 ,安徽省农业科学院水稻研究所,安徽省水稻遗传育种重点实验室,合肥 230031
,安徽省农业科学院水稻研究所,安徽省水稻遗传育种重点实验室,合肥 230031Prime editing creates a novel dimension of plant precise genome editing
Ruiying Qin, Pengcheng Wei,Rice Institute, Anhui Academy of Agriculture Science, Anhui Key Laboratory of Rice Genetics and Breeding, Hefei 230031, China通讯作者: 魏鹏程,博士,研究员,研究方向:作物遗传与育种。E-mail:weipengcheng@gmail.com
编委: 韩玉波
收稿日期:2020-05-2修回日期:2020-05-20网络出版日期:2020-06-20
| 基金资助: |
Editorial board:
Received:2020-05-2Revised:2020-05-20Online:2020-06-20
| Fund supported: |
作者简介 About authors
秦瑞英,硕士,副研究员,研究方向:作物遗传与育种。E-mail:laorui_1023@163.com。

摘要
由于植物细胞内同源重组频率较低、供体传递受限等原因,对植物基因组进行精准编辑十分困难。近期,中国科学院遗传与发育生物学研究所高彩霞团队构建了适用于植物的引导编辑器(plant prime editor, PPE)系统,并在重要作物水稻和小麦中完成了引导编辑。该系统不产生DNA双链断裂,仍可高度准确实现所有可能的12种单碱基替换、多碱基替换及片段缺失插入,从而为植物基因组精确编辑提供了多用途工具。本文介绍了PPE的组成结构和编辑能力,同时也结合其他研究组随后发表的报告综述了植物引导编辑器的优化探索,为合理使用PPEs和继续开展优化工作提供帮助。
关键词:
Abstract
The precise genome editing has not been well established in plants, largely because of the limited frequency of homology recombination and the delivery barrier of donor templates. Recently, Dr. Caixia Gao’s group from the Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, developed a series of plant prime editors (PPEs), which mediats the prime editing in the genomes of rice and wheat. The PPE systems are able to generate all 12 kinds of programmable base substitutions, as well desired multiplex nucleotide substitutions and small deletions or insertions without DNA double-strand breaks, thus providing versatile tools for precise plant genome editing. Herein, we introduce the structure and the editing capacity of the PPEs. The attemp on efficiency enhancements of PPEs and other PPEs are also discussed, which may provide a reference for appropriate application of PPEs in plants and also for continuous optimization of the editing tools.
Keywords:
PDF (443KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
秦瑞英, 魏鹏程. Prime editing引导植物基因组精确编辑新局面. 遗传[J], 2020, 42(6): 519-523 doi:10.16288/j.yczz.20-125
Ruiying Qin.
CRISPR-Cas (clustered regularly interspaced short palindromic repeats/CRISPR-associated proteins)基因编辑系统正日益深刻地影响着植物学研究的发展。该系统利用在植物基因组特定位点产生DNA双链断裂(double strand break, DSB),通过错误倾向性的非同源末端连接(non-homologous end joining, NHEJ)机制可修复失活的目标基因[1]。由于NHEJ修复介导的碱基插入缺失(InDel)存在一定的随机性,因此难以实现精确的基因组编辑。尽管借助于同源介导修复(homology-directed repair, HDR)机制,CRISPR-Cas系统可在外源DNA供体(donor)的指导下实现精准的碱基替换或片段插入缺失[2],但是在植物细胞中,由于同源重组频率偏低和DNA供体递送困难,CRISPR介导的HDR效率显著受限,往往也难以实现高效的基因组精确编辑[1,3]。碱基编辑(base editing)是新近发展的不依赖于HDR而介导基因组精确碱基替换的新型基因编辑技术[4]。目前碱基编辑工具包括胞嘧啶碱基编辑器(cytosine base editor, CBE)和腺嘌呤碱基编辑器(adenine base editor, ABE)两类。CBE工具是以剪切活性受损的Cas蛋白(如Cas9 nickase, nCas9或catalytically dead Cas12a, dCas12a等)融合胞嘧啶脱氨酶(如rAPOBEC1、PmCDA、hAID1或hAPOBEC3等)发展而来;而ABE工具则多是利用定向进化的腺嘌呤脱氨酶TadA*7.10或TadA-8e融合nCas9构成。在多种植物中,CBE和ABE均可有效介导G∙C-to-A∙T的碱基转换,为植物基因组精确编辑提供了新的有效策略[1]。但是,碱基编辑系统也存在着一定问题和局限性[5]。一方面,现有的碱基编辑器仅能完成特定的4种形式的碱基转换,缺乏完成精确可控的碱基颠换或者片段插入缺失等其他基因组编辑的能力;另一方面,碱基编辑往往仅能在向导RNA (guide RNA)识别区域中有限编辑窗口内发挥高效作用,编辑区域受到编辑器Cas蛋白PAM (protospacer adjacent motif)识别能力和编辑器窗口范围的双重限制,因此无法编辑基因组中一些重要位点。此外,碱基编辑器自身编辑的精确性还有待进一步优化。例如,编辑器脱氨酶,特别是CBE的胞嘧啶脱氨酶,存在序列非特异活性,可能在全基因组范围出现脱靶[6,7];同时当编辑窗口内存在多个可编辑碱基时,编辑器在编辑目标碱基时也会造成其他非靶向的碱基突变(bystander mutation)[8]。
2019年底,美国哈佛大学David Liu研究组报道了不同于碱基编辑的基因组精确编辑技术,即引导编辑(prime editing)系统[8]。该系统利用nSpCas9 (H840A)和工程化改造M-MLV RT逆转录酶(moloney murine leukemia virus reverse transcriptase)融合构建引导编辑器(prime editor),由pegRNA (prime editing guide RNA)中guide RNA部分引导在人细胞基因组靶位置附近形成编辑链上的单链切口,进而通过pegRNA中的PBS(primer binding site)序列引导以含有目标编辑序列的逆转录模板(RT template)将突变精确导入基因组中(图1)。相对于CRISPR介导、依赖于HDR的精确编辑,引导编辑的效率更高,副产物更少。而相对于碱基编辑,引导编辑也表现出明显的优势:(1)引导编辑可完成的编辑类型更为广泛,不仅可以实现12种任意类型的碱基替换,还可精确高效地导入44 bp以内的小片段插入、80 bp以内的片段缺失以及复杂形式混合突变;(2)引导编辑受PAM的限制相对较小,在长达33 nt的PAM远端序列内,多个位点都可以高效编辑;(3)引导编辑更为精准,当在目标位点附近有多个相同碱基时,不会受到碱基编辑器bystander mutation问题的困扰。在此基础上,中国科学院遗传与发育生物学研究所高彩霞团队构建了适于植物表达的引导编辑工具,并成功地在水稻(Oryza sativa L.)和小麦(Triticum aestivum L.)中完成了DNA精确编辑[3]。这项工作创制了灵活、多用途的植物精确编辑工具,为植物基因组编辑提供了新路径。
图1
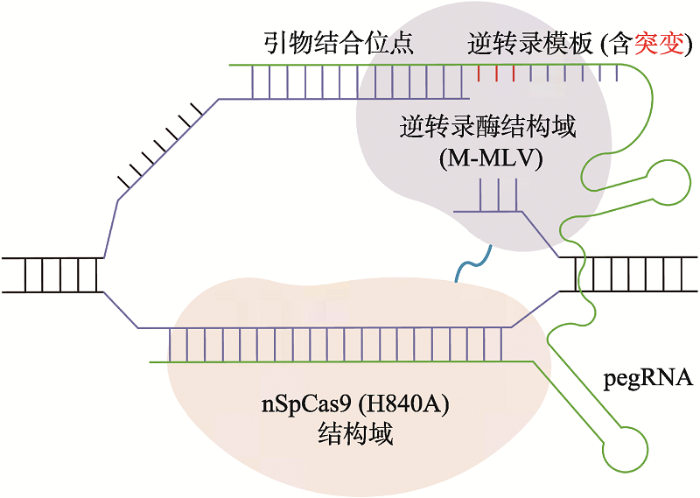 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1引导编辑工作模型
引导编辑器由DNA切割活性部分缺失的nSpCas9 (H840A)和通过逆转录负责修复的M-MLV两个结构域融合而成。在非靶标链产生单链缺口后,引导编辑器在引物结合位点序列帮助下结合在断裂位点,利用逆转录活性在逆转录模板的指导下修复单链断裂并导入目标突变。
Fig. 1Model of prime editing
1 引导编辑可用于植物细胞内DNA编辑
植物引导编辑器(plant prime editor, PPE)由nCas9 (H840A)与M-MLV RT变体(M-MLV RT D200N/L603W/T330P/T306K/W313F)的融合基因经植物偏好密码子优化而来。与用于动物细胞编辑的引导编辑系统类似[8],PPE系统可根据表达向导 RNA的不同分为PPE2、PPE3和PPE3b三类。PPE2仅表达pegRNA用于定位靶点和引导编辑;PPE3则同时表达pegRNA和额外的、靶向非编辑链的sgRNA (nicking sgRNA),在PPE导入突变形成不完全配对的DNA双链后,nicking sgRNA可通过剪切非编辑链提高以编辑链为模板修复的概率从而增强编辑效率。由于nicking sgRNA引导的单链缺刻可能产生插入缺失突变而降低了编辑的精确度,因此利用非完全匹配sgRNA构建的PPE3b系统则在增强引导编辑效率的同时避免InDel的过多产生。利用水稻原生质体瞬时表达系统,PPE3b可精确编辑蓝色荧光蛋白(blue fluorescent protein, BFP)产生CC-to-GT突变而将其转换为绿色荧光蛋白(green fluorescent protein, GFP)。在该系统中,PPE3b的编辑效率约为4.4%。PPE2和PPE3/3b也可编辑植物基因组位点。在水稻原生质体中,利用PPE精确导入的6 bp片段删除效率可达8.2%;而3 bp碱基插入的效率约为2.0%。在水稻或小麦基因组的多个位点,PPE均能完成不同形式的碱基转换或颠换,充分展示了引导编辑的广适性和灵活性。有趣的是,与人源细胞不同,植物细胞中的PPE3/3b的效率较PPE2并没有显著增强,暗示着植物引导编辑可能具有一定特殊性。进一步在水稻原生质体中的研究利用 PPE3在4个基因组位点实现了所有12种可能的碱基替换,效率在0.2%~8.0%之间。此外,PPEs也可在水稻基因组不同位点完成多种不同的多碱基替换、小片段缺失和插入等形式的精确编辑。利用农杆菌介导的稳定转化,PPE3可在水稻再生株系中精确导入单碱基、多碱基替换、目标序列插入或删除。因此,PPE系统是有效的精准编辑工具,为不受限制的开展植物基因组精确编辑提供了新路径。值得注意的是,成都电子科技大学张勇团队、中国农业科学院作物研究所夏兰琴团队、北京市农林科学院杨进孝团队、中国科学院上海植物逆境生物学研究中心朱健康团队、沙特阿拉伯阿卜杜拉国王科技大学Mahfouz团队以及本团队也陆续报道了类似结构的植物PE编辑工具[9,10,11,12,13,14]。通过瞬时表达或稳定表达,这些工具在植物细胞中也均可精确的编辑外源转入DNA或内源基因组靶标,为PPE系统的植物基因组编辑能力提供了补充和验证。
2 PPE的优化探索
编辑系统的产物纯度和效率直接影响方法的实用性,是需要着重优化的指标。例如,早期开发的CBE类碱基编辑工具常产生InDel或非C-to-T的编辑副产物,严重影响了编辑能力[15]。PPE也可能产生编辑副产物,主要体现为pegRNA骨架序列插入或替换编辑位点序列,但一般频率不高(0.5%~4.9%),对于植物基因组精确编辑影响不大。而编辑效率是PPE系统迫切需要改进的主要方面。一方面,在原生质体瞬时表达系统中,PPE对BFP上CC-to-GT突变的诱导率(4.4%)小于植物碱基编辑器PBE (plant base editing) (6.6%),在水稻基因组位点上PPEs的效率也低于PBE,暗示着植物引导编辑系统效率相对受限。由PPE3诱导的稳定编辑株系均为嵌合突变而没有纯合突变,也表明PPE的效率需要继续优化。另一方面,与动物引导编辑系统类似,植物PPE效率与PBS长度、RT template长度、nicking sgRNA的选择乃至导入突变类型等参数高度相关[3,12,13],使用现有效率相对受限的PPE工具编辑基因组特定位点需要构建和测试不同结构的pegRNA和nicking RNA,因此急需高效稳定的PPE系统降低时间和经济成本。实验证据表明,在一些Cas9可高效编辑的位点PPE效率反而相对受限[6]。因此,可以怀疑在RT活性不足导致精确编辑效率偏低。由于植物的培养温度往往低于M-MLV的最适反应温度,因此在编辑过程中提高温度可能增加效率。 相对于常规的26℃培养,升温至37℃能够显著的提升PPE3b在一些靶点的编辑效率[3]。但是也有独立报道表明,相对于32℃,37℃培养并不能明显提升编辑效率[12],这可能是由于处理方式、材料自身条件、靶位点序列结构等多种原因造成。这些结果也暗示着通过优化培养条件提高prime editing效率是可行的,但仍然需要大量探索。利用不同来源或进化版本的脱氨酶是提高碱基编辑器编辑能力的常用策略[15]。与之类似,尝试不同的RT显然是优化PPE的思路之一。然而,尽管利用花椰菜花叶病毒逆转录酶(cauliflower mosaic virus RT, RT-CaMV)或大肠杆菌retron-derived RT (RT-retron)替代PPE中M-MLV RT构建的PPE- CaMV和PPE-retron可将外源转入的BFP编辑为GFP,但其效率仅接近或略低于PPE3b;同时在水稻基因组靶点上,PPE-CaMV也并没有表现出稳定的高出PPE的活性。
在植物CRISPR系统中,利用Pol II启动子驱动的基于核酶(ribozyme)加工的sgRNA表达策略往往能获得较高编辑效率[16];在植物引导编辑中利用该策略构建了PPE-ribozyme (PPE-R)系统。在水稻基因组的部分位点,PPE-R也可显著提高编辑效率,但是在有些位点编辑反而受到抑制。单一Pol III启动子驱动tRNA串联sgRNA阵列也是植物CRISPR基因编辑常用的sgRNA表达优化手段[17];这种模式也被用于植物引导编辑系统中pegRNA和nicking sgRNA的表达[11,12]。与PPE-R相似,tRNA串联在部分靶点具有一定的效率提升作用,但是并不能在所有位点将植物引导编辑提升到较高水平。另一方面,植物基因编辑效率与编辑器蛋白表达量也密切相关。基于失活抗性标记的替代编辑系统(surrogate system)或编辑器-抗性标记融合表达系统(如ABE-HPT)可通过抗性筛选富集可能出现编辑的植物细胞,从而提高获得碱基编辑株系的概率[18,19]。在部分位点,这些策略同样能一定程度的富集引导编辑株系。但是可能由于引导编辑器活性低于碱基编辑器,富集策略对植物引导编辑的帮助远低于碱基编辑[13,14]。
3 结语与展望
2013年以来,尽管植物基因编辑技术已取得了长足进步,但多数编辑案例是通过靶向突变实现的。相对于高效稳定的基因组靶向突变工具,植物精准编辑工具往往在编辑形式或效率上存在明显缺陷。PPE系统在植物中实现了其他编辑工具无法完成的多种精准突变,极大增强了植物基因组编辑能力,使得在植物功能基因组研究和作物定向育种改良广泛应用精准编辑成为可能。目前,植物引导编辑器在部分基因组位点上还存在效率不足等问题,但这些问题可能从优化编辑器表达条件、筛选或进化更加适于植物系统的逆转录工具、以及适配pegRNA结构相对简单的CRISPR-Cas系统等角度加以解决。相信随着编辑条件的持续优化和编辑器系统的反复迭代,植物引导编辑系统将快速实用化,为在植物基因组精确编辑技术的开发和利用打开新局面。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 3]
[本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 3]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 2]
[本文引用: 4]
URL [本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}