Cis-regulatory mechanisms and biological effects of translation elongation
Taolan Zhao1, Shuo Zhang1,2, Wenfeng Qian,1,2 1. State Key Laboratory of Plant Genomics, Institute of Genetics and Developmental Biology, Innovation Academy for Seed Design, Chinese Academy of Sciences, Beijing 100101, China 2. University of Chinese Academy of Sciences, Beijing 100049, China
Supported by the National Key R&D Program of China No.2019YFA0508700 the National Natural Science Foundation of China Nos.31900455 the National Natural Science Foundation of China Nos.91331112
作者简介 About authors 肇涛澜,博士,研究方向:翻译调控。E-mail:tlzhao@genetics.ac.cn。
Abstract Proteins are biological macromolecules essential for cells to maintain their metabolic activities. Proteins are synthesized during translation elongation, a synergistic process in which ribosomes decode the genetic information transmitted in mRNA, using tRNA. Numerous human diseases, such as neurodegenerative diseases and cancers, are known to be related to abnormal translation elongation. Translation elongation, as one of the two critical steps for the central dogma, used to be the focus of research in molecular biology. However, limitations in methodology had hindered further investigations on the dynamic process of translation elongation and its regulation. Recently, breakthroughs in methodology have revived this research field. Studies in the past decade or so have revealed that, beyond simple decoding of genetic information in mRNA, translation elongation entails sophisticated regulatory mechanisms and multifaceted biological consequences; such insights have provided a novel theoretical framework for understanding the maintenance of protein homeostasis and the development of diseases. In this review, we summarize the most updated methods that can be used to investigate the processes of translation elongation and highlight the mechanisms by which mRNA and protein sequences modulate the local rate of translation elongation. We further enumerate the consequences of dysregulation in translation elongation, from various aspects such as mRNA stability, protein synthesis and degradation, protein subcellular localization, and co-translational protein folding. We anticipate that this review will serve to draw the attention of scholars in various research fields to participate in the study of translation elongation. Keywords:translation elongation;ribosome;ribo-seq;co-translational protein folding;codon usage bias;RNA secondary structure;nascent polypeptide
PDF (932KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 肇涛澜, 张硕, 钱文峰. 翻译延伸的顺式调控机理与生物学效应. 遗传[J], 2020, 42(7): 613-631 doi:10.16288/j.yczz.20-074 Taolan Zhao. Cis-regulatory mechanisms and biological effects of translation elongation. Hereditas(Beijing)[J], 2020, 42(7): 613-631 doi:10.16288/j.yczz.20-074
翻译过程通常包括4个步骤:起始(initiation)、延伸(elongation)、终止(termination)和核糖体循环利用(recycling)。其中,翻译起始长久以来被认为是一条mRNA单位时间合成蛋白质数量的主要调节因素[1],受到翻译起始因子、Shine-Dalgarno (SD)序列等多方面的调控。然而近年来的研究表明,翻译延伸——核糖体从mRNA的5′端到3′端定向移动的同时将三联体核苷酸密码子的信息解码为氨基酸序列的过程——同样受到精细且严格的调控。翻译延伸的异常将引发mRNA的降解、错误的蛋白质亚细胞定位和折叠以及蛋白质的非生理性聚集[2],进而阻碍胚胎发育与神经系统的功能维持,研究显示这些影响与包括脆性X染色体综合征(fragile X syndrome)、神经退行性疾病(neurodegenerative diseases)和癌症在内的多种人类疾病的发生有关[3]。
由于氨基酸侧链的空间结构和化学性质存在差异,其形成肽键的速率各不相同(图3C)。脯氨酸是20种生物体主要氨基酸中最为特殊的一种,其氨基(-NH2)与侧链成环后仅存余亚氨基(-NH-)结构,因此准确地说是亚氨基酸。这使得其在位于核糖体A位点处时成为肽键形成的弱受体[20,71,72]。近期,A位点含有脯氨酰-tRNA类似物的核糖体结构进一步显示,脯氨酸作为底物在肽基转移酶中心(peptidyl transferase center, PTC)内处于不利的空间位置[73]。此外,不利的空间位置也使得位于核糖体P位点处的脯氨酸成为肽键形成的弱供体[74]。由于脯氨酸既是肽键形成的弱供体也是弱受体,当其串联出现时,对翻译延伸的抑制作用尤为明显[74,75,76,77,78,79]。
新生肽链的正确折叠对蛋白质正常执行其分子功能至关重要,而蛋白质的错误折叠和聚集则可导致多种严重的人类疾病,如阿尔茨海默病(Alzheimer’s disease, AD)、朊病毒(prion)相关疾病和帕金森病(Parkinson’s disease, PD)等[134,135,136]。模拟分析结果显示,近1/3的胞质蛋白质存在共翻译折叠现象(co-translational protein folding)[137],暗示了翻译延伸速率可能对蛋白质折叠发挥重要的调控作用(图3J)。
核糖体在同一条mRNA上延伸速率的异质性可能直接影响新合成蛋白质的构象。迄今为止,大多数相关例证均与密码子使用偏好性有关[138]。例如,稀有密码子被发现在蛋白质结构域边界区成簇出现,由其导致的翻译暂停一般被认为可为其上游蛋白质结构域的正确折叠提供充足时间[119,139~142]。通过改变同义密码子使用的方式改变翻译延伸速率可生成构象和功能异常的蛋白质[143,144,145,146]。除同义密码子的使用之外,一些基因组证据显示mRNA二级结构也可通过调控翻译延伸速率影响蛋白质折叠[147]。例如,拟南芥的mRNA二级结构倾向于在蛋白质结构域连接区或无序区增强,这可能会通过减缓翻译延伸保证新生肽链有充足的时间折叠出正确的蛋白质结构域[63];美国国家生物技术信息中心(National Center for Biotechnology Information, NCBI) Koonin课题组通过分析两个真核生物和3个原核生物的蛋白质结构数据也得到了类似的结论[148]。
蛋白质的共翻译折叠在新生肽链进入核糖体肽链输出通道的一刻即可开始[10]。核糖体肽链输出通道可容纳30~40个氨基酸长度的新生肽链,其宽度(10~20 Å)一般认为足够容纳α-螺旋(α-helix)等小型蛋白质二级结构在其内部折叠[149,150,151,152,153]。肽链输出通道出口外的新生肽链则可以与核糖体表面相互作用,这也可能调控蛋白质的折叠[154,155,156]。位于肽链输出通道外部的新生肽链还会接触到大量细胞质中的核糖体关联蛋白因子(ribosome-associated protein factor)与分子伴侣(chaperone),这些蛋白因子与核糖体及新生肽链的动态结合也被认为对蛋白质的共翻译折叠起到重要的调控作用[157,158,159]。例如,Liu等[160]研究显示,细菌中唯一的核糖体相关分子伴侣——触发因子(trigger factor)可通过结合EF-G的新生肽链来抑制结构域之间的错误相互作用,并防止未折叠区域的多肽对已折叠结构域的构象破坏。
ZhengCX, MaXF, ZhangYH, LiHJ, ZhangGF . Research progress in the mechanism of translation initiation of eukaryotic mRNAs Hereditas(Beijing), 2018,40(8):607-619. [本文引用: 1]
SteinKC, FrydmanJ . The stop-and-go traffic regulating protein biogenesis: how translation kinetics controls proteostasis J Biol Chem, 2019,294(6):2076-2084. [本文引用: 1]
RichterJD, CollerJ . Pausing on polyribosomes: make way for elongation in translational control Cell, 2015,163(2):292-300. [本文引用: 3]
ChoiJ, GroselyR, PrabhakarA, LapointeCP, WangJf, PuglisiJD . How messenger RNA and nascent chain sequences regulate translation elongation Annu Rev Biochem, 2018,87:421-449. [本文引用: 1]
BuskirkAR, GreenR . Ribosome pausing, arrest and rescue in bacteria and eukaryotes Philos Trans R Soc Lond B Biol Sci, 2017,372(1716):20160183. [本文引用: 4]
VoorheesRM, RamakrishnanV . Structural basis of the translational elongation cycle Annu Rev Biochem, 2013,82:203-236. [本文引用: 1]
LinJZ, GagnonMG, BulkleyD, SteitzTA . Conformational changes of elongation factor G on the ribosome during tRNA translocation Cell, 2015,160(0):219-227. [本文引用: 1]
GagnonMG, LinJZ, BulkleyD, SteitzTA . Crystal structure of elongation factor 4 bound to a clockwise ratcheted ribosome Science, 2014,345(6197):684-687. [本文引用: 1]
MustoeAM, BrooksCL, Al-HashimiHM . Hierarchy of RNA functional dynamics Annu Rev Biochem, 2014,83:441-466. [本文引用: 1]
JavedA, ChristodoulouJ, CabritaLD, OrlovaEV . The ribosome and its role in protein folding: looking through a magnifying glass Acta Crystallogr D Struct Biol, 2017,73(pt 6):509-521. [本文引用: 2]
BaiXC, FernandezIS, McMullan G, ScheresSH. Ribosome structures to near-atomic resolution from thirty thousand cryo-EM particles eLife, 2013,2:e00461. [本文引用: 1]
AmuntsA, BrownA, BaiXC, LlacerJL, HussainT, EmsleyP, LongF, MurshudovG, ScheresSHW, RamakrishnanV . Structure of the yeast mitochondrial large ribosomal subunit Science, 2014,343(6178):1485-1489. [本文引用: 1]
LiuZ, Gutierrez-VargasC, WeiJ, GrassucciRA, SunM, EspinaN, Madison-AntenucciS, TongL, FrankJ . Determination of the ribosome structure to a resolution of 2.5 ? by single-particle cryo-EM Protein Sci, 2017,26(1):82-92. [本文引用: 1]
LovelandAB, DemoG, GrigorieffN, KorostelevAA . Ensemble cryo-EM elucidates the mechanism of translation fidelity Nature, 2017,546(7656):113-117. [本文引用: 1]
FischerN, NeumannP, KonevegaAL, BockLV, FicnerR, RodninaMV, StarkH . Structure of the E. coli ribosome-EF-Tu complex at <3 ? resolution by Cs-corrected cryo-EM Nature, 2015,520(7548):567-570. [本文引用: 1]
ZhangYQ, MandavaCS, CaoW, LiXJ, ZhangDJ, LiNN, ZhangYX, ZhangXX, QinY, MiKX, LeiJL, SanyaS, GaoN . HflX is a ribosome-splitting factor rescuing stalled ribosomes under stress conditions Nat Struct Mol Biol, 2015,22(11):906-913. [本文引用: 1]
JohanssonM, BouakazE, LovmarM, EhrenbergM . The kinetics of ribosomal peptidyl transfer revisited Mol Cell, 2008,30(5):589-598. [本文引用: 1]
WangJF, KwiatkowskiM, ForsterAC . Kinetics of ribosome-catalyzed polymerization using artificial aminoacyl-tRNA substrates clarifies inefficiencies and improvements ACS Chem Biol, 2015,10(10):2187-2192. [本文引用: 1]
WohlgemuthI, BrennerS, BeringerM, RodninaMV . Modulation of the rate of peptidyl transfer on the ribosome by the nature of substrates J Biol Chem, 2008,283(47):32229-32235. [本文引用: 2]
ChuD, KazanaE, BellangerN, SinghT , Tuite MF, von der Haar T. Translation elongation can control translation initiation on eukaryotic mRNAs EMBO J, 2014,33(1):21-34. [本文引用: 2]
MorisakiT, StasevichTJ . Quantifying single mRNA translation kinetics in living cells Cold Spring Harb Perspect Biol, 2018,10(11):a032078. [本文引用: 1]
YanXW, HoekTA, ValeRD, TanenbaumME . Dynamics of translation of single mRNA molecules in vivo. Cell, 2016,165(4):976-989. [本文引用: 3]
WuB, EliscovichC, YoonYJ, SingerRH . Translation dynamics of single mRNAs in live cells and neurons Science, 2016,352(6292):1430-1435. [本文引用: 1]
MorisakiT, LyonK, DeLuca KF, DeLucaJG, EnglishBP, ZhangZJ, LavisLD, GrimmJB, ViswanathanS, LoogerLL, LionnetT, StasevichTJ. Real-time quantification of single RNA translation dynamics in living cells Science, 2016,352(6292):1425-1429. [本文引用: 1]
SteitzJA . Polypeptide chain initiation: nucleotide sequences of the three ribosomal binding sites in bacteriophage R17 RNA Nature, 1969,224(5223):957-964. [本文引用: 1]
IngoliaNT, GhaemmaghamiS, NewmanJR, WeissmanJS . Genome-wide analysis in vivo of translation with nucleotide resolution using ribosome profiling Science, 2009,324(5924):218-223. [本文引用: 2]
MichelAM, BaranovPV . Ribosome profiling: a Hi-Def monitor for protein synthesis at the genome-wide scale Wiley Interdiscip Rev RNA, 2013,4(5):473-490. [本文引用: 1]
GreletS, HowePH,. hnRNP E1 at the crossroads of translational regulation of epithelial-mesenchymal transition J Cancer Metastasis Treat, 2019,5:16. [本文引用: 1]
DesnoyersG, BouchardMP, MasseE . New insights into small RNA-dependent translational regulation in prokaryotes Trends Genet, 2013,29(2):92-98. [本文引用: 1]
AmbrosV, ChenXM . The regulation of genes and genomes by small RNAs Development, 2007,134(9):1635-1641. [本文引用: 1]
GranthamR, GautierC, GouyM, MercierR, PavéA . Codon catalog usage and the genome hypothesis Nucleic Acids Res, 1980,8(1):r49-r62. [本文引用: 1]
IkemuraT . Correlation between the abundance of Escherichia coli transfer RNAs and the occurrence of the respective codons in its protein genes: A proposal for a synonymous codon choice that is optimal for the E. coli translational system J Mol Biol, 1981,151(3):389-409. [本文引用: 2]
HershbergR, PetrovDA . Selection on codon bias Annu Rev Genet, 2008,42:287-299. [本文引用: 1]
PlotkinJB, KudlaG . Synonymous but not the same: the causes and consequences of codon bias Nat Rev Genet, 2011,12(1):32-42. [本文引用: 2]
GingoldH, PilpelY . Determinants of translation efficiency and accuracy Mol Syst Biol, 2011,7:481. [本文引用: 1]
AkashiH . Synonymous codon usage in Drosophila melanogaster: natural selection and translational accuracy Genetics, 1994,136(3):927-935. [本文引用: 1]
StoletzkiN, Eyre-WalkerA . Synonymous codon usage in Escherichia coli: selection for translational accuracy Mol Biol Evol, 2007,24(2):374-381. [本文引用: 1]
IkemuraT . Correlation between the abundance of yeast transfer RNAs and the occurrence of the respective codons in protein genes. Differences in synonymous codon choice patterns of yeast and Escherichia coli with reference to the abundance of isoaccepting transfer RNAs J Mol Biol, 1982,158(4):573-597. [本文引用: 1]
IkemuraT . Codon usage and tRNA content in unicellular and multicellular organisms Mol Biol Evol, 1985,2(1):13-34. [本文引用: 1]
NovoaEM, Ribas de Pouplana L. Speeding with control: codon usage, tRNAs, and ribosomes Trends Genet, 2012,28(11):574-581. [本文引用: 1]
BuhrF, JhaS, ThommenM, MittelstaetJ, KutzF, SchwalbeH, RodninaMV, KomarAA . Synonymous codons direct cotranslational folding toward different protein conformations Mol Cell, 2016,61(3):341-351. [本文引用: 1]
CurranJF, YarusM . Rates of aminoacyl-tRNA selection at 29 sense codons in vivo J Mol Biol, 1989,209(1):65-77. [本文引用: 1]
GardinJ, YeasminR, YurovskyA, CaiY, SkienaS, FutcherB . Measurement of average decoding rates of the 61 sense codons in vivo eLife, 2014,3:e03735. [本文引用: 1]
KerteszM, IovinoN, UnnerstallU, GaulU, SegalE . The role of site accessibility in microRNA target recognition Nat Genet, 2007,39(10):1278-1284. [本文引用: 1]
RouskinS, ZubradtM, WashietlS, KellisM, WeissmanJS . Genome-wide probing of RNA structure reveals active unfolding of mRNA structures in vivo Nature, 2014,505(7485):701-705. [本文引用: 1]
ChartrandP, MengXH, HuttelmaierS, DonatoD, SingerRH . Asymmetric sorting of ash1p in yeast results from inhibition of translation by localization elements in the mRNA Mol Cell, 2002,10(6):1319-1330. [本文引用: 1]
QuXH, WenJD, LancasterL, NollerHF, BustamanteC, TinocoI . The ribosome uses two active mechanisms to unwind messenger RNA during translation Nature, 2011,475(7354):118-121. [本文引用: 1]
KerteszM, WanY, MazorE, RinnJL, NutterRC, ChangHY, SegalE . Genome-wide measurement of RNA secondary structure in yeast Nature, 2010,467(7311):103-107. [本文引用: 1]
TangY, AssmannSM, BevilacquaPC . Protein structure is related to RNA structural reactivity in vivo. J Mol Biol, 2016,428(5):758-766. [本文引用: 3]
WattsJM, DangKK, GorelickRJ, LeonardCW, BessJW, SwanstromR, BurchCL, WeeksKM . Architecture and secondary structure of an entire HIV-1 RNA genome Nature, 2009,460(7256):711-716. [本文引用: 1]
DinmanJD . Mechanisms and implications of programmed translational frameshifting Wiley Interdiscip Rev RNA, 2012,3(5):661-673. [本文引用: 1]
BiswasP, JiangX, PacchiaAL, DoughertyJP, PeltzSW . The human immunodeficiency virus type 1 ribosomal frameshifting site is an invariant sequence determinant and an important target for antiviral therapy J Virol, 2004,78(4):2082-2087. [本文引用: 1]
WuF, ZhaoS, YuB, ChenYM, WangW, SongZG, HuY, TaoZW, TianJH, PeiYY, YuanML, ZhangYL, DaiFH, LiuY, WangQM, ZhengJJ, XuL, HolmesEC, ZhangYZ . A new coronavirus associated with human respiratory disease in China Nature, 2020,579(7798):265-269. [本文引用: 1]
CaliskanN, PeskeF, RodninaMV . Changed in translation: mRNA recoding by -1 programmed ribosomal frameshifting Trends Biochem Sci, 2015,40(5):265-274. [本文引用: 1]
PavlovMY, WattsRE, TanZ, CornishVW, EhrenbergM, ForsterAC . Slow peptide bond formation by proline and other N-alkylamino acids in translation Proc Natl Acad Sci USA, 2009,106(1):50-54. [本文引用: 2]
MutoH, ItoK . Peptidyl-prolyl-tRNA at the ribosomal P-site reacts poorly with puromycin Biochem Biophys Res Commun, 2008,366(4):1043-1047. [本文引用: 1]
MelnikovS, MailliotJ, RiggerL, NeunerS, ShinBS, YusupovaG, DeverTE, MicuraR, YusupovM . Molecular insights into protein synthesis with proline residues EMBO Rep, 2016,17(12):1776-1784. [本文引用: 1]
DoerfelLK, WohlgemuthI, KotheC, PeskeF, UrlaubH, RodninaMV . EF-P is essential for rapid synthesis of proteins containing consecutive proline residues Science, 2013,339(6115):85-88. [本文引用: 2]
WoolstenhulmeCJ, ParajuliS, HealeyDW, ValverdeDP, PetersenEN, StarostaAL, GuydoshNR, JohnsonWE, WilsonDN, BuskirkAR . Nascent peptides that block protein synthesis in bacteria Proc Natl Acad Sci USA, 2013,110(10):E878-E887. [本文引用: 3]
StarostaAL, LassakJ, PeilL, AtkinsonGC, Virum?eK, TensonT, RemmeJ, JungK, WilsonDN . Translational stalling at polyproline stretches is modulated by the sequence context upstream of the stall site Nucleic Acids Res, 2014,42(16):10711-10719. [本文引用: 1]
PeilL, StarostaAL, LassakJ, AtkinsonGC, Virum?eK, SpitzerM, TensonT, JungK, RemmeJ, WilsonDN . Distinct XPPX sequence motifs induce ribosome stalling, which is rescued by the translation elongation factor EF-P Proc Natl Acad Sci USA, 2013,110(38):15265-15270. [本文引用: 1]
RodninaMV . The ribosome in action: tuning of translational efficiency and protein folding Protein Sci, 2016,25(8):1390-1406. [本文引用: 1]
ItoK, ChibaS . Arrest peptides: cis-acting modulators of translation Annu Rev Biochem, 2013,82:171-202. [本文引用: 1]
TensonT, EhrenbergM . Regulatory nascent peptides in the ribosomal tunnel Cell, 2002,108(5):591-594. [本文引用: 2]
NakatogawaH, ItoK . The ribosomal exit tunnel functions as a discriminating gate Cell, 2002,108(5):629-636. [本文引用: 1]
MutoH, NakatogawaH, ItoK . Genetically encoded but nonpolypeptide prolyl-tRNA functions in the A site for SecM-mediated ribosomal stall Mol Cell, 2006,22(4):545-552. [本文引用: 1]
NavonSP, KornbergG, ChenJ, SchwartzmanT, TsaiA, PuglisiEV, PuglisiJD, AdirN . Amino acid sequence repertoire of the bacterial proteome and the occurrence of untranslatable sequences Proc Natl Acad Sci USA, 2016,113(26):7166-7170. [本文引用: 1]
LuJL, KobertzWR, DeutschC . Mapping the electrostatic potential within the ribosomal exit tunnel J Mol Biol, 2007,371(5):1378-1391. [本文引用: 1]
LuJL, DeutschC . Electrostatics in the ribosomal tunnel modulate chain elongation rates J Mol Biol, 2008,384(1):73-86. [本文引用: 2]
KoutmouKS, SchullerAP, BrunelleJL, RadhakrishnanA, DjuranovicS, GreenR . Ribosomes slide on lysine-encoding homopolymeric A stretches eLife, 2015,4:e05534. [本文引用: 1]
CharneskiCA, HurstLD . Positively charged residues are the major determinants of ribosomal velocity PLoS Biol, 2013,11(3):e1001508. [本文引用: 1]
ArtieriCG, FraserHB . Accounting for biases in riboprofiling data indicates a major role for proline in stalling translation Genome Res, 2014,24(12):2011-2021. [本文引用: 2]
SabiR, TullerT . A comparative genomics study on the effect of individual amino acids on ribosome stalling BMC Genomics, 2015,16(Suppl.10):S5. [本文引用: 1]
IkeuchiK, IzawaT, InadaT . Recent progress on the molecular mechanism of quality controls induced by ribosome stalling Front Genet, 2018,9:743. [本文引用: 2]
SharmaAK, O'BrienEP. Non-equilibrium coupling of protein structure and function to translation-elongation kinetics Curr Opin Struct Biol, 2018,49:94-103. [本文引用: 1]
Zurita RendonO, FredricksonEK, HowardCJ, Van VrankenJ, FogartyS, TolleyND, KaliaR, OsunaBA, ShenPS, HillCP, FrostA, RutterJ . Vms1p is a release factor for the ribosome-associated quality control complex Nat Commun, 2018,9(1):2197. [本文引用: 1]
DefenouillèreQ, YaoYH, MouaikelJ, NamaneA, GalopierA, DecourtyL, DoyenA, MalabatC, SaveanuC, JacquierA, Fromont-RacineM . Cdc48-associated complex bound to 60S particles is required for the clearance of aberrant translation products Proc Natl Acad Sci USA, 2013,110(13):5046-5051. [本文引用: 1]
VermaR, OaniaRS, KolawaNJ, DeshaiesRJ . Cdc48/p97 promotes degradation of aberrant nascent polypeptides bound to the ribosome eLife, 2013,2:e00308. [本文引用: 1]
DomaMK, ParkerR . Endonucleolytic cleavage of eukaryotic mRNAs with stalls in translation elongation Nature, 2006,440(7083):561-564. [本文引用: 2]
ShoemakerCJ, EylerDE, GreenR . Dom34:Hbs1 promotes subunit dissociation and peptidyl-tRNA drop-off to initiate no-go decay Science, 2010,330(6002):369-372. [本文引用: 1]
ShoemakerCJ, GreenR . Kinetic analysis reveals the ordered coupling of translation termination and ribosome recycling in yeast Proc Natl Acad Sci USA, 2011,108(51):E1392-1398. [本文引用: 1]
PisarevaVP, SkabkinMA, HellenCU, PestovaTV, PisarevAV . Dissociation by Pelota, Hbs1 and ABCE1 of mammalian vacant 80S ribosomes and stalled elongation complexes EMBO J, 2011,30(9):1804-1817. [本文引用: 1]
DimitrovaLN, KurohaK, TatematsuT, InadaT . Nascent peptide-dependent translation arrest leads to Not4p-mediated protein degradation by the proteasome J Biol Chem, 2009,284(16):10343-10352. [本文引用: 1]
JuszkiewiczS, Hegde RS. Initiation of quality control during poly(A)translation requires site-specific ribosome ubiquitination Mol Cell, 2017, 65(4): 743-750.e744. [本文引用: 1]
BengtsonMH, JoazeiroCAP . Role of a ribosome- associated E3 ubiquitin ligase in protein quality control Nature, 2010,467(7314):470-473. [本文引用: 1]
LetzringDP, WolfAS, BruleCE, GrayhackEJ . Translation of CGA codon repeats in yeast involves quality control components and ribosomal protein L1 RNA, 2013,19(9):1208-1217. [本文引用: 1]
SimmsCL, HudsonBH, MosiorJW, RangwalaAS, ZaherHS . An active role for the ribosome in determining the fate of oxidized Mrna Cell Rep, 2014,9(4):1256-1264. [本文引用: 3]
IshimuraR, NagyG, DotuI, ZhouHH, YangXL, SchimmelP, SenjuS, NishimuraY, ChuangJH, AckermanSL . RNA function. Ribosome stalling induced by mutation of a CNS-specific tRNA causes neurodegeneration Science, 2014,345(6195):455-459. [本文引用: 1]
ShahP, DingY, NiemczykM, KudlaG, PlotkinJB . Rate-limiting steps in yeast protein translation Cell, 2013,153(7):1589-1601. [本文引用: 1]
CarliniDB, StephanW . In vivo introduction of unpreferred synonymous codons into the Drosophila Adh gene results in reduced levels of ADH protein Genetics, 2003,163(1):239-243. [本文引用: 1]
TorrentM, ChalanconG, de GrootNS, WusterA, Madan Babu M,. Cells alter their tRNA abundance to selectively regulate protein synthesis during stress conditions Sci Signal, 2018,11(546): eaat6409. [本文引用: 1]
ZhaoTL, HuanQ, SunJ, LiuCY, HouXL, YuX, SilvermanIM, ZhangY, GregoryBD, LiuCM, QianWF, CaoXF . Impact of poly(A)-tail G-content on Arabidopsis PAB binding and their role in enhancing translational efficiency Genome Biol, 2019,20:189. [本文引用: 1]
ChenSY, LiK, CaoWQ, WangJ, ZhaoT, HuanQ, YangYF, WuSH, QianWF . Codon-resolution analysis reveals a direct and context-dependent impact of individual synonymous mutations on mRNA level Mol Biol Evol, 2017,34(11):2944-2958. [本文引用: 2]
Schikora-TamaritMA, CareyLB . Poor codon optimality as a signal to degrade transcripts with frameshifts Transcription, 2018,9(5):327-333. [本文引用: 1]
BazziniAA, Del VisoF, Moreno-MateosMA, JohnstoneTG, VejnarCE, QinYD, YaoJ, KhokhaMK, GiraldezAJ . Codon identity regulates mRNA stability and translation efficiency during the maternal-to-zygotic transition EMBO J, 2016,35(19):2087-2103. [本文引用: 1]
PresnyakV, AlhusainiN, ChenYH, MartinS, MorrisN, KlineN, OlsonS, WeinbergD, BakerKE, GraveleyBR, CollerJ . Codon optimality is a major determinant of mRNA stability Cell, 2015,160(6):1111-1124. [本文引用: 1]
RadhakrishnanA, ChenYH, MartinS, AlhusainN, GreenR, Coller J. The DEAD-box protein Dhh1p couples mRNA decay and translation by monitoring codon optimality Cell, 2016, 167(1): 122-132. e129. [本文引用: 1]
MishimaY, TomariY . Codon usage and 3' UTR length determine maternal mRNA stability in zebrafish Mol Cell, 2016,61(6):874-885. [本文引用: 1]
SiegelV, WalterP . Each of the activities of signal recognition particle (SRP) is contained within a distinct domain: analysis of biochemical mutants of SRP Cell, 1988,52(1):39-49. [本文引用: 1]
HalicM, BeckeT, PoolMR, SpahnCM, GrassucciRA, FrankJ, BeckmannR . Structure of the signal recognition particle interacting with the elongation-arrested ribosome Nature, 2004,427(6977):808-814. [本文引用: 1]
CrickSL, RuffKM, GaraiK, FriedenC, PappuRV . Unmasking the roles of N- and C-terminal flanking sequences from exon 1 of huntingtin as modulators of polyglutamine aggregation Proc Natl Acad Sci USA, 2013,110(50):20075-20080. [本文引用: 1]
BrinkmanRR, MezeiMM, TheilmannJ, AlmqvistE, HaydenMR . The likelihood of being affected with Huntington disease by a particular age, for a specific CAG size Am J Hum Genet, 1997,60(5):1202-1210. [本文引用: 1]
LiSH, LiXJ . Aggregation of N-terminal huntingtin is dependent on the length of its glutamine repeats Hum Mol Genet, 1998,7(5):777-782. [本文引用: 1]
LeeJM, RamosEM, LeeJH, GillisT, MysoreJS, HaydenMR, WarbySC, MorrisonP, NanceM, RossCA, MargolisRL, SquitieriF, OrobelloS, Di DonatoS, Gomez-TortosaE, AyusoC, SuchowerskyO, TrentRJ, McCusker E, NovellettoA, FrontaliM, JonesR, AshizawaT, FrankS, Saint-HilaireMH, HerschSM, RosasHD, LucenteD, HarrisonMB, ZankoA, AbramsonRK, MarderK, SequeirosJ, PaulsenJS, LandwehrmeyerGB, MyersRH, MacDonaldME, GusellaJF. CAG repeat expansion in Huntington disease determines age at onset in a fully dominant fashion Neurology, 2012,78(9):690-695. [本文引用: 1]
RockabrandE, SlepkoN, PantaloneA, NukalaVN, KazantsevA, MarshJL, SullivanPG, SteffanJS, SensiSL, ThompsonLM . The first 17 amino acids of Huntingtin modulate its sub-cellular localization, aggregation and effects on calcium homeostasis Hum Mol Genet, 2007,16(1):61-77. [本文引用: 1]
NissleyDA, O'Brien EP. Altered co-translational processing plays a role in Huntington's pathogenesis—a hypothesis Front Mol Neurosci, 2016,9:54. [本文引用: 1]
HartlFU . Protein misfolding diseases Annu Rev Biochem, 2017,86(1):21-26. [本文引用: 1]
ChitiF, DobsonCM . Protein misfolding, amyloid formation, and human disease: a summary of progress over the last decade Annu Rev Biochem, 2017,86:27-68. [本文引用: 1]
SotoC, PritzkowS . Protein misfolding, aggregation, and conformational strains in neurodegenerative diseases Nat Neurosci, 2018,21(10):1332-1340. [本文引用: 1]
CiryamP, MorimotoRI, VendruscoloM, DobsonCM, O'Brien EP. In vivo translation rates can substantially delay the cotranslational folding of the Escherichia coli cytosolic proteome Proc Natl Acad Sci USA, 2013,110(2):E132-140. [本文引用: 1]
KomarAA . Synonymous codon usage-a guide for co-translational protein folding in the cell Mol Biol (Mosk), 2019,53(6):883-898. [本文引用: 1]
KomarAA, LesnikT, ReissC . Synonymous codon substitutions affect ribosome traffic and protein folding during in vitro translation FEBS Lett, 1999,462(3):387-391.
ZhangG, IgnatovaZ . Folding at the birth of the nascent chain: coordinating translation with co-translational folding Curr Opin Struct Biol, 2011,21(1):25-31.
RodninaMV, WintermeyerW . Protein elongation, co-translational folding and targeting J Mol Biol, 2016,428(10):2165-2185. [本文引用: 1]
CortazzoP, Cerve?anskyC, MarínM, ReissC, EhrlichR, DeanaA . Silent mutations affect in vivo protein folding in Escherichia coli Biochem Biophys Res Commun, 2002,293(1):537-541. [本文引用: 1]
Kimchi-SarfatyC, OhJM, KimIW, SaunaZE, CalcagnoAM, AmbudkarSV, GottesmanMM . A “silent” polymorphism in the MDR1 gene changes substrate specificity Science, 2007,315(5811):525-528. [本文引用: 1]
WalshIM, BowmanMA, Soto SantarriagaIF, RodriguezA, ClarkPL . Synonymous codon substitutions perturb cotranslational protein folding in vivo and impair cell fitness Proc Natl Acad Sci USA, 2020,117(7):3528-3534. [本文引用: 1]
AgasheD, SaneM, PhalnikarK, DiwanGD, HabibullahA, Martinez-GomezNC, SahasrabuddheV, PolachekW, WangJ, ChubizLM, MarxCJ . Large-effect beneficial synonymous mutations mediate rapid and parallel adaptation in a bacterium Mol Biol Evol, 2016,33(6):1542-1553. [本文引用: 1]
YangJR . Does mRNA structure contain genetic information for regulating co-translational protein folding? Zool Res, 2017,38(1):36-43. [本文引用: 1]
FaureG, OgurtsovAY, ShabalinaSA, KooninEV . Role of mRNA structure in the control of protein folding Nucleic Acids Res, 2016,44(22):10898-10911. [本文引用: 1]
VossNR, GersteinM, SteitzTA, MoorePB . The geometry of the ribosomal polypeptide exit tunnel J Mol Biol, 2006,360(4):893-906. [本文引用: 1]
BhushanS, GartmannM, HalicM, ArmacheJP, JaraschA, MielkeT, BerninghausenO, WilsonDN, BeckmannR . Alpha-Helical nascent polypeptide chains visualized within distinct regions of the ribosomal exit tunnel Nat Struct Mol Biol, 2010,17(3):313-317. [本文引用: 1]
NilssonOB, HedmanR, MarinoJ, WicklesS, BischoffL, JohanssonM, Müller-LucksA, TrovatoF, PuglisiJD, O'Brien EP, BeckmannR, vonHeijne G. Cotranslational protein folding inside the ribosome exit tunnel Cell Rep, 2015,12(10):1533-1540. [本文引用: 1]
LuJL, DeutschC . Folding zones inside the ribosomal exit tunnel Nat Struct Mol Biol, 2005,12(12):1123-1129. [本文引用: 1]
WoolheadCA, McCormick PJ, JohnsonAE. Nascent membrane and secretory proteins differ in FRET-detected folding far inside the ribosome and in their exposure to ribosomal proteins Cell, 2004,116(5):725-736. [本文引用: 1]
KaiserCM, GoldmanDH, ChoderaJD, TinocoI, BustamanteC . The ribosome modulates nascent protein folding Science, 2011,334(6063):1723-1727. [本文引用: 1]
CabritaLD, CassaignauAME, LaunayHMM, WaudbyCA, WlodarskiT, CamilloniC, KaryadiME, RobertsonAL, WangXL, WentinkAS, GoodsellL, WoolheadCA, VendruscoloM, DobsonCM, ChristodoulouJ . A structural ensemble of a ribosome-nascent chain complex during cotranslational protein folding Nat Struct Mol Biol, 2016,23(4):278-285. [本文引用: 1]
CabritaLD, HsuST, LaunayH, DobsonCM, ChristodoulouJ . Probing ribosome-nascent chain complexes produced in vivo by NMR spectroscopy Proc Natl Acad Sci USA, 2009,106(2):22239-22244. [本文引用: 1]
SandikciA, GlogeF, MartinezM, MayerMP, WadeR, BukauB, KramerG . Dynamic enzyme docking to the ribosome coordinates N-terminal processing with polypeptide folding Nat Struct Mol Biol, 2013,20(7):843-850. [本文引用: 1]
WaudbyCA, DobsonCM, ChristodoulouJ . Nature and regulation of protein folding on the ribosome Trends Biochem Sci, 2019,44(11):914-926. [本文引用: 1]
LiuKX, MaciubaK, Kaiser CM, . The ribosome cooperates with a chaperone to guide multi-domain protein folding Mol Cell, 2019, 74(2): 310-319.e7. [本文引用: 1]
 ,1,2
,1,2

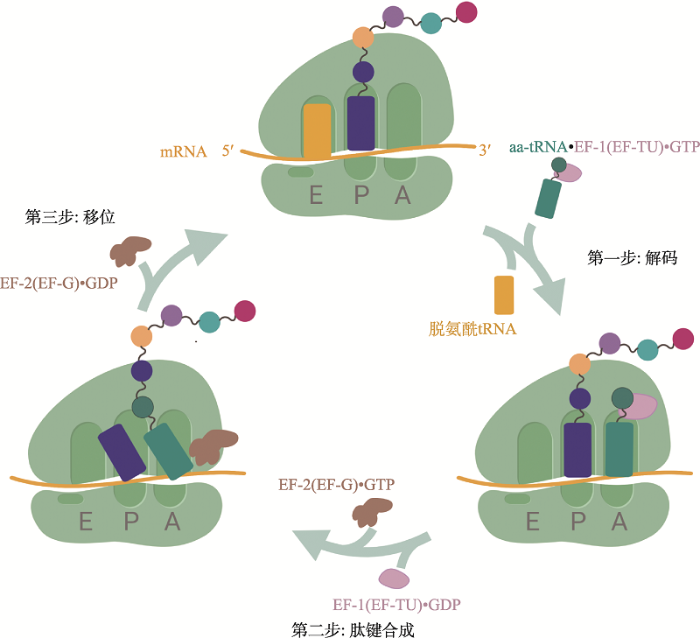 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT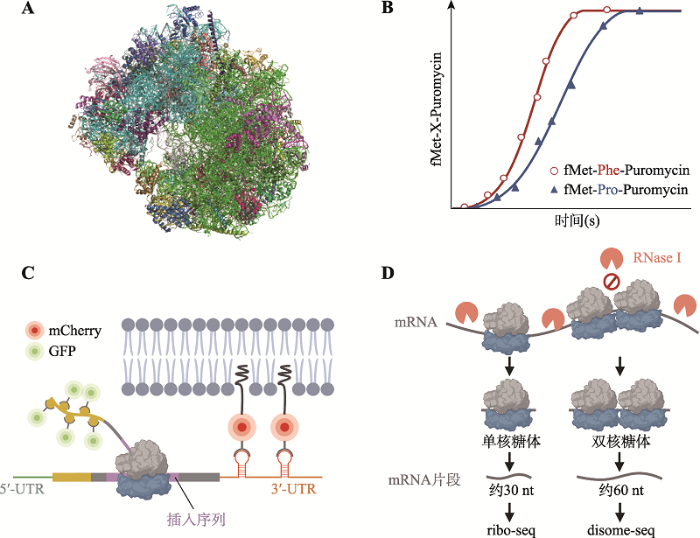 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT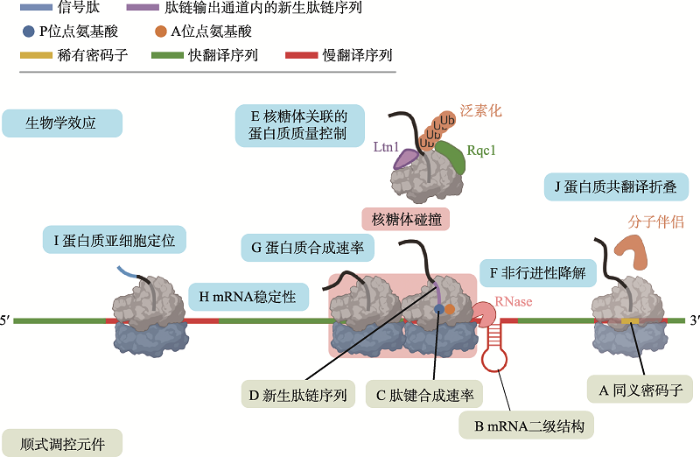 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}