嵴细胞向黑素细胞的分化选择过程至关重要。MITF在黑素细胞发育过程中扮演着交通枢纽角色,可以和多种转录因子结合进而调节上游或下游信号通路。在上游信号通路中,MITF-M启动子与LEF1/TCF结合激活MITF表达,激活Wnt/β-catenin通路转导促进黑素细胞谱系的建立[49]。CREB是一种bZIP转录因子,可识别MITF启动子中的TGACGTCA基序促进MITF-M的表达,进而响应MC1R下游的cAMP浓度变化,最终促进黑色素的沉着。SOX10能响应cAMP信号而激活MITF-M,而PAX3通过与MITF基因的启动子结合来调节MITF的表达;因此,PAX3与SOX10对MITF的表达有协同作用,这对黑色素细胞的发育和黑素细胞干细胞活化的调节是必需的。FOXD3通过抑制PAX3与MITF启动子的结合,降低MITF的表达最终维持黑素细胞的存活[48?~50]。在下游信号通路中,MITF能够识别并结合TYR家族启动子中的E-box序列,指导该家族成员(TYR、TYPR1、DCT)在黑素细胞中的特异性表达,从而产生黑色素[51](图1)。
1.2黑素体形态发生的调控
黑素体在黑素细胞中的形态发生和成熟主要包括4个阶段(I-IV期)。I期黑素体是圆形无定型的基质小囊泡,细胞器腔内开始出现早期的蛋白原纤维。II期黑素体包含有序的结构化纤维基质,黑素体呈椭圆形,无色素沉着。III期TYR催化L-酪氨酸氧化形成多巴,多巴经过氧化形成多巴醌,后者再经过一系列反应在黑素体中生成黑色素。IV期黑色素沉积在蛋白纤维上并充满整个黑素体[52??~55]。在黑素体形成前期(I-II期)中,PMEL基因编码一种黑素细胞特异性蛋白,并形成生理淀粉样蛋白纤维。前黑素体蛋白(PMEL)首先在内质网中合成,经过高尔基体和高尔基体网状结构加工修饰后进入黑素体(I期)。其进入方式有两种:(1) PMEL与衔接蛋白-1(AP1)首先形成复合物,之后通过网格蛋白包被的囊泡直接分选进入黑素体;(2) AP2与PMEL形成的复合物通过间接途径先分选至质膜,再重新回到黑素体[56],在II期形成纤维状结构,黑色素最终沉积在黑素体的管腔中。PMEL基因失活会导致黑素体的形态发生改变,导致毛色被稀释[57]。除PMEL外,MART1、GPNMB也是黑素体形成过程中重要的功能基因。MART1能与PMEL形成复合物,维持PMEL在黑素体中稳定表达。将MART1进行siRNA处理后,黑素细胞中PMEL的稳定性受影响,影响其加工与运输[58]。MART1基因敲除的小鼠体系中,小鼠的毛色淡化,黑素细胞中的黑素体形态结构也发生改变[59]。GPNMB是一种糖基化的跨膜蛋白,与PMEL的氨基酸序列具有高同源性且在黑素体形成的整个过程中表达,将GPNMB在黑素细胞中敲除后黑素细胞中黑素体减少,表明GPNMB可促进早期黑素体的形成[60]。
在黑素体合成后期(III-IV期)中,TYRP1和TYR从循环内体(recycling endosome)到黑素体中的运输有两条不同的路径,TYR依赖于AP-3运输,而TYPR1依赖于BLOC-1或BLOC-2运输[61]。在该过程中,Mahanty等[62]研究表明,Rab9A、BLOC-3、Rab32/38或VARP通过调节STX13介导的循环内体来控制TYRP1和TYR在黑素体中的转运从而影响黑素体的成熟。在黑素体成熟过程中,OCA2、SLC45A2、SLC24A5基因编码的蛋白维持着黑素体内的环境稳态。TYR在pH为中性时活性最高,有利于黑色素的合成[63]。OCA2和SLC45A2基因编码的蛋白均作为一个Na+/H+交换器,将H+从黑素体中泵出去,将Na+泵进来维持体内的pH保持在中性。SLC24A5编码的离子泵偶联V-ATPase可调控黑素体内的阳离子浓度(K+/Na+/Ca+/H+)[64](图1)。
1.3黑色素合成的调控
黑素皮质受体1(MC1R)属于G蛋白偶联受体家族,主要在黑素细胞中表达,是调节哺乳动物毛色的重要受体。MC1R通过与α-MSH和ACTH结合,激活细胞膜上腺苷酸环化酶系统使三磷酸腺苷(ATP)转变为环腺苷酸(cAMP);之后MITF启动子区的cAMP反应元件(cAMP-response element, CRE)与cAMP效应元件结合蛋白(cAMP response element-binding protein, CREB)结合而上调MITF基因的表达,最终激活TYR而促进真黑素产生[65]。此外,角质细胞由于UVB照射导致的DNA损伤可激活P53反应机制而上调促黑皮质激素基因(Pro-opiomelanocortin,POMC)的转录表达。该基因是α-MSH和ATCH的前体,α-MSH与MC1R结合激活cAMP,进一步激活PKA导致MITF表达上调最终促进黑色素的合成[66]。ASIP是MC1R的拮抗剂,ASIP可与α-MSH竞争性地结合MC1R而抑制TYR表达,从而阻碍真黑素的合成而激活褐黑素合成[67]。酪氨酸酶TYR是黑色素合成代谢过程中的关键酶,它能催化酪氨酸羟基化为多巴并随后形成DQ。哺乳动物的毛色类型与真黑素和褐黑素二者的比例有关,多巴醌是生成这两种黑色素的共同前体物,因此TYR控制着黑色素的生成,在毛色形成过程中扮演着不可缺少的角色。除了TYR,其酪氨酸酶基因家族还有TYRP1和TYRP2,二者在催化真黑素合成过程中也扮演着重要角色,催化多巴醌进一步合成真黑素[68](图1)。
1.4黑色素的胞外转运调控
黑素体成熟后被转移至表皮或毛囊中的角质细胞中,以此吸收紫外线保护细胞核免受损害[69]。目前,成熟黑素体的转运主要有4种模型和假说:(1)吞噬作用模型中,黑素细胞通过延长树突与角质细胞接触,角质细胞将黑素细胞的树突包围、掐断并吞噬;(2)膜融合模型中,黑素细胞丝状伪足与角质细胞膜融合形成瞬时膜导管,黑素体通过该瞬时膜导管转移至角质细胞中;(3)脱落-吞噬作用模型中,黑素体沿着黑素细胞伸出的丝状伪足迁移至顶端聚集成色素小球并脱落,之后角质细胞将色素小球包裹并吞噬;(4)胞吐-内吞模型中,黑素体膜与黑素细胞质膜融合被分泌至细胞间隙,随后周围的角质细胞通过吞噬作用将黑素体吞噬内化[70]。角质细胞中黑素体的数量、分布能影响毛发、皮肤的色素沉着。RAB27A、MYO5A、MLPH等基因编码的蛋白复合物是黑素体转运所必需的,可协调黑素体沿微管的长距离转运和沿肌动蛋白丝的短距离转运,并在随后的色素沉着中也起到关键作用[71,72],这些基因的遗传突变均会导致小鼠的毛色被稀释[73]。在模型(1)、(2)和(4)中,黑素体的转移均需要角质细胞的吞噬完成,该作用受蛋白酶激活受体2 (recombinant protease activated receptor 2,Par-2)调节,Par-2的活化可增强吞噬作用和皮肤色素沉着[74]。1.5miRNA调节黑色素合成
除毛色相关基因调节哺乳动物的色素化过程外,研究发现miRNA的表达模式也与黑色素合成有关[21]。miRNA是一类长约22 nt的非编码RNA,它们能与细胞中其他因子结合形成沉默复合体(RNA- induced silencing complex, RISC),引起靶基因RNA的降解或翻译抑制而表现出细胞功能活性[75]。miRNA不仅可以通过与Wnt/β-catenin信号通路、TGF-β信号通路和cAMP信号通路中关键基因RNA结合而调控黑色素沉积,还能够通过调控MITF、TYR、DCT等基因表达模式而影响黑色素合成代谢[21,26]。2毛色适应性进化的遗传基础
2.1毛色表型与伪装行为的适应性进化
一般而言,哺乳动物成年个体的毛色是相对稳定的,不会出现明显的变化,但也有特例。由于真黑素/褐黑素的含量变化,使其毛色能够随着季节而变化。例如雪鞋兔(Lepus americanus)为了适应冬季的冰雪环境以此达到躲避天敌的伪装目的,它们在冬季褪去棕色的外衣转变为白色[76]。随着全球变暖,部分地区降雪减少,雪鞋兔为躲避捕食者,它们打破在冬季换装的规律,仍保留其棕色毛发。Jones等[77]研究发现北美雪鞋兔不同毛发颜色的转换与ASIP基因的顺式调控作用相关。白色毛发和棕色毛发中ASIP基因的表达量存在显著差异,白色雪鞋兔5?非编码区存在1Kb左右的碱基插入,而北美雪鞋兔的棕色表型则是黑尾长耳大野兔基因适应性渗透的结果。北极狐(Alopex lagopus)的毛色也能够随季节而变化,在冬季其毛色由棕灰色毛发蜕变为白色毛发以便更好地在雪地里伪装。有****通过白色北极狐和其蓝色变种来研究其毛色变化机制,发现MC1R基因的遗传突变在其毛色调控中发挥了主要作用,其中两个非同义突变位点(p.C5G、p.C280F)影响黑色素的合成水平[78]。非洲条纹鼠(Rhabdomys pumilio)背部有两对深色浅色交替分布的条纹,其背部的条纹可能有利于躲避捕食者[79],研究发现背部的深浅条纹受Alx3调控,Alx3在浅色条纹中高水平表达,Alx3通过与MITF的启动子结合,抑制MITF活性,干扰黑素细胞分化以及黑色素的合成[80]。岩小囊鼠的毛色与其栖息地背景高度匹配,火山熔岩附近区域生活的岩小囊鼠有着深色皮毛,而生活在较远且遍布浅色花岗岩地区的群体其被毛颜色则相对较浅[8]。Nachman等[81]对不同来自栖息地的岩小囊鼠的ASIP和MCIR基因进行测序分析发现,栖息于亚利桑那州中南部Pinacate地区熔岩中的岩小囊鼠其黑色毛发受MC1R基因调控(p.R18C、p.R109W、p.R160W和p.Q233H) (表1)。Table 1
表1
表1毛色表型与适应性进化遗传基础研究总结
Table 1
| 适应性状 | 物种 | 表型特征 | 基因 | 参考文献 | |
|---|---|---|---|---|---|
| 躲避天敌 | 雪鞋兔(Lepus americanus) | 棕色 | ASIP | [76, 77] | |
| 北极狐(Alopex lagopus) | 白色 | MC1R | [78] | ||
| 非洲条纹鼠(Rhabdomys pumilio) | 深浅交替分布的条纹 | Alx3 | [79, 80] | ||
| 岩小囊鼠(Chaetodipus intermedius) | 深色毛发 | MC1R | [8, 81] | ||
| 捕食 | 白灵熊(Ursus americanus kermodei) | 白色 | MC1R | [82, 83] | |
| 非洲猎犬(Lycaon pictus) | 黑色、白色、黄色 | MYO5A、HPS6、PAH | [84] | ||
| 猫科动物(Felidae) | 黑、黄斑纹 | Taqpep、EDN3 | [85, 86] | ||
| 紫外辐射 | 猪(Sus scrofa domesticus) | 藏猪 | 黑色 | MC1R | [16] |
| 山羊(Capra aegagrus hircus) | 西藏绒山羊 | 黑色 | KITLG | [91] | |
| 人工选择 | 猪(Sus scrofa domesticus) | 通城猪 | 两头乌 | MITF、EDNRB | [96] |
| 滇南小耳猪 | 六白 | EDNRB、CNTLN、PINK1 | [97] | ||
| 水牛(Bubalus bubalis) | 白色 | ASIP | [98] | ||
| 白色斑点 | MITF | [99] | |||
| 家犬(Canis lupus familiaris) | 白色、浅黄色、紫貂色 | ASIP | [100, 101] | ||
| 棕色 | TYRP1 | [100, 101] | |||
| 黑色 | CBD103 | [100, 101] | |||
| 毛色稀释 | MLPH | [100, 101] | |||
| 斑点 | SILV、MITF | [100, 101] | |||
| 黄色 | MC1R | [102] | |||
| 家马(Equus ferus caballus) | 黑色、栗色、骝毛 | MC1R、ASIP | [103~ 105] | ||
| 奶酪色、珍珠色、 白斑 | SLC45A2、PMEL、 KIT、EDNRB、MITF | [103~ 105] | |||
| 家兔(Oryctolagus cuniculus f.domesticus) | 灰色 | TYR | [106] | ||
| 獭兔(Oryctolagus cuniculus) | 白色、黑色、棕色、 灰色、灰黄色 | KIT | [111] | ||
| 绵羊(Ovis aries) | 黑色、白色 | TYR、TYRP1 | [107] | ||
| 山羊(Capra aegagrus hircus) | 南江黄羊 | 黄色/黄褐色, 背脊有黑色条带 | RALY、EIF2S2 | [91] | |
| 美姑山羊 | 黑色 | IRF4、EXOC2 | [91] | ||
| 美洲驼(Lama glama) | 白色斑点 | MITF | [108] | ||
| 阿拉伯骆驼(Camelus dromedarius) | 白斑 | KIT | [109] | ||
| 羊驼(Vicugna pacos) | 棕色 | miR-211、miR-184、miR-486、 miR-885、 miR-451、miR-451 | [113] | ||
| 白色 | miR-202、miR-542-5p、miR-424、miR-370、miR-22-3p、miR-143-5p、miR-101a-3p、miR-144a-3p、miR-380-3p | [113~ 116] | |||
| 蓝狐(Alopex lagopus) | 白色 | KIT | [110] | ||
| 驴(Equus asinus) | 黑色、栗色 | TBX3 | [112] | ||
新窗口打开|下载CSV
2.2毛色表型与捕食行为的适应性进化
白灵熊(Ursus americanus kermodei)MC1R的错义突变(p.Y298C)使白灵熊呈现出白色毛发[82],这让它在森林和灌丛中显得格外醒目。但白灵熊主要以鲑鱼为食,白色毛发更能与天空背景融为一体,便于捕食蛙鱼,有报道也证实了在白天白色毛发的白灵熊比黑熊更容易捕食到鲑鱼[83]。非洲野犬(Lycaon pictus)毛色多样,通常由黑色、白色和黄色组成形态各异的斑纹。其多样的毛色可能与伪装有关,有利于其捕食高角羚(Aepyceros melampus)等猎物。Chavez等[84]研究发现其毛色受MYO5A、HPS6和PAH三个基因的调节,其中MYO5A和HPS6通过调节黑素体的转运从而调节黑、白毛色,而PAH通过调节黑色素的合成从而调节黄、黑色毛发。Allen等[85]表明猫科(Felidae)动物的毛色与其生存环境息息相关,其斑点图案有助于在树木、灌木丛中伪装,便于捕食猎物。Kaelin等[86]发现Taqpep和EDN3基因共同调控猫科动物的斑纹形成,Taqpep在毛囊发育前期决定深浅斑纹形成区域,EDN3负责该区域黑、黄毛色的形成(表1)。2.3毛色表型与抵御紫外线辐射的适应性进化
紫外线对皮肤有穿透作用,皮肤在紫外线强烈照射下会导致皮肤变黑、老化、炎症甚至引发皮肤癌[87]。强紫外线照射能诱导环丁烷嘧啶二聚体(cyclobutane pyrimidine dimers, CPD)和嘧啶(6-4)嘧啶酮光产物(6,4-photoproducts,(6-4)PP)的产生而造成DNA损伤[88],也会诱导皮肤中活性氧自由基(ROS)过量产生对细胞及DNA结构造成损伤而导致细胞癌变[89]。对哺乳动物而言毛发中黑色素的沉积是哺乳动物抵御紫外线损伤的第一道防线,黑色素能够减少紫外线对皮肤的穿透作用而有效降低紫外辐射对细胞的损伤[90]。青藏高原高海拔地区有较强的紫外辐射,Li等[16]认为栖息于青藏高原的藏猪的黑色毛发可能用于抵御紫外辐射,避免紫外辐射引发的DNA损伤,作者基于基因组分析从藏猪中筛选出了多个与抵御紫外线相关的适应性进化基因,其中包含与黑色素合成有关的MC1R基因。Guo等[91]通过全基因组分析筛选出调控波尔山羊、美姑山羊、金堂黑山羊、南江黄羊、臧山羊和西藏绒山羊6种山羊毛色、高海拔适应、生长繁殖的正选择基因,其中在西藏绒山羊中筛选出可能调控其黑色毛发的KITLG基因,KITLG可能通过调控黑素细胞的分化和迁移来调控黑色素的生成,从而帮助西藏绒山羊抵御青藏高原强烈的紫外线辐射(表1)。2.4毛色表型与人工选择下的适应性进化
家养动物类群的毛色也是人类高度关注的经济表型性状,家养动物的毛色高度多样化很大程度上反应了人们的喜好差异,或者某些特殊毛色因为和某些关键驯化性状偶联,包括温顺、产崽率以及生长发育速率等而被搭载选择,导致群体/品系固定。例如,敲除Agouti基因的黑色鹿鼠(Peromyscus maniculatus)与正常黄色鹿鼠相比,non-Agouti基因的鹿鼠表现得更温顺,攻击性也较低[92];几乎所有KIT/KITL突变的纯合白斑小鼠都是不育的[93];Avy基因型小鼠有黄色、黑斑、鼠灰色3种表型,而鼠灰色小鼠体重增加速率明显低于其他两种表型[94]。人工选择是家养动物表型进化的主要驱动力,家养动物相较于野生哺乳动物类群往往呈现出了更多的毛色多态性。在中国家猪品系中,有黑色、白色和黑白花色等多种表型,其中黑色家猪在中国较为常见,该毛色也被认为是驯化出来的性状。Li等[95]认为黑色家猪的驯化与中国特殊的祭祀文化有关。与野猪(Sus scrofa)种群相比,中国家猪在瓶颈效应与人工选择共同作用下导致了中国家猪MClR基因区遗传多样性的显著下降,最终使MC1R的两个错义突变(p.V95M和p.L102P)在家猪中固定。除了黑或白的纯色家猪,黑眼圈白毛、乌云盖雪、两头乌等家猪品系均在中国有分布。通过全基因组关联分析发现两头乌群体与非两头乌群体间在MITF和EDNRB基因处有显著信号和遗传差异[96]。对中国滇南小耳猪的全基因组测序和分析,发现其六白毛色表型(四蹄、头和尾尖白色)与EDNRB、CNTLN、PINK1等基因区的遗传突变相关。其中,EDNRB基因上游调控序列中的CEBPB识别基序发生突变,影响了该基因的表达;PINK1基因上一个高度保守的非同义替换位点突变(p.E396D)可能消除PINK1功能,通过介导黑素细胞的凋亡而影响黑色素的合成[97]。与黑色家猪相似,受葬礼文化或宗教的影响,白色水牛(Bubalus bubalis)在中国等亚洲国家也受到特别的偏爱。基于全基因组测序分析发现,ASIP基因上游LINE-1转座子的插入使ASIP基因强表达,使黑色素合成受阻导致水牛表现出白色[98]。Yusnizar等[99]研究发现在水牛中MITF基因突变(c.328C>T;c.840+2T>A)会造成白斑表型。此外,狗(Canis lupus familiaris)是最早被驯化的家养动物,目前已有超过400个不同性状的家犬品系,毛色丰富的家犬受多种色素相关基因如MC1R、TYRP1、ASIP、MLPH和MITF等调控[100,101]。例如昆明犬有狼青、黑背和草黄3种毛色表型,通过对昆明犬MC1R、CBD103和ASIP三个色素调控基因进行测序和进化分析,发现MC1R(p.S90G、p.A105T、p.P159Q、p.M264V)在昆明犬狼青和黑背这两个群体中具有高度杂合性,MC1R可能通过平衡选择作用导致昆明犬的毛色差异[102]。另外,家马(Equus ferus caballus)大约包含900个品种,其黑色、栗色和骝毛等基础毛色主要受MC1R、ASIP基因调节,而奶酪色、珍珠毛色等淡化毛色则由SLC45A2、PMEL17、KIT、EDNRB等相关基因的不同突变类型调节[103?~105]。家兔(Oryctolagus cuniculus f. domesticus)TYR基因3?UTR的缺失使黑色素含量减少,导致灰色毛色[106]。Yao等[107]发现TYR基因和TYPR1基因在黑色绵羊和白色绵羊中表现出显著差异。美洲驼(Lama glama)中MITF基因的突变(c.532T>C)造成白色斑点[108]。阿拉伯骆驼(Camelus dromedarius)KIT中的一个移码突变导致基因转录-翻译提前终止,而出现白斑表型[109]。蓝狐KIT基因中第12内含子的第一个碱基由G突变为T,改变mRNA剪切方式从而导致该基因的第12号外显子缺失,蓝狐出现了显性白色表型[110]。KIT基因在不同毛色的獭兔(Oryctolagus cuniculus)中差异表达,在黑色毛发中表达量最高,并且KIT基因对其他毛色相关基因如TYR、MITF、PMEL和DCT的表达也有影响[111]。野驴的毛色主要为灰色,在人工选择下出现了黑色和栗色的家驴,Wang等[112]基于全基因组数据分析发现家驴的黑色和栗色毛色的出现是由TBX3基因的转录因子结合位点下游约18.6 kb处有1 bp缺失所导致。Guo等[91]通过全基因组分析在南江黄羊(RALY和EIF2S2)和美姑山羊(IRF4和EXOC2)中筛选出了分别调控其黄色/黄褐色、黑色毛发的正选择基因。除了基因突变以外,在羊驼(Vicugna pacos)中,Tian等[113]基于Illumina测序技术对白色羊驼和棕色羊驼皮肤中的miRNA全面分析,发现分别有35个和13个miRNA在白色羊驼和棕色羊驼中高表达。Ji等[114]研究发现miR-143-5p可通过靶向TAK1的3′UTR区域调节TGF-β激活酶1信号通路的表达,抑制TAK1表达从而促进羊驼黑素细胞的增殖和迁移。Zhu等[115]通过实验验证发现miR-101a-3p和miR-144a-3p通过与靶基因MITF的3′UTR结合,调节MITF及其下游基因(TYR、TYRP1和DCT)的表达,抑制黑色素的形成。Liu等[116]研究发现SOX6是miR-380-3p的靶基因,miR-380-3p通过下调SOX6影响β-catenin和MITF的转录和翻译,进而影响下游基因TYR、TYRP1和DCT的表达抑制黑色素的形成(表1)。综上所述,在人工选择过程中,家养动物的毛色受多个基因的调控,但在黑素细胞发育、黑色素合成过程中主要受MITF、KIT、MC1R、ASIP等主效基因调控,此外在黑色素转运过程中相关基因突变(如SLC45A2)也能够导致家养动物毛色改变(表1)。
毛色相关基因的突变一方面能够导致毛色表型呈现多样化,实现对周围生存环境的适应或应对极端的人工选择压力,另一方面,越来越多的研究也发现哺乳动物中一些毛色相关基因的突变往往会偶联着一些比较明显的生理缺陷和病症。比如:在视觉上,TYR基因外显子2的第975位胞嘧啶缺失,导致密码子提前终止使猫(Felis catus)患有白化病,表现为粉红色眼睛和白色毛发[117];日本黑牛(Bos taurus)LYST基因(p.H2015R)突变引起Chediak- Higashi综合征症,表现为眼皮肤白化病,伴有免疫缺陷,并且毛色变浅[118]。在听觉上,斑点狗MITF基因的非编码区突变与先天性神经性耳聋(canine congenital sensorineural deafness, CCSD)有关[119]。毛色相关基因的突变还与癌症偶联,黑色素瘤是由黑素细胞恶变引发的肿瘤,紫外线在黑色素瘤的发展中起着重要作用[81]。灰色马匹的毛色会随着年龄的增长而逐渐变白,该表型与STX17内含子6的4.6 kb重复有关,STX17的突变可能通过调节黑素体的产生或是运输而影响黑色素的沉着,从而导致马毛色变白。体内缺乏黑色素的马匹因而不能有效抵御紫外线的照射,增加了其患黑色素瘤的风险[120,121]。毛色相关基因突变还能够引发皮肤疾病,例如MLPH基因第2外显子的突变会导致杜宾犬毛色被稀释,同时,还伴随着色素稀释性脱毛(color dilution alopecia, CDA),表现为毛发逐渐脱落,并有时伴有毛囊炎[122]。此外,毛色相关基因的突变对哺乳动物的存活也有影响。EDNRB除了参与黑素细胞的发育调控外,在肠神经元发育中也具有重要作用。在家马中,EDNRB基因的突变与致死性白色马驹综合症密切相关,EDNRB基因编码的第118位氨基酸由异亮氨酸变为赖氨酸,会导致患病的小马驹毛色全白或几乎全白,并且在几天内因肠神经节病变而死亡[123]。同样,在小鼠中敲除EDNRB基因会引起小鼠出现白斑表型并伴随着巨结肠[124]。
3结语与展望
目前,关于哺乳动物毛色性状的遗传学和进化生物学研究主要基于全基因组或CDS区上的点突变位点遗传标记来展开。通过对某物种/群体的基因组进行测序分析,筛选出单核苷酸多态性(single nucleotide polymorphisms,SNPs)位点进行保守性分析、功能富集等推测出影响该物种毛色变化的毛色相关基因。但对基于小片段插入或缺失(insertion-deletions, Indels)或大片段的基因组结构性变异(structural variations, SVs)展开的毛色多态性分子机制研究则相对较少;同时,大多数的研究都主要集中于模式物种类群,迄今为止在小鼠中已经发现有171个毛色相关基因(378个突变位点) (随着全基因组测序技术的不断发展和普及,针对模式和非模式物种,基于更多种类分子标记如插入缺失、拷贝数变异(copy number variations, CNVs)与基因组结构性变异的比较基因组学或群体基因组学研究将是哺乳动物毛色多样化研究的重要发展方向;此外,基因组编辑(如CRISPR/Cas9)等技术的日趋完善和成熟,有望从细胞水平或个体水平来深入探究候选基因不同突变类型在细胞层面或个体层面导致的细胞色素化或毛色差异,而不再仅局限于分析某不同毛色的物种中毛色相关基因在其组织中的相对表达量差异,可通过编辑细胞对非模式物种的毛色相关基因进行研究。
目前,对哺乳动物的毛色研究主要体现在两方面:从进化生物学角度探究哺乳动物毛色形成模式的作用和从分子生物学角度探究哺乳动物毛色形成的分子机制,但将二者结合进行研究的相对较少。我们既需要在已知哺乳动物毛色进化意义的情况下,进一步从分子水平层面探究其分子机制(如岩小囊鼠[81]);也需要在已知其分子机制情况下探究其毛色形成的演化意义(如黑色家猪[95])。
此外,人工选择在促进了家养动物毛色多样性的同时,也伴随着部分生理缺陷或疾病的产生,导致出现视觉障碍、听觉障碍、癌症等[117,119,121]。而在人类中,也发现了由毛色相关基因突变引发的类似疾病,比如TYR基因突变常引起眼皮肤白化病I型(OCA1),表现为皮肤、头发和眼睛中的黑色素减少或者丧失,还会伴随着眼球震颤、视力下降的症状[106]。其他毛色相关基因如OCA2、TYPR1和SLC45A2能分别导致眼皮肤白化病II、III和IV型的产生[125?~127]。因此,对不同哺乳动物类群毛色多态性分子调控机制的揭示,也能够为人们探究人群中某些重大遗传疾病的发病机理提供重要线索和启发。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1038/cr.2017.32URLPMID:28281538 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:22997338 [本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
URLPMID:31138092 [本文引用: 1]
[本文引用: 1]
DOI:10.1111/mec.12935URLPMID:25251393 [本文引用: 1]

Genetic colour polymorphisms are widespread across animals and often subjected to complex selection regimes. Traditionally, colour morphs were used as simple visual markers to measure allele frequency changes in nature, selection, population divergence and speciation. With advances in sequencing technology and analysis methods, several model systems are emerging where the molecular targets of selection are being described. Here, we discuss recent studies on the genetics of sexually selected colour polymorphisms, aiming at (i) reviewing the evidence of sexual selection on colour polymorphisms, (ii) highlighting the genetic architecture, molecular and developmental basis underlying phenotypic colour diversification and (iii) discuss how the maintenance of such polymorphisms might be facilitated or constrained by these. Studies of the genetic architecture of colour polymorphism point towards the importance of tight clustering of colour loci with other trait loci, such as in the case of inversions and supergene structures. Other interesting findings include linkage between colour loci and mate preferences or sex determination, and the role of introgression and regulatory variation in fuelling polymorphisms. We highlight that more studies are needed that explicitly integrate fitness consequences of sexual selection on colour with the underlying molecular targets of colour to gain insights into the evolutionary consequences of sexual selection on polymorphism maintenance.
[本文引用: 1]
[本文引用: 1]
URLPMID:24162736 [本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 4]
[本文引用: 4]
[本文引用: 2]
[本文引用: 2]
URLPMID:23433358 [本文引用: 1]
DOI:10.1034/j.1600-0749.2001.140105.xURLPMID:11277491 [本文引用: 1]
Melanocytes produce two chemically distinct types of melanin pigments, eumelanin and pheomelanin. These pigments can be quantitatively analyzed by acidic permanganate oxidation or reductive hydrolysis with hydriodic acid to form pyrrole-2,3,5-tricarboxylic acid or aminohydroxyphenylalanine, respectively. About 30 coat color genes in mice have been cloned, and functions of many of those genes have been elucidated. However, little is known about the interacting functions of these loci. In this study, we used congenic mice to eliminate genetic variability, and analyzed eumelanin and pheomelanin contents of hairs from mice mutant at one or more of the major pigment loci, i.e., the albino (C) locus that encodes tyrosinase, the slaty (Slt) locus that encodes tyrosinase-related protein 2 (TRP2 also known as dopachrome tautomerase, DCT), the brown (B) locus that encodes TRP1, the silver (Si) locus that encodes a melanosomal silver protein, the agouti (A) locus that encodes agouti signaling protein (ASP), the extension (E) locus that encodes melanocortin-1 receptor, and the mahogany (Mg) locus that encodes attractin. We also measured total melanin contents after solubilization of hairs in hot Soluene-350 plus water. Hairs were shaved from 2-3-month-old congenic C57BL/6J mice. The chinchilla (c(ch)) allele is known to encode tyrosinase, whose activity is about one third that of wild type (C). Phenotypes of chinchilla (c(ch)/c(ch)) mice that are wild type or mutant at the brown and/or slaty, loci indicate that functioning TRP2 and TRP1 are necessary, in addition to high levels of tyrosinase, for a full production of eumelanin. The chinchilla allele was found to reduce the amount of pheomelanin in lethal yellow and recessive yellow mice to less than one fifth of that in congenic yellow mice that were wild type at the albino locus. This indicates that reduction in tyrosinase activity affects pheomelanogenesis more profoundly compared with eumelanogenesis. Hairs homozygous for mutation at the slaty locus contain 5,6-dihydroxyindole-2-carboxylic acid (DHICA)-poor melanin, and this chemical phenotype was retained in hairs that were mutant at both the brown locus and the slaty locus. Hair from mice mutant at the brown locus, but not at the slaty locus, do not contain DHICA-poor melanin. This indicates that the proportion of DHICA in eumelanin is determined by TRP2, but not by TRP1. Mutation at the slaty locus (Slt(lt)) was found to have no effect on pheomelanogenesis, supporting a role of TRP2 only in eumelanogenesis. The mutation at silver (si) locus showed an effect similar to brown, a partial suppression of eumelanogenesis. The mutation at mahogany (mg) locus partially suppressed the effect of lethal yellow (Ay) on pheomelanogenesis, supporting a role of mahogany in interfering with agouti signaling. These results show that combination of double mutation study of congenic mice with chemical analysis of melanins is useful in evaluating the interaction of pigment gene functions.
[本文引用: 2]
[本文引用: 3]
[本文引用: 3]
URLPMID:21443614 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.0022-202X.2004.23528.xURLPMID:15654948 [本文引用: 2]
Hair shaft melanin components (eu- or/and pheomelanin) are a long-lived record of precise interactions in the hair follicle pigmentary unit, e.g., between follicular melanocytes, keratinocytes, and dermal papilla fibroblasts. Follicular melanogenesis (FM) involves sequentially the melanogenic activity of follicular melanocytes, the transfer of melanin granules into cortical and medulla keratinocytes, and the formation of pigmented hair shafts. This activity is in turn regulated by an array of enzymes, structural and regulatory proteins, transporters, and receptors and their ligands, acting on the developmental stages, cellular, and hair follicle levels. FM is stringently coupled to the anagen stage of the hair cycle, being switched-off in catagen to remain absent through telogen. At the organ level FM is precisely coupled to the life cycle of melanocytes with changes in their compartmental distribution and accelerated melanoblast/melanocyte differentiation with enhanced secretory activity. The melanocyte compartments in the upper hair follicle also provides a reservoir for the repigmentation of epidermis and, for the cyclic formation of new anagen hair bulbs. Melanin synthesis and pigment transfer to bulb keratinocytes are dependent on the availability of melanin precursors, and regulation by signal transduction pathways intrinsic to skin and hair follicle, which are both receptor dependent and independent, act through auto-, para- or intracrine mechanisms and can be modified by hormonal signals. The important regulators are MC1 receptor its and adrenocorticotropic hormone, melanocyte stimulating hormone, agouti protein ligands (in rodents), c-Kit, and the endothelin receptors with their ligands. Melanin itself has a wide range of bioactivities that extend far beyond its determination of hair color.
DOI:10.1007/s00018-012-1112-4URLPMID:22915137 [本文引用: 1]
Melanoblasts are a particular type of cell that displays extensive cellular proliferation during development to contribute to the skin. There are only a few melanoblast founders, initially located just dorsal to the neural tube, and they sequentially colonize the dermis, epidermis, and hair follicles. In each compartment, melanoblasts are exposed to a wide variety of developmental cues that regulate their expansion. The colonization of the dermis and epidermis by melanoblasts involves substantial proliferation to generate thousands of cells or more from a few founders within a week of development. This review addresses the cellular and molecular events occurring during melanoblast development. We focus on intrinsic and extrinsic factors that control melanoblast proliferation. We also present a robust mathematical model for estimating the doubling-time of dermal and epidermal melanoblasts for all coat color phenotypes from black to white.
[本文引用: 1]
URLPMID:28911859 [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:21040502 [本文引用: 1]
DOI:10.1016/j.jdermsci.2013.08.006URLPMID:24016750 [本文引用: 1]
BACKGROUND: Melanin for skin pigmentation is synthesized from tyrosine via an enzymatic cascade that is controlled by tyrosinase (TYR), tyrosinase-related protein 1 (TRP1), and dopachrome tautomerase/tyrosinase related protein 2 (Dct/TRP2), which are the targets of microphthalmia-associated transcription factor (MITF). MITF is a master regulator of pigmentation and a target of beta-catenin in Wnt/beta-catenin signaling during melanocyte differentiation. Stem cells have been used in skin pigmentation studies, but the mechanisms were not determined for the conditioned medium (CM)-mediated effects. OBJECTIVES: In this study, the inhibition and mechanisms of melanin synthesis were elucidated in B16 melanoma cells and UV-B irradiated C57/BL-6 mice that were treated with human neural stem cell-conditioned medium (NSC-CM). METHODS: B16-F10 melanoma cells (1.5x10(4)cells/well) and the shaved dorsal skin of mice were pretreated with various amount (5, 10, 20, 50, and 100%) of NSC-CM. Melanin contents and TYR activity were measured by a Spectramax spectrophotometer. The expression of TYR, TRP1, Dct/TRP2, MITF, beta-catenin and Wnt inhibitors were evaluated by RT-PCR and western blot. The dorsal skin samples were analyzed by immunofluorescence with various antibodies and compared with that control of tissues. RESULTS: Marked decreases were evident in melanin content and TYR, TRP1, DCT/TRP2, MITF, and beta-catenin expression in B16 cells and C57/BL-6 mice. NSC-CM negatively regulated Wnt/beta-catenin signaling by decreasing the expression of beta-catenin protein, which resulted from robust expression of Wnt inhibitors Dickkopf-1 (DKK1) and secreted frizzled-related protein 2 (sFRP2). CONCLUSIONS: These results demonstrate that NSC-CM suppresses melanin production in vitro and in vivo, suggesting that factors in NSC-CM may play an important role in deregulation of epidermal melanogenesis.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:12789276 [本文引用: 1]
[本文引用: 1]
DOI:10.1046/j.0022-202x.2001.00010.xURLPMID:11764295 [本文引用: 2]
Microphthalmia-associated transcription factor (MITF) regulates the differentiation and development of melanocytes and retinal pigment epithelium and is also responsible for pigment cell-specific transcription of the melanogenesis enzyme genes. Heterozygous mutations in the MITF gene cause auditory-pigmentary syndromes. MITF consists of at least five isoforms, MITF-A, MITF-B, MITF-C, MITF-H, and MITF-M, differing at their N-termini and expression patterns. Here we show a remarkable similarity between the N-terminal domain of MITF-A and cytoplasmic retinoic acid-binding proteins. To date, four isoform-specific first exons have been identified in the MITF gene: exons 1A, 1H, 1B, and 1M in the 5' to 3' direction, each of which encodes the unique N-terminus of a given isoform. The 5'-flanking regions of these isoform-specific exons are termed A, H, B, and M promoters, respectively. Among these promoters, the M promoter has received particular attention, because it is functional only in melanocyte-lineage cells and is upregulated by Wnt signaling via the functional LEF-1-binding site. Moreover, the M promoter is upregulated by other transcription factors, PAX3, SOX10, and CREB. The activity and degradation of MITF-M are regulated by extracellular signals via protein phosphorylation, such as c-Kit signaling. Together, multiple signals appear to converge on the M promoter as well as on MITF proteins, leading to the proper regulation of MITF-M in melanocytes and other MITF isoforms in many cell types.
[本文引用: 1]
URLPMID:11076759 [本文引用: 1]
URLPMID:28263292 [本文引用: 2]
[本文引用: 2]
[本文引用: 1]
DOI:10.1016/j.molmed.2006.07.008URLPMID:16899407 [本文引用: 1]
Microphthalmia-associated transcription factor (MITF) acts as a master regulator of melanocyte development, function and survival by modulating various differentiation and cell-cycle progression genes. It has been demonstrated that MITF is an amplified oncogene in a fraction of human melanomas and that it also has an oncogenic role in human clear cell sarcoma. However, MITF also modulates the state of melanocyte differentiation. Several closely related transcription factors also function as translocated oncogenes in various human malignancies. These data place MITF between instructing melanocytes towards terminal differentiation and/or pigmentation and, alternatively, promoting malignant behavior. In this review, we survey the roles of MITF as a master lineage regulator in melanocyte development and its emerging activities in malignancy. Understanding the molecular function of MITF and its associated pathways will hopefully shed light on strategies for improving therapeutic approaches for these diseases.
[本文引用: 1]
URLPMID:30425278 [本文引用: 1]
DOI:10.1016/j.bbrc.2018.07.012URLPMID:30208522 [本文引用: 1]
The functionally specialized melanosome is a membrane-enclosed lysosome-related organelle, which coexists with lysosomes in melanocytes. Pre-melanosomal protein (PMEL) initiates pre-melanosome morphogenesis and is the only cell-specific pigment protein required for the formation of fibrils on which melanin is deposited in melanosomes. But the effects of PMEL on melanin synthesis and lysosome activity remain unclear. In the study, PMEL was silenced in human epidermal melanocytes by siRNA transfection. Compared to the non-treated group, melanin content in the transfected cells was greatly reduced. Real-time qPCR, Western blotting and immunofluorescence analyses all showed that PMEL-siRNA transfection reduced protein level of tyrosinase, a key enzyme in melanogenesis, but it does not affect tyrosinase gene expression. Moreover, in the absence of PMEL, lysosomal activation was manifested by an increase in the number of lysosomes and activity of hydrolysis enzymes. The lysosome inhibitors restored tyrosinase expression after PMEL silencing, indicating that tyrosinase was degradated by lysosomes. The data collectively showed that silencing of PMEL suppressed melanization through activating lysosomes and degradation of tyrosinase by lysosomes. Our findings provide novel insight into the interaction between the melanosome and its related organelle, the lysosome, supplying a new idea for the pathogenesis and clinical treatment of pigmented diseases.
DOI:10.1152/physrev.00059.2017URLPMID:30255724 [本文引用: 1]
Melanosomes are organelles that produce and store melanin, a widespread biological pigment with a unique suite of properties including high refractive index, semiconducting capabilities, material stiffness, and high fossilization potential. They are involved in numerous critical biological functions in organisms across the tree of life. Individual components such as melanin chemistry and melanosome development have recently been addressed, but a broad synthesis is needed. Here, we review the hierarchical structure, development, functions, and evolution of melanosomes. We highlight variation in melanin chemistry and melanosome morphology and how these may relate to function. For example, we review what is known of the chemical differences between different melanin types (eumelanin, pheomelanin, allomelanin) and whether/how melanosome morphology relates to chemistry and color. We integrate the distribution of melanin across living organisms with what is known from the fossil record and produce hypotheses on its evolution. We suggest that melanin was present in life forms early in evolutionary history and that melanosomes evolved at the origin of organelles. Throughout, we discuss the (sometimes gaping) holes in our knowledge and suggest areas that need particular attention as we move forward in our understanding of these still-mysterious organelles and the materials that they contain.
DOI:10.1242/jcs.02804URLPMID:16492709 [本文引用: 1]
Adaptor proteins (AP) play important roles in the sorting of proteins from the trans-Golgi network, but how they function in the sorting of various melanosome-specific proteins such as Pmel17, an essential structural component of melanosomes, in melanocytes is unknown. We characterized the processing and trafficking of Pmel17 via adaptor protein complexes within melanocytic cells. Proteomics analysis detected Pmel17, AP1 and AP2, but not AP3 or AP4 in early melanosomes. Real-time PCR, immunolabeling and tissue in-situ hybridization confirmed the coexpression of AP1 isoforms mu1A and mu1B (expressed only in polarized cells) in melanocytes and keratinocytes, but expression of mu1B is missing in some melanoma cell lines. Transfection with AP1 isoforms (mu1A or mu1B) showed two distinct distribution patterns that involved Pmel17, and only mu1B was able to restore the sorting of Pmel17 to the plasma membrane in cells lacking mu1B expression. Finally, we established that expression of mu1B is regulated physiologically in melanocytes by UV radiation or DKK1. These results show that Pmel17 is sorted to melanosomes by various intracellular routes, directly or indirectly through the plasma membrane, and the presence of basolateral elements in melanocytes suggests their polarized nature.
DOI:10.1371/journal.pgen.1002285URLPMID:21949658 [本文引用: 1]
PMEL is an amyloidogenic protein that appears to be exclusively expressed in pigment cells and forms intralumenal fibrils within early stage melanosomes upon which eumelanins deposit in later stages. PMEL is well conserved among vertebrates, and allelic variants in several species are associated with reduced levels of eumelanin in epidermal tissues. However, in most of these cases it is not clear whether the allelic variants reflect gain-of-function or loss-of-function, and no complete PMEL loss-of-function has been reported in a mammal. Here, we have created a mouse line in which the Pmel gene has been inactivated (Pmel(-)/(-)). These mice are fully viable, fertile, and display no obvious developmental defects. Melanosomes within Pmel(-)/(-) melanocytes are spherical in contrast to the oblong shape present in wild-type animals. This feature was documented in primary cultures of skin-derived melanocytes as well as in retinal pigment epithelium cells and in uveal melanocytes. Inactivation of Pmel has only a mild effect on the coat color phenotype in four different genetic backgrounds, with the clearest effect in mice also carrying the brown/Tyrp1 mutation. This phenotype, which is similar to that observed with the spontaneous silver mutation in mice, strongly suggests that other previously described alleles in vertebrates with more striking effects on pigmentation are dominant-negative mutations. Despite a mild effect on visible pigmentation, inactivation of Pmel led to a substantial reduction in eumelanin content in hair, which demonstrates that PMEL has a critical role for maintaining efficient epidermal pigmentation.
DOI:10.1074/jbc.M413692200URLPMID:15695812 [本文引用: 1]
More than 125 genes that regulate pigmentation have been identified to date. Of those, MART-1 has been widely studied as a melanoma-specific antigen and as a melanosome-specific marker. Whereas the functions of other melanosomal proteins, such as tyrosinase, tyrosinase-related protein-1, dopachrome tautomerase, and Pmel17, are known, the function of MART-1 in melanogenesis, is unclear. A role for MART-1 in pigmentation is expected because its expression pattern and subcellular distribution is quite similar to the other melanosomal proteins and usually correlates with melanin content. We investigated the function of MART-1 using a multidisciplinary approach, including the use of siRNA to inhibit MART-1 function and the use of transfection to re-express MART-1 in MART-1-negative cells. We show that MART-1 forms a complex with Pmel17 and affects its expression, stability, trafficking, and the processing which is required for melanosome structure and maturation. We conclude that MART-1 is indispensable for Pmel17 function and thus plays an important role in regulating mammalian pigmentation.
[本文引用: 1]
DOI:10.1371/journal.pone.0042955URLPMID:22912767 [本文引用: 1]
BACKGROUND: Melanosomes are specialized membrane-surrounded organelles, which are involved in the synthesis, storage and transport of melanin. Glycoprotein (transmembrane) non-metastatic melanoma protein b (GPNMB), a melanosome-specific structural protein, shares significant amino acid sequence homology with Pmel-17. Proteomic analysis demonstrated that GPNMB is present in all stages (I-IV) of melanosomes. However, little is known about the role of GPNMB in melanosomes. METHODOLOGY/PRINCIPAL FINDINGS: Using real-time quantitative PCR, Western blotting and immunofluorescence analysis, we demonstrated that the expression of GPNMB in PIG1 melanocytes was up-regulated by ultraviolet B (UVB) radiation. Transmission electron microscopy analysis showed that the total number of melanosomes in PIG1 melanocytes was sharply reduced by GPNMB-siRNA transfection. Simultaneously, the expression levels of tyrosinase (Tyr), tyrosinase related protein 1 (Trp1), Pmel17/gp100 and ocular albinism type 1 protein (OA1) were all significantly attenuated. But the expression of microphthalmia-associated transcription factor (MITF) was up-regulated. Intriguingly, in GPNMB silenced PIG1 melanocytes, UVB radiation sharply reduced MITF expression. CONCLUSION: Our present work revealed that the GPNMB was critical for the formation of melanosomes. And GPNMB expression down-regulation attenuated melanosome formation in a MITF-independent fashion.
[本文引用: 1]
DOI:10.1111/pcmr.12434URLPMID:26527546 [本文引用: 1]
Melanosomes are a type of lysosome-related organelle that is commonly defective in Hermansky-Pudlak syndrome. Biogenesis of melanosomes is regulated by BLOC-1, -2, -3, or AP-1, -3 complexes, which mediate cargo transport from recycling endosomes to melanosomes. Although several Rab GTPases have been shown to regulate these trafficking steps, the precise role of Rab9A remains unknown. Here, we found that a cohort of Rab9A associates with the melanosomes and its knockdown in melanocytes results in hypopigmented melanosomes due to mistargeting of melanosomal proteins to lysosomes. In addition, the Rab9A-depletion phenotype resembles Rab38/32-inactivated or BLOC-3-deficient melanocytes, suggesting that Rab9A works in line with BLOC-3 and Rab38/32 during melanosome cargo transport. Furthermore, silencing of Rab9A, Rab38/32 or its effector VARP, or BLOC-3-deficiency in melanocytes decreased the length of STX13-positive recycling endosomal tubules and targeted the SNARE to lysosomes. This result indicates a defect in directing recycling endosomal tubules to melanosomes. Thus, Rab9A and its co-regulatory GTPases control STX13-mediated cargo delivery to maturing melanosomes.
DOI:10.1006/excr.2001.5251URLPMID:11461115 [本文引用: 1]
The skin pigment melanin is produced in melanocytes in highly specialized organelles known as melanosomes. Melanosomes are related to the organelles of the endosomal/lysosomal pathway and can have a low internal pH. In the present study we have shown that melanin synthesis in human pigment cell lysates is maximal at pH 6.8. We therefore investigated the role of intramelanosomal pH as a possible control mechanism for melanogenesis. To do this we examined the effect of neutralizing melanosomal pH on tyrosinase activity and melanogenesis in 11 human melanocyte cultures and in 3 melanoma lines. All melanocyte cultures (9 of 9) from Caucasian skin as well as two melanoma cell lines with comparable melanogenic activity showed rapid (within 24 h) increases in melanogenesis in response to neutralization of melanosomal pH. Chemical analysis of total melanin indicated a preferential increase in eumelanin production. Electron microscopy revealed an accumulation of melanin and increased maturation of melanosomes in response to pH neutralization. In summary, our findings show that: (i) near neutral melanosomal pH is optimal for human tyrosinase activity and melanogenesis; (ii) melanin production in Caucasian melanocytes is suppressed by low melanosomal pH; (iii) the ratio of eumelanin/phaeomelanin production and maturation rate of melanosomes can be regulated by melanosomal pH. We conclude that melanosomal pH is an essential factor which regulates multiple stages of melanin production. Furthermore, since we have recently identified that pink locus product (P protein) mediates neutralization of melanosomal pH, we propose that P protein is a key control point for skin pigmentation. We would further propose that the wide variations in both constitutive and facultative skin pigmentation seen in the human population could be associated with the high degree of P-locus polymorphism.
DOI:10.1074/jbc.M707521200URLPMID:18166528 [本文引用: 1]
A non-synonymous single nucleotide polymorphism in the human SLC24A5 gene is associated with natural human skin color variation. Multiple sequence alignments predict that this gene encodes a member of the potassium-dependent sodium-calcium exchanger family denoted NCKX5. In cultured human epidermal melanocytes we show using affinity-purified antisera that native human NCKX5 runs as a triplet of approximately 43 kDa on SDS-PAGE and is partially localized to the trans-Golgi network. Removal of the NCKX5 protein through small interfering RNA-mediated knockdown disrupts melanogenesis in human and murine melanocytes, causing a significant reduction in melanin pigment production. Using a heterologous expression system, we confirm for the first time that NCKX5 possesses the predicted exchanger activity. Site-directed mutagenesis of NCKX5 and NCKX2 in this system reveals that the non-synonymous single nucleotide polymorphism in SLC24A5 alters a residue that is important for NCKX5 and NCKX2 activity. We suggest that NCKX5 directly regulates human epidermal melanogenesis and natural skin color through its intracellular potassium-dependent exchanger activity.
DOI:10.1016/j.bbadis.2017.02.027URLPMID:28259754 [本文引用: 1]
The melanocortin-1 receptor (MC1R) preferentially expressed in melanocytes is best known as a key regulator of the synthesis of epidermal melanin pigments. Its paracrine stimulation by keratinocyte-derived melanocortins also activates DNA repair pathways and antioxidant defenses to build a complex, multifaceted photoprotective response. Many MC1R actions rely on cAMP-dependent activation of two transcription factors, MITF and PGC1alpha, but pleiotropic MC1R signaling also involves activation of mitogen-activated kinases and AKT. MC1R partners such as beta-arrestins, PTEN and the E3 ubiquitin ligase MGRN1 differentially regulate these pathways. The MC1R gene is complex and polymorphic, with frequent variants associated with skin phenotypes and increased cancer risk. We review current knowledge of signaling from canonical MC1R, its splice isoforms and natural polymorphic variants. Recently discovered intracellular targets and partners are also discussed, to highlight the diversity of mechanisms that may contribute to normal and pathological variation of pigmentation and sensitivity to solar radiation-induced damage. This article is part of a Special Issue entitled: Melanocortin Receptors - edited by Ya-Xiong Tao.
DOI:10.3390/ijms140612222URLPMID:23749111 [本文引用: 1]
UV radiation (UV) is classified as a
DOI:10.1093/molbev/msm134URLPMID:17609536 [本文引用: 1]
We have characterized the biochemical function of the melanocortin 1 receptor (MC1R), a critical regulator of melanin synthesis, from 9 phylogenetically diverse primate species with varying coat colors. There is substantial diversity in melanocyte-stimulating hormone (MSH) binding affinity and basal levels of activity in the cloned MC1Rs. MSH binding was lost independently in lemur and New World monkey lineages, whereas high basal levels of MC1R activity occur in lemurs and some New World monkeys and Old World monkeys. Highest levels of basal activity were found in the MC1R of ruffed lemurs, which have the E94K mutation that leads to constitutive activation in other species. In 3 species (2 lemurs and the howler monkey), we report the novel finding that binding and inhibition of MC1R by agouti signaling protein (ASIP) can occur when MSH binding has been lost, thus enabling continuing regulation of the melanin type via ASIP expression. Together, these findings can explain the previous paradox of a predominantly pheomelanic coat in the red ruffed lemur (Varecia rubra). The presence of a functional, MSH-responsive MC1R in orangutan demonstrates that the mechanism of red hair generation in this ape is different from the prevalent mechanism in European human populations. Overall, we have found unexpected diversity in MC1R function among primates and show that the evolution of the regulatory control of MC1R activity occurs by independent variation of 3 distinct mechanisms: basal MC1R activity, MSH binding and activation, and ASIP binding and inhibition. This diversity of function is broadly associated with primate phylogeny and does not have a simple relation to coat color phenotype within primate clades.
DOI:10.1111/ics.12466URLPMID:29752874 [本文引用: 1]
In human skin, melanogenesis is a tightly regulated process. Indeed, several extracellular signals are transduced via dedicated signalling pathways and mostly converge to MITF, a transcription factor integrating upstream signalling and regulating downstream genes involved in the various inherent mechanisms modulating melanogenesis. The synthesis of melanin pigments occurs in melanocytes inside melanosomes where melanogenic enzymes (tyrosinase and related proteins) are addressed with the help of specific protein complexes. The melanosomes loaded with melanin are then transferred to keratinocytes. A more elaborate level of melanogenesis regulation comes into play via the action of non-coding RNAs (microRNAs, lncRNAs). Besides this canonical regulation, melanogenesis can also be modulated by other non-specific intrinsic pathways (hormonal environment, inflammation) and by extrinsic factors (solar irradiation such as ultraviolet irradiation, environmental pollution). We developed a bioinformatic interaction network gathering the multiple aspects of melanogenesis and skin pigmentation as a resource to better understand and study skin pigmentation biology.
[本文引用: 1]
DOI:10.1016/j.ceb.2014.02.003URL [本文引用: 1]
The pigmentation of skin and hair in mammals is driven by the creation within melanocytes of melanosomes, a specialized pigment-producing organelle, and the subsequent intercellular transfer of this organelle to keratinocytes. This latter process is absolutely required for visible pigmentation and effective photo-protection because it serves to disperse the pigment in skin and hair. Therefore, the transfer of melanosomes from the melanocyte to the keratinocyte is as important for the biological endpoint of mammalian pigmentation as the biogenesis of this fascinating organelle. Here we review new findings that shed light on, and raise additional questions about, the mechanism of this enigmatic process.
[本文引用: 1]
DOI:10.1371/journal.pone.0041160URLPMID:22844437 [本文引用: 1]
Human skin color is predominantly determined by melanin produced in melanosomes within melanocytes and subsequently distributed to keratinocytes. There are many studies that have proposed mechanisms underlying ethnic skin color variations, whereas the processes involved from melanin synthesis in melanocytes to the transfer of melanosomes to keratinocytes are common among humans. Apart from the activities in the melanogenic rate-limiting enzyme, tyrosinase, in melanocytes and the amounts and distribution patterns of melanosomes in keratinocytes, the abilities of the actin-associated factors in charge of melanosome transport within melanocytes also regulate pigmentation. Mutations in genes encoding melanosome transport-related molecules, such as MYO5A, RAB27A and SLAC-2A, have been reported to cause a human pigmentary disease known as Griscelli syndrome, which is associated with diluted skin and hair color. Thus we hypothesized that process might play a role in modulating skin color variations. To address that hypothesis, the correlations of expression of RAB27A and its specific effector, SLAC2-A, to melanogenic ability were evaluated in comparison with tyrosinase, using human melanocytes derived from 19 individuals of varying skin types. Following the finding of the highest correlation in RAB27A expression to the melanogenic ability, darkly-pigmented melanocytes with significantly higher RAB27A expression were found to transfer significantly more melanosomes to keratinocytes than lightly-pigmented melanocytes in co-culture and in human skin substitutes (HSSs) in vivo, resulting in darker skin color in concert with the difference observed in African-descent and Caucasian skins. Additionally, RAB27A knockdown by a lentivirus-derived shRNA in melanocytes concomitantly demonstrated a significantly reduced number of transferred melanosomes to keratinocytes in co-culture and a significantly diminished epidermal melanin content skin color intensity (DeltaL* = 4.4) in the HSSs. These data reveal the intrinsically essential role of RAB27A in human ethnic skin color determination and provide new insights for the fundamental understanding of regulatory mechanisms underlying skin pigmentation.
DOI:10.1002/biof.29URLPMID:19449448 [本文引用: 1]
More than 150 genes have been identified that affect skin color either directly or indirectly, and we review current understanding of physiological factors that regulate skin pigmentation. We focus on melanosome biogenesis, transport and transfer, melanogenic regulators in melanocytes, and factors derived from keratinocytes, fibroblasts, endothelial cells, hormones, inflammatory cells, and nerves. Enzymatic components of melanosomes include tyrosinase, tyrosinase-related protein 1, and dopachrome tautomerase, which depend on the functions of OA1, P, MATP, ATP7A, and BLOC-1 to synthesize eumelanins and pheomelanins. The main structural component of melanosomes is Pmel17/gp100/Silv, whose sorting involves adaptor protein 1A (AP1A), AP1B, AP2, and spectrin, as well as a chaperone-like component, MART-1. During their maturation, melanosomes move from the perinuclear area toward the plasma membrane. Microtubules, dynein, kinesin, actin filaments, Rab27a, melanophilin, myosin Va, and Slp2-a are involved in melanosome transport. Foxn1 and p53 up-regulate skin pigmentation via bFGF and POMC derivatives including alpha-MSH and ACTH, respectively. Other critical factors that affect skin pigmentation include MC1R, CREB, ASP, MITF, PAX3, SOX9/10, LEF-1/TCF, PAR-2, DKK1, SCF, HGF, GM-CSF, endothelin-1, prostaglandins, leukotrienes, thromboxanes, neurotrophins, and neuropeptides. UV radiation up-regulates most factors that increase melanogenesis. Further studies will elucidate the currently unknown functions of many other pigment genes/proteins. (c) 2009 International Union of Biochemistry and Molecular Biology, Inc.
DOI:10.1016/j.gde.2017.05.001URLPMID:28605672 [本文引用: 1]
The intercellular transfer of the melanin-producing organelle, called melanosome, from melanocytes to adjacent keratinocytes, is largely responsible for the coat colors and skin pigmentation of amniotes (birds, reptiles, and mammals). Although several hypotheses of melanin-transfer were proposed mainly by in vitro studies and electron microscopies, how the melanosome transfer takes place in the actual skin remained unclear. With advances in technologies of gene manipulations and high-resolution microscopy that allow direct visualization of plasma membrane, we are beginning to understand the amazing behaviors and dynamics of melanocytes. Studies in melanosome transfer further provide a clue to understand a general principle of intercellular organelle transport, including the intercellular translocations of mitochondria.
[本文引用: 1]
[本文引用: 1]
DOI:10.1126/science.aan8097URLPMID:29449510 [本文引用: 2]
Maintenance of biodiversity in a rapidly changing climate will depend on the efficacy of evolutionary rescue, whereby population declines due to abrupt environmental change are reversed by shifts in genetically driven adaptive traits. However, a lack of traits known to be under direct selection by anthropogenic climate change has limited the incorporation of evolutionary processes into global conservation efforts. In 21 vertebrate species, some individuals undergo a seasonal color molt from summer brown to winter white as camouflage against snow, whereas other individuals remain brown. Seasonal snow duration is decreasing globally, and fitness is lower for winter white animals on snowless backgrounds. Based on 2713 georeferenced samples of known winter coat color-from eight species across trophic levels-we identify environmentally driven clinal gradients in winter coat color, including polymorphic zones where winter brown and white morphs co-occur. These polymorphic zones, underrepresented by existing global protected area networks, indicate hot spots for evolutionary rescue in a changing climate.
DOI:10.1126/science.aar5273URLPMID:29930138 [本文引用: 2]
Snowshoe hares (Lepus americanus) maintain seasonal camouflage by molting to a white winter coat, but some hares remain brown during the winter in regions with low snow cover. We show that cis-regulatory variation controlling seasonal expression of the Agouti gene underlies this adaptive winter camouflage polymorphism. Genetic variation at Agouti clustered by winter coat color across multiple hare and jackrabbit species, revealing a history of recurrent interspecific gene flow. Brown winter coats in snowshoe hares likely originated from an introgressed black-tailed jackrabbit allele that has swept to high frequency in mild winter environments. These discoveries show that introgression of genetic variants that underlie key ecological traits can seed past and ongoing adaptation to rapidly changing environments.
DOI:10.1016/j.peptides.2004.11.040URLPMID:15982782 [本文引用: 2]
We have characterized two mutations in the MC1R gene of the blue variant of the arctic fox (Alopex lagopus) that both incorporate a novel cysteine residue into the receptor. A family study in farmed arctic foxes verified that the dominant expression of the blue color phenotype cosegregates completely with the allele harboring these two mutations. Additionally to the altered pigment synthesis, the blue fox allele suppresses the seasonal change in coat color found in the native arctic fox. Consequently, these findings suggest that the MC1R/agouti regulatory system is involved in the seasonal changes of coat color found in arctic fox.
DOI:10.1016/j.cub.2018.02.009URLPMID:29614283 [本文引用: 2]
Mallarino et al. introduce the African striped mouse, which is being used in a number of fields of research, including animal behavior, evolutionary developmental biology, and chronobiology.
DOI:10.1038/nature20109URLPMID:27806375 [本文引用: 2]
Mammalian colour patterns are among the most recognizable characteristics found in nature and can have a profound impact on fitness. However, little is known about the mechanisms underlying the formation and subsequent evolution of these patterns. Here we show that, in the African striped mouse (Rhabdomys pumilio), periodic dorsal stripes result from underlying differences in melanocyte maturation, which give rise to spatial variation in hair colour. We identify the transcription factor ALX3 as a regulator of this process. In embryonic dorsal skin, patterned expression of Alx3 precedes pigment stripes and acts to directly repress Mitf, a master regulator of melanocyte differentiation, thereby giving rise to light-coloured hair. Moreover, Alx3 is upregulated in the light stripes of chipmunks, which have independently evolved a similar dorsal pattern. Our results show a previously undescribed mechanism for modulating spatial variation in hair colour and provide insights into how phenotypic novelty evolves.
[本文引用: 4]
DOI:10.1016/S0960-9822(01)00448-1URL [本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOI:10.1098/rspb.2010.1734URLPMID:20961899 [本文引用: 2]
A complete explanation of the diversity of animal colour patterns requires an understanding of both the developmental mechanisms generating them and their adaptive value. However, only two previous studies, which involved computer-generated evolving prey, have attempted to make this link. This study examines variation in the camouflage patterns displayed on the flanks of many felids. After controlling for the effects of shared ancestry using a fully resolved molecular phylogeny, this study shows how phenotypes from plausible felid coat pattern generation mechanisms relate to ecology. We found that likelihood of patterning and pattern attributes, such as complexity and irregularity, were related to felids' habitats, arboreality and nocturnality. Our analysis also indicates that disruptive selection is a likely explanation for the prevalence of melanistic forms in Felidae. Furthermore, we show that there is little phylogenetic signal in the visual appearance of felid patterning, indicating that camouflage adapts to ecology over relatively short time scales. Our method could be applied to any taxon with colour patterns that can reasonably be matched to reaction-diffusion and similar models, where the kinetics of the reaction between two or more initially randomly dispersed morphogens determines the outcome of pattern development.
DOI:10.1126/science.1220893URLPMID:22997338 [本文引用: 2]
Color markings among felid species display both a remarkable diversity and a common underlying periodicity. A similar range of patterns in domestic cats suggests a conserved mechanism whose appearance can be altered by selection. We identified the gene responsible for tabby pattern variation in domestic cats as Transmembrane aminopeptidase Q (Taqpep), which encodes a membrane-bound metalloprotease. Analyzing 31 other felid species, we identified Taqpep as the cause of the rare king cheetah phenotype, in which spots coalesce into blotches and stripes. Histologic, genomic expression, and transgenic mouse studies indicate that paracrine expression of Endothelin3 (Edn3) coordinates localized color differences. We propose a two-stage model in which Taqpep helps to establish a periodic pre-pattern during skin development that is later implemented by differential expression of Edn3.
[本文引用: 1]
DOI:10.1111/j.1600-0749.2006.00358.xURLPMID:17250543 [本文引用: 1]
Pigmentation of human skin is closely involved in protection against environmental stresses, in particular exposure to ultraviolet (UV) radiation. It is well known that darker skin is significantly more resistant to the damaging effects of UV, such as photocarcinogenesis and photoaging, than is lighter skin. Constitutive skin pigmentation depends on the amount of melanin and its distribution in that tissue. Melanin is significantly photoprotective and epidermal cells in darker skin incur less DNA damage than do those in lighter skin. This review summarizes current understanding of the regulation of constitutive human skin pigmentation and responses to UV radiation, with emphasis on physiological factors that influence those processes. Further research is needed to characterize the role of skin pigmentation to reduce photocarcinogenesis and to develop effective strategies to minimize such risks.
DOI:10.3390/biom5020545URLPMID:25906193 [本文引用: 1]
Oxidative stress in skin plays a major role in the aging process. This is true for intrinsic aging and even more for extrinsic aging. Although the results are quite different in dermis and epidermis, extrinsic aging is driven to a large extent by oxidative stress caused by UV irradiation. In this review the overall effects of oxidative stress are discussed as well as the sources of ROS including the mitochondrial ETC, peroxisomal and ER localized proteins, the Fenton reaction, and such enzymes as cyclooxygenases, lipoxygenases, xanthine oxidases, and NADPH oxidases. Furthermore, the defense mechanisms against oxidative stress ranging from enzymes like superoxide dismutases, catalases, peroxiredoxins, and GSH peroxidases to organic compounds such as L-ascorbate, alpha-tocopherol, beta-carotene, uric acid, CoQ10, and glutathione are described in more detail. In addition the oxidative stress induced modifications caused to proteins, lipids and DNA are discussed. Finally age-related changes of the skin are also a topic of this review. They include a disruption of the epidermal calcium gradient in old skin with an accompanying change in the composition of the cornified envelope. This modified cornified envelope also leads to an altered anti-oxidative capacity and a reduced barrier function of the epidermis.
DOI:10.1016/j.fsc.2016.08.011URLPMID:27888889 [本文引用: 1]
Skin coloration is highly diverse, partly due to the presence of pigmentation. Color variation is related to the extent of ultraviolet radiation exposure, as well as other factors. Inherent skin coloration arises from differences in basal epidermal melanin amount and type. Skin color is influenced by both the quantity and distribution of melanocytes. The effectiveness of inherent pigmentation for protecting living cells also varies. This article discusses skin color, pigmentation, and ethnicity in relation to clinical practice. Color perception, skin typing/classification, and quantitation of pigmentation are reviewed in relation to ethnicity, environmental stresses/irritants, and potential treatment effects.
DOI:10.1038/s41598-018-28719-wURLPMID:29991772 [本文引用: 5]
Comparative population genomics analysis is an effective approach to identify selection signatures in farm animals. In this study, we systematically investigated the selection signatures in six phenotypically diverse goat breeds using SNPs obtained from pooled whole-genome resequencing data. More than 95.5% of 446-642 million clean reads were mapped to the latest reference goat genome, which generated a sequencing depth ranging from 22.30 to 31.75-fold for each breed. A total of 5,802,307, 6,794,020, 7,562,312, 5,325,119, 8,764,136, and 9,488,057 putative SNPs were detected in Boer, Meigu, Jintang Black, Nanjiang Yellow, Tibetan, and Tibetan cashmere goats, respectively. Based on the genome-wide FST and expected heterozygosity scores along 100-kb sliding windows, 68, 89, 44, 44, 19, and 35 outlier windows were deemed as the selection signatures in the six goat breeds. After genome annotation, several genes within the selection signals were found to be possibly associated with important traits in goats, such as coat color (IRF4, EXOC2, RALY, EIF2S2, and KITLG), high-altitude adaptation (EPAS1), growth (LDB2), and reproduction traits (KHDRBS2). In summary, we provide an improved understanding of the genetic diversity and the genomic footprints under positive selection or the adaptations to the local environments in the domestic goat genome.
DOI:10.1037/0735-7036.111.4.419URLPMID:9419886 [本文引用: 1]
The agouti locus influences coat color by antagonizing melanocyte-stimulating hormone (MSH) at its receptor on pigment cells and may antagonize MSH in neural tissue. This study replicates work on rats to assess whether behavioral (neural) effects of the agouti locus are as similar across mammals as those on coat color. Handling, open-field, platform jump, and food-novelty tests were conducted on agouti and nonagouti deer mice (Peromyscus maniculatus) following protocols in C. A. Cottle and E. O. Price (1987). As with rats, nonagouti deer mice were less aggressive, less active, and easier to handle compared with their agouti counterparts. Nonagouti deer mice also groomed more than agouti subjects. Thus, behavioral effects of the agouti locus are conservative, and agouti may be an important modulator of melanocortins in neural as well as integumentary tissue.
DOI:10.1095/biolreprod.103.019323URLPMID:12890724 [本文引用: 1]
Mutations in the dominant-white spotting (W; c-kit) and stem cell factor (Sl; SCF) genes, which encode the transmembrane tyrosine kinase receptor and its ligand, respectively, affect both the proliferation and differentiation of many types of stem cells. Almost all homozygous W or Sl mutant mice are sterile because of the lack of differentiated germ cells or spermatogonial stem cells. To characterize spermatogenesis in c-kit/SCF mutants and to understand the role of c-kit signal transduction in spermatogonial stem cells, the existence, proliferation, and differentiation of spermatogonia were examined in the W/Wv mutant mouse testis. In the present study, some of the W/Wv mutant testes completely lacked spermatogonia, and many of the remaining testes contained only a few spermatogonia. Examination of the proliferative activity of the W/Wv mutant spermatogonia by transplantation of enhanced green fluorescent protein (eGFP)-labeled W/Wv spermatogonia into the seminiferous tubules of normal SCF (W/Wv) or SCF mutant (Sl/Sld) mice demonstrated that the W/Wv spermatogonia had the ability to settle and proliferate, but not to differentiate, in the recipient seminiferous tubules. Although the germ cells in the adult W/Wv testis were c-kit-receptor protein-negative undifferentiated type A spermatogonia, the juvenile germ cells were able to differentiate into spermatogonia that expressed the c-kit-receptor protein. Furthermore, differentiated germ cells with the c-kit-receptor protein on the cell surface could be induced by GnRH antagonist treatment, even in the adult W/Wv testis. These results indicate that all the spermatogonial stem cell characteristics of settlement, proliferation, and differentiation can be demonstrated without stimulating the c-kit-receptor signal. The c-kit/SCF signal transduction system appears to be necessary for the maintenance and proliferation of differentiated c-kit receptor-positive spermatogonia but not for the initial step of spermatogonial cell differentiation.
DOI:10.1126/science.147.3662.1145URLPMID:14242032 [本文引用: 1]
The
[本文引用: 2]
DOI:10.1111/1755-0998.12311URLPMID:25132237 [本文引用: 2]
Pigs from Asia and Europe were independently domesticated from c. 9000 years ago. During this period, strong artificial selection has led to dramatic phenotypic changes in domestic pigs. However, the genetic basis underlying these morphological and behavioural adaptations is relatively unknown, particularly for indigenous Chinese pigs. Here, we performed a genome-wide analysis to screen 196 regions with selective sweep signals in Tongcheng pigs, which are a typical indigenous Chinese breed. Genes located in these regions have been found to be involved in lipid metabolism, melanocyte differentiation, neural development and other biological processes, which coincide with the evolutionary phenotypic changes in this breed. A synonymous substitution, c.669T>C, in ESR1, which colocalizes with a major quantitative trait locus for litter size, shows extreme differences in allele frequency between Tongcheng pigs and wild boars. Notably, the variant C allele in this locus exhibits high allele frequency in most Chinese populations, suggesting a consequence of positive selection. Five genes (PRM1, PRM2, TNP2, GPR149 and JMJD1C) related to reproductive traits were found to have high haplotype similarity in Chinese breeds. Two selected genes, MITF and EDNRB, are implied to shape the two-end black colour trait in Tongcheng pig. Subsequent SNP microarray studies of five Chinese white-spotted breeds displayed a concordant signature at both loci, suggesting that these two genes are responsible for colour variations in Chinese breeds. Utilizing massively parallel sequencing, we characterized the candidate sites that adapt to artificial and environmental selections during the Chinese pig domestication. This study provides fundamental proof for further research on the evolutionary adaptation of Chinese pigs.
DOI:10.1038/srep27534URLPMID:27270507 [本文引用: 2]
A common phenotypic difference among domestic animals is variation in coat color. Six-white-point is a pigmentation pattern observed in varying pig breeds, which seems to have evolved through several different mechanistic pathways. Herein, we re-sequenced whole genomes of 31 Diannan small-ear pigs from China and found that the six-white-point coat color in Diannan small-ear pigs is likely regulated by polygenic loci, rather than by the MC1R locus. Strong associations were observed at three loci (EDNRB, CNTLN, and PINK1), which explain about 20 percent of the total coat color variance in the Diannan small-ear pigs. We found a mutation that is highly differentiated between six-white-point and black Diannan small-ear pigs, which is located in a conserved noncoding sequence upstream of the EDNRB gene and is a putative binding site of the CEBPB protein. This study advances our understanding of coat color evolution in Diannan small-ear pigs and expands our traditional knowledge of coat color being a monogenic trait.
DOI:10.1093/molbev/msaa279URLPMID:33212507 [本文引用: 2]
Visible pigmentation phenotypes can be used to explore the regulation of gene expression and the evolution of coat color patterns in animals. Here, we performed whole-genome and RNA sequencing and applied genome-wide association study, comparative population genomics and biological experiments to show that the 2,809-bp-long LINE-1 insertion in the ASIP (agouti signaling protein) gene is the causative mutation for the white coat phenotype in swamp buffalo (Bubalus bubalis). This LINE-1 insertion (3' truncated and containing only 5' UTR) functions as a strong proximal promoter that leads to a 10-fold increase in the transcription of ASIP in white buffalo skin. The 165 bp of 5' UTR transcribed from the LINE-1 is spliced into the first coding exon of ASIP, resulting in a chimeric transcript. The increased expression of ASIP prevents melanocyte maturation, leading to the absence of pigment in white buffalo skin and hairs. Phylogenetic analyses indicate that the white buffalo-specific ASIP allele originated from a recent genetic transposition event in swamp buffalo. Interestingly, as a similar LINE-1 insertion has been identified in the cattle ASIP gene, we discuss the convergent mechanism of coat color evolution in the Bovini tribe.
DOI:10.1111/age.12334URLPMID:26417640 [本文引用: 2]
A candidate gene analysis of the microphthalmia-associated transcription factor (MITF) gene was used in an attempt to identify the genetic basis for a white-spotted coat color phenotype in the Asian swamp buffalo (Bubalus bubalis carabanensis). Ninety-three buffaloes-32 solid, 38 spotted and 23 white individuals-were Sanger-sequenced for all MITF exons as well as highly conserved intronic and flanking regions. MITF cDNA representing skin and iris tissue from six spotted, nine solid and one white buffaloes was also Sanger-sequenced to confirm detected mutations. Two independent loss-of-function mutations, a premature stop codon (c.328C>T, p.Arg110*) and a donor splice-site mutation (c.840+2T>A, p.Glu281_Leu282Ins8), both of which cause white-spotted coat color in swamp buffaloes, were identified. The nonsense mutation leads to a premature stop codon in exon 3, and likely removal of the resulting mRNA via nonsense-mediated decay pathway, whereas the donor splice-site mutation leads to aberrant splicing of exon 8 that encodes part of a highly conserved region of MITF. The resulting insertion of eight amino acid residues is expected to perturb the leucine zipper part in the basic helix-loop-helix leucine zipper (bHLH-Zip) domain and will most likely influence dimerization and DNA binding capacity. Electrophoretic mobility shift assay was performed using mutant and wild-type MITF proteins and showed that the mutant MITF protein resulting from the splice-site mutation decreased in vitro DNA binding capacity compared to wild-type MITF. White-spotted buffalo bulls are sacrificed in funeral ceremonies in Tana Toraja, Indonesia, because they are considered holy, and our results show that genetic variation causes a tie to the cultural use of these buffaloes.
DOI:10.1093/jhered/esm022URLPMID:17548862 [本文引用: 6]
New techniques allow fast genotyping of large numbers of single-nucleotide polymorphisms (SNPs) of the genome. These techniques are used to map disorders with complex inheritance patterns and require large study groups. Linkage analysis of monogenetic traits exploits close family relationships between relatively small numbers of cases and controls. Linkage studies are typically performed with a set of microsatellite markers spaced at 10 cM. We were interested to test whether SNP typing could be applied in genome-wide linkage analysis because of the speed of the procedure. White spotting in Boxer dogs was chosen as a model because it is a semidominant trait, allowing the assignment of locus genotypes to each phenotyped dog. A set of just more than 1500 SNPs were typed in 5 families with heterozygous parents and offspring that included 11 white, 6 brown, and 19 spotted dogs. Multipoint linkage analysis was performed and a LOD score of 12.1 was obtained on canine chromosome 20. The CFA20 region was the only region with a positive LOD score. The gene MITF, coding for a transcription factor implicated in Waardenburg syndrome in humans, is located in the region close to a SNP that is in apparent linkage disequilibrium with the white spotting locus. Thus, MITF is a likely candidate for involvement in white spotting in boxers. We conclude that SNPs, spaced at an average distance of 1.6 Mb, are highly informative in linkage analysis of monogenic traits and are a powerful alternative to microsatellite markers.
DOI:10.1111/j.1365-2052.2007.01664.xURLPMID:18052939 [本文引用: 6]
Tremendous progress has been made in identifying genes involved in pigmentation in dogs in the past few years. Comparative genomics has both aided and benefited from these findings. Seven genes that cause specific coat colours and/or patterns in dogs have been identified: melanocortin 1 receptor, tyrosinase related protein 1, agouti signal peptide, melanophilin, SILV (formerly PMEL17), microphthalmia-associated transcription factor and beta-defensin 103. Although not all alleles have been yet identified at each locus, DNA tests are available for many. The identification of these alleles has provided information on interactions in this complex set of genes involved in both pigmentation and neurological development. The review also discusses pleiotropic effects of some coat colour genes as they relate to disease. The alleles found in various breeds have shed light on some potential breed development histories and phylogenetic relationships. The information is of value to dog breeders who have selected for and against specific colours since breed standards and dog showing began in the late 1800s. Because coat colour is such a visible trait, this information will also be a valuable teaching resource.
DOI:10.1371/journal.pone.0055469URLPMID:23424634 [本文引用: 2]
Coat color in dog breeds is an excellent character for revealing the power of artificial selection, as it is extremely diverse and likely the result of recent domestication. Coat color is generated by melanocytes, which synthesize pheomelanin (a red or yellow pigment) or eumelanin (a black or brown pigment) through the pigment type-switching pathway, and is regulated by three genes in dogs: MC1R (melanocortin receptor 1), CBD103 (beta-defensin 103), and ASIP (agouti-signaling protein precursor). The genotypes of these three gene loci in dog breeds are associated with coat color pattern. Here, we resequenced these three gene loci in two Kunming dog populations and analyzed these sequences using population genetic approaches to identify evolutionary patterns that have occurred at these loci during the recent domestication and breeding of the Kunming dog. The analysis showed that MC1R undergoes balancing selection in both Kunming dog populations, and that the Fst value for MC1R indicates significant genetic differentiation across the two populations. In contrast, similar results were not observed for CBD103 or ASIP. These results suggest that high heterozygosity and allelic differences at the MC1R locus may explain both the mixed color coat, of yellow and black, and the difference in coat colors in both Kunming dog populations.
DOI:10.1111/j.1365-2052.2009.01893.xURLPMID:19456317 [本文引用: 3]
White coat colour in horses is inherited as a monogenic autosomal dominant trait showing a variable expression of coat depigmentation. Mutations in the KIT gene have previously been shown to cause white coat colour phenotypes in pigs, mice and humans. We recently also demonstrated that four independent mutations in the equine KIT gene are responsible for the dominant white coat colour phenotype in various horse breeds. We have now analysed additional horse families segregating for white coat colour phenotypes and report seven new KIT mutations in independent Thoroughbred, Icelandic Horse, German Holstein, Quarter Horse and South German Draft Horse families. In four of the seven families, only one single white horse, presumably representing the founder for each of the four respective mutations, was available for genotyping. The newly reported mutations comprise two frameshift mutations (c.1126_1129delGAAC; c.2193delG), two missense mutations (c.856G>A; c.1789G>A) and three splice site mutations (c.338-1G>C; c.2222-1G>A; c.2684+1G>A). White phenotypes in horses show a remarkable allelic heterogeneity. In fact, a higher number of alleles are molecularly characterized at the equine KIT gene than for any other known gene in livestock species.
DOI:10.1111/age.12528URLPMID:28084638 [本文引用: 1]
[本文引用: 3]
[本文引用: 3]
DOI:10.1038/s41598-017-01727-yURLPMID:28484254 [本文引用: 3]
The 3' untranslated regions (UTRs), located at the end of mRNA molecules, are believed to play a role in RNA replication and/or protein translation. Mutations in the tyrosinase (Tyr) gene are known to cause recessive albinism in humans and other species. In this study, to test whether the CRISPR/Cas9 system works on the mutation of the UTRs regulatory region in rabbit, the 3' UTR of the rabbit Tyr gene was deleted by a dual sgRNA directed CRISPR/Cas9 system. As expected, gray coat color and reduced melanin in hair follicles and irises was found in the mutated rabbit, thus increasing confidence in the association of the mutation of the Tyr 3' UTR with graying in rabbit. The graying phenotype was also found in the F1 generation, suggesting that the mutated allele can be stably inherited by the offspring. Thus, we provide the first evidence that reduced melanin and graying can be caused by deletion of the Tyr 3' UTR in rabbits. Additionally, CRISPR/Cas9-mediated large fragment deletions can facilitate genotype to phenotype studies of UTRs or non-coding RNAs in future.
DOI:10.7717/peerj.8077URLPMID:31772839 [本文引用: 2]
Background: To investigate the molecular mechanisms determining the coat color of native breed sheep in Xinjiang. Methods: Bashibai sheep, Yemule white sheep and Tulufan black sheep were selected. Illumina HiSeq X Ten sequencing technology was used to detect the genes responsible for the white, light brown, black and cyan gray coat colors in sheep. Sequence analysis and functional gene annotation analysis were performed to analyze the results. The signal pathways and differentially expressed genes related to sheep hair color production regulation were screened and finally verified by real-time polymerase chain reaction. Results: Functional annotation by Kyoto Encyclopedia of Genes and Genomes analysis revealed significant differences in enrichment of immunity-related pathways as well as melanogenesis synthetic and tyrosine metabolism pathways. Our results showed that the DCT, TYR, TYRP1, PMEL, SLC45A2 and MLANA six genes may be associated with the regulation of coat color development and provide a theoretical basis for selecting natural coat colors of sheep.
DOI:10.1111/age.12769URLPMID:30730042 [本文引用: 2]
The llama (Lama glama) is a fiber-producing species that presents a wide range of coat colors, among which white is one of the most important for the textile industry. However, there is little information about the molecular mechanisms that control the white phenotype in this species. In domestic mammals, a white coat is usually produced by mutations in the KIT proto-oncogene receptor tyrosine kinase (KIT) and microphthalmia-associated transcription factor (MITF) genes. In this work we have sequenced and described the coding regions of KIT and MITF-M, the melanocyte-specific isoform, and the two transcriptional variants MITF-M(-) and MITF-M(+). Moreover, we studied the expression of these genes in the skin of white and colored llamas. Although no variants were revealed to be associated with white coat color, significant differences between phenotypes were observed in the expression levels of KIT and MITF-M. Interestingly, white llamas expressed less MITF-M(+) than did colored ones, which is consistent with a consequent reduction in the synthesis of melanin. Even though our results indicate that downregulation of KIT and MITF-M expression is involved in white phenotype production in llamas, the causative gene of white coat color remains unknown.
[本文引用: 2]
DOI:10.1111/age.12105URL [本文引用: 2]
The dominant white coat colour of farmed blue fox is inherited as a monogenic autosomal dominant trait and is suggested to be embryonic lethal in the homozygous state. In this study, the transcripts of KIT were identified by RT-PCR for a dominant white fox and a normal blue fox. Sequence analysis showed that the KIT transcript in normal blue fox contained the full-length coding sequence of 2919bp (GenBank Acc. No ), but in the dominant white individual, a truncated isoform lacking the entire exon 12 specifically co-expressed with the normal transcript. Genomic DNA sequencing revealed that a single nucleotide polymorphism (c.1867+1G>T) in intron 12 appeared only in the dominant white individuals and a 1-bp ins/del polymorphism in the same intron showed in individuals representing two different coat colours. Genotyping results of the SNP with PCR-RFLP in 185 individuals showed all 90 normal blue foxes were homozygous for the G allele, and all dominant white individuals were heterozygous. Due to the truncated protein with a deletion of 35 amino acids and an amino acid replacement (p.Pro623Ala) located in the conserved ATP binding domain, we propose that the mutant receptor had absent tyrosine kinase activity. These findings reveal that the base substitution at the first nucleotide of intron 12 of KIT gene, resulting in skipping of exon 12, is a causative mutation responsible for the dominant white phenotype of blue fox.
DOI:10.7717/peerj.9402URLPMID:32596061 [本文引用: 2]
DOI:10.1038/s41467-020-19813-7URLPMID:33293529 [本文引用: 2]
Current knowledge about the evolutionary history of donkeys is still incomplete due to the lack of archeological and whole-genome diversity data. To fill this gap, we have de novo assembled a chromosome-level reference genome of one male Dezhou donkey and analyzed the genomes of 126 domestic donkeys and seven wild asses. Population genomics analyses indicate that donkeys were domesticated in Africa and conclusively show reduced levels of Y chromosome variability and discordant paternal and maternal histories, possibly reflecting the consequences of reproductive management. We also investigate the genetic basis of coat color. While wild asses show diluted gray pigmentation (Dun phenotype), domestic donkeys display non-diluted black or chestnut coat colors (non-Dun) that were probably established during domestication. Here, we show that the non-Dun phenotype is caused by a 1 bp deletion downstream of the TBX3 gene, which decreases the expression of this gene and its inhibitory effect on pigment deposition.
DOI:10.1186/1471-2164-13-555URLPMID:23067000 [本文引用: 3]
UNLABELLED: AB BACKGROUND: MicroRNAs (miRNAs) are small, non-coding 21-25 nt RNA molecules that play an important role in regulating gene expression. Little is known about the expression profiles and functions of miRNAs in skin and their role in pigmentation. Alpacas have more than 22 natural coat colors, more than any other fiber producing species. To better understand the role of miRNAs in control of coat color we performed a comprehensive analysis of miRNA expression profiles in skin of white versus brown alpacas. RESULTS: Two small RNA libraries from white alpaca (WA) and brown alpaca (BA) skin were sequenced with the aid of Illumina sequencing technology. 272 and 267 conserved miRNAs were obtained from the WA and BA skin libraries, respectively. Of these conserved miRNAs, 35 and 13 were more abundant in WA and BA skin, respectively. The targets of these miRNAs were predicted and grouped based on Gene Ontology and KEGG pathway analysis. Many predicted target genes for these miRNAs are involved in the melanogenesis pathway controlling pigmentation. In addition to the conserved miRNAs, we also obtained 22 potentially novel miRNAs from the WA and BA skin libraries. CONCLUSION: This study represents the first comprehensive survey of miRNAs expressed in skin of animals of different coat colors by deep sequencing analysis. We discovered a collection of miRNAs that are differentially expressed in WA and BA skin. The results suggest important potential functions of miRNAs in coat color regulation.
[本文引用: 1]
DOI:10.1186/s12867-019-0137-8URLPMID:31412786 [本文引用: 1]
BACKGROUND: Many miRNA functions have been revealed to date. Single miRNAs can participate in life processes by regulating more than one target gene, and more than one miRNA can also simultaneously act on one target mRNA. Thus, a complex regulatory network involved in many processes can be formed. Herein, the pigmentation regulation mechanism of miR-101a-3p and miR-144a-3p was studied at the cellular level by the overexpression and equal overexpression of miR-101a-3p and miR-144a-3p. RESULTS: Results revealed that miR-101a-3p and miR-144a-3p directly regulated the expression of microphthalmia-associated transcription factor, thereby affecting melanin synthesis. CONCLUSIONS: The two miRNAs with the same binding site in the same gene independently excreted each other's function. However, the inhibitory effect of miR-144a-3p was stronger than that of miR-101-3p in alpaca melanocytes, although both decreased melanin production.
DOI:10.1186/s12864-019-6343-4URLPMID:31823726 [本文引用: 2]
BACKGROUND: Melanocytes are derived from neural crest stem cells in the embryonic stage. In mature melanocytes, a series of complex enzyme-catalyzed reactions leads to the production of melanins, which determine the hair and skin colors of animals. The process of melanogenesis is complex and can be regulated by mRNA, microRNAs (miRNAs) and long noncoding RNAs (lncRNAs) genes. MiRNAs are a type of endogenous noncoding RNA approximately 22 nt in size that predominantly regulate gene expression by inhibiting translation. miR-380-3p is a candidate miRNA potentially related to melanogenesis. To better understand the mechanism of miR-380-3p melanogenesis regulation, plasmids to overexpress or knockdown miR-380-3p were transfected into alpaca melanocytes, and their effects on melanogenesis were evaluated. RESULTS: In situ hybridization identified a positive miR-380-3p signal in alpaca melanocyte cytoplasm. Luciferase activity assays confirmed that SOX6 is targeted by miR-380-3p. miR-380-3p overexpression and knockdown in alpaca melanocytes respectively downregulated and upregulated SOX6 expression at the mRNA and protein levels. Additionally, miR-380-3p overexpression and knockdown, respectively, in alpaca melanocytes decreased and increased the mRNA levels of melanin transfer-related genes, including microphthalmia-associated transcription factor (MITF), tyrosinase (TYR), tyrosine-related protein-1 (TYRP1), and dopachrome tautomerase (DCT). In contrast, miR-380-3p overexpression and knockdown respectively increased and decreased the mRNA levels of beta-catenin. Additionally, the effect of miR-380-3p on melanogenesis was assessed by Masson-Fontana melanin staining. CONCLUSIONS: The results demonstrated that miR-380-3p targeted SOX6 to regulate melanogenesis by influencing beta-catenin and MITF transcription and translation, which reduced the expression of downstream genes, including TYR, TYRP1, and DCT. These results provide insights into the mechanisms through which miR-380-3p controls melanogenesis.
DOI:10.1111/j.1365-2052.2005.01409.xURLPMID:16573534 [本文引用: 2]
Albino phenotypes are documented in a variety of species including the domestic cat. As albino phenotypes in other species are associated with tyrosinase (TYR) mutations, TYR was proposed as a candidate gene for albinism in cats. An Oriental and Colourpoint Shorthair cat pedigree segregating for albinism was analysed for association with TYR by linkage and sequence analyses. Microsatellite FCA931, which is closely linked to TYR and TYR sequence variants were tested for segregation with the albinism phenotype. Sequence analysis of genomic DNA from wild-type and albino cats identified a cytosine deletion in TYR at position 975 in exon 2, which causes a frame shift resulting in a premature stop codon nine residues downstream from the mutation. The deletion mutation in TYR and an allele of FCA931 segregated concordantly with the albino phenotype. Taken together, our results suggest that the TYR gene corresponds to the colour locus in cats and its alleles, from dominant to recessive, are as follows: C (full colour) > c(b) (burmese) > or = c(s) (siamese) > c (albino).
DOI:10.1007/s003359901181URLPMID:10594238 [本文引用: 1]
An inheritable bleeding disorder with light coat color caused by an autosomal recessive gene has been reported in a population of Japanese black cattle. The disease has been diagnosed as Chediak-Higashi Syndrome (CHS) of cattle which correspond to a human inheritable disorder caused by mutation in LYST gene. To characterize the molecular lesion causing CHS in cattle, cDNAs encoding bovine LYST were isolated from a bovine brain cDNA library. The nucleotide and deduced amino acid sequences of bovine LYST had 89.6 and 90.2% identity with those of the human LYST gene, respectively. In order to identify the mutation within the LYST gene causing CHS in cattle, cDNA fragments of the LYST gene were amplified from an affected animal by RT-PCR and their nucleotide sequences were completely determined. Notably, a nucleotide substitution of A to G transition, resulting in an amino acid substitution of histidine to arginine (H2015R) was identified in the affected animal. The presence of the substitution was completely corresponding with the occurrence of the CHS phenotype among 105 members of pedigrees of the Japanese black cattle and no cattle of other populations had this substitution. These findings strongly suggested that H2015R is the causative mutation in CHS of Japanese black cattle.
DOI:10.1111/j.1439-0388.2008.00761.xURLPMID:19207931 [本文引用: 2]
Microphthalmia-associated transcription factor (MITF) is involved in white spotting and deafness associated with lack of pigmentation in human and mice. In the present study, we employed MITF-associated markers to evaluate MITF as a candidate for canine congenital sensorineural deafness (CCSD) in Dalmatian dogs. We performed an association study using MITF flanking and intragenic markers for 88 Dalmatian dogs of different hearing and eye pigmentation status. A significant association was identified for MITF-related markers with CCSD and blue iris colour. We conclude that MITF might play a role in CCSD and blue eye colour in Dalmatian dogs.
DOI:10.1038/ng.185URLPMID:18641652 [本文引用: 1]
In horses, graying with age is an autosomal dominant trait associated with a high incidence of melanoma and vitiligo-like depigmentation. Here we show that the Gray phenotype is caused by a 4.6-kb duplication in intron 6 of STX17 (syntaxin-17) that constitutes a cis-acting regulatory mutation. Both STX17 and the neighboring NR4A3 gene are overexpressed in melanomas from Gray horses. Gray horses carrying a loss-of-function mutation in ASIP (agouti signaling protein) had a higher incidence of melanoma, implying that increased melanocortin-1 receptor signaling promotes melanoma development in Gray horses. The Gray horse provides a notable example of how humans have cherry-picked mutations with favorable phenotypic effects in domestic animals.
DOI:10.1111/jvim.12133URL [本文引用: 2]
DOI:10.1186/1471-2156-6-34URLPMID:15960853 [本文引用: 1]
BACKGROUND: Pinschers and other dogs with coat color dilution show a characteristic pigmentation phenotype. The fur colors are a lighter shade, e.g. silvery grey (blue) instead of black and a sandy color (Isabella fawn) instead of red or brown. In some dogs the coat color dilution is sometimes accompanied by hair loss and recurrent skin inflammation, the so called color dilution alopecia (CDA) or black hair follicular dysplasia (BHFD). In humans and mice a comparable pigmentation phenotype without any documented hair loss is caused by mutations within the melanophilin gene (MLPH). RESULTS: We sequenced the canine MLPH gene and performed a mutation analysis of the MLPH exons in 6 Doberman Pinschers and 5 German Pinschers. A total of 48 sequence variations was identified within and between the breeds. Three families of dogs showed co-segregation for at least one polymorphism in an MLPH exon and the dilute phenotype. No single polymorphism was identified in the coding sequences or at splice sites that is likely to be causative for the dilute phenotype of all dogs examined. In 18 German Pinschers a mutation in exon 7 (R199H) was consistently associated with the dilute phenotype. However, as this mutation was present in homozygous state in four dogs of other breeds with wildtype pigmentation, it seems unlikely that this mutation is truly causative for coat color dilution. In Doberman Pinschers as well as in Large Munsterlanders with BHFD, a set of single nucleotide polymorphisms (SNPs) around exon 2 was identified that show a highly significant association to the dilute phenotype. CONCLUSION: This study provides evidence that coat color dilution is caused by one or more mutations within or near the MLPH gene in several dog breeds. The data on polymorphisms that are strongly associated with the dilute phenotype will allow the genetic testing of Pinschers to facilitate the breeding of dogs with defined coat colors and to select against Large Munsterlanders carrying BHFD.
DOI:10.1007/s003359900790URLPMID:9585428 [本文引用: 1]
Lethal White Foal Syndrome is a disease associated with horse breeds that register white coat spotting patterns. Breedings between particular spotted horses, generally described as frame overo, produce some foals that, in contrast to their parents, are all white or nearly all white and die shortly after birth of severe intestinal blockage. These foals have aganglionosis characterized by a lack of submucosal and myenteric ganglia from the distal small intestine to the large intestine, similar to human Hirschsprung Disease. Some sporadic and familial cases of Hirschsprung Disease are due to mutations in the endothelin B receptor gene (EDNRB). In this study, we investigate the role of EDNRB in Lethal White Foal Syndrome. A cDNA for the wild-type horse endothelin-B receptor gene was cloned and sequenced. In three unrelated lethal white foals, the EDNRB gene contained a 2-bp nucleotide change leading to a missense mutation (I118K) in the first transmembrane domain of the receptor, a highly conserved region of this protein among different species. Seven additional unrelated lethal white foal samples were found to be homozygous for this mutation. No other homozygotes were identified in 138 samples analyzed, suggesting that homozygosity was restricted to lethal white foals. All (40/40) horses with the frame overo pattern (a distinct coat color pattern that is a subset of overo horses) that were tested were heterozygous for this allele, defining a heterozygous coat color phenotype for this mutation. Horses with tobiano markings included some carriers, indicating that tobiano is epistatic to frame overo. In addition, horses were identified that were carriers but had no recognized overo coat pattern phenotype, demonstrating the variable penetrance of the mutation. The test for this mutant allele can be utilized in all breeds where heterozygous animals may be unknowingly bred to each other including the Paint Horse, Pinto horse, Quarter Horse, Miniature Horse, and Thoroughbred.
DOI:10.1016/0092-8674(94)90018-3URLPMID:8001160 [本文引用: 1]
Defects in the gene encoding the endothelin-B receptor produce aganglionic megacolon and pigmentary disorders in mice and humans. We report that a targeted disruption of the mouse endothelin-3 ligand (EDN3) gene produces a similar recessive phenotype of megacolon and coat color spotting. A natural recessive mutation that results in the same developmental defects in mice, lethal spotting (ls), failed to complement the targeted EDN3 allele. The ls mice carry a point mutation of the EDN3 gene, which replaces the Arg residue at the C-terminus of the inactive intermediate big EDN3 with a Trp residue. This mutation prevents the proteolytic activation of big EDN3 by ECE-1. These findings indicate that interaction of EDN3 with the endothelin-B receptor is essential in the development of neural crest-derived cell lineages. We postulate that defects in the human EDN3 gene may cause Hirschsprung's disease.
DOI:10.1007/s00347-007-1590-1URL [本文引用: 1]
DOI:10.1371/journal.pone.0125651URLPMID:25919014 [本文引用: 1]
BACKGROUND: Oculocutaneous albinism (OCA) is an autosomal recessive disorder. The most common type OCA1 and OCA2 are caused by homozygous or compound heterozygous mutations in the tyrosinase gene (TYR) and OCA2 gene, respectively. OBJECTIVE: The purpose of this study was to evaluate the molecular basis of oculocutaneous albinism in four Chinese families. PATIENTS AND METHODS: Four non-consanguineous OCA families were included in the study. The TYR and OCA2 genes of all individuals were amplified by polymerase chain reaction (PCR), sequenced and compared with a reference database. RESULTS: Four patients with a diagnosis of oculocutaneous albinism, presented with milky skin, white or light brown hair and nystagmus. Genetic analyses demonstrated that patient A was compound heterozygous for c.1037-7T.A, c.1037-10_11delTT and c.1114delG mutations in the TYR gene; patient B was heterozygous for c.593C>T and c.1426A>G mutations in the OCA2 gene, patients C and D were compound heterozygous mutations in the TYR gene (c.549_550delGT and c.896G>A, c.832C>T and c.985T>C, respectively). The heterozygous c.549_550delGT and c.1114delG alleles in the TYR gene were two novel mutations. Interestingly, heterozygous members in these pedigrees who carried c.1114delG mutations in the TYR gene or c.1426A>G mutations in the OCA2 gene presented with blond or brown hair and pale skin, but no ocular disorders when they were born; the skin of these patients accumulated pigment over time and with sun exposure. CONCLUSION: This study expands the mutation spectrum of oculocutaneous albinism. It is the first time, to the best of our knowledge, to report that c.549_550delGT and c.1114delG mutations in the TYR gene were associated with OCA. The two mutations (c.1114delG in the TYR gene and c.1426A>G in the OCA2 gene) may be responsible for partial clinical manifestations of OCA.
DOI:10.1002/mgg3.687URLPMID:31199599 [本文引用: 1]
BACKGROUND: Oculocutaneous albinism (OCA) is a group of heterogeneous autosomal recessive genetic disorder of melanin synthesis results in hypopigmented hair, skin, and eyes. OCA type 1, OCA type 2, and OCA type 4, which are respectively caused by mutations in TYR, OCA2, and SLC45A2 have high morbidity rates in Asia. METHODS: TYR, OCA2, and SLC45A2 mutation analysis was carried out on 18 nonconsanguineous OCA patients and four fetuses were included for prenatal diagnose. Three genes of all individuals were amplified by polymerase chain reaction and examined by Sanger sequencing. The pathogenicity of the detected mutations were analyzed by Mutation Taster, PolyPhen 2, and SIFT software, and the conservation of the substituted amino acids were analyzed by MEGA software. RESULTS: Eleven TYR mutations, three OCA2 mutations, and two SLC45A2 mutations were identified in 14 OCA type 1 patients, two OCA type 2 patients, and two OCA type 4 patients. c.1021A>G, p.R341G in TYR, c.1096_1104del, p.V366*, and c.1079C>T, p.S360F in OCA2 were novel. One of the four fetuses carried compound heterozygous mutation of TYR and became spontaneous abortion, the other three carried no mutations and appeared normal at birth. CONCLUSION: In this study, specific clinical characteristics of OCA patients were described. Three novel pathogenic mutations were identified which will enrich the mutation spectrum of OCA, and the prenatal genetic screening in fetus at risk of OCA can provide vital information for genetic counseling.

{kind=link}
{kind=link}