Abstract To provide genetic information suitable for improving stigma-related traits of genic male sterile lines in rice. We investigated four stigma-related traits of 7001S (japonica genic male sterile line, P1), Z913S (indica genic male sterile line, P2), F1 (P1×P2), F2 and F2:3. Coefficients of linear correlation among four stigma-related traits were calculated, and genetic patterns were analyzed using mixed major-gene plus polygene inheritance models. We found an extremely significant linear positive correlation among four stigma traits, and the correlation coefficients were between 0.274 and 0.897. Genetic analysis revealed that, style length (SYL) and percentage of exserted stigma (PES) were controlled by two major genes with additive-dominant effects and one major gene with additive-equal-negative-dominant effects plus polygenes in F2:3, respectively. Stigma length (STL), SYL (except F2:3), the sum of stigma and style length (TSSL), and PES (except F2:3) were controlled by two major genes plus polygenes in F2 and F2:3. The two major genes expressed additive-dominant-epistatic effects but expressed additive-dominant effects of SYL in the F2:3 populations. Four stigma traits were mainly governed by major genes in two generations. Keywords:rice;genic male sterile line;stigma trait;major gene plus polygene model;genetic analysis
PDF (2225KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 江建华, 党小景, 姚文豪, 胡梦竹, 王雨停, 胡长敏, 张瑛, 王德正. 水稻核不育系4个柱头性状的遗传分析. 植物学报, 2021, 56(3): 284-295 doi:10.11983/CBB20200 Jiang Jianhua, Dang Xiaojing, Yao Wenhao, Hu Mengzhu, Wang Yuting, Hu Changmin, Zhang Ying, Wang Dezheng. Genetic Analysis of Four Stigma Traits with Genic Male Sterile Line in Rice (Oryza sativa). Chinese Bulletin of Botany, 2021, 56(3): 284-295 doi:10.11983/CBB20200
水稻(Oryza sativa)是最主要的粮食作物, 世界一半以上的人口以稻米为主食(袁隆平, 2010)。始于我国20世纪70年代的杂交水稻较同熟期常规稻增产20%以上, 杂交水稻的推广种植使水稻生产力大幅提升(Qian et al., 2016)。当前, 我国杂交水稻年种植面积约1.67×107 hm2, 以杂交籼稻为主, 而杂交粳稻的种植面积不足杂交水稻种植面积的3% (曹黎明等, 2018)。制种产量低是制约杂交粳稻发展的重要因素, 提高粳型不育系的异交结实率是提高杂交粳稻制种产量的关键环节(杨振玉等, 2016)。异交结实率主要取决于水稻花器性状和开花习性, 花器性状主要包括柱头长度(stigma length, STL)、花柱长度(style length, SYL)、柱头外露率(percentage of exserted stigma, PES)和开颖角度(glume opening angle, GOA)等(计慎敏和张大兵, 2007; 罗伟雄等, 2011)。廖佩言等(1986)研究发现, 柱头-花柱总长度(以下简称柱花总长度) (the sum of stigma and style length, TSSL)和柱头单外露率对异交结实率的贡献率分别为17.86%和7.9%。杨保汉(1997)研究表明, 柱头外露率和制种产量呈极显著正相关, 不育系的柱头外露率每增加1个百分点, 其异交结实率可提高0.74%- 0.92%, 制种产量可增加47-68 kg?hm-2。可见, 柱头性状与水稻异交结实率密切相关。
(A) 7001S和Z913S的柱头长度比较; (B) 水稻雌蕊各部位名称(STL: 柱头长度; SYL: 花柱长度; TSSL: 柱花总长度); (C) 水稻柱头无外露、单外露以及双外露的表型; (D) Z913S田间柱头外露表型。(A), (B) Bars=1 mm; (C), (D) Bars=1 cm Figure 1Phenotypes of pistil and stigma of 7001S and Z913S
(A) Stigma morphology of 7001S and Z913S; (B) Names of rice pistil parts defined in this study (STL: Stigma length; SYL: Style length; TSSL: The sum of stigma and style length); (C) Phenotype of single, dual, and no stigma exsertion in a spikelet; (D) Phenotype of exserted stigma in Z913S. (A), (B) Bars=1 mm; (C), (D) Bars=1 cm
Table 1 表1 表1水稻4个柱头性状间的相关性系数 Table 1Correlation coefficients among four stigma traits of rice
Traits
Stigma length
Style length
The sum of stigma and style length
Percentage of exserted stigma
Stigma length
-
0.432**
0.897**
0.298**
Style length
0.526**
-
0.847**
0.320**
The sum of stigma and style length
0.792**
0.893**
-
0.365**
Percentage of exserted stigma
0.386**
0.274**
0.383**
-
Lower left triangle and upper right triangle of “-” indicates F2 and F2:3, respectively; ** indicate significant differences at 1% level. “-”的左下角和右上角分别表示F2和F2:3的相关性系数; **表示在1%水平差异显著。
(A) 柱头长度; (B) 花柱长度; (C) 柱花总长度; (D) 柱头外露率。** 表示在1%水平差异显著。 Figure 3Phenotypic evaluations of four stigma traits in two environments of rice 7001S and Z913S
(A) Stigma length; (B) Style length; (C) The sum of stigma and style length; (D) Percentage of exserted stigma. ** indicate significant differences at 1% level.
Table 3 表3 表3水稻7001S、Z913S和7001S/Z913S F1 4个柱头性状的联合方差分析 Table 3Joint analysis of variance for four stigma traits of rice 7001S, Z913S, and 7001S/Z913S F1
Traits
Source of variation
Degrees of freedom
Sum of squares
Mean square
F value
F0.05
F0.01
Stigma length
Genotypes
2
6.34
3.17
876.00**
3.10
4.85
Environments
1
0.02
0.02
6.83*
3.95
6.93
Genotype × Environment
2
0.01
0.01
1.51
3.10
4.85
Style length
Genotypes
2
1.10
0.55
86.84**
3.10
4.85
Environments
1
0.08
0.08
12.93**
3.95
6.93
Genotype × Environment
2
0.04
0.02
3.19
3.10
4.85
The sum of stigma and style length
Genotypes
2
12.67
6.33
1174.95**
3.10
4.85
Environments
1
0.20
0.20
36.45**
3.95
6.93
Genotype × Environment
2
0.01
0.00
0.86
3.10
4.85
Percentage of exserted stigma
Genotypes
2
16677.93
8338.96
278.09**
3.10
4.85
Environments
1
136.20
136.20
4.54*
3.95
6.93
Genotype × Environment
2
327.42
163.71
5.46**
3.10
4.85
*和**分别表示在P<0.05和P<0.01水平差异显著。* and ** indicate significant differences at P<0.05 and P<0.01, respectively.
Table 4 表4 表4水稻4个柱头相关性状备选模型配合表型分布的极大似然函数值(MLV)和AIC值(IECM算法) Table 4Max log likelihood value (MLV) and Akaike’s Information Criterion (AIC) values of candidate models calculated with IECM method for four stigma related traits of rice
Trait
Environment
Candidate model
MLV value
AIC value
Test of goodness-of-fita
Stigma length
E1
PG-ADI
246.62
-481.23
0/0/0/0/0
MX1-AD-ADI
251.95
-487.89
0/0/0/0/0
MX2-ADI-ADI
257.05
-490.09
0/0/0/0/0
MX2-ADI-AD
256.33
-494.66
0/0/0/0/0
E2
PG-ADI
210.90
-409.81
0/0/0/1/0
MX1-AD-ADI
210.90
-407.80
0/0/0/1/0
MX2-ADI-AD
211.65
-397.30
0/0/0/0/0
MX2-AD-AD
207.63
-397.25
0/0/0/0/0
Style length
E1
MX2-ADI-ADI
322.90
-621.79
0/0/2/0/0
MX2-ADI-AD
322.69
-627.37
0/0/2/0/0
MX2-AD-AD
316.56
-623.12
0/1/1/1/0
MX2-EEAD-AD
313.03
-622.05
0/1/1/1/0
E2
1MG-A
107.98
-211.96
1/1/0/1/0
2MG-AD
111.92
-211.84
0/0/0/0/0
2MG-A
111.21
-214.41
0/0/0/0/0
MX1-A-AD
112.06
-214.11
0/0/0/0/0
The sum of stigma and style length
E1
2MG-ADI
90.19
-158.38
0/0/0/1/0
MX1-AD-ADI
92.18
-168.37
0/0/0/1/0
MX2-ADI-ADI
97.25
-170.51
0/0/0/1/0
MX2-ADI-AD
95.63
-173.27
0/0/0/1/0
E2
2MG-EEAD
-12.03
32.05
0/0/1/0/0
MX1-A-AD
-11.29
32.59
1/1/1/1/0
MX2-AD-AD
-2.11
22.23
0/0/0/0/0
MX2-EEAD-AD
-7.10
26.19
0/0/0/0/0
Percentage of exserted stigma
E1
MX2-ADI-ADI
-1538.19
3100.38
0/0/2/0/0
MX2-ADI-AD
-1538.18
3094.36
0/0/2/0/0
MX2-AD-AD
-1540.16
3090.32
0/0/2/0/0
MX2-EEAD-AD
-1545.89
3095.79
0/0/1/0/0
E2
2MG-AD
-1425.36
2862.71
0/0/2/0/0
2MG-A
-1426.26
2860.51
0/0/2/1/0
MX1-AD-AD
-1423.53
2861.07
0/0/2/0/0
MX1-AEND-AD
-1425.67
2861.34
0/0/2/0/0
a Five digitals in test of goodness-of-fit column denote numbers of significant statistic parameters among U12, U22, U32, nW2, and Dn. MG: Major gene model; PG: Polygene model; MX: Mixed major gene and polygene model; A: Additive effect; D: Dominance effect; I: Epistasis; E: Equal; N: Negative. Bold letter indicates the optimal model. a 适合性检验中5个数字分别指在U12、U22、U32、nW2和Dn适合性检验中达到显著水平的统计量个数。MG: 主基因模型; PG: 多基因模型; MX: 主基因+多基因模型; A: 加性效应; D: 显性效应; I: 上位性; E: 等效; N: 负向。粗体表示最适模型。
(A), (B) F2和F2:3群体中柱头长度分布; (C), (D) F2和F2:3群体中花柱长度分布; (E), (F) F2和F2:3群体中柱花总长度分布; (G), (H) F2和F2:3群体中柱头外露率分布 Figure 4Frequency distribution, fitted mixed distribution and its component distribution for four stigma traits of F2 and F2:3 populations in the cross of rice 7001S/Z913S
(A), (B) Distribution in F2 and F2:3 populations of stigma length; (C), (D) Distribution in F2 and F2:3 populations of style length; (E), (F) Distribution in F2 and F2:3 populations of the sum of stigma and style length; (G), (H) Distribution in F2 and F2:3 populations of percentage of exserted stigma
Table 5 表5 表5水稻4个柱头性状遗传参数估计值 Table 5Estimates of genetic parameters for four stigma traits of rice
Genetic parameter
Trait
Stigma length
Style length
The sum of stigma and style length
Percentage of exserted stigma
E1
E2
E1
E2
E1
E2
E1
E2
MX2-ADI- AD
MX2-ADI- AD
MX2-ADI- AD
2MG-AD
MX2-ADI- AD
MX2-EEAD-AD
MX2-EEAD-AD
MX1-AEND-AD
Univalent parameter
da(d)
0.06
0.002
0.11
0.05
0.14
-0.26
-10.61
-17.87
db
0.06
0.001
0.03
-0.20
0.02
-
-
-
ha(h)
-0.12
-0.05
-0.12
0.04
-0.29
-
-
-
hb
-0.10
-0.05
-0.06
-0.005
-0.21
-
-
-
i
0.12
0.02
0.05
-
0.17
-
-
-
jab
0.09
0.002
0.02
-
0.10
-
-
-
jba
-0.06
0.001
-0.02
-
-0.01
-
-
-
l
0.13
0.36
0.08
-
0.24
-
-
-
[d]
-0.45
-0.30
-0.24
-
-0.59
0.06
3.01
-10.51
[h]
0.20
-0.24
0.17
-
0.45
0.54
24.67
0.14
Bivalent parameter
σ2p
0.020
0.022
0.014
0.040
0.053
0.089
232.04
325.28
σ2mg
0.013
0.014
0.010
0.033
0.039
0.082
155.72
216.11
σ2pg
0.005
0.005
-
-
0.010
-
40.91
80.75
h2mg (%)
65.00
63.64
71.43
82.50
73.58
92.13
67.11
66.44
h2pg (%)
25.00
22.73
-
-
18.87
-
17.63
24.82
d: Main gene additive effect value; da: Additive effect of the first pair of major genes; db: Additive effect of the second pair of major genes; h: Main gene dominant effect value; ha: Dominant effect of the first pair of major genes; hb: Dominant effect of the second pair of major genes; i: Epistatic effect value between da and db; jab: Epistatic effect value between da and hb; jba: Epistatic effect value between ha and db; l: Epistatic effect value between ha and hb; [d]: Additive effect of polygene; [h]: Dominant effect of polygene; σ2p: Phenotypic variance; σ2mg: Major gene variance; σ2pg: Polygene variance; h2mg(%): Heritability of major gene; h2pg(%): Heritability of polygene. - indicates the value is absent. d: 主基因加性效应值; da: 第1对主基因的加性效应; db: 第2对主基因的加性效应; h: 主基因显性效应值; ha: 第1对主基因的显性效应; hb: 第2对主基因的显性效应; i: 加性×加性互作; jab: 加性×显性互作; jba: 显性×加性互作; l: 显性×显性互作; [d]: 多基因加性效应; [h]: 多基因显性效应; σ2p: 表型方差; σ2mg: 主基因方差; σ2pg: 多基因方差; h2mg(%): 主基因遗传率; h2pg(%): 多基因遗传率。-表示空缺。
BaktiC, TanakaJ (2019). Detection of dominant QTLs for stigma exsertion ratio in rice derived from Oryza rufipogon accession ‘W0120’ Breed Sci 69, 143-150. DOI:10.1270/jsbbs.18139URL [本文引用: 2]
DangXJ, LiuEB, LiangYF, LiuQM, BreriaCM, HongDL (2016). QTL detection and elite alleles mining for stigma traits in Oryza sativa by association mapping Front Plant Sci 7, 1188. [本文引用: 8]
LiuQM, QinJC, LiTW, LiuEB, FanDJ, EdzesiWM, LiuJH, JiangJH, LiuXL, XiaoLJ, LiuLL, HongDL (2015). Fine mapping and candidate gene analysis of qSTL3, a stigma length-conditioning locus in rice(Oryza sativa L.) PLoS One 10, e0127938. DOI:10.1371/journal.pone.0127938URL [本文引用: 3]
LiuY, ZhangAN, WangFM, KongDY, LiMS, BiJG, ZhangFY, WangJH, LuoXX, PanZQ, YuXQ, LiuGL, LuoLJ (2019). Fine mapping a quantitative trait locus, qSER-7, that controls stigma exsertion rate in rice(Oryza sativa L.) Rice 12, 46. DOI:10.1186/s12284-019-0304-zPMID:31289958 [本文引用: 4] Stigma exsertion rate (SER) is a key determinant of outcrossing in hybrid rice seed production. A quantitative trait locus (QTL) for stigma exsertion rate in rice, qSER-7, has previously been detected on chromosome 7 by using a F population derived from two indica cytoplasmic male sterility (CMS) maintainers, Huhan 1B and II-32B.The chromosomal location of qSER-7 was precisely delimited by fine-scale mapping. Near-isogenic lines (NILs) were established, one of which isolated the locus in the qSER-7 line, which contains an introgressed segment of II-32B in the Huhan 1B genetic background, and exhibits a significantly higher stigma exsertion rate than that of the recurrent parent. Using 3192 individuals from the BCF segregation population, the QTL qSER-7 was narrowed down to a 28.4-kb region between the markers RM3859 and Indel4373 on chromosome 7. According to the rice genome annotation database, three genes were predicted within the target region. Real-time PCR analysis showed significantly higher expression levels of LOC_Os07g15370 and LOC_Os07g15390 in II-32B than in Huhan 1B. LOC_Os07g15370(OsNRAMP5) was a previously reported gene for Mn and Cd transporter. The stigma exertion rates of OsNRAMP5-overexpressing plants were significantly higher than that of wild type plants, in contrast, a T-DNA insertion mutant osnramp5 showed a lower stigma exertion rate.In the present study, the QTL qSER-7 was isolated to a region between the markers RM3859 and Indel4373. Two candidate genes were selected based on the expression difference between the two parents, which can facilitate the further cloning of the gene underlying the quantitative trait associated with qSER-7 as well as the marker-assisted transfer of desirable genes for stigma exsertion rate improvement in rice.
MarathiB, RamosJ, HechanovaSL, OaneRH, JenaKK (2015). SNP genotyping and characterization of pistil traits revealing a distinct phylogenetic relationship among the species of Oryza Euphytica 201, 131-148. DOI:10.1007/s10681-014-1213-2URL [本文引用: 7]
MiyataM, YamamotoT, KomoriT, NittaN (2007). Marker- assisted selection and evaluation of the QTL for stigma exsertion under japonica rice genetic background Theor Appl Genet 114, 539-548. PMID:17131105 [本文引用: 5] Stigma exsertion is one of the important traits which contribute to the efficient improvement of commercial seed production in hybrid rice. In order to understand the genetic factors involved in the stigma exsertion of an indica variety--IR24--a QTL analysis was conducted using the F2 population between a japonica variety--Koshihikari--and a breeding line showing exserted stigma selected from the backcross population between IR24 as a donor and japonica varieties. As a result, a highly significant QTL (qES3), which had been predicted in the recombinant inbred population of IR24, was confirmed at the centromeric region on chromosome 3. qES3 increases about 20% of the frequency of the exserted stigmas at the IR24 allele and explains about 32% of the total phenotypic variance. A QTL near-isogenic line for qES3 increased the frequency of the exserted stigma by 36% compared to that of Koshihikari in a field evaluation, which suggests that qES3 is a promising QTL for the development of a maternal line for hybrid rice.
QianQ, GuoLB, SmithSM, LiJY (2016). Breeding high-yield superior quality hybrid super rice by rational design Natl Sci Rev 3, 283-294. DOI:10.1093/nsr/nww006URL [本文引用: 1]
RahmanMH, YuP, ZhangYX, SunLP, WuWX, ShenXH, ZhanXD, ChenDB, CaoLY, ChengSH (2016). Quantitative trait loci mapping of the stigma exertion rate and spikelet number per panicle in rice (Oryza sativa L.) Genet Mol Res 15, gmr15048432. [本文引用: 2]
RahmanMH, ZhangYX, ZhangKQ, RahmanMS, BarmanHN, RiazA, ChenYY, WuWX, ZhanXD, CaoLY, ChengSH (2017). Genetic dissection of the major quantitative trait locus (qSE11), and its validation as the major influence on the rate of stigma exsertion in rice(Oryza sativa L.) Front Plant Sci 8, 1818. DOI:10.3389/fpls.2017.01818URL [本文引用: 3]
SasakiA, AshikariM, Ueguchi-TanakaM, ItohH, NishimuraA, SwapanD, IshiyamaK, SaitoT, KobayashiM, KhushGS, KitanoH, MatsuokaM (2002). Green revolution: a mutant gibberellin-synthesis gene in rice Nature 416, 701-702. PMID:11961544 [本文引用: 1] The chronic food shortage that was feared after the rapid expansion of the world population in the 1960s was averted largely by the development of a high-yielding semi-dwarf variety of rice known as IR8, the so-called rice 'green revolution'. The short stature of IR8 is due to a mutation in the plant's sd1 gene, and here we identify this gene as encoding an oxidase enzyme involved in the biosynthesis of gibberellin, a plant growth hormone. Gibberellin is also implicated in green-revolution varieties of wheat, but the reduced height of those crops is conferred by defects in the hormone's signalling pathway.
Takano-KaiN, Doi K, Yoshimura A(2011). GS3 participates in stigma exsertion as well as seed length in rice Breed Sci 61, 244-250. DOI:10.1270/jsbbs.61.244URL [本文引用: 1]
UgaY, FukutaY, CaiHW, IwataH, OhsawaR, MorishimaH, FujimuraT (2003). Mapping QTLs influencing rice floral morphology using recombinant inbred lines derived from a cross between Oryza sativa L. and Oryza rufipogon Griff Theor Appl Genet 107, 218-226. PMID:12845437 [本文引用: 5] To understand the genetic basis of floral traits associated with the mating system in rice, we analyzed pistil, stamen and glume traits using a recombinant inbred line population, derived from a cross between an Asian cultivated rice ( Oryza sativa L.), Pei-kuh, and a wild rice ( Oryza rufipogon Griff.), W1944. Quantitative trait loci (QTLs) affecting floral morphology were detected by composite interval mapping using a linkage map constructed using 147 markers, mostly RFLPs. A total of 7, 4, 14 and 6 QTLs were detected for traits related to pistil, stamen, and size and shape of the glume, respectively. Comparison of 31 QTLs affecting these organs revealed ten QTLs affecting the different organs in four adjacent regions on chromosomes 2, 4, 5 and 10, but most QTLs (68%) were located separately on the whole chromosomes. Although four QTLs for stigma breadth, anther length and thickness of lemma and palea explained more than 25% of the total phenotypic variance, most QTLs (87%) had smaller effects. These results suggest that quantitative variation observed for pistil, stamen and glume traits is controlled by several distinct genes with small effects.
UgaY, SiangliwM, NagamineT, OhsawaR, FujimuraT, FukutaY (2010). Comparative mapping of QTLs determining glume, pistil and stamen sizes in cultivated rice ( Oryza sativa L.) Plant Breed 129, 657-669. DOI:10.1111/j.1439-0523.2009.01765.xURL [本文引用: 4]
WangSK, WuK, YuanQB, LiuXY, LiuZB, LinXY, ZengRZ, ZhuHT, DongGJ, QianQ, ZhangGQ, FuXD (2012). Control of grain size, shape and quality by OsSPL16 in rice Nat Genet 44, 950-955. DOI:10.1038/ng.2327URL [本文引用: 1]
XingYZ, TangWJ, XueWY, XuCG, ZhangQF (2008). Fine mapping of a major quantitative trait loci, qSSP7, controlling the number of spikelets per panicle as a single Mendelian factor in rice Theor Appl Genet 116, 789-796. DOI:10.1007/s00122-008-0711-9PMID:18219477 [本文引用: 1] In our previous studies, one putative QTL affecting number of spikelets per panicle (SPP) was identified in the pericentromeric region of rice chromosome 7 using a recombinant inbred population. In order to define the QTL (qSPP7), RI50, a recombinant inbred line with 70% of genetic background same as the female parent of Zhenshan 97, was selected to produce near-isogenic lines for the target region in the present study. In a BC(2)F(2) population consisting of 190 plants, the frequency distribution of SPP was shown to be discontinuous and followed the expected Mendelian ratios (1:2:1 by progeny test) for single locus segregation. qSPP7 was mapped to a 0.4 cM region between SSR marker RM3859 and RFLP marker C39 based on tests of the BC(2)F(2) population and its progeny. Its additive and dominant effects on SPP were 51.1 and 24.9 spikelets, respectively. Of great interest, the QTL region also had effects on grain yield per plant (YD), 1,000 grain weight (GW), tillers per plant (TPP) and seed setting ratio (SR). Significant correlations were observed between SPP and YD (r = 0.66) and between SPP and SR (r = -0.29) in the progeny test. 1082 extremely small panicle plants of a BC(3)F(2) population containing 8,400 individuals were further used to fine map the QTL. It turns out that qSPP7 co-segregated with two markers, RM5436 and RM5499 spanning a physical distance of 912.4 kb. Overall results suggested that recombination suppression occurred in the region and positional cloning strategy is infeasible for qSPP7 isolation. The higher grain yield of Minghui 63 homozygote as compared to the heterozygote suggested that Minghui 63 homozygote at qSPP7 in hybrid rice could further improve its yield.
XiongLX, LiuKD, DaiXK, XuCG, ZhangQ (1999). Identification of genetic factors controlling domestication-related traits of rice using an F2 population of a cross between Oryza sativa and O. rufipogon Theor Appl Genet 98, 243-251. DOI:10.1007/s001220051064URL [本文引用: 2]
XueWY, XingYZ, WengXY, ZhaoY, TangWJ, WangL, ZhouHJ, YuSB, XuCG, LiXH, ZhangQF (2008). Na- tural variation in Ghd7 is an important regulator of heading date and yield potential in rice Nat Genet 40, 761-767. DOI:10.1038/ng.143URL [本文引用: 1]
YanWG, LiY, AgramaHA, LuoDG, GaoFY, LuXJ, RenGJ (2009). Association mapping of stigma and spikelet characteristics in rice (Oryza sativa L.) Mol Breed 24, 277-292. DOI:10.1007/s11032-009-9290-yURL [本文引用: 5]
YuXQ, MeiHW, LuoLJ, LiuGL, LiuHY, ZouGH, HuSP, LiMS, WuJH (2006). Dissection of additive, epistatic effect and Q×E interaction of quantitative trait loci influencing stigma exsertion under water stress in rice Acta Genet Sin 33, 542-550. DOI:10.1016/S0379-4172(06)60083-8URL [本文引用: 2]
ZhangKQ, ZhangYX, WuWX, ZhanXD, Bakr AnisG, RahmanMH, HongYB, RiazA, ZhuAK, CaoYR, SunLP, YangZF, YangQQ, CaoLY, ChengSH (2018). qSE7 is a major quantitative trait locus (QTL) influencing stigma exsertion rate in rice (Oryza sativa L.) Sci Rep 8, 14523. DOI:10.1038/s41598-018-32629-2URL [本文引用: 3]
ZhouH, LiPB, XieWB, HussainS, LiYB, XiaD, ZhaoH, SunSY, ChenJX, YeH, HouJ, ZhaoD, GaoGJ, ZhangQL, WangGW, LianXM, XiaoJH, YuSB, LiXH, HeYQ (2017). Genome-wide association analyses reveal the genetic basis of stigma exsertion in rice Mol Plant 10, 634-644. DOI:S1674-2052(17)30002-3PMID:28110091 [本文引用: 9] Stigma exsertion, a key determinant of the rice mating system, greatly contributes to the application of heterosis in rice. Although a few quantitative trait loci associated with stigma exsertion have been fine mapped or cloned, the underlying genetic architecture remains unclear. We performed a genome-wide association study on stigma exsertion and related floral traits using 6.5 million SNPs characterized in 533 diverse accessions of Oryza sativa. We identified 23 genomic loci that are significantly associated with stigma exsertion and related traits, three of which are co-localized with three major grain size genes GS3, GW5, and GW2. Further analyses indicated that these three genes affected the stigma exsertion by controlling the size and shape of the spikelet and stigma. Combinations of GS3 and GW5 largely defined the levels of stigma exsertion and related traits. Selections of these two genes resulted in specific distributions of floral traits among subpopulations of O.?sativa. The low stigma exsertion combination gw5GS3 existed in half of the cultivated rice varieties; therefore, introducing the GW5gs3 combination into male sterile lines is of high potential for improving the seed production of hybrid rice.Copyright ? 2017 The Author. Published by Elsevier Inc. All rights reserved. 上海杂交粳稻产业发展与展望 1 2018
... 水稻(Oryza sativa)是最主要的粮食作物, 世界一半以上的人口以稻米为主食(袁隆平, 2010).始于我国20世纪70年代的杂交水稻较同熟期常规稻增产20%以上, 杂交水稻的推广种植使水稻生产力大幅提升(Qian et al., 2016).当前, 我国杂交水稻年种植面积约1.67×107 hm2, 以杂交籼稻为主, 而杂交粳稻的种植面积不足杂交水稻种植面积的3% (曹黎明等, 2018).制种产量低是制约杂交粳稻发展的重要因素, 提高粳型不育系的异交结实率是提高杂交粳稻制种产量的关键环节(杨振玉等, 2016).异交结实率主要取决于水稻花器性状和开花习性, 花器性状主要包括柱头长度(stigma length, STL)、花柱长度(style length, SYL)、柱头外露率(percentage of exserted stigma, PES)和开颖角度(glume opening angle, GOA)等(计慎敏和张大兵, 2007; 罗伟雄等, 2011).廖佩言等(1986)研究发现, 柱头-花柱总长度(以下简称柱花总长度) (the sum of stigma and style length, TSSL)和柱头单外露率对异交结实率的贡献率分别为17.86%和7.9%.杨保汉(1997)研究表明, 柱头外露率和制种产量呈极显著正相关, 不育系的柱头外露率每增加1个百分点, 其异交结实率可提高0.74%- 0.92%, 制种产量可增加47-68 kg?hm-2.可见, 柱头性状与水稻异交结实率密切相关. ...
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
水稻花器性状对异交结实影响的初步分析 1 1986
... 水稻(Oryza sativa)是最主要的粮食作物, 世界一半以上的人口以稻米为主食(袁隆平, 2010).始于我国20世纪70年代的杂交水稻较同熟期常规稻增产20%以上, 杂交水稻的推广种植使水稻生产力大幅提升(Qian et al., 2016).当前, 我国杂交水稻年种植面积约1.67×107 hm2, 以杂交籼稻为主, 而杂交粳稻的种植面积不足杂交水稻种植面积的3% (曹黎明等, 2018).制种产量低是制约杂交粳稻发展的重要因素, 提高粳型不育系的异交结实率是提高杂交粳稻制种产量的关键环节(杨振玉等, 2016).异交结实率主要取决于水稻花器性状和开花习性, 花器性状主要包括柱头长度(stigma length, STL)、花柱长度(style length, SYL)、柱头外露率(percentage of exserted stigma, PES)和开颖角度(glume opening angle, GOA)等(计慎敏和张大兵, 2007; 罗伟雄等, 2011).廖佩言等(1986)研究发现, 柱头-花柱总长度(以下简称柱花总长度) (the sum of stigma and style length, TSSL)和柱头单外露率对异交结实率的贡献率分别为17.86%和7.9%.杨保汉(1997)研究表明, 柱头外露率和制种产量呈极显著正相关, 不育系的柱头外露率每增加1个百分点, 其异交结实率可提高0.74%- 0.92%, 制种产量可增加47-68 kg?hm-2.可见, 柱头性状与水稻异交结实率密切相关. ...
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... ; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
... 水稻(Oryza sativa)是最主要的粮食作物, 世界一半以上的人口以稻米为主食(袁隆平, 2010).始于我国20世纪70年代的杂交水稻较同熟期常规稻增产20%以上, 杂交水稻的推广种植使水稻生产力大幅提升(Qian et al., 2016).当前, 我国杂交水稻年种植面积约1.67×107 hm2, 以杂交籼稻为主, 而杂交粳稻的种植面积不足杂交水稻种植面积的3% (曹黎明等, 2018).制种产量低是制约杂交粳稻发展的重要因素, 提高粳型不育系的异交结实率是提高杂交粳稻制种产量的关键环节(杨振玉等, 2016).异交结实率主要取决于水稻花器性状和开花习性, 花器性状主要包括柱头长度(stigma length, STL)、花柱长度(style length, SYL)、柱头外露率(percentage of exserted stigma, PES)和开颖角度(glume opening angle, GOA)等(计慎敏和张大兵, 2007; 罗伟雄等, 2011).廖佩言等(1986)研究发现, 柱头-花柱总长度(以下简称柱花总长度) (the sum of stigma and style length, TSSL)和柱头单外露率对异交结实率的贡献率分别为17.86%和7.9%.杨保汉(1997)研究表明, 柱头外露率和制种产量呈极显著正相关, 不育系的柱头外露率每增加1个百分点, 其异交结实率可提高0.74%- 0.92%, 制种产量可增加47-68 kg?hm-2.可见, 柱头性状与水稻异交结实率密切相关. ...
中国杂交粳稻发展与展望 1 2016
... 水稻(Oryza sativa)是最主要的粮食作物, 世界一半以上的人口以稻米为主食(袁隆平, 2010).始于我国20世纪70年代的杂交水稻较同熟期常规稻增产20%以上, 杂交水稻的推广种植使水稻生产力大幅提升(Qian et al., 2016).当前, 我国杂交水稻年种植面积约1.67×107 hm2, 以杂交籼稻为主, 而杂交粳稻的种植面积不足杂交水稻种植面积的3% (曹黎明等, 2018).制种产量低是制约杂交粳稻发展的重要因素, 提高粳型不育系的异交结实率是提高杂交粳稻制种产量的关键环节(杨振玉等, 2016).异交结实率主要取决于水稻花器性状和开花习性, 花器性状主要包括柱头长度(stigma length, STL)、花柱长度(style length, SYL)、柱头外露率(percentage of exserted stigma, PES)和开颖角度(glume opening angle, GOA)等(计慎敏和张大兵, 2007; 罗伟雄等, 2011).廖佩言等(1986)研究发现, 柱头-花柱总长度(以下简称柱花总长度) (the sum of stigma and style length, TSSL)和柱头单外露率对异交结实率的贡献率分别为17.86%和7.9%.杨保汉(1997)研究表明, 柱头外露率和制种产量呈极显著正相关, 不育系的柱头外露率每增加1个百分点, 其异交结实率可提高0.74%- 0.92%, 制种产量可增加47-68 kg?hm-2.可见, 柱头性状与水稻异交结实率密切相关. ...
水稻柱头外露率QTL定位 2 2014
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
发展杂交水稻保障粮食安全 1 2010
... 水稻(Oryza sativa)是最主要的粮食作物, 世界一半以上的人口以稻米为主食(袁隆平, 2010).始于我国20世纪70年代的杂交水稻较同熟期常规稻增产20%以上, 杂交水稻的推广种植使水稻生产力大幅提升(Qian et al., 2016).当前, 我国杂交水稻年种植面积约1.67×107 hm2, 以杂交籼稻为主, 而杂交粳稻的种植面积不足杂交水稻种植面积的3% (曹黎明等, 2018).制种产量低是制约杂交粳稻发展的重要因素, 提高粳型不育系的异交结实率是提高杂交粳稻制种产量的关键环节(杨振玉等, 2016).异交结实率主要取决于水稻花器性状和开花习性, 花器性状主要包括柱头长度(stigma length, STL)、花柱长度(style length, SYL)、柱头外露率(percentage of exserted stigma, PES)和开颖角度(glume opening angle, GOA)等(计慎敏和张大兵, 2007; 罗伟雄等, 2011).廖佩言等(1986)研究发现, 柱头-花柱总长度(以下简称柱花总长度) (the sum of stigma and style length, TSSL)和柱头单外露率对异交结实率的贡献率分别为17.86%和7.9%.杨保汉(1997)研究表明, 柱头外露率和制种产量呈极显著正相关, 不育系的柱头外露率每增加1个百分点, 其异交结实率可提高0.74%- 0.92%, 制种产量可增加47-68 kg?hm-2.可见, 柱头性状与水稻异交结实率密切相关. ...
Detection of dominant QTLs for stigma exsertion ratio in rice derived from Oryza rufipogon accession ‘W0120’ 2 2019
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
QTL detection and elite alleles mining for stigma traits in Oryza sativa by association mapping 8 2016
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... ; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... ; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
... ; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
OsSYL2 AA, an allele identified by gene-based association, increases style length in rice (Oryza sativa L.) 2 2020
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
QTL analysis of floral traits of rice (Oryza sativa L.) under well-watered and drought stress conditions 2 2009
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Genetic mapping and validation of quantitative trait loci for stigma exsertion rate in rice 2 2014
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Natural variation in PTB1 regulates rice seed setting rate by controlling pollen tube growth 1 2013
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
Fine mapping and candidate gene analysis of qSTL3, a stigma length-conditioning locus in rice(Oryza sativa L.) 3 2015
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Fine mapping a quantitative trait locus, qSER-7, that controls stigma exsertion rate in rice(Oryza sativa L.) 4 2019
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
SNP genotyping and characterization of pistil traits revealing a distinct phylogenetic relationship among the species of Oryza 7 2015
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... ; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... ; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 柱头长度(STL)指水稻雌蕊上毛刷状部分的长度, 花柱长度(SYL)指毛刷状底部与子房交界处之间的长度(Marathi et al., 2015; Zhou et al., 2017), 柱花总长度(TSSL)指柱头长度与花柱长度之和(图1B). ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
... )、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Marker- assisted selection and evaluation of the QTL for stigma exsertion under japonica rice genetic background 5 2007
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... ; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Breeding high-yield superior quality hybrid super rice by rational design 1 2016
... 水稻(Oryza sativa)是最主要的粮食作物, 世界一半以上的人口以稻米为主食(袁隆平, 2010).始于我国20世纪70年代的杂交水稻较同熟期常规稻增产20%以上, 杂交水稻的推广种植使水稻生产力大幅提升(Qian et al., 2016).当前, 我国杂交水稻年种植面积约1.67×107 hm2, 以杂交籼稻为主, 而杂交粳稻的种植面积不足杂交水稻种植面积的3% (曹黎明等, 2018).制种产量低是制约杂交粳稻发展的重要因素, 提高粳型不育系的异交结实率是提高杂交粳稻制种产量的关键环节(杨振玉等, 2016).异交结实率主要取决于水稻花器性状和开花习性, 花器性状主要包括柱头长度(stigma length, STL)、花柱长度(style length, SYL)、柱头外露率(percentage of exserted stigma, PES)和开颖角度(glume opening angle, GOA)等(计慎敏和张大兵, 2007; 罗伟雄等, 2011).廖佩言等(1986)研究发现, 柱头-花柱总长度(以下简称柱花总长度) (the sum of stigma and style length, TSSL)和柱头单外露率对异交结实率的贡献率分别为17.86%和7.9%.杨保汉(1997)研究表明, 柱头外露率和制种产量呈极显著正相关, 不育系的柱头外露率每增加1个百分点, 其异交结实率可提高0.74%- 0.92%, 制种产量可增加47-68 kg?hm-2.可见, 柱头性状与水稻异交结实率密切相关. ...
Quantitative trait loci mapping of the stigma exertion rate and spikelet number per panicle in rice (Oryza sativa L.) 2 2016
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Genetic dissection of the major quantitative trait locus (qSE11), and its validation as the major influence on the rate of stigma exsertion in rice(Oryza sativa L.) 3 2017
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Green revolution: a mutant gibberellin-synthesis gene in rice 1 2002
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
GS3 participates in stigma exsertion as well as seed length in rice 1 2011
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
Mapping QTLs influencing rice floral morphology using recombinant inbred lines derived from a cross between Oryza sativa L. and Oryza rufipogon Griff 5 2003
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... ))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... )在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
... )和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Comparative mapping of QTLs determining glume, pistil and stamen sizes in cultivated rice ( Oryza sativa L.) 4 2010
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... , 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Genetic variability in floral characteristics influencing outcrossing in Oryza sativa L 1 1973
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
Control of grain size, shape and quality by OsSPL16 in rice 1 2012
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
Fine mapping of a major quantitative trait loci, qSSP7, controlling the number of spikelets per panicle as a single Mendelian factor in rice 1 2008
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Identification of genetic factors controlling domestication-related traits of rice using an F2 population of a cross between Oryza sativa and O. rufipogon 2 1999
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Na- tural variation in Ghd7 is an important regulator of heading date and yield potential in rice 1 2008
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Association mapping of stigma and spikelet characteristics in rice (Oryza sativa L.) 5 2009
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... )以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
... ; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
Dissection of additive, epistatic effect and Q×E interaction of quantitative trait loci influencing stigma exsertion under water stress in rice 2 2006
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
qSE7 is a major quantitative trait locus (QTL) influencing stigma exsertion rate in rice (Oryza sativa L.) 3 2018
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
The evolution and pathogenic mechanisms of the rice sheath blight pathogen 1 2013
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
Genome-wide association analyses reveal the genetic basis of stigma exsertion in rice 9 2017
... 柱头性状主要包括柱头长度、花柱长度、柱花总长度和柱头外露率(单外露率、双外露率和总外露率)等.研究发现, 柱头性状间存在紧密联系, 如柱花总长度与柱头外露率呈显著(Virmani and Athwal, 1973; Miyata et al., 2007; Takano-Kai et al., 2011)或极显著(刘强明, 2015; Zhou et al., 2017)正相关, 柱头长度、花柱长度和柱花总长度间也表现出极显著正相关(Dang et al., 2016; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... ; Zhou et al., 2017).但当前对于柱头性状的研究远远落后于水稻其它性状(如米质性状) (Wang et al., 2012)、农艺性状(Li et al., 2013; Zhou et al., 2013)以及抗性(Zheng et al., 2013).目前, 对柱头性状的研究多集中于柱头外露率的QTL定位, 研究者利用不同群体检测到112个控制柱头外露率的QTLs (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; Li et al., 2014; 尹成等, 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而其中仅3个QTLs (qSE7 (Rahman et al., 2017)、qSE11 (Zhang et al., 2018)和qSER-7 (Liu et al., 2019))被精细定位.对于柱头长度相关性状的报道则较少.研究者利用重组自交系(recombination inbred line, RIL) (Uga et al., 2003, 2010), F2、回交重组自交系(backcross inbred line, BIL)和双单倍体(doubled haploid line, DH)群体(Uga et al., 2010)以及自然群体(Yan et al., 2009; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... ; Zhou et al., 2017)在水稻的11条染色体(除第11号染色体外)上检测到28个控制柱头长度的QTLs, 在水稻第1、2、3、4、6、7、10和11号染色体上检测到21个控制花柱长度的QTLs (Uga et al., 2003, 2010; Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... ; Zhou et al., 2017), 在水稻的9条染色体(除第8、9和10号染色体外)上检测到16个控制柱花总长度的QTLs (陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016).仅见Liu等(2015)将qSTL3精细定位于水稻第3号染色体短臂19.8 kb区间, Dang等(2020)将控制花柱长度的基因OsSYL2AA克隆.尚未见柱头性状基因功能分析等深层次的研究报道, 说明当前对于柱头性状的研究少而浅, 这可能正是导致至今未见分子标记辅助选择(marked-assisted selection, MAS)选育长柱头、高外露率不育系等育种材料报道的主要原因. ...
... 柱头长度(STL)指水稻雌蕊上毛刷状部分的长度, 花柱长度(SYL)指毛刷状底部与子房交界处之间的长度(Marathi et al., 2015; Zhou et al., 2017), 柱花总长度(TSSL)指柱头长度与花柱长度之和(图1B). ...
... ; Zhou et al., 2017).本研究还表明STL与PES间也表现极显著正相关.由于毛刷状柱头(柱头长度部分)是柱头接收外来花粉的器官, 因此柱头长度增加及柱头外露率提高必然导致水稻异交结实率增加, 从而促进杂交水稻制种产量增加(Miyata et al., 2007; 吴爽等, 2015). ...
... 目前, 对于柱头性状的研究主要集中于PES (Xiong et al., 1999; Uga et al., 2003; Yu et al., 2006; Miyata et al., 2007; Hu et al., 2009; Yan et al., 2009; 尹成等, 2014; Li et al., 2014; Rahman et al., 2016, 2017; 李威等, 2017; Zhang et al., 2018; Bakti and Tanaka, 2019; Liu et al., 2019), 而对柱头长度相关性状(STL (Marathi et al., 2015; Dang et al., 2016; Zhou et al., 2017)、SYL (Marathi et al., 2015; Dang et al., 2016, 2020; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
... ; Zhou et al., 2017)和TSSL (Uga et al., 2003, 2010; Yan et al., 2009; 陈兰等, 2012; 刘强明, 2015; Liu et al., 2015; Marathi et al., 2015; Dang et al., 2016))的报道较少.因此, 对于4个柱头性状遗传效应的研究也主要集中于PES.从本研究的一阶遗传参数来看, PES以加性效应为主, 这与Yan等(2009)的研究结果一致.而STL、SYL和TSSL在不同群体中表现不尽相同.例如, STL在2个群体中均表现出以上位性效应为主, 加性效应和显性效应较小.对于SYL而言, F2群体中加性、显性和上位性效应并重, 而在F2:3群体中则表现出以加性效应为主.对于TSSL, F2群体中表现出以显性效应和上位性效应为主, 而在F2:3群体中则以加性效应为主, 这与刘强明(2015)利用CSSL群体分析得出TSSL以加性效应遗传为主的结果不尽一致, 表明柱头长度性状的遗传较PES更复杂.从二阶遗传参数来看, 4个性状在2个世代中的主基因遗传率介于63.64% (F2:3群体SYL)-92.13% (F2:3群体TSSL)之间, 多基因遗传率介于17.63% (F2群体PES)-25.00% (F2群体STL)之间, 表明这4个性状均以主基因遗传为主.但本研究同时也发现, 4个柱头性状中的多基因效应(加性效应和显性效应)也起较大作用, 如F2群体的TSSL, 其主基因效应(加性效应、显性效应和上位性效应)绝对值之和为1.18, 而其多基因效应(加性效应和显性效应)绝对值之和为1.04, 两者基本一致, 其它性状类似.以上结果说明柱头性状虽然属于数量性状, 受主效基因(主基因)和微效基因(多基因)共同控制, 但其同时又与水稻其它数量性状如株高(sd1) (Sasaki et al., 2002)、穗粒数(qSSP7) (Xing et al., 2008)和生育期(Ghd7) (Xue et al., 2008)存在差异, 即本研究中的4个柱头性状其主基因(主效基因)和多基因(微效基因)间的差异并不是很大, 而sd1、qSSP7和Ghd7等主效基因与之对应的微效基因相比其效应差异明显, 这也可能是导致当前柱头性状研究滞后的原因之一.在育种实践中, 籼型水稻的异交率远高于粳型, 尤其是在不育系方面, 这也是限制杂交粳稻和籼粳交杂交水稻推广种植的关键性因素.本研究利用核不育系籼粳交后代群体对4个柱头性状进行遗传解析, 为粳型不育系的改良提供了技术路线和理论基础.即育种家在高异交率粳型不育系选育过程中, 可将肉眼易辨的田间柱头外露率的直接观察与柱头长度性状的室内调查相结合, 通过筛选长柱头、高外露率进而提高粳型不育系的异交率.同时, 要统筹考虑主效基因和微效基因的影响, 及时开展早期有效的单株选择, 重点关注高代单株柱头性状的表现, 降低环境因素对柱头性状选择带来的干扰.4个柱头相关性状的主基因+多基因遗传分析是对其遗传组成、遗传规律特征及遗传参数进行全面剖析, 不仅可为粳型水稻柱头性状改良提供有益的指导, 也可为柱头性状的基因挖掘、功能分析和分子设计育种提供重要的理论依据. ...
 ,1,*
,1,*
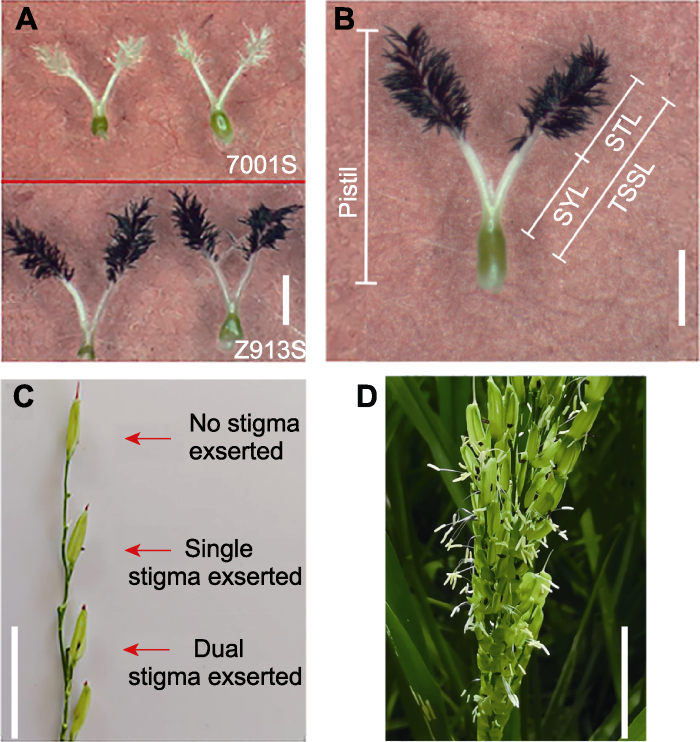 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT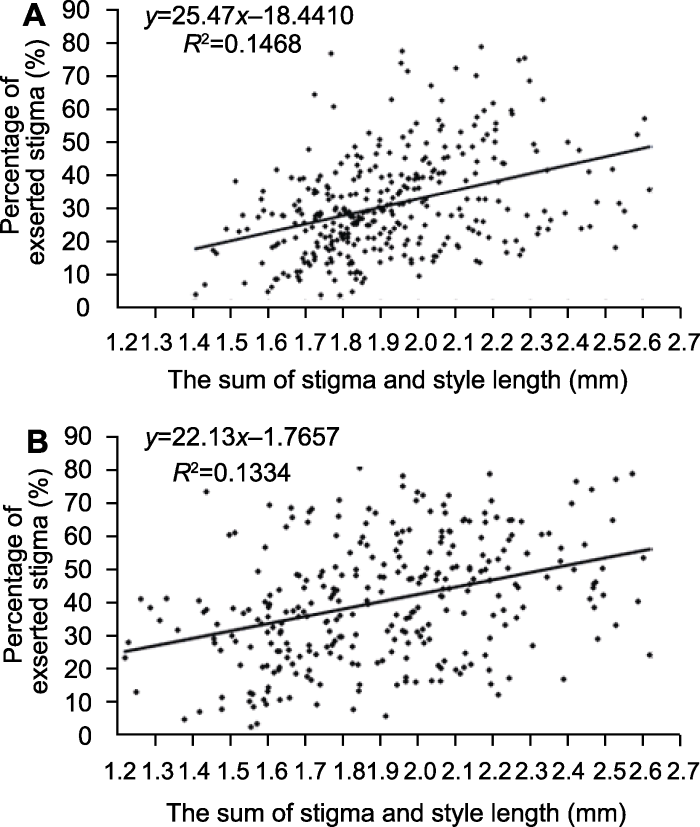 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT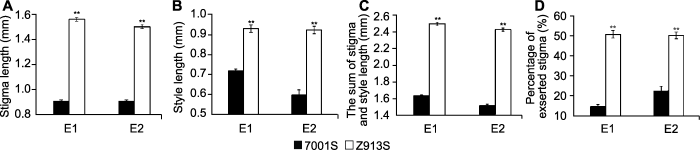 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT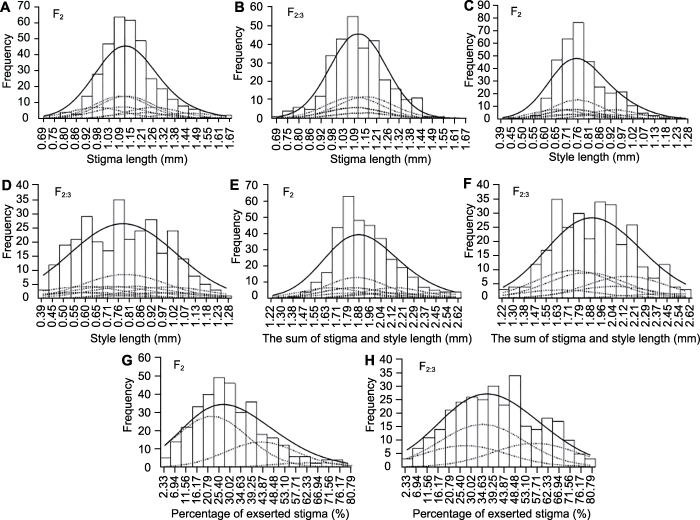 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}