Research Advances in Oxysterol-binding Protein-related Proteins
Wenjiao Zou1,3, Lei Ge2, Qian Yu,2,*1State Key Laboratory of Crop Biology, College of Life Sciences, Shandong Agricultural University, Tai’an 271018, China 2Center for Crop Panomics, College of Agronomy, Qingdao Agriculture University, Qingdao 266109, China 3Innovation Institute of Traditional Chinese Medicine, Shandong University of Traditional Chinese Medicine, Jinan 250355, China
Abstract Membrane lipids not only are important components of cell membranes, but also participate in signal transduction as signal molecules. The uneven distribution of lipid molecules in membranes requires specific types of transport channels and transporters for each lipid type. Oxysterol-binding protein (OSBP)-related proteins (ORPs) are a highly conserved family of lipid transport proteins that recognize and transport phosphoinositides and sterols, which are involved in many physiological processes including signal transduction, vesicle transport, lipid metabolism and non-vesicle transport, and hence play a very important role in the growth and development of individual organism. In recent years, a series of important findings have been made on the structure and function of ORPs in mammals and yeasts, but the advances in plants are relatively slow. In this paper, we review the progress of ORPs research in mammals, yeasts and plants, analyze the structural domains of ORPs in plants and the phylogenetical relationship to their homologs in mammals and yeasts, and also provide perspectives on the directions of plant ORPs research in the future. Keywords:oxysterol-binding protein-related proteins;lipid transport;membrane contact site
PDF (1444KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 邹文娇, 葛磊, 予茜. 氧化甾醇结合蛋白相关蛋白家族的研究进展. 植物学报, 2021, 56(5): 627-640 doi:10.11983/CBB21045 Zou Wenjiao, Ge Lei, Yu Qian. Research Advances in Oxysterol-binding Protein-related Proteins. Chinese Bulletin of Botany, 2021, 56(5): 627-640 doi:10.11983/CBB21045
细胞膜包括细胞质膜, 以及细胞核、线粒体、质体、内质网(endoplasmic reticulum, ER)、高尔基体、液泡等细胞器膜和胞内运输所需要的各种类型囊泡膜所组成的膜系统。脂质是细胞膜的基本组分, 包含脂肪(fats)和类脂(lipids)。类脂又分为磷脂(phospholipids)、糖脂(glycolipids)、脂蛋白(lipoproteins)和固醇类(steroid)。不同物种、不同类型的细胞以及不同细胞器的细胞膜所含脂质的种类、分布和含量都不尽相同。不同的脂质分子在细胞生命活动过程中起着非常重要的作用。脂质不仅是细胞膜的结构成分, 而且参与能量储存以及维持细胞膜的流动性和通透性, 特别是作为信号转导的关键组分参与生物体的个体发育和生理过程。然而, 膜脂信号在胞内的传送需要脂类运输蛋白(lipid transfer proteins, LTPs)的充分配合。正是由于LTPs在胞内信号转导中具有不可或缺的作用, 其生理功能备受研究者的关注, 相关研究在近几年也取得了重要进展(Pietrangelo and Ridgway, 2018; Balla et al., 2019; Wong et al., 2019)。
脂质在细胞膜上的分布不均一(van Meer et al., 2008)。大多数脂质主要在内质网中合成, 随后被运输到各类细胞膜(质膜上的脂类最多), 以维持脂质的特定区域化(Moreau et al., 1998; van Meer et al., 2008)。脂类具有不溶于水的疏水特性。脂质是如何从合成部位(内质网)转移到各种细胞器的? 细胞如何维持固醇分子在不同细胞器膜中的不均匀分布? 这些都是具有科学意义的重要问题。脂类分子的运输对此过程起到关键作用。胞内脂类运输主要包括3种机制: 囊泡运输、非囊泡运输及膜与膜之间的交融扩散。由LTPs参与的非囊泡运输是脂类运输的主要方式。LTPs主要有3方面功能。(1) 作为脂类的运输蛋白, 从一种膜撷取脂类运输到另一种膜。(2) 作为脂类的感受器, 通过与某种蛋白互作来结合或释放某种脂类。(3) 作为脂类的提供者, 如Sec14p (secretion 14 protein), 能够把磷脂酰肌醇运送至磷脂酰肌醇激酶 (Schaaf et al., 2008; Raychaudhuri and Prinz, 2010)。 LTPs是含有许多成员的大家族, 根据其脂质结合域可细分为SEC14、PITP (phosphatidylinositol transfer protein)、START (steroidogenic acute regulatory protein (StAR)-related transfer family)、GLTP (glycolipid transfer protein)、SCP-2 (sterol carrier protein 2)和OSBP/ORPs等家族(D'Angelo et al., 2008)。
FFAT基序的氨基酸序列一般为-EFFDAXE-, 是内质网定位的决定性区域。人类细胞中除了ORP5和ORP8, 其它成员都含有FFAT基序。酵母Osh1-3含有FFAT基序(Raychaudhuri and Prinz, 2010)。FFAT基序能够与VAPs (vesicle-associated membrane protein (VAMP)-associated proteins)蛋白互作, 而VAPs是一类内质网定位的囊泡运输相关蛋白(Skehel et al., 1995), 因此FFAT基序能够指导ORPs定位到内质网上(Loewen et al., 2003)。但FFAT基序介导并不是ORPs定位到内质网上的唯一方式。例如, ORP5和ORP8虽然没有FFAT基序, 但是可通过C端的跨膜区定位到内质网膜(Yan et al., 2008; Du et al., 2011)。Osh4p没有PH蛋白结构域和FFAT基序, 依然在内质网-质膜(plasma membrane) (ER-PM)上定位(Schulz et al., 2009)。目前, 在植物界还未见FFAT基序或相似基序的功能报道。
ANK蛋白结构域是一种特殊的锚蛋白重复结构域。目前, 只发现在人类ORP1和酵母Osh1p-Osh2p中含有ANK蛋白结构域。Osh1p的ANK蛋白结构域通过与核-液泡连接蛋白1 (nuclear-vacuole junction protein 1, NVJ1)互作使其定位在细胞核和液泡之间(Manik et al., 2017)。全长ORP1L (ORP1 large)通过ANK蛋白结构域与GTP结合将Rab7蛋白定位于晚期核内体与溶酶体之间, 可以干扰内膜的转运过程(Johansson et al., 2005; van der Kant et al., 2013)。ANK结构域在Osh2p中的具体功能还不清楚。
OSBP通过FFAT基序与VAP27-A互作, 定位于内质网, 通过PH蛋白结构域定位在高尔基体。OSBP一般通过Coiled-coil结构域形成二聚体行使功能。ORD蛋白结构域在磷脂酰肌醇-4-磷酸(PI4P)逆行转运的驱动下, 将胆固醇从内质网转移到高尔基体。 Figure 1Working model of oxysterol-binding protein (OSBP) function in cholesterol transportation at membrane contact site (MCS) between endoplasmic reticulum (ER) and trans-Golgi (modified from Mesmin et al., 2013; Pietrangelo and Ridgway, 2018)
OSBP is located in ER by FFAT motif interacting with VAP27-A and in trans-Golgi by the PH domain. OSBP usually forms dimers through the Coiled-coil domain to play its role. The ORD domain could move cholesterol from the ER to trans-Golgi, driven by the retrograde transport of phosphatidylinositol-4-phosphate (PI4P).
拟南芥ORPs包括ORP1A、ORP1B、ORP1C、ORP1D、ORP2A、ORP2B、ORP3A、ORP3B、ORP3C、ORP4A、ORP4B和ORP4C; 酵母ORPs包括Osh1-7p; 人类ORPs包括OSBP和ORP1-11。 Figure 3Phylogenetic analysis of ORPs in Arabidopsis thaliana, human and yeast
Arabidopsis ORPs include ORP1A, ORP1B, ORP1C, ORP1D, ORP2A, ORP2B, ORP3A, ORP3B, ORP3C, ORP4A, ORP4B, and ORP4C; Yeast ORPs include Osh1-7p; Human ORPs include OSBP and ORP1-11.
虽然植物中的ORPs基因早已被克隆, 但关于植物ORPs的功能却少有研究。目前仅有研究表明ORP3A可通过与VAP33家族成员PVA12相互作用而定位于内质网(Saravanan et al., 2009)。BIFC实验证明ORP2A与CPK3相互作用, 但在酵母双杂交实验中未检测到二者互作(Berendzen et al., 2012)。PiORP1通过与矮牵牛(Petunia hybrida)花粉管质膜上的PRK1受体激酶相互作用参与花粉的生长与发育(Skirpan et al., 2006)。在高盐胁迫条件下大豆(Glycine max) GmOSBP表达被抑制, 而在衰老叶片中表达被诱导, 说明GmOSBP可能参与应激反应和细胞衰老过程(Li et al., 2008)。与人类或酵母ORPs的研究相比, 植物ORPs影响细胞内脂质转运的机制尚不清楚。
8 研究展望
目前, 大量关于人和酵母ORPs功能的研究证实了其在细胞生命活动中发挥重要作用, 但是依然存在一些基本问题亟待解决。例如, 同一ORPs蛋白如何实现多种定位? PH蛋白结构域实现多种膜定位的机制是什么? 同一ORD蛋白结构域如何识别并转运不同脂类? 相比于动物, 针对植物体中ORPs的研究尚处于起步阶段。近年来, 植物细胞学得到了长足发展, 已发现众多的细胞器定位标记蛋白(Geldner et al., 2009; Zhu et al., 2020)。植物细胞取材方便, 有利于进行ORPs蛋白家族成员功能研究。目前, 我们需要在以下几方面对植物ORPs蛋白的功能进行更深入的探索。
AmarilioR, RamachandranS, SabanayH, LevS (2005). Differential regulation of endoplasmic reticulum structure through VAP-Nir protein interaction J Biol Chem 280, 5934-5944. PMID:15545272 [本文引用: 1] The endoplasmic reticulum (ER) exhibits a characteristic tubular structure that is dynamically rearranged in response to specific physiological demands. However, the mechanisms by which the ER maintains its characteristic structure are largely unknown. Here we show that the integral ER-membrane protein VAP-B causes a striking rearrangement of the ER through interaction with the Nir2 and Nir3 proteins. We provide evidence that Nir (Nir1, Nir2, and Nir3)-VAP-B interactions are mediated through the conserved FFAT (two phenylalanines (FF) in acidic tract) motif present in Nir proteins. However, each interaction affects the structural integrity of the ER differently. Whereas the Nir2-VAP-B interaction induces the formation of stacked ER membrane arrays, the Nir3-VAP-B interaction leads to a gross remodeling of the ER and the bundling of thick microtubules along the altered ER membranes. In contrast, the Nir1-VAP-B interaction has no apparent effect on ER structure. We also show that the Nir2-VAP-B interaction attenuates protein export from the ER. These results demonstrate new mechanisms for the regulation of ER structure, all of which are mediated through interaction with an identical integral ER-membrane protein.
AnnissAM, ApostolopoulosJ, DworkinS, PurtonLE, SparrowRL (2002). An oxysterol-binding protein family identified in the mouse DNA Cell Biol 21, 571-580. DOI:10.1089/104454902320308942URL [本文引用: 2]
BallaT, BondevaT, VarnaiP (2000). How accurately can we image inositol lipids in living cells? Trends Pharmacol Sci 21, 238-241. PMID:10871889 [本文引用: 1]
BallaT, KimYJ, Alvarez-PratsA, PembertonJ (2019). Lipid dynamics at contact sites between the endoplasmic reticulum and other organelles Annu Rev Cell Dev Biol 35, 85-109. DOI:10.1146/cellbio.2019.35.issue-1URL [本文引用: 1]
BehCT, CoolL, PhillipsJ, RineJ (2001). Overlapping functions of the yeast oxysterol-binding protein homologues Genetics 157, 1117-1140. PMID:11238399 [本文引用: 3] The Saccharomyces cerevisiae genome encodes seven homologues of the mammalian oxysterol-binding protein (OSBP), a protein implicated in lipid trafficking and sterol homeostasis. To determine the functions of the yeast OSBP gene family (OSH1-OSH7), we used a combination of genetics, genomics, and sterol lipid analysis to characterize OSH deletion mutants. All 127 combinations and permutations of OSH deletion alleles were constructed. Individual OSH genes were not essential for yeast viability, but the elimination of the entire gene family was lethal. Thus, the family members shared an essential function. In addition, the in vivo depletion of all Osh proteins disrupted sterol homeostasis. Like mutants that affect ergosterol production, the viable combinations of OSH deletion alleles exhibited specific sterol-related defects. Although none of the single OSH deletion mutants was defective for growth, gene expression profiles revealed that each mutant had a characteristic molecular phenotype. Therefore, each gene performed distinct nonessential functions and contributed to a common essential function. Our findings indicated that OSH genes performed a multitude of nonessential roles defined by specific subsets of the genes and that most shared at least one essential role potentially linked to changes in sterol lipid levels.
BehCT, RineJ (2004). A role for yeast oxysterol-binding protein homologs in endocytosis and in the maintenance of intracellular sterol-lipid distribution J Cell Sci 117, 2983-2996. DOI:10.1242/jcs.01157URL [本文引用: 1]
BerendzenKW, BöhmerM, WallmerothN, PeterS, VesićM, ZhouY, TieslerFKE, SchleifenbaumF, HarterK (2012). Screening for in planta protein-protein interactions combining bimolecular fluorescence complementation with flow cytometry Plant Methods 8, 25. PMID:22789293 [本文引用: 1] Understanding protein and gene function requires identifying interaction partners using biochemical, molecular or genetic tools. In plants, searching for novel protein-protein interactions is limited to protein purification assays, heterologous in vivo systems such as the yeast-two-hybrid or mutant screens. Ideally one would be able to search for novel protein partners in living plant cells. We demonstrate that it is possible to screen for novel protein-protein interactions from a random library in protoplasted Arabidopsis plant cells and recover some of the interacting partners. Our screen is based on capturing the bi-molecular complementation of mYFP between an YN-bait fusion partner and a completely random prey YC-cDNA library with FACS. The candidate interactions were confirmed using in planta BiFC assays and in planta FRET-FLIM assays. From this work, we show that the well characterized protein Calcium Dependent Protein Kinase 3 (CPK3) interacts with APX3, HMGB5, ORP2A and a ricin B-related lectin domain containing protein At2g39050. This is one of the first randomin planta screens to be successfully employed.
BouttéY, GrebeM (2009). Cellular processes relying on sterol function in plants Curr Opin Plant Biol 12, 705-713. DOI:10.1016/j.pbi.2009.09.013URL [本文引用: 1]
ChenHJ, AnagnostouG, ChaiA, WithersJ, MorrisA, AdhikareeJ, PennettaG, de BellerocheJS (2010). Characterization of the properties of a novel mutation in VAPB in familial amyotrophic lateral sclerosis J Biol Chem 285, 40266-40281. DOI:10.1074/jbc.M110.161398URL [本文引用: 1]
ChungJ, TortaF, MasaiK, LucastL, CzaplaH, TannerLB, NarayanaswamyP, WenkMR, NakatsuF, De CamilliP (2015). PI4P/phosphatidylserine countertransport at ORP5- and ORP8-mediated ER-plasma membrane contacts Science 349, 428-432. [本文引用: 4]
D'AngeloG, VicinanzaM, De MatteisMA (2008). Lipid- transfer proteins in biosynthetic pathways Curr Opin Cell Biol 20, 360-370. DOI:10.1016/j.ceb.2008.03.013URL [本文引用: 1]
De CraeneJO, BertazziDL, BärS, FriantS (2017). Phosphoinositides, major actors in membrane trafficking and lipid signaling pathways Int J Mol Sci 18, 634. DOI:10.3390/ijms18030634URL [本文引用: 2]
De Saint-JeanM, DelfosseV, DouguetD, ChicanneG, PayrastreB, BourguetW, AntonnyB, DrinG (2011). Osh4p exchanges sterols for phosphatidylinositol 4-phosphate between lipid bilayers J Cell Biol 195, 965-978. DOI:10.1083/jcb.201104062PMID:22162133 [本文引用: 2] Osh/Orp proteins transport sterols between organelles and are involved in phosphoinositide metabolism. The link between these two aspects remains elusive. Using novel assays, we address the influence of membrane composition on the ability of Osh4p/Kes1p to extract, deliver, or transport dehydroergosterol (DHE). Surprisingly, phosphatidylinositol 4-phosphate (PI(4)P) specifically inhibited DHE extraction because PI(4)P was itself efficiently extracted by Osh4p. We solve the structure of the Osh4p-PI(4)P complex and reveal how Osh4p selectively substitutes PI(4)P for sterol. Last, we show that Osh4p quickly exchanges DHE for PI(4)P and, thereby, can transport these two lipids between membranes along opposite routes. These results suggest a model in which Osh4p transports sterol from the ER to late compartments pinpointed by PI(4)P and, in turn, transports PI(4)P backward. Coupled to PI(4)P metabolism, this transport cycle would create sterol gradients. Because the residues that recognize PI(4)P are conserved in Osh4p homologues, other Osh/Orp are potential sterol/phosphoinositol phosphate exchangers.
DeeksMJ, CalcuttJR, IngleEKS, HawkinsTJ, ChapmanS, RichardsonAC, MentlakDA, DixonMR, CartwrightF, SmertenkoAP, OparkaK, HusseyPJ (2012). A superfamily of actin-binding proteins at the actin-membrane nexus of higher plants Curr Biol 22, 1595-1600. DOI:10.1016/j.cub.2012.06.041URL [本文引用: 1]
FriedmanJR, VoeltzGK (2011). The ER in 3D: a multifunctional dynamic membrane network Trends Cell Biol 21, 709-717. DOI:10.1016/j.tcb.2011.07.004URL [本文引用: 1]
GalmesR, HoucineA, van VlietAR, AgostinisP, JacksonCL, GiordanoF (2016). ORP5/ORP8 localize to endoplasmic reticulum-mitochondria contacts and are involved in mitochondrial function EMBO Rep 17, 800-810. DOI:10.15252/embr.201541108URL [本文引用: 1]
GhaiR, DuXM, WangH, DongJQ, FergusonC, BrownAJ, PartonRG, WuJW, YangHY (2017). ORP5 and ORP8 bind phosphatidylinositol-4,5-biphosphate (PtdIns (4,5)P2) and regulate its level at the plasma membrane Nat Commun 8, 757. DOI:10.1038/s41467-017-00861-5URL [本文引用: 4]
GodiA, Di CampliA, KonstantakopoulosA, Di TullioG, AlessiDR, KularGS, DanieleT, MarraP, LucocqJM, De MatteisMA (2004). FAPPs control Golgi-to-cell-surface membrane traffic by binding to ARF and PtdIns(4)P Nat Cell Biol 6, 393-404. DOI:10.1038/ncb1119URL [本文引用: 1]
GuimilS, DunandC (2007). Cell growth and differentiation in Arabidopsis epidermal cells J Exp Bot 58, 3829-3840. PMID:18162628 [本文引用: 1] Plant epidermal cells are morphologically diverse, differing in size, shape, and function. Their unique morphologies reflect the integral function each cell performs in the organ to which it belongs. Cell morphogenesis involves multiple cellular processes acting in concert to create specialized shapes. The Arabidopsis epidermis contains numerous cell types greatly differing in shape, size, and function. Work on three types of epidermal cells, namely trichomes, root hairs, and pavement cells, has made significant progress towards understanding how plant cells reach their final morphology. These three cell types have highly distinct morphologies and each has become a model cell for the study of morphological processes. A growing body of knowledge is creating a picture of how endoreduplication, cytoskeletal dynamics, vesicle transport, and small GTPase signalling, work in concert to create specialized shapes. Similar mechanisms that determine cell shape and polarity are shared between these cell types, while certain mechanisms remain specific to each.
HelleSCJ, KanferG, KolarK, LangA, MichelAH, KornmannB (2013). Organization and function of membrane contact sites Biochim Biophys Acta Mol Cell Res 1833, 2526-2541. DOI:10.1016/j.bbamcr.2013.01.028URL [本文引用: 2]
HongYY, ZhaoJ, GuoL, KimSC, DengXJ, WangGL, ZhangGY, LiMY, WangXM (2016). Plant phospholipases D and C and their diverse functions in stress responses Prog Lipid Res 62, 55-74. DOI:10.1016/j.plipres.2016.01.002URL [本文引用: 1]
HynynenR, SuchanekM, SpandlJ, BäckN, ThieleC, OlkkonenVM (2009). OSBP-related protein 2 is a sterol receptor on lipid droplets that regulates the metabolism of neutral lipids J Lipid Res 50, 1305-1315. DOI:10.1194/jlr.M800661-JLR200PMID:19224871 [本文引用: 1] Oxysterol binding protein-related protein 2 (ORP2) is a member of the oxysterol binding protein family, previously shown to bind 25-hydroxycholesterol and implicated in cellular cholesterol metabolism. We show here that ORP2 also binds 22(R)-hydroxycholesterol [22(R)OHC], 7-ketocholesterol, and cholesterol, with 22(R)OHC being the highest affinity ligand of ORP2 (K(d) 1.4 x 10(-8) M). We report the localization of ORP2 on cytoplasmic lipid droplets (LDs) and its function in neutral lipid metabolism using the human A431 cell line as a model. The ORP2 LD association depends on sterol binding: Treatment with 5 microM 22(R)OHC inhibits the LD association, while a mutant defective in sterol binding is constitutively LD bound. Silencing of ORP2 using RNA interference slows down cellular triglyceride hydrolysis. Furthermore, ORP2 silencing increases the amount of [(14)C]cholesteryl esters but only under conditions in which lipogenesis and LD formation are enhanced by treatment with oleic acid. The results identify ORP2 as a sterol receptor present on LD and provide evidence for its role in the regulation of neutral lipid metabolism, possibly as a factor that integrates the cellular metabolism of triglycerides with that of cholesterol.
IshikawaS, NagaiY, MasudaT, KogaY, NakamuraT, ImamuraY, TakamoriH, HirotaM, FunakosiA, FukushimaM, BabaH (2010). The role of oxysterol binding protein-related protein 5 in pancreatic cancer Cancer Sci 101, 898-905. DOI:10.1111/j.1349-7006.2009.01475.xURL [本文引用: 1]
JansenM, OhsakiY, RegaLR, BittmanR, OlkkonenVM, IkonenE (2011). Role of ORPs in sterol transport from plasma membrane to ER and lipid droplets in mammalian cells Traffic 12, 218-231. DOI:10.1111/j.1600-0854.2010.01142.xURL [本文引用: 1]
JaworskiCJ, MoreiraE, LiAG, LeeR, RodriguezIR (2001). A family of 12 human genes containing oxysterol-binding domains Genomics 78, 185-196. PMID:11735225 [本文引用: 1] Oxysterol-binding proteins (OSBPs) have been described in a wide range of eukaryotes, and are often found to be part of a multi-gene family. We have used bioinformatics and data mining as a starting point for identifying new family members in humans based on the presence of the OSBP signature EQVSHHPP. In addition to OSBP and the recently reported OSBP2, we have found 10 other genes encoding oxysterol-binding domains. Here, we report cDNA and deduced peptide sequences of the previously unknown OSBPs and compare the peptides and genes. All of the genes encode a pleckstrin homology domain, except OSBPL2. However, two of the peptides, OSBPL2 and OSBPL1A, consist of the OSBP domain only. A second OSBPL1 transcript (OSBPL1B) contains 15 additional upstream exons, with a deduced peptide containing a pleckstrin homology domain. Cladistic analysis divides the human OSBP genes into five groups, whose members share similarities in sequence and gene structure; RT-PCR analysis indicates that expression patterns among group members vary widely.
JiaQ, KongDF, LiQH, SunS, SongJL, ZhuYB, LiangKL, KeQM, LinWX, HuangJW (2019). The function of inositol phosphatases in plant tolerance to abiotic stress Int J Mol Sci 20, 3999. DOI:10.3390/ijms20163999URL [本文引用: 1]
JohanssonM, LehtoM, TanhuanpääK, CoverTL, OlkkonenVM (2005). The oxysterol-binding protein homologue ORP1L interacts with Rab7 and alters functional properties of late endocytic compartments Mol Biol Cell 16, 5480-5492. PMID:16176980 [本文引用: 1] ORP1L is a member of the human oxysterol-binding protein (OSBP) family. ORP1L localizes to late endosomes (LEs)/lysosomes, colocalizing with the GTPases Rab7 and Rab9 and lysosome-associated membrane protein-1. We demonstrate that ORP1L interacts physically with Rab7, preferentially with its GTP-bound form, and provide evidence that ORP1L stabilizes GTP-bound Rab7 on LEs/lysosomes. The Rab7-binding determinant is mapped to the ankyrin repeat (ANK) region of ORP1L. The pleckstrin homology domain (PHD) of ORP1L binds phosphoinositides with low affinity and specificity. ORP1L ANK- and ANK+PHD fragments induce perinuclear clustering of LE/lysosomes. This is dependent on an intact microtubule network and a functional dynein/dynactin motor complex. The dominant inhibitory Rab7 mutant T22N reverses the LE clustering, suggesting that the effect is dependent on active Rab7. Transport of fluorescent dextran to LEs is inhibited by overexpression of ORP1L. Overexpression of ORP1L, and in particular the N-terminal fragments of ORP1L, inhibits vacuolation of LE caused by Helicobacter pylori toxin VacA, a process also involving Rab7. The present study demonstrates that ORP1L binds to Rab7, modifies its functional cycle, and can interfere with LE/lysosome organization and endocytic membrane trafficking. This is the first report of a direct connection between the OSBP-related protein family and the Rab GTPases.
JohanssonM, RochaN, ZwartW, JordensI, JanssenL, KuijlC, OlkkonenVM, NeefjesJ (2007). Activation of endosomal dynein motors by stepwise assembly of Rab7- RILP-p150Glued, ORP1L, and the receptorβlll spectrin J Cell Biol 176, 459-471. PMID:17283181 [本文引用: 1] The small GTPase Rab7 controls late endocytic transport by the minus end-directed motor protein complex dynein-dynactin, but how it does this is unclear. Rab7-interacting lysosomal protein (RILP) and oxysterol-binding protein-related protein 1L (ORP1L) are two effectors of Rab7. We show that GTP-bound Rab7 simultaneously binds RILP and ORP1L to form a RILP-Rab7-ORP1L complex. RILP interacts directly with the C-terminal 25-kD region of the dynactin projecting arm p150(Glued), which is required for dynein motor recruitment to late endocytic compartments (LEs). Still, p150(Glued) recruitment by Rab7-RILP does not suffice to induce dynein-driven minus-end transport of LEs. ORP1L, as well as betaIII spectrin, which is the general receptor for dynactin on vesicles, are essential for dynein motor activity. Our results illustrate that the assembly of microtubule motors on endosomes involves a cascade of linked events. First, Rab7 recruits two effectors, RILP and ORP1L, to form a tripartite complex. Next, RILP directly binds to the p150(Glued) dynactin subunit to recruit the dynein motor. Finally, the specific dynein motor receptor Rab7-RILP is transferred by ORP1L to betaIII spectrin. Dynein will initiate translocation of late endosomes to microtubule minus ends only after interacting with betaIII spectrin, which requires the activities of Rab7-RILP and ORP1L.
KabashiE, El OussiniH, BercierV, Gros-LouisF, ValdmanisPN, McDearmidJ, MejierIA, DionPA, DupreN, HollingerD, SinnigerJ, Dirrig-GroschS, CamuW, MeiningerV, LoefflerJP, RenéF, DrapeauP, RouleauGA, DupuisL (2013). Investigating the contribution of VAPB/ALS8 loss of function in amyotrophic lateral sclerosis Hum Mol Genet 22, 2350-2360. DOI:10.1093/hmg/ddt080PMID:23446633 [本文引用: 1] The mutations P56S and T46I in the gene encoding vesicle-associated membrane protein-associated protein B/C (VAPB) cause ALS8, a familial form of amyotrophic lateral sclerosis (ALS). Overexpression of mutant forms of VAPB leads to cytosolic aggregates, suggesting a gain of function of the mutant protein. However, recent work suggested that the loss of VAPB function could be the major mechanism leading to ALS8. Here, we used multiple genetic and experimental approaches to study whether VAPB loss of function might be sufficient to trigger motor neuron degeneration. In order to identify additional ALS-associated VAPB mutations, we screened the entire VAPB gene in a cohort of ALS patients and detected two mutations (A145V and S160Δ). To directly address the contribution of VAPB loss of function in ALS, we generated zebrafish and mouse models with either a decreased or a complete loss of Vapb expression. Vapb knockdown in zebrafish led to swimming deficits. Mice knocked-out for Vapb showed mild motor deficits after 18 months of age yet had innervated neuromuscular junctions (NMJs). Importantly, overexpression of VAPB mutations were unable to rescue the motor deficit caused by Vapb knockdown in zebrafish and failed to cause a toxic gain-of-function defect on their own. Thus, Vapb loss of function weakens the motor system of vertebrate animal models but is on its own unable to lead to a complete ALS phenotype. Our findings are consistent with the notion that VAPB mutations constitute a risk factor for motor neuron disease through a loss of VAPB function.
KagiwadaS, HashimotoM (2007). The yeast VAP homolog Scs2p has a phosphoinositide-binding ability that is correlated with its activity Biochem Biophys Res Commun 364, 870-876. DOI:10.1016/j.bbrc.2007.10.079URL [本文引用: 1]
KagiwadaS, ZenR (2003). Role of the yeast VAP homolog, Scs2p, in INO1 expression and phospholipid metabolism J Biochem 133, 515-522. PMID:12761300 [本文引用: 2] In the yeast Saccharomyces cerevisiae, the expression of phospholipid biosynthetic genes, including the INO1 gene (encoding inositol-1-phosphate synthase), is coordinately regulated by a cis-acting transcriptional element, UAS(INO) (inositol-sensitive upstream activating sequence). For this paper we studied the effect of SCS2 disruption on INO1 expression. SCS2 encodes a type II membrane protein and its deletion leads to inositol auxotrophy at temperatures above 34 degrees C. We found that the expression of the INO1 gene was reduced in the scs2Delta strain even when the cells were cultured under derepressing conditions for INO1 expression. However, the beta-galactosidase gene fused with the INO1 promoter region was expressed normally in the scs2Delta strain. The phospholipid composition of scs2Delta cells was not dramatically changed compared with wild-type cells at 28 degrees C, but the phosphatidylinositol level was reduced in scs2Delta cells cultured at 34 degrees C. In addition, elevated phosphatidylcholine synthesis through the CDP-choline pathway was observed in the scs2Delta strain, and the disruption of genes involved in the CDP-choline pathway rescued the INO1 expression defect of the scs2Delta strain. These results indicate that Scs2p can contribute to coordinated phospholipid metabolism including INO1 expression by regulating phosphatidylcholine synthesis through the CDP-choline pathway.
KandutschAA, ThompsonEB (1980). Cytosolic proteins that bind oxygenated sterols. Cellular distribution, specificity, and some properties J Biol Chem 255, 10813-10821. PMID:7430156 [本文引用: 1] The differential binding of cholesterol and 25-hydroxycholesterol to cytosolic proteins in various types of cells was investigated. 25-Hydroxycholesterol taken up by six different established cell culture lines and by mouse spleen cells in primary culture was bound to cytosolic components which, during velocity gradient centrifugation, displayed sedimentation coefficients of approximately 5 S and 8 S. In contrast, cholesterol taken up by the cells was concentrated near the bottom (approximately 20 S) of the sucrose density gradient but was distributed throughout. Results with primary cultures of mouse fetal liver differed from thos obtained with other cell cultures in that both sterols appeared principally in a 5 S band. Further characterization of the binding components from intact L cells indicated that binding of 25-hydroxycholesterol to the 8 S fraction was saturable and reversible, whereas binding to the 5 S band was not saturable. The 8 S 25-hydroxycholesterol complex involved a protein with a relatively long half-life. The complex was essentially stable at 0 degrees C but dissociated slowly at 25 degrees C. Sulfhydryl functions were not required for sterol binding, and formation of the complex was not dependent upon cAMP. Competition studies with intact cells and with isolated cytosol indicated that a number of other oxygenated sterols bind to 8 S sites occupated by 25-hydroxycholesterol. Those sterols which are potent suppressors of 3-hydroxy-3-methylglutaryl coenzyme A reductase (EC 1.1.1.34) activity competed for these binding sites, while those which do not suppress the reductase did not compete for them. These studies suggest that the binding of oxygenated sterols to the 8 S cytosolic component may be in some sense specific, while that to the 5 S component is nonspecific. The kinetics of formation and dissociation of the 8 S 25-hydroxy-cholesterol.protein complex and of the suppression of 3-hydroxy-3-methylglutaryl-CoA reductase in the presence of the diol was also consistent with a postulated role for the complex in the regulation of the enzyme.
KawanoM, KumagaiK, NishijimaM, HanadaK (2006). Efficient trafficking of ceramide from the endoplasmic reticulum to the Golgi apparatus requires a VAMP-associated protein-interacting FFAT motif of CERT J Biol Chem 281, 30279-30288. DOI:10.1074/jbc.M605032200URL [本文引用: 2]
KetelaarT, AllwoodEG, HusseyPJ (2007). Actin organization and root hair development are disrupted by ethanol-induced overexpression of Arabidopsis actin interacting protein 1 (AIP1) New Phytol 174, 57-62. DOI:10.1111/j.1469-8137.2007.01979.xPMID:17335497 [本文引用: 1] * Actin organization and dynamics are essential for cell division, growth and cytoplasmic streaming. Here we analyse the effects of the overexpression of Actin Interacting Protein 1 (AIP1) on Arabidopsis development. * Arabidopsis plants were transformed with an ethanol-inducible AIP1 construct and the characteristics of these plants were analysed after induction. * When AIP1 was increased to approx. 90% above wild-type values, root hair development and actin organization in all cell types examined were disrupted. * Our data demonstrate that AIP1 is a key regulator of actin organization and that its regulation is essential for normal plant cell morphogenesis.
LagaceTA, ByersDM, CookHW, RidgwayND (1997). Altered regulation of cholesterol and cholesteryl ester synthesis in Chinese-hamster ovary cells overexpressing the oxysterol-binding protein is dependent on the pleckstrin homology domain Biochem J 326(Pt1), 205-213. DOI:10.1042/bj3260205URL [本文引用: 1]
LehtoM, LaitinenS, ChinettiG, JohanssonM, EhnholmC, StaelsB, IkonenE, OlkkonenVM (2001). The OSBP-related protein family in humans J Lipid Res 42, 1203-1213. PMID:11483621 [本文引用: 1] Oxysterols are oxygenated derivatives of cholesterol that have a number of biological effects and play a key role in the maintenance of the body cholesterol balance. In this study, we describe the cDNA sequences and genomic structures of the recently identified human oxysterol-binding protein (OSBP)-related protein (ORP) family (Laitinen, S. et al. 1999. J. Lipid Res. 40: 2204-2211). The family now includes 12 genes/proteins, which can be divided into six distinct subfamilies. The ORP have two major structural features: a highly conserved OSBP-type sterol-binding domain in the C-terminal half and a pleckstrin homology domain present in the N-terminal region of most family members. Several ORP genes are present in S. cerevisiae, D. melanogaster, and C. elegans, suggesting that the protein family has functions of fundamental importance in the eukaryotic kingdom. Analysis of ORP mRNA levels in unloaded or acetylated LDL-loaded human macrophages revealed that the expression of ORP genes was not significantly affected by the loading, with the exception of ORP6, which was up-regulated 2-fold. The present study summarizes the basic characteristics of the OSBP-related gene/protein family in humans, and provides tools for functional analysis of the encoded proteins.
LemmonMA (2003). Phosphoinositide recognition domains Traffic 4, 201-213. PMID:12694559 [本文引用: 1] Domains or modules known to bind phosphoinositides have increased dramatically in number over the past few years, and are found in proteins involved in intracellular trafficking, cellular signaling, and cytoskeletal remodeling. Analysis of lipid binding by these domains and its structural basis has provided significant insight into the mechanism of membrane recruitment by the different cellular phosphoinositides. Domains that target only the rare (3-phosphorylated) phosphoinositides must bind with very high affinity, and with exquisite specificity. This is achieved solely by headgroup interactions in the case of certain pleckstrin homology (PH) domains [which bind PtdIns(3,4,5)P3 and/or PtdIns(3,4)P2], but requires an additional membrane-insertion and/or oligomerization component in the case of the PtdIns(3)P-targeting phox homology (PX) and FYVE domains. Domains that target PtdIns(4,5)P2, which is more abundant by some 25-fold, do not require the same stringent affinity and specificity characteristics, and tend to be more diverse in structure. The mode of phosphoinositide binding by different domains also appears to reflect their distinct functions. For example, pleckstrin homology domains that serve as simple targeting domains recognize only phosphoinositide headgroups. By contrast, certain other domains, notably the epsin ENTH domain, appear to promote bilayer curvature by inserting into the membrane upon binding.
LessmannE, NgoM, LeitgesM, MinguetS, RidgwayND, HuberM (2007). Oxysterol-binding protein-related protein (ORP) 9 is a PDK-2 substrate and regulates Akt phosphorylation Cell Signal 19, 384-392. PMID:16962287 [本文引用: 1] The oxysterol-binding protein and oxysterol-binding protein-related protein family has been implicated in lipid transport and metabolism, vesicle trafficking and cell signaling. While investigating the phosphorylation of Akt/protein kinase B in stimulated bone marrow-derived mast cells, we observed that a monoclonal antibody directed against phospho-S473 Akt cross-reacted with oxysterol-binding protein-related protein 9 (ORP9). Further analysis revealed that mast cells exclusively express ORP9S, an N-terminal truncated version of full-length ORP9L. A PDK-2 consensus phosphorylation site in ORP9L and OPR9S at S287 (VPEFS(287)Y) was confirmed by site-directed mutagenesis. In contrast to Akt, increased phosphorylation of ORP9S S287 in stimulated mast cells was independent of phosphatidylinositol 3-kinase but sensitive to inhibition of conventional PKC isotypes. PKC-beta dependence was confirmed by lack of ORP9S phosphorylation at S287 in PKC-beta-deficient, but not PKC-alpha-deficient, mast cells. Moreover, co-immunoprecipitation of PKC-beta and ORP9S, and in vitro phosphorylation of ORP9S in this complex, argued for direct phosphorylation of ORP9S by PKC-beta, introducing ORP9S as a novel PKC-beta substrate. Akt was also detected in a PKC-beta/ORP9S immune complex and phosphorylation of Akt on S473 was delayed in PKC-deficient mast cells. In HEK293 cells, RNAi experiments showed that depletion of ORP9L increased Akt S473 phosphorylation 3-fold without affecting T308 phosphorylation in the activation loop. Furthermore, mammalian target of rapamycin was implicated in ORP9L phosphorylation in HEK293 cells. These studies identify ORP9 as a PDK-2 substrate and negative regulator of Akt phosphorylation at the PDK-2 site.
LevS, HalevyDB, PerettiD, DahanN (2008). The VAP protein family: from cellular functions to motor neuron disease Trends Cell Biol 18, 282-290. DOI:10.1016/j.tcb.2008.03.006URL [本文引用: 2]
LevineT (2004). Short-range intracellular trafficking of small molecules across endoplasmic reticulum junctions Trends Cell Biol 14, 483-490. PMID:15350976 [本文引用: 1] Intracellular trafficking is not mediated exclusively by vesicles. Additional, non-vesicular mechanisms transport material, in particular small molecules such as lipids and Ca(2+) ions, from one organelle to another. This transport occurs at narrow cytoplasmic gaps called membrane contact sites (MCSs), at which two organelles come into close apposition. Despite the conservation of these structures throughout evolution, little is known about this transport, largely because of a lack of knowledge of almost all molecular components of MCSs. Recently, this situation has started to change because the structural proteins that bridge an MCS are now known in a single case, and proteins implicated in lipid trafficking have been localized to MCSs. In the light of these advances, I hypothesize that the endoplasmic reticulum has a central role in the trafficking of lipids and ions by forming a network of MCSs with most other intracellular organelles.
LevineTP, MunroS (2001). Dual targeting of Osh1p, a yeast homologue of oxysterol-binding protein, to both the Golgi and the nucleus-vacuole junction Mol Biol Cell 12, 1633-1644. PMID:11408574 [本文引用: 2] Oxysterol binding protein (OSBP) is the only protein known to bind specifically to the group of oxysterols with potent effects on cholesterol homeostasis. Although the function of OSBP is currently unknown, an important role is implicated by the existence of multiple homologues in all eukaryotes so far examined. OSBP and a subset of homologues contain pleckstrin homology (PH) domains. Such domains are responsible for the targeting of a wide range of proteins to the plasma membrane. In contrast, OSBP is a peripheral protein of Golgi membranes, and its PH domain targets to the trans-Golgi network of mammalian cells. In this article, we have characterized Osh1p, Osh2p, and Osh3p, the three homologues of OSBP in Saccharomyces cerevisiae that contain PH domains. Examination of a green fluorescent protein (GFP) fusion to Osh1p revealed a striking dual localization with the protein present on both the late Golgi, and in the recently described nucleus-vacuole (NV) junction. Deletion mapping revealed that the PH domain of Osh1p specified targeting to the late Golgi, and an ankyrin repeat domain targeting to the NV junction, the first such targeting domain identified for this structure. GFP fusions to Osh2p and Osh3p showed intracellular distributions distinct from that of Osh1p, and their PH domains appear to contribute to their differing localizations.
LiuXW, RidgwayND (2014). Characterization of the sterol and phosphatidylinositol 4-phosphate binding properties of Golgi-associated OSBP-related protein 9 (ORP9) PLoS One 9, e108368. DOI:10.1371/journal.pone.0108368URL [本文引用: 1]
LiuY, SuY, WangXM (2013). Phosphatidic acid-mediated signaling Adv Exp Med Biol 991, 159-176. DOI:10.1007/978-94-007-6331-9_9PMID:23775695 [本文引用: 1] Phosphatidic acid (PA) is recognized as an important class of lipid messengers. The cellular PA levels are dynamic; PA is produced and metabolized by several enzymatic reactions, including different phospholipases, lipid kinases, and phosphatases. PA interacts with various proteins and the interactions may modulate enzyme catalytic activities and/or tether proteins to membranes. The PA-protein interactions are impacted by changes in cellular pH and other effectors, such as cations. PA is involved in a wide range of cellular processes, including vesicular trafficking, cytoskeletal organization, secretion, cell proliferation, and survival. Manipulations of different PA production reactions alter cellular and organismal response to a wide range of abiotic and biotic stresses. Further investigations of PA's function and mechanisms of action will advance not only the understanding of cell signaling networks but also may lead to biotechnological and pharmacological applications.
LoewenCJR, RoyA, LevineTP (2003). A conserved ER targeting motif in three families of lipid binding proteins and in Opi1p binds VAP EMBO J 22, 2025-2035. DOI:10.1093/emboj/cdg201URL [本文引用: 3]
LundbækJA, CollingwoodSA, IngólfssonHI, KapoorR, AndersenOS (2010). Lipid bilayer regulation of membrane protein function: gramicidin channels as molecular force probes J R Soc Interface 7, 373-395. DOI:10.1098/rsif.2009.0443URL [本文引用: 1]
LuoJ, YangHY, SongBL (2020). Mechanisms and regulation of cholesterol homeostasis Nat Rev Mol Cell Biol 21, 225-245. DOI:10.1038/s41580-019-0190-7URL [本文引用: 1]
MaZG, LiuZH, HuangX (2010). OSBP- and FAN-mediated sterol requirement for spermatogenesis in Drosophila Development 137, 3775-3784. DOI:10.1242/dev.049312URL [本文引用: 1]
MalinskyJ, OpekarovaM, GrossmannG, TannerW (2013). Membrane microdomains, rafts, and detergent- resistant membranes in plants and fungi Annu Rev Plant Biol 64, 501-529. DOI:10.1146/annurev-arplant-050312-120103PMID:23638827 [本文引用: 1] The existence of specialized microdomains in plasma membranes, postulated for almost 25 years, has been popularized by the concept of lipid or membrane rafts. The idea that detergent-resistant membranes are equivalent to lipid rafts, which was generally abandoned after a decade of vigorous data accumulation, contributed to intense discussions about the validity of the raft concept. The existence of membrane microdomains, meanwhile, has been verified by unequivocal independent evidence. This review summarizes the current state of research in plants and fungi with respect to common aspects of both kingdoms. In these organisms, principally immobile microdomains large enough for microscopic detection have been visualized. These microdomains are found in the context of cell-cell interactions (plant symbionts and pathogens), membrane transport, stress, and polarized growth, and the data corroborate at least three mechanisms of formation. As documented in this review, modern methods of visualization of lateral membrane compartments are also able to uncover the functional relevance of membrane microdomains.
ManikMK, YangH, TongJ, ImYJ (2017). Structure of yeast OSBP-related protein OSH1 reveals key determinants for lipid transport and protein targeting at the nucleus-vacuole junction Structure 25, 617-629. DOI:10.1016/j.str.2017.02.010URL [本文引用: 2]
MaoDX, LinG, TepeB, ZuoZY, TanKL, SenturkM, ZhangS, ArenkielBR, SardielloM, BellenHJ (2019). VAMP associated proteins are required for autophagic and lysosomal degradation by promoting a PtdIns4P-medIated endosomal pathway Autophagy 15, 1214-1233. DOI:10.1080/15548627.2019.1580103URL [本文引用: 2]
MoreauP, HartmannMA, PerretAM, Sturbois-BalcerzakB, CassagneC (1998). Transport of sterols to the plasma membrane of leek seedlings Plant Physiol 117, 931-937. PMID:9662535 [本文引用: 1] To investigate the intracellular transport of sterols in etiolated leek (Allium porrum L.) seedlings, in vivo pulse-chase experiments with [1-14C]acetate were performed. Then, endoplasmic reticulum-, Golgi-, and plasma membrane (PM)-enriched fractions were prepared and analyzed for the radioactivity incorporated into free sterols. In leek seedlings sterols are present as a mixture in which (24R)-24-ethylcholest-5-en-3beta-ol is by far the major compound (around 60%). The other sterols are represented by cholest-5-en-3beta-ol, 24-methyl-cholest-5-en-3beta-ol, (24S)-24-ethylcholesta-5,22E-dien-3beta-ol, and stigmasta-5, 24(24(1))Z-dien-3beta-ol. These compounds are shown to reside mainly in the PM. Our results clearly indicate that free sterols are actively transported from the endoplasmic reticulum to the PM during the first 60 min of chase, with kinetics very similar to that of phosphatidylserine. Such a transport was found to be decreased at low temperature (12 degreesC) and following treatment with monensin and brefeldin A. These data are consistent with a membrane-mediated process for the intracellular transport of sterols to the PM, which likely involves the Golgi apparatus.
MurphySE, LevineTP (2016). VAP, a versatile access point for the endoplasmic reticulum: review and analysis of FFAT-like motifs in the VAPome Biochim Biophys Acta Mol Cell Biol Lipids 1861, 952-961. [本文引用: 1]
NgoM, RidgwayND (2009). Oxysterol binding protein- related protein 9 (ORP9) is a cholesterol transfer protein that regulates Golgi structure and function Mol Biol Cell 20, 1388-1399. DOI:10.1091/mbc.e08-09-0905URL [本文引用: 1]
NishimuraAL, Mitne-NetoM, SilvaHCA, Richieri-CostaA, MiddletonS, CascioD, KokF, OliveiraJRM, GillingwaterT, WebbJ, SkehelP, ZatzM (2004). A mutation in the vesicle-trafficking protein VAPB causes late-onset spinal muscular atrophy and amyotrophic lateral sclerosis Am J Hum Genet 75, 822-831. PMID:15372378 [本文引用: 1] Motor neuron diseases (MNDs) are a group of neurodegenerative disorders with involvement of upper and/or lower motor neurons, such as amyotrophic lateral sclerosis (ALS), spinal muscular atrophy (SMA), progressive bulbar palsy, and primary lateral sclerosis. Recently, we have mapped a new locus for an atypical form of ALS/MND (atypical amyotrophic lateral sclerosis [ALS8]) at 20q13.3 in a large white Brazilian family. Here, we report the finding of a novel missense mutation in the vesicle-associated membrane protein/synaptobrevin-associated membrane protein B (VAPB) gene in patients from this family. Subsequently, the same mutation was identified in patients from six additional kindreds but with different clinical courses, such as ALS8, late-onset SMA, and typical severe ALS with rapid progression. Although it was not possible to link all these families, haplotype analysis suggests a founder effect. Members of the vesicle-associated proteins are intracellular membrane proteins that can associate with microtubules and that have been shown to have a function in membrane transport. These data suggest that clinically variable MNDs may be caused by a dysfunction in intracellular membrane trafficking.
NishimuraY, HayashiM, InadaH, TanakaT (1999). Molecular cloning and characterization of mammalian homologues of vesicle-associated membrane protein-assocIated (VAMP-associated) proteins Biochem Biophys Res Commun 254, 21-26. DOI:10.1006/bbrc.1998.9876URL [本文引用: 2]
PietrangeloA, RidgwayND (2018). Bridging the molecular and biological functions of the oxysterol-binding protein family Cell Mol Life Sci 75, 3079-3098. DOI:10.1007/s00018-018-2795-yPMID:29536114 [本文引用: 3] Oxysterol-binding protein (OSBP) and OSBP-related proteins (ORPs) constitute a large eukaryotic gene family that transports and regulates the metabolism of sterols and phospholipids. The original classification of the family based on oxysterol-binding activity belies the complex dual lipid-binding specificity of the conserved OSBP homology domain (OHD). Additional protein- and membrane-interacting modules mediate the targeting of select OSBP/ORPs to membrane contact sites between organelles, thus positioning the OHD between opposing membranes for lipid transfer and metabolic regulation. This unique subcellular location, coupled with diverse ligand preferences and tissue distribution, has identified OSBP/ORPs as key arbiters of membrane composition and function. Here, we will review how molecular models of OSBP/ORP-mediated intracellular lipid transport and regulation at membrane contact sites relate to their emerging roles in cellular and organismal functions.
RaychaudhuriS, ImYJ, HurleyJH, PrinzWA (2006). Nonvesicular sterol movement from plasma membrane to ER requires oxysterol-binding protein-related proteins and phosphoinositides J Cell Biol 173, 107-119. PMID:16585271 [本文引用: 1] Sterols are moved between cellular membranes by nonvesicular pathways whose functions are poorly understood. In yeast, one such pathway transfers sterols from the plasma membrane (PM) to the endoplasmic reticulum (ER). We show that this transport requires oxysterol-binding protein (OSBP)-related proteins (ORPs), which are a large family of conserved lipid-binding proteins. We demonstrate that a representative member of this family, Osh4p/Kes1p, specifically facilitates the nonvesicular transfer of cholesterol and ergosterol between membranes in vitro. In addition, Osh4p transfers sterols more rapidly between membranes containing phosphoinositides (PIPs), suggesting that PIPs regulate sterol transport by ORPs. We confirmed this by showing that PM to ER sterol transport slows dramatically in mutants with conditional defects in PIP biosynthesis. Our findings argue that ORPs move sterols among cellular compartments and that sterol transport and intracellular distribution are regulated by PIPs.
RaychaudhuriS, PrinzWA (2010). The diverse functions of oxysterol-binding proteins Annu Rev Cell Dev Biol 26, 157-177. DOI:10.1146/annurev.cellbio.042308.113334PMID:19575662 [本文引用: 3] Oxysterol-binding protein (OSBP)-related proteins (ORPs) are lipid-binding proteins that are conserved from yeast to humans. They are implicated in many cellular processes including signaling, vesicular trafficking, lipid metabolism, and nonvesicular sterol transport. All ORPs contain an OSBP-related domain (ORD) that has a hydrophobic pocket that binds a single sterol. ORDs also contain additional membrane-binding surfaces, some of which bind phosphoinositides and may regulate sterol binding. Studies in yeast suggest that ORPs function as sterol transporters, perhaps in regions where organelle membranes are closely apposed. Yeast ORPs also participate in vesicular trafficking, although their role is unclear. In mammalian cells, some ORPs function as sterol sensors that regulate the assembly of protein complexes in response to changes in cholesterol levels. This review will summarize recent advances in our understanding of how ORPs bind lipids and membranes and how they function in diverse cellular processes.
RochaN, KuijlC, van der KantR, JanssenL, HoubenD, JanssenH, ZwartW, NeefjesJ (2009). Cholesterol sensor ORP1L contacts the ER protein VAP to control Rab7-RILP-p150Glued and late endosome positioning J Cell Biol 185, 1209-1225. DOI:10.1083/jcb.200811005PMID:19564404 [本文引用: 1] Late endosomes (LEs) have characteristic intracellular distributions determined by their interactions with various motor proteins. Motor proteins associated to the dynactin subunit p150(Glued) bind to LEs via the Rab7 effector Rab7-interacting lysosomal protein (RILP) in association with the oxysterol-binding protein ORP1L. We found that cholesterol levels in LEs are sensed by ORP1L and are lower in peripheral vesicles. Under low cholesterol conditions, ORP1L conformation induces the formation of endoplasmic reticulum (ER)-LE membrane contact sites. At these sites, the ER protein VAP (VAMP [vesicle-associated membrane protein]-associated ER protein) can interact in trans with the Rab7-RILP complex to remove p150(Glued) and associated motors. LEs then move to the microtubule plus end. Under high cholesterol conditions, as in Niemann-Pick type C disease, this process is prevented, and LEs accumulate at the microtubule minus end as the result of dynein motor activity. These data explain how the ER and cholesterol control the association of LEs with motor proteins and their positioning in cells.
RoyA, LevineTP (2004). Multiple pools of phosphatidylinositol 4-phosphate detected using the pleckstrin homology domain of Osh2p J Biol Chem 279, 44683-44689. DOI:10.1074/jbc.M401583200URL [本文引用: 1]
RustenTE, StenmarkH (2006). Analyzing phosphoinositides and their interacting proteins Nat Methods 3, 251-258. PMID:16554828 [本文引用: 1] Phosphorylated derivatives of phosphatidylinositol (PtdIns), known as phosphoinositides (PIs), are essential regulators of nuclear functions, cytoskeletal dynamics, cell signaling and membrane trafficking. These lipids are found on the cytosolic face of intracellular membranes but can also be detected in membrane-free regions of the nucleoplasm. Their downstream effectors include several proteins that contain various PI-specific domains. Because impaired PI metabolism is associated with disorders such as cancer, cardiovascular disease and immune dysfunction, there is currently great interest in studying PIs and their metabolic enzymes. Here we describe strategies and techniques for quantitative and qualitative measurement of PIs, for characterization of specific PI-binding proteins and for determination of PI kinase and phosphatase activities in vitro and in vivo.
SaravananRS, SlabaughE, SinghVR, LapidusLJ, HaasT, BrandizziF (2009). The targeting of the oxysterol- binding protein ORP3a to the endoplasmic reticulum relies on the plant VAP33 homolog PVA12 Plant J 58, 817-830. DOI:10.1111/tpj.2009.58.issue-5URL [本文引用: 1]
SchaafG, OrtlundEA, TyeryarKR, MousleyCJ, IleKE, GarrettTA, RenJH, WoollsMJ, RaetzCRH, RedinboMR, BankaitisVA (2008). Functional anatomy of phospholipid binding and regulation of phosphoinositide homeostasis by proteins of the sec14 superfamily Mol Cell 29, 191-206. DOI:10.1016/j.molcel.2007.11.026URL [本文引用: 1]
SchulzTA, ChoiMG, RaychaudhuriS, MearsJA, GhirlandoR, HinshawJE, PrinzWA (2009). Lipid-regulated sterol transfer between closely apposed membranes by oxysterol-binding protein homologues J Cell Biol 187, 889-903. DOI:10.1083/jcb.200905007URL [本文引用: 6]
SkehelPA, MartinKC, KandelER, BartschD (1995). A VAMP-binding protein from Aplysia required for neurotransmitter release Science 269, 1580-1583. PMID:7667638 [本文引用: 1] Before the fusion of synaptic vesicles with the plasma membrane, a protein complex is thought to form between VAMP--an integral membrane protein of the vesicle--and two proteins associated with the plasma membrane, SNAP-25 and syntaxin. The yeast two-hybrid interaction cloning system has now been used to identify additional proteins from Aplysia that interact directly with VAMP. A 33-kilodalton membrane protein, termed VAP-33 (VAMP-associated protein of 33 kilodaltons), was identified whose corresponding messenger RNA was detected only in the central nervous system and the gill of Aplysia. Presynaptic injection of antibodies specific for VAP-33 inhibited synaptic transmission, which suggests that VAP-33 is required for the exocytosis of neurotransmitter.
SkirpanAL, DowdPE, SijacicP, JaworskiCJ, GilroyS, KaoTH (2006). Identification and characterization of PiORP1, a Petunia oxysterol-binding-protein related protein involved in receptor-kinase mediated signaling in pollen, and analysis of the ORP gene family in Arabidopsis Plant Mol Biol 61, 553-565. PMID:16897474 [本文引用: 5] Oxysterol-binding proteins (OSBPs) and oxysterol-binding-protein related proteins (ORPs) are encoded by most eukaryotic genomes examined to date; however, they have not yet been characterized in plants. Here we report the identification and characterization of PiORP1, an ORP of Petunia inflata that interacts with the cytoplasmic kinase domain of a receptor-like kinase, named PRK1, of P. inflata. PiORP1 is phosphorylated by PRK1 in vitro and therefore may be involved in PRK1 signaling during pollen development and growth. RNA gel blot analysis showed that PiORP1 and PRK1 had very similar expression patterns in developing pollen, mature pollen and pollen tubes. GFP fusion proteins of PiORP1 localized in the plasma membrane of pollen tubes at distinct foci and its PH domain alone was sufficient to mediate this localization. The sequence for the oxysterol-binding domain of PiORP1 was used to search the genome of Arabidopsis; 12 ORPs were identified and phylogenetic analysis revealed that they fell into two distinct clades, consistent with the ORPs of other eukaryotes. RT-PCR analysis showed that all 12 Arabidopsis ORPs were expressed; 10 were expressed in most of the tissues examined under normal growth conditions, but only three were expressed in pollen.
SoussanL, BurakovD, DanielsMP, Toister-AchituvM, PoratA, YardenY, ElazarZ (1999). ERG30, a VAP-33- related protein, functions in protein transport mediated by COPI vesicles J Cell Biol 146, 301-312. PMID:10427086 [本文引用: 1] Intracellular transport of newly synthesized and mature proteins via vesicles is controlled by a large group of proteins. Here we describe a ubiquitous rat protein-endoplasmic reticulum (ER) and Golgi 30-kD protein (ERG30)-which shares structural characteristics with VAP-33, a 33-kD protein from Aplysia californica which was shown to interact with the synaptic protein VAMP. The transmembrane topology of the 30-kD ERG30 corresponds to a type II integral membrane protein, whose cytoplasmic NH(2) terminus contains a predicted coiled-coil motif. We localized ERG30 to the ER and to pre-Golgi intermediates by biochemical and immunocytochemical methods. Consistent with a role in vesicular transport, anti-ERG30 antibodies specifically inhibit intra-Golgi transport in vitro, leading to significant accumulation of COPI-coated vesicles. It appears that ERG30 functions early in the secretory pathway, probably within the Golgi and between the Golgi and the ER.
StefanoG, RennaL, WormsbaecherC, GambleJ, ZienkiewiczK, BrandizziF (2018). Plant endocytosis requires the ER membrane-anchored proteins VAP27-1 and VAP27-3 Cell Rep 23, 2299-2307. DOI:10.1016/j.celrep.2018.04.091URL [本文引用: 2]
TakáčT, NovákD, ŠamajJ (2019). Recent advances in the cellular and developmental biology of phospholipases in plants Front Plant Sci 10, 362. DOI:10.3389/fpls.2019.00362URL [本文引用: 1]
TakahashiD, KawamuraY, UemuraM (2013). Changes of detergent-resistant plasma membrane proteins in oat and rye during cold acclimation: association with differential freezing tolerance J Proteome Res 12, 4998-5011. DOI:10.1021/pr400750gPMID:24111712 [本文引用: 1] Cold acclimation (CA) results in an increase in freezing tolerance of plants, which is closely associated to functional changes of the plasma membrane (PM). Although proteomic studies have revealed compositional changes of the PM during CA, there has been no large-scale study of how the microdomains in the PM, which contains specific lipids and proteins, change during CA. Therefore, we conducted semiquantitative shotgun proteomics using microdomain-enriched detergent-resistant membrane (DRM) fractions extracted from low freezing-tolerant oat and highly freezing-tolerant rye. We identified 740 and 809 DRM proteins in oat and rye, respectively. Among the proteins identified, the abundances of a variety of proteins, such as P-type ATPase and aquaporins, were affected by CA in both oat and rye. Some CA-responsive proteins in the DRM fractions, such as heat shock protein 70, changed differently in oat and rye. In addition, changes in lipocalins and sugar transporters in the DRM fractions were different from those found in total PM fraction during CA. This is the first report to describe compositional changes in the DRM during CA. The proteomic profiles obtained in the present study hint at many possible microdomain functions associated with CA and freezing tolerance.
TapkenW, MurphyAS (2015). Membrane nanodomains in plants: capturing form, function, and movement J Exp Bot 66, 1573-1586. DOI:10.1093/jxb/erv054URL [本文引用: 1]
TaylorFR, KandutschAA (1985). [2] Use of oxygenated sterols to probe the regulation of 3-hydroxy-3-methylglutaryl-CoA reductase and sterologenesis Methods Enzymol 110, 9-19. [本文引用: 1]
TaylorFR, SaucierSE, ShownEP, ParishEJ, KandutschAA (1984). Correlation between oxysterol binding to a cytosolic binding protein and potency in the repression of hydroxymethylglutaryl coenzyme a reductase J Biol Chem 259, 12382-12387. PMID:6490619 [本文引用: 1] Support for the role of a cytosolic oxysterol-binding protein in the regulation of 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase was obtained by correlating the relative binding affinities of a wide range of oxysterols to their potency in suppressing HMG-CoA reductase activity in mouse fibroblast cell cultures. Forty-seven oxysterols encompassing a 100-fold range of activity in both assays were tested and the two parameters were closely correlated for 35 of the sterols. Twelve sterols showed poor binding when compared to their ability to suppress HMG-CoA reductase activity in cell cultures. Among these were seven sterols with a ketone function at C-3. For this group, the discrepancy could be explained by their rapid conversion within cells to the 3 beta-hydroxy derivatives which have a much higher affinity for the binding protein. One sterol with 3-keto-4-ene grouping was not reduced to its 3 beta-hydroxy derivative in cells and thereby showed no discrepancy in the two assays. The remaining five sterols exhibiting discordant activities in the two tests contained 4,4-dimethyl moieties and were relatively weak suppressors of HMG-CoA reductase activity. Cellular metabolism of these sterols was not detected. Possible reasons for their apparent inactivity in the binding assay are discussed.
TongJS, YangH, YangHY, EomSH, ImYJ (2013). Structure of Osh3 reveals a conserved mode of phosphoinositide binding in oxysterol-binding proteins Structure 21, 1203-1213. DOI:10.1016/j.str.2013.05.007URL [本文引用: 1]
van der KantR, FishA, JanssenL, JanssenH, KromS, HoN, BrummelkampT, CaretteJ, RochaN, NeefjesJ (2013). Late endosomal transport and tethering are coupled processes controlled by RILP and the cholesterol sensor ORP1L J Cell Sci 126, 3462-3474. DOI:10.1242/jcs.129270PMID:23729732 [本文引用: 1] Late endosomes and lysosomes are dynamic organelles that constantly move and fuse to acquire cargo from early endosomes, phagosomes and autophagosome. Defects in lysosomal dynamics cause severe neurodegenerative and developmental diseases, such as Niemann-Pick type C disease and ARC syndrome, yet little is known about the regulation of late endosomal fusion in a mammalian system. Mammalian endosomes destined for fusion need to be transported over very long distances before they tether to initiate contact. Here, we describe that lysosomal tethering and transport are combined processes co-regulated by one multi-protein complex: RAB7-RILP-ORP1L. We show that RILP directly and concomitantly binds the tethering HOPS complex and the p150(Glued) subunit of the dynein motor. ORP1L then functions as a cholesterol-sensing switch controlling RILP-HOPS-p150(Glued) interactions. We show that RILP and ORP1L control Ebola virus infection, a process dependent on late endosomal fusion. By combining recruitment and regulation of both the dynein motor and HOPS complex into a single multiprotein complex, the RAB7-RILP-ORP1L complex efficiently couples and regulates the timing of microtubule minus-end transport and fusion, two major events in endosomal biology.
van MeerG, VoelkerDR, FeigensonGW (2008). Membrane lipids: where they are and how they behave Nat Rev Mol Cell Biol 9, 112-124. DOI:10.1038/nrm2330URL [本文引用: 2]
VihervaaraT, UronenRL, WohlfahrtG, BjörkhemI, IkonenE, OlkkonenVM (2011). Sterol binding by OSBP- related protein 1L regulates late endosome motility and function Cell Mol Life Sci 68, 537-551. DOI:10.1007/s00018-010-0470-zPMID:20690035 [本文引用: 1] ORP1L is an oxysterol binding homologue that regulates late endosome (LE) positioning. We show that ORP1L binds several oxysterols and cholesterol, and characterize a mutant, ORP1L Δ560-563, defective in oxysterol binding. While wild-type ORP1L clusters LE, ORP1L Δ560-563 induces LE scattering, which is reversed by disruption of the endoplasmic reticulum (ER) targeting FFAT motif, suggesting that it is due to enhanced LE-ER interactions. Endosome motility is reduced upon overexpression of ORP1L. Both wild-type ORP1L and the Δ560-563 mutant induce the recruitment of both dynactin and kinesin-2 on LE. Most of the LE decorated by overexpressed ORP1L fail to accept endocytosed dextran or EGF, and the transfected cells display defective degradation of internalized EGF. ORP1L silencing in macrophage foam cells enhances endosome motility and results in inhibition of [(3)H]cholesterol efflux to apolipoprotein A-I. These data demonstrate that LE motility and functions in both protein and lipid transport are regulated by ORP1L.
von FilseckJM, ČopičA, DelfosseV, VanniS, JacksonCL, BourguetW, DrinG (2015a). Phosphatidylserine transport by ORP/Osh proteins is driven by phosphatidylinositol 4-phosphate Science 349, 432-436. [本文引用: 1]
von FilseckJM, VanniS, MesminB, AntonnyB, DrinG (2015b). A phosphatidylinositol-4-phosphate powered exchange mechanism to create a lipid gradient between membranes Nat Commun 6, 6671. DOI:10.1038/ncomms7671URL [本文引用: 2]
WangH, MaQL, QiYF, DongJQ, DuXM, RaeJ, WangJ, WuWF, BrownAJ, PartonRG, WuJW, YangHY (2019a). ORP2 delivers cholesterol to the plasma membrane in exchange for phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2) Mol Cell 73, 458-473. DOI:10.1016/j.molcel.2018.11.014URL [本文引用: 1]
WangPH, DuanW, MunnAL, YangHY (2005a). Molecular characterization of Osh6p, an oxysterol binding protein homolog in the yeast Saccharomyces cerevisiae FEBS J 272, 4703-4715. DOI:10.1111/ejb.2005.272.issue-18URL [本文引用: 1]
WangPW, HawkinsTJ, RichardsonC, CumminsI, DeeksMJ, SparkesI, HawesC, HusseyPJ (2014). The plant cytoskeleton, NET3C, and VAP27 mediate the link between the plasma membrane and endoplasmic reticulum Curr Biol 24, 1397-1405. DOI:10.1016/j.cub.2014.05.003URL [本文引用: 1]
WangPW, RichardsonC, HawkinsTJ, SparkesI, HawesC, HusseyPJ (2016a). Plant VAP27 proteins: domain characterization, intracellular localization and role in plant development New Phytol 210, 1311-1326. DOI:10.1111/nph.2016.210.issue-4URL [本文引用: 3]
WangPY, WengJ, AndersonRGW (2005b). OSBP is a cholesterol-regulated scaffolding protein in control of ERK1/2 activation Science 307, 1472-1476. DOI:10.1126/science.1107710URL [本文引用: 1]
WangWM, LiuPQ, XuYJ, XiaoSY (2016b). Protein trafficking during plant innate immunity J Integr Plant Biol 58, 284-298. DOI:10.1111/jipb.v58.4URL [本文引用: 2]
Weber-BoyvatM, KentalaH, PeränenJ, OlkkonenVM (2015b). Ligand-dependent localization and function of ORP-VAP complexes at membrane contact sites Cell Mol Life Sci 72, 1967-1987. DOI:10.1007/s00018-014-1786-xPMID:25420878 [本文引用: 2] Oxysterol-binding protein/OSBP-related proteins (ORPs) constitute a conserved family of sterol/phospholipid-binding proteins with lipid transporter or sensor functions. We investigated the spatial occurrence and regulation of the interactions of human OSBP/ORPs or the S. cerevisiae orthologs, the Osh (OSBP homolog) proteins, with their endoplasmic reticulum (ER) anchors, the VAMP-associated proteins (VAPs), by employing bimolecular fluorescence complementation and pull-down set-ups. The ORP-VAP interactions localize frequently at distinct subcellular sites, shown in several cases to represent membrane contact sites (MCSs). Using established ORP ligand-binding domain mutants and pull-down assays with recombinant proteins, we show that ORP liganding regulates the ORP-VAP association, alters the subcellular targeting of ORP-VAP complexes, or modifies organelle morphology. There is distinct protein specificity in the effects of the mutants on subcellular targeting of ORP-VAP complexes. We provide evidence that complexes of human ORP2 and VAPs at ER-lipid droplet interfaces regulate the hydrolysis of triglycerides and lipid droplet turnover. The data suggest evolutionarily conserved, complex ligand-dependent functions of ORP-VAP complexes at MCSs, with implications for cellular lipid homeostasis and signaling.
WijdevenRH, JanssenH, NahidiazarL, JanssenL, JalinkK, BerlinI, NeefjesJ (2016). Cholesterol and ORP1L- mediated ER contact sites control autophagosome transport and fusion with the endocytic pathway Nat Commun 7, 11808. DOI:10.1038/ncomms11808PMID:27283760 [本文引用: 1] Autophagy is the main homeostatic pathway guiding cytosolic materials for degradation by the lysosome. Maturation of autophagosomes requires their transport towards the perinuclear region of the cell, with key factors underlying both processes still poorly understood. Here we show that transport and positioning of late autophagosomes depends on cholesterol by way of the cholesterol-sensing Rab7 effector ORP1L. ORP1L localizes to late autophagosomes and-under low-cholesterol conditions-contacts the ER protein VAP-A, forming ER-autophagosome contact sites, which prevent minus-end transport by the Rab7-RILP-dynein complex. ORP1L-mediated contact sites also inhibit localization of PLEKHM1 to Rab7. PLEKHM1, together with RILP, then recruits the homotypic fusion and vacuole protein-sorting ( HOPS) complex for fusion of autophagosomes with late endosomes and lysosomes. Thus, ORP1L, via its liganding by lipids and the formation of contacts between autophagic vacuoles and the ER, governs the last steps in autophagy that lead to the lysosomal degradation of cytosolic material.
WongLH, GattaAT, LevineTP (2019). Lipid transfer proteins: the lipid commute via shuttles, bridges and tubes Nat Rev Mol Cell Biol 20, 85-101. DOI:10.1038/s41580-018-0071-5URL [本文引用: 1]
WylesJP, McMasterCR, RidgwayND (2002). Vesicle- associated membrane protein-associated protein-A (VAP- A) interacts with the oxysterol-binding protein to modify export from the endoplasmic reticulum J Biol Chem 277, 29908-29918. DOI:10.1074/jbc.M201191200URL [本文引用: 1]
WylesJP, PerryRJ, RidgwayND (2007). Characterization of the sterol-binding domain of oxysterol-binding protein (OSBP)-related protein 4 reveals a novel role in vimentin organization Exp Cell Res 313, 1426-1437. DOI:10.1016/j.yexcr.2007.01.018URL [本文引用: 2]
WylesJP, RidgwayND (2004). VAMP-associated protein-A regulates partitioning of oxysterol-binding protein-related protein-9 between the endoplasmic reticulum and Golgi apparatus Exp Cell Res 297, 533-547. DOI:10.1016/j.yexcr.2004.03.052URL [本文引用: 1]
XingJJ, ZhangL, DuanZK, LinJX (2021). Coordination of phospholipid-based signaling and membrane trafficking in plant immunity Trends Plant Sci 26, 407-420. DOI:10.1016/j.tplants.2020.11.010URL [本文引用: 2]
XueHW, ChenX, MeiY (2009). Function and regulation of phospholipid signaling in plants Biochem J 421, 145-156. DOI:10.1042/BJ20090300URL [本文引用: 1]
YanDG, LehtoM, RasilainenL, MetsoJ, EhnholmC, Ylä-HerttualaS, JauhiainenM, OlkkonenVM (2007). Oxysterol binding protein induces upregulation of SREBP- 1c and enhances hepatic lipogenesis Arterioscler Thromb Vasc Biol 27, 1108-1114. DOI:10.1161/ATVBAHA.106.138545URL [本文引用: 1]
ZhuDM, ZhangMD, GaoCJ, ShenJB (2020). Protein trafficking in plant cells: tools and markers Sci China Life Sci 63, 343-363. DOI:10.1007/s11427-019-9598-3URL [本文引用: 1]
ZhuangXH, ChungKP, CuiY, LinWL, GaoCJ, KangBH, JiangLW (2017). ATG9 regulates autophagosome progression from the endoplasmic reticulum in Arabidopsis Proc Natl Acad Sci USA 114, E426-E435. DOI:10.1073/pnas.1616299114URL [本文引用: 1]
Differential regulation of endoplasmic reticulum structure through VAP-Nir protein interaction 1 2005
... VAPs是一类在真核生物中非常保守的囊泡相关膜蛋白(Nishimura et al., 1999).动物VAPs家族成员众多且分布广泛, 主要通过FFAT基序定位在内质网和内质网与其它膜形成的MCS (Murphy and Levine, 2016).VAPs一般含有MSP蛋白结构域(major sperm protein)、Coiled-coil结构域和C端跨膜结构域(C-terminal transmembrane domain, TMD).尚未在酵母中发现可以形成二聚体的Coiled-coil结构域或其它类似结构域.VAPs主要与3类蛋白互作: 突触囊泡SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptors)、病毒性蛋白和含有FFAT基序的蛋白(Lev et al., 2008).VAPs与含有FFAT基序的蛋白互作发现于Scs2p与Opi1p相互关系的研究(Loewen et al., 2003).通过FFAT基序与VAPs互作的脂类运输相关蛋白主要包括ORPs、CERT (ceramide transport protein)和Nirs (PYK2 N-terminal domain-interacting receptors) 3个蛋白家族(Wyles and Ridgway, 2004; Amarilio et al., 2005; Kawano et al., 2006).除了典型的FFAT基序, ORP3还含有2个FFAT-like基序(HFFSGST和NYSDGSE), 介导与VAPs的互作(Weber-Boyvat et al., 2015a, 2015b). ...
An oxysterol-binding protein family identified in the mouse 2 2002
... ORPs家族的第1个成员OSBP最初是作为氧化甾醇的受体被发现, 其通过抑制3-羟基-3-甲基戊二酰辅酶A还原酶(HMG-CoA)的活性抑制胆固醇的合成(Kandutsch and Thompson, 1980; Taylor et al., 1984; Taylor and Kandutsch, 1985).在不同物种中ORPs家族成员的数目不尽相同, 但功能域保守性很强.在人类细胞中, ORPs包含12个成员(Jaworski et al., 2001).老鼠细胞也包含12个成员(Anniss et al., 2002).在酵母中, ORPs包含7个成员(Beh et al., 2001).模式植物拟南芥(Arabidopsis thaliana)与人类和老鼠一样, 其ORPs包含12个成员(Skirpan et al., 2006).ORPs一般含有结合磷脂酰肌醇的PH蛋白结构域、介导二聚化的coiled-coil蛋白结构域、FFAT基序(two phenylalanine in an acidic tract motif)以及含高度保守的EQVSHHPP序列的可结合脂类的ORD蛋白结构域(OSBP-related domain). ...
... 在众多LTPs中, PH蛋白结构域相当保守, 且在人类基因组中含量很高.约10% PH蛋白结构域与磷酸肌醇强特异性结合, 从而使蛋白结合到特定膜上, 其余90% PH蛋白结构域功能还不清楚(Lemmon, 2007).在人类细胞12个成员中, 除了ORP2和ORP9, 其余成员均含有PH蛋白结构域(Lehto et al., 2001).在老鼠细胞12个成员中, 7个含有PH蛋白结构域(Anniss et al., 2002).在酵母7个成员中, Osh1p- Osh3p含有PH蛋白结构域(Beh et al., 2001).在拟南芥12个成员中, 5个成员含有PH蛋白结构域(Skirpan et al., 2006). ...
How accurately can we image inositol lipids in living cells? 1 2000
... 由于PH蛋白结构域与磷酸肌醇结合具有特异性, PH-GFP常用来指示特定磷酸肌醇的定位(Balla et al., 2000; Rusten and Stenmark, 2006).ORP5和ORP8的PH蛋白结构域通过与N端相邻的多碱基序列识别PI4P和PI(4,5)P2而将其定位于质膜(Ghai et al., 2017).虽然磷酸肌醇有助于PH蛋白结构域结合到膜上, 但是它们之间的定位不特异.例如, Osh1p通过与PI4P互作定位到高尔基体膜, Osh2p通过与PI4P互作不仅定位在高尔基体膜, 也定位在质膜(Roy and Levine, 2004; Yu et al., 2004).因此, ORPs的特异性定位不仅依赖于PH蛋白结构域, 还需要多种因子的参与.例如, OSBP定位在高尔基体膜不仅依赖于PH蛋白结构域与高尔基体膜上PI4P的结合, 还需要ARF (GTPase ADP-ribosylation factor)蛋白的参与(Levine and Munro, 2002; Godi et al., 2004). ...
Lipid dynamics at contact sites between the endoplasmic reticulum and other organelles 1 2019
... 细胞膜包括细胞质膜, 以及细胞核、线粒体、质体、内质网(endoplasmic reticulum, ER)、高尔基体、液泡等细胞器膜和胞内运输所需要的各种类型囊泡膜所组成的膜系统.脂质是细胞膜的基本组分, 包含脂肪(fats)和类脂(lipids).类脂又分为磷脂(phospholipids)、糖脂(glycolipids)、脂蛋白(lipoproteins)和固醇类(steroid).不同物种、不同类型的细胞以及不同细胞器的细胞膜所含脂质的种类、分布和含量都不尽相同.不同的脂质分子在细胞生命活动过程中起着非常重要的作用.脂质不仅是细胞膜的结构成分, 而且参与能量储存以及维持细胞膜的流动性和通透性, 特别是作为信号转导的关键组分参与生物体的个体发育和生理过程.然而, 膜脂信号在胞内的传送需要脂类运输蛋白(lipid transfer proteins, LTPs)的充分配合.正是由于LTPs在胞内信号转导中具有不可或缺的作用, 其生理功能备受研究者的关注, 相关研究在近几年也取得了重要进展(Pietrangelo and Ridgway, 2018; Balla et al., 2019; Wong et al., 2019). ...
Overlapping functions of the yeast oxysterol-binding protein homologues 3 2001
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
... ORPs家族的第1个成员OSBP最初是作为氧化甾醇的受体被发现, 其通过抑制3-羟基-3-甲基戊二酰辅酶A还原酶(HMG-CoA)的活性抑制胆固醇的合成(Kandutsch and Thompson, 1980; Taylor et al., 1984; Taylor and Kandutsch, 1985).在不同物种中ORPs家族成员的数目不尽相同, 但功能域保守性很强.在人类细胞中, ORPs包含12个成员(Jaworski et al., 2001).老鼠细胞也包含12个成员(Anniss et al., 2002).在酵母中, ORPs包含7个成员(Beh et al., 2001).模式植物拟南芥(Arabidopsis thaliana)与人类和老鼠一样, 其ORPs包含12个成员(Skirpan et al., 2006).ORPs一般含有结合磷脂酰肌醇的PH蛋白结构域、介导二聚化的coiled-coil蛋白结构域、FFAT基序(two phenylalanine in an acidic tract motif)以及含高度保守的EQVSHHPP序列的可结合脂类的ORD蛋白结构域(OSBP-related domain). ...
... 在众多LTPs中, PH蛋白结构域相当保守, 且在人类基因组中含量很高.约10% PH蛋白结构域与磷酸肌醇强特异性结合, 从而使蛋白结合到特定膜上, 其余90% PH蛋白结构域功能还不清楚(Lemmon, 2007).在人类细胞12个成员中, 除了ORP2和ORP9, 其余成员均含有PH蛋白结构域(Lehto et al., 2001).在老鼠细胞12个成员中, 7个含有PH蛋白结构域(Anniss et al., 2002).在酵母7个成员中, Osh1p- Osh3p含有PH蛋白结构域(Beh et al., 2001).在拟南芥12个成员中, 5个成员含有PH蛋白结构域(Skirpan et al., 2006). ...
A role for yeast oxysterol-binding protein homologs in endocytosis and in the maintenance of intracellular sterol-lipid distribution 1 2004
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
Screening for in planta protein-protein interactions combining bimolecular fluorescence complementation with flow cytometry 1 2012
... 虽然植物中的ORPs基因早已被克隆, 但关于植物ORPs的功能却少有研究.目前仅有研究表明ORP3A可通过与VAP33家族成员PVA12相互作用而定位于内质网(Saravanan et al., 2009).BIFC实验证明ORP2A与CPK3相互作用, 但在酵母双杂交实验中未检测到二者互作(Berendzen et al., 2012).PiORP1通过与矮牵牛(Petunia hybrida)花粉管质膜上的PRK1受体激酶相互作用参与花粉的生长与发育(Skirpan et al., 2006).在高盐胁迫条件下大豆(Glycine max) GmOSBP表达被抑制, 而在衰老叶片中表达被诱导, 说明GmOSBP可能参与应激反应和细胞衰老过程(Li et al., 2008).与人类或酵母ORPs的研究相比, 植物ORPs影响细胞内脂质转运的机制尚不清楚. ...
Cellular processes relying on sterol function in plants 1 2009
... 固醇类也是细胞膜的结构成分之一, 作为信号物质在细胞膜或细胞核内对生物体的生长发育起至关重要的作用(Boutté and Grebe, 2009; Ferrer et al., 2017).有些固醇既存在于动物中, 也存在于植物中, 如胆固醇、谷甾醇和菜油甾醇(Tarkowská, 2019).人体细胞内胆固醇水平的变化能够引起心血管疾病、神经性疾病和相关癌症(Luo et al., 2020).目前植物固醇的分子功能研究相对较少.在植物中, 油菜素内酯(brassinosteroids, BRs)是一种重要的植物固醇类激素(Geuns, 1983), 通过与膜受体BRI1蛋白结合, 调控细胞的伸长、分裂和分化(Li and He, 2020).对动物与酵母的研究表明, 类固醇分子能够与鞘脂类结合形成一种称为脂筏(lipid rafts)的微区域, 介导细胞间的识别、脂膜和蛋白质的细胞内运输、信号转导、逆境反应和极性生长等多种生物学过程(Malinsky et al., 2013; Tapken and Murphy, 2015).植物中也存在类似的富含固醇和鞘脂的细胞膜微区域, 被称为变性剂不溶或者不敏感膜区(detergent-insoluble (resistant) membranes, DIMs/DRMs), 其中所含蛋白质可对外界刺激产生动态应答(Takahashi et al., 2013; Zauber et al., 2014).由此表明, 植物细胞膜上形成的固醇类富集的细胞膜微区域可能对植物发育和环境信号应答具有重要作用. ...
Characterization of the properties of a novel mutation in VAPB in familial amyotrophic lateral sclerosis 1 2010
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
PI4P/phosphatidylserine countertransport at ORP5- and ORP8-mediated ER-plasma membrane contacts 4 2015
... ORPs运输脂类的过程一般发生在MCS.例如, OSBP在内质网-高尔基体之间传递胆固醇与PI4P (Mesmin et al., 2013).Osh6p、ORP5和ORP8均在内质网-质膜之间传递磷脂酰丝氨酸与PI4P (Maeda et al., 2013; Chung et al., 2015).目前已报道的ORPs的定位及功能见表1. ...
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
... ORP5和ORP8可以在EPCS区(ER-PM contact site)转运PI4P和磷脂酰丝氨酸(Chung et al., 2015).研究表明, PI(4,5)P2也可以作为ORP5和ORP8的供体(Ghai et al., 2017).ORP5和ORP8还定位于内质网-线粒体和内质网-后期囊泡(late endosome)的MCS, 调节胆固醇的再分配(Ishikawa et al., 2010; Galmes et al., 2016). ...
How phosphoinositides shape autophagy in plant cells 1 2019
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
Lipid- transfer proteins in biosynthetic pathways 1 2008
... 脂质在细胞膜上的分布不均一(van Meer et al., 2008).大多数脂质主要在内质网中合成, 随后被运输到各类细胞膜(质膜上的脂类最多), 以维持脂质的特定区域化(Moreau et al., 1998; van Meer et al., 2008).脂类具有不溶于水的疏水特性.脂质是如何从合成部位(内质网)转移到各种细胞器的? 细胞如何维持固醇分子在不同细胞器膜中的不均匀分布? 这些都是具有科学意义的重要问题.脂类分子的运输对此过程起到关键作用.胞内脂类运输主要包括3种机制: 囊泡运输、非囊泡运输及膜与膜之间的交融扩散.由LTPs参与的非囊泡运输是脂类运输的主要方式.LTPs主要有3方面功能.(1) 作为脂类的运输蛋白, 从一种膜撷取脂类运输到另一种膜.(2) 作为脂类的感受器, 通过与某种蛋白互作来结合或释放某种脂类.(3) 作为脂类的提供者, 如Sec14p (secretion 14 protein), 能够把磷脂酰肌醇运送至磷脂酰肌醇激酶 (Schaaf et al., 2008; Raychaudhuri and Prinz, 2010). LTPs是含有许多成员的大家族, 根据其脂质结合域可细分为SEC14、PITP (phosphatidylinositol transfer protein)、START (steroidogenic acute regulatory protein (StAR)-related transfer family)、GLTP (glycolipid transfer protein)、SCP-2 (sterol carrier protein 2)和OSBP/ORPs等家族(D'Angelo et al., 2008). ...
Phosphoinositides, major actors in membrane trafficking and lipid signaling pathways 2 2017
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
... (3) 植物ORPs是否参与植物免疫及对外界环境因子的应激反应.磷脂是磷脂双分子层的重要组成部分, 在信号转导中发挥重要作用(Takáč et al., 2019).磷脂的膜分布可影响膜的弯曲, 进而调节膜运输, 此过程对植物免疫应答反应至关重要(Wang et al., 2016b; De Craene et al., 2017).近期有综述阐明基于磷脂的信号转导和膜转运与植物免疫有密切联系(Xing et al., 2021).因此, 探讨脂类运输蛋白ORPs是否通过膜脂运输影响植物应对病原菌侵害和不适环境变化具有重要意义. ...
Osh4p exchanges sterols for phosphatidylinositol 4-phosphate between lipid bilayers 2 2011
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
... 在酵母细胞中, Osh1-3含有PH和ORD蛋白结构域, Osh4-7只含有ORD蛋白结构域(Raychaudhuri and Prinz, 2010).Osh1可以通过ANK蛋白结构域定位于细胞核与液泡之间的MCS, 转运磷脂和固醇, 进而促进细胞内脂质代谢(Levine and Munro, 2001; Manik et al., 2017).只有ORD蛋白结构域的Osh4p依然可通过形成PI4P浓度梯度在ER-PM之间运输固醇(de Saint-Jean et al., 2011; von Filseck et al., 2015b).与哺乳动物一样, 酵母细胞中固醇的转运也通过PI4P-磷酸酶Sac1水解PI4P形成一定浓度梯度来维持.Osh6p和Osh7p可以将磷脂酰丝氨酸从内质网转运到质膜(Maeda et al., 2013; von Filseck et al., 2015a). ...
A superfamily of actin-binding proteins at the actin-membrane nexus of higher plants 1 2012
... 拟南芥VAPs含有10个成员, 由于发现的第1个VAP分子量只有27 kDa, 故命名该家族为VAP27 (Nishimura et al., 1999).VAP27分为3个亚家族, 亚家族I包括5个成员(VAP27-1、VAP27-3、VAP27-5、VAP27-6和VAP27-7), 亚家族II包括3个成员(VAP27-8、VAP27-9和VAP27-10), 亚家族III包含2个成员(VAP27-2和VAP27-4).拟南芥VAPs同样含有MSD蛋白结构域、Coiled-coil蛋白结构域和跨膜区.VAP27蛋白主要定位在内质网和EPCS (Wang et al., 2016a).跨膜区的存在是VAPs定位在内质网的决定性区域.VAP27-1定位在EPCS还需要NET3C的介导(Wang et al., 2014).NET3C属于植物特异的肌动蛋白结合蛋白NET (networked)家族(Deeks et al., 2012).NET与肌动蛋白结合影响细胞骨架的运动, 因此VAP27蛋白可能通过NET来影响细胞骨架的运动. ...
Endosome-ER contacts control actin nucleation and retromer function through VAP-dependent regulation of PI4P 1 2016
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
A role for oxysterol-binding protein-related protein 5 in endosomal cholesterol trafficking 2 2011
... FFAT基序的氨基酸序列一般为-EFFDAXE-, 是内质网定位的决定性区域.人类细胞中除了ORP5和ORP8, 其它成员都含有FFAT基序.酵母Osh1-3含有FFAT基序(Raychaudhuri and Prinz, 2010).FFAT基序能够与VAPs (vesicle-associated membrane protein (VAMP)-associated proteins)蛋白互作, 而VAPs是一类内质网定位的囊泡运输相关蛋白(Skehel et al., 1995), 因此FFAT基序能够指导ORPs定位到内质网上(Loewen et al., 2003).但FFAT基序介导并不是ORPs定位到内质网上的唯一方式.例如, ORP5和ORP8虽然没有FFAT基序, 但是可通过C端的跨膜区定位到内质网膜(Yan et al., 2008; Du et al., 2011).Osh4p没有PH蛋白结构域和FFAT基序, 依然在内质网-质膜(plasma membrane) (ER-PM)上定位(Schulz et al., 2009).目前, 在植物界还未见FFAT基序或相似基序的功能报道. ...
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
... ORP5和ORP8可以在EPCS区(ER-PM contact site)转运PI4P和磷脂酰丝氨酸(Chung et al., 2015).研究表明, PI(4,5)P2也可以作为ORP5和ORP8的供体(Ghai et al., 2017).ORP5和ORP8还定位于内质网-线粒体和内质网-后期囊泡(late endosome)的MCS, 调节胆固醇的再分配(Ishikawa et al., 2010; Galmes et al., 2016). ...
FAPPs control Golgi-to-cell-surface membrane traffic by binding to ARF and PtdIns(4)P 1 2004
... 由于PH蛋白结构域与磷酸肌醇结合具有特异性, PH-GFP常用来指示特定磷酸肌醇的定位(Balla et al., 2000; Rusten and Stenmark, 2006).ORP5和ORP8的PH蛋白结构域通过与N端相邻的多碱基序列识别PI4P和PI(4,5)P2而将其定位于质膜(Ghai et al., 2017).虽然磷酸肌醇有助于PH蛋白结构域结合到膜上, 但是它们之间的定位不特异.例如, Osh1p通过与PI4P互作定位到高尔基体膜, Osh2p通过与PI4P互作不仅定位在高尔基体膜, 也定位在质膜(Roy and Levine, 2004; Yu et al., 2004).因此, ORPs的特异性定位不仅依赖于PH蛋白结构域, 还需要多种因子的参与.例如, OSBP定位在高尔基体膜不仅依赖于PH蛋白结构域与高尔基体膜上PI4P的结合, 还需要ARF (GTPase ADP-ribosylation factor)蛋白的参与(Levine and Munro, 2002; Godi et al., 2004). ...
Cell growth and differentiation in Arabidopsis epidermal cells 1 2007
... 在酵母细胞中, VAPs通过与ORPs互作介导脂类运输过程(Mesmin et al., 2013).但在植物中, 关于VAP27的功能及其作用机理的研究相对较少.已有研究表明, 拟南芥中鞘氨醇转移蛋白ACD11可以与VAP27-1相互作用(Petersen et al., 2009).VAP27- 1-GFP和VAP27-3-GFP转基因株系和功能缺失RNAi株系在根毛分叉表型上存在缺陷(Wang et al., 2016a).根毛分叉的表型与肌动蛋白互作蛋白AIP1突变体以及F-Actin2调节蛋白ROP2突变体的表型类似, 进一步说明VAP27可能通过调节肌动蛋白来调节根毛的发育(Guimil and Dunand, 2007; Ketelaar et al., 2007).VAP27-1和VAP27-3通过PIPs与网格蛋白的相互作用调节植物的内吞过程(Stefano et al., 2018).但是植物VAP27蛋白家族成员能否通过与ORP蛋白家族成员相互结合参与膜脂运输还未见报道. ...
ORP3 phosphorylation regulates phosphatidylinositol 4-phosphate and Ca2+ dynamics at plasma membrane-ER contact sites 1 2020
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
Understanding the diversity of membrane lipid composition 1 2018
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
Phosphoinositide signaling in plant development 1 2016
A family of 12 human genes containing oxysterol-binding domains 1 2001
... ORPs家族的第1个成员OSBP最初是作为氧化甾醇的受体被发现, 其通过抑制3-羟基-3-甲基戊二酰辅酶A还原酶(HMG-CoA)的活性抑制胆固醇的合成(Kandutsch and Thompson, 1980; Taylor et al., 1984; Taylor and Kandutsch, 1985).在不同物种中ORPs家族成员的数目不尽相同, 但功能域保守性很强.在人类细胞中, ORPs包含12个成员(Jaworski et al., 2001).老鼠细胞也包含12个成员(Anniss et al., 2002).在酵母中, ORPs包含7个成员(Beh et al., 2001).模式植物拟南芥(Arabidopsis thaliana)与人类和老鼠一样, 其ORPs包含12个成员(Skirpan et al., 2006).ORPs一般含有结合磷脂酰肌醇的PH蛋白结构域、介导二聚化的coiled-coil蛋白结构域、FFAT基序(two phenylalanine in an acidic tract motif)以及含高度保守的EQVSHHPP序列的可结合脂类的ORD蛋白结构域(OSBP-related domain). ...
The function of inositol phosphatases in plant tolerance to abiotic stress 1 2019
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
The oxysterol-binding protein homologue ORP1L interacts with Rab7 and alters functional properties of late endocytic compartments 1 2005
... ANK蛋白结构域是一种特殊的锚蛋白重复结构域.目前, 只发现在人类ORP1和酵母Osh1p-Osh2p中含有ANK蛋白结构域.Osh1p的ANK蛋白结构域通过与核-液泡连接蛋白1 (nuclear-vacuole junction protein 1, NVJ1)互作使其定位在细胞核和液泡之间(Manik et al., 2017).全长ORP1L (ORP1 large)通过ANK蛋白结构域与GTP结合将Rab7蛋白定位于晚期核内体与溶酶体之间, 可以干扰内膜的转运过程(Johansson et al., 2005; van der Kant et al., 2013).ANK结构域在Osh2p中的具体功能还不清楚. ...
Activation of endosomal dynein motors by stepwise assembly of Rab7- RILP-p150Glued, ORP1L, and the receptorβlll spectrin 1 2007
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
Investigating the contribution of VAPB/ALS8 loss of function in amyotrophic lateral sclerosis 1 2013
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
The yeast VAP homolog Scs2p has a phosphoinositide-binding ability that is correlated with its activity 1 2007
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
Role of the yeast VAP homolog, Scs2p, in INO1 expression and phospholipid metabolism 2 2003
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
... 的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
Cytosolic proteins that bind oxygenated sterols. Cellular distribution, specificity, and some properties 1 1980
... ORPs家族的第1个成员OSBP最初是作为氧化甾醇的受体被发现, 其通过抑制3-羟基-3-甲基戊二酰辅酶A还原酶(HMG-CoA)的活性抑制胆固醇的合成(Kandutsch and Thompson, 1980; Taylor et al., 1984; Taylor and Kandutsch, 1985).在不同物种中ORPs家族成员的数目不尽相同, 但功能域保守性很强.在人类细胞中, ORPs包含12个成员(Jaworski et al., 2001).老鼠细胞也包含12个成员(Anniss et al., 2002).在酵母中, ORPs包含7个成员(Beh et al., 2001).模式植物拟南芥(Arabidopsis thaliana)与人类和老鼠一样, 其ORPs包含12个成员(Skirpan et al., 2006).ORPs一般含有结合磷脂酰肌醇的PH蛋白结构域、介导二聚化的coiled-coil蛋白结构域、FFAT基序(two phenylalanine in an acidic tract motif)以及含高度保守的EQVSHHPP序列的可结合脂类的ORD蛋白结构域(OSBP-related domain). ...
Efficient trafficking of ceramide from the endoplasmic reticulum to the Golgi apparatus requires a VAMP-associated protein-interacting FFAT motif of CERT 2 2006
... VAPs是一类在真核生物中非常保守的囊泡相关膜蛋白(Nishimura et al., 1999).动物VAPs家族成员众多且分布广泛, 主要通过FFAT基序定位在内质网和内质网与其它膜形成的MCS (Murphy and Levine, 2016).VAPs一般含有MSP蛋白结构域(major sperm protein)、Coiled-coil结构域和C端跨膜结构域(C-terminal transmembrane domain, TMD).尚未在酵母中发现可以形成二聚体的Coiled-coil结构域或其它类似结构域.VAPs主要与3类蛋白互作: 突触囊泡SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptors)、病毒性蛋白和含有FFAT基序的蛋白(Lev et al., 2008).VAPs与含有FFAT基序的蛋白互作发现于Scs2p与Opi1p相互关系的研究(Loewen et al., 2003).通过FFAT基序与VAPs互作的脂类运输相关蛋白主要包括ORPs、CERT (ceramide transport protein)和Nirs (PYK2 N-terminal domain-interacting receptors) 3个蛋白家族(Wyles and Ridgway, 2004; Amarilio et al., 2005; Kawano et al., 2006).除了典型的FFAT基序, ORP3还含有2个FFAT-like基序(HFFSGST和NYSDGSE), 介导与VAPs的互作(Weber-Boyvat et al., 2015a, 2015b). ...
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
Actin organization and root hair development are disrupted by ethanol-induced overexpression of Arabidopsis actin interacting protein 1 (AIP1) 1 2007
... 在酵母细胞中, VAPs通过与ORPs互作介导脂类运输过程(Mesmin et al., 2013).但在植物中, 关于VAP27的功能及其作用机理的研究相对较少.已有研究表明, 拟南芥中鞘氨醇转移蛋白ACD11可以与VAP27-1相互作用(Petersen et al., 2009).VAP27- 1-GFP和VAP27-3-GFP转基因株系和功能缺失RNAi株系在根毛分叉表型上存在缺陷(Wang et al., 2016a).根毛分叉的表型与肌动蛋白互作蛋白AIP1突变体以及F-Actin2调节蛋白ROP2突变体的表型类似, 进一步说明VAP27可能通过调节肌动蛋白来调节根毛的发育(Guimil and Dunand, 2007; Ketelaar et al., 2007).VAP27-1和VAP27-3通过PIPs与网格蛋白的相互作用调节植物的内吞过程(Stefano et al., 2018).但是植物VAP27蛋白家族成员能否通过与ORP蛋白家族成员相互结合参与膜脂运输还未见报道. ...
Multivesicular body formation requires OSBP-related proteins and cholesterol 1 2010
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
A systematic screening to identify de novo mutations causing sporadic early-onset Parkinson’s disease 1 2015
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
Altered regulation of cholesterol and cholesteryl ester synthesis in Chinese-hamster ovary cells overexpressing the oxysterol-binding protein is dependent on the pleckstrin homology domain 1 1997
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
The OSBP-related protein family in humans 1 2001
... 在众多LTPs中, PH蛋白结构域相当保守, 且在人类基因组中含量很高.约10% PH蛋白结构域与磷酸肌醇强特异性结合, 从而使蛋白结合到特定膜上, 其余90% PH蛋白结构域功能还不清楚(Lemmon, 2007).在人类细胞12个成员中, 除了ORP2和ORP9, 其余成员均含有PH蛋白结构域(Lehto et al., 2001).在老鼠细胞12个成员中, 7个含有PH蛋白结构域(Anniss et al., 2002).在酵母7个成员中, Osh1p- Osh3p含有PH蛋白结构域(Beh et al., 2001).在拟南芥12个成员中, 5个成员含有PH蛋白结构域(Skirpan et al., 2006). ...
Pleckstrin homology (PH) domains and phosphoinositides 1 2007
... 在众多LTPs中, PH蛋白结构域相当保守, 且在人类基因组中含量很高.约10% PH蛋白结构域与磷酸肌醇强特异性结合, 从而使蛋白结合到特定膜上, 其余90% PH蛋白结构域功能还不清楚(Lemmon, 2007).在人类细胞12个成员中, 除了ORP2和ORP9, 其余成员均含有PH蛋白结构域(Lehto et al., 2001).在老鼠细胞12个成员中, 7个含有PH蛋白结构域(Anniss et al., 2002).在酵母7个成员中, Osh1p- Osh3p含有PH蛋白结构域(Beh et al., 2001).在拟南芥12个成员中, 5个成员含有PH蛋白结构域(Skirpan et al., 2006). ...
Oxysterol-binding protein-related protein (ORP) 9 is a PDK-2 substrate and regulates Akt phosphorylation 1 2007
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
The VAP protein family: from cellular functions to motor neuron disease 2 2008
... VAPs是一类在真核生物中非常保守的囊泡相关膜蛋白(Nishimura et al., 1999).动物VAPs家族成员众多且分布广泛, 主要通过FFAT基序定位在内质网和内质网与其它膜形成的MCS (Murphy and Levine, 2016).VAPs一般含有MSP蛋白结构域(major sperm protein)、Coiled-coil结构域和C端跨膜结构域(C-terminal transmembrane domain, TMD).尚未在酵母中发现可以形成二聚体的Coiled-coil结构域或其它类似结构域.VAPs主要与3类蛋白互作: 突触囊泡SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptors)、病毒性蛋白和含有FFAT基序的蛋白(Lev et al., 2008).VAPs与含有FFAT基序的蛋白互作发现于Scs2p与Opi1p相互关系的研究(Loewen et al., 2003).通过FFAT基序与VAPs互作的脂类运输相关蛋白主要包括ORPs、CERT (ceramide transport protein)和Nirs (PYK2 N-terminal domain-interacting receptors) 3个蛋白家族(Wyles and Ridgway, 2004; Amarilio et al., 2005; Kawano et al., 2006).除了典型的FFAT基序, ORP3还含有2个FFAT-like基序(HFFSGST和NYSDGSE), 介导与VAPs的互作(Weber-Boyvat et al., 2015a, 2015b). ...
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
Short-range intracellular trafficking of small molecules across endoplasmic reticulum junctions 1 2004
... 膜结合位点(membrane contact site, MCS)指2个细胞器膜之间距离小于30 nm的区域(Helle et al., 2013).小分子物质的交换以及信号的传递往往发生于MCS.内质网是细胞内分布最广的细胞器, 能够与多种细胞器形成MCS, 如质膜、高尔基体、核内体、线粒体及脂滴(Levine, 2004; Friedman and Voeltz, 2011; Helle et al., 2013).其它2种膜之间也可以形成MCS, 如核膜与液泡膜, 线粒体与液泡膜, 溶酶体与过氧化物酶体(Eisenberg-Bord et al., 2016; Saheki and De Camilli, 2017). ...
Dual targeting of Osh1p, a yeast homologue of oxysterol-binding protein, to both the Golgi and the nucleus-vacuole junction 2 2001
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
... 在酵母细胞中, Osh1-3含有PH和ORD蛋白结构域, Osh4-7只含有ORD蛋白结构域(Raychaudhuri and Prinz, 2010).Osh1可以通过ANK蛋白结构域定位于细胞核与液泡之间的MCS, 转运磷脂和固醇, 进而促进细胞内脂质代谢(Levine and Munro, 2001; Manik et al., 2017).只有ORD蛋白结构域的Osh4p依然可通过形成PI4P浓度梯度在ER-PM之间运输固醇(de Saint-Jean et al., 2011; von Filseck et al., 2015b).与哺乳动物一样, 酵母细胞中固醇的转运也通过PI4P-磷酸酶Sac1水解PI4P形成一定浓度梯度来维持.Osh6p和Osh7p可以将磷脂酰丝氨酸从内质网转运到质膜(Maeda et al., 2013; von Filseck et al., 2015a). ...
Targeting of Golgi-specific pleckstrin homology domains involves both PtdIns 4-kinase- dependent and-independent components 1 2002
... 由于PH蛋白结构域与磷酸肌醇结合具有特异性, PH-GFP常用来指示特定磷酸肌醇的定位(Balla et al., 2000; Rusten and Stenmark, 2006).ORP5和ORP8的PH蛋白结构域通过与N端相邻的多碱基序列识别PI4P和PI(4,5)P2而将其定位于质膜(Ghai et al., 2017).虽然磷酸肌醇有助于PH蛋白结构域结合到膜上, 但是它们之间的定位不特异.例如, Osh1p通过与PI4P互作定位到高尔基体膜, Osh2p通过与PI4P互作不仅定位在高尔基体膜, 也定位在质膜(Roy and Levine, 2004; Yu et al., 2004).因此, ORPs的特异性定位不仅依赖于PH蛋白结构域, 还需要多种因子的参与.例如, OSBP定位在高尔基体膜不仅依赖于PH蛋白结构域与高尔基体膜上PI4P的结合, 还需要ARF (GTPase ADP-ribosylation factor)蛋白的参与(Levine and Munro, 2002; Godi et al., 2004). ...
Molecular characterization of a novel salt-inducible gene for an OSBP (oxysterol-binding protein)-homologue from soybean 1 2008
... 虽然植物中的ORPs基因早已被克隆, 但关于植物ORPs的功能却少有研究.目前仅有研究表明ORP3A可通过与VAP33家族成员PVA12相互作用而定位于内质网(Saravanan et al., 2009).BIFC实验证明ORP2A与CPK3相互作用, 但在酵母双杂交实验中未检测到二者互作(Berendzen et al., 2012).PiORP1通过与矮牵牛(Petunia hybrida)花粉管质膜上的PRK1受体激酶相互作用参与花粉的生长与发育(Skirpan et al., 2006).在高盐胁迫条件下大豆(Glycine max) GmOSBP表达被抑制, 而在衰老叶片中表达被诱导, 说明GmOSBP可能参与应激反应和细胞衰老过程(Li et al., 2008).与人类或酵母ORPs的研究相比, 植物ORPs影响细胞内脂质转运的机制尚不清楚. ...
Roles of brassinosteroids in plant reproduction 1 2020
... 固醇类也是细胞膜的结构成分之一, 作为信号物质在细胞膜或细胞核内对生物体的生长发育起至关重要的作用(Boutté and Grebe, 2009; Ferrer et al., 2017).有些固醇既存在于动物中, 也存在于植物中, 如胆固醇、谷甾醇和菜油甾醇(Tarkowská, 2019).人体细胞内胆固醇水平的变化能够引起心血管疾病、神经性疾病和相关癌症(Luo et al., 2020).目前植物固醇的分子功能研究相对较少.在植物中, 油菜素内酯(brassinosteroids, BRs)是一种重要的植物固醇类激素(Geuns, 1983), 通过与膜受体BRI1蛋白结合, 调控细胞的伸长、分裂和分化(Li and He, 2020).对动物与酵母的研究表明, 类固醇分子能够与鞘脂类结合形成一种称为脂筏(lipid rafts)的微区域, 介导细胞间的识别、脂膜和蛋白质的细胞内运输、信号转导、逆境反应和极性生长等多种生物学过程(Malinsky et al., 2013; Tapken and Murphy, 2015).植物中也存在类似的富含固醇和鞘脂的细胞膜微区域, 被称为变性剂不溶或者不敏感膜区(detergent-insoluble (resistant) membranes, DIMs/DRMs), 其中所含蛋白质可对外界刺激产生动态应答(Takahashi et al., 2013; Zauber et al., 2014).由此表明, 植物细胞膜上形成的固醇类富集的细胞膜微区域可能对植物发育和环境信号应答具有重要作用. ...
Characterization of the sterol and phosphatidylinositol 4-phosphate binding properties of Golgi-associated OSBP-related protein 9 (ORP9) 1 2014
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
A conserved ER targeting motif in three families of lipid binding proteins and in Opi1p binds VAP 3 2003
... FFAT基序的氨基酸序列一般为-EFFDAXE-, 是内质网定位的决定性区域.人类细胞中除了ORP5和ORP8, 其它成员都含有FFAT基序.酵母Osh1-3含有FFAT基序(Raychaudhuri and Prinz, 2010).FFAT基序能够与VAPs (vesicle-associated membrane protein (VAMP)-associated proteins)蛋白互作, 而VAPs是一类内质网定位的囊泡运输相关蛋白(Skehel et al., 1995), 因此FFAT基序能够指导ORPs定位到内质网上(Loewen et al., 2003).但FFAT基序介导并不是ORPs定位到内质网上的唯一方式.例如, ORP5和ORP8虽然没有FFAT基序, 但是可通过C端的跨膜区定位到内质网膜(Yan et al., 2008; Du et al., 2011).Osh4p没有PH蛋白结构域和FFAT基序, 依然在内质网-质膜(plasma membrane) (ER-PM)上定位(Schulz et al., 2009).目前, 在植物界还未见FFAT基序或相似基序的功能报道. ...
... VAPs是一类在真核生物中非常保守的囊泡相关膜蛋白(Nishimura et al., 1999).动物VAPs家族成员众多且分布广泛, 主要通过FFAT基序定位在内质网和内质网与其它膜形成的MCS (Murphy and Levine, 2016).VAPs一般含有MSP蛋白结构域(major sperm protein)、Coiled-coil结构域和C端跨膜结构域(C-terminal transmembrane domain, TMD).尚未在酵母中发现可以形成二聚体的Coiled-coil结构域或其它类似结构域.VAPs主要与3类蛋白互作: 突触囊泡SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptors)、病毒性蛋白和含有FFAT基序的蛋白(Lev et al., 2008).VAPs与含有FFAT基序的蛋白互作发现于Scs2p与Opi1p相互关系的研究(Loewen et al., 2003).通过FFAT基序与VAPs互作的脂类运输相关蛋白主要包括ORPs、CERT (ceramide transport protein)和Nirs (PYK2 N-terminal domain-interacting receptors) 3个蛋白家族(Wyles and Ridgway, 2004; Amarilio et al., 2005; Kawano et al., 2006).除了典型的FFAT基序, ORP3还含有2个FFAT-like基序(HFFSGST和NYSDGSE), 介导与VAPs的互作(Weber-Boyvat et al., 2015a, 2015b). ...
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
Lipid bilayer regulation of membrane protein function: gramicidin channels as molecular force probes 1 2010
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
OSBP- and FAN-mediated sterol requirement for spermatogenesis in Drosophila 1 2010
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
Interactome map uncovers phosphatidylserine transport by oxysterol-binding proteins 5 2013
... ORPs运输脂类的过程一般发生在MCS.例如, OSBP在内质网-高尔基体之间传递胆固醇与PI4P (Mesmin et al., 2013).Osh6p、ORP5和ORP8均在内质网-质膜之间传递磷脂酰丝氨酸与PI4P (Maeda et al., 2013; Chung et al., 2015).目前已报道的ORPs的定位及功能见表1. ...
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
... 在酵母细胞中, Osh1-3含有PH和ORD蛋白结构域, Osh4-7只含有ORD蛋白结构域(Raychaudhuri and Prinz, 2010).Osh1可以通过ANK蛋白结构域定位于细胞核与液泡之间的MCS, 转运磷脂和固醇, 进而促进细胞内脂质代谢(Levine and Munro, 2001; Manik et al., 2017).只有ORD蛋白结构域的Osh4p依然可通过形成PI4P浓度梯度在ER-PM之间运输固醇(de Saint-Jean et al., 2011; von Filseck et al., 2015b).与哺乳动物一样, 酵母细胞中固醇的转运也通过PI4P-磷酸酶Sac1水解PI4P形成一定浓度梯度来维持.Osh6p和Osh7p可以将磷脂酰丝氨酸从内质网转运到质膜(Maeda et al., 2013; von Filseck et al., 2015a). ...
Membrane microdomains, rafts, and detergent- resistant membranes in plants and fungi 1 2013
... 固醇类也是细胞膜的结构成分之一, 作为信号物质在细胞膜或细胞核内对生物体的生长发育起至关重要的作用(Boutté and Grebe, 2009; Ferrer et al., 2017).有些固醇既存在于动物中, 也存在于植物中, 如胆固醇、谷甾醇和菜油甾醇(Tarkowská, 2019).人体细胞内胆固醇水平的变化能够引起心血管疾病、神经性疾病和相关癌症(Luo et al., 2020).目前植物固醇的分子功能研究相对较少.在植物中, 油菜素内酯(brassinosteroids, BRs)是一种重要的植物固醇类激素(Geuns, 1983), 通过与膜受体BRI1蛋白结合, 调控细胞的伸长、分裂和分化(Li and He, 2020).对动物与酵母的研究表明, 类固醇分子能够与鞘脂类结合形成一种称为脂筏(lipid rafts)的微区域, 介导细胞间的识别、脂膜和蛋白质的细胞内运输、信号转导、逆境反应和极性生长等多种生物学过程(Malinsky et al., 2013; Tapken and Murphy, 2015).植物中也存在类似的富含固醇和鞘脂的细胞膜微区域, 被称为变性剂不溶或者不敏感膜区(detergent-insoluble (resistant) membranes, DIMs/DRMs), 其中所含蛋白质可对外界刺激产生动态应答(Takahashi et al., 2013; Zauber et al., 2014).由此表明, 植物细胞膜上形成的固醇类富集的细胞膜微区域可能对植物发育和环境信号应答具有重要作用. ...
Structure of yeast OSBP-related protein OSH1 reveals key determinants for lipid transport and protein targeting at the nucleus-vacuole junction 2 2017
... ANK蛋白结构域是一种特殊的锚蛋白重复结构域.目前, 只发现在人类ORP1和酵母Osh1p-Osh2p中含有ANK蛋白结构域.Osh1p的ANK蛋白结构域通过与核-液泡连接蛋白1 (nuclear-vacuole junction protein 1, NVJ1)互作使其定位在细胞核和液泡之间(Manik et al., 2017).全长ORP1L (ORP1 large)通过ANK蛋白结构域与GTP结合将Rab7蛋白定位于晚期核内体与溶酶体之间, 可以干扰内膜的转运过程(Johansson et al., 2005; van der Kant et al., 2013).ANK结构域在Osh2p中的具体功能还不清楚. ...
... 在酵母细胞中, Osh1-3含有PH和ORD蛋白结构域, Osh4-7只含有ORD蛋白结构域(Raychaudhuri and Prinz, 2010).Osh1可以通过ANK蛋白结构域定位于细胞核与液泡之间的MCS, 转运磷脂和固醇, 进而促进细胞内脂质代谢(Levine and Munro, 2001; Manik et al., 2017).只有ORD蛋白结构域的Osh4p依然可通过形成PI4P浓度梯度在ER-PM之间运输固醇(de Saint-Jean et al., 2011; von Filseck et al., 2015b).与哺乳动物一样, 酵母细胞中固醇的转运也通过PI4P-磷酸酶Sac1水解PI4P形成一定浓度梯度来维持.Osh6p和Osh7p可以将磷脂酰丝氨酸从内质网转运到质膜(Maeda et al., 2013; von Filseck et al., 2015a). ...
VAMP associated proteins are required for autophagic and lysosomal degradation by promoting a PtdIns4P-medIated endosomal pathway 2 2019
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
... ).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
PI(4,5)P2-binding effector proteins for vesicle exocytosis 1 2015
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
Sterol transfer, PI4P consumption, and control of membrane lipid order by endogenous OSBP 1 2017
... 氧化固醇结合蛋白(OSBP)在内质网和高尔基体之间膜结合位点转运胆固醇的功能模式图(改自Mesmin et al., 2013; Pietrangelo and Ridgway, 2018) OSBP通过FFAT基序与VAP27-A互作, 定位于内质网, 通过PH蛋白结构域定位在高尔基体.OSBP一般通过Coiled-coil结构域形成二聚体行使功能.ORD蛋白结构域在磷脂酰肌醇-4-磷酸(PI4P)逆行转运的驱动下, 将胆固醇从内质网转移到高尔基体. ...
... OSBP通过FFAT基序与VAP27-A互作, 定位于内质网, 通过PH蛋白结构域定位在高尔基体.OSBP一般通过Coiled-coil结构域形成二聚体行使功能.ORD蛋白结构域在磷脂酰肌醇-4-磷酸(PI4P)逆行转运的驱动下, 将胆固醇从内质网转移到高尔基体. Working model of oxysterol-binding protein (OSBP) function in cholesterol transportation at membrane contact site (MCS) between endoplasmic reticulum (ER) and <i>trans</i>-Golgi (modified from <xref ref-type="bibr" rid="b72">Mesmin et al., 2013</xref>; <xref ref-type="bibr" rid="b80">Pietrangelo and Ridgway, 2018</xref>) OSBP is located in ER by FFAT motif interacting with VAP27-A and in trans-Golgi by the PH domain. OSBP usually forms dimers through the Coiled-coil domain to play its role. The ORD domain could move cholesterol from the ER to trans-Golgi, driven by the retrograde transport of phosphatidylinositol-4-phosphate (PI4P). ...
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
... 在酵母细胞中, VAPs通过与ORPs互作介导脂类运输过程(Mesmin et al., 2013).但在植物中, 关于VAP27的功能及其作用机理的研究相对较少.已有研究表明, 拟南芥中鞘氨醇转移蛋白ACD11可以与VAP27-1相互作用(Petersen et al., 2009).VAP27- 1-GFP和VAP27-3-GFP转基因株系和功能缺失RNAi株系在根毛分叉表型上存在缺陷(Wang et al., 2016a).根毛分叉的表型与肌动蛋白互作蛋白AIP1突变体以及F-Actin2调节蛋白ROP2突变体的表型类似, 进一步说明VAP27可能通过调节肌动蛋白来调节根毛的发育(Guimil and Dunand, 2007; Ketelaar et al., 2007).VAP27-1和VAP27-3通过PIPs与网格蛋白的相互作用调节植物的内吞过程(Stefano et al., 2018).但是植物VAP27蛋白家族成员能否通过与ORP蛋白家族成员相互结合参与膜脂运输还未见报道. ...
Oxysterol-binding protein- related protein (ORP) 6 localizes to the ER and ER- plasma membrane contact sites and is involved in the turnover of PI4P in cerebellar granule neurons 1 2018
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
Transport of sterols to the plasma membrane of leek seedlings 1 1998
... 脂质在细胞膜上的分布不均一(van Meer et al., 2008).大多数脂质主要在内质网中合成, 随后被运输到各类细胞膜(质膜上的脂类最多), 以维持脂质的特定区域化(Moreau et al., 1998; van Meer et al., 2008).脂类具有不溶于水的疏水特性.脂质是如何从合成部位(内质网)转移到各种细胞器的? 细胞如何维持固醇分子在不同细胞器膜中的不均匀分布? 这些都是具有科学意义的重要问题.脂类分子的运输对此过程起到关键作用.胞内脂类运输主要包括3种机制: 囊泡运输、非囊泡运输及膜与膜之间的交融扩散.由LTPs参与的非囊泡运输是脂类运输的主要方式.LTPs主要有3方面功能.(1) 作为脂类的运输蛋白, 从一种膜撷取脂类运输到另一种膜.(2) 作为脂类的感受器, 通过与某种蛋白互作来结合或释放某种脂类.(3) 作为脂类的提供者, 如Sec14p (secretion 14 protein), 能够把磷脂酰肌醇运送至磷脂酰肌醇激酶 (Schaaf et al., 2008; Raychaudhuri and Prinz, 2010). LTPs是含有许多成员的大家族, 根据其脂质结合域可细分为SEC14、PITP (phosphatidylinositol transfer protein)、START (steroidogenic acute regulatory protein (StAR)-related transfer family)、GLTP (glycolipid transfer protein)、SCP-2 (sterol carrier protein 2)和OSBP/ORPs等家族(D'Angelo et al., 2008). ...
VAP, a versatile access point for the endoplasmic reticulum: review and analysis of FFAT-like motifs in the VAPome 1 2016
... VAPs是一类在真核生物中非常保守的囊泡相关膜蛋白(Nishimura et al., 1999).动物VAPs家族成员众多且分布广泛, 主要通过FFAT基序定位在内质网和内质网与其它膜形成的MCS (Murphy and Levine, 2016).VAPs一般含有MSP蛋白结构域(major sperm protein)、Coiled-coil结构域和C端跨膜结构域(C-terminal transmembrane domain, TMD).尚未在酵母中发现可以形成二聚体的Coiled-coil结构域或其它类似结构域.VAPs主要与3类蛋白互作: 突触囊泡SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptors)、病毒性蛋白和含有FFAT基序的蛋白(Lev et al., 2008).VAPs与含有FFAT基序的蛋白互作发现于Scs2p与Opi1p相互关系的研究(Loewen et al., 2003).通过FFAT基序与VAPs互作的脂类运输相关蛋白主要包括ORPs、CERT (ceramide transport protein)和Nirs (PYK2 N-terminal domain-interacting receptors) 3个蛋白家族(Wyles and Ridgway, 2004; Amarilio et al., 2005; Kawano et al., 2006).除了典型的FFAT基序, ORP3还含有2个FFAT-like基序(HFFSGST和NYSDGSE), 介导与VAPs的互作(Weber-Boyvat et al., 2015a, 2015b). ...
Oxysterol binding protein- related protein 9 (ORP9) is a cholesterol transfer protein that regulates Golgi structure and function 1 2009
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
A mutation in the vesicle-trafficking protein VAPB causes late-onset spinal muscular atrophy and amyotrophic lateral sclerosis 1 2004
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
Molecular cloning and characterization of mammalian homologues of vesicle-associated membrane protein-assocIated (VAMP-associated) proteins 2 1999
... VAPs是一类在真核生物中非常保守的囊泡相关膜蛋白(Nishimura et al., 1999).动物VAPs家族成员众多且分布广泛, 主要通过FFAT基序定位在内质网和内质网与其它膜形成的MCS (Murphy and Levine, 2016).VAPs一般含有MSP蛋白结构域(major sperm protein)、Coiled-coil结构域和C端跨膜结构域(C-terminal transmembrane domain, TMD).尚未在酵母中发现可以形成二聚体的Coiled-coil结构域或其它类似结构域.VAPs主要与3类蛋白互作: 突触囊泡SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptors)、病毒性蛋白和含有FFAT基序的蛋白(Lev et al., 2008).VAPs与含有FFAT基序的蛋白互作发现于Scs2p与Opi1p相互关系的研究(Loewen et al., 2003).通过FFAT基序与VAPs互作的脂类运输相关蛋白主要包括ORPs、CERT (ceramide transport protein)和Nirs (PYK2 N-terminal domain-interacting receptors) 3个蛋白家族(Wyles and Ridgway, 2004; Amarilio et al., 2005; Kawano et al., 2006).除了典型的FFAT基序, ORP3还含有2个FFAT-like基序(HFFSGST和NYSDGSE), 介导与VAPs的互作(Weber-Boyvat et al., 2015a, 2015b). ...
... 拟南芥VAPs含有10个成员, 由于发现的第1个VAP分子量只有27 kDa, 故命名该家族为VAP27 (Nishimura et al., 1999).VAP27分为3个亚家族, 亚家族I包括5个成员(VAP27-1、VAP27-3、VAP27-5、VAP27-6和VAP27-7), 亚家族II包括3个成员(VAP27-8、VAP27-9和VAP27-10), 亚家族III包含2个成员(VAP27-2和VAP27-4).拟南芥VAPs同样含有MSD蛋白结构域、Coiled-coil蛋白结构域和跨膜区.VAP27蛋白主要定位在内质网和EPCS (Wang et al., 2016a).跨膜区的存在是VAPs定位在内质网的决定性区域.VAP27-1定位在EPCS还需要NET3C的介导(Wang et al., 2014).NET3C属于植物特异的肌动蛋白结合蛋白NET (networked)家族(Deeks et al., 2012).NET与肌动蛋白结合影响细胞骨架的运动, 因此VAP27蛋白可能通过NET来影响细胞骨架的运动. ...
Identification of proteins interacting with Arabidopsis ACD11 1 2009
... 在酵母细胞中, VAPs通过与ORPs互作介导脂类运输过程(Mesmin et al., 2013).但在植物中, 关于VAP27的功能及其作用机理的研究相对较少.已有研究表明, 拟南芥中鞘氨醇转移蛋白ACD11可以与VAP27-1相互作用(Petersen et al., 2009).VAP27- 1-GFP和VAP27-3-GFP转基因株系和功能缺失RNAi株系在根毛分叉表型上存在缺陷(Wang et al., 2016a).根毛分叉的表型与肌动蛋白互作蛋白AIP1突变体以及F-Actin2调节蛋白ROP2突变体的表型类似, 进一步说明VAP27可能通过调节肌动蛋白来调节根毛的发育(Guimil and Dunand, 2007; Ketelaar et al., 2007).VAP27-1和VAP27-3通过PIPs与网格蛋白的相互作用调节植物的内吞过程(Stefano et al., 2018).但是植物VAP27蛋白家族成员能否通过与ORP蛋白家族成员相互结合参与膜脂运输还未见报道. ...
Bridging the molecular and biological functions of the oxysterol-binding protein family 3 2018
... 细胞膜包括细胞质膜, 以及细胞核、线粒体、质体、内质网(endoplasmic reticulum, ER)、高尔基体、液泡等细胞器膜和胞内运输所需要的各种类型囊泡膜所组成的膜系统.脂质是细胞膜的基本组分, 包含脂肪(fats)和类脂(lipids).类脂又分为磷脂(phospholipids)、糖脂(glycolipids)、脂蛋白(lipoproteins)和固醇类(steroid).不同物种、不同类型的细胞以及不同细胞器的细胞膜所含脂质的种类、分布和含量都不尽相同.不同的脂质分子在细胞生命活动过程中起着非常重要的作用.脂质不仅是细胞膜的结构成分, 而且参与能量储存以及维持细胞膜的流动性和通透性, 特别是作为信号转导的关键组分参与生物体的个体发育和生理过程.然而, 膜脂信号在胞内的传送需要脂类运输蛋白(lipid transfer proteins, LTPs)的充分配合.正是由于LTPs在胞内信号转导中具有不可或缺的作用, 其生理功能备受研究者的关注, 相关研究在近几年也取得了重要进展(Pietrangelo and Ridgway, 2018; Balla et al., 2019; Wong et al., 2019). ...
Working model of oxysterol-binding protein (OSBP) function in cholesterol transportation at membrane contact site (MCS) between endoplasmic reticulum (ER) and <i>trans</i>-Golgi (modified from <xref ref-type="bibr" rid="b72">Mesmin et al., 2013</xref>; <xref ref-type="bibr" rid="b80">Pietrangelo and Ridgway, 2018</xref>) OSBP is located in ER by FFAT motif interacting with VAP27-A and in trans-Golgi by the PH domain. OSBP usually forms dimers through the Coiled-coil domain to play its role. The ORD domain could move cholesterol from the ER to trans-Golgi, driven by the retrograde transport of phosphatidylinositol-4-phosphate (PI4P). ...
Nonvesicular sterol movement from plasma membrane to ER requires oxysterol-binding protein-related proteins and phosphoinositides 1 2006
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
The diverse functions of oxysterol-binding proteins 3 2010
... 脂质在细胞膜上的分布不均一(van Meer et al., 2008).大多数脂质主要在内质网中合成, 随后被运输到各类细胞膜(质膜上的脂类最多), 以维持脂质的特定区域化(Moreau et al., 1998; van Meer et al., 2008).脂类具有不溶于水的疏水特性.脂质是如何从合成部位(内质网)转移到各种细胞器的? 细胞如何维持固醇分子在不同细胞器膜中的不均匀分布? 这些都是具有科学意义的重要问题.脂类分子的运输对此过程起到关键作用.胞内脂类运输主要包括3种机制: 囊泡运输、非囊泡运输及膜与膜之间的交融扩散.由LTPs参与的非囊泡运输是脂类运输的主要方式.LTPs主要有3方面功能.(1) 作为脂类的运输蛋白, 从一种膜撷取脂类运输到另一种膜.(2) 作为脂类的感受器, 通过与某种蛋白互作来结合或释放某种脂类.(3) 作为脂类的提供者, 如Sec14p (secretion 14 protein), 能够把磷脂酰肌醇运送至磷脂酰肌醇激酶 (Schaaf et al., 2008; Raychaudhuri and Prinz, 2010). LTPs是含有许多成员的大家族, 根据其脂质结合域可细分为SEC14、PITP (phosphatidylinositol transfer protein)、START (steroidogenic acute regulatory protein (StAR)-related transfer family)、GLTP (glycolipid transfer protein)、SCP-2 (sterol carrier protein 2)和OSBP/ORPs等家族(D'Angelo et al., 2008). ...
... FFAT基序的氨基酸序列一般为-EFFDAXE-, 是内质网定位的决定性区域.人类细胞中除了ORP5和ORP8, 其它成员都含有FFAT基序.酵母Osh1-3含有FFAT基序(Raychaudhuri and Prinz, 2010).FFAT基序能够与VAPs (vesicle-associated membrane protein (VAMP)-associated proteins)蛋白互作, 而VAPs是一类内质网定位的囊泡运输相关蛋白(Skehel et al., 1995), 因此FFAT基序能够指导ORPs定位到内质网上(Loewen et al., 2003).但FFAT基序介导并不是ORPs定位到内质网上的唯一方式.例如, ORP5和ORP8虽然没有FFAT基序, 但是可通过C端的跨膜区定位到内质网膜(Yan et al., 2008; Du et al., 2011).Osh4p没有PH蛋白结构域和FFAT基序, 依然在内质网-质膜(plasma membrane) (ER-PM)上定位(Schulz et al., 2009).目前, 在植物界还未见FFAT基序或相似基序的功能报道. ...
... 在酵母细胞中, Osh1-3含有PH和ORD蛋白结构域, Osh4-7只含有ORD蛋白结构域(Raychaudhuri and Prinz, 2010).Osh1可以通过ANK蛋白结构域定位于细胞核与液泡之间的MCS, 转运磷脂和固醇, 进而促进细胞内脂质代谢(Levine and Munro, 2001; Manik et al., 2017).只有ORD蛋白结构域的Osh4p依然可通过形成PI4P浓度梯度在ER-PM之间运输固醇(de Saint-Jean et al., 2011; von Filseck et al., 2015b).与哺乳动物一样, 酵母细胞中固醇的转运也通过PI4P-磷酸酶Sac1水解PI4P形成一定浓度梯度来维持.Osh6p和Osh7p可以将磷脂酰丝氨酸从内质网转运到质膜(Maeda et al., 2013; von Filseck et al., 2015a). ...
Cholesterol sensor ORP1L contacts the ER protein VAP to control Rab7-RILP-p150Glued and late endosome positioning 1 2009
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
... 膜结合位点(membrane contact site, MCS)指2个细胞器膜之间距离小于30 nm的区域(Helle et al., 2013).小分子物质的交换以及信号的传递往往发生于MCS.内质网是细胞内分布最广的细胞器, 能够与多种细胞器形成MCS, 如质膜、高尔基体、核内体、线粒体及脂滴(Levine, 2004; Friedman and Voeltz, 2011; Helle et al., 2013).其它2种膜之间也可以形成MCS, 如核膜与液泡膜, 线粒体与液泡膜, 溶酶体与过氧化物酶体(Eisenberg-Bord et al., 2016; Saheki and De Camilli, 2017). ...
The targeting of the oxysterol- binding protein ORP3a to the endoplasmic reticulum relies on the plant VAP33 homolog PVA12 1 2009
... 虽然植物中的ORPs基因早已被克隆, 但关于植物ORPs的功能却少有研究.目前仅有研究表明ORP3A可通过与VAP33家族成员PVA12相互作用而定位于内质网(Saravanan et al., 2009).BIFC实验证明ORP2A与CPK3相互作用, 但在酵母双杂交实验中未检测到二者互作(Berendzen et al., 2012).PiORP1通过与矮牵牛(Petunia hybrida)花粉管质膜上的PRK1受体激酶相互作用参与花粉的生长与发育(Skirpan et al., 2006).在高盐胁迫条件下大豆(Glycine max) GmOSBP表达被抑制, 而在衰老叶片中表达被诱导, 说明GmOSBP可能参与应激反应和细胞衰老过程(Li et al., 2008).与人类或酵母ORPs的研究相比, 植物ORPs影响细胞内脂质转运的机制尚不清楚. ...
Functional anatomy of phospholipid binding and regulation of phosphoinositide homeostasis by proteins of the sec14 superfamily 1 2008
... 脂质在细胞膜上的分布不均一(van Meer et al., 2008).大多数脂质主要在内质网中合成, 随后被运输到各类细胞膜(质膜上的脂类最多), 以维持脂质的特定区域化(Moreau et al., 1998; van Meer et al., 2008).脂类具有不溶于水的疏水特性.脂质是如何从合成部位(内质网)转移到各种细胞器的? 细胞如何维持固醇分子在不同细胞器膜中的不均匀分布? 这些都是具有科学意义的重要问题.脂类分子的运输对此过程起到关键作用.胞内脂类运输主要包括3种机制: 囊泡运输、非囊泡运输及膜与膜之间的交融扩散.由LTPs参与的非囊泡运输是脂类运输的主要方式.LTPs主要有3方面功能.(1) 作为脂类的运输蛋白, 从一种膜撷取脂类运输到另一种膜.(2) 作为脂类的感受器, 通过与某种蛋白互作来结合或释放某种脂类.(3) 作为脂类的提供者, 如Sec14p (secretion 14 protein), 能够把磷脂酰肌醇运送至磷脂酰肌醇激酶 (Schaaf et al., 2008; Raychaudhuri and Prinz, 2010). LTPs是含有许多成员的大家族, 根据其脂质结合域可细分为SEC14、PITP (phosphatidylinositol transfer protein)、START (steroidogenic acute regulatory protein (StAR)-related transfer family)、GLTP (glycolipid transfer protein)、SCP-2 (sterol carrier protein 2)和OSBP/ORPs等家族(D'Angelo et al., 2008). ...
Lipid-regulated sterol transfer between closely apposed membranes by oxysterol-binding protein homologues 6 2009
... FFAT基序的氨基酸序列一般为-EFFDAXE-, 是内质网定位的决定性区域.人类细胞中除了ORP5和ORP8, 其它成员都含有FFAT基序.酵母Osh1-3含有FFAT基序(Raychaudhuri and Prinz, 2010).FFAT基序能够与VAPs (vesicle-associated membrane protein (VAMP)-associated proteins)蛋白互作, 而VAPs是一类内质网定位的囊泡运输相关蛋白(Skehel et al., 1995), 因此FFAT基序能够指导ORPs定位到内质网上(Loewen et al., 2003).但FFAT基序介导并不是ORPs定位到内质网上的唯一方式.例如, ORP5和ORP8虽然没有FFAT基序, 但是可通过C端的跨膜区定位到内质网膜(Yan et al., 2008; Du et al., 2011).Osh4p没有PH蛋白结构域和FFAT基序, 依然在内质网-质膜(plasma membrane) (ER-PM)上定位(Schulz et al., 2009).目前, 在植物界还未见FFAT基序或相似基序的功能报道. ...
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
A VAMP-binding protein from Aplysia required for neurotransmitter release 1 1995
... FFAT基序的氨基酸序列一般为-EFFDAXE-, 是内质网定位的决定性区域.人类细胞中除了ORP5和ORP8, 其它成员都含有FFAT基序.酵母Osh1-3含有FFAT基序(Raychaudhuri and Prinz, 2010).FFAT基序能够与VAPs (vesicle-associated membrane protein (VAMP)-associated proteins)蛋白互作, 而VAPs是一类内质网定位的囊泡运输相关蛋白(Skehel et al., 1995), 因此FFAT基序能够指导ORPs定位到内质网上(Loewen et al., 2003).但FFAT基序介导并不是ORPs定位到内质网上的唯一方式.例如, ORP5和ORP8虽然没有FFAT基序, 但是可通过C端的跨膜区定位到内质网膜(Yan et al., 2008; Du et al., 2011).Osh4p没有PH蛋白结构域和FFAT基序, 依然在内质网-质膜(plasma membrane) (ER-PM)上定位(Schulz et al., 2009).目前, 在植物界还未见FFAT基序或相似基序的功能报道. ...
Identification and characterization of PiORP1, a Petunia oxysterol-binding-protein related protein involved in receptor-kinase mediated signaling in pollen, and analysis of the ORP gene family in Arabidopsis 5 2006
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
... ORPs家族的第1个成员OSBP最初是作为氧化甾醇的受体被发现, 其通过抑制3-羟基-3-甲基戊二酰辅酶A还原酶(HMG-CoA)的活性抑制胆固醇的合成(Kandutsch and Thompson, 1980; Taylor et al., 1984; Taylor and Kandutsch, 1985).在不同物种中ORPs家族成员的数目不尽相同, 但功能域保守性很强.在人类细胞中, ORPs包含12个成员(Jaworski et al., 2001).老鼠细胞也包含12个成员(Anniss et al., 2002).在酵母中, ORPs包含7个成员(Beh et al., 2001).模式植物拟南芥(Arabidopsis thaliana)与人类和老鼠一样, 其ORPs包含12个成员(Skirpan et al., 2006).ORPs一般含有结合磷脂酰肌醇的PH蛋白结构域、介导二聚化的coiled-coil蛋白结构域、FFAT基序(two phenylalanine in an acidic tract motif)以及含高度保守的EQVSHHPP序列的可结合脂类的ORD蛋白结构域(OSBP-related domain). ...
... 在众多LTPs中, PH蛋白结构域相当保守, 且在人类基因组中含量很高.约10% PH蛋白结构域与磷酸肌醇强特异性结合, 从而使蛋白结合到特定膜上, 其余90% PH蛋白结构域功能还不清楚(Lemmon, 2007).在人类细胞12个成员中, 除了ORP2和ORP9, 其余成员均含有PH蛋白结构域(Lehto et al., 2001).在老鼠细胞12个成员中, 7个含有PH蛋白结构域(Anniss et al., 2002).在酵母7个成员中, Osh1p- Osh3p含有PH蛋白结构域(Beh et al., 2001).在拟南芥12个成员中, 5个成员含有PH蛋白结构域(Skirpan et al., 2006). ...
... 虽然植物中的ORPs基因早已被克隆, 但关于植物ORPs的功能却少有研究.目前仅有研究表明ORP3A可通过与VAP33家族成员PVA12相互作用而定位于内质网(Saravanan et al., 2009).BIFC实验证明ORP2A与CPK3相互作用, 但在酵母双杂交实验中未检测到二者互作(Berendzen et al., 2012).PiORP1通过与矮牵牛(Petunia hybrida)花粉管质膜上的PRK1受体激酶相互作用参与花粉的生长与发育(Skirpan et al., 2006).在高盐胁迫条件下大豆(Glycine max) GmOSBP表达被抑制, 而在衰老叶片中表达被诱导, 说明GmOSBP可能参与应激反应和细胞衰老过程(Li et al., 2008).与人类或酵母ORPs的研究相比, 植物ORPs影响细胞内脂质转运的机制尚不清楚. ...
ERG30, a VAP-33- related protein, functions in protein transport mediated by COPI vesicles 1 1999
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
Plant endocytosis requires the ER membrane-anchored proteins VAP27-1 and VAP27-3 2 2018
... 在酵母细胞中, VAPs通过与ORPs互作介导脂类运输过程(Mesmin et al., 2013).但在植物中, 关于VAP27的功能及其作用机理的研究相对较少.已有研究表明, 拟南芥中鞘氨醇转移蛋白ACD11可以与VAP27-1相互作用(Petersen et al., 2009).VAP27- 1-GFP和VAP27-3-GFP转基因株系和功能缺失RNAi株系在根毛分叉表型上存在缺陷(Wang et al., 2016a).根毛分叉的表型与肌动蛋白互作蛋白AIP1突变体以及F-Actin2调节蛋白ROP2突变体的表型类似, 进一步说明VAP27可能通过调节肌动蛋白来调节根毛的发育(Guimil and Dunand, 2007; Ketelaar et al., 2007).VAP27-1和VAP27-3通过PIPs与网格蛋白的相互作用调节植物的内吞过程(Stefano et al., 2018).但是植物VAP27蛋白家族成员能否通过与ORP蛋白家族成员相互结合参与膜脂运输还未见报道. ...
... (2) 植物不同ORPs蛋白的生理机制及其在发育过程中的作用.例如, 它们是否参与细胞自噬; 在动物和酵母中, LTPs在MCS通过介导脂类从内质网至自噬体膜的运输, 参与自噬体的形成(Ye et al., 2020).在低胆固醇条件下, 人类ORP1L通过与VAP-A相互作用形成内质网-自噬体接触位点, 介导自噬体的运输(Wijdeven et al., 2016).VAP27-1和VAP27-3已被证明参与植物内吞过程(Wang et al., 2016a; Stefano et al., 2018).VAP27-1也可以与拟南芥EH蛋白(AtEH1/Pan1和AtEH2/Pan2)互作, 调节自噬体的形成(Wang et al., 2019).然而植物ORPs是否通过VAP27参与细胞自噬过程仍需更多实验验证. ...
Recent advances in the cellular and developmental biology of phospholipases in plants 1 2019
... (3) 植物ORPs是否参与植物免疫及对外界环境因子的应激反应.磷脂是磷脂双分子层的重要组成部分, 在信号转导中发挥重要作用(Takáč et al., 2019).磷脂的膜分布可影响膜的弯曲, 进而调节膜运输, 此过程对植物免疫应答反应至关重要(Wang et al., 2016b; De Craene et al., 2017).近期有综述阐明基于磷脂的信号转导和膜转运与植物免疫有密切联系(Xing et al., 2021).因此, 探讨脂类运输蛋白ORPs是否通过膜脂运输影响植物应对病原菌侵害和不适环境变化具有重要意义. ...
Changes of detergent-resistant plasma membrane proteins in oat and rye during cold acclimation: association with differential freezing tolerance 1 2013
... 固醇类也是细胞膜的结构成分之一, 作为信号物质在细胞膜或细胞核内对生物体的生长发育起至关重要的作用(Boutté and Grebe, 2009; Ferrer et al., 2017).有些固醇既存在于动物中, 也存在于植物中, 如胆固醇、谷甾醇和菜油甾醇(Tarkowská, 2019).人体细胞内胆固醇水平的变化能够引起心血管疾病、神经性疾病和相关癌症(Luo et al., 2020).目前植物固醇的分子功能研究相对较少.在植物中, 油菜素内酯(brassinosteroids, BRs)是一种重要的植物固醇类激素(Geuns, 1983), 通过与膜受体BRI1蛋白结合, 调控细胞的伸长、分裂和分化(Li and He, 2020).对动物与酵母的研究表明, 类固醇分子能够与鞘脂类结合形成一种称为脂筏(lipid rafts)的微区域, 介导细胞间的识别、脂膜和蛋白质的细胞内运输、信号转导、逆境反应和极性生长等多种生物学过程(Malinsky et al., 2013; Tapken and Murphy, 2015).植物中也存在类似的富含固醇和鞘脂的细胞膜微区域, 被称为变性剂不溶或者不敏感膜区(detergent-insoluble (resistant) membranes, DIMs/DRMs), 其中所含蛋白质可对外界刺激产生动态应答(Takahashi et al., 2013; Zauber et al., 2014).由此表明, 植物细胞膜上形成的固醇类富集的细胞膜微区域可能对植物发育和环境信号应答具有重要作用. ...
Membrane nanodomains in plants: capturing form, function, and movement 1 2015
... 固醇类也是细胞膜的结构成分之一, 作为信号物质在细胞膜或细胞核内对生物体的生长发育起至关重要的作用(Boutté and Grebe, 2009; Ferrer et al., 2017).有些固醇既存在于动物中, 也存在于植物中, 如胆固醇、谷甾醇和菜油甾醇(Tarkowská, 2019).人体细胞内胆固醇水平的变化能够引起心血管疾病、神经性疾病和相关癌症(Luo et al., 2020).目前植物固醇的分子功能研究相对较少.在植物中, 油菜素内酯(brassinosteroids, BRs)是一种重要的植物固醇类激素(Geuns, 1983), 通过与膜受体BRI1蛋白结合, 调控细胞的伸长、分裂和分化(Li and He, 2020).对动物与酵母的研究表明, 类固醇分子能够与鞘脂类结合形成一种称为脂筏(lipid rafts)的微区域, 介导细胞间的识别、脂膜和蛋白质的细胞内运输、信号转导、逆境反应和极性生长等多种生物学过程(Malinsky et al., 2013; Tapken and Murphy, 2015).植物中也存在类似的富含固醇和鞘脂的细胞膜微区域, 被称为变性剂不溶或者不敏感膜区(detergent-insoluble (resistant) membranes, DIMs/DRMs), 其中所含蛋白质可对外界刺激产生动态应答(Takahashi et al., 2013; Zauber et al., 2014).由此表明, 植物细胞膜上形成的固醇类富集的细胞膜微区域可能对植物发育和环境信号应答具有重要作用. ...
Plants are capable of synthesizing animal steroid hormones 1 2019
... 固醇类也是细胞膜的结构成分之一, 作为信号物质在细胞膜或细胞核内对生物体的生长发育起至关重要的作用(Boutté and Grebe, 2009; Ferrer et al., 2017).有些固醇既存在于动物中, 也存在于植物中, 如胆固醇、谷甾醇和菜油甾醇(Tarkowská, 2019).人体细胞内胆固醇水平的变化能够引起心血管疾病、神经性疾病和相关癌症(Luo et al., 2020).目前植物固醇的分子功能研究相对较少.在植物中, 油菜素内酯(brassinosteroids, BRs)是一种重要的植物固醇类激素(Geuns, 1983), 通过与膜受体BRI1蛋白结合, 调控细胞的伸长、分裂和分化(Li and He, 2020).对动物与酵母的研究表明, 类固醇分子能够与鞘脂类结合形成一种称为脂筏(lipid rafts)的微区域, 介导细胞间的识别、脂膜和蛋白质的细胞内运输、信号转导、逆境反应和极性生长等多种生物学过程(Malinsky et al., 2013; Tapken and Murphy, 2015).植物中也存在类似的富含固醇和鞘脂的细胞膜微区域, 被称为变性剂不溶或者不敏感膜区(detergent-insoluble (resistant) membranes, DIMs/DRMs), 其中所含蛋白质可对外界刺激产生动态应答(Takahashi et al., 2013; Zauber et al., 2014).由此表明, 植物细胞膜上形成的固醇类富集的细胞膜微区域可能对植物发育和环境信号应答具有重要作用. ...
[2] Use of oxygenated sterols to probe the regulation of 3-hydroxy-3-methylglutaryl-CoA reductase and sterologenesis 1 1985
... ORPs家族的第1个成员OSBP最初是作为氧化甾醇的受体被发现, 其通过抑制3-羟基-3-甲基戊二酰辅酶A还原酶(HMG-CoA)的活性抑制胆固醇的合成(Kandutsch and Thompson, 1980; Taylor et al., 1984; Taylor and Kandutsch, 1985).在不同物种中ORPs家族成员的数目不尽相同, 但功能域保守性很强.在人类细胞中, ORPs包含12个成员(Jaworski et al., 2001).老鼠细胞也包含12个成员(Anniss et al., 2002).在酵母中, ORPs包含7个成员(Beh et al., 2001).模式植物拟南芥(Arabidopsis thaliana)与人类和老鼠一样, 其ORPs包含12个成员(Skirpan et al., 2006).ORPs一般含有结合磷脂酰肌醇的PH蛋白结构域、介导二聚化的coiled-coil蛋白结构域、FFAT基序(two phenylalanine in an acidic tract motif)以及含高度保守的EQVSHHPP序列的可结合脂类的ORD蛋白结构域(OSBP-related domain). ...
Correlation between oxysterol binding to a cytosolic binding protein and potency in the repression of hydroxymethylglutaryl coenzyme a reductase 1 1984
... ORPs家族的第1个成员OSBP最初是作为氧化甾醇的受体被发现, 其通过抑制3-羟基-3-甲基戊二酰辅酶A还原酶(HMG-CoA)的活性抑制胆固醇的合成(Kandutsch and Thompson, 1980; Taylor et al., 1984; Taylor and Kandutsch, 1985).在不同物种中ORPs家族成员的数目不尽相同, 但功能域保守性很强.在人类细胞中, ORPs包含12个成员(Jaworski et al., 2001).老鼠细胞也包含12个成员(Anniss et al., 2002).在酵母中, ORPs包含7个成员(Beh et al., 2001).模式植物拟南芥(Arabidopsis thaliana)与人类和老鼠一样, 其ORPs包含12个成员(Skirpan et al., 2006).ORPs一般含有结合磷脂酰肌醇的PH蛋白结构域、介导二聚化的coiled-coil蛋白结构域、FFAT基序(two phenylalanine in an acidic tract motif)以及含高度保守的EQVSHHPP序列的可结合脂类的ORD蛋白结构域(OSBP-related domain). ...
Structure of Osh3 reveals a conserved mode of phosphoinositide binding in oxysterol-binding proteins 1 2013
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
Late endosomal transport and tethering are coupled processes controlled by RILP and the cholesterol sensor ORP1L 1 2013
... ANK蛋白结构域是一种特殊的锚蛋白重复结构域.目前, 只发现在人类ORP1和酵母Osh1p-Osh2p中含有ANK蛋白结构域.Osh1p的ANK蛋白结构域通过与核-液泡连接蛋白1 (nuclear-vacuole junction protein 1, NVJ1)互作使其定位在细胞核和液泡之间(Manik et al., 2017).全长ORP1L (ORP1 large)通过ANK蛋白结构域与GTP结合将Rab7蛋白定位于晚期核内体与溶酶体之间, 可以干扰内膜的转运过程(Johansson et al., 2005; van der Kant et al., 2013).ANK结构域在Osh2p中的具体功能还不清楚. ...
Membrane lipids: where they are and how they behave 2 2008
... 脂质在细胞膜上的分布不均一(van Meer et al., 2008).大多数脂质主要在内质网中合成, 随后被运输到各类细胞膜(质膜上的脂类最多), 以维持脂质的特定区域化(Moreau et al., 1998; van Meer et al., 2008).脂类具有不溶于水的疏水特性.脂质是如何从合成部位(内质网)转移到各种细胞器的? 细胞如何维持固醇分子在不同细胞器膜中的不均匀分布? 这些都是具有科学意义的重要问题.脂类分子的运输对此过程起到关键作用.胞内脂类运输主要包括3种机制: 囊泡运输、非囊泡运输及膜与膜之间的交融扩散.由LTPs参与的非囊泡运输是脂类运输的主要方式.LTPs主要有3方面功能.(1) 作为脂类的运输蛋白, 从一种膜撷取脂类运输到另一种膜.(2) 作为脂类的感受器, 通过与某种蛋白互作来结合或释放某种脂类.(3) 作为脂类的提供者, 如Sec14p (secretion 14 protein), 能够把磷脂酰肌醇运送至磷脂酰肌醇激酶 (Schaaf et al., 2008; Raychaudhuri and Prinz, 2010). LTPs是含有许多成员的大家族, 根据其脂质结合域可细分为SEC14、PITP (phosphatidylinositol transfer protein)、START (steroidogenic acute regulatory protein (StAR)-related transfer family)、GLTP (glycolipid transfer protein)、SCP-2 (sterol carrier protein 2)和OSBP/ORPs等家族(D'Angelo et al., 2008). ...
... ; van Meer et al., 2008).脂类具有不溶于水的疏水特性.脂质是如何从合成部位(内质网)转移到各种细胞器的? 细胞如何维持固醇分子在不同细胞器膜中的不均匀分布? 这些都是具有科学意义的重要问题.脂类分子的运输对此过程起到关键作用.胞内脂类运输主要包括3种机制: 囊泡运输、非囊泡运输及膜与膜之间的交融扩散.由LTPs参与的非囊泡运输是脂类运输的主要方式.LTPs主要有3方面功能.(1) 作为脂类的运输蛋白, 从一种膜撷取脂类运输到另一种膜.(2) 作为脂类的感受器, 通过与某种蛋白互作来结合或释放某种脂类.(3) 作为脂类的提供者, 如Sec14p (secretion 14 protein), 能够把磷脂酰肌醇运送至磷脂酰肌醇激酶 (Schaaf et al., 2008; Raychaudhuri and Prinz, 2010). LTPs是含有许多成员的大家族, 根据其脂质结合域可细分为SEC14、PITP (phosphatidylinositol transfer protein)、START (steroidogenic acute regulatory protein (StAR)-related transfer family)、GLTP (glycolipid transfer protein)、SCP-2 (sterol carrier protein 2)和OSBP/ORPs等家族(D'Angelo et al., 2008). ...
Sterol binding by OSBP- related protein 1L regulates late endosome motility and function 1 2011
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
Phosphatidylserine transport by ORP/Osh proteins is driven by phosphatidylinositol 4-phosphate 1 2015
... 在酵母细胞中, Osh1-3含有PH和ORD蛋白结构域, Osh4-7只含有ORD蛋白结构域(Raychaudhuri and Prinz, 2010).Osh1可以通过ANK蛋白结构域定位于细胞核与液泡之间的MCS, 转运磷脂和固醇, 进而促进细胞内脂质代谢(Levine and Munro, 2001; Manik et al., 2017).只有ORD蛋白结构域的Osh4p依然可通过形成PI4P浓度梯度在ER-PM之间运输固醇(de Saint-Jean et al., 2011; von Filseck et al., 2015b).与哺乳动物一样, 酵母细胞中固醇的转运也通过PI4P-磷酸酶Sac1水解PI4P形成一定浓度梯度来维持.Osh6p和Osh7p可以将磷脂酰丝氨酸从内质网转运到质膜(Maeda et al., 2013; von Filseck et al., 2015a). ...
A phosphatidylinositol-4-phosphate powered exchange mechanism to create a lipid gradient between membranes 2 2015
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
... 在酵母细胞中, Osh1-3含有PH和ORD蛋白结构域, Osh4-7只含有ORD蛋白结构域(Raychaudhuri and Prinz, 2010).Osh1可以通过ANK蛋白结构域定位于细胞核与液泡之间的MCS, 转运磷脂和固醇, 进而促进细胞内脂质代谢(Levine and Munro, 2001; Manik et al., 2017).只有ORD蛋白结构域的Osh4p依然可通过形成PI4P浓度梯度在ER-PM之间运输固醇(de Saint-Jean et al., 2011; von Filseck et al., 2015b).与哺乳动物一样, 酵母细胞中固醇的转运也通过PI4P-磷酸酶Sac1水解PI4P形成一定浓度梯度来维持.Osh6p和Osh7p可以将磷脂酰丝氨酸从内质网转运到质膜(Maeda et al., 2013; von Filseck et al., 2015a). ...
ORP2 delivers cholesterol to the plasma membrane in exchange for phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2) 1 2019
... (2) 植物不同ORPs蛋白的生理机制及其在发育过程中的作用.例如, 它们是否参与细胞自噬; 在动物和酵母中, LTPs在MCS通过介导脂类从内质网至自噬体膜的运输, 参与自噬体的形成(Ye et al., 2020).在低胆固醇条件下, 人类ORP1L通过与VAP-A相互作用形成内质网-自噬体接触位点, 介导自噬体的运输(Wijdeven et al., 2016).VAP27-1和VAP27-3已被证明参与植物内吞过程(Wang et al., 2016a; Stefano et al., 2018).VAP27-1也可以与拟南芥EH蛋白(AtEH1/Pan1和AtEH2/Pan2)互作, 调节自噬体的形成(Wang et al., 2019).然而植物ORPs是否通过VAP27参与细胞自噬过程仍需更多实验验证. ...
Molecular characterization of Osh6p, an oxysterol binding protein homolog in the yeast Saccharomyces cerevisiae 1 2005
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
Plant VAP27 proteins: domain characterization, intracellular localization and role in plant development 3 2016
... 拟南芥VAPs含有10个成员, 由于发现的第1个VAP分子量只有27 kDa, 故命名该家族为VAP27 (Nishimura et al., 1999).VAP27分为3个亚家族, 亚家族I包括5个成员(VAP27-1、VAP27-3、VAP27-5、VAP27-6和VAP27-7), 亚家族II包括3个成员(VAP27-8、VAP27-9和VAP27-10), 亚家族III包含2个成员(VAP27-2和VAP27-4).拟南芥VAPs同样含有MSD蛋白结构域、Coiled-coil蛋白结构域和跨膜区.VAP27蛋白主要定位在内质网和EPCS (Wang et al., 2016a).跨膜区的存在是VAPs定位在内质网的决定性区域.VAP27-1定位在EPCS还需要NET3C的介导(Wang et al., 2014).NET3C属于植物特异的肌动蛋白结合蛋白NET (networked)家族(Deeks et al., 2012).NET与肌动蛋白结合影响细胞骨架的运动, 因此VAP27蛋白可能通过NET来影响细胞骨架的运动. ...
... 在酵母细胞中, VAPs通过与ORPs互作介导脂类运输过程(Mesmin et al., 2013).但在植物中, 关于VAP27的功能及其作用机理的研究相对较少.已有研究表明, 拟南芥中鞘氨醇转移蛋白ACD11可以与VAP27-1相互作用(Petersen et al., 2009).VAP27- 1-GFP和VAP27-3-GFP转基因株系和功能缺失RNAi株系在根毛分叉表型上存在缺陷(Wang et al., 2016a).根毛分叉的表型与肌动蛋白互作蛋白AIP1突变体以及F-Actin2调节蛋白ROP2突变体的表型类似, 进一步说明VAP27可能通过调节肌动蛋白来调节根毛的发育(Guimil and Dunand, 2007; Ketelaar et al., 2007).VAP27-1和VAP27-3通过PIPs与网格蛋白的相互作用调节植物的内吞过程(Stefano et al., 2018).但是植物VAP27蛋白家族成员能否通过与ORP蛋白家族成员相互结合参与膜脂运输还未见报道. ...
... (2) 植物不同ORPs蛋白的生理机制及其在发育过程中的作用.例如, 它们是否参与细胞自噬; 在动物和酵母中, LTPs在MCS通过介导脂类从内质网至自噬体膜的运输, 参与自噬体的形成(Ye et al., 2020).在低胆固醇条件下, 人类ORP1L通过与VAP-A相互作用形成内质网-自噬体接触位点, 介导自噬体的运输(Wijdeven et al., 2016).VAP27-1和VAP27-3已被证明参与植物内吞过程(Wang et al., 2016a; Stefano et al., 2018).VAP27-1也可以与拟南芥EH蛋白(AtEH1/Pan1和AtEH2/Pan2)互作, 调节自噬体的形成(Wang et al., 2019).然而植物ORPs是否通过VAP27参与细胞自噬过程仍需更多实验验证. ...
OSBP is a cholesterol-regulated scaffolding protein in control of ERK1/2 activation 1 2005
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
Protein trafficking during plant innate immunity 2 2016
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
... (3) 植物ORPs是否参与植物免疫及对外界环境因子的应激反应.磷脂是磷脂双分子层的重要组成部分, 在信号转导中发挥重要作用(Takáč et al., 2019).磷脂的膜分布可影响膜的弯曲, 进而调节膜运输, 此过程对植物免疫应答反应至关重要(Wang et al., 2016b; De Craene et al., 2017).近期有综述阐明基于磷脂的信号转导和膜转运与植物免疫有密切联系(Xing et al., 2021).因此, 探讨脂类运输蛋白ORPs是否通过膜脂运输影响植物应对病原菌侵害和不适环境变化具有重要意义. ...
OSBP-related protein 3 (ORP3) coupling with VAMP-associated protein A regulates R-Ras activity 2 2015
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
... VAPs是一类在真核生物中非常保守的囊泡相关膜蛋白(Nishimura et al., 1999).动物VAPs家族成员众多且分布广泛, 主要通过FFAT基序定位在内质网和内质网与其它膜形成的MCS (Murphy and Levine, 2016).VAPs一般含有MSP蛋白结构域(major sperm protein)、Coiled-coil结构域和C端跨膜结构域(C-terminal transmembrane domain, TMD).尚未在酵母中发现可以形成二聚体的Coiled-coil结构域或其它类似结构域.VAPs主要与3类蛋白互作: 突触囊泡SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptors)、病毒性蛋白和含有FFAT基序的蛋白(Lev et al., 2008).VAPs与含有FFAT基序的蛋白互作发现于Scs2p与Opi1p相互关系的研究(Loewen et al., 2003).通过FFAT基序与VAPs互作的脂类运输相关蛋白主要包括ORPs、CERT (ceramide transport protein)和Nirs (PYK2 N-terminal domain-interacting receptors) 3个蛋白家族(Wyles and Ridgway, 2004; Amarilio et al., 2005; Kawano et al., 2006).除了典型的FFAT基序, ORP3还含有2个FFAT-like基序(HFFSGST和NYSDGSE), 介导与VAPs的互作(Weber-Boyvat et al., 2015a, 2015b). ...
Cholesterol and ORP1L- mediated ER contact sites control autophagosome transport and fusion with the endocytic pathway 1 2016
... (2) 植物不同ORPs蛋白的生理机制及其在发育过程中的作用.例如, 它们是否参与细胞自噬; 在动物和酵母中, LTPs在MCS通过介导脂类从内质网至自噬体膜的运输, 参与自噬体的形成(Ye et al., 2020).在低胆固醇条件下, 人类ORP1L通过与VAP-A相互作用形成内质网-自噬体接触位点, 介导自噬体的运输(Wijdeven et al., 2016).VAP27-1和VAP27-3已被证明参与植物内吞过程(Wang et al., 2016a; Stefano et al., 2018).VAP27-1也可以与拟南芥EH蛋白(AtEH1/Pan1和AtEH2/Pan2)互作, 调节自噬体的形成(Wang et al., 2019).然而植物ORPs是否通过VAP27参与细胞自噬过程仍需更多实验验证. ...
Lipid transfer proteins: the lipid commute via shuttles, bridges and tubes 1 2019
... 细胞膜包括细胞质膜, 以及细胞核、线粒体、质体、内质网(endoplasmic reticulum, ER)、高尔基体、液泡等细胞器膜和胞内运输所需要的各种类型囊泡膜所组成的膜系统.脂质是细胞膜的基本组分, 包含脂肪(fats)和类脂(lipids).类脂又分为磷脂(phospholipids)、糖脂(glycolipids)、脂蛋白(lipoproteins)和固醇类(steroid).不同物种、不同类型的细胞以及不同细胞器的细胞膜所含脂质的种类、分布和含量都不尽相同.不同的脂质分子在细胞生命活动过程中起着非常重要的作用.脂质不仅是细胞膜的结构成分, 而且参与能量储存以及维持细胞膜的流动性和通透性, 特别是作为信号转导的关键组分参与生物体的个体发育和生理过程.然而, 膜脂信号在胞内的传送需要脂类运输蛋白(lipid transfer proteins, LTPs)的充分配合.正是由于LTPs在胞内信号转导中具有不可或缺的作用, 其生理功能备受研究者的关注, 相关研究在近几年也取得了重要进展(Pietrangelo and Ridgway, 2018; Balla et al., 2019; Wong et al., 2019). ...
Vesicle- associated membrane protein-associated protein-A (VAP- A) interacts with the oxysterol-binding protein to modify export from the endoplasmic reticulum 1 2002
... VAPs参与调控许多细胞内的生物过程, 如膜运输、脂类代谢和运输以及微管的组织(Soussan et al., 1999; Wyles et al., 2002; Kagiwada and Zen, 2003; Kawano et al., 2006).Scs2是酵母中很重要的VAPs之一.Scs2p参与脂类感知、磷脂代谢、脂类运输以及未折叠蛋白反应(unfolded protein response, UPR)和端粒沉默等过程(Lev et al., 2008).Scs2p可与PI4P和PI(4,5)P2结合, 磷酸肌醇含量的增加可抑制Scs2p与Opi1p的结合(Kagiwada and Hashimoto, 2007).Scs2p功能缺失抑制肌醇-1-磷酸合成酶基因INO1的表达, 降低磷脂酰肌醇的合成, 促进磷脂酰胆碱的合成(Kagiwada and Zen, 2003).VAPs与LTPs互作介导脂类运输是VAPs的重要功能.Scs2p可与酵母Osh1p、Osh2p和Osh3p互作介导MCS之间脂类的运输(Loewen et al., 2003).与此类似, 人类VAP-A与OSBP蛋白互作使其定位在内质网, 介导MCS间胆固醇的运输(Mesmin et al., 2013).VAPs功能缺失引起高尔基体PI4P水平升高, 使高尔基体产生的内吞体增加, 最终导致产生非正常溶酶体和自噬体(Mao et al., 2019).人类VAPs功能缺失可引起一系列神经性疾病.例如, VAP-BP56S的突变引起罕见的遗传性运动神经元疾病脊髓性肌萎缩和肌萎缩侧索硬化(Nishimura et al., 2004; Chen et al., 2010; Kabashi et al., 2013); VAP-B∆V25的突变引起帕金森病(Kun-Rodrigues et al., 2015).VAP-B的突变导致内质网不能栓系到高尔基体上, 引发高尔基体PI4P水平升高, 继而导致非功能溶酶体的积累, 使正常自噬溶酶体的降解速率下降(Mao et al., 2019). ...
Characterization of the sterol-binding domain of oxysterol-binding protein (OSBP)-related protein 4 reveals a novel role in vimentin organization 2 2007
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
VAMP-associated protein-A regulates partitioning of oxysterol-binding protein-related protein-9 between the endoplasmic reticulum and Golgi apparatus 1 2004
... VAPs是一类在真核生物中非常保守的囊泡相关膜蛋白(Nishimura et al., 1999).动物VAPs家族成员众多且分布广泛, 主要通过FFAT基序定位在内质网和内质网与其它膜形成的MCS (Murphy and Levine, 2016).VAPs一般含有MSP蛋白结构域(major sperm protein)、Coiled-coil结构域和C端跨膜结构域(C-terminal transmembrane domain, TMD).尚未在酵母中发现可以形成二聚体的Coiled-coil结构域或其它类似结构域.VAPs主要与3类蛋白互作: 突触囊泡SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptors)、病毒性蛋白和含有FFAT基序的蛋白(Lev et al., 2008).VAPs与含有FFAT基序的蛋白互作发现于Scs2p与Opi1p相互关系的研究(Loewen et al., 2003).通过FFAT基序与VAPs互作的脂类运输相关蛋白主要包括ORPs、CERT (ceramide transport protein)和Nirs (PYK2 N-terminal domain-interacting receptors) 3个蛋白家族(Wyles and Ridgway, 2004; Amarilio et al., 2005; Kawano et al., 2006).除了典型的FFAT基序, ORP3还含有2个FFAT-like基序(HFFSGST和NYSDGSE), 介导与VAPs的互作(Weber-Boyvat et al., 2015a, 2015b). ...
Coordination of phospholipid-based signaling and membrane trafficking in plant immunity 2 2021
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
... (3) 植物ORPs是否参与植物免疫及对外界环境因子的应激反应.磷脂是磷脂双分子层的重要组成部分, 在信号转导中发挥重要作用(Takáč et al., 2019).磷脂的膜分布可影响膜的弯曲, 进而调节膜运输, 此过程对植物免疫应答反应至关重要(Wang et al., 2016b; De Craene et al., 2017).近期有综述阐明基于磷脂的信号转导和膜转运与植物免疫有密切联系(Xing et al., 2021).因此, 探讨脂类运输蛋白ORPs是否通过膜脂运输影响植物应对病原菌侵害和不适环境变化具有重要意义. ...
Function and regulation of phospholipid signaling in plants 1 2009
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
Oxysterol binding protein induces upregulation of SREBP- 1c and enhances hepatic lipogenesis 1 2007
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
OSBP-related protein 8 (ORP8) suppresses ABCA1 expression and cholesterol efflux from macrophages 2 2008
... 氧化固醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)是一类重要的LTPs, 在各类生物中广泛存在, 并含有众多家族成员(Skirpan et al., 2006).许多研究表明, OSBP/ ORPs不仅可以运输脂类, 还参与囊泡运输(Dong et al., 2016)、脂质代谢(Lagace et al., 1997; Beh et al., 2001; Beh and Rine, 2004; Ma et al., 2010)、信号转导(Wang et al., 2005b; Lessmann et al., 2007)、细胞骨架动力学(Johansson et al., 2007; Wyles et al., 2007)和基因表达等生理过程(Yan et al., 2007, 2008).尽管近2年ORPs相关研究在人类(Homo sapiens)和酵母(Saccharomyces cerevisiae)中取得了重要进展, 我们对植物中ORPs的生物学功能研究仍不充分, 对其认识较为模糊.揭示植物中ORPs的分子功能将会在生物化学、细胞学、生理学和发育学层面拓宽我们对膜蛋白信号转导的认识, 因此, 本文从8个方面对ORP蛋白家族成员的研究进展进行阐述. ...
... FFAT基序的氨基酸序列一般为-EFFDAXE-, 是内质网定位的决定性区域.人类细胞中除了ORP5和ORP8, 其它成员都含有FFAT基序.酵母Osh1-3含有FFAT基序(Raychaudhuri and Prinz, 2010).FFAT基序能够与VAPs (vesicle-associated membrane protein (VAMP)-associated proteins)蛋白互作, 而VAPs是一类内质网定位的囊泡运输相关蛋白(Skehel et al., 1995), 因此FFAT基序能够指导ORPs定位到内质网上(Loewen et al., 2003).但FFAT基序介导并不是ORPs定位到内质网上的唯一方式.例如, ORP5和ORP8虽然没有FFAT基序, 但是可通过C端的跨膜区定位到内质网膜(Yan et al., 2008; Du et al., 2011).Osh4p没有PH蛋白结构域和FFAT基序, 依然在内质网-质膜(plasma membrane) (ER-PM)上定位(Schulz et al., 2009).目前, 在植物界还未见FFAT基序或相似基序的功能报道. ...
Membrane contact sites and organelles interaction in plant autophagy 1 2020
... (2) 植物不同ORPs蛋白的生理机制及其在发育过程中的作用.例如, 它们是否参与细胞自噬; 在动物和酵母中, LTPs在MCS通过介导脂类从内质网至自噬体膜的运输, 参与自噬体的形成(Ye et al., 2020).在低胆固醇条件下, 人类ORP1L通过与VAP-A相互作用形成内质网-自噬体接触位点, 介导自噬体的运输(Wijdeven et al., 2016).VAP27-1和VAP27-3已被证明参与植物内吞过程(Wang et al., 2016a; Stefano et al., 2018).VAP27-1也可以与拟南芥EH蛋白(AtEH1/Pan1和AtEH2/Pan2)互作, 调节自噬体的形成(Wang et al., 2019).然而植物ORPs是否通过VAP27参与细胞自噬过程仍需更多实验验证. ...
Genome-wide analysis of membrane targeting by S. cerevisiae pleckstrin homology domains 1 2004
... 由于PH蛋白结构域与磷酸肌醇结合具有特异性, PH-GFP常用来指示特定磷酸肌醇的定位(Balla et al., 2000; Rusten and Stenmark, 2006).ORP5和ORP8的PH蛋白结构域通过与N端相邻的多碱基序列识别PI4P和PI(4,5)P2而将其定位于质膜(Ghai et al., 2017).虽然磷酸肌醇有助于PH蛋白结构域结合到膜上, 但是它们之间的定位不特异.例如, Osh1p通过与PI4P互作定位到高尔基体膜, Osh2p通过与PI4P互作不仅定位在高尔基体膜, 也定位在质膜(Roy and Levine, 2004; Yu et al., 2004).因此, ORPs的特异性定位不仅依赖于PH蛋白结构域, 还需要多种因子的参与.例如, OSBP定位在高尔基体膜不仅依赖于PH蛋白结构域与高尔基体膜上PI4P的结合, 还需要ARF (GTPase ADP-ribosylation factor)蛋白的参与(Levine and Munro, 2002; Godi et al., 2004). ...
Plasma membrane lipid-protein interactions affect signaling processes in sterol-biosynthesis mutants in Arabidopsis thaliana 1 2014
... 固醇类也是细胞膜的结构成分之一, 作为信号物质在细胞膜或细胞核内对生物体的生长发育起至关重要的作用(Boutté and Grebe, 2009; Ferrer et al., 2017).有些固醇既存在于动物中, 也存在于植物中, 如胆固醇、谷甾醇和菜油甾醇(Tarkowská, 2019).人体细胞内胆固醇水平的变化能够引起心血管疾病、神经性疾病和相关癌症(Luo et al., 2020).目前植物固醇的分子功能研究相对较少.在植物中, 油菜素内酯(brassinosteroids, BRs)是一种重要的植物固醇类激素(Geuns, 1983), 通过与膜受体BRI1蛋白结合, 调控细胞的伸长、分裂和分化(Li and He, 2020).对动物与酵母的研究表明, 类固醇分子能够与鞘脂类结合形成一种称为脂筏(lipid rafts)的微区域, 介导细胞间的识别、脂膜和蛋白质的细胞内运输、信号转导、逆境反应和极性生长等多种生物学过程(Malinsky et al., 2013; Tapken and Murphy, 2015).植物中也存在类似的富含固醇和鞘脂的细胞膜微区域, 被称为变性剂不溶或者不敏感膜区(detergent-insoluble (resistant) membranes, DIMs/DRMs), 其中所含蛋白质可对外界刺激产生动态应答(Takahashi et al., 2013; Zauber et al., 2014).由此表明, 植物细胞膜上形成的固醇类富集的细胞膜微区域可能对植物发育和环境信号应答具有重要作用. ...
OSBP-related protein 11 (ORP11) dimerizes with ORP9 and localizes at the Golgi- late endosome interface 1 2010
... Subcellular localization and function of ORPs in animal and yeast Table 1
蛋白
膜结合位点
运输脂类
参考文献
OSBP
ER (endoplasmic reticulum)- trans-Golgi
Cholesterol, PI4P
Mesmin et al., 2013
ORP1
ER-late endosome/lysomes
Cholesterol
Rocha et al., 2009; Kobuna et al., 2010; Vihervaara et al., 2011
ORP2
ER-lipid droplet, ER-PM (plasma membrane)
Cholesterol, PI(4,5)P2
Hynynen et al., 2009; Jansen et al., 2011; Weber-Boyvat et al., 2015b; Wang et al., 2019
ORP3
ER-PM
PI4P
Weber-Boyvat et al., 2015a; Gulyás et al., 2020
ORP4
Unknown
Cholesterol
Wyles et al., 2007
ORP5
ER-PM, ER-late endosome?
PS, PI4P, cholesterol, PI(4,5)P2
Du et al., 2011; Chung et al., 2015; Ghai et al., 2017
ORP6
ER-PM
PI4P
Mochizuki et al., 2018
ORP7
Unknown
Unknown
None
ORP8
ER-PM, ER-mitochondrion
PS, PI4P
Chung et al., 2015; Ghai et al., 2017
ORP9
ER-trans-Golgi
Cholesterol, PI4P
Ngo and Ridgway, 2009; Liu and Ridgway, 2014
ORP10
Unknown
PS
Maeda et al., 2013
ORP11
trans-Golgi-late endosome
Unknown
Zhou et al., 2010
Osh1p
Nucleus-vacuole junction
Cholesterol, ergosterol, PI4P
Levine and Munro, 2001; Schulz et al., 2009
Osh2p
ER-PM
Cholesterol
Schulz et al., 2009
Osh3p
ER-PM
PI4P, Cholesterol
Schulz et al., 2009; Tong et al., 2013
Osh4p
ER-PM, ER-mitochondrion
Sterol, PI4P, PI(4,5) P2
Raychaudhuri et al., 2006; Schulz et al., 2009; de Saint-Jean et al., 2011; von Filseck et al., 2015b
ATG9 regulates autophagosome progression from the endoplasmic reticulum in Arabidopsis 1 2017
... 磷脂分子参与调控许多生理过程, 包括根的发育、花粉发育、微管发育、激素信号、膜脂运输、自噬反应、植物免疫以及一些生物和非生物胁迫反应(Xue et al., 2009; Harayama and Riezman, 2018; Jia et al., 2019; Xing et al., 2021).在植物中, 免疫系统依赖膜脂运输, 将防御相关因子转运到特定区域(Wang et al., 2016b), 而磷脂分子对于膜脂运输至关重要(Liu et al., 2013).例如, PI(4,5)P2是囊泡运输过程中的关键调控因子之一(Martin, 2015; De Craene et al., 2017).磷酸肌醇也参与细胞内自噬体的形成(Chung, 2019).自噬相关基因(autophagy related genes, ATG) VPS34 (vacuolar protein sorting 34)影响自噬小体发生部位PtdIns3P的产生.ATG9和ATG18a为植物细胞内质网自噬体形成所必需, ATG9依赖PtdIns3P调控ATG18a在自噬体膜上的转运(Zhuang et al., 2017).磷酸肌醇也是离子通道的重要调节因子.PI(4,5)P2可通过调节K+通道调节气孔运动(Ma et al., 2009). ...
 ,2,*
,2,*
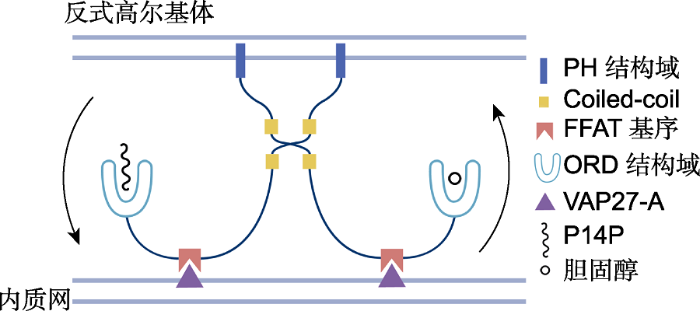 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT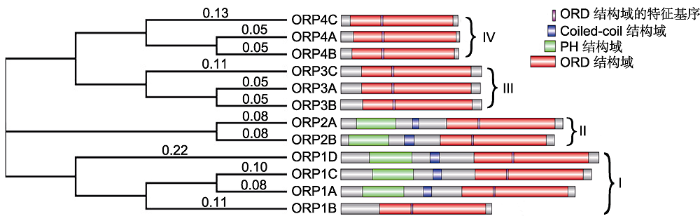 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}