 ,先正达北京创新中心,北京 102206
,先正达北京创新中心,北京 102206Genome editing in plants directed by CRISPR/Cas ribonucleoprotein complexes
Xia Li, Wan Shi, Lizhao Geng, Jianping Xu,Syngenta Beijing Innovation Center, Beijing 102206, China通讯作者: 许建平,博士,资深首席科学家,先正达全球基因编辑技术科学主管,研究方向:植物生物技术和植物基因组编辑。E-mail:jianping.xu@syngenta.com
编委: 高彩霞
收稿日期:2020-01-13修回日期:2020-04-27网络出版日期:2020-06-20
Editorial board:
Received:2020-01-13Revised:2020-04-27Online:2020-06-20
作者简介 About authors
李霞,博士,研究方向:植物遗传转化和植物基因组编辑。E-mail:46831067@qq.com。

摘要
CRISPR/Cas (clustered regularly interspaced short palindromic repeats/CRISPR-associated proteins)系统作为一种重要的基因编辑工具,自诞生以来被广泛应用于作物的性状改良。与CRISPR/Cas DNA载体介导的植物基因组编辑相比,CRISPR/Cas核糖核蛋白(CRISPR/Cas ribonucleoprotein, CRISPR/Cas RNP)介导的植物基因组编辑具有作用迅速、脱靶率低和无外源DNA插入(DNA-free)等优点,因而无需清除CRISPR编辑工具而更容易获得纯合的编辑体。但是,由于植物细胞转化方法和细胞再生技术的限制,不借助筛选标记的辅助将CRISPR/Cas RNP直接导入植物细胞并获得高效基因编辑仍比较困难,直接限制了CRISPR/Cas RNP在植物基因组编辑中的广泛应用。本文系统介绍了CRISPR/Cas RNP 基因组编辑技术的分子作用机理及其优势,并总结了CRISPR/Cas RNP导入植物细胞的方法,最后对CRISPR/Cas RNP在植物基因组编辑中的新应用和新思路进行了展望,以期为进一步改进CRISPR/Cas RNP基因组编辑技术和扩大其在作物改良中的应用提供参考。
关键词:
Abstract
The CRISPR/Cas system is the most popular genome editing technology in recent years and has been widely used in crop improvement. Compared with introducing the CRISPR/Cas system into plant cells with DNA constructs, introducing CRISPR/Cas ribonucleoprotein (RNP) to perform genome editing excels in rapid action, low off-target rates and is free of DNA insertions in editing plants. However, efficient delivery of CRISPR/Cas RNP into plant cells and achieving high editing frequency are still very challenging, which limits the extensive implementation of CRISPR/Cas RNP-mediated genome editing in plants. In this review, we summarize the progress of protein and RNP delivery methods in plant cells, and provide new perspectives of further development and future applications of the CRISPR/Cas RNP technology in plant genome editing.
Keywords:
PDF (509KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
李霞, 施皖, 耿立召, 许建平. CRISPR/Cas核糖核蛋白介导的植物基因组编辑. 遗传[J], 2020, 42(6): 556-564 doi:10.16288/j.yczz.20-017
Xia Li.
实现精准、高效的基因组修饰是当今生命科学领域的研究目标之一。近年来,基因组编辑技术,尤其是成簇的规律间隔的短回文重复序列及其相关系统(clustered regularly interspaced short palindromic repeats/CRISPR-associated proteins, CRISPR/Cas)成为实现该目标的最重要工具[1]。CRISPR/Cas系统具有灵活、高效、操作简单等特点,因此在农业生物技术领域被广泛应用于创造新的植物突变群体[2]。但是,利用传统的植物遗传转化方法,如农杆菌或基因枪介导的CRISPR/Cas DNA的转化,在目标基因被编辑的编辑体基因组中通常留有随机插入编码CRISPR/Cas的外源DNA片段,对基因编辑产品的安全和政府监管提出了较大的挑战[3]。因此,无外源DNA(DNA-free)的基因组编辑技术在简化政府监管以及提高大众对基因编辑产品的接受度等方面具有独特的优势,在农业生物技术中具有极高应用价值[4,5]。
CRISPR/Cas核糖核蛋白(CRISPR/Cas ribonucleoprotein, CRISPR/Cas RNP)复合体介导的基因组编辑技术是当前最重要的无外源DNA整合在植物基因组上的编辑技术之一。该技术摒弃了编码Cas蛋白和向导RNA的DNA载体,在试管中将纯化的Cas蛋白和向导RNA预先组装成具备完整活性的CRISPR/Cas RNP复合体,再将CRISPR/Cas RNP通过物理或者化学的方法直接导入植物细胞,从源头上避免了在植物基因组中插入外源DNA[6]。同时,因为导入的是具有完整活性的CRISPR/Cas RNP复合体,CRISPR/Cas RNP进入细胞后可以立即发挥功能,迅速切割靶位点的基因组DNA;又由于Cas蛋白和向导RNA的半衰期均较短,细胞中又没有新的Cas蛋白和向导RNA持续合成,所以CRISPR/Cas RNP对基因组DNA的切割被限定在一个短暂的时间内,得到的编辑产物在嵌合率和脱靶率方面均优于使用编码Cas蛋白和向导RNA的DNA载体得到的编辑产物[6,7]。但是,由于目前植物细胞转化方法和细胞再生技术的限制,在没有筛选标记的情况下,将CRISPR/Cas RNP直接导入植物细胞并高效获得基因编辑体仍比较困难,成本较高,限制了CRISPR/ Cas RNP在植物基因组编辑中的广泛应用[5,8]。本文主要总结了CRISPR/Cas RNP基因组编辑技术的分子机制及优势、CRISPR/Cas RNP导入植物细胞的方法,并对CRISPR/Cas RNP的应用前景进行了展望。
1 CRISPR/Cas RNP基因组编辑技术分子机制
CRISPR/Cas系统广泛存在于原核生物基因组中,是细菌和古细菌为应对病毒攻击而演化形成的获得性免疫防御机制[9,10]。CRISPR序列中存储着若干一定长度的、病毒来源的DNA片段,可经转录产生前体CRISPR RNA (pre-crRNA)。Pre-crRNA进一步被加工成一系列短的含有保守重复序列和间隔区的成熟CRISPR RNA (crRNA),能通过碱基配对识别入侵的病毒DNA。Cas是CRISPR相关蛋白,一些特定的Cas蛋白是受RNA引导的核酸内切酶,即能在crRNA的引导下识别并切割病毒基因组中的特定核酸片段,被切割的病毒基因组受到宿主细胞内核酸酶的影响而被降解或失去功能,使自身免于遭受病毒侵染[1,9,11]。由于CRISPR/Cas系统能够根据需求而设计识别和切割特定的目标DNA序列,近年来成为高效的真核生物基因组编辑工具[12]。在目前已经发现的多个CRISPR/Cas系统中,CRISPR/Cas9[13]和CRISPR/Cas12a (旧称Cpf1[14,15])是在基因组编辑领域应用最广泛的两套系统。1.1 CRISPR/Cas9 RNP系统分子机制
CRISPR/Cas9属于II-A型CRISPR系统[16],具有双分子的向导RNA[17,18]。crRNA通过碱基配对与反式激活CRISPR RNA (trans-activating crRNA, tracrRNA)结合形成杂合双链RNA,可引导Cas9蛋白识别并切断靶标双链DNA。在实际应用中,可将tracrRNA和crRNA人工融合成一条嵌合的单分子向导RNA (single guide RNA, sgRNA)[19],sgRNA能代替天然的双分子向导RNA起到相同的引导作用[18],并大幅提高了切割效率。Cas:sgRNA二元复合体首先扫描目标DNA分子,寻找特定的前间隔序列邻近基序(protospacer adjacent motif, PAM)。该二元复合体定位并结合到PAM上游后,会打开DNA双链形成R环(R-loop),由crRNA的间隔序列(spacer)与目标DNA的前间隔序列(protospacer)配对进行互补性检测。当间隔序列和前间隔序列形成足够稳定的互补配对,Cas9的构象将发生改变,激活两个切割DNA单链的催化域——HNH催化域切割DNA与crRNA互补的链(又称靶向链),同时类RuvC催化域切割非互补链(又称非靶向链),结果造成PAM上游第3和第4个碱基对间引入平末端的DNA双链断裂[18,20]。CRISPR/Cas9 RNP系统即是Cas9:sgRNA二元复合体,CRISPR/Cas9 RNP的构成及其识别和切割双链DNA的机理如图1A所示。在真核细胞中,基因组DNA的双链断裂会被非同源末端连接(non- homologous end-joining, NHEJ)或同源重组介导的修复(homology-directed repair, HDR)途径修复[13]。通过CRISPR/Cas9的定向切割产生的DNA双链断裂和生物体本身的修复途径,可以方便快捷地实现基因敲除、敲入、定点突变等基因组编辑[4,13]。1.2 CRISPR/Cas12a RNP系统分子机制
CRISPR/Cas12a属于V-A型CRISPR系统[16],与CRISPR/Cas9相似,可以定点引入DNA双链断裂,因而也被改造成基因组编辑工具[17]。与CRISPR/ Cas9系统相比,CRISPR/Cas12a系统主要有以下特点:(1) Cas12a天然使用单分子导向RNA,不需要tracrRNA,只需要一个crRNA分子(42个核苷酸),比Cas9的sgRNA (100个核苷酸左右)短很多,因此Cas12a的crRNA更容易通过化学合成获得[14]; (2) Cas12a的基因较小,DNA载体更容易操作; (3) Cas12a切割双链DNA后产生4~5 bp的5′突出末端(staggered overhang),有研究表明这种粘性末端的修复更有利于HDR途径,利于实现HDR介导的精准基因编辑[21];(4)与Cas9识别高GC含量的PAM(NGG)互为补充,Cas12a识别高AT含量的PAM (TTTV或TTV),大大扩展了基因组编辑靶点的范围[22,23];(5) Cas12a同时具有核糖核酸酶(RNase)活性,能对自身的pre-crRNA进行识别并加工获得成熟crRNA,使得在对多个基因组位点进行同时编辑时,向导RNA阵列的设计和实现更简便[24]。与CRISPR/Cas9 RNP相似,CRISPR/Cas12a RNP系统是Cas12a蛋白与crRNA的复合体,CRISPR/Cas12a RNP构成及其识别和切割双链DNA的机理如图1B所示。2 CRISPR/Cas RNP技术应用于植物基因组编辑的优势
目前CRISPR/Cas基因组编辑技术已被广泛应用于作物的性状改良,最常用的技术路线是首先采用农杆菌转化或基因枪轰击的方法将编码CRISPR/ Cas系统的DNA载体导入植物细胞,由DNA载体在植物细胞内转录翻译组装出CRISPR/Cas复合体进行基因编辑,再通过筛选、再生、鉴定得到编辑体植株[4]。在这些转化过程中,CRISPR/Cas系统的编码序列一般会整合到植物基因组中,加之Cas基因多由组成型启动子驱动,这样会导致编辑系统在各个细胞中持续表达,从而增加非靶标位点被突变的风险,即增加了脱靶突变的几率;同时也会增加不同细胞中发生不同类型突变,即增加嵌合突变体的几率[6,25]。CRISPR/Cas RNP则以具有活性的完整复合体直接导入细胞,进入细胞核后即可迅速发挥功能;另一方面由于没有持续的基因表达,导入细胞的CRISPR/Cas RNP会在较短时间内降解,不会随细胞分裂而增殖,并且CRISPR/Cas RNP切割染色体靶位点作用迅速而短暂,能使脱靶率和嵌合率都显著降低[6,7,26,27]。图1
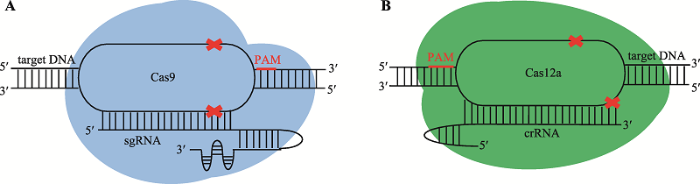 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1CRISPR/Cas RNP系统构成和作用机理
A:CRISPR/Cas9 RNP系统构成和作用机理;B:CRISPR/Cas12a RNP系统构成和作用机理。
Fig. 1Schematics of the components of CRISPR/Cas RNP complexes and the mechanisms of action
此外,DNA载体的片段还可能随机插入到植物基因组各处,产生非预期的基因序列改变[28,29,30],造成潜在的非预期的产品安全问题。多数作物必须额外花费一代或多代的时间,通过自交或回交将这些非预期的外源序列分离出去。而葡萄(Vitis vinifera L.)、马铃薯(Solanum tuberosum L.)和香蕉(Musa acuminata Colla)等无性繁殖的作物,则无法通过自交或杂交分离外源DNA[6]。由于CRISPR/Cas RNP不含任何外源DNA的组分,从源头上避免了外源DNA插入基因组的可能,大大节省了获得无外源DNA插入的基因组编辑植株所需的时间,也使无性繁殖作物中得到无外源DNA插入的基因组编辑植株成为可能[6,30]。
在基因组编辑试剂的前期准备方面,CRISPR/ Cas RNP也比传统的DNA载体更为简便。Cas蛋白可以在大肠杆菌中大量表达和纯化,向导RNA可以通过化学合成或体外转录迅速制备,甚至可以直接订购商业化的成品蛋白和向导RNA,省去了构建和验证载体的大量繁琐工作[31,32]。这一优势在对多靶点同时进行编辑时尤为突出,可直接将Cas蛋白和多条向导RNA按一定比例在室温下混匀孵育后即可进入转化流程,具有极强的灵活性[33,34]。
3 CRISPR/Cas RNP导入植物细胞的方法
蛋白质由于分子量大、结构复杂,且必须维持正确的三维结构才能保证活性,因此将活性蛋白质导入细胞的难度较大。CRISPR/Cas RNP是蛋白质与核酸的功能复合体,空间结构更加复杂,因而导入植物细胞更有挑战性[35]。电穿孔和显微注射技术可以把CRISPR/Cas RNP直接导入动物细胞[33,36,37]。最近研究表明,用于核酸转染细胞的阳离子脂质体也可协助将CRISPR/Cas RNP导入动物细胞[35]:向导RNA所带的强负电荷使CRISPR/Cas RNP整体带负电,与阳离子脂质体通过静电作用结合,形成被脂质体包裹的颗粒,通过胞吞作用进入细胞[35,38]。该方法的优势在于对细胞伤害小,且脂质体包裹保护了CRISPR/Cas RNP的完整性和活性[35]。植物细胞比动物细胞多出一道物理屏障——细胞壁,这就决定了将生物大分子导入植物细胞必须采用与导入动物细胞截然不同的策略[26]。回顾目前已发表的将CRISPR/Cas RNP用于植物基因组编辑的研究,大多数先通过酶解消化细胞壁得到原生质体,然后通过聚乙二醇(polyethylene glycol, PEG)介导的原生质体转化将CRISPR/Cas RNP导入。该方法已经在多种植物细胞中成功实现了CRISPR/Cas RNP介导的基因组编辑[6,39~43](表1)。但由于技术的限制,仅生菜(Lactuca sativa L.)[6]、马铃薯[43]等少数植物种类的原生质体可以再生,大多数植物的原生质体还无法再生成为完整植株[44,45],也就无法通过原生质体得到能够稳定遗传的基因组编辑完整植物株系。但也有研究另辟蹊径,将体外分离的水稻(Oryza sativa L. cv. Nipponbare)精细胞和卵细胞电击融合产生无细胞壁的合子,再以PEG介导将CRISPR/Cas RNP导入进行基因组编辑,被编辑的合子可继续发育成可稳定遗传的完整植株[46]。
除了以PEG介导的RNP复合物导入原生质体的方法外,基因枪也是常用的将CRISPR/Cas RNP复合物导入植物细胞的方法之一。将生物大分子如CRISPR/Cas RNP复合物通过特定的方法包裹在微米甚至纳米级的金粉微粒上,通过基因枪将金粉轰击到玉米(Zea mays L.)[26]、小麦(Triticum aestivum L.)[30]等单子叶作物幼胚或预诱导的愈伤组织中,能够在7~9周内获得再生的由RNP基因编辑的完整植株[47]。阳离子脂质体被认为可以帮助CRISPR/Cas RNP形成更大的复合体,有助于RNP复合体与金粉颗粒的结合,从而提高导入效率[26]。但也有研究表明CRISPR/Cas RNP不需要额外的介质(如阳离子脂质体)协助也能有效结合到金粉微粒上,可以进一步简化实验操作及降低成本[30]。
Table 1
表1
表1利用CRISPR/Cas RNP介导的植物基因组编辑研究
Table 1
| 性状基因 | 细胞类型 | 导入方法 | 编辑工具 | 编辑结果 | 文献来源 |
|---|---|---|---|---|---|
| 油菜素内酯信号途径负调控因子基因(brassinosteroid insensitive 2, BIN2)的同源基因 | 生菜(L. sativa)原生质体Lactuca sativa | PEG | Cas9 | 编辑体占再生植株的46% | [6] |
| 茉莉酸合成途径丙二烯氧化物环化酶基因(allen oxide cyclase, AOC) | 烟草(Nicotiana tabacum L.)原生质体 | PEG | Cas9 | 深度测序编辑频率44% | [6] |
| 光敏色素B基因(phytochrome B, PHYB),油菜素内酯信号途径负调控因子基因(brassinosteroid insensitive 1, BRI1) | 拟南芥(Arabidopsis thaliana L. Heynh.) 原生质体 | PEG | Cas9 | PHYB:深度测序编辑频率16%;BRI1:两条sgRNA同时引导切割,在两个靶点间产生223 bp缺失 | [6]] |
| 细胞色素基因P450,DDB1结合WD40蛋白编码基因DWD1 | 水稻(O. sativa cv. Dongjin)原生质体 | PEG | Cas9 | 深度测序编辑频率:19%(P450)和8.4%(DWD1) | [6]] |
| 硝酸还原酶基因(nitrate reductase, NR) | 矮牵牛(Petunia × hybrida hort. ex E. Vilm.)原生质体 | PEG | Cas9 | 深度测序编辑频率5.3%~17.8% | [39]] |
| 苹果火疫病易感基因DIPM-1,2,4 | 苹果(Malus domestica Borkh. cv. Golden delicious)原生质体 | PEG | Cas9 | 深度测序编辑频率:6.7%(DIPM-1),0.5-3.3%(DIPM-2),2.5-6.9%(DIPM-3) | [40]] |
| 葡萄白粉病易感基因(mildew resistance locus O-7, MLO-7) | 葡萄(V. vinifera cv. Chardonnay)原生质体 | PEG | Cas9 | 深度测序编辑频率0.1% | [40] |
| 叶舌发育相关基因(ligulelss1, LIG1),乙酰乳酸合成酶基因(acetolactate synthase 2, ALS2),雄性不育基因MS26、MS45 | 玉米(Z. mays L.)幼胚 | 基因枪加阳离子脂质体 | Cas9 | 编辑体分别占再生植株的4.0%(MS45)、9.7%(LIG1)和2.4%(MS26);两棵植株在ALS2位点发生同源重组介导的氨基酸精准替换 | [26] |
| 籽粒粒重相关基因(grain width 2, TaGW2),籽粒粒长相关基因TaGASR7 | 小麦(T. aestivum cv. Kenong 199和cv. YZ814)幼胚 | 基因枪 | Cas9 | 编辑体分别占总外植体的4%~5%(cv. Kenong 199)和1%~2%(cv. YZ814) | [30] |
| 脂肪酸去饱和酶基因(fatty acid desaturase, FAD)2-1A、FAD2-1B | 大豆(Glycine max L. Merr.)原生质体 | PEG | Cas12a | 深度测序编辑频率:FAD2-1A 0.0%~11.7%,FAD2-1B 0.0%~9.1% | [41] |
| 丙二烯氧化物环化酶基因AOC | 野生烟草(Nicotiana attenuata Torr. ex S.Watson)原生质体 | PEG | Cas12a | 深度测序编辑频率0.08%~0.8% | [41] |
| 类胡萝卜素合成途径八氢番茄红素脱氢酶基因(phytoene desaturase, PDS)和春化作用调控基因(frigida ,FRI) | 卷心菜(Brassica oleracea var. capitata L. f. alba)、大白菜(B. rapa subsp. pekinensis (Lour.) Hanelt ex Mansf.)和油菜(B. napus L. cv. Topaz)原生质体 | PEG | Cas9 | 深度测序编辑频率:卷心菜0.09%~2.25%(FRI)和0.14%~ 1.33% (PDS),大白菜1.15%~ 24.51%(FRI)和3.78%~24.51% (PDS),油菜0% | [42] |
| 结合态淀粉合成酶基因(granule-bound starch synthase, GBSS) | 马铃薯(S. tuberosum cv. Kuras)原生质体 | PEG | Cas9 | 编辑体占再生植株的:1%~9% (化学合成sgRNA)和22%~ 25% (体外转录sgRNA);4条等位基因全部被编辑的植株占再生植株的2%~3% | [43] |
| 红色荧光蛋白报告基因DsRed2,中脉形成和心皮发育调控基因(drooping leaf, DL),籽粒粒形相关基因(grain width 7, GW7),精卵融合调控基因(generative cell specific-1, GCS1) | 水稻(O. sativa cv. Nipponbare)合子 | PEG | Cas9 | 编辑体植株分别占PEG转化合子数的25% (DsRed2)、13.6%~14.3% (DL)、21.4% (GW7)和64% (GCS1) | [46] |
| 八氢番茄红素脱氢酶基因OsPDS | 水稻(O. sativa cv. Nipponbare)幼胚 | 基因枪 | Cas9 | 编辑体占再生植株(经共转化筛选标记筛选)的33.8% | [8] |
| 八氢番茄红素脱氢酶基因OsPDS | 水稻(O. sativa cv. Nipponbare)幼胚 | 基因枪 | Cas9和Cas12a | 编辑体分别占再生植株(经共转化筛选标记筛选)的3.6%(野生型Cas9)、8.8% (HiFi Cas9)、0.0% (AsCas12a)和23.8% (LbCas12a) | [31] |
| 橙色荧光蛋白报告基因pporRFP | 烟草(N. tabacum cv. Bright Yellow-2)BY-2悬浮细胞系 | 阳离子脂质体和基因枪 | Cas9 | 编辑体占随机挑选细胞团的6% (阳离子脂质体)和3% (基因枪) | [48] |
新窗口打开|下载CSV
4 结语与展望
近年来,随着细胞生物学和材料科学的不断发展,植物细胞转化的新方法层出不穷,这意味着会有更多、更有效的CRISPR/Cas RNP导入细胞的方法。最近新发现的跨细胞膜转运蛋白,如细胞穿透肽(cell-penetrating peptides, CPPs)可能是CRISPR/ Cas RNP导入植物细胞更有效的新方法[49,50]。CPP是一系列长度一般不超过30个氨基酸的短肽,通常净带正电,同时兼有亲水和疏水两性基团。CPP介导的跨膜转运机制目前尚不明确,有时会部分通过胞吞途径,但似乎不依赖于受体和能量。CPP介导的转运优势在于低毒性和无细胞类型的限制,协助转运的大小范围可以从小分子化合物到蛋白甚至是超大分子粒子[51]。目前已有研究利用CPP将β-葡萄糖苷酸酶(β-glucuronidase, GUS)蛋白成功导入到黑小麦(× Triticosecale Witt.)的小孢子中[52],但还未见利用CPP将CRISPR/Cas RNP导入植物细胞的报道。纳米材料也日益成为递送生物分子的常用手段[53]。目前已有研究将纳米材料表面铺敷精氨酸,使纳米材料带上正电,并将谷氨酸肽(E-tag)融合到Cas9蛋白的N端,使Cas9带上负电。带电荷的纳米材料和Cas9-E-tag融合蛋白通过正负电荷作用结合,可携带融合蛋白在一系列不同的细胞类型中获得至高可达90%的导入效率,基因编辑效率最高可达30%[38]。碳纳米管是另一种新兴的纳米材料,能够以无机械辅助的方式高效地将DNA递送到多种植物细胞中,实现无转基因整合的DNA递送和瞬时基因表达[54],结合CRISPR/Cas系统能达到导入CRISPR/Cas RNP同样的效果,也可能成为未来植物遗传转化和基因组编辑的方向之一。
CRISPR/Cas RNP的应用范围也将不断拓宽,不再局限于定点进入DNA双链断裂、后经NHEJ途径修复得到定点突变或经HDR途径修复得到精准编辑。所有CRISPR/Cas系统能够实现的功能,都可以通过向细胞内导入CRISPR/Cas RNP实现,如基于CRISPR/Cas的单碱基编辑器、基因表达调控器等[12,34,55]。目前已有研究将高保真的CRISPR/Cas单碱基编辑器以RNP形式导入哺乳动物细胞中,建立DNA-free的单碱基编辑系统,比质粒转染具有更高的特异性,脱靶率也更低[55,56]。
任何技术都不可避免存在缺陷,CRISPR/Cas RNP介导的植物基因组编辑技术首先亟待解决的问题是实现可稳定遗传的编辑效率仍然较低,这一不足有望通过进一步改造Cas蛋白和向导RNA来提高CRISPR/Cas RNP的稳定性和生物活性[57],以及进一步提高CRISPR/Cas导入细胞特别是细胞核的效率来改进。多项在动物细胞中的研究提示增加融合在Cas蛋白上的核定位信号(nuclear localization signal, NLS)的数量和优化NLS的位置编排可以有效改善CRISPR/Cas RNP的编辑效率[58,59]。另外,CRISPR/Cas RNP的DNA-free特性使得转化过程缺乏有效的筛选标记,后期需再生大量植株,导致组织培养和分子检测的工作量较大[8,26,30]。近期一项在硅藻(Phaeodactylum tricornutum Bohlin.)中的研究巧妙利用CRISPR/Cas RNP多位点同时编辑的灵活性,在编辑目的性状基因的同时编辑内源基因作为筛选标记,为解决DNA-free的筛选问题提示了新方向[60]。随着CRISPR/Cas RNP研究的不断深入和技术的不断改进,其应用会越来越广,必将会给植物基因组编辑技术带来革命性的进步。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 2]
[本文引用: 12]
[本文引用: 2]
[本文引用: 3]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 3]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 6]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 6]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}