The Measurement Principles, Methods and Applications of P515
Chunyan Zhang,1,2,*, Xiaojie Pang21Plant Science Facility of Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2Key Laboratory of Photobiology, Chinese Academy of Sciences, Beijing 100093, China
Abstract The spectral techniques have been widely used in the field of photosynthesis research, such as the light absorption signals P515 and P700 redox kinetics, and chlorophyll fluorescence, which can detect the photosynthetic activities of plants quickly and accurately. P515 signal is widely present in higher plants and algae, which is caused by the shift of absorption spectrum of pigments on thylakoid membrane. We can detect the ratio of PSI to PSII reaction center, the proton conductivity of chloroplast ATP synthase, the cyclic electron flow rate around PSI, the proton motive force (pmf) and its components by the P515 fast and slow kinetics, and study the photoprotective mechanism by simultaneous detection of P515 signal and chlorophyll fluorescence. In this paper, we summarize the main measurement methods of P515, expound its principles, and the applications. The aim is to provide technical supports for further study on the mechanism of photosynthesis. Keywords:P515;ATP synthase;cyclic electron transport;proton motive force
PDF (964KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张春艳, 庞肖杰. P515的测量原理、方法和应用. 植物学报, 2021, 56(5): 594-604 doi:10.11983/CBB21052 Zhang Chunyan, Pang Xiaojie. The Measurement Principles, Methods and Applications of P515. Chinese Bulletin of Botany, 2021, 56(5): 594-604 doi:10.11983/CBB21052
在光合作用中, 放氧光合生物吸收太阳能并裂解水分子, 在类囊体囊腔释放大量的H+并驱动电子在膜蛋白复合物和电子载体间传递, 最终生成还原型烟酰胺腺嘌呤二核苷酸磷酸(辅酶II) (NADPH)。同时, 光合电子的传递导致质子从叶绿体基质向类囊体囊腔传递并伴随多种离子的跨膜转运, 随后卡尔文循环消耗质子进行固碳, 又导致叶绿体基质碱化, 这些因素共同诱导形成跨膜电势(ΔΨ)和质子梯度(ΔpH), 即质子动力势(proton motive force, pmf) (Cruz et al., 2001; Carraretto et al., 2016)。质子动力势驱动ATP合酶产生腺苷三磷酸(ATP), ATP和NADPH为光合作用中卡尔文循环及其它代谢途径提供能量和还原力(Kramer et al., 2003; Takizawa et al., 2007)。
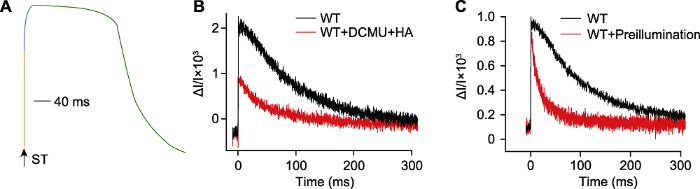
(A) 饱和闪光诱导的P515快速动力学示意图, 包括快速上升阶段(橙)、缓慢上升阶段(蓝)和缓慢下降阶段(绿), 箭头表示1次单周转饱和闪光(ST); (B) 饱和闪光诱导莱茵衣藻在520-550 nm的快速吸收变化(单周转饱和闪光波长635 nm, 光强200000 μmol·m-2·s-1, 持续时间5 μs), 莱茵衣藻野生型(WT) (浓度约25 μg·mL-1)加入(红)和不加入(黑) 10 µmol·L-1二氯苯基二甲脲(DCMU)和1 mmol·L-1羟胺(HA), 测量前莱茵衣藻暗适应20分钟; (C) 预照光对P515快速动力学的影响, 莱茵衣藻野生型在暗适应20分钟(黑)以及在光下持续照光2分钟(光强40 μmol·m-2·s-1), 黑暗1分钟(红)后的P515快速动力学。 Figure 1The P515 fast kinetics induced by a single turnover saturating flash
(A) The diagram of P515 fast kinetics induced by a single turnover saturating flash, rapid ascent stage (orange), slow ascent stage (blue) and slow descent stage (green), the arrow indicated a single turnover saturating flash (ST); (B) The rapid absorption changes of Chlamydomonas reinhardtii cells (WT) at 520-550 nm induced by a ST (the wavelength of ST was 635 nm, the light intensity was 200000 μmol·m-2·s-1 and the duration was 5 μs), WT (concentration 25 μg·mL-1) was incubated with (red) and without (black) 10 μmol·L-1 3-(3,4-dichlorophenyl)-1,1-dimethylurea (DCMU) and 1 mmol·L-1 hydroxylamine (HA) for 20 min in darkness; (C) The affection of preillumination to the P515 fast kinetics, WT algae was incubated in darkness for 20 min (black) and preilluminated for 2 min at 40 μmol·m-2·s-1 followed by 1 min dark (red).
值得注意的是, 饱和闪光可能会诱导PSI反应中心发生2次电荷分离, 从而导致电荷分离的反应中心数量被高估(Bailleul et al., 2010)。在植物光适应的分子机理研究方面, PSI和PSII反应中心的比例关系为其研究提供了重要的数据支持, 如捕光叶绿素蛋白生物合成缺陷的莱茵衣藻BF4和p71突变体, 其PSI和PSII的天线大小均有不同程度的减小, 而BF4突变体由于PSII的生物发生受阻, 导致PSI/PSII反应中心比例增加至1.25 (Bujaldon et al., 2020)。虽然PSI和PSII的天线大小未发生改变, 但莱茵衣藻还可通过降低PSI/PSII反应中心的比值适应强光变化(Bonente et al., 2012)。缺失质体蓝素(PC)的莱茵衣藻突变体在状态转换过程中可达到完全的状态1, 其PSI/PSII比值比野生型下降10% (Nawrocki et al., 2016)。
拟南芥离体叶片在光强为500 μmol·m-2·s-1的活化光(635 nm)下照光10分钟, 照光期间提供稳定的CO2 (380 μmol·mol-1)、湿度(60%大气湿度)和温度(23°C)。检测黑暗250 ms的P515衰减动力学, 重复100次后取平均值。ATP合酶的质子传导率为P515衰减动力学曲线半衰期(t1/2)的倒数, 即94.3·s-1。箭头表示活化光关闭。 Figure 2Detection the H+ conductivity of chloroplast ATP synthase in Arabidopsis thaliana
The detached leaf of Arabidopsis thaliana was exposed to light (635 nm, 500 μmol·m-2·s-1) for 10 min, and provided stable CO2 (380 μmol·mol-1), humidity (60% atmospheric humidity) and temperature (23°C) during illumination. Detected the P515 decay kinetics curve in darkness that lasted 250 ms, the curve corresponded to 100 independent biological replicates. The H+ conductivity of ATP synthase was 94.3·s-1, which is the reciprocal of the half-time (t1/2) of P515 decay kinetics curve. The arrow indicated light off.
莱茵衣藻野生型(WT)和PGR5突变体(Mutant)在光强为80 μmol·m-2·s-1的活化光(635 nm)下照光5分钟, 关光后检测围绕PSI的环式电子传递速率, 黑暗持续时间为250 ms。测量前, 藻细胞(浓度约25 μg·mL-1)加入10 µmol·L-1二氯苯基二甲脲(DCMU)、1 mmol·L-1羟胺(HA)和20%的聚蔗糖, 暗适应20分钟。箭头表示活化光关闭。 Figure 3Detection the cyclic electron flow rate around photosystem I in Chlamydomonas reinhardtii
The C. reinhardtii cells (WT) and PGR5 mutant (Mutant) were exposed to light (635 nm, 80 μmol·m-2·s-1) for 5 min. The cyclic electron flow rate was detected after the light was turned off and the darkness duration was 250 ms. Before measurement, algal cells (about 25 μg·mL-1) were incubated with 10 μmol·L-1 3-(3,4-dichlorophenyl)-1,1-dimethylurea (DCMU), 1 mmol·L-1 hydroxylamine (HA) and 20% ficoll for 20 min in darkness. The arrow indicated light off.
拟南芥离体叶片在光强为100 μmol·m-2·s-1 (WT 100, 黑)和750 μmol·m-2·s-1 (WT 750, 红)的活化光(635 nm)下照光3分钟, 照光期间提供稳定的CO2 (380 μmol·mol-1)、湿度(60%大气湿度)和温度(23°C)。黑暗时间60秒, 曲线为3次重复的平均值。测量前, 拟南芥叶片暗适应至少3小时。向上箭头表示活化光打开, 向下箭头表示活化光关闭。 Figure 4Detection the proton motive force (pmf) and its two components (Δψ and ΔpH) in Arabidopsis thaliana
The detached leaf of Arabidopsis thaliana was exposed to light (635 nm, 100 μmol·m-2·s-1, labeled as WT 100, black and 750 μmol·m-2·s-1, labeled as WT 750, red) for 3 min, and provided stable CO2 (380 μmol·mol-1), humidity (60% atmospheric humidity) and temperature (23°C) during illumination. The dark time was 60 s and the curve corresponded to 3 independent biological replicates. The Arabidopsis leaf was darkly adapted for at least 3 hours before measurement. The up arrow indicated light on, and the down arrow indicated light off.
将莱茵衣藻野生型(浓度约30 μg·mL-1)暗适应20分钟, 放入荧光仪的测量室, 分别打开P515/535模块和叶绿素荧光模块的测量光, 测定初始荧光产量(Fo)。打开饱和闪光(635 nm, 6000 μmol·m-2·s-1, 持续时间250 ms), 测定黑暗状态下的最大荧光产量(Fm)。之后打开活化光(635 nm, 1500 μmol·m-2·s-1), 持续照光140秒, 期间每隔60秒施加1次饱和闪光, 测定光下最大荧光产量(Fm′)。活化光关闭后测定质子动力势(pmf)及其组分(ΔΨ和ΔpH), 与此同时每隔60秒施加1次饱和闪光, 检测黑暗状态下的荧光产量是否恢复到最大荧光产量(Fm)。Fo: 初始荧光产量; Fm: 最大荧光产量; Fm′: 光下最大荧光产量; Fs: 稳态荧光产量; Actinic light: 活化光; Saturating pulse: 饱和闪光; pmf: 质子动力势; ΔΨ: 跨膜电势; ΔpH: 跨膜质子梯度 Figure 5Simultaneous measurement of the P515 signal and fluorescence in Chlamydomonas reinhardtii
The C. reinhardtii cells (WT, about 30 μg·mL-1) were placed in the chamber of the chlorophyll fluorometer after dark adaptation for 20 minutes. The measuring lights of the P515/535 module and the chlorophyll fluorescence module were turned on respectively to determine the original fluorescence yield (Fo). Then a saturating pulse (635 nm, 6000 μmol·m-2·s-1, duration 250 ms) was turned on to detect the maximal fluorescence yield (Fm) in darkness. Then an actinic light (635 nm, 1500 μmol·m-2·s-1) was turned on for 140 seconds, during which a saturating flash was applied every 60 seconds to detect the maximum fluorescence yield (Fm') under light. The proton motive force (pmf) and its components (ΔΨ and ΔpH) were measured after the actinic light was turned off, meanwhile a saturating pulse was applied every 60 seconds to check whether the fluorescence yield was restored to the maximum fluorescence yield (Fm) in darkness. Fo: Original fluorescence yield; Fm: Maximal fluorescence yield; Fm′: Maximal fluorescence yield under light; Fs: Steady state fluorescence yield; pmf: Proton motive force; ΔΨ: Transmembrane potential; ΔpH: Transmembrane proton gradient
AllorentG, ByrdinM, CarrarettoL, MorosinottoT, SzaboI, FinazziG (2018). Global spectroscopic analysis to study the regulation of the photosynthetic proton motive force: a critical reappraisal Biochim Biophys Acta Bioenerg 1859, 676-683. DOI:10.1016/j.bbabio.2018.07.001URL [本文引用: 1]
AlricJ (2014). Redox and ATP control of photosynthetic cyclic electron flow in Chlamydomonas reinhardtii: (II) involvement of the PGR5-PGRL1 pathway under anaerobic conditions Biochim Biophys Acta Bioenerg 1837, 825-834. DOI:10.1016/j.bbabio.2014.01.024URL [本文引用: 5]
BailleulB, CardolP, BreytonC, FinazziG (2010). Electrochromism: a useful probe to study algal photosynthesis Photosynth Res 106, 179-189. DOI:10.1007/s11120-010-9579-zURL [本文引用: 3]
BakerNR, HarbinsonJ, KramerDM (2007). Determining the limitations and regulation of photosynthetic energy transduction in leaves Plant Cell Environ 30, 1107-1125. PMID:17661750 [本文引用: 1] The light-dependent production of ATP and reductants by the photosynthetic apparatus in vivo involves a series of electron and proton transfers. Consideration is given as to how electron fluxes through photosystem I (PSI), using absorption spectroscopy, and through photosystem II (PSII), using chlorophyll fluorescence analyses, can be estimated in vivo. Measurements of light-induced electrochromic shifts using absorption spectroscopy provide a means of analyzing the proton fluxes across the thylakoid membranes in vivo. Regulation of these electron and proton fluxes is required for the thylakoids to meet the fluctuating metabolic demands of the cell. Chloroplasts exhibit a wide and flexible range of mechanisms to regulate electron and proton fluxes that enable chloroplasts to match light use for ATP and reductant production with the prevailing metabolic requirements. Non-invasive probing of electron fluxes through PSI and PSII, and proton fluxes across the thylakoid membranes can provide insights into the operation of such regulatory processes in vivo.
BonenteG, PippaS, CastellanoS, BassiR, BallottariM (2012). Acclimation of Chlamydomonas reinhardtii to different growth irradiances J Biol Chem 287, 5833-5847. DOI:10.1074/jbc.M111.304279PMID:22205699 [本文引用: 1] We report on the changes the photosynthetic apparatus of Chlamydomonas reinhardtii undergoes upon acclimation to different light intensity. When grown in high light, cells had a faster growth rate and higher biomass production compared with low and control light conditions. However, cells acclimated to low light intensity are indeed able to produce more biomass per photon available as compared with high light-acclimated cells, which dissipate as heat a large part of light absorbed, thus reducing their photosynthetic efficiency. This dissipative state is strictly dependent on the accumulation of LhcSR3, a protein related to light-harvesting complexes, responsible for nonphotochemical quenching in microalgae. Other changes induced in the composition of the photosynthetic apparatus upon high light acclimation consist of an increase of carotenoid content on a chlorophyll basis, particularly zeaxanthin, and a major down-regulation of light absorption capacity by decreasing the chlorophyll content per cell. Surprisingly, the antenna size of both photosystem I and II is not modulated by acclimation; rather, the regulation affects the PSI/PSII ratio. Major effects of the acclimation to low light consist of increased activity of state 1 and 2 transitions and increased contributions of cyclic electron flow.
BujaldonS, KodamaN, RathodMK, TourasseN, OzawaSI, SellésJ, VallonO, TakahashiY, WollmanFA (2020). The BF4 and p71 antenna mutants from Chlamydomonas reinhardtii Biochim Biophys Acta Bioenerg 1861, 148085. DOI:10.1016/j.bbabio.2019.148085URL [本文引用: 1]
CarrarettoL, FormentinE, TeardoE, ChecchettoV, TomizioliM, MorosinottoT, GiacomettiGM, FinazziG, SzabóI (2013). A thylakoid-located two-pore K+ channel controls photosynthetic light utilization in plants Science 342, 114-118. DOI:10.1126/science.1242113PMID:24009357 [本文引用: 1] The size of the light-induced proton motive force (pmf) across the thylakoid membrane of chloroplasts is regulated in response to environmental stimuli. Here, we describe a component of the thylakoid membrane, the two-pore potassium (K(+)) channel TPK3, which modulates the composition of the pmf through ion counterbalancing. Recombinant TPK3 exhibited potassium-selective channel activity sensitive to Ca(2+) and H(+). In Arabidopsis plants, the channel is found in the thylakoid stromal lamellae. Arabidopsis plants silenced for the TPK3 gene display reduced growth and altered thylakoid membrane organization. This phenotype reflects an impaired capacity to generate a normal pmf, which results in reduced CO2 assimilation and deficient nonphotochemical dissipation of excess absorbed light. Thus, the TPK3 channel manages the pmf necessary to convert photochemical energy into physiological functions.
ChecchettoV, TeardoE, CarrarettoL, LeanzaL, SzaboI (2016). Physiology of intracellular potassium channels: a unifying role as mediators of counterion fluxes? Biochim Biophys Acta Bioenerg 1857, 1258-1266. DOI:10.1016/j.bbabio.2016.03.011URL [本文引用: 2]
CruzJA, SackstederCA, KanazawaA, KramerDM (2001). Contribution of electric field (Δψ) to steady-state transthylakoid proton motive force (pmf) in vitro and in vivo. Control of pmf parsing into Δψ and ΔpH by ionic strength Biochemistry 40, 1226-1237. PMID:11170448 [本文引用: 4] The observed levels of Delta G(ATP) in chloroplasts, as well as the activation behavior of the CF(1)CF(0)-ATP synthase, suggest a minimum transthylakoid proton motive force (pmf) equivalent to a Delta pH of approximately 2.5 units. If, as is commonly believed, all transthylakoid pmf is stored as Delta pH, this would indicate a lumen pH of less than approximately 5. In contrast, we have presented evidence that the pH of the thylakoid lumen does not drop below pH approximately 5.8 [Kramer, D. M., Sacksteder, C. A., and Cruz, J. A. (1999) Photosynth. Res. 60, 151-163], leading us to propose that Delta psi can contribute to steady-state pmf. In this work, it is demonstrated, through assays on isolated thylakoids and computer simulations, that thylakoids can store a substantial fraction of pmf as Delta psi, provided that the activities of ions permeable to the thylakoid membrane in the chloroplast stromal compartment are relatively low and the buffering capacity (beta) for protons of the lumen is relatively high. Measurements of the light-induced electrochromic shift (ECS) confirm the ionic strength behavior of steady-state Delta psi in isolated, partially uncoupled thylakoids. Measurements of the ECS in intact plants illuminated for 65 s were consistent with low concentrations of permeable ions and approximately 50% storage of pmf as Delta psi. We propose that the plant cell, possibly at the level of the inner chloroplast envelope, can control the parsing of pmf into Delta psi and Delta pH by regulating the ionic strength and balance of the chloroplast. In addition, this work demonstrates that, under certain conditions, the kinetics of the light-induced ECS can be used to estimate the fractions of pmf stored as Delta psi and Delta pH both in vitro and in vivo.
DavisGA, KanazawaA, SchöttlerMA, KohzumaK, FroehlichJE, RutherfordAW, Satoh-CruzM, MinhasD, TietzS, DhingraA, KramerDM (2016). Limitations to photosynthesis by proton motive force-induced photosystem II photodamage eLife 5, e16921. DOI:10.7554/eLife.16921URL [本文引用: 1]
DavisGA, RutherfordAW, KramerDM (2017). Hacking the thylakoid proton motive force for improved photosynthesis: modulating ion flux rates that control proton motive force partitioning into Δψ and ΔpH Philos Trans R Soc Lond B Biol Sci 372, 20160381. DOI:10.1098/rstb.2016.0381URL [本文引用: 1]
de BianchiS, BallottariM, Dall'OstoL, BassiR (2010). Regulation of plant light harvesting by thermal dissipation of excess energy Biochem Soc Trans 38, 651-660. DOI:10.1042/BST0380651URL [本文引用: 1]
Demmig-AdamsB, CohuCM, MullerO, Adams WWIII (2012). Modulation of photosynthetic energy conversion efficiency in nature: from seconds to seasons Photosynth Res 113, 75-88. DOI:10.1007/s11120-012-9761-6URL [本文引用: 1]
DuanZK, KongFN, ZhangL, LiWJ, ZhangJ, PengLW (2016). A bestrophin-like protein modulates the proton motive force across the thylakoid membrane in Arabidopsis J Integr Plant Biol 58, 848-858. DOI:10.1111/jipb.12475URL [本文引用: 2]
DuysensLNM (1954). Reversible changes in the absorption spectrum of Chlorella upon irradiation Science 120, 353-354. PMID:17753561 [本文引用: 1]
ForkDC, AmeszJ (1967). Light-induced shifts in the absorption spectrum of carotenoids in red and brown algae Photochem Photobiol 6, 913-918. DOI:10.1111/j.1751-1097.1967.tb09656.xURL [本文引用: 1]
FreseRN, PalaciosMA, AzziziA, van StokkumIHM, KruipJ, RögnerM, KarapetyanNV, SchlodderE, van GrondelleR, DekkerJP (2002). Electric field effects on red chlorophylls, β-carotenes and P700 in cyanobacterial Photosystem I complexes Biochim Biophys Acta Bioenerg 1554, 180-191. DOI:10.1016/S0005-2728(02)00242-6URL [本文引用: 1]
FristedtR, MartinsNF, StrenkertD, ClarkeCA, SuchoszekM, ThieleW, SchöttlerMA, MerchantSS (2015). The thylakoid membrane protein CGL160 supports CF1CFo ATP synthase accumulation in Arabidopsis thaliana PLoS One 10, e0121658. DOI:10.1371/journal.pone.0121658URL [本文引用: 1]
HindG, NakataniHY, IzawaS (1974). Light-dependent redistribution of ions in suspensions of chloroplast thylakoid membranes Proc Natl Acad Sci USA 71, 1484-1488. DOI:10.1073/pnas.71.4.1484URL [本文引用: 1]
HiranoM, KatohS (1981). Electrochromic band shifts of carotenoid in a blue-green alga Photochem Photobiol 34, 637-643. DOI:10.1111/php.1981.34.issue-5URL [本文引用: 1]
HolmesNG, HunterCN, NiedermanRA, CroftsAR (1980). Identification of the pigment pool responsible for the flash-induced carotenoid band shift in Rhodopseudomonas sphaeroides chromatophores FEBS Lett 115, 43-48. DOI:10.1016/0014-5793(80)80723-XURL [本文引用: 1]
JoliotP, JoliotA (2002). Cyclic electron transfer in plant leaf Proc Natl Acad Sci USA 99, 10209-10214. DOI:10.1073/pnas.102306999URL [本文引用: 3]
JungeW, WittHT (1968). On the ion transport system of photosynthesis: investigations on a molecular level Z Naturforsch B 23, 244-254. DOI:10.1515/znb-1968-0222URL [本文引用: 1]
KlughammerC, SiebkeK, SchreiberU (2013). Continuous ECS-indicated recording of the proton-motive charge flux in leaves Photosynth Res 117, 471-487. DOI:10.1007/s11120-013-9884-4URL [本文引用: 1]
KramerDM, AvensonTJ, EdwardsGE (2004). Dynamic flexibility in the light reactions of photosynthesis governed by both electron and proton transfer reactions Trends Plant Sci 9, 349-357. PMID:15231280 [本文引用: 1] Plant photosynthesis performs the remarkable feat of converting light energy into usable chemical forms, which involves taming highly reactive intermediates without harming plant cells. This requires an apparatus that is not only efficient and robust but also flexible in its responses to changing environmental conditions. It also requires that the output of the energy-storing reactions be matched with the demands of metabolism. This article addresses the mechanisms by which this flexibility is achieved for short-term environmental changes. We argue that chloroplasts need two types of flexible mechanisms: one for modulating the output ratio of ATP:NADPH, which involves cyclic electron flux around photosystem I; and another for changing the regulatory sensitivity of the light-harvesting antenna to electron (and proton) flow.
KramerDM, CruzJA, KanazawaA (2003). Balancing the central roles of the thylakoid proton gradient Trends Plant Sci 8, 27-32. PMID:12523997 [本文引用: 1] The photosynthetic electron transfer chain generates proton motive force (pmf), composed of both electric field (Deltapsi) and concentration (DeltapH) gradients. Both components can drive ATP synthesis, whereas the DeltapH component alone can trigger feedback regulation of the antenna. It has often been suggested that a relatively large pmf is needed to sustain the energetic contributions of the ATP synthase reaction (DeltaG(ATP)), and that the Deltapsi component is dissipated during illumination, leading to an acidic lumen in the light. We suggest that this is incompatible with the stabilities of lumenal components and the observed activation of downregulation. Recent work on the chloroplast ATP synthase suggests that a more moderate pmf can sustain DeltaG(ATP). In addition, in vivo probes suggest that a substantial fraction of pmf can be stored as Deltapsi. Together, these factors should allow sufficient DeltaG(ATP) to maintain lumen pH in a range where lumenal enzyme activities are nearly optimal, and where the level of NPQ is regulated.
KramerDM, EvansJR (2011). The importance of energy balance in improving photosynthetic productivity Plant Physiol 155, 70-78. DOI:10.1104/pp.110.166652PMID:21078862 [本文引用: 1]
KramerH, MathisP (1980). Quantum yield and rate of formation of the carotenoid triplet state in photosynthetic structures Biochim Biophys Acta Bioenerg 593, 319-329. DOI:10.1016/0005-2728(80)90069-9URL [本文引用: 1]
LiguoriN, RoyLM, OpacicM, DurandG, CroceR (2013). Regulation of light harvesting in the green alga Chlamydomonas reinhardtii: the C-terminus of LHCSR is the knob of a dimmer switch J Am Chem Soc 135, 18339-18342. DOI:10.1021/ja4107463URL [本文引用: 1]
LuckerB, KramerDM (2013). Regulation of cyclic electron flow in Chlamydomonas reinhardtii under fluctuating carbon availability Photosynth Res 117, 449-459. DOI:10.1007/s11120-013-9932-0URL [本文引用: 5]
NandhaB, FinazziG, JoliotP, HaldS, JohnsonGN (2007). The role of PGR5 in the redox poising of photosynthetic electron transport Biochim Biophys Acta Bioenerg 1767, 1252-1259. DOI:10.1016/j.bbabio.2007.07.007URL [本文引用: 1]
NawrockiWJ, SantabarbaraS, MosebachL, WollmanFA, RappaportF (2016). State transitions redistribute rather than dissipate energy between the two photosystems in Chlamydomonas Nat Plants 2, 16031. DOI:10.1038/nplants.2016.31URL [本文引用: 1]
NiyogiKK, GrossmanAR, BjörkmanO (1998). Arabidopsis mutants define a central role for the xanthophyll cycle in the regulation of photosynthetic energy conversion Plant Cell 10, 1121-1134. PMID:9668132 [本文引用: 1] A conserved regulatory mechanism protects plants against the potentially damaging effects of excessive light. Nearly all photosynthetic eukaryotes are able to dissipate excess absorbed light energy in a process that involves xanthophyll pigments. To dissect the role of xanthophylls in photoprotective energy dissipation in vivo, we isolated Arabidopsis xanthophyll cycle mutants by screening for altered nonphotochemical quenching of chlorophyll fluorescence. The npq1 mutants are unable to convert violaxanthin to zeaxanthin in excessive light, whereas the npq2 mutants accumulate zeaxanthin constitutively. The npq2 mutants are new alleles of aba1, the zeaxanthin epoxidase gene. The high levels of zeaxanthin in npq2 affected the kinetics of induction and relaxation but not the extent of nonphotochemical quenching. Genetic mapping, DNA sequencing, and complementation of npq1 demonstrated that this mutation affects the structural gene encoding violaxanthin deepoxidase. The npq1 mutant exhibited greatly reduced nonphotochemical quenching, demonstrating that violaxanthin deepoxidation is required for the bulk of rapidly reversible nonphotochemical quenching in Arabidopsis. Altered regulation of photosynthetic energy conversion in npq1 was associated with increased sensitivity to photoinhibition. These results, in conjunction with the analysis of npq mutants of Chlamydomonas, suggest that the role of the xanthophyll cycle in nonphotochemical quenching has been conserved, although different photosynthetic eukaryotes rely on the xanthophyll cycle to different extents for the dissipation of excess absorbed light energy.
PottosinII, SchönknechtG (1996). Ion channel permeable for divalent and monovalent cations in native spinach thylakoid membranes J Membr Biol 152, 223-233. DOI:10.1007/s002329900100URL [本文引用: 1]
RottM, MartinsNF, ThieleW, LeinW, BockR, KramerDM, SchöttlerMA (2011). ATP synthase repression in tobacco restricts photosynthetic electron transport, CO2 assimilation, and plant growth by overacidification of the thylakoid lumen Plant Cell 23, 304-321. DOI:10.1105/tpc.110.079111URL [本文引用: 2]
RubanAV, PascalAA, RobertB, HortonP (2002). Activation of zeaxanthin is an obligatory event in the regulation of photosynthetic light harvesting J Biol Chem 277, 7785-7789. DOI:10.1074/jbc.M110693200URL [本文引用: 1]
SackstederCA, KanazawaA, JacobyME, KramerDM (2000). The proton to electron stoichiometry of steady- state photosynthesis in living plants: a proton-pumping Q cycle is continuously engaged Proc Natl Acad Sci USA 97, 14283-14288. DOI:10.1073/pnas.97.26.14283URL [本文引用: 1]
SackstederCA, KramerDM (2000). Dark-interval relaxation kinetics (DIRK) of absorbance changes as a quantitative probe of steady-state electron transfer Photosynth Res 66, 145-158. DOI:10.1023/A:1010785912271URL [本文引用: 2]
SchmidtS, ReichR, WittHT (1971). Electrochromism of chlorophylls and carotenoids in multilayers and in chloroplasts Naturwissenschaften 58, 414. [本文引用: 1]
SchönknechtG, HedrichR, JungeW, RaschkeK (1988). A voltage-dependent chloride channel in the photosynthetic membrane of a higher plant Nature 336, 589-592. DOI:10.1038/336589a0URL [本文引用: 1]
SchreiberU, KlughammerC (2008). New accessory for the DUAL-PAM-100: the P515/535 module and examples of its application PAM Appl Notes 1, 1-10. [本文引用: 6]
SpeteaC, HerdeanA, AllorentG, CarrarettoL, FinazziG, SzaboI (2017). An update on the regulation of photosynthesis by thylakoid ion channels and transporters in Arabidopsis Physiol Plant 161, 16-27. DOI:10.1111/ppl.2017.161.issue-1URL [本文引用: 1]
SukhovV, SurovaL, MorozovaE, SherstnevaO, VodeneevV (2016). Changes in H+-ATP synthase activity, proton electrochemical gradient, and pH inPea chloroplast can be connected with variation potential Front Plant Sci 7, 1092. [本文引用: 2]
TakahashiH, ClowezS, WollmanFA, VallonO, RappaportF (2013). Cyclic electron flow is redox-controlled but independent of state transition Nat Commun 4, 1954. DOI:10.1038/ncomms2954PMID:23760547 [本文引用: 1] Photosynthesis is the biological process that feeds the biosphere with reduced carbon. The assimilation of CO2 requires the fine tuning of two co-existing functional modes: linear electron flow, which provides NADPH and ATP, and cyclic electron flow, which only sustains ATP synthesis. Although the importance of this fine tuning is appreciated, its mechanism remains equivocal. Here we show that cyclic electron flow as well as formation of supercomplexes, thought to contribute to the enhancement of cyclic electron flow, are promoted in reducing conditions with no correlation with the reorganization of the thylakoid membranes associated with the migration of antenna proteins towards Photosystems I or II, a process known as state transition. We show that cyclic electron flow is tuned by the redox power and this provides a mechanistic model applying to the entire green lineage including the vast majority of the cases in which state transition only involves a moderate fraction of the antenna.
TakizawaK, CruzJA, KanazawaA, KramerDM (2007). The thylakoid proton motive force in vivo. Quantitative, non-invasive probes, energetics, and regulatory consequences of light-induced pmf Biochim Biophys Acta Bioenerg 1767, 1233-1244. DOI:10.1016/j.bbabio.2007.07.006URL [本文引用: 1]
TolleterD, GhyselsB, AlricJ, PetroutsosD, TolstyginaI, KrawietzD, HappeT, AuroyP, AdrianoJM, BeylyA, CuinéS, PletJ, ReiterIM, GentyB, CournacL, HipplerM, PeltierG (2011). Control of hydrogen photoproduction by the proton gradient generated by cyclic electron flow in Chlamydomonas reinhardtii Plant Cell 23, 2619-2630. DOI:10.1105/tpc.111.086876URL [本文引用: 1]
ViolaS, BailleulB, YuJF, NixonP, SellésJ, JoliotP, WollmanFA (2019). Probing the electric field across thylakoid membranes in cyanobacteria Proc Natl Acad Sci USA 116, 21900-21906. DOI:10.1073/pnas.1913099116URL [本文引用: 2]
WangP, DuanW, TakabayashiA, EndoT, ShikanaiT, YeJY, MiHL (2006). Chloroplastic NAD(P)H dehydrogenase in tobacco leaves functions in alleviation of oxidative damage caused by temperature stress Plant Physiol 141, 465-474. DOI:10.1104/pp.105.070490URL [本文引用: 2]
WittHT (1979). Energy conversion in the functional membrane of photosynthesis. Analysis by light pulse and electric pulse methods: the central role of the electric field Biochim Biophys Acta 505, 355-427. PMID:35227 [本文引用: 1]
YamoriW, TakahashiS, MakinoA, PriceGD, BadgerMR, von CaemmererS (2011). The roles of ATP synthase and the cytochrome b 6/f complexes in limiting chloroplast electron transport and determining photosynthetic capacity Plant Physiol 155, 956-962. DOI:10.1104/pp.110.168435URL [本文引用: 1]
Global spectroscopic analysis to study the regulation of the photosynthetic proton motive force: a critical reappraisal 1 2018
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
Redox and ATP control of photosynthetic cyclic electron flow in Chlamydomonas reinhardtii: (II) involvement of the PGR5-PGRL1 pathway under anaerobic conditions 5 2014
... 值得注意的是, 饱和闪光可能会诱导PSI反应中心发生2次电荷分离, 从而导致电荷分离的反应中心数量被高估(Bailleul et al., 2010).在植物光适应的分子机理研究方面, PSI和PSII反应中心的比例关系为其研究提供了重要的数据支持, 如捕光叶绿素蛋白生物合成缺陷的莱茵衣藻BF4和p71突变体, 其PSI和PSII的天线大小均有不同程度的减小, 而BF4突变体由于PSII的生物发生受阻, 导致PSI/PSII反应中心比例增加至1.25 (Bujaldon et al., 2020).虽然PSI和PSII的天线大小未发生改变, 但莱茵衣藻还可通过降低PSI/PSII反应中心的比值适应强光变化(Bonente et al., 2012).缺失质体蓝素(PC)的莱茵衣藻突变体在状态转换过程中可达到完全的状态1, 其PSI/PSII比值比野生型下降10% (Nawrocki et al., 2016). ...
Determining the limitations and regulation of photosynthetic energy transduction in leaves 1 2007
... 当植物受到环境胁迫时, 环式电子传递的主要作用可能是通过增加ATP的供给保障光合作用高效运行(Kramer et al., 2004; Nandha et al., 2007).例如, 当烟草受到热胁迫且CO2同化受阻时, NDH (NAD (P)H dehydrogenase)介导的环式电子传递会被快速激活, 从而启动环式光合磷酸化产生ATP (Wang et al., 2006).由于ATP与NADPH的产生和消耗紧密相关, 因此围绕PSI的环式电子传递可平衡ATP与NADPH的比值(Kramer and Evans, 2011).例如, 在低浓度无机碳条件下, 莱茵衣藻可能通过增大环式电子传递速率满足对ATP/NADPH增加的需求(Lucker and Kramer, 2013).该方法的缺点是样品经过DCMU等抑制剂处理后, 会影响细胞内的氧化还原状态或者代谢状态, 这些因素可能会激活CEF的调控机制, 进而影响CEF检测的准确性(Lucker and Kramer, 2013).在不使用PSII抑制剂的条件下, 可利用gH+与线式电子传递速率的比值估算环式电子的相对传递速率, 其中线式电子传递速率可通过叶绿素荧光检测PSII的光化学量子产量, 该量子产量乘以光强得到.该方法的优点是不会破坏样品正常的生理状态(Sacksteder and Kramer, 2000; Joliot and Joliot, 2002; Baker et al., 2007; Lucker and Kramer, 2013). ...
Chlororespiration revisited: mitochondrial-plastid interactions in Chlamydomonas 1 1994
... P515分钟级的慢速动力学比较复杂, 既反映质子动力势的生成, 也反映玉米黄素的生成.质子动力势(proton motive force, pmf)驱动ATP合酶产生ATP, 而pmf的组分——ΔpH会诱导光保护机制, 将过剩的光能以热能的形式耗散, 保护PSII免受光损伤(Niyogi et al., 1998; de Bianchi et al., 2010; Checchetto et al., 2013, 2016), 还可通过降低电子在Cytb6f复合物的传递速率, 保护PSI免受光抑制(Sonoike, 2011; Yamori et al., 2011).跨膜电势(ΔΨ)也可调节PSII的光损伤.研究发现, 高水平的ΔΨ不仅降低NPQ的诱导效率(Duan et al., 2016)、增加PSII的光敏感性(Davis et al., 2016), 还导致PSII反应中心发生电荷重组, 促使单线态氧水平升高, 造成严重的光损伤(Bennoun, 1994).我们的研究结果显示, 拟南芥在100 μmol·m-2·s-1的光强下, pmf为1.3, ΔpH约占56%, 而当光强达到750 μmol·m-2·s-1时, pmf增至7.2, ΔpH所占比例增至68%, ΔΨ减少至32% (图4), 说明ΔpH可能在光保护中起更关键的作用. ...
Acclimation of Chlamydomonas reinhardtii to different growth irradiances 1 2012
... 值得注意的是, 饱和闪光可能会诱导PSI反应中心发生2次电荷分离, 从而导致电荷分离的反应中心数量被高估(Bailleul et al., 2010).在植物光适应的分子机理研究方面, PSI和PSII反应中心的比例关系为其研究提供了重要的数据支持, 如捕光叶绿素蛋白生物合成缺陷的莱茵衣藻BF4和p71突变体, 其PSI和PSII的天线大小均有不同程度的减小, 而BF4突变体由于PSII的生物发生受阻, 导致PSI/PSII反应中心比例增加至1.25 (Bujaldon et al., 2020).虽然PSI和PSII的天线大小未发生改变, 但莱茵衣藻还可通过降低PSI/PSII反应中心的比值适应强光变化(Bonente et al., 2012).缺失质体蓝素(PC)的莱茵衣藻突变体在状态转换过程中可达到完全的状态1, 其PSI/PSII比值比野生型下降10% (Nawrocki et al., 2016). ...
The BF4 and p71 antenna mutants from Chlamydomonas reinhardtii 1 2020
... 值得注意的是, 饱和闪光可能会诱导PSI反应中心发生2次电荷分离, 从而导致电荷分离的反应中心数量被高估(Bailleul et al., 2010).在植物光适应的分子机理研究方面, PSI和PSII反应中心的比例关系为其研究提供了重要的数据支持, 如捕光叶绿素蛋白生物合成缺陷的莱茵衣藻BF4和p71突变体, 其PSI和PSII的天线大小均有不同程度的减小, 而BF4突变体由于PSII的生物发生受阻, 导致PSI/PSII反应中心比例增加至1.25 (Bujaldon et al., 2020).虽然PSI和PSII的天线大小未发生改变, 但莱茵衣藻还可通过降低PSI/PSII反应中心的比值适应强光变化(Bonente et al., 2012).缺失质体蓝素(PC)的莱茵衣藻突变体在状态转换过程中可达到完全的状态1, 其PSI/PSII比值比野生型下降10% (Nawrocki et al., 2016). ...
A thylakoid-located two-pore K+ channel controls photosynthetic light utilization in plants 1 2013
... 虽然ΔpH和ΔΨ在质子动力势中的分配机制还不清楚, 但是多种离子通道或转运蛋白参与调节两个组分的分配, 进而影响光合效率(Checchetto et al., 2013, 2016; Davis et al., 2017; Spetea et al., 2017).例如, 双孔K+通道蛋白TPK3缺失的拟南芥突变体, 由于K+外排受阻导致ΔpH的形成受损, 同时NPQ诱导效率降低(Carraretto et al., 2013); Cl-通道蛋白AtBest缺失会导致拟南芥pmf和ΔΨ水平显著升高, 说明Cl-向囊腔内的转运可消散ΔΨ, 从而调控pmf的分配(Duan et al., 2016). ...
Ion channels in plant bioenergetic organelles, chloroplasts and mitochondria: from molecular identification to function 1 2016
... 在光合作用中, 放氧光合生物吸收太阳能并裂解水分子, 在类囊体囊腔释放大量的H+并驱动电子在膜蛋白复合物和电子载体间传递, 最终生成还原型烟酰胺腺嘌呤二核苷酸磷酸(辅酶II) (NADPH).同时, 光合电子的传递导致质子从叶绿体基质向类囊体囊腔传递并伴随多种离子的跨膜转运, 随后卡尔文循环消耗质子进行固碳, 又导致叶绿体基质碱化, 这些因素共同诱导形成跨膜电势(ΔΨ)和质子梯度(ΔpH), 即质子动力势(proton motive force, pmf) (Cruz et al., 2001; Carraretto et al., 2016).质子动力势驱动ATP合酶产生腺苷三磷酸(ATP), ATP和NADPH为光合作用中卡尔文循环及其它代谢途径提供能量和还原力(Kramer et al., 2003; Takizawa et al., 2007). ...
Regulation of photosynthesis by ion channels in cyanobacteria and higher plants 2 2013
... P515分钟级的慢速动力学比较复杂, 既反映质子动力势的生成, 也反映玉米黄素的生成.质子动力势(proton motive force, pmf)驱动ATP合酶产生ATP, 而pmf的组分——ΔpH会诱导光保护机制, 将过剩的光能以热能的形式耗散, 保护PSII免受光损伤(Niyogi et al., 1998; de Bianchi et al., 2010; Checchetto et al., 2013, 2016), 还可通过降低电子在Cytb6f复合物的传递速率, 保护PSI免受光抑制(Sonoike, 2011; Yamori et al., 2011).跨膜电势(ΔΨ)也可调节PSII的光损伤.研究发现, 高水平的ΔΨ不仅降低NPQ的诱导效率(Duan et al., 2016)、增加PSII的光敏感性(Davis et al., 2016), 还导致PSII反应中心发生电荷重组, 促使单线态氧水平升高, 造成严重的光损伤(Bennoun, 1994).我们的研究结果显示, 拟南芥在100 μmol·m-2·s-1的光强下, pmf为1.3, ΔpH约占56%, 而当光强达到750 μmol·m-2·s-1时, pmf增至7.2, ΔpH所占比例增至68%, ΔΨ减少至32% (图4), 说明ΔpH可能在光保护中起更关键的作用. ...
... 虽然ΔpH和ΔΨ在质子动力势中的分配机制还不清楚, 但是多种离子通道或转运蛋白参与调节两个组分的分配, 进而影响光合效率(Checchetto et al., 2013, 2016; Davis et al., 2017; Spetea et al., 2017).例如, 双孔K+通道蛋白TPK3缺失的拟南芥突变体, 由于K+外排受阻导致ΔpH的形成受损, 同时NPQ诱导效率降低(Carraretto et al., 2013); Cl-通道蛋白AtBest缺失会导致拟南芥pmf和ΔΨ水平显著升高, 说明Cl-向囊腔内的转运可消散ΔΨ, 从而调控pmf的分配(Duan et al., 2016). ...
Physiology of intracellular potassium channels: a unifying role as mediators of counterion fluxes? 2 2016
... P515分钟级的慢速动力学比较复杂, 既反映质子动力势的生成, 也反映玉米黄素的生成.质子动力势(proton motive force, pmf)驱动ATP合酶产生ATP, 而pmf的组分——ΔpH会诱导光保护机制, 将过剩的光能以热能的形式耗散, 保护PSII免受光损伤(Niyogi et al., 1998; de Bianchi et al., 2010; Checchetto et al., 2013, 2016), 还可通过降低电子在Cytb6f复合物的传递速率, 保护PSI免受光抑制(Sonoike, 2011; Yamori et al., 2011).跨膜电势(ΔΨ)也可调节PSII的光损伤.研究发现, 高水平的ΔΨ不仅降低NPQ的诱导效率(Duan et al., 2016)、增加PSII的光敏感性(Davis et al., 2016), 还导致PSII反应中心发生电荷重组, 促使单线态氧水平升高, 造成严重的光损伤(Bennoun, 1994).我们的研究结果显示, 拟南芥在100 μmol·m-2·s-1的光强下, pmf为1.3, ΔpH约占56%, 而当光强达到750 μmol·m-2·s-1时, pmf增至7.2, ΔpH所占比例增至68%, ΔΨ减少至32% (图4), 说明ΔpH可能在光保护中起更关键的作用. ...
... 虽然ΔpH和ΔΨ在质子动力势中的分配机制还不清楚, 但是多种离子通道或转运蛋白参与调节两个组分的分配, 进而影响光合效率(Checchetto et al., 2013, 2016; Davis et al., 2017; Spetea et al., 2017).例如, 双孔K+通道蛋白TPK3缺失的拟南芥突变体, 由于K+外排受阻导致ΔpH的形成受损, 同时NPQ诱导效率降低(Carraretto et al., 2013); Cl-通道蛋白AtBest缺失会导致拟南芥pmf和ΔΨ水平显著升高, 说明Cl-向囊腔内的转运可消散ΔΨ, 从而调控pmf的分配(Duan et al., 2016). ...
Contribution of electric field (Δψ) to steady-state transthylakoid proton motive force (pmf) in vitro and in vivo. Control of pmf parsing into Δψ and ΔpH by ionic strength 4 2001
... 在光合作用中, 放氧光合生物吸收太阳能并裂解水分子, 在类囊体囊腔释放大量的H+并驱动电子在膜蛋白复合物和电子载体间传递, 最终生成还原型烟酰胺腺嘌呤二核苷酸磷酸(辅酶II) (NADPH).同时, 光合电子的传递导致质子从叶绿体基质向类囊体囊腔传递并伴随多种离子的跨膜转运, 随后卡尔文循环消耗质子进行固碳, 又导致叶绿体基质碱化, 这些因素共同诱导形成跨膜电势(ΔΨ)和质子梯度(ΔpH), 即质子动力势(proton motive force, pmf) (Cruz et al., 2001; Carraretto et al., 2016).质子动力势驱动ATP合酶产生腺苷三磷酸(ATP), ATP和NADPH为光合作用中卡尔文循环及其它代谢途径提供能量和还原力(Kramer et al., 2003; Takizawa et al., 2007). ...
... 将暗适应3小时(为了降低玉米黄素的含量)的拟南芥离体叶片放入双通道调制叶绿素荧光仪叶室内, 打开测量光, 待基线平稳后, 打开活化光(635 nm, 750 μmol·m-2·s-1), PSII和PSI反应中心发生原初电荷分离, 从而导致P515信号迅速增加.在持续照光(3 分钟)过程中, 类囊体腔内不断积累的质子逐渐被流入的阴离子(主要是Cl-)中和, P515信号达到相对稳态水平(Cruz et al., 2001; Schreiber and Klughammer, 2008).活化光关闭瞬间, 类囊体腔内积累的质子以不变的速率通过ATP合酶快速释放到类囊体基质, 造成囊腔内阴离子过剩, 从而引起信号反转, 表现为P515信号迅速下降, 下降的最大幅度即质子动力势(pmf).活化光关闭后, 随着类囊体腔内阴离子(主要是Cl-)的外流, 基质中阳离子(主要是K+和Mg2+)内流, 囊腔内外离子浓度逐渐达到平衡(Hind et al.,1974; Schönknecht et al., 1988; Pottosin and Schönknecht, 1996), 反转的P515信号又慢慢恢复, 表现为信号缓慢上升并逐渐达到“暗基线”(Cruz et al., 2001; Schreiber and Klughammer, 2008).pmf及其组分Δψ和ΔpH的计算方法依据文献(Schreiber and Klughammer, 2008). ...
... ), 反转的P515信号又慢慢恢复, 表现为信号缓慢上升并逐渐达到“暗基线”(Cruz et al., 2001; Schreiber and Klughammer, 2008).pmf及其组分Δψ和ΔpH的计算方法依据文献(Schreiber and Klughammer, 2008). ...
... 当植物处于稳态的光合作用时, 通过光合电子传递引起的质子流入类囊体囊腔的速率与通过ATP合酶的质子流出速率相等(Sacksteder and Kramer, 2000).DIRK (dark internal relaxation kinetics)技术是利用短暂的黑暗(<500 ms)干扰处于稳态光合的电子和质子流, 检测样品在520 nm处的光吸收(Sacksteder et al., 2000).该方法可估算叶绿体ATP合酶的活性(Cruz et al., 2001).其原理是当ATP合酶活性降低时, 通过ATP合酶的质子流出速率也会相应降低.拟南芥野生型在500 μmol·m-2·s-1的光照强度下, 叶绿体ATP合酶的质子传导率(gH+)为94.3·s-1 (图2).拟南芥bfa1和bfa3是两个ATP合酶突变体, 在不同光照强度下, 其gH+分别下降至野生型的30%和50%, 同时均表现出较高的NPQ (Zhang et al., 2016, 2018).这是因为当ATP合酶活性降低时, 通过ATP合酶的质子流出效率降低, 导致质子在类囊体囊腔累积, 囊腔过度酸化会抑制NPQ衰减, 从而表现出高水平的NPQ.烟草(Nicotiana tabacum) GTG-atpB突变体的gH+降至野生型的25%, 且突变体的碳同化能力、线式电子传递速率和P700+再还原能力也显著降低(Rott et al., 2011).P515与叶绿素荧光、P700氧化还原动力学的结合使用在烟草、玉米(Zea mays)、拟南芥和豌豆(Pisum sativum)等叶绿体ATP合酶突变体的表型筛选和功能研究方面发挥了重要作用(Rott et al., 2011; Zoschke et al., 2012; Fristedt et al., 2015; Sukhov et al., 2016; Zhang et al., 2016). ...
Limitations to photosynthesis by proton motive force-induced photosystem II photodamage 1 2016
... P515分钟级的慢速动力学比较复杂, 既反映质子动力势的生成, 也反映玉米黄素的生成.质子动力势(proton motive force, pmf)驱动ATP合酶产生ATP, 而pmf的组分——ΔpH会诱导光保护机制, 将过剩的光能以热能的形式耗散, 保护PSII免受光损伤(Niyogi et al., 1998; de Bianchi et al., 2010; Checchetto et al., 2013, 2016), 还可通过降低电子在Cytb6f复合物的传递速率, 保护PSI免受光抑制(Sonoike, 2011; Yamori et al., 2011).跨膜电势(ΔΨ)也可调节PSII的光损伤.研究发现, 高水平的ΔΨ不仅降低NPQ的诱导效率(Duan et al., 2016)、增加PSII的光敏感性(Davis et al., 2016), 还导致PSII反应中心发生电荷重组, 促使单线态氧水平升高, 造成严重的光损伤(Bennoun, 1994).我们的研究结果显示, 拟南芥在100 μmol·m-2·s-1的光强下, pmf为1.3, ΔpH约占56%, 而当光强达到750 μmol·m-2·s-1时, pmf增至7.2, ΔpH所占比例增至68%, ΔΨ减少至32% (图4), 说明ΔpH可能在光保护中起更关键的作用. ...
Hacking the thylakoid proton motive force for improved photosynthesis: modulating ion flux rates that control proton motive force partitioning into Δψ and ΔpH 1 2017
... 虽然ΔpH和ΔΨ在质子动力势中的分配机制还不清楚, 但是多种离子通道或转运蛋白参与调节两个组分的分配, 进而影响光合效率(Checchetto et al., 2013, 2016; Davis et al., 2017; Spetea et al., 2017).例如, 双孔K+通道蛋白TPK3缺失的拟南芥突变体, 由于K+外排受阻导致ΔpH的形成受损, 同时NPQ诱导效率降低(Carraretto et al., 2013); Cl-通道蛋白AtBest缺失会导致拟南芥pmf和ΔΨ水平显著升高, 说明Cl-向囊腔内的转运可消散ΔΨ, 从而调控pmf的分配(Duan et al., 2016). ...
Regulation of plant light harvesting by thermal dissipation of excess energy 1 2010
... P515分钟级的慢速动力学比较复杂, 既反映质子动力势的生成, 也反映玉米黄素的生成.质子动力势(proton motive force, pmf)驱动ATP合酶产生ATP, 而pmf的组分——ΔpH会诱导光保护机制, 将过剩的光能以热能的形式耗散, 保护PSII免受光损伤(Niyogi et al., 1998; de Bianchi et al., 2010; Checchetto et al., 2013, 2016), 还可通过降低电子在Cytb6f复合物的传递速率, 保护PSI免受光抑制(Sonoike, 2011; Yamori et al., 2011).跨膜电势(ΔΨ)也可调节PSII的光损伤.研究发现, 高水平的ΔΨ不仅降低NPQ的诱导效率(Duan et al., 2016)、增加PSII的光敏感性(Davis et al., 2016), 还导致PSII反应中心发生电荷重组, 促使单线态氧水平升高, 造成严重的光损伤(Bennoun, 1994).我们的研究结果显示, 拟南芥在100 μmol·m-2·s-1的光强下, pmf为1.3, ΔpH约占56%, 而当光强达到750 μmol·m-2·s-1时, pmf增至7.2, ΔpH所占比例增至68%, ΔΨ减少至32% (图4), 说明ΔpH可能在光保护中起更关键的作用. ...
Modulation of photosynthetic energy conversion efficiency in nature: from seconds to seasons 1 2012
A bestrophin-like protein modulates the proton motive force across the thylakoid membrane in Arabidopsis 2 2016
... P515分钟级的慢速动力学比较复杂, 既反映质子动力势的生成, 也反映玉米黄素的生成.质子动力势(proton motive force, pmf)驱动ATP合酶产生ATP, 而pmf的组分——ΔpH会诱导光保护机制, 将过剩的光能以热能的形式耗散, 保护PSII免受光损伤(Niyogi et al., 1998; de Bianchi et al., 2010; Checchetto et al., 2013, 2016), 还可通过降低电子在Cytb6f复合物的传递速率, 保护PSI免受光抑制(Sonoike, 2011; Yamori et al., 2011).跨膜电势(ΔΨ)也可调节PSII的光损伤.研究发现, 高水平的ΔΨ不仅降低NPQ的诱导效率(Duan et al., 2016)、增加PSII的光敏感性(Davis et al., 2016), 还导致PSII反应中心发生电荷重组, 促使单线态氧水平升高, 造成严重的光损伤(Bennoun, 1994).我们的研究结果显示, 拟南芥在100 μmol·m-2·s-1的光强下, pmf为1.3, ΔpH约占56%, 而当光强达到750 μmol·m-2·s-1时, pmf增至7.2, ΔpH所占比例增至68%, ΔΨ减少至32% (图4), 说明ΔpH可能在光保护中起更关键的作用. ...
... 虽然ΔpH和ΔΨ在质子动力势中的分配机制还不清楚, 但是多种离子通道或转运蛋白参与调节两个组分的分配, 进而影响光合效率(Checchetto et al., 2013, 2016; Davis et al., 2017; Spetea et al., 2017).例如, 双孔K+通道蛋白TPK3缺失的拟南芥突变体, 由于K+外排受阻导致ΔpH的形成受损, 同时NPQ诱导效率降低(Carraretto et al., 2013); Cl-通道蛋白AtBest缺失会导致拟南芥pmf和ΔΨ水平显著升高, 说明Cl-向囊腔内的转运可消散ΔΨ, 从而调控pmf的分配(Duan et al., 2016). ...
Reversible changes in the absorption spectrum of Chlorella upon irradiation 1 1954
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
Light-induced shifts in the absorption spectrum of carotenoids in red and brown algae 1 1967
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
Electric field effects on red chlorophylls, β-carotenes and P700 in cyanobacterial Photosystem I complexes 1 2002
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
The thylakoid membrane protein CGL160 supports CF1CFo ATP synthase accumulation in Arabidopsis thaliana 1 2015
... 当植物处于稳态的光合作用时, 通过光合电子传递引起的质子流入类囊体囊腔的速率与通过ATP合酶的质子流出速率相等(Sacksteder and Kramer, 2000).DIRK (dark internal relaxation kinetics)技术是利用短暂的黑暗(<500 ms)干扰处于稳态光合的电子和质子流, 检测样品在520 nm处的光吸收(Sacksteder et al., 2000).该方法可估算叶绿体ATP合酶的活性(Cruz et al., 2001).其原理是当ATP合酶活性降低时, 通过ATP合酶的质子流出速率也会相应降低.拟南芥野生型在500 μmol·m-2·s-1的光照强度下, 叶绿体ATP合酶的质子传导率(gH+)为94.3·s-1 (图2).拟南芥bfa1和bfa3是两个ATP合酶突变体, 在不同光照强度下, 其gH+分别下降至野生型的30%和50%, 同时均表现出较高的NPQ (Zhang et al., 2016, 2018).这是因为当ATP合酶活性降低时, 通过ATP合酶的质子流出效率降低, 导致质子在类囊体囊腔累积, 囊腔过度酸化会抑制NPQ衰减, 从而表现出高水平的NPQ.烟草(Nicotiana tabacum) GTG-atpB突变体的gH+降至野生型的25%, 且突变体的碳同化能力、线式电子传递速率和P700+再还原能力也显著降低(Rott et al., 2011).P515与叶绿素荧光、P700氧化还原动力学的结合使用在烟草、玉米(Zea mays)、拟南芥和豌豆(Pisum sativum)等叶绿体ATP合酶突变体的表型筛选和功能研究方面发挥了重要作用(Rott et al., 2011; Zoschke et al., 2012; Fristedt et al., 2015; Sukhov et al., 2016; Zhang et al., 2016). ...
Light-dependent redistribution of ions in suspensions of chloroplast thylakoid membranes 1 1974
... 将暗适应3小时(为了降低玉米黄素的含量)的拟南芥离体叶片放入双通道调制叶绿素荧光仪叶室内, 打开测量光, 待基线平稳后, 打开活化光(635 nm, 750 μmol·m-2·s-1), PSII和PSI反应中心发生原初电荷分离, 从而导致P515信号迅速增加.在持续照光(3 分钟)过程中, 类囊体腔内不断积累的质子逐渐被流入的阴离子(主要是Cl-)中和, P515信号达到相对稳态水平(Cruz et al., 2001; Schreiber and Klughammer, 2008).活化光关闭瞬间, 类囊体腔内积累的质子以不变的速率通过ATP合酶快速释放到类囊体基质, 造成囊腔内阴离子过剩, 从而引起信号反转, 表现为P515信号迅速下降, 下降的最大幅度即质子动力势(pmf).活化光关闭后, 随着类囊体腔内阴离子(主要是Cl-)的外流, 基质中阳离子(主要是K+和Mg2+)内流, 囊腔内外离子浓度逐渐达到平衡(Hind et al.,1974; Schönknecht et al., 1988; Pottosin and Schönknecht, 1996), 反转的P515信号又慢慢恢复, 表现为信号缓慢上升并逐渐达到“暗基线”(Cruz et al., 2001; Schreiber and Klughammer, 2008).pmf及其组分Δψ和ΔpH的计算方法依据文献(Schreiber and Klughammer, 2008). ...
Electrochromic band shifts of carotenoid in a blue-green alga 1 1981
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
Identification of the pigment pool responsible for the flash-induced carotenoid band shift in Rhodopseudomonas sphaeroides chromatophores 1 1980
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
Cyclic electron transfer in plant leaf 3 2002
... 在高等植物和真核藻类中存在线式(LEF)和环式(CEF) 2种电子传递途径, 环式电子传递对光合作用的功能调控已成为研究热点(Joliot and Joliot, 2002; Wang et al., 2006; Lucker and Kramer, 2013; Takahashi et al., 2013; Alric, 2014).目前检测CEF的常用方法是将PSII光合活性抑制后(加入DCMU和HA), 通过P700+的再还原速率计算环式电子传递速率.上文提到利用DIRK技术可测量通过ATP合酶的质子流出速率, 而该速率等于光驱动的通过PSI和PSII的总电子传递速率(Joliot and Joliot, 2002).因此, 将PSII光合活性抑制后, 通过ATP合酶的质子外流速率等于围绕PSI的环式电子传递速率.已有研究表明, 将莱茵衣藻PSII光合活性抑制后, P515与P700氧化还原动力学检测到的围绕PSI的环式电子传递速率呈线性关系(Lucker and Kramer, 2013; Alric, 2014). ...
... 的再还原速率计算环式电子传递速率.上文提到利用DIRK技术可测量通过ATP合酶的质子流出速率, 而该速率等于光驱动的通过PSI和PSII的总电子传递速率(Joliot and Joliot, 2002).因此, 将PSII光合活性抑制后, 通过ATP合酶的质子外流速率等于围绕PSI的环式电子传递速率.已有研究表明, 将莱茵衣藻PSII光合活性抑制后, P515与P700氧化还原动力学检测到的围绕PSI的环式电子传递速率呈线性关系(Lucker and Kramer, 2013; Alric, 2014). ...
... 当植物受到环境胁迫时, 环式电子传递的主要作用可能是通过增加ATP的供给保障光合作用高效运行(Kramer et al., 2004; Nandha et al., 2007).例如, 当烟草受到热胁迫且CO2同化受阻时, NDH (NAD (P)H dehydrogenase)介导的环式电子传递会被快速激活, 从而启动环式光合磷酸化产生ATP (Wang et al., 2006).由于ATP与NADPH的产生和消耗紧密相关, 因此围绕PSI的环式电子传递可平衡ATP与NADPH的比值(Kramer and Evans, 2011).例如, 在低浓度无机碳条件下, 莱茵衣藻可能通过增大环式电子传递速率满足对ATP/NADPH增加的需求(Lucker and Kramer, 2013).该方法的缺点是样品经过DCMU等抑制剂处理后, 会影响细胞内的氧化还原状态或者代谢状态, 这些因素可能会激活CEF的调控机制, 进而影响CEF检测的准确性(Lucker and Kramer, 2013).在不使用PSII抑制剂的条件下, 可利用gH+与线式电子传递速率的比值估算环式电子的相对传递速率, 其中线式电子传递速率可通过叶绿素荧光检测PSII的光化学量子产量, 该量子产量乘以光强得到.该方法的优点是不会破坏样品正常的生理状态(Sacksteder and Kramer, 2000; Joliot and Joliot, 2002; Baker et al., 2007; Lucker and Kramer, 2013). ...
On the ion transport system of photosynthesis: investigations on a molecular level 1 1968
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
Continuous ECS-indicated recording of the proton-motive charge flux in leaves 1 2013
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
Dynamic flexibility in the light reactions of photosynthesis governed by both electron and proton transfer reactions 1 2004
... 当植物受到环境胁迫时, 环式电子传递的主要作用可能是通过增加ATP的供给保障光合作用高效运行(Kramer et al., 2004; Nandha et al., 2007).例如, 当烟草受到热胁迫且CO2同化受阻时, NDH (NAD (P)H dehydrogenase)介导的环式电子传递会被快速激活, 从而启动环式光合磷酸化产生ATP (Wang et al., 2006).由于ATP与NADPH的产生和消耗紧密相关, 因此围绕PSI的环式电子传递可平衡ATP与NADPH的比值(Kramer and Evans, 2011).例如, 在低浓度无机碳条件下, 莱茵衣藻可能通过增大环式电子传递速率满足对ATP/NADPH增加的需求(Lucker and Kramer, 2013).该方法的缺点是样品经过DCMU等抑制剂处理后, 会影响细胞内的氧化还原状态或者代谢状态, 这些因素可能会激活CEF的调控机制, 进而影响CEF检测的准确性(Lucker and Kramer, 2013).在不使用PSII抑制剂的条件下, 可利用gH+与线式电子传递速率的比值估算环式电子的相对传递速率, 其中线式电子传递速率可通过叶绿素荧光检测PSII的光化学量子产量, 该量子产量乘以光强得到.该方法的优点是不会破坏样品正常的生理状态(Sacksteder and Kramer, 2000; Joliot and Joliot, 2002; Baker et al., 2007; Lucker and Kramer, 2013). ...
Balancing the central roles of the thylakoid proton gradient 1 2003
... 在光合作用中, 放氧光合生物吸收太阳能并裂解水分子, 在类囊体囊腔释放大量的H+并驱动电子在膜蛋白复合物和电子载体间传递, 最终生成还原型烟酰胺腺嘌呤二核苷酸磷酸(辅酶II) (NADPH).同时, 光合电子的传递导致质子从叶绿体基质向类囊体囊腔传递并伴随多种离子的跨膜转运, 随后卡尔文循环消耗质子进行固碳, 又导致叶绿体基质碱化, 这些因素共同诱导形成跨膜电势(ΔΨ)和质子梯度(ΔpH), 即质子动力势(proton motive force, pmf) (Cruz et al., 2001; Carraretto et al., 2016).质子动力势驱动ATP合酶产生腺苷三磷酸(ATP), ATP和NADPH为光合作用中卡尔文循环及其它代谢途径提供能量和还原力(Kramer et al., 2003; Takizawa et al., 2007). ...
The importance of energy balance in improving photosynthetic productivity 1 2011
... 当植物受到环境胁迫时, 环式电子传递的主要作用可能是通过增加ATP的供给保障光合作用高效运行(Kramer et al., 2004; Nandha et al., 2007).例如, 当烟草受到热胁迫且CO2同化受阻时, NDH (NAD (P)H dehydrogenase)介导的环式电子传递会被快速激活, 从而启动环式光合磷酸化产生ATP (Wang et al., 2006).由于ATP与NADPH的产生和消耗紧密相关, 因此围绕PSI的环式电子传递可平衡ATP与NADPH的比值(Kramer and Evans, 2011).例如, 在低浓度无机碳条件下, 莱茵衣藻可能通过增大环式电子传递速率满足对ATP/NADPH增加的需求(Lucker and Kramer, 2013).该方法的缺点是样品经过DCMU等抑制剂处理后, 会影响细胞内的氧化还原状态或者代谢状态, 这些因素可能会激活CEF的调控机制, 进而影响CEF检测的准确性(Lucker and Kramer, 2013).在不使用PSII抑制剂的条件下, 可利用gH+与线式电子传递速率的比值估算环式电子的相对传递速率, 其中线式电子传递速率可通过叶绿素荧光检测PSII的光化学量子产量, 该量子产量乘以光强得到.该方法的优点是不会破坏样品正常的生理状态(Sacksteder and Kramer, 2000; Joliot and Joliot, 2002; Baker et al., 2007; Lucker and Kramer, 2013). ...
Quantum yield and rate of formation of the carotenoid triplet state in photosynthetic structures 1 1980
... 当植物受到环境胁迫时, 环式电子传递的主要作用可能是通过增加ATP的供给保障光合作用高效运行(Kramer et al., 2004; Nandha et al., 2007).例如, 当烟草受到热胁迫且CO2同化受阻时, NDH (NAD (P)H dehydrogenase)介导的环式电子传递会被快速激活, 从而启动环式光合磷酸化产生ATP (Wang et al., 2006).由于ATP与NADPH的产生和消耗紧密相关, 因此围绕PSI的环式电子传递可平衡ATP与NADPH的比值(Kramer and Evans, 2011).例如, 在低浓度无机碳条件下, 莱茵衣藻可能通过增大环式电子传递速率满足对ATP/NADPH增加的需求(Lucker and Kramer, 2013).该方法的缺点是样品经过DCMU等抑制剂处理后, 会影响细胞内的氧化还原状态或者代谢状态, 这些因素可能会激活CEF的调控机制, 进而影响CEF检测的准确性(Lucker and Kramer, 2013).在不使用PSII抑制剂的条件下, 可利用gH+与线式电子传递速率的比值估算环式电子的相对传递速率, 其中线式电子传递速率可通过叶绿素荧光检测PSII的光化学量子产量, 该量子产量乘以光强得到.该方法的优点是不会破坏样品正常的生理状态(Sacksteder and Kramer, 2000; Joliot and Joliot, 2002; Baker et al., 2007; Lucker and Kramer, 2013). ...
... ).该方法的缺点是样品经过DCMU等抑制剂处理后, 会影响细胞内的氧化还原状态或者代谢状态, 这些因素可能会激活CEF的调控机制, 进而影响CEF检测的准确性(Lucker and Kramer, 2013).在不使用PSII抑制剂的条件下, 可利用gH+与线式电子传递速率的比值估算环式电子的相对传递速率, 其中线式电子传递速率可通过叶绿素荧光检测PSII的光化学量子产量, 该量子产量乘以光强得到.该方法的优点是不会破坏样品正常的生理状态(Sacksteder and Kramer, 2000; Joliot and Joliot, 2002; Baker et al., 2007; Lucker and Kramer, 2013). ...
... ; Lucker and Kramer, 2013). ...
The role of PGR5 in the redox poising of photosynthetic electron transport 1 2007
... 当植物受到环境胁迫时, 环式电子传递的主要作用可能是通过增加ATP的供给保障光合作用高效运行(Kramer et al., 2004; Nandha et al., 2007).例如, 当烟草受到热胁迫且CO2同化受阻时, NDH (NAD (P)H dehydrogenase)介导的环式电子传递会被快速激活, 从而启动环式光合磷酸化产生ATP (Wang et al., 2006).由于ATP与NADPH的产生和消耗紧密相关, 因此围绕PSI的环式电子传递可平衡ATP与NADPH的比值(Kramer and Evans, 2011).例如, 在低浓度无机碳条件下, 莱茵衣藻可能通过增大环式电子传递速率满足对ATP/NADPH增加的需求(Lucker and Kramer, 2013).该方法的缺点是样品经过DCMU等抑制剂处理后, 会影响细胞内的氧化还原状态或者代谢状态, 这些因素可能会激活CEF的调控机制, 进而影响CEF检测的准确性(Lucker and Kramer, 2013).在不使用PSII抑制剂的条件下, 可利用gH+与线式电子传递速率的比值估算环式电子的相对传递速率, 其中线式电子传递速率可通过叶绿素荧光检测PSII的光化学量子产量, 该量子产量乘以光强得到.该方法的优点是不会破坏样品正常的生理状态(Sacksteder and Kramer, 2000; Joliot and Joliot, 2002; Baker et al., 2007; Lucker and Kramer, 2013). ...
State transitions redistribute rather than dissipate energy between the two photosystems in Chlamydomonas 1 2016
... 值得注意的是, 饱和闪光可能会诱导PSI反应中心发生2次电荷分离, 从而导致电荷分离的反应中心数量被高估(Bailleul et al., 2010).在植物光适应的分子机理研究方面, PSI和PSII反应中心的比例关系为其研究提供了重要的数据支持, 如捕光叶绿素蛋白生物合成缺陷的莱茵衣藻BF4和p71突变体, 其PSI和PSII的天线大小均有不同程度的减小, 而BF4突变体由于PSII的生物发生受阻, 导致PSI/PSII反应中心比例增加至1.25 (Bujaldon et al., 2020).虽然PSI和PSII的天线大小未发生改变, 但莱茵衣藻还可通过降低PSI/PSII反应中心的比值适应强光变化(Bonente et al., 2012).缺失质体蓝素(PC)的莱茵衣藻突变体在状态转换过程中可达到完全的状态1, 其PSI/PSII比值比野生型下降10% (Nawrocki et al., 2016). ...
Arabidopsis mutants define a central role for the xanthophyll cycle in the regulation of photosynthetic energy conversion 1 1998
... P515分钟级的慢速动力学比较复杂, 既反映质子动力势的生成, 也反映玉米黄素的生成.质子动力势(proton motive force, pmf)驱动ATP合酶产生ATP, 而pmf的组分——ΔpH会诱导光保护机制, 将过剩的光能以热能的形式耗散, 保护PSII免受光损伤(Niyogi et al., 1998; de Bianchi et al., 2010; Checchetto et al., 2013, 2016), 还可通过降低电子在Cytb6f复合物的传递速率, 保护PSI免受光抑制(Sonoike, 2011; Yamori et al., 2011).跨膜电势(ΔΨ)也可调节PSII的光损伤.研究发现, 高水平的ΔΨ不仅降低NPQ的诱导效率(Duan et al., 2016)、增加PSII的光敏感性(Davis et al., 2016), 还导致PSII反应中心发生电荷重组, 促使单线态氧水平升高, 造成严重的光损伤(Bennoun, 1994).我们的研究结果显示, 拟南芥在100 μmol·m-2·s-1的光强下, pmf为1.3, ΔpH约占56%, 而当光强达到750 μmol·m-2·s-1时, pmf增至7.2, ΔpH所占比例增至68%, ΔΨ减少至32% (图4), 说明ΔpH可能在光保护中起更关键的作用. ...
Ion channel permeable for divalent and monovalent cations in native spinach thylakoid membranes 1 1996
... 将暗适应3小时(为了降低玉米黄素的含量)的拟南芥离体叶片放入双通道调制叶绿素荧光仪叶室内, 打开测量光, 待基线平稳后, 打开活化光(635 nm, 750 μmol·m-2·s-1), PSII和PSI反应中心发生原初电荷分离, 从而导致P515信号迅速增加.在持续照光(3 分钟)过程中, 类囊体腔内不断积累的质子逐渐被流入的阴离子(主要是Cl-)中和, P515信号达到相对稳态水平(Cruz et al., 2001; Schreiber and Klughammer, 2008).活化光关闭瞬间, 类囊体腔内积累的质子以不变的速率通过ATP合酶快速释放到类囊体基质, 造成囊腔内阴离子过剩, 从而引起信号反转, 表现为P515信号迅速下降, 下降的最大幅度即质子动力势(pmf).活化光关闭后, 随着类囊体腔内阴离子(主要是Cl-)的外流, 基质中阳离子(主要是K+和Mg2+)内流, 囊腔内外离子浓度逐渐达到平衡(Hind et al.,1974; Schönknecht et al., 1988; Pottosin and Schönknecht, 1996), 反转的P515信号又慢慢恢复, 表现为信号缓慢上升并逐渐达到“暗基线”(Cruz et al., 2001; Schreiber and Klughammer, 2008).pmf及其组分Δψ和ΔpH的计算方法依据文献(Schreiber and Klughammer, 2008). ...
ATP synthase repression in tobacco restricts photosynthetic electron transport, CO2 assimilation, and plant growth by overacidification of the thylakoid lumen 2 2011
... 当植物处于稳态的光合作用时, 通过光合电子传递引起的质子流入类囊体囊腔的速率与通过ATP合酶的质子流出速率相等(Sacksteder and Kramer, 2000).DIRK (dark internal relaxation kinetics)技术是利用短暂的黑暗(<500 ms)干扰处于稳态光合的电子和质子流, 检测样品在520 nm处的光吸收(Sacksteder et al., 2000).该方法可估算叶绿体ATP合酶的活性(Cruz et al., 2001).其原理是当ATP合酶活性降低时, 通过ATP合酶的质子流出速率也会相应降低.拟南芥野生型在500 μmol·m-2·s-1的光照强度下, 叶绿体ATP合酶的质子传导率(gH+)为94.3·s-1 (图2).拟南芥bfa1和bfa3是两个ATP合酶突变体, 在不同光照强度下, 其gH+分别下降至野生型的30%和50%, 同时均表现出较高的NPQ (Zhang et al., 2016, 2018).这是因为当ATP合酶活性降低时, 通过ATP合酶的质子流出效率降低, 导致质子在类囊体囊腔累积, 囊腔过度酸化会抑制NPQ衰减, 从而表现出高水平的NPQ.烟草(Nicotiana tabacum) GTG-atpB突变体的gH+降至野生型的25%, 且突变体的碳同化能力、线式电子传递速率和P700+再还原能力也显著降低(Rott et al., 2011).P515与叶绿素荧光、P700氧化还原动力学的结合使用在烟草、玉米(Zea mays)、拟南芥和豌豆(Pisum sativum)等叶绿体ATP合酶突变体的表型筛选和功能研究方面发挥了重要作用(Rott et al., 2011; Zoschke et al., 2012; Fristedt et al., 2015; Sukhov et al., 2016; Zhang et al., 2016). ...
... )等叶绿体ATP合酶突变体的表型筛选和功能研究方面发挥了重要作用(Rott et al., 2011; Zoschke et al., 2012; Fristedt et al., 2015; Sukhov et al., 2016; Zhang et al., 2016). ...
Activation of zeaxanthin is an obligatory event in the regulation of photosynthetic light harvesting 1 2002
... 传导性, 从而导致缓慢下降阶段显著加快, 因此通过缓慢下降阶段的衰减速率可判断类囊体膜的通透性(Schreiber and Klughammer, 2008). ...
Photoinhibition of photosystem I 1 2011
... P515分钟级的慢速动力学比较复杂, 既反映质子动力势的生成, 也反映玉米黄素的生成.质子动力势(proton motive force, pmf)驱动ATP合酶产生ATP, 而pmf的组分——ΔpH会诱导光保护机制, 将过剩的光能以热能的形式耗散, 保护PSII免受光损伤(Niyogi et al., 1998; de Bianchi et al., 2010; Checchetto et al., 2013, 2016), 还可通过降低电子在Cytb6f复合物的传递速率, 保护PSI免受光抑制(Sonoike, 2011; Yamori et al., 2011).跨膜电势(ΔΨ)也可调节PSII的光损伤.研究发现, 高水平的ΔΨ不仅降低NPQ的诱导效率(Duan et al., 2016)、增加PSII的光敏感性(Davis et al., 2016), 还导致PSII反应中心发生电荷重组, 促使单线态氧水平升高, 造成严重的光损伤(Bennoun, 1994).我们的研究结果显示, 拟南芥在100 μmol·m-2·s-1的光强下, pmf为1.3, ΔpH约占56%, 而当光强达到750 μmol·m-2·s-1时, pmf增至7.2, ΔpH所占比例增至68%, ΔΨ减少至32% (图4), 说明ΔpH可能在光保护中起更关键的作用. ...
An update on the regulation of photosynthesis by thylakoid ion channels and transporters in Arabidopsis 1 2017
... 虽然ΔpH和ΔΨ在质子动力势中的分配机制还不清楚, 但是多种离子通道或转运蛋白参与调节两个组分的分配, 进而影响光合效率(Checchetto et al., 2013, 2016; Davis et al., 2017; Spetea et al., 2017).例如, 双孔K+通道蛋白TPK3缺失的拟南芥突变体, 由于K+外排受阻导致ΔpH的形成受损, 同时NPQ诱导效率降低(Carraretto et al., 2013); Cl-通道蛋白AtBest缺失会导致拟南芥pmf和ΔΨ水平显著升高, 说明Cl-向囊腔内的转运可消散ΔΨ, 从而调控pmf的分配(Duan et al., 2016). ...
Changes in H+-ATP synthase activity, proton electrochemical gradient, and pH inPea chloroplast can be connected with variation potential 2 2016
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
... 当植物处于稳态的光合作用时, 通过光合电子传递引起的质子流入类囊体囊腔的速率与通过ATP合酶的质子流出速率相等(Sacksteder and Kramer, 2000).DIRK (dark internal relaxation kinetics)技术是利用短暂的黑暗(<500 ms)干扰处于稳态光合的电子和质子流, 检测样品在520 nm处的光吸收(Sacksteder et al., 2000).该方法可估算叶绿体ATP合酶的活性(Cruz et al., 2001).其原理是当ATP合酶活性降低时, 通过ATP合酶的质子流出速率也会相应降低.拟南芥野生型在500 μmol·m-2·s-1的光照强度下, 叶绿体ATP合酶的质子传导率(gH+)为94.3·s-1 (图2).拟南芥bfa1和bfa3是两个ATP合酶突变体, 在不同光照强度下, 其gH+分别下降至野生型的30%和50%, 同时均表现出较高的NPQ (Zhang et al., 2016, 2018).这是因为当ATP合酶活性降低时, 通过ATP合酶的质子流出效率降低, 导致质子在类囊体囊腔累积, 囊腔过度酸化会抑制NPQ衰减, 从而表现出高水平的NPQ.烟草(Nicotiana tabacum) GTG-atpB突变体的gH+降至野生型的25%, 且突变体的碳同化能力、线式电子传递速率和P700+再还原能力也显著降低(Rott et al., 2011).P515与叶绿素荧光、P700氧化还原动力学的结合使用在烟草、玉米(Zea mays)、拟南芥和豌豆(Pisum sativum)等叶绿体ATP合酶突变体的表型筛选和功能研究方面发挥了重要作用(Rott et al., 2011; Zoschke et al., 2012; Fristedt et al., 2015; Sukhov et al., 2016; Zhang et al., 2016). ...
Cyclic electron flow is redox-controlled but independent of state transition 1 2013
... 在高等植物和真核藻类中存在线式(LEF)和环式(CEF) 2种电子传递途径, 环式电子传递对光合作用的功能调控已成为研究热点(Joliot and Joliot, 2002; Wang et al., 2006; Lucker and Kramer, 2013; Takahashi et al., 2013; Alric, 2014).目前检测CEF的常用方法是将PSII光合活性抑制后(加入DCMU和HA), 通过P700+的再还原速率计算环式电子传递速率.上文提到利用DIRK技术可测量通过ATP合酶的质子流出速率, 而该速率等于光驱动的通过PSI和PSII的总电子传递速率(Joliot and Joliot, 2002).因此, 将PSII光合活性抑制后, 通过ATP合酶的质子外流速率等于围绕PSI的环式电子传递速率.已有研究表明, 将莱茵衣藻PSII光合活性抑制后, P515与P700氧化还原动力学检测到的围绕PSI的环式电子传递速率呈线性关系(Lucker and Kramer, 2013; Alric, 2014). ...
The thylakoid proton motive force in vivo. Quantitative, non-invasive probes, energetics, and regulatory consequences of light-induced pmf 1 2007
... 在光合作用中, 放氧光合生物吸收太阳能并裂解水分子, 在类囊体囊腔释放大量的H+并驱动电子在膜蛋白复合物和电子载体间传递, 最终生成还原型烟酰胺腺嘌呤二核苷酸磷酸(辅酶II) (NADPH).同时, 光合电子的传递导致质子从叶绿体基质向类囊体囊腔传递并伴随多种离子的跨膜转运, 随后卡尔文循环消耗质子进行固碳, 又导致叶绿体基质碱化, 这些因素共同诱导形成跨膜电势(ΔΨ)和质子梯度(ΔpH), 即质子动力势(proton motive force, pmf) (Cruz et al., 2001; Carraretto et al., 2016).质子动力势驱动ATP合酶产生腺苷三磷酸(ATP), ATP和NADPH为光合作用中卡尔文循环及其它代谢途径提供能量和还原力(Kramer et al., 2003; Takizawa et al., 2007). ...
Control of hydrogen photoproduction by the proton gradient generated by cyclic electron flow in Chlamydomonas reinhardtii 1 2011
Chloroplastic NAD(P)H dehydrogenase in tobacco leaves functions in alleviation of oxidative damage caused by temperature stress 2 2006
... 在高等植物和真核藻类中存在线式(LEF)和环式(CEF) 2种电子传递途径, 环式电子传递对光合作用的功能调控已成为研究热点(Joliot and Joliot, 2002; Wang et al., 2006; Lucker and Kramer, 2013; Takahashi et al., 2013; Alric, 2014).目前检测CEF的常用方法是将PSII光合活性抑制后(加入DCMU和HA), 通过P700+的再还原速率计算环式电子传递速率.上文提到利用DIRK技术可测量通过ATP合酶的质子流出速率, 而该速率等于光驱动的通过PSI和PSII的总电子传递速率(Joliot and Joliot, 2002).因此, 将PSII光合活性抑制后, 通过ATP合酶的质子外流速率等于围绕PSI的环式电子传递速率.已有研究表明, 将莱茵衣藻PSII光合活性抑制后, P515与P700氧化还原动力学检测到的围绕PSI的环式电子传递速率呈线性关系(Lucker and Kramer, 2013; Alric, 2014). ...
... 当植物受到环境胁迫时, 环式电子传递的主要作用可能是通过增加ATP的供给保障光合作用高效运行(Kramer et al., 2004; Nandha et al., 2007).例如, 当烟草受到热胁迫且CO2同化受阻时, NDH (NAD (P)H dehydrogenase)介导的环式电子传递会被快速激活, 从而启动环式光合磷酸化产生ATP (Wang et al., 2006).由于ATP与NADPH的产生和消耗紧密相关, 因此围绕PSI的环式电子传递可平衡ATP与NADPH的比值(Kramer and Evans, 2011).例如, 在低浓度无机碳条件下, 莱茵衣藻可能通过增大环式电子传递速率满足对ATP/NADPH增加的需求(Lucker and Kramer, 2013).该方法的缺点是样品经过DCMU等抑制剂处理后, 会影响细胞内的氧化还原状态或者代谢状态, 这些因素可能会激活CEF的调控机制, 进而影响CEF检测的准确性(Lucker and Kramer, 2013).在不使用PSII抑制剂的条件下, 可利用gH+与线式电子传递速率的比值估算环式电子的相对传递速率, 其中线式电子传递速率可通过叶绿素荧光检测PSII的光化学量子产量, 该量子产量乘以光强得到.该方法的优点是不会破坏样品正常的生理状态(Sacksteder and Kramer, 2000; Joliot and Joliot, 2002; Baker et al., 2007; Lucker and Kramer, 2013). ...
Energy conversion in the functional membrane of photosynthesis. Analysis by light pulse and electric pulse methods: the central role of the electric field 1 1979
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
The roles of ATP synthase and the cytochrome b 6/f complexes in limiting chloroplast electron transport and determining photosynthetic capacity 1 2011
... P515分钟级的慢速动力学比较复杂, 既反映质子动力势的生成, 也反映玉米黄素的生成.质子动力势(proton motive force, pmf)驱动ATP合酶产生ATP, 而pmf的组分——ΔpH会诱导光保护机制, 将过剩的光能以热能的形式耗散, 保护PSII免受光损伤(Niyogi et al., 1998; de Bianchi et al., 2010; Checchetto et al., 2013, 2016), 还可通过降低电子在Cytb6f复合物的传递速率, 保护PSI免受光抑制(Sonoike, 2011; Yamori et al., 2011).跨膜电势(ΔΨ)也可调节PSII的光损伤.研究发现, 高水平的ΔΨ不仅降低NPQ的诱导效率(Duan et al., 2016)、增加PSII的光敏感性(Davis et al., 2016), 还导致PSII反应中心发生电荷重组, 促使单线态氧水平升高, 造成严重的光损伤(Bennoun, 1994).我们的研究结果显示, 拟南芥在100 μmol·m-2·s-1的光强下, pmf为1.3, ΔpH约占56%, 而当光强达到750 μmol·m-2·s-1时, pmf增至7.2, ΔpH所占比例增至68%, ΔΨ减少至32% (图4), 说明ΔpH可能在光保护中起更关键的作用. ...
Biogenesis factor required for ATP synthase 3 facilitates assembly of the chloroplast ATP synthase complex 2 2016
... 当植物处于稳态的光合作用时, 通过光合电子传递引起的质子流入类囊体囊腔的速率与通过ATP合酶的质子流出速率相等(Sacksteder and Kramer, 2000).DIRK (dark internal relaxation kinetics)技术是利用短暂的黑暗(<500 ms)干扰处于稳态光合的电子和质子流, 检测样品在520 nm处的光吸收(Sacksteder et al., 2000).该方法可估算叶绿体ATP合酶的活性(Cruz et al., 2001).其原理是当ATP合酶活性降低时, 通过ATP合酶的质子流出速率也会相应降低.拟南芥野生型在500 μmol·m-2·s-1的光照强度下, 叶绿体ATP合酶的质子传导率(gH+)为94.3·s-1 (图2).拟南芥bfa1和bfa3是两个ATP合酶突变体, 在不同光照强度下, 其gH+分别下降至野生型的30%和50%, 同时均表现出较高的NPQ (Zhang et al., 2016, 2018).这是因为当ATP合酶活性降低时, 通过ATP合酶的质子流出效率降低, 导致质子在类囊体囊腔累积, 囊腔过度酸化会抑制NPQ衰减, 从而表现出高水平的NPQ.烟草(Nicotiana tabacum) GTG-atpB突变体的gH+降至野生型的25%, 且突变体的碳同化能力、线式电子传递速率和P700+再还原能力也显著降低(Rott et al., 2011).P515与叶绿素荧光、P700氧化还原动力学的结合使用在烟草、玉米(Zea mays)、拟南芥和豌豆(Pisum sativum)等叶绿体ATP合酶突变体的表型筛选和功能研究方面发挥了重要作用(Rott et al., 2011; Zoschke et al., 2012; Fristedt et al., 2015; Sukhov et al., 2016; Zhang et al., 2016). ...
... ; Zhang et al., 2016). ...
Nucleus-encoded protein BFA1 promotes efficient assembly of the chloroplast ATP synthase coupling factor 1 1 2018
... 当植物处于稳态的光合作用时, 通过光合电子传递引起的质子流入类囊体囊腔的速率与通过ATP合酶的质子流出速率相等(Sacksteder and Kramer, 2000).DIRK (dark internal relaxation kinetics)技术是利用短暂的黑暗(<500 ms)干扰处于稳态光合的电子和质子流, 检测样品在520 nm处的光吸收(Sacksteder et al., 2000).该方法可估算叶绿体ATP合酶的活性(Cruz et al., 2001).其原理是当ATP合酶活性降低时, 通过ATP合酶的质子流出速率也会相应降低.拟南芥野生型在500 μmol·m-2·s-1的光照强度下, 叶绿体ATP合酶的质子传导率(gH+)为94.3·s-1 (图2).拟南芥bfa1和bfa3是两个ATP合酶突变体, 在不同光照强度下, 其gH+分别下降至野生型的30%和50%, 同时均表现出较高的NPQ (Zhang et al., 2016, 2018).这是因为当ATP合酶活性降低时, 通过ATP合酶的质子流出效率降低, 导致质子在类囊体囊腔累积, 囊腔过度酸化会抑制NPQ衰减, 从而表现出高水平的NPQ.烟草(Nicotiana tabacum) GTG-atpB突变体的gH+降至野生型的25%, 且突变体的碳同化能力、线式电子传递速率和P700+再还原能力也显著降低(Rott et al., 2011).P515与叶绿素荧光、P700氧化还原动力学的结合使用在烟草、玉米(Zea mays)、拟南芥和豌豆(Pisum sativum)等叶绿体ATP合酶突变体的表型筛选和功能研究方面发挥了重要作用(Rott et al., 2011; Zoschke et al., 2012; Fristedt et al., 2015; Sukhov et al., 2016; Zhang et al., 2016). ...
Moderate heat stress reduces the pH component of the transthylakoid proton motive force in light-adapted, intact tobacco leaves 1 2009
... 在光合作用中, 类囊体膜产生的跨膜电势会改变某些光合色素的吸收光谱(Witt, 1979).这一现象最早由Duysens (1954)发现, 小球藻(Chlorella)在照光条件下的吸收光谱呈双波形状且最大吸收峰在515 nm处, 即P515信号(也称ECS信号, electrochromic shift).Junge和Witt (1968)发现, P515信号与类囊体跨膜电势呈线性关系.后来发现P515信号普遍存在于高等植物和藻类中, 即在光合作用过程中, 类囊体膜上的叶绿素b和类胡萝卜素分子吸收光能后发生能级跃迁, 而电势的产生导致能级跃迁发生改变, 从而使其吸收光谱发生位移(Fork and Amesz, 1967; Schmidt et al., 1971; Holmes et al., 1980; Hirano and Katoh, 1981; Frese et al., 2002; Zhang et al., 2009; Bailleul et al., 2010; Klughammer et al., 2013; Sukhov et al., 2016; Allorent et al., 2018; Viola et al., 2019). ...
The pentatricopeptide repeat-SMR protein ATP4 promotes translation of the chloroplast atpB/E mRNA 1 2012
... 当植物处于稳态的光合作用时, 通过光合电子传递引起的质子流入类囊体囊腔的速率与通过ATP合酶的质子流出速率相等(Sacksteder and Kramer, 2000).DIRK (dark internal relaxation kinetics)技术是利用短暂的黑暗(<500 ms)干扰处于稳态光合的电子和质子流, 检测样品在520 nm处的光吸收(Sacksteder et al., 2000).该方法可估算叶绿体ATP合酶的活性(Cruz et al., 2001).其原理是当ATP合酶活性降低时, 通过ATP合酶的质子流出速率也会相应降低.拟南芥野生型在500 μmol·m-2·s-1的光照强度下, 叶绿体ATP合酶的质子传导率(gH+)为94.3·s-1 (图2).拟南芥bfa1和bfa3是两个ATP合酶突变体, 在不同光照强度下, 其gH+分别下降至野生型的30%和50%, 同时均表现出较高的NPQ (Zhang et al., 2016, 2018).这是因为当ATP合酶活性降低时, 通过ATP合酶的质子流出效率降低, 导致质子在类囊体囊腔累积, 囊腔过度酸化会抑制NPQ衰减, 从而表现出高水平的NPQ.烟草(Nicotiana tabacum) GTG-atpB突变体的gH+降至野生型的25%, 且突变体的碳同化能力、线式电子传递速率和P700+再还原能力也显著降低(Rott et al., 2011).P515与叶绿素荧光、P700氧化还原动力学的结合使用在烟草、玉米(Zea mays)、拟南芥和豌豆(Pisum sativum)等叶绿体ATP合酶突变体的表型筛选和功能研究方面发挥了重要作用(Rott et al., 2011; Zoschke et al., 2012; Fristedt et al., 2015; Sukhov et al., 2016; Zhang et al., 2016). ...
 ,1,2,*, 庞肖杰2
,1,2,*, 庞肖杰2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT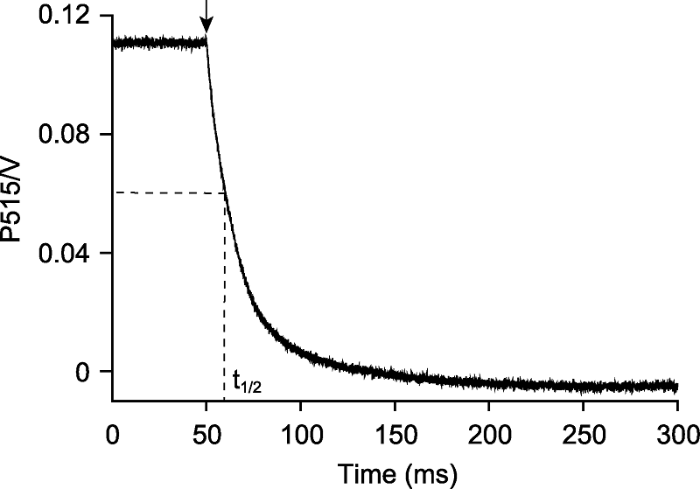 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT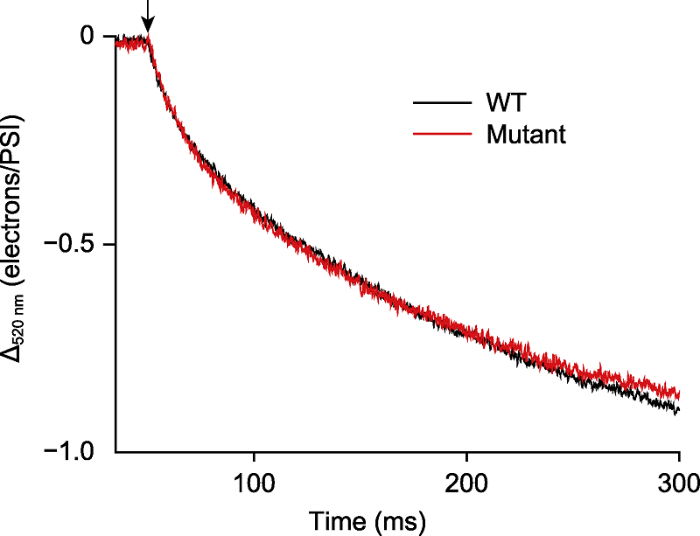 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT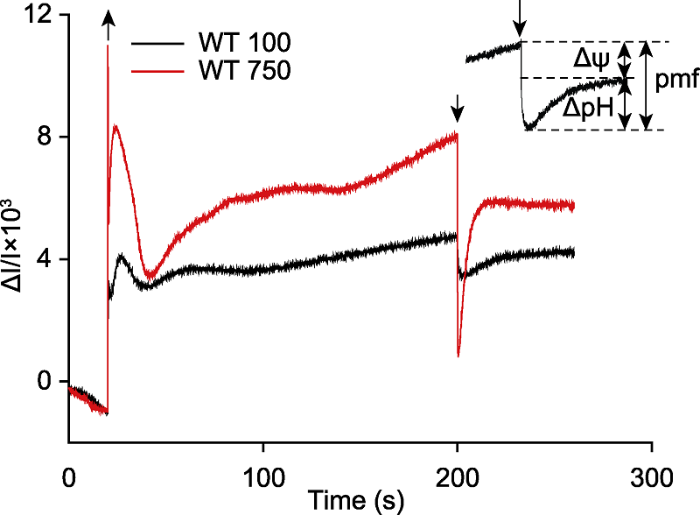 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT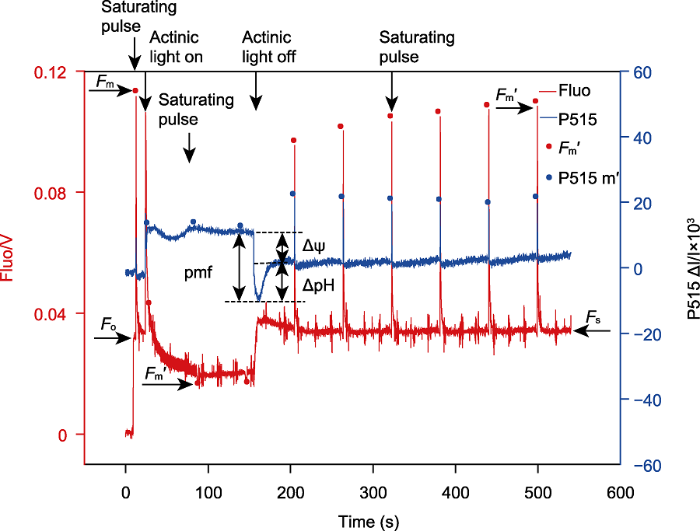 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}