 ,11.
,11. 2.
3.
4.
Pedigree and development of wheat varieties in Sichuan Province
Jianmin Zheng1, Jiangtao Luo1, Hongshen Wan1, Shizhao Li1, Manyu Yang1, Jun Li1, Ennian Yang1, Yun Jiang2, Yubin Liu3, Xiangquan Wang4, Zongjun Pu,11. 2.
3.
4.
通讯作者:
第一联系人:
编委: 夏先春
收稿日期:2019-03-8修回日期:2019-06-4网络出版日期:2019-07-20
| 基金资助: |
Received:2019-03-8Revised:2019-06-4Online:2019-07-20
| Fund supported: |
作者简介 About authors
郑建敏,硕士,助理研究员,研究方向:小麦遗传育种与资源利用E-mail:jianminxingzheng@yeah.net。
罗江陶,博士,助理研究员,研究方向:小麦遗传育种与细胞学研究E-mail:715644470@qq.com。

摘要
关键词:
Abstract
Keywords:
PDF (439KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
郑建敏, 罗江陶, 万洪深, 李式昭, 杨漫宇, 李俊, 杨恩年, 蒋云, 刘于斌, 王相权, 蒲宗君. 四川省小麦育成品种系谱分析及发展进程[J]. 遗传, 2019, 41(7): 599-610 doi:10.16288/j.yczz.19-081
Jianmin Zheng, Jiangtao Luo, Hongshen Wan, Shizhao Li, Manyu Yang, Jun Li, Ennian Yang, Yun Jiang, Yubin Liu, Xiangquan Wang, Zongjun Pu.
分析育成品种的系谱,可以明确品种的遗传基础,揭示亲缘关系及演变特点,总结育种经验,发现选育规律,指导育种实践,提高选择效率。1983年,金善宝院士主编的专著《中国小麦品种及其系谱》[1],对新中国成立至1980年间我国利用国内外种质选育小麦(Triticum aestivum Linn.)品种在生产上的应用情况和品种系谱组成、亲缘关系及部分目标性状的遗传传递脉络等进行了详述。2002年,庄巧生院士在延承前书脉络的基础上,主编《中国小麦品种改良及系谱分析》[2]一书,补充了我国早期小麦育种工作简况,重点从亲本取材、组合选配和遗传资源利用角度阐明我国小麦品种改良所走过的历程,概述了各大麦区生态环境和生产情况,介绍了品种更替和系谱渊源,并对我国小麦育种工作的主旋律即抗病育种、产量提高、加工品质改良进行了专题论述。前人从育种实践和科学实验中提炼出的经验与认识,对今后的小麦育种实践具有积极的指导意义。
四川省比较系统的小麦改良工作始于20世纪30年代。1998年,余遥研究员主编《四川小麦》[3]一书,系统总结了20世纪30年代以来四川省小麦生产发展的技术经验、育种、栽培和相关科研取得的成果与进展,提出今后的发展方向和路径。2010年,任正隆教授等在《雨养农业区的小麦育种》[4]一书中,总结了四川省小麦育种攻关30年的成果与经验。2018年,李生荣研究员等对绵阳市农业科学院小麦育种及相关研究工作进行了回顾总结,出版专著《绵阳号小麦品种改良的实践与发展》[5],对品种改良具有重要参考价值。
本文通过系谱梳理,对四川省小麦育种发展进程进行了分析,从不同的分析角度,总结成功的育种利用经验,探讨存在的共性问题,为今后小麦育种提供参考。
1 四川小麦育种和生产概况
自20世纪30年代至今,四川小麦育种已有80余年历史。全省平均产量从30年代的50 kg/亩(即750 kg/ha)左右上升到2016年的250 kg/亩(即3750 kg/ha)左右(参考2017年四川省统计年鉴,数据来自四川统计局网站http://tjj.sc.gov.cn/tjcbw/tjnj/);目前,成都平原大面积生产产量平均在350 kg/亩(即5250 kg/ha,数据来自广汉等地种植大户)左右,省区域试验高产水平在400 kg/亩(即6000 kg/ha,参考2010~2018年区试总结)左右,高产栽培示范最高纪录为710.7 kg/亩(即10 660.5 kg/ha,2010年,江油大堰乡泉水村,数据来自《农民日报》及网站http:// www.moa.gov.cn/ztzl/qnszcgdfs/201006/t20100602_1497968.htm)。80余年间,全省小麦总面积呈先上升后下降趋势,高峰时达到3480余万亩(即232万公顷,1979~1983年间)[3],目前稳定在1600万亩左右(即106.7万ha,参考2017年四川省统计年鉴)。育种主体目标从重点关注产量拓展到产量品质兼顾;由于地域气候生态特点,虽有强筋型品种,但以中筋、弱筋型为主[6]。规范的区域试验始于1980年冬季,最初由四川省农科院作物所负责安排实施,后由四川省种子站接管,1984年初有品种通过审(认)定。1982年3月,在四川省科学技术委员会领导下正式成立小麦育种攻关组,由四川省人民政府划拨专项科研经费支持,开展全省小麦育种联合攻关工作,极大推动了四川省小麦育种长期稳定地发展。从《四川小麦》[3]、《雨养农业区的小麦育种》[4]、《绵阳号小麦品种改良的实践与发展》[5]、《中国小麦品种改良及系谱分析》[2]、四川省种子站区试报告、四川省农作物品种审定公告及各个育种单位专家发表的文献资料[7,8,9,10,11,12]中,本文收集汇总了四川省1936~2017年总计326个品种(含5个地方品种、6个引进品种和315个育成品种),通过育种方式、亲本构成、高频亲本及骨干亲本变化、遗传贡献、易位系及人工合成种质分布情况等方面进行梳理,对四川省小麦育种发展进程进行了分析。
因育种家对系谱编写规则理解的不同,系谱书写格式存在差异。为研究分析方便,本文在假设所有育种家遵循相同规则下编写系谱,并默认:(1)系谱中F6及以上代次材料为稳定种质/亲本,如川育12资料所列系谱为:(983/高加索)F7//(980/3130)F8,则认为川育12为直接亲本1(983/高加索)和直接亲本2(980/3130)的单交;(2)选系或辐射育种材料系谱同原材料系谱,如绵阳21为绵阳11选系,则其系谱与绵阳11相同。统计育成品种直接亲本数量,分析育成品种直接亲本构成等。
2 育种方式
2.1 育种方式统计
根据初步统计结果,四川省育成品种的育种方式有杂交育种(基因重组)、诱变育种(基因突变)和杂交诱变结合3种方式。为更清晰地将育种方式呈现给读者,遂将杂交育种继续细分为:选系(因被选系均为通过杂交育种方式选育,遂将选系归入杂交育种)、单交、三交、双交、回交和其他复交;诱变育种主要是辐射诱变;杂交诱变结合均为单交+辐射诱变。如表1所示,315个育成品种中,通过杂交育种方式选育的有300个,占95.24%;通过诱变方式选育的有7个,通过杂交诱变结合方式选育的有8个。统计结果表明,杂交育种是四川省品种选育的主要方式,其中通过单交方式选育的品种最多,占66.98%。
Table 1
表1
表1 四川省品种育种方式统计表
Table 1
| 育种方式 | 品种数 | 比例(%) | ||
|---|---|---|---|---|
| 杂交育种 | 选系 | 17 | 5.40 | |
| 单交 | 211 | 66.98 | ||
| 复交 | 三交 | 43 | 13.65 | |
| 双交 | 8 | 2.54 | ||
| 其他 | 13 | 4.13 | ||
| 回交 | 8 | 2.54 | ||
| 诱变育种 | 辐射 | 7 | 2.22 | |
| 杂交诱变 | 单交+辐射 | 8 | 2.54 | |
新窗口打开|下载CSV
2.2 育种方式对品种的影响
作物育种是利用变异(自然突变或人为创造)现象,在各种选择压力下筛选符合目标要求的类型,满足生产和生活的需要。自然突变是产生物种变异的重要方式,是进化的主要动力,但发生频率较低,无法满足人类社会的发展需求。人工诱变(化学、物理、太空环境等)是产生变异的方式之一,因操作方法的繁杂、对人体存在有害影响及有利变异频率较低等情况,未能成为作物育种利用的主要方式。杂交是人为创造变异、产生新材料(品种)简单易行的方式。杂交育种是利用现有材料或品种,通过有限杂交实现基因重组产生新变异类型进行选择,是在已有材料和品种的基础上不断提升抗性、品质、产量等性状的改良过程。育种目标的确立是战略性考虑,育种方式是达到目标途径的战术性选择,目标的设定需要有高度和远见,途径的选择要具体可行且注重实效。杂交育种根据亲本数量和组配方式的不同,可分为单交、复交(两个以上亲本间杂交)、回交(存在轮回亲本)等。假设不同亲本间遗传物质存在较大差异,根据遗传学经典定律,则亲本数量越多,组配方式越复杂,后代变异的类型就越丰富,可能选育的品种类型就越多。对自然进化过程而言,更多的变异有利于存留更多生存适应型个体;人工选择偏向于符合生产生活需求的具体性状选择,易淘汰和丢失目标性状表现平平而综合适应性好的个体,且受选择复杂程度影响,变异类型的多样性与选育品种数量并不成正比。从四川育成小麦品种来看,杂交育种是小麦选育的主要方式;单交选育的品种数达211个,占66.98%,是杂交选育的主要方式,该结果与林作揖等[13]和方正等 [14]的结论一致。单交亲本数量少,后代选择目标性状明确,易选出优良品种;复交遗传基础复杂,性状间的组合多样,增加选择难度;针对优异品种的少数性状进行改良,选择有限回交能更快获得具备目标性状的材料。从理论上也不难推断,随着杂交亲本的增加,遗传组成势必变得复杂,变异类型多样化,材料稳定时间变长,增加优异基因丢失风险(尤其是在观察选育情况下),因此增加选择的复杂性。
在育种方式中,选择的路径多样化,不拘泥于某种形式,只要能选育出好品种便是好路径。在杂交育种方面,四川小麦生产历史上推广面积较大的品种中,山农205、阿勃、绵阳11、川麦107和川麦104来自单交组合,南大2419、川麦28和川麦42来自三交组合,繁6来自聚合杂交组合。随着育种技术和生物技术的发展,以及小麦基因组工作取得的巨大成绩,人们对小麦农艺性状基因表达及调控的理解越来越深入,可利用的连锁及功能标记逐渐丰富,传统育种路径正在逐渐转变,表型选择与分子标记筛选结合更为紧密,优异基因聚合及不良连锁基因断裂难度缩小,设计育种将由设想逐步趋向现实,未来育种方式将产生颠覆性的变化。此外,品种选育效率不仅受育种方式的影响,还受选择压力等多种因素的制约;有研究表明,不同选择压(环境)的交替选择,是选育优良品种的一个有效途径[14]。随着科技的发展,人们改造自然进而改变世界所受的束缚将逐渐减少,创造变异可以通过人工杂交、理化诱变、定点编辑等多种方式,选择压力可以人为创造、诱导发生,表型数据可以激光扫描、智能获取,性状表达可以人工智能、大数据模拟。至此,育种可以外包和程序化,通过相关操作说明,育种工作者可以精确地获得与育种目标一致的新品种。
3 亲本构成、贡献及变化规律
3.1 直接亲本统计
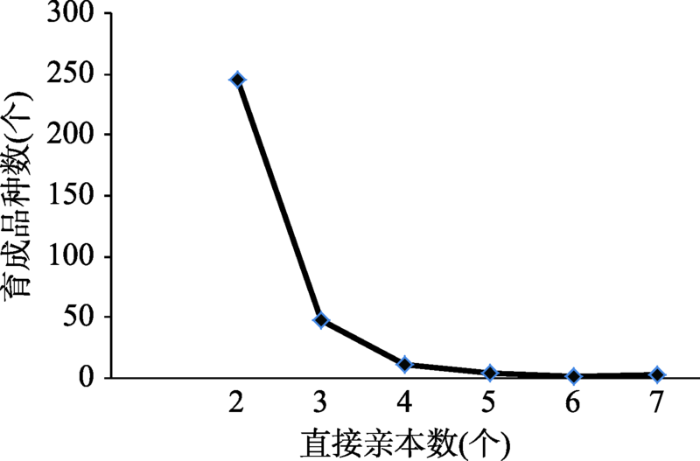
育成品种中,因西辐6号是以育种材料西辐3号通过辐射诱变方式选育而成,西辐3号无法获悉系谱信息,未纳入育成品种中分析直接亲本构成。统计表明,314个育成品种直接亲本有387个,系谱组合256个;其中由2个直接亲本培育的品种最多(累计达246个,占78.10%),随直接亲本数量的增加,育成品种数呈递减趋势(图1)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1育成品种直接亲本数统计
Fig. 1Statistical of direct parental number and its varieties
统计结果表明:在育成品种的256个组合中,选育1个品种的组合有217个;选育2个品种的组合有23个;选育3个品种的组合有12个;选育出3个以上品种的组合有3个,分别为:组合01-3570/ R138选育出4个品种,组合70-5858/繁6选育出5个品种,组合Autonomia/Fontarionco选育出6个品种。
3.2 亲本贡献
参照赵团结等[15]、盖钧镒等[16]方法计算直接亲本细胞核遗传贡献,同时追溯原始细胞质构成。每个育成品种细胞核遗传贡献值为1,细胞核贡献值按育成品种的直接亲本提供均等遗传贡献计算。例如由单交育成品种的双亲核遗传贡献各为0.5,3个亲本三交育成品种(假设系谱为:亲本1/亲本2//亲本3)的3个亲本遗传贡献分别为0.25、0.25和0.5;4个亲本双交育成品种(假设系谱为:亲本1/亲本2//亲本3/亲本4)的4个亲本遗传贡献值均为0.25;以此类推,其他复交方式育成品种依其杂交先后次序及组合,推算各个亲本遗传贡献值。遗传贡献率=贡献值/总贡献值×100%。育成品种细胞质与其杂交母本细胞质相同,将母本按其系谱,在所收集的327个品种系谱中追溯到无法再溯源为止,溯源母本胞质贡献为1,每个育成品种细胞质贡献为1。如表2所示,在387个直接亲本中,普通小麦亲本遗传贡献值累计达313.4375,遗传贡献率为99.82%,硬粒小麦、圆锥小麦等非普通小麦亲本遗传贡献值仅0.5625,遗传贡献率为0.18%;普通小麦亲本中选育材料遗传贡献(232.75,74.12%)>引进材料(71.4375,22.75%)>地方品种(4.9375,1.57%)>未知材料(4.3125,1.37%);选育材料中,中间材料贡献>选育品种,引进材料以国外引进材料为主。
Table 2
表2
表2 普通小麦和非普通小麦亲本来源及细胞核遗传贡献
Table 2
| 亲本来源 | 分类 | 遗传贡献值 | 遗传贡献率(%) | |
|---|---|---|---|---|
| 普通小麦 | 地方品种 | 4.9375 | 1.57 | |
| 选育 材料 | 选育品种 | 98.1875 | 31.49 | |
| 中间材料 | 134.5625 | 42.72 | ||
| 引进 材料 | 国外引进 | 51.625 | 16.39 | |
| 省外引进 | 19.8125 | 6.29 | ||
| 未知材料 | 4.3125 | 1.37 | ||
| 非普通小麦 | 硬粒小麦、圆锥小麦 | 0.5625 | 0.18 | |
新窗口打开|下载CSV
314个育成品种可追溯到124个细胞质材料,其中具国外引进材料胞质有150个品种,遗传贡献率为47.77%;具中间材料胞质的有117个,遗传贡献率为37.26%;具省外引进材料胞质的有24个,遗传贡献率为7.64%;具地方品种胞质的有10个,遗传贡献率为3.18%;具非普通小麦胞质的有6个,遗传贡献率为1.91%;其他未知来源胞质的有7个,遗传贡献率为2.23%。在124个细胞质中,育成品种数超过10个的均为国外引进材料,其中育成品种最多的细胞质材料为IBO1828,共育成59个品种,遗传贡献率为18.79%。
3.3 高使用频率亲本及骨干亲本变化
在分析直接亲本构成基础上,计算直接亲本育成品种数量,筛选出育成品种≥4个的亲本。从首个培育品种年份到截止目前最后一个培育品种年份排列,提炼出育成品种≥7个(高频亲本平均培育品种数)的骨干亲本进行分析。如表3统计显示,四川省育成品种数≥4的亲本有碧玉麦、中农28、成都光头等40个材料。结果表明,20世纪中期到20世纪末高使用频率亲本主要是国外引进材料和少数地方品种;21世纪开始,高使用频率亲本才逐渐转变成育成品种或中间材 料,这与我国的小麦育种发展过程相似,经历从无到有,从主要靠引进材料、地方品种到自主创制材料的过程。
Table 3
表3
表3 高使用频率亲本统计
Table 3
| 直接亲本名称 | 育成品种数 | 遗传贡献值 | 遗传贡献率(%) | 品种年份 |
|---|---|---|---|---|
| 碧玉麦 | 4 | 1.5000 | 0.48 | 1943~1966 |
| 中农28/中农28B | 6 | 1.6875 | 0.54 | 1943~1989 |
| 成都光头/成都光头分枝麦 | 6 | 1.4375 | 0.46 | 1947~1989 |
| 南大2419/南大2419选系* | 14 | 6.2500 | 1.98 | 1958~2006 |
| 51麦* | 8 | 2.8750 | 0.91 | 1961~1969 |
| 山农205 | 5 | 2.5000 | 0.79 | 1962~1967 |
| 玛拉* | 14 | 7.0000 | 2.22 | 1962~1989 |
| Autonomia | 6 | 3.0000 | 0.95 | 1965~1973 |
| Fontarionco | 6 | 3.0000 | 0.95 | 1965~1973 |
| IBO1828 | 5 | 1.3125 | 0.42 | 1965~1989 |
| NP824 | 5 | 1.5000 | 0.48 | 1965~1989 |
| 阿夫 | 5 | 1.7500 | 0.56 | 1966~1989 |
| 阿勃* | 7 | 3.5000 | 1.11 | 1969~2006 |
| 繁6* | 11 | 5.5000 | 1.75 | 1976~1989 |
| 高加索 | 4 | 1.6250 | 0.52 | 1978~1997 |
| 70-5858 | 5 | 2.5000 | 0.79 | 1984~1988 |
| 西昌反修麦 | 5 | 2.0625 | 0.65 | 1985~2004 |
| 77中2882* | 7 | 3.2500 | 1.03 | 1988~1996 |
| 绵阳11 * | 9 | 4.2500 | 1.35 | 1988~2000 |
| 绵阳19 | 5 | 2.5000 | 0.79 | 1989~1997 |
| 巴麦18 | 5 | 2.2500 | 0.71 | 1989~2001 |
| 1900 | 4 | 2.2500 | 0.71 | 1991~2001 |
| 绵阳15 | 4 | 2.0000 | 0.63 | 1995~1999 |
| 绵阳01821 | 5 | 2.0000 | 0.63 | 1995~2009 |
| 10927 (川育12) | 4 | 2.0000 | 0.63 | 2000~2017 |
| 绵阳26* | 9 | 4.7500 | 1.51 | 2002~2009 |
| SW1862 | 4 | 2.2500 | 0.71 | 2002~2013 |
| Syn-CD769 | 3 | 0.7500 | 0.24 | 2003~2005 |
| SW3243 (川麦30)* | 10 | 3.7500 | 1.19 | 2003~2017 |
| 贵农19/贵农19-4* | 7 | 3.2500 | 1.03 | 2004~2011 |
| 贵农21/贵农21-1* | 14 | 5.7500 | 1.83 | 2004~2017 |
| 生核3295 Shenghe3295 | 4 | 1.5000 | 0.48 | 2006~2014 |
| SW8688 (川麦36) | 5 | 2.2500 | 0.71 | 2006~2017 |
| 99-1572 (川麦42)* | 17 | 8.0000 | 2.55 | 2008~2017 |
| 1275-1 (绵麦37) | 5 | 2.0000 | 0.63 | 2010~2015 |
| 99-1522 (川麦43)* | 10 | 4.7500 | 1.51 | 2010~2017 |
| 攀早抗 Panzaokang | 4 | 2.0000 | 0.63 | 2013~2014 |
| 98-1231 (川农16) | 6 | 3.0000 | 0.95 | 2013~2017 |
| R138 (川农23) | 4 | 2.0000 | 0.63 | 2014~2016 |
| 01-3570 (川麦44)* | 9 | 4.2500 | 1.35 | 2014~2016 |
新窗口打开|下载CSV
在高使用频率亲本基础上,以育成品种数均值(6.75)为界线,筛选出14个骨干亲本,分别为:南大2419、51麦、玛拉、阿勃、繁6、77中2882、绵阳11、绵阳26、川麦30、贵农19、贵农21、川麦42、川麦43和川麦44;其中直接育成品种数超过10个的有南大2419 (14个)、玛拉(14个)、繁6 (11个)、贵农21 (14个)和川麦42 (17个)。
3.4 骨干亲本的存在与遗传多样性
目前作物育种多是跟踪式/递进式的,一般每一个时期选育的品种主要是利用前一时期选育的品种/材料做亲本,下一个时期品种又多是利用这一时期的品种/材料来培育;在此育种模式下,性状的综合改良推进到一定程度后往往会出现瓶颈问题,需要创新材料或创新技术来打破障碍,实现突破。在这样的育种过程中,农艺性状好、抗性强、品质优、产量高的材料常被偏向性选择利用,选育出品种的概率被人为增加,因此更有可能成为骨干亲本。本文在统计筛选高使用频率亲本基础上,提炼出14个骨干亲本,按其育成品种时间先后顺序分别为:南大2419、51麦、玛拉、阿勃、繁6、77中2882、绵阳11、绵阳26、川麦30、贵农19、贵农21、川麦42、川麦43和川麦44。不难发现,骨干亲本随育种的推进而逐渐变化,从主要为国外引进材料转变到育成品种。从我国小麦育种历史来看,早期也是利用国外引进种质及地方品种进行组配育种,其后才利用培育的品种/中间材料作为亲本进行杂交选育,与此同时不断引进新种质开展遗传育种改良工作[1,2,17]。育种发展经历了从无到有,从依赖引进材料、地方品种到自主创制材料的过程。
骨干亲本的存在是品种选育的结果,也是导致遗传多样性散失的主要原因之一。长期以来,小麦育种目的是满足人们生产和生活的需求,其育种目标是高产、优质、广适、抗病、抗逆等,这些一致性的需求和目标导致育种过程中选择性牵连效应[18]产生和少数骨干亲本的反复使用,进而不可避免地出现遗传基础单一化现象;并且,可以推断控制质量性状区段的单一化速度较数量性状区段快。作物育种导致品种遗传基础单一化、多样性散失现象普遍存在,如水稻(Oryza sativa L.)[18,19,20]、玉米(Zea mays L.)[21,22]、小麦[23,24]、马铃薯(Solanum tuberosum L.)[25]、谷子(Setaria italica)[26]、甘薯(Dioscorea esculenta)[27]、大麦(Hordeum vulgare L.)[28]、花生(Arachis hypogaea L.)[29]和甘蔗(Saccharum officinarum L.)[30]等作物中均有相关报道。遗传多样性散失是育种目标一致性的必然结果。假如作物肩负有改变空气质量、园艺观赏、建筑填充等多样化的需求任务,育种目标也将随之多样化,其遗传基础自然随目标多样化而变得丰富。
目前拓宽作物遗传基础、增加品种遗传多样性的主要途径有:(1)开展资源收集、保存、评价、管理和利用工作,如收集近缘种质开展远缘杂交等;(2)采用新的技术手段,如基因编辑等[31,32],创制更丰富的遗传变异。作物种质收集、保存、评价、管理和利用是防止资源丧失及物种灭绝的关键,也是避免因品种单一化、遗传基础狭窄带来负面影响的重要举措,具有长远的战略意义。遗传多样性散失是基因型频率单一化,基因本身并未消失或减少,采用新的技术方法编辑和改变基因序列可以创制新的变异类型,从而改变遗传结构;创新技术是社会科技发展的必然需求和生产发展的重要动力。
4 易位系/人工合成种质育种贡献
普通小麦近缘种,如黑麦(Secale cereale L.)、簇毛麦(Haynaldia villosa L.)、粗山羊草(Aegilops tauschii)等,携带的抗病、抗逆、优质及高产基因对改良小麦品种农艺性状具有巨大的潜在价值[33,34,35]。为确定近缘种质对四川小麦育种的贡献,本文从系谱来源上,统计易位系/人工合成种质衍生品种关系及数量。本文讨论的易位系主要指国内育种家创制并用于育种的易位系,直接来源于国外的易位系未做详细统计。人工合成种质则包含直接来源国外的种质和国内育种家创制的材料。4.1 1RS.1BL易位系
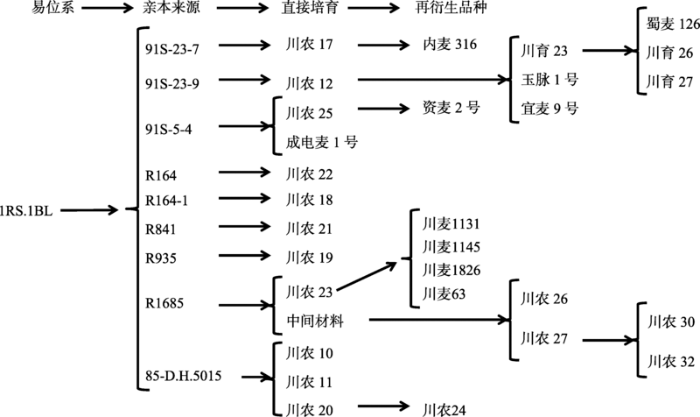
小麦-黑麦染色体易位所形成的1RS·1BL易位系是黑麦染色体存在于小麦染色体组的最常见形式。目前全世界用于小麦生产的新品种中,超过50%的品种含有1RS·1BL易位染色体[36,37,38]。源自德国的1RS·1BL易位材料被广泛应用于我国小麦育种中,该易位在我国北部麦区、黄淮麦区、长江中下游麦区及西南麦区均有较高分布率[37]。四川小麦1RS.1BL易位系由四川农业大学任正隆课题组创制并应用于育种[39,40,41],其1RS.1BL易位系有两种类型:其一源于苏联品种Aurora,含有抗小麦条锈病基因Yr9和抗白粉病基因Pm8,如85-D.H.5015;另一类型为该课题组创制的新1RS.1BL易位系,如91S-23-7、91S-23-7、91S-4-5等。如图2所示,截止目前,1RS.1BL易位系直接培育品种13个,间接培育品种17个,累计30个,占育成品种的9.52%。研究表明,新1RS·1BL易位系在改良抗病性,改善品质、提高叶片光合功能(功能性延绿)上具有重要作用[40,41]。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图21RS.1BL易位系及其培育品种
Fig. 21RS.1BL translocation lines and derived cultivars
4.2 6VS.6AL易位系
四川小麦中的小麦-簇毛麦6VS.6AL易位系引自南京农业大学,由刘大均等[42,43,44,45]利用普通小麦-簇毛麦代换系与扬麦5号杂交、辐射选育而成,该易位系对小麦白粉病反应型为免疫、近免疫。刘大钧 等[42,43,44,45]育成的92R系列小麦-簇毛麦易位系材料含抗白粉病基因Pm21和抗条锈病基因Yr26。如图3所示,利用该易位系直接培育品种8个,间接培育品种2个,累计10个,占育成品种的3.17%,该易位系已成为四川省育种和生产利用的重要白粉病抗源。图3
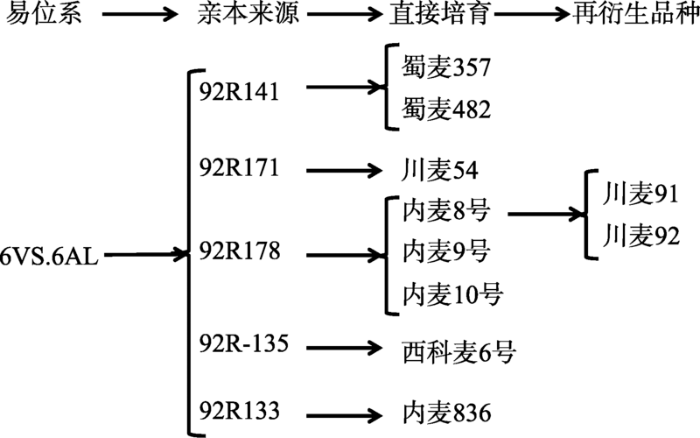 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图36VS.6AL易位系及其培育品种
Fig. 36VS.6AL translocation lines and derived cultivars
4.3 人工合成种质
利用人工合成种质是拓展小麦遗传基础、丰富遗传背景、突破育种瓶颈极为重要的材料。四川小麦人工合成种质主要有两种来源:(1)引自国际玉米小麦改良中心,如Syn-CD769、Syn-CD786等[46,47]。四川省农业科学院作物研究所杨武云研究员利用引进的人工合成种质培育出川麦38、川麦42、川麦43、川麦47等一批优异新品种,获得国家科技进步奖二等奖,成为国际上人工合成种质利用的典范。研究表明,川麦42不仅含有5+10优质基因、抗条锈基因,另外还发现了对产量提升具有重要作用的位点[48,49]。在这些利用人工合成小麦衍生品种川麦42育成的后代品种中,几乎所有的品种都继承了川麦42具有叶鞘绒毛这一特性,该野生特性来源于人工合成小麦D基因组,其在高育种选择压力下被保留下来,是因为该性状与高产性状相关QTL紧密连锁而受到了选择牵连[50],其大穗特性构成了目前四川小麦基本骨架,其在后代品种中得到了很好的传递[51]。川麦42、川麦43及其衍生品种将四川小麦产量水平推进到一个新的高度,对推动小麦育种与生产发展发挥了重大作用;(2)由四川农业大学小麦研究所刘登才团队利用圆锥小麦(T. turgidum, AABB)和粗山羊草(Ae. tauschii,DD)合成的新人工种质SHW-L1[52],其衍生品种蜀麦969和蜀麦830具有大穗、多小穗、高产等优良特征。目前,利用这两种人工合成种质直接培育品种6个,间接培育品种25个,累计31个,占总育成品种的9.84% (图4)。图4
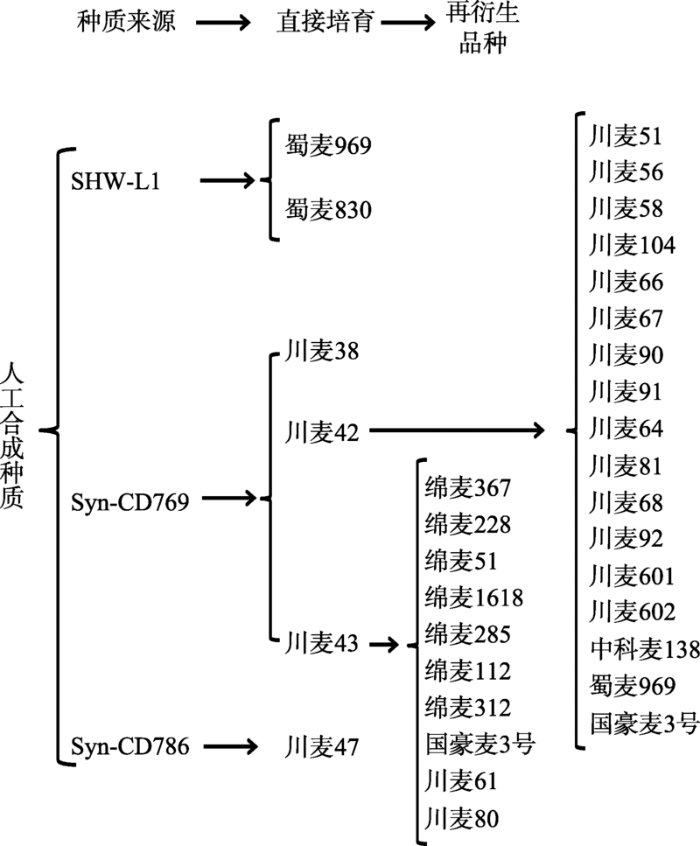 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4人工合成种质及其培育品种
Fig. 4Synthetic germplasm and derived cultivars
4 结语与展望
过去的80余年间,四川省育种家们主要采用杂交育种方式,培育了314个品种,这些品种来自于387个直接亲本和256个组合。四川省小麦育种经历从无到有,从主要依靠引进材料、地方品种到依靠自主创制材料的过程;骨干亲本随育种进程推进而逐渐变化。易位系和人工合成种质对四川省小麦育种贡献巨大。但是,育种目标的一致性及跟踪式的育种进程,必然导致遗传多样性散失和遗传基础脆弱性,今后重点应加强资源的保护和利用。随着生物科技发展和调控机理研究的深入,作物育种技术正在逐渐转变,聚集优质高产抗病抗逆的品种成为可能,但要实现四川省微生态环境多样条件下的大面积高产稳产(即6000 kg/ha),还需栽培、植保、土肥、农田水利等多学科协作;同时,根据市场需求,培育出品质类型多样、优质专用型品种,整体推进“川麦(面)”优化升级。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 2]
[本文引用: 3]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]

采用微卫星分子标记 (SSR)对四川省近 5 0年以来年推广面积达 6 6 70 0 hm2 (10 0万亩 )以上的 4 0个主栽小麦品种的遗传多样性进行了研究。结果发现 ,在小麦全基因组 4 2条染色体臂上的 4 6个 SSR位点上 30个 SSR位点 (6 5 .2 2 % )具有多态性。这 4 6个位点共检测到 110个等位变异 ,每个 SSR位点能检测到 1~ 8个 ,平均为 2 .4个。聚类分析表明 ,SSR标记能将 4 0个品种相互区分开。品种间遗传相似系数 (GS)变幅为 0 .4 5 1~ 0 .76 7,平均 GS值为0 .6 0 1。据此认为 ,SSR标记揭示出四川主栽小麦品种具有较高的遗传多样性。各年代间 GS值变化趋势分析表明 ,2 0世纪 70年代后 ,四川小麦的遗传多样性呈明显的下降趋势
Magsci [本文引用: 1]
采用微卫星分子标记 (SSR)对四川省近 5 0年以来年推广面积达 6 6 70 0 hm2 (10 0万亩 )以上的 4 0个主栽小麦品种的遗传多样性进行了研究。结果发现 ,在小麦全基因组 4 2条染色体臂上的 4 6个 SSR位点上 30个 SSR位点 (6 5 .2 2 % )具有多态性。这 4 6个位点共检测到 110个等位变异 ,每个 SSR位点能检测到 1~ 8个 ,平均为 2 .4个。聚类分析表明 ,SSR标记能将 4 0个品种相互区分开。品种间遗传相似系数 (GS)变幅为 0 .4 5 1~ 0 .76 7,平均 GS值为0 .6 0 1。据此认为 ,SSR标记揭示出四川主栽小麦品种具有较高的遗传多样性。各年代间 GS值变化趋势分析表明 ,2 0世纪 70年代后 ,四川小麦的遗传多样性呈明显的下降趋势
Magsci [本文引用: 1]
本课题组使用染色体工程方法选育了高产抗病的小麦新品系R164-1和A302-1。1993年用它们做亲本杂交。经过冬播和夏繁的交替选择,选出R25。该品种在四川省区试中表现优秀,2003年通过四川省品种审定委员会审定,并定名为川农18。该新品种分蘖力很强,成穗率特高,在小麦育种上突破了四川省的“生态穗容量”,具穗大兼穗数多的优良特性。这与现在所有的小麦品种类型不同,因而被称之为“协调型”新品种。2003年获国家品种权保护。被四川省政府推荐为重点推广品种。
Magsci [本文引用: 1]
本课题组使用染色体工程方法选育了高产抗病的小麦新品系R164-1和A302-1。1993年用它们做亲本杂交。经过冬播和夏繁的交替选择,选出R25。该品种在四川省区试中表现优秀,2003年通过四川省品种审定委员会审定,并定名为川农18。该新品种分蘖力很强,成穗率特高,在小麦育种上突破了四川省的“生态穗容量”,具穗大兼穗数多的优良特性。这与现在所有的小麦品种类型不同,因而被称之为“协调型”新品种。2003年获国家品种权保护。被四川省政府推荐为重点推广品种。
Magsci [本文引用: 1]
绵阳33号和绵阳35号是绵阳市农业科学研究所1992和1993年以自育中间材料为主要亲本杂交育成,具有绵阳系列品种特色的大穗型小麦新品种。主要持点是丰产性好,抗病力强,品质优良,适应性广泛。综合农艺性状优良,株高适中,穗层整产,穗大粒多,籽粒白皮大粒,饱满均匀,商品性突出,适应市场需求,具有良好的利用前景。两品种于2003年10月通过四川省品种审定,定名推广。
Magsci [本文引用: 1]
绵阳33号和绵阳35号是绵阳市农业科学研究所1992和1993年以自育中间材料为主要亲本杂交育成,具有绵阳系列品种特色的大穗型小麦新品种。主要持点是丰产性好,抗病力强,品质优良,适应性广泛。综合农艺性状优良,株高适中,穗层整产,穗大粒多,籽粒白皮大粒,饱满均匀,商品性突出,适应市场需求,具有良好的利用前景。两品种于2003年10月通过四川省品种审定,定名推广。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]
<div ><span >基于已获得的控制小麦条锈病成株抗性“一致性”</span><span >QTL</span><span >区段</span><span >80</span><span >个</span><span >SSR</span><span >标记,结合小麦骨干亲本繁</span><span >6</span><span >及其衍生的</span><span >39</span><span >个后代小麦品种进行田间条锈病成株期抗性表型鉴定,揭示了骨干亲本繁</span><span >6</span><span >遗传物质及其成株抗性在其衍生品种的遗传规律。结果表明,骨干亲本繁</span><span >6</span><span >在条锈病条中</span><span >31</span><span >、</span><span >32</span><span >和</span><span >33</span><span >混合生理小种诱导下表现成株抗性,</span><span >7</span><span >个衍生后代品种表现全生育期抗性;用控制小麦条锈病成株抗性</span><span >QTL</span><span >区段的</span><span >80</span><span >个</span><span >SSR</span><span >标记对繁</span><span >6</span><span >及其后代衍生品种的其他亲本进行分子扫描,共发现</span><span >9</span><span >个来自繁</span><span >6</span><span >基因组的特异</span><span >SSR</span><span >标记,即</span><em><span >Xwmc631</span></em><span >、</span><em><span >Xgwm359</span></em><span >、</span><em><span >Xwmc407</span></em><span >、</span><em><span >Xgwm501</span></em><span >、</span><em><span >Xgwm148</span></em><span >、</span><em><span >Xgwm539</span></em><span >、</span><em><span >Xgwm533</span></em><span >、</span><em><span >Xgwm299</span></em><span >和</span><em><span >Xgwm639</span></em><span >,其中,</span><em><span >Xwmc631</span></em><span >、</span><em><span >Xgwm359</span></em><span >、</span><em><span >Xgwm501</span></em><span >、</span><em><span >Xgwm299</span></em><span >和</span><em><span >Xgwm639</span></em><span >在繁</span><span >6</span><span >衍生后代的</span><span >4</span><span >个子代中表现较高的遗传贡献率。以</span><span >SSR</span><span >标记与小麦条锈病成株抗性的关联分析发现</span><span >6</span><span >个</span><span >SSR</span><span >标记与小麦条锈病成株抗性显著相关,其中来自繁</span><span >6</span><span >的特异</span><span >SSR</span><span >等位变异</span><em><span >Xgwm539-2D</span></em><span >和</span><em><span >Xgwm299-3B</span></em><span >与严重度、反应型、普遍率、病情指数及病程曲线下面积</span><span >(AUDPC)</span><span >均具显著相关性,表明繁</span><span >6</span><span >的成株抗性及其控制遗传位点在其衍生后代品种选育过程中得到了很好的定向选择,并在西南麦区小麦条锈病抗性育种中发挥了重要作用。</span></div>
Magsci [本文引用: 1]
<div ><span >基于已获得的控制小麦条锈病成株抗性“一致性”</span><span >QTL</span><span >区段</span><span >80</span><span >个</span><span >SSR</span><span >标记,结合小麦骨干亲本繁</span><span >6</span><span >及其衍生的</span><span >39</span><span >个后代小麦品种进行田间条锈病成株期抗性表型鉴定,揭示了骨干亲本繁</span><span >6</span><span >遗传物质及其成株抗性在其衍生品种的遗传规律。结果表明,骨干亲本繁</span><span >6</span><span >在条锈病条中</span><span >31</span><span >、</span><span >32</span><span >和</span><span >33</span><span >混合生理小种诱导下表现成株抗性,</span><span >7</span><span >个衍生后代品种表现全生育期抗性;用控制小麦条锈病成株抗性</span><span >QTL</span><span >区段的</span><span >80</span><span >个</span><span >SSR</span><span >标记对繁</span><span >6</span><span >及其后代衍生品种的其他亲本进行分子扫描,共发现</span><span >9</span><span >个来自繁</span><span >6</span><span >基因组的特异</span><span >SSR</span><span >标记,即</span><em><span >Xwmc631</span></em><span >、</span><em><span >Xgwm359</span></em><span >、</span><em><span >Xwmc407</span></em><span >、</span><em><span >Xgwm501</span></em><span >、</span><em><span >Xgwm148</span></em><span >、</span><em><span >Xgwm539</span></em><span >、</span><em><span >Xgwm533</span></em><span >、</span><em><span >Xgwm299</span></em><span >和</span><em><span >Xgwm639</span></em><span >,其中,</span><em><span >Xwmc631</span></em><span >、</span><em><span >Xgwm359</span></em><span >、</span><em><span >Xgwm501</span></em><span >、</span><em><span >Xgwm299</span></em><span >和</span><em><span >Xgwm639</span></em><span >在繁</span><span >6</span><span >衍生后代的</span><span >4</span><span >个子代中表现较高的遗传贡献率。以</span><span >SSR</span><span >标记与小麦条锈病成株抗性的关联分析发现</span><span >6</span><span >个</span><span >SSR</span><span >标记与小麦条锈病成株抗性显著相关,其中来自繁</span><span >6</span><span >的特异</span><span >SSR</span><span >等位变异</span><em><span >Xgwm539-2D</span></em><span >和</span><em><span >Xgwm299-3B</span></em><span >与严重度、反应型、普遍率、病情指数及病程曲线下面积</span><span >(AUDPC)</span><span >均具显著相关性,表明繁</span><span >6</span><span >的成株抗性及其控制遗传位点在其衍生后代品种选育过程中得到了很好的定向选择,并在西南麦区小麦条锈病抗性育种中发挥了重要作用。</span></div>
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]
在对1923~1995年育成的651个中国大豆品种系谱分析的基础上,计算每一育成品种祖先亲本的细胞核和细胞质遗传贡献值。在全国347个细胞核祖先亲本和214个细胞质祖先亲本中,来自东北、黄淮海、南方及国外的祖先亲本对全国育成品种细胞核与细胞质遗传贡献分别为48.23%和50.54%,25.50%和29.49%,13.74%和16.28%,11.95%和2.92%。进一步分析了东北、黄淮海、南方三大生态区大豆育成品种的核、质来源,包括国外引种所占的份额及其特点。评选出38个对全国和三大生态区域遗传贡献最大的种质。
Magsci [本文引用: 1]
在对1923~1995年育成的651个中国大豆品种系谱分析的基础上,计算每一育成品种祖先亲本的细胞核和细胞质遗传贡献值。在全国347个细胞核祖先亲本和214个细胞质祖先亲本中,来自东北、黄淮海、南方及国外的祖先亲本对全国育成品种细胞核与细胞质遗传贡献分别为48.23%和50.54%,25.50%和29.49%,13.74%和16.28%,11.95%和2.92%。进一步分析了东北、黄淮海、南方三大生态区大豆育成品种的核、质来源,包括国外引种所占的份额及其特点。评选出38个对全国和三大生态区域遗传贡献最大的种质。
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 2]
作物在长期进化过程中,除自然选择外,还经历了两次大的人工选择,即人工驯化选择和育种选择,使栽培种与野生种之间、现代品种与古老的地方品种之间在群体遗传结构及性状上形成了很大的差异;在基因组中,一些承受强选择作用的基因在群体中的多样性显著降低,同时这些基因附近区域的遗传多样性也明显下降。在遗传学中将这种对个别基因的选择导致其侧翼区域遗传多样性降低的现象称之为选择牵连效应(Hitchhiking effect,也称选择搭载效应)。通过大群体多位点的扫描分析,可找到一些发生选择牵连效应的基因组区段,利用标记/性状关联分析(Marker/Trait association analysis),就可发现这些区段所控制的重要性状;对这些区段进行精细扫描和分析,即可找到一些决定重要农艺性状的基因,并发现优异等位变异,从而为重要基因的克隆和作物品种的分子设计奠定基础。各大作物高密度分子标记连锁图谱的绘制完成、高通量基因型分析技术体系的建立,为利用选择牵连效应分析、通过标记/性状之间的关联,寻找和定位一些重要基因奠定了基础。本文在查阅文献的基础上,结合作者的研究,以小麦株高、千粒重、磷的吸收和利用等性状为例,就选择牵连效应分析的基本思路、方法进行了讨论,以起到抛砖引玉之作用。
Magsci [本文引用: 2]
作物在长期进化过程中,除自然选择外,还经历了两次大的人工选择,即人工驯化选择和育种选择,使栽培种与野生种之间、现代品种与古老的地方品种之间在群体遗传结构及性状上形成了很大的差异;在基因组中,一些承受强选择作用的基因在群体中的多样性显著降低,同时这些基因附近区域的遗传多样性也明显下降。在遗传学中将这种对个别基因的选择导致其侧翼区域遗传多样性降低的现象称之为选择牵连效应(Hitchhiking effect,也称选择搭载效应)。通过大群体多位点的扫描分析,可找到一些发生选择牵连效应的基因组区段,利用标记/性状关联分析(Marker/Trait association analysis),就可发现这些区段所控制的重要性状;对这些区段进行精细扫描和分析,即可找到一些决定重要农艺性状的基因,并发现优异等位变异,从而为重要基因的克隆和作物品种的分子设计奠定基础。各大作物高密度分子标记连锁图谱的绘制完成、高通量基因型分析技术体系的建立,为利用选择牵连效应分析、通过标记/性状之间的关联,寻找和定位一些重要基因奠定了基础。本文在查阅文献的基础上,结合作者的研究,以小麦株高、千粒重、磷的吸收和利用等性状为例,就选择牵连效应分析的基本思路、方法进行了讨论,以起到抛砖引玉之作用。
Magsci [本文引用: 1]
利用36个微卫星标记和42个表型性状对453份选育品种进行分析, 研究中国水稻选育品种的遗传多样性地理分布及其近50年的变化趋势. 结果表明, 微卫星标记和表型性状分析的遗传多样性具有较高的相似性; 籼稻品种的遗传多样性大于粳稻品种的遗传多样性; 从20世纪50年代到80年代, 选育品种的遗传多样性一直下降, 80年代降低到最低水平, 90年代又有显著提高; 在地理上华中稻区的选育品种遗传多样性最大, 东北稻区和西北稻区遗传多样性最小. 位于长江中下游的江苏、江西和西南地区的四川等地是中国水稻选育品种遗传多样性最大的地区. 东北地区作为重要的粳稻生产基地, 遗传基础非常狭窄, 应该发掘新的种质资源拓宽品种的遗传多样性.
Magsci [本文引用: 1]
利用36个微卫星标记和42个表型性状对453份选育品种进行分析, 研究中国水稻选育品种的遗传多样性地理分布及其近50年的变化趋势. 结果表明, 微卫星标记和表型性状分析的遗传多样性具有较高的相似性; 籼稻品种的遗传多样性大于粳稻品种的遗传多样性; 从20世纪50年代到80年代, 选育品种的遗传多样性一直下降, 80年代降低到最低水平, 90年代又有显著提高; 在地理上华中稻区的选育品种遗传多样性最大, 东北稻区和西北稻区遗传多样性最小. 位于长江中下游的江苏、江西和西南地区的四川等地是中国水稻选育品种遗传多样性最大的地区. 东北地区作为重要的粳稻生产基地, 遗传基础非常狭窄, 应该发掘新的种质资源拓宽品种的遗传多样性.
Magsci [本文引用: 1]
采用40个SSR标记,比较分析了151份20世纪50年代(78份)和近10年(73份)我国常规稻主栽品种的遗传差异,发现有39个标记具有多态性,多态性位点共检测到213个等位基因,每个位点2~11个,平均5.5个;平均Nei基因多样性指数(He)为0.649,范围在0.309(RM174)~0.869(RM418)。籼粳亚种间SSR多样性差异明显,籼稻平均等位基因数(Na)和Nei基因多样性指数(Na = 4.4,He = 0.458)均高于粳稻品种(Na = 4.0,He = 0.395)。比较了78份20世纪50年代与73份近10年水稻主栽品种的遗传多样性,籼、粳亚种表现出相近的变化趋势,即Nei多样性指数和等位基因数20世纪50年代主栽品种高于近10年的。虽然Nei基因多样性指数的变化并不显著(籼稻:z= 1.471,P=0.141;粳稻:z= 1.932,P=0.053),但等位基因数目的变化达到显著水平(籼稻:z= 2.677,P=0.007;粳稻:z= 3.441,P=0.001)。分子方差分析(AMOVA)表明,遗传变异绝大部分存在于两时期内,尽管时期间平均贡献的遗传变异仅占1.9%,但仍然达到5%的显著水平;籼、粳亚种两时期间平均贡献的遗传变异高于整个分析样本,分别为5.0%和8.2%;籼、粳亚种不同位点的遗传分化程度也各不相同,籼稻和粳稻品种分别有13个(占33.3%)和11个(占28.2%)SSR位点的等位基因在两时期间差异显著,而其余位点的遗传变异则是因时期内品种间的差异引起的。研究表明近10年我国常规稻主栽品种丢失了一部分等位基因,水稻育种仍应加强更广泛的种质亲本的选择。
Magsci [本文引用: 1]
采用40个SSR标记,比较分析了151份20世纪50年代(78份)和近10年(73份)我国常规稻主栽品种的遗传差异,发现有39个标记具有多态性,多态性位点共检测到213个等位基因,每个位点2~11个,平均5.5个;平均Nei基因多样性指数(He)为0.649,范围在0.309(RM174)~0.869(RM418)。籼粳亚种间SSR多样性差异明显,籼稻平均等位基因数(Na)和Nei基因多样性指数(Na = 4.4,He = 0.458)均高于粳稻品种(Na = 4.0,He = 0.395)。比较了78份20世纪50年代与73份近10年水稻主栽品种的遗传多样性,籼、粳亚种表现出相近的变化趋势,即Nei多样性指数和等位基因数20世纪50年代主栽品种高于近10年的。虽然Nei基因多样性指数的变化并不显著(籼稻:z= 1.471,P=0.141;粳稻:z= 1.932,P=0.053),但等位基因数目的变化达到显著水平(籼稻:z= 2.677,P=0.007;粳稻:z= 3.441,P=0.001)。分子方差分析(AMOVA)表明,遗传变异绝大部分存在于两时期内,尽管时期间平均贡献的遗传变异仅占1.9%,但仍然达到5%的显著水平;籼、粳亚种两时期间平均贡献的遗传变异高于整个分析样本,分别为5.0%和8.2%;籼、粳亚种不同位点的遗传分化程度也各不相同,籼稻和粳稻品种分别有13个(占33.3%)和11个(占28.2%)SSR位点的等位基因在两时期间差异显著,而其余位点的遗传变异则是因时期内品种间的差异引起的。研究表明近10年我国常规稻主栽品种丢失了一部分等位基因,水稻育种仍应加强更广泛的种质亲本的选择。
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]
<P><FONT face=Verdana>【目的】研究十年来中国主要玉米杂交种遗传基础的变化趋势及分析四平头、Reid和Lancaster三大种质类群等位基因的变异,为中国玉米育种提供一些理论依据。【方法】利用111个SSR标记对1991—2001年间大面积推广的66个杂交种的84份亲本自交系进行分析。【结果】①在66个杂交种中共检测到659个SSR等位基因,平均每个位点检测到5.95个等位基因。在过去的10年间,大面积推广的杂交种Nei遗传多样性指数由0.67增加到0.69,可检测到的等位基因的数量逐年增加,由603增加到653个等位基因,2001年大面积推广的杂交种与1991年的杂交种相比,缺失的等位基因仅有0.9%,而新增加了8.5%的等位基因;②对其中的四平头类群自交系16份,Reid类群15份自交系和Lancaster类群8份自交系进一步分析发现,在39个玉米自交系中共检测到583个等位基因,占检测到的SSR位点等位基因总数的88.3%。其中四平头类群检测到的等位基因位点最多,占检测到的等位基因总数的77.4%,Reid类群和Lancaster类群差别不大,分别为61.5%和60%。Reid与四平头及Lancaster与四平头的等位基因差异均在32%左右,而Reid与Lancaster间等位基因的差异仅为25%。而且Reid与Lancaster类群提供的特有等位基因比率分别为7.9%和7.3%,大部分的特有等位基因存在于四平头类群,约为24%。【结论】随着新的种质资源的引入和对原有资源的改造,与20世纪90年代初相比,中国的玉米资源遗传多样性有了一定的增加;地方种质含有大量的特有等位基因,这是其与国外种质组配优异杂交种的重要基础,应当加强地方种质的改良和利用。<BR></FONT></P>
Magsci [本文引用: 1]
<P><FONT face=Verdana>【目的】研究十年来中国主要玉米杂交种遗传基础的变化趋势及分析四平头、Reid和Lancaster三大种质类群等位基因的变异,为中国玉米育种提供一些理论依据。【方法】利用111个SSR标记对1991—2001年间大面积推广的66个杂交种的84份亲本自交系进行分析。【结果】①在66个杂交种中共检测到659个SSR等位基因,平均每个位点检测到5.95个等位基因。在过去的10年间,大面积推广的杂交种Nei遗传多样性指数由0.67增加到0.69,可检测到的等位基因的数量逐年增加,由603增加到653个等位基因,2001年大面积推广的杂交种与1991年的杂交种相比,缺失的等位基因仅有0.9%,而新增加了8.5%的等位基因;②对其中的四平头类群自交系16份,Reid类群15份自交系和Lancaster类群8份自交系进一步分析发现,在39个玉米自交系中共检测到583个等位基因,占检测到的SSR位点等位基因总数的88.3%。其中四平头类群检测到的等位基因位点最多,占检测到的等位基因总数的77.4%,Reid类群和Lancaster类群差别不大,分别为61.5%和60%。Reid与四平头及Lancaster与四平头的等位基因差异均在32%左右,而Reid与Lancaster间等位基因的差异仅为25%。而且Reid与Lancaster类群提供的特有等位基因比率分别为7.9%和7.3%,大部分的特有等位基因存在于四平头类群,约为24%。【结论】随着新的种质资源的引入和对原有资源的改造,与20世纪90年代初相比,中国的玉米资源遗传多样性有了一定的增加;地方种质含有大量的特有等位基因,这是其与国外种质组配优异杂交种的重要基础,应当加强地方种质的改良和利用。<BR></FONT></P>
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]
<p><span >为对马铃薯品种鉴别、优良杂交组合选配提供分子水平上的依据,利用</span><span lang="EN-US" >SSR</span><span >标记构建了中国</span><span lang="EN-US" >2000—2007</span><span >年审定的</span><span lang="EN-US" >88</span><span >个马铃薯品种的指纹图谱并进行了遗传多样性分析。以</span><span lang="EN-US" >138</span><span >对</span><span lang="EN-US" >SSR</span><span >引物对</span><span lang="EN-US" >16</span><span >份遗传差异较大的马铃薯材料的基因组</span><span lang="EN-US" >DNA</span><span >进行了扩增,筛选出</span><span lang="EN-US" >10</span><span >对多态性高、谱带清晰的引物。利用</span><span lang="EN-US" >10</span><span >对</span><span lang="EN-US" >SSR</span><span >引物对全部供试材料进行扩增及电泳检测,共检测到</span><span lang="EN-US" >135</span><span >个等位位点,其中</span><span lang="EN-US" >133</span><span >个为多态性位点,多态性比率达</span><span lang="EN-US" >98.52%</span><span >。每对</span><span lang="EN-US" >SSR</span><span >引物扩增出的等位位点数</span><span lang="EN-US" >7~22</span><span >个,平均</span><span lang="EN-US" >13.5</span><span >个,多态性信息量变化范围为</span><span lang="EN-US" >0.7604~0.9375</span><span >,平均</span><span lang="EN-US" >0.8501</span><span >。通过对电泳检测结果的统计分析,利用</span><span lang="EN-US" >S180</span><span >、</span><span lang="EN-US" >S25</span><span >、</span><span lang="EN-US" >S7</span><span >、</span><span lang="EN-US" >S151</span><span >、</span><span lang="EN-US" >S184</span><span >及</span><span lang="EN-US" >S192</span><span >等</span><span lang="EN-US" >6</span><span >对引物构建了</span><span lang="EN-US" >88</span><span >份供试材料的</span><span lang="EN-US" >SSR</span><span >指纹图谱。聚类分析表明,在相似系数</span><span lang="EN-US" >0.620</span><span >处,所有供试材料被被聚为一类,在相似系数</span><span lang="EN-US" >0.652</span><span >处,</span><span lang="EN-US" >81.8%</span><span >的材料仍然聚在一起,从分子水平上表明供试材料遗传基础非常狭窄。</span><span >聚类分析结果与供试材料系谱来源有较好一致性,同一栽培区域育成的品种在不同程度上聚在一类。</span></p>
Magsci [本文引用: 1]
<p><span >为对马铃薯品种鉴别、优良杂交组合选配提供分子水平上的依据,利用</span><span lang="EN-US" >SSR</span><span >标记构建了中国</span><span lang="EN-US" >2000—2007</span><span >年审定的</span><span lang="EN-US" >88</span><span >个马铃薯品种的指纹图谱并进行了遗传多样性分析。以</span><span lang="EN-US" >138</span><span >对</span><span lang="EN-US" >SSR</span><span >引物对</span><span lang="EN-US" >16</span><span >份遗传差异较大的马铃薯材料的基因组</span><span lang="EN-US" >DNA</span><span >进行了扩增,筛选出</span><span lang="EN-US" >10</span><span >对多态性高、谱带清晰的引物。利用</span><span lang="EN-US" >10</span><span >对</span><span lang="EN-US" >SSR</span><span >引物对全部供试材料进行扩增及电泳检测,共检测到</span><span lang="EN-US" >135</span><span >个等位位点,其中</span><span lang="EN-US" >133</span><span >个为多态性位点,多态性比率达</span><span lang="EN-US" >98.52%</span><span >。每对</span><span lang="EN-US" >SSR</span><span >引物扩增出的等位位点数</span><span lang="EN-US" >7~22</span><span >个,平均</span><span lang="EN-US" >13.5</span><span >个,多态性信息量变化范围为</span><span lang="EN-US" >0.7604~0.9375</span><span >,平均</span><span lang="EN-US" >0.8501</span><span >。通过对电泳检测结果的统计分析,利用</span><span lang="EN-US" >S180</span><span >、</span><span lang="EN-US" >S25</span><span >、</span><span lang="EN-US" >S7</span><span >、</span><span lang="EN-US" >S151</span><span >、</span><span lang="EN-US" >S184</span><span >及</span><span lang="EN-US" >S192</span><span >等</span><span lang="EN-US" >6</span><span >对引物构建了</span><span lang="EN-US" >88</span><span >份供试材料的</span><span lang="EN-US" >SSR</span><span >指纹图谱。聚类分析表明,在相似系数</span><span lang="EN-US" >0.620</span><span >处,所有供试材料被被聚为一类,在相似系数</span><span lang="EN-US" >0.652</span><span >处,</span><span lang="EN-US" >81.8%</span><span >的材料仍然聚在一起,从分子水平上表明供试材料遗传基础非常狭窄。</span><span >聚类分析结果与供试材料系谱来源有较好一致性,同一栽培区域育成的品种在不同程度上聚在一类。</span></p>
Magsci [本文引用: 1]
<p><span >通过遗传种质的多样性评估</span><span >可以指导深入研究资源和育种中优异互补亲本的选择</span><span >进而提高优异基因的交流累加和新品种培育的效率。本研究选用了来自世界各地的</span><span >份谷子核心种质通过</span><span >个表型性状的综合鉴定</span><span >评估遗传多样性和筛选优异种质资源</span><span >结果表明</span><span >我国谷子资源的表型遗传多样性丰富</span><span >单穗粒重、穗长、穗粗、株高、茎节数和生育期均表现了丰富的变异</span><span new="">谷子育成品种遗传多样性相比农家品种下降明显</span><span >育种的遗传增益主要体现在株高和穗长的适度减低,以及茎粗、茎节数、穗粗、单穗粒重、单穗重及生育期的适度增加</span><span new="">系统聚类分析将谷子资源分成</span><span >类</span><span >第</span><span >类以来源为东北欧国家的品种为主</span><span >第</span><span >类以北美和非洲的品种为主</span><span >第</span><span >类以东亚、南亚的品种为主</span><span new="">我国种质可划分为春播型、春夏兼播型和南方型</span><span >类型</span><span new="">采用主成分分析法和逐步回归分析法综合评判表明</span><span >叶鞘色、刚毛长度、粒色、米色、株高、穗长、茎粗和单穗粒重</span><span >个性状可作为谷子表型鉴定的主要指标。</span></p>
Magsci [本文引用: 1]
<p><span >通过遗传种质的多样性评估</span><span >可以指导深入研究资源和育种中优异互补亲本的选择</span><span >进而提高优异基因的交流累加和新品种培育的效率。本研究选用了来自世界各地的</span><span >份谷子核心种质通过</span><span >个表型性状的综合鉴定</span><span >评估遗传多样性和筛选优异种质资源</span><span >结果表明</span><span >我国谷子资源的表型遗传多样性丰富</span><span >单穗粒重、穗长、穗粗、株高、茎节数和生育期均表现了丰富的变异</span><span new="">谷子育成品种遗传多样性相比农家品种下降明显</span><span >育种的遗传增益主要体现在株高和穗长的适度减低,以及茎粗、茎节数、穗粗、单穗粒重、单穗重及生育期的适度增加</span><span new="">系统聚类分析将谷子资源分成</span><span >类</span><span >第</span><span >类以来源为东北欧国家的品种为主</span><span >第</span><span >类以北美和非洲的品种为主</span><span >第</span><span >类以东亚、南亚的品种为主</span><span new="">我国种质可划分为春播型、春夏兼播型和南方型</span><span >类型</span><span new="">采用主成分分析法和逐步回归分析法综合评判表明</span><span >叶鞘色、刚毛长度、粒色、米色、株高、穗长、茎粗和单穗粒重</span><span >个性状可作为谷子表型鉴定的主要指标。</span></p>
Magsci [本文引用: 1]
用ISSR标记分析了中国62份甘薯主要亲本的遗传多样性, 明确了其遗传差异。结果表明, 17个ISSR引物共检测出490条多态性谱带, 平均每条引物检测出28.8条多态性谱带, 说明ISSR标记是评价甘薯遗传多样性的有效途径之一。62份中国甘薯主要亲本遗传距离为0.158~0.924, 平均为0.574, 通过UPGMA法, 可以聚为2大类, 一类为国内自育亲本, 一类为外引亲本, 说明中国甘薯主要亲本遗传多样性较丰富, 其中自育亲本与外引亲本之间遗传距离较远; 亚洲亲本遗传多样性高于非洲和美洲亲本, 并且与其他亲本间遗传距离较远; 亚洲品种中, 中国大陆亲本遗传距离最小, 为0.419, 与来自中国台湾的亲本差异较小, 但与外引亚洲亲本遗传距离较远。因此, 中国在未来甘薯育种中, 可以国内自育亲本与外引亲本以及外引亚洲亲本与外引其他亲本配制组合, 拓宽中国甘薯品种的遗传背景。
Magsci [本文引用: 1]
用ISSR标记分析了中国62份甘薯主要亲本的遗传多样性, 明确了其遗传差异。结果表明, 17个ISSR引物共检测出490条多态性谱带, 平均每条引物检测出28.8条多态性谱带, 说明ISSR标记是评价甘薯遗传多样性的有效途径之一。62份中国甘薯主要亲本遗传距离为0.158~0.924, 平均为0.574, 通过UPGMA法, 可以聚为2大类, 一类为国内自育亲本, 一类为外引亲本, 说明中国甘薯主要亲本遗传多样性较丰富, 其中自育亲本与外引亲本之间遗传距离较远; 亚洲亲本遗传多样性高于非洲和美洲亲本, 并且与其他亲本间遗传距离较远; 亚洲品种中, 中国大陆亲本遗传距离最小, 为0.419, 与来自中国台湾的亲本差异较小, 但与外引亚洲亲本遗传距离较远。因此, 中国在未来甘薯育种中, 可以国内自育亲本与外引亲本以及外引亚洲亲本与外引其他亲本配制组合, 拓宽中国甘薯品种的遗传背景。
Magsci [本文引用: 1]
【目的】分析大麦亲本材料的遗传多样性,寻找与部分农艺性状相关联的分子标记,为大麦杂交组合的配置及分子标记辅助育种提供依据。【方法】利用86个SSR标记对113份大麦亲本材料进行多态性扫描,并进行遗传多样性分析。挑选57个标记进行群体遗传结构分析,在此基础上采用Tassel 2.1 GLM(general linear model)和MLM(mixed linear model)方法进行标记与农艺性状的关联分析。【结果】86个标记共检测出200个等位变异,变异范围为1—5个;基因频率的变异范围为0.0088—1.0000,Shannon指数变异范围为0.0000—1.2236;遗传相似系数(GS)变异范围在0.5504—0.9897,平均值为0.7477。通过群体遗传结构分析将供试材料分为4个亚群。以GLM分析,发现9个与株高、穗长、芒长、穗粒数和小穗着生密度相关联的标记,各标记对表型变异的解释率在0.0507—0.2766;以MLM分析,发现6个与株高、芒长和小穗着生密度相关的标记,各标记对表型变异的解释率在0.0238—0.1999。【结论】利用SSR标记分析了113份大麦亲本材料的遗传多样性及群体遗传结构,并通过2种关联分析模型,分别寻找到了9个与株高、穗长、芒长、穗粒数相关联,6个与株高、芒长和小穗着生密度相关联的标记,这些标记位于1H、2H、3H、4H和7H染色体。
Magsci [本文引用: 1]
【目的】分析大麦亲本材料的遗传多样性,寻找与部分农艺性状相关联的分子标记,为大麦杂交组合的配置及分子标记辅助育种提供依据。【方法】利用86个SSR标记对113份大麦亲本材料进行多态性扫描,并进行遗传多样性分析。挑选57个标记进行群体遗传结构分析,在此基础上采用Tassel 2.1 GLM(general linear model)和MLM(mixed linear model)方法进行标记与农艺性状的关联分析。【结果】86个标记共检测出200个等位变异,变异范围为1—5个;基因频率的变异范围为0.0088—1.0000,Shannon指数变异范围为0.0000—1.2236;遗传相似系数(GS)变异范围在0.5504—0.9897,平均值为0.7477。通过群体遗传结构分析将供试材料分为4个亚群。以GLM分析,发现9个与株高、穗长、芒长、穗粒数和小穗着生密度相关联的标记,各标记对表型变异的解释率在0.0507—0.2766;以MLM分析,发现6个与株高、芒长和小穗着生密度相关的标记,各标记对表型变异的解释率在0.0238—0.1999。【结论】利用SSR标记分析了113份大麦亲本材料的遗传多样性及群体遗传结构,并通过2种关联分析模型,分别寻找到了9个与株高、穗长、芒长、穗粒数相关联,6个与株高、芒长和小穗着生密度相关联的标记,这些标记位于1H、2H、3H、4H和7H染色体。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[J]
Magsci [本文引用: 1]
小麦近缘种是改良小麦的一个重要基因库, 具有许多栽培小麦所不具备的优良特性。我们通过远缘杂交、染色体工程的方法创制了一大批不同类型的材料, 经基因组原位杂交GISH、多色FISH 和特异分子标记鉴定, 抗条锈病、白粉病、叶锈病鉴定, 品质、营养性状以及产量性状鉴定, 共选育出10 类可为育种家利用的抗病、优质、富含微量营养元素、氮高效、丰产性状优良的远缘杂交新种质和新不育系种质; 开发了414对黑麦基因组专化的EST 引物, 31 个黑麦染色体(臂)专化的EST 分子标记, 可应用于分子标记辅助育种, 或追踪检测小麦背景中的黑麦染色体或染色体片段; 进行了抗病新基因的遗传分析和分子标记定位工作。利用新种质, 选育出了一批表现突出的抗病、营养高效的小麦-黑麦、小麦-冰草远缘杂交新品系
Magsci [本文引用: 1]
小麦近缘种是改良小麦的一个重要基因库, 具有许多栽培小麦所不具备的优良特性。我们通过远缘杂交、染色体工程的方法创制了一大批不同类型的材料, 经基因组原位杂交GISH、多色FISH 和特异分子标记鉴定, 抗条锈病、白粉病、叶锈病鉴定, 品质、营养性状以及产量性状鉴定, 共选育出10 类可为育种家利用的抗病、优质、富含微量营养元素、氮高效、丰产性状优良的远缘杂交新种质和新不育系种质; 开发了414对黑麦基因组专化的EST 引物, 31 个黑麦染色体(臂)专化的EST 分子标记, 可应用于分子标记辅助育种, 或追踪检测小麦背景中的黑麦染色体或染色体片段; 进行了抗病新基因的遗传分析和分子标记定位工作。利用新种质, 选育出了一批表现突出的抗病、营养高效的小麦-黑麦、小麦-冰草远缘杂交新品系
Magsci [本文引用: 1]
<p>发掘小麦近缘种低分子量麦谷蛋白基因, 可为小麦品质改良提供更多的基因资源。文章利用<em>Glu-B3</em>位点特异性标记LB1F/LB1R、LB2F/LB2R、LB3F/LB3R和 LB4F/LB4R, 对普通小麦B染色体组的7个可能供体近缘种, 即硬粒小麦(<em>T. durum</em>)、栽培二粒小麦(<em>T. dicoccum</em>)、野生二粒小麦(<em>T. dicoccoides</em>)、拟斯卑尔脱山羊草(<em>Ae. speltoides</em>)、高大山羊草(<em>Ae. longissima</em>)、西尔斯山羊草(<em>Ae. searsii</em>)和双角山羊草(<em>Ae. bicornis</em>)共20份材料进行PCR扩增, 克隆小麦近缘种中<em>GluB3-1、GluB3-2、GluB3-3</em>和<em>GluB3-4</em>基因的等位变异, 并对<em>Glu-B3</em>位点基因进行系统发育分析。共获得16个新等位变异, 其中<em>GluB3-1</em>基因的新等位变异1个, 命名为<em>GluB3-16</em>, 其推导氨基酸分子量为39.2 kDa; <em>GluB3-3</em>的新等位变异有3个, 分别命名为<em>GluB3-35、GluB3-36</em>和<em>GluB3-37</em>, 其推导氨基酸分子量为44.5 kDa(<em>GluB3-36</em>)或44.6 kDa(<em>GluB3-35</em>和<em>GluB3-37</em>); <em>GluB3-4</em>的新等位变异12个, 分别命名为<em>GluB3-46、GluB3-47、GluB3-48、GluB3-49、GluB3-410、GluB3-411、GluB3-412、GluB3-413、GluB3-414、GluB3-415、GluB3-416</em>和<em>GluB3-417</em>, 其推导氨基酸分子量变化在38.6(<em>GluB3-414</em>)~ 42.5 kDa(<em>GluB3-413</em>)之间; 16个新等位变异都包含单一的完整开放阅读框, 具有低分子量麦谷蛋白亚基的典型结构。文章进一步拓展了低分子量麦谷蛋白基因资源, 揭示不同<em>Glu-B3</em>基因的进化过程不完全相同, 为有效地利用小麦近缘种材料和转基因育种提供了新的基因资源。</p>
Magsci [本文引用: 1]
<p>发掘小麦近缘种低分子量麦谷蛋白基因, 可为小麦品质改良提供更多的基因资源。文章利用<em>Glu-B3</em>位点特异性标记LB1F/LB1R、LB2F/LB2R、LB3F/LB3R和 LB4F/LB4R, 对普通小麦B染色体组的7个可能供体近缘种, 即硬粒小麦(<em>T. durum</em>)、栽培二粒小麦(<em>T. dicoccum</em>)、野生二粒小麦(<em>T. dicoccoides</em>)、拟斯卑尔脱山羊草(<em>Ae. speltoides</em>)、高大山羊草(<em>Ae. longissima</em>)、西尔斯山羊草(<em>Ae. searsii</em>)和双角山羊草(<em>Ae. bicornis</em>)共20份材料进行PCR扩增, 克隆小麦近缘种中<em>GluB3-1、GluB3-2、GluB3-3</em>和<em>GluB3-4</em>基因的等位变异, 并对<em>Glu-B3</em>位点基因进行系统发育分析。共获得16个新等位变异, 其中<em>GluB3-1</em>基因的新等位变异1个, 命名为<em>GluB3-16</em>, 其推导氨基酸分子量为39.2 kDa; <em>GluB3-3</em>的新等位变异有3个, 分别命名为<em>GluB3-35、GluB3-36</em>和<em>GluB3-37</em>, 其推导氨基酸分子量为44.5 kDa(<em>GluB3-36</em>)或44.6 kDa(<em>GluB3-35</em>和<em>GluB3-37</em>); <em>GluB3-4</em>的新等位变异12个, 分别命名为<em>GluB3-46、GluB3-47、GluB3-48、GluB3-49、GluB3-410、GluB3-411、GluB3-412、GluB3-413、GluB3-414、GluB3-415、GluB3-416</em>和<em>GluB3-417</em>, 其推导氨基酸分子量变化在38.6(<em>GluB3-414</em>)~ 42.5 kDa(<em>GluB3-413</em>)之间; 16个新等位变异都包含单一的完整开放阅读框, 具有低分子量麦谷蛋白亚基的典型结构。文章进一步拓展了低分子量麦谷蛋白基因资源, 揭示不同<em>Glu-B3</em>基因的进化过程不完全相同, 为有效地利用小麦近缘种材料和转基因育种提供了新的基因资源。</p>
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
Magsci [本文引用: 2]
采用SDS-PAGE和SCAR标记对我国小麦主产区近30年来主要推广品种和新近育成的部分品系共179份进行了1BL/1RS鉴定,结果表明:我国20世纪80年代后育成的小麦品种中约38%为1BL/1RS品种,其中北方冬麦区和黄淮冬麦区频率较高,分别为59%和42%;长江中下游冬麦区和西南冬麦区频率较低,均为20%;东北春麦区未发现1BL/1RS品种。大多
Magsci [本文引用: 2]
采用SDS-PAGE和SCAR标记对我国小麦主产区近30年来主要推广品种和新近育成的部分品系共179份进行了1BL/1RS鉴定,结果表明:我国20世纪80年代后育成的小麦品种中约38%为1BL/1RS品种,其中北方冬麦区和黄淮冬麦区频率较高,分别为59%和42%;长江中下游冬麦区和西南冬麦区频率较低,均为20%;东北春麦区未发现1BL/1RS品种。大多
Magsci [本文引用: 1]
世界上广泛使用的1RS·1BL易位系遗传基础单一,已不能满足世界小麦育种的需要。本课题组培育了不同遗传基础的1RS·1BL易位系,并按照高产优质抗病的选育方向,用于“川农”系列小麦新品种的选育。为了深入了解1RS·1BL易位染色体的传递规律和育种价值,使用酸性聚丙烯酰胺凝胶电泳(A PAGE)、Giemsa C 带技术、DNA原位杂交技术对13个川农号小麦新品种及4个进入区域试验的新品系进行了检测,并对这17份供试材料的抗病性、农艺性状和主要品质进行了鉴定。结果证明川农10号、川农11、川农20含有来源于Aurora 的1RS·1BL易位染色体,川农12、川农17、川农18和R291是含新合成的1RS·1BL易位染色体的新品种(系)。分析结果表明,目前利用外源种质的小麦育种中,决定1RS·1BL易位能否存在于新品种中的第一选择压仍是抗病性,农艺和品质性状则是第二、三位的选择因素。本研究指出,持续利用1RS·1BL易位染色体培育高产优质抗病的小麦新品种的关键问题是发掘黑麦1RS本身的基因资源,在优先创建1RS·1BL易位系抗病性的遗传多样性的同时,创建农艺与品质性状的遗传多样性。
Magsci [本文引用: 1]
世界上广泛使用的1RS·1BL易位系遗传基础单一,已不能满足世界小麦育种的需要。本课题组培育了不同遗传基础的1RS·1BL易位系,并按照高产优质抗病的选育方向,用于“川农”系列小麦新品种的选育。为了深入了解1RS·1BL易位染色体的传递规律和育种价值,使用酸性聚丙烯酰胺凝胶电泳(A PAGE)、Giemsa C 带技术、DNA原位杂交技术对13个川农号小麦新品种及4个进入区域试验的新品系进行了检测,并对这17份供试材料的抗病性、农艺性状和主要品质进行了鉴定。结果证明川农10号、川农11、川农20含有来源于Aurora 的1RS·1BL易位染色体,川农12、川农17、川农18和R291是含新合成的1RS·1BL易位染色体的新品种(系)。分析结果表明,目前利用外源种质的小麦育种中,决定1RS·1BL易位能否存在于新品种中的第一选择压仍是抗病性,农艺和品质性状则是第二、三位的选择因素。本研究指出,持续利用1RS·1BL易位染色体培育高产优质抗病的小麦新品种的关键问题是发掘黑麦1RS本身的基因资源,在优先创建1RS·1BL易位系抗病性的遗传多样性的同时,创建农艺与品质性状的遗传多样性。
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
Magsci [本文引用: 2]
用抗白粉病的普通小麦-簇毛麦6VS/6AL易位系与普通小麦品种扬麦5号、普通小麦-簇毛麦6V代换系和中国春6A双端二体以及6V代换系和6A双端二体配制了4个测交组合,分析了这4个杂交组合F1|PMC’s MI C-分带的减数分裂构型。在(6VS/6AL易位系×扬麦5号)和(6VS/6AL易位系×6V代换系)的F1|PMC ’s MI, 分别观察到由易位染色体与6A染色体和6V染色体配对形成的具有特定C-分带带型的棒状二价体,杂种中的棒状二价体数目高于各自亲本中的棒状二价体数。在(易位系×C.S.d.d.t 6A)F1中,在87.9%的PMC中观察到由易位染色体长臂与6AL端体配对形成的异形二价体(tl’’)。而在6V代换系×C.S.d.d .t6a(F1)中,96.68%的PMC具有两个单价端体(t’,t’)。该结果进一步证实易位涉及簇毛麦染色体6VS和小麦染色体6AL,易位断点靠近着丝粒。在减数分裂中,易位染色体的正常配对和分离,保证了6VS上白粉病抗性基因Pm21的正常传递,为这一新抗源在小麦育种中的应用奠定了细胞学基础。
Magsci [本文引用: 2]
用抗白粉病的普通小麦-簇毛麦6VS/6AL易位系与普通小麦品种扬麦5号、普通小麦-簇毛麦6V代换系和中国春6A双端二体以及6V代换系和6A双端二体配制了4个测交组合,分析了这4个杂交组合F1|PMC’s MI C-分带的减数分裂构型。在(6VS/6AL易位系×扬麦5号)和(6VS/6AL易位系×6V代换系)的F1|PMC ’s MI, 分别观察到由易位染色体与6A染色体和6V染色体配对形成的具有特定C-分带带型的棒状二价体,杂种中的棒状二价体数目高于各自亲本中的棒状二价体数。在(易位系×C.S.d.d.t 6A)F1中,在87.9%的PMC中观察到由易位染色体长臂与6AL端体配对形成的异形二价体(tl’’)。而在6V代换系×C.S.d.d .t6a(F1)中,96.68%的PMC具有两个单价端体(t’,t’)。该结果进一步证实易位涉及簇毛麦染色体6VS和小麦染色体6AL,易位断点靠近着丝粒。在减数分裂中,易位染色体的正常配对和分离,保证了6VS上白粉病抗性基因Pm21的正常传递,为这一新抗源在小麦育种中的应用奠定了细胞学基础。
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
URL [本文引用: 1]
Magsci [本文引用: 1]
<p><span >川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂来源于人工合成小麦亲本</span><span >Syn769</span><span >。利用川麦</span><span >42</span><span >与含</span><span >1BL/1RS</span><span >易位系的四川小麦品种川农</span><span >16</span><span >构建的</span><span >127</span><span >个重组自交系</span><span >(RIL, F<sub>8</sub>)</span><span >,经</span><span >3</span><span >年</span><span >4</span><span >个环境的遗传评价,比较了川麦</span><span >42</span><span >的</span><span >1BS</span><span >和川农</span><span >16</span><span >的</span><span >1RS</span><span >染色体臂对小麦产量构成因子和产量的遗传效应。结果表明,</span><span >RIL</span><span >群体中川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂株系和川农</span><span >16</span><span >的</span><span >1RS</span><span >染色体臂株系在分蘖力、成穗率、全生育期、小穗数、收获指数和籽粒产量</span><span >6</span><span >个性状上存在显著差异</span><span >; 1BS</span><span >染色体臂有利于提高成穗率和收获指数,而</span><span >1RS</span><span >染色体臂有利于提高分蘖能力和增加小穗数,</span><span >1BS</span><span >株系的籽粒平均产量比</span><span >1RS</span><span >株系增加</span><span >2.91%</span><span >。鉴于</span><span >1RS</span><span >染色体臂上的抗条锈病基因</span><span >丧失抗性,其携带的黑麦碱基因对加工品质有明显的负向作用,而川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂携带高抗条锈病基因</span><em><span >YrCH42</span></em><span >, </span><span >并对小麦籽粒产量有正向作用,因此建议在小麦遗传改良中利用川麦</span><span >42</span><span >的</span><span >1BS</span><span >替换</span><span >1RS</span><span >染色体臂。</span></p>
Magsci [本文引用: 1]
<p><span >川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂来源于人工合成小麦亲本</span><span >Syn769</span><span >。利用川麦</span><span >42</span><span >与含</span><span >1BL/1RS</span><span >易位系的四川小麦品种川农</span><span >16</span><span >构建的</span><span >127</span><span >个重组自交系</span><span >(RIL, F<sub>8</sub>)</span><span >,经</span><span >3</span><span >年</span><span >4</span><span >个环境的遗传评价,比较了川麦</span><span >42</span><span >的</span><span >1BS</span><span >和川农</span><span >16</span><span >的</span><span >1RS</span><span >染色体臂对小麦产量构成因子和产量的遗传效应。结果表明,</span><span >RIL</span><span >群体中川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂株系和川农</span><span >16</span><span >的</span><span >1RS</span><span >染色体臂株系在分蘖力、成穗率、全生育期、小穗数、收获指数和籽粒产量</span><span >6</span><span >个性状上存在显著差异</span><span >; 1BS</span><span >染色体臂有利于提高成穗率和收获指数,而</span><span >1RS</span><span >染色体臂有利于提高分蘖能力和增加小穗数,</span><span >1BS</span><span >株系的籽粒平均产量比</span><span >1RS</span><span >株系增加</span><span >2.91%</span><span >。鉴于</span><span >1RS</span><span >染色体臂上的抗条锈病基因</span><span >丧失抗性,其携带的黑麦碱基因对加工品质有明显的负向作用,而川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂携带高抗条锈病基因</span><em><span >YrCH42</span></em><span >, </span><span >并对小麦籽粒产量有正向作用,因此建议在小麦遗传改良中利用川麦</span><span >42</span><span >的</span><span >1BS</span><span >替换</span><span >1RS</span><span >染色体臂。</span></p>
Magsci [本文引用: 1]
人工合成小麦是改良现代小麦的重要基因资源。川麦42是人工合成小麦与普通小麦杂交育成的高产、抗条锈、广适小麦新品种。利用小麦全基因组的1029个SSR标记扫描,检测了川麦42遗传背景中人工合成小麦导入位点,并利用川麦42与四川小麦品种川农16构建的127个重组自交系(RIL, F<sub>8</sub>),在4年6个环境下种植获得的农艺性状数据,分析了人工合成小麦导入位点对小麦产量和产量构成因子的遗传效应,在川麦42遗传背景中发现一个高产的人工合成小麦导入位点Barc1183。根据Barc1183分子标记,将RIL群体中的127个株系分为川麦42基因型(具人工合成小麦导入位点)和川农16基因型(具川农16位点)两组,前者的人工合成小麦导入位点能促进分蘖能力,提高有效穗数、每平方米粒数,增加收获指数、籽粒生产率,在4年6个环境下较后者平均增产达8.92%,Barc1183为一高产的人工合成小麦导入位点。利用中国春双端体和硬粒小麦Longdon的D染色体代换系验证,将其定位于小麦4D染色体长臂。川麦42遗传背景中的高产人工合成小麦导入位点Barc1183,对于进一步开展小麦高产育种研究具有重要价值。
Magsci [本文引用: 1]
人工合成小麦是改良现代小麦的重要基因资源。川麦42是人工合成小麦与普通小麦杂交育成的高产、抗条锈、广适小麦新品种。利用小麦全基因组的1029个SSR标记扫描,检测了川麦42遗传背景中人工合成小麦导入位点,并利用川麦42与四川小麦品种川农16构建的127个重组自交系(RIL, F<sub>8</sub>),在4年6个环境下种植获得的农艺性状数据,分析了人工合成小麦导入位点对小麦产量和产量构成因子的遗传效应,在川麦42遗传背景中发现一个高产的人工合成小麦导入位点Barc1183。根据Barc1183分子标记,将RIL群体中的127个株系分为川麦42基因型(具人工合成小麦导入位点)和川农16基因型(具川农16位点)两组,前者的人工合成小麦导入位点能促进分蘖能力,提高有效穗数、每平方米粒数,增加收获指数、籽粒生产率,在4年6个环境下较后者平均增产达8.92%,Barc1183为一高产的人工合成小麦导入位点。利用中国春双端体和硬粒小麦Longdon的D染色体代换系验证,将其定位于小麦4D染色体长臂。川麦42遗传背景中的高产人工合成小麦导入位点Barc1183,对于进一步开展小麦高产育种研究具有重要价值。
Magsci [本文引用: 1]
<p><span >川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂来源于人工合成小麦亲本</span><span >Syn769</span><span >。利用川麦</span><span >42</span><span >与含</span><span >1BL/1RS</span><span >易位系的四川小麦品种川农</span><span >16</span><span >构建的</span><span >127</span><span >个重组自交系</span><span >(RIL, F<sub>8</sub>)</span><span >,经</span><span >3</span><span >年</span><span >4</span><span >个环境的遗传评价,比较了川麦</span><span >42</span><span >的</span><span >1BS</span><span >和川农</span><span >16</span><span >的</span><span >1RS</span><span >染色体臂对小麦产量构成因子和产量的遗传效应。结果表明,</span><span >RIL</span><span >群体中川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂株系和川农</span><span >16</span><span >的</span><span >1RS</span><span >染色体臂株系在分蘖力、成穗率、全生育期、小穗数、收获指数和籽粒产量</span><span >6</span><span >个性状上存在显著差异</span><span >; 1BS</span><span >染色体臂有利于提高成穗率和收获指数,而</span><span >1RS</span><span >染色体臂有利于提高分蘖能力和增加小穗数,</span><span >1BS</span><span >株系的籽粒平均产量比</span><span >1RS</span><span >株系增加</span><span >2.91%</span><span >。鉴于</span><span >1RS</span><span >染色体臂上的抗条锈病基因</span><span >丧失抗性,其携带的黑麦碱基因对加工品质有明显的负向作用,而川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂携带高抗条锈病基因</span><em><span >YrCH42</span></em><span >, </span><span >并对小麦籽粒产量有正向作用,因此建议在小麦遗传改良中利用川麦</span><span >42</span><span >的</span><span >1BS</span><span >替换</span><span >1RS</span><span >染色体臂。</span></p>
Magsci [本文引用: 1]
<p><span >川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂来源于人工合成小麦亲本</span><span >Syn769</span><span >。利用川麦</span><span >42</span><span >与含</span><span >1BL/1RS</span><span >易位系的四川小麦品种川农</span><span >16</span><span >构建的</span><span >127</span><span >个重组自交系</span><span >(RIL, F<sub>8</sub>)</span><span >,经</span><span >3</span><span >年</span><span >4</span><span >个环境的遗传评价,比较了川麦</span><span >42</span><span >的</span><span >1BS</span><span >和川农</span><span >16</span><span >的</span><span >1RS</span><span >染色体臂对小麦产量构成因子和产量的遗传效应。结果表明,</span><span >RIL</span><span >群体中川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂株系和川农</span><span >16</span><span >的</span><span >1RS</span><span >染色体臂株系在分蘖力、成穗率、全生育期、小穗数、收获指数和籽粒产量</span><span >6</span><span >个性状上存在显著差异</span><span >; 1BS</span><span >染色体臂有利于提高成穗率和收获指数,而</span><span >1RS</span><span >染色体臂有利于提高分蘖能力和增加小穗数,</span><span >1BS</span><span >株系的籽粒平均产量比</span><span >1RS</span><span >株系增加</span><span >2.91%</span><span >。鉴于</span><span >1RS</span><span >染色体臂上的抗条锈病基因</span><span >丧失抗性,其携带的黑麦碱基因对加工品质有明显的负向作用,而川麦</span><span >42</span><span >的</span><span >1BS</span><span >染色体臂携带高抗条锈病基因</span><em><span >YrCH42</span></em><span >, </span><span >并对小麦籽粒产量有正向作用,因此建议在小麦遗传改良中利用川麦</span><span >42</span><span >的</span><span >1BS</span><span >替换</span><span >1RS</span><span >染色体臂。</span></p>
URL [本文引用: 1]
Magsci [本文引用: 1]
【目的】解析突破性高产小麦新品种川麦104的遗传构成,探讨双亲川麦42和川农16对其高产特性的贡献。【方法】利用已构建的遗传连锁图谱上的176个SSR和683个DArT标记对川麦104及其亲本进行分析,了解川麦104的遗传构成;根据已定位到的产量性状QTL,分析来源于双亲的染色体区段对川麦104产量相关性状的贡献。【结果】在川麦104的双亲具有差异的859个多态位点中(22个位点缺失),有522个位点上的等位基因来源于川麦42,315个位点上的等位基因来源于川农16;川麦104更多地继承了川麦42的遗传成分(60.8%);川麦104中来源于双亲的遗传位点在A、B和D基因组分布不同,来源于川麦42的等位位点在A、B和D基因组所占比例分别为55.00%、60.20%和67.27%;川麦104中来源于双亲的等位位点在21条染色体上的分布也不同,来源于川麦42的等位位点主要分布于3A、5A、7A、1B、5B、7B、3D、4D、5D和7D染色体上,来源于川农16的等位位点主要分布于4A、3B、4B、6B、1D、2D和6D染色体上。川麦104来源于双亲的染色体区段(遗传距离大于5 cM)共68个,总长度为3 089.6 cM;来源于川麦42和川农16的染色体区段分别为36和32个,来源于川麦42的染色体区段主要分布在3D、5D、7A、7B和7D染色体上,来源于川农16的染色体区段主要分布在3B、4B和6D染色体上;在A和D基因组川麦104来源于川麦42的染色体区段比川农16的多,B基因组中来源于川农16的染色体区段比川麦42的多。在1B、1D、2B、4A、4D、5A、5B、5D和7A染色体上,9个来源于川麦42的染色体区段以及5个来源于川农16的染色体区段富集了与产量性状相关的QTL,其中,在1BS和4A染色体上来源于川麦42的染色体区段携带增加穗粒数的QTL等位位点;在1D、2B和4A染色体上来源于川农16的染色体区段携带增加单位面积穗数的QTL等位位点;5B染色体上来源于川麦42的染色体区段和4A、4D染色体上来源于川农16的染色体区段均携带增加千粒重的QTL等位位点,这些QTL的聚合对川麦104的产量三因素有增效作用。【结论】小麦新品种川麦104的高穗粒数特性来源于川麦42,多穗数特性来源于川农16,其千粒重特性双亲均有贡献,表明双亲的正效产量性状QTL重组是川麦104的高产遗传基础。
Magsci [本文引用: 1]
【目的】解析突破性高产小麦新品种川麦104的遗传构成,探讨双亲川麦42和川农16对其高产特性的贡献。【方法】利用已构建的遗传连锁图谱上的176个SSR和683个DArT标记对川麦104及其亲本进行分析,了解川麦104的遗传构成;根据已定位到的产量性状QTL,分析来源于双亲的染色体区段对川麦104产量相关性状的贡献。【结果】在川麦104的双亲具有差异的859个多态位点中(22个位点缺失),有522个位点上的等位基因来源于川麦42,315个位点上的等位基因来源于川农16;川麦104更多地继承了川麦42的遗传成分(60.8%);川麦104中来源于双亲的遗传位点在A、B和D基因组分布不同,来源于川麦42的等位位点在A、B和D基因组所占比例分别为55.00%、60.20%和67.27%;川麦104中来源于双亲的等位位点在21条染色体上的分布也不同,来源于川麦42的等位位点主要分布于3A、5A、7A、1B、5B、7B、3D、4D、5D和7D染色体上,来源于川农16的等位位点主要分布于4A、3B、4B、6B、1D、2D和6D染色体上。川麦104来源于双亲的染色体区段(遗传距离大于5 cM)共68个,总长度为3 089.6 cM;来源于川麦42和川农16的染色体区段分别为36和32个,来源于川麦42的染色体区段主要分布在3D、5D、7A、7B和7D染色体上,来源于川农16的染色体区段主要分布在3B、4B和6D染色体上;在A和D基因组川麦104来源于川麦42的染色体区段比川农16的多,B基因组中来源于川农16的染色体区段比川麦42的多。在1B、1D、2B、4A、4D、5A、5B、5D和7A染色体上,9个来源于川麦42的染色体区段以及5个来源于川农16的染色体区段富集了与产量性状相关的QTL,其中,在1BS和4A染色体上来源于川麦42的染色体区段携带增加穗粒数的QTL等位位点;在1D、2B和4A染色体上来源于川农16的染色体区段携带增加单位面积穗数的QTL等位位点;5B染色体上来源于川麦42的染色体区段和4A、4D染色体上来源于川农16的染色体区段均携带增加千粒重的QTL等位位点,这些QTL的聚合对川麦104的产量三因素有增效作用。【结论】小麦新品种川麦104的高穗粒数特性来源于川麦42,多穗数特性来源于川农16,其千粒重特性双亲均有贡献,表明双亲的正效产量性状QTL重组是川麦104的高产遗传基础。
URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}