Progress on spermatogonial stem cells of large animals
Xin Zhao, Huaqiang Yang,National Engineering Research Center for Breeding Swine Industry, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
Abstract Spermatogonial stem cells (SSCs) are male germline stem cells that reside in the basement membrane of the seminiferous tubule in the testis. SSCs are characterized by their capability of self-renewal to maintain the stem cell pool throughout the lifespan and commitments to germ line after puberty, thus transmitting the genetic information from parents to the SSC-derived progenies. SSCs can be isolated from testis, propagated in vitro, and induced to differentiate into varied germ cells. Although significant progress has been made in the field of rodent SSCs, the SSCs of large animals have advanced slowly. Studies on SSCs of large animal models can offer insights into the physiological and pathological mechanism of human reproduction. Moreover, SSCs of agricultural large animals can be used as an essential tool for multiplication of elite animal individuals, and generation of genetically modified livestock with valuable economic traits. In this review, we summarize the recent progress on SSCs of large animal models for agricultural and medical purposes, and discuss the present problems and future prospects. This review can give an overall view of large animal SSCs as respect to their applications in novel alternative reproductive technologies, generation of transgenic animals, treatment of male infertility and regenerative medicine. Keywords:spermatogonial stem cell;large animals;cell transplantation;spermatogenesis
PDF (563KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 赵鑫, 杨化强. 大动物精原干细胞研究进展[J]. 遗传, 2019, 41(8): 686-702 doi:10.16288/j.yczz.19-167 Xin Zhao, Huaqiang Yang. Progress on spermatogonial stem cells of large animals[J]. Hereditas(Beijing), 2019, 41(8): 686-702 doi:10.16288/j.yczz.19-167
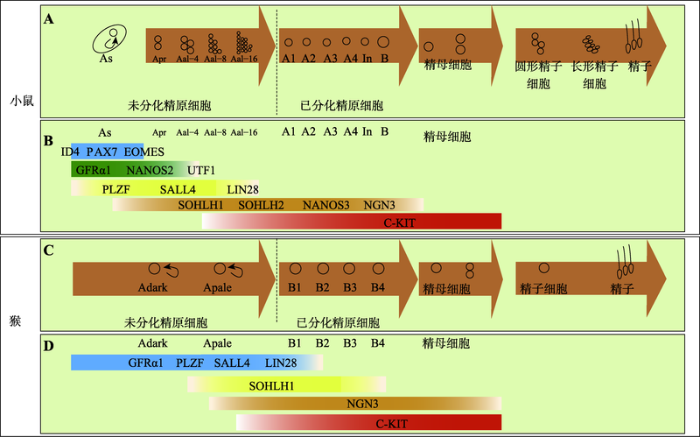
A:小鼠精原干细胞的体内发育谱系;B:小鼠体内精原干细胞不同发育阶段相对应的分子标记;C:猴精原干细胞的体内发育谱系;D:猴体内精原干细胞不同发育阶段相对应的分子标记。PGCs在胚胎的发育过程中迁移至生殖嵴后分化为生殖母细胞,生殖母细胞增殖一段时间后进入静息状态,在出生后第1周作为SSCs恢复生理活动。随着生长发育的进行,SSCs不断地增殖与分化以维持精子发生的稳态。根据参考文献[2, 4, 8, 10, 11]修改绘制。 Fig. 1Schematic of typical spermatogenesis and corresponding molecular markers in some animals
ZhengY, ZhangY, QuR, HeY, TianX, ZengW . Spermatogonial stem cells from domestic animals: progress and prospects Reproduction, 2014,147(3):R65-74. [本文引用: 6]
SahareMG, Suyatno, ImaiH . Recent advances of in vitro culture systems for spermatogonial stem cells in mammals Reprod Med Biol, 2018,17(2):134-142. [本文引用: 3]
GonzálezR, DobrinskiI . Beyond the mouse monopoly: studying the male germ line in domestic animal models ILAR J, 2015,56(1):83-98. [本文引用: 3]
FayomiAP, OrwigKE . Spermatogonial stem cells and spermatogenesis in mice, monkeys and men Stem Cell Res, 2018,29:207-214. [本文引用: 6]
StaubC, JohnsonL . Review: Spermatogenesis in the bull Animal, 2018,12(s1):s27-s35. [本文引用: 1]
McleanDJ . Spermatogonial stem cell transplantation and testicular function Cell Tissue Res, 2005,322(1):21-31. [本文引用: 1]
De RooijDG . The nature and dynamics of spermatogonial stem cells Development, 2017,144(17):3022-3030. [本文引用: 2]
OliveV, CuzinF . The spermatogonial stem cell: from basic knowledge to transgenic technology Int J Biochem Cell Biol, 2005,37(2):246-250. [本文引用: 1]
BoitaniC, Di PersioS, EspositoV, ViciniE . Spermatogonial cells: mouse, monkey and man comparison Semin Cell Dev Biol, 2016,59:79-88. [本文引用: 2]
SuzukiH, AhnHW, ChuT, BowdenW, GasseiK, OrwigK, RajkovicA . SOHLH1 and SOHLH2 coordinate spermatogonial differentiation Dev Biol, 2012,361(2):301-312. [本文引用: 1]
HuleihelM, NourashrafeddinS, PlantTM . Application of three-dimensional culture systems to study mammalian spermatogenesis, with an emphasis on the rhesus monkey (Macaca mulatta) Asian J Androl, 2015,17(6):972-980.
AziziH, Ghasemi HamidabadiH, SkutellaT . Differential proliferation effects after Short-Term cultivation of mouse spermatogonial stem cells on different feeder layers Cell J, 2019,21(2):186-193. [本文引用: 1]
GoodyearS, BrinsterR . Isolation of the spermatogonial stem cell-containing fraction from testes Cold Spring Harb Protoc, 2017, 2017(4): pdb. prot094185. [本文引用: 1]
BuaasFW, KirshAL, SharmaM, McleanDJ, MorrisJL, GriswoldMD, De RooijDG, BraunRE . Plzf is required in adult male germ cells for stem cell self-renewal Nat Genet, 2004,36(6):647-652. [本文引用: 1]
GoelS, SugimotoM, MinamiN, YamadaM, KumeS, ImaiH . Identification, isolation, and in vitro culture of porcine gonocytes Biol Reprod, 2007,77(1):127-137. [本文引用: 1]
RedingSC, StepnoskiAL, CloningerEW, OatleyJM . THY1 is a conserved marker of undifferentiated spermatogonia in the pre-pubertal bull testis Reproduction, 2010,139(5):893-903. [本文引用: 2]
BorjiginU, DaveyR, HuttonK, HerridM . Expression of promyelocytic leukaemia zinc-finger in ovine testis and its application in evaluating the enrichment efficiency of differential plating Reprod Fertil Dev, 2010,22(5):733-742. [本文引用: 1]
SongW, ZhuH, LiM, LiN, WuJ, MuH, YaoX, HanW, LiuW, HuaJ . Promyelocytic leukaemia zinc finger maintains self-renewal of male germline stem cells (mGSCs) and its expression pattern in dairy goat testis Cell Prolif, 2013,46(4):457-468. [本文引用: 1]
LuoJ, MegeeS, DobrinskiI . Asymmetric distribution of UCH-L1 in spermatogonia is associated with maintenance and differentiation of spermatogonial stem cells J Cell Physiol, 2009,220(2):460-468. [本文引用: 1]
ZhouQ, GuoY, ZhengB, ShaoB, JiangM, WangG, ZhouT, WangL, ZhouZ, GuoX, HuangX . Establishment of a proteome profile and identification of molecular markers for mouse spermatogonial stem cells J Cell Mol Med, 2015,19(3):521-534. [本文引用: 1]
ZhangP, QinY, ZhengY, ZengW . Phospholipase D family member 6 is a surface marker for enrichment of undifferentiated spermatogonia in prepubertal boars Stem Cells Dev, 2018,27(1):55-64. [本文引用: 2]
BuageawA, SukhwaniM, Ben-YehudahA, EhmckeJ, RaweVY, PholpramoolC, OrwigKE, SchlattS . GDNF family receptor alpha1 phenotype of spermatogonial stem cells in immature mouse testes Biol Reprod, 2005,73(5):1011-1016. [本文引用: 1]
KimYH, ChoiYR, KimBJ, JungSE, KimSM, JinJH, YunMH, KimSU, KimYH, HwangS, PangMG, RyuBY . GDNF family receptor alpha 1 is a reliable marker of undifferentiated germ cells in bulls Theriogenology, 2019,132:172-181. [本文引用: 1]
LeeR, LeeWY, ParkHJ, HaWT, WooJS, ChungHJ, LeeJH, HongK, SongH . Stage-specific expression of DDX4 and c-kit at different developmental stages of the porcine testis Anim Reprod Sci, 2018,190:18-26. [本文引用: 1]
AlmuniaJ, NakamuraK, MurakamiM, TakashimaS, TakasuM . Characterization of domestic pig spermatogenesis using spermatogonial stem cell markers in the early months of life Theriogenology, 2018,107:154-161.
FujiharaM, KimSM, MinamiN, YamadaM, ImaiH . Characterization and in vitro culture of male germ cells from developing bovine testis J Reprod Dev, 2011,57(3):355-364.
GoelS, ReddyN, MandalS, FujiharaM, KimSM, ImaiH . Spermatogonia-specific proteins expressed in prepubertal buffalo (Bubalus bubalis) testis and their utilization for isolation and in vitro cultivation of spermatogonia Theriogenology, 2010,74(7):1221-1232. [本文引用: 1]
HerridM, NagyP, JuhaszJ, MorrellJM, BillahM, KhazanehdariK, SkidmoreJA . Donor sperm production in heterologous recipients by testis germ cell transplantation in the dromedary camel Reprod Fertil Dev, 2019,31(3):538-546. [本文引用: 1]
KimYH, KangHG, KimBJ, JungSE, KarmakarPC, KimSM, HwangS, RyuBY . Enrichment and in vitro culture of spermatogonial stem cells from Pre-pubertal monkey testes Tissue Eng Regen Med, 2017,14(5):557-566. [本文引用: 2]
LeeKH, LeeWY, KimJH, YoonMJ, KimNH, KimJH, UhmSJ, KimDH, ChungHJ, SongH . Characterization of GFRα-1-positive and GFRα-1-negative spermatogonia in neonatal pig testis Reprod Domest Anim, 2013,48(6):954-960.
ZhengY, HeY, AnJ, QinJ, WangY, ZhangY, TianX, ZengW . THY1 is a surface marker of porcine gonocytes Reprod Fertil Dev, 2014,26(4):533-539.
RafeeqiT, KaulG . Isolation and enrichment of type A spermatogonia from pre-pubertal buffalo (Bubalus bubalis) testis Andrologia, 2013,45(3):195-203.
AbbasiH, TahmoorespurM, HosseiniSM, NasiriZ, BahadoraniM, HajianM, NasiriMR, Nasr-EsfahaniMH . THY1 as a reliable marker for enrichment of undifferentiated spermatogonia in the goat Theriogenology, 2013,80(8):923-932.
KimYH, KimBJ, KimBG, LeeYA, KimKJ, ChungHJ, HwangS, WooJS, ParkJK, SchmidtJA, Pang MG, RyuBY . Stage-specific embryonic antigen-1 expression by undifferentiated spermatogonia in the prepubertal boar testis J Anim Sci, 2013,91(7):3143-3154. [本文引用: 1]
GoelS, FujiharaM, TsuchiyaK, TakagiY, MinamiN, YamadaM, ImaiH . Multipotential ability of primitive germ cells from neonatal pig testis cultured in vitro Reprod Fertil Dev, 2009,21(5):696-708.
De BarrosFR, WorstRA, SaurinGC, MendesCM, AssumpçãoME, VisintinJA . α-6 integrin expression in bovine spermatogonial cells purified by discontinuous Percoll density gradient Reprod Domest Anim, 2012,47(6):887-890.
LuoJ, MegeeS, RathiR, DobrinskiI . Protein gene product 9.5 is a spermatogonia-specific marker in the pig testis: Application to enrichment and culture of porcine spermatogonia Mol Reprod Dev, 2006,73(12):1531-1540.
HerridM, DaveyRJ, HillJR . Characterization of germ cells from pre-pubertal bull calves in preparation for germ cell transplantation Cell Tissue Res, 2007,330(2):321-329.
Rodriguez-SosaJR, DobsonH, HahnelA . Isolation and transplantation of spermatogonia in sheep Theriogenology, 2006,66(9):2091-2103.
HeidariB, Rahmati-AhmadabadiM, AkhondiMM, ZarnaniAH, Jeddi-TehraniM, ShiraziA, NaderiMM, BehzadiB . Isolation, identification, and culture of goat spermatogonial stem cells using c-kit and PGP9.5 markers J Assist Reprod Genet, 2012,29(10):1029-1038.
ZengW, TangL, BondarevaA, LuoJ, MegeeSO, ModelskiM, BlashS, MelicanDT, DestrempesMM, OvertonSA, GavinWG, AyresS, EchelardY, DobrinskiI . Non-viral transfection of goat germline stem cells by nucleofection results in production of transgenic sperm after germ cell transplantation Mol Reprod Dev, 2012,79(4):255-261.
ParkMH, ParkJE, KimMS, LeeKY, ParkHJ, YunJI, ChoiJH, LeeE, LeeST . Development of a high-yield technique to isolate spermatogonial stem cells from porcine testes J Assist Reprod Genet, 2014,31(8):983-991.
OatleyJM, BrinsterRL . The germline stem cell niche unit in mammalian testes Physiol Rev, 2012,92(2):577-595. [本文引用: 2]
ShinoharaT, OrwigKE, AvarbockMR, BrinsterRL . Remodeling of the postnatal mouse testis is accompanied by dramatic changes in stem cell number and niche accessibility Proc Natl Acad Sci USA, 2001,98(11):6186-6191. [本文引用: 1]
KokkinakiM, LeeTL, HeZ, JiangJ, GolestanehN, HofmannMC, ChanWY, DymM . The molecular signature of spermatogonial stem/progenitor cells in the 6-day-old mouse testis Biol Reprod, 2009,80(4):707-717. [本文引用: 1]
SachulaWu, GuTY, HeZY, HerridM, WangX, UyunbiligBorjigin . Advances in the research of mammalian spermatogonial stem cells Chin J Cell Biol, 2014,36(3):392-399. [本文引用: 1]
GriswoldMD . 50 years of spermatogenesis: Sertoli cells and their interactions with germ cells Biol Reprod, 2018,99(1):87-100. [本文引用: 1]
OatleyJM, OatleyMJ, AvarbockMR, TobiasJW, BrinsterRL . Colony stimulating factor 1 is an extrinsic stimulator of mouse spermatogonial stem cell self-renewal Development, 2009,136(7):1191-1199. [本文引用: 2]
PotterSJ, DefalcoT . Role of the testis interstitial compartment in spermatogonial stem cell function Reproduction, 2017,153(4):R151-R162. [本文引用: 1]
MeistrichML, ShettyG . The new director of "the spermatogonial niche": introducing the peritubular macrophage Cell Rep, 2015,12(7):1069-1070. [本文引用: 1]
IzadyarF, Den OudenK, CreemersLB, PosthumaG, ParvinenM, De RooijDG . Proliferation and differentiation of bovine type A spermatogonia during long-term culture Biol Reprod, 2003,68(1):272-281. [本文引用: 1]
TiptanavattanaN, TechakumphuM, TharasanitT . Simplified isolation and enrichment of spermatogonial stem-like cells from pubertal domestic cats (Felis catus) J Vet Med Sci, 2015,77(11):1347-1353. [本文引用: 1]
KubotaH, BrinsterRL . Culture of rodent spermatogonial stem cells, male germline stem cells of the postnatal animal Methods Cell Biol, 2008,86:59-84. [本文引用: 1]
Kanatsu-ShinoharaM, OgonukiN, InoueK, MikiH, OguraA, ToyokuniS, ShinoharaT . Long-term proliferation in culture and germline transmission of mouse male germline stem cells Biol Reprod, 2003,69(2):612-616. [本文引用: 1]
WeiX, JiaY, XueY, GengL, WangM, LiL, WangM, ZhangX, WuX . GDNF-expressing STO feeder layer supports the long-term propagation of undifferentiated mouse spermatogonia with stem cell properties Sci Rep, 2016,6:36779. [本文引用: 1]
ClouthierDE, AvarbockMR, MaikaSD, HammerRE, BrinsterRL . Rat spermatogenesis in mouse testis Nature, 1996,381(6581):418-421. [本文引用: 2]
DobrinskiI, AvarbockMR, BrinsterRL . Germ cell transplantation from large domestic animals into mouse testes Mol Reprod Dev, 2000,57(3):270-279. [本文引用: 2]
OgawaT, DobrinskiI, AvarbockMR, BrinsterRL . Xenogeneic spermatogenesis following transplantation of hamster germ cells to mouse testes Biol Reprod, 1999,60(2):515-521. [本文引用: 1]
KaurG, LongCR, DufourJM . Genetically engineered immune privileged sertoli cells: a new road to cell based gene therapy Spermatogenesis, 2012,2(1):23-31. [本文引用: 1]
SavvulidiF, PtacekM, Savvulidi VargovaK, StadnikL . Manipulation of spermatogonial stem cells in livestock species J Anim Sci Biotechno, 2019,10:46. [本文引用: 2]
GiassettiMI, CiccarelliM, OatleyJM . Spermatogonial stem cell transplantation: insights and outlook for domestic animals Annu Rev Anim Biosci, 2019,7:385-401. [本文引用: 3]
OgawaT, AréchagaJM, AvarbockMR, BrinsterRL . Transplantation of testis germinal cells into mouse seminiferous tubules Int J Dev Biol, 1997,41(1):111-122. [本文引用: 2]
HonaramoozA, BehboodiE, HauslerCL, BlashS, AyresS, AzumaC, EchelardY, DobrinskiI . Depletion of endogenous germ cells in male pigs and goats in preparation for germ cell transplantation J Androl, 2005,26(6):698-705. [本文引用: 2]
IzadyarF, Den OudenK, StoutTA, StoutJ, CoretJ, LankveldDP, SpoormakersTJ, ColenbranderB, OldenbroekJK, Van DerPloeg KD, WoeldersH, KalHB, De RooijDG . Autologous and homologous transplantation of bovine spermatogonial stem cells Reproduction, 2003,126(6):765-774.
SchlattS, RosiepenG, WeinbauerGF, RolfC, BrookPF, NieschlagE . Germ cell transfer into rat, bovine, monkey and human testes Hum Reprod, 1999,14(1):144-150. [本文引用: 1]
OatleyJM . Recent advances for spermatogonial stem cell transplantation in livestock Reprod Fertil Dev, 2017,30(1):44-49. [本文引用: 3]
OgawaT, DobrinskiI, BrinsterRL . Recipient preparation is critical for spermatogonial transplantation in the rat Tissue Cell, 1999,31(5):461-472. [本文引用: 2]
LinZ, BaoJ, KongQ, BaiY, LuoF, SongyangZ, WuY, HuangJ . Effective production of recipient male pigs for spermatogonial stem cell transplantation by intratesticular injection with busulfan Theriogenology, 2017, 89: 365-373. e2. [本文引用: 1]
CreemersLB, MengX, Den OudenK, van PeltAM, IzadyarF, SantoroM, SariolaH, de RooijDG . Transplantation of germ cells from glial cell line-derived neurotrophic factor-overexpressing mice to host testes depleted of endogenous spermatogenesis by fractionated irradiation Biol Reprod, 2002,66(6):1579-1584. [本文引用: 1]
ParkKE, KaucherAV, PowellA, WaqasMS, SandmaierSE, OatleyMJ, ParkCH, TibaryA, DonovanDM, BlombergLA, LillicoSG, WhitelawCB, MilehamA, TeluguBP, OatleyJM . Generation of germline ablated male pigs by CRISPR/Cas9 editing of the NANOS2 gene Sci Rep, 2017,7:40176. [本文引用: 2]
TsudaM, SasaokaY, KisoM, AbeK, HaraguchiS, KobayashiS, SagaY . Conserved role of nanos proteins in germ cell development Science, 2003,301(5637):1239-1241. [本文引用: 1]
SuzukiA, TsudaM, SagaY . Functional redundancy among Nanos proteins and a distinct role of Nanos2 during male germ cell development Development, 2007,134(1):77-83. [本文引用: 1]
WangX, ChenT, ZhangY, LiB, XuQ, SongC . Isolation and culture of pig spermatogonial stem cells and their in vitro differentiation into Neuron-like cells and adipocytes Int J Mol Sci, 2015,16(11):26333-26346. [本文引用: 2]
ParkJE, ParkMH, KimMS, ParkYR, YunJI, CheongHT, KimM, ChoiJH, LeeE, LeeST . Porcine spermatogonial stem cells self-renew effectively in a three dimensional culture microenvironment Cell Biol Int, 2017,41(12):1316-1324. [本文引用: 1]
KimBG, KimYH, LeeYA, KimBJ, KimKJ, JungSE, ChungHJ, HwangS, ChoiSH, Kim MJ, KimDH, KimIC, KimMK, KimNH, KimCG, RyuBY . Production of transgenic spermatozoa by lentiviral transduction and transplantation of porcine spermatogonial stem cells Tissue Eng Regen Med, 2014,11(6):458-466. [本文引用: 1]
ApontePM, SodaT, TeerdsKJ, MizrakSC, van de KantHJ, de RooijDG . Propagation of bovine spermatogonial stem cells in vitro Reproduction, 2008,136(5):543-557. [本文引用: 1]
HerridM, VignarajanS, DaveyR, DobrinskiI, HillJR . Successful transplantation of bovine testicular cells to heterologous recipients Reproduction, 2006,132(4):617-624. [本文引用: 1]
StockwellS, HerridM, DaveyR, BrownleeA, HuttonK, HillJR . Microsatellite detection of donor-derived sperm DNA following germ cell transplantation in cattle Reprod Fertil Dev, 2009,21(3):462-468. [本文引用: 1]
HonaramoozA, BehboodiE, MegeeSO, OvertonSA, Galantino-HomerH, EchelardY, DobrinskiI . Fertility and germline transmission of donor haplotype following germ cell transplantation in immunocompetent goats Biol Reprod, 2003,69(4):1260-1264. [本文引用: 1]
O'brienSJ, JohnsonW, DriscollC, PontiusJ, Pecon-SlatteryJ, Menotti-RaymondM . State of cat genomics Trends Genet, 2008,24(6):268-279. [本文引用: 1]
SilvaAF, Escada-RebeloS, AmaralS, TavaresRS, SchlattS, Ramalho-SantosJ, MotaPC . Can we induce spermatogenesis in the domestic cat using an in vitro tissue culture approach? PLoS One, 2018,13(2):e0191912. [本文引用: 1]
SilvaRC, CostaGM, LacerdaSM, BatlouniSR, SoaresJM, AvelarGF, BöttgerKB, Silva SFJr, NogueiraMS, AndradeLM, FrançaLR . Germ cell transplantation in felids: a potential approach to preserving endangered species J Androl, 2012,33(2):264-276. [本文引用: 1]
Bedford-Guaus SJ, KimS, MuleroL, VaqueroJM, MoreraC, Adan-MilanèsR, VeigaA, Rayaá . Molecular markers of putative spermatogonial stem cells in the domestic cat Reprod Domest Anim, 2017,52(Suppl.2):177-186. [本文引用: 1]
TsaiKL, ClarkLA, MurphyKE . Understanding hereditary diseases using the dog and human as companion model systems Mamm Genome, 2007,18(6-7):444-451. [本文引用: 1]
KimY, TurnerD, NelsonJ, DobrinskiI, McenteeM, TravisAJ . Production of donor-derived sperm after spermatogonial stem cell transplantation in the dog Reproduction, 2008,136(6):823-831. [本文引用: 1]
AnandS, BhartiyaD, SriramanK, MallickA . Underlying mechanisms that restore spermatogenesis on transplanting healthy Niche cells in busulphan treated mouse testis Stem Cell Rev, 2016,12(6):682-697. [本文引用: 1]
BhartiyaD, AnandS . Effects of oncotherapy on testicular stem cells and niche Mol Hum Reprod, 2017,23(9):654-655. [本文引用: 1]
MulderCL, ZhengY, JanSZ, StruijkRB, ReppingS, HamerG, van PeltM . Spermatogonial stem cell autotransplantation and germline genomic editing: a future cure for spermatogenic failure and prevention of transmission of genomic diseases Hum Reprod Update, 2016,22(5):561-573. [本文引用: 1]
EasleyCA, PhillipsBT, McguireMM, BarringerJM, ValliH, HermannBP, SimerlyCR, RajkovicA, MikiT, OrwigKE, SchattenGP . Direct differentiation of human pluripotent stem cells into haploid spermatogenic cells Cell Rep, 2012,2(3):440-446. [本文引用: 1]
LorzadehN, KazemiradN . Embryonic stem cells and infertility Am J Perinatol, 2018,35(10):925-930. [本文引用: 2]
 ,华南农业大学 动物科学学院,国家生猪种业工程技术研究中心,广州 510642
,华南农业大学 动物科学学院,国家生猪种业工程技术研究中心,广州 510642
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}