Yuqing Lin1, Yanhua Qi,1,2,*1State Key Laboratory of Plant Physiology and Biochemistry, Institute of Plant Biology, College of Life Sciences, Zhejiang University, Hangzhou 310058, China 2Key Laboratory of Herbage and Endemic Crop Biotechnology, Ministry of Education, School of Life Sciences, Inner Mongolia University, Huhehot 010000, China
Abstract Auxin polar transport regulates plant growth and development. The polar transport of auxin mainly depends on three transporters: AUX/LAX, PIN and ABCB protein families. The direction of auxin flow between cells is closely related to the polar localization of PIN proteins in cells. The PIN protein contains a central hydrophilic loop (HL) and two separated hydrophobic regions, and the multiple phosphorylation sites on HL are the targets of protein kinases. The PIN proteins are fine-tuned at multiple levels, including transcriptional regulation, post-transcriptional modification, intracellular recycling and vacuolar trafficking for degradation, in response to endogenous and exogenous signals. Using genome-wide analysis, 12, 15 and 11 PIN like genes have been identified in rice, maize and sorghum, respectively, but the functions of only a few genes have been reported. Here we reviewed the research progress of PIN protein in Arabidopsis thalianaand cereal crops from the aspects of protein structure, activity regulation and functional verification to provide new ideas and clues for exploring the auxin polar transport mediated by PIN protein family. Keywords:PIN protein;auxin polar transport;Arabidopsis thaliana;cereal crops
PDF (1304KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 林雨晴, 齐艳华. 生长素输出载体PIN家族研究进展. 植物学报, 2021, 56(2): 151-165 doi:10.11983/CBB20198 Lin Yuqing, Qi Yanhua. Advances in Auxin Efflux Carrier PIN Proteins. Bulletin of Botany, 2021, 56(2): 151-165 doi:10.11983/CBB20198
生长素是最早被发现的植物激素, 参与植物生长发育的各个阶段(Enders and Strader, 2015)。生长素在植物组织和器官中建立的不对称分布是其行使功能的重要基础。例如, 调控根分生组织的活性、器官的形成和维管组织分化等, 是长期适应的结果(Sabatini et al.1999; Benkováet al.2003; Mattsson et al.2003)。生长素从合成部位(如幼叶和顶芽)运输到作用部位主要有2条途径: 一是在整个植株范围内, 以不固定方向被动运输的方式, 通过韧皮部完成长距离运输; 二是需要载体参与的细胞与细胞之间短距离极性运输, 包括从茎尖向根的单向运输和组织间的局部单向运输( Petrá?ek and Friml,2009)。极性运输对建立和维持生长素的浓度梯度至关重要, 进而特异性调节植物的生长发育及其对外界环境变化的响应。
生长素的极性运输需要载体, 目前已证实有家族蛋白具有生长素转运功能(Rubery and Sheldrake, 1974; Bennett et al.1996; G?lweileret al.1998; Sidler et al.1998)。生长素在细胞间的运输主要依赖3种膜定位蛋白, 分别为生长素输入载体AUX/LAX (AUXIN1/LIKE-AUX1)、生长素输出载体PIN (PIN- FORMED)以及ABCB/PGP (ATP-Binding Cassette subfamily B/P-glycoprotein)转运体(Péretet al.2012; Geisler et al.2017; Zhou and Luo, 2018)。此外, 在拟南芥(Arabidopsis thaliana)中, 硝酸盐转运体NRT1.1也具有转运生长素的能力, 低氮条件下其通过促进生长素的向基运输抑制侧根的生长(Krouk et al.2010)。生长素在细胞内的运输则依赖定位于胞内的PILS (PIN-Like)蛋白家族、液泡生长素转运蛋白WAT1 (WALLS ARE THIN 1)以及具有短亲水环结构的部分PIN蛋白(Barbez et al.2012; Ranocha et al.2013)。
根据化学渗透偶联学说, 生长素输出载体在细胞膜上的不对称分布是生长素极性运输的关键(Friml, 2010; Peer et al.2011)。已有研究表明, PIN蛋白在质膜上的极性定位关系到生长素的运输方向, 是生长素在植物体内不对称分布的重要原因(Benkováet al.2003; Wi?niewska,2006; Vieten et al.2007)。运用全基因组关联分析(genome wide association study, GWAS)方法, 目前已在30多种植物中鉴定出不同数量的PIN基因(Zhou and Luo, 2018)。例如, 在咖啡(Coffea arabica)、玉米(Zea mays)以及高粱(Sorghum bicolor)中分别发现了17、15和11个PIN基因(Shen et al.2010; Yue et al.2015; Huang et al.2020)。模式植物拟南芥中有8个PIN蛋白成员, 其生物学功能已有相关报道。 然而对水稻(Oryza sativa)、小麦(Triticum aestivum)和番茄(Solanum lycopersicum)等作物以及裸子植物中的PIN基因功能还知之甚少, 尚待进一步研究和探索(K?e?eket al.2009)。本文主要综述了PIN蛋白的分子结构、极性定位的分子调控机理及其在拟南芥和禾谷类作物中的研究进展。
1 PIN蛋白的发现和分子结构
PIN家族最早被发现的基因是PIN1。该突变体的特征是花序茎上几乎无叶和花等器官, 形状类似大头针, 因此命名为pin1 (pin-formed1)突变体(Goto et al.1987)。研究发现, 在添加生长素极性运输抑制剂培养基上生长的野生型拟南芥会表现出与pin1突变体相似的表型, 且pin1突变体中生长素的极性运输活性降低, 因此推测PIN1参与花序发育中生长素的极性运输(Okada et al.1991)。1998年, PIN1基因被克隆出来, 进一步研究表明, PIN1的蛋白结构类似于细菌和真核生物中的载体蛋白, 且极性定位于维管组织细胞的细胞膜基部, 与化学渗透假说推测的生长素输出载体相符(G?lweileret al.1998)。
EIR1基因的克隆早于PIN1, Luschnig等(1998)研究发现, 表达EIR1蛋白的酵母(Saccharomyces cerevisiae)能将有毒的IAA类似物5-氟-吲哚运出细胞, 从而表现出对该化合物的抗性, 表明EIR1蛋白具有生长素输出功能。在此之前, 通过不同途径鉴定到的根向地性缺失突变体agr1和wavy6、根特异的乙烯不敏感突变体eir1的突变基因, 以及同年由其它3个实验室从根向地性缺失突变体中分离克隆的基因, 经序列分析表明为同一基因, 该基因与PIN1同源, 命名为PIN2(Bell and Maher, 1990; Okada and Shimura, 1990; Roman et al.1995; Chen et al.1998; Mülleret al.1998; Utsuno et al.1998)。不同角度的研究结果共同证实了PIN的生长素输出功能, 最直接的证据是当在植物细胞(如烟草(Nicotiana tabacum) BY-2细胞和拟南芥悬浮细胞)和异源表达系统(如爪蟾(Xenopus laevis)卵母细胞和酵母)中表达PIN蛋白时, 其均能表现出外排生长素的能力(Petráseket al.2006; Yang and Murphy, 2009)。
(A) 以拟南芥PIN1为代表的长亲水环PIN蛋白结构模型及亲水环上的磷酸化位点; (B) 短亲水环PIN蛋白结构模型。 Figure 1Structural model of PIN proteins
(A)Structural model of PIN protein with a long hydrophilic loop (HL) and the phosphorylation sites on the HL (with AtPIN1 as an example); (B)Structural model of PIN protein with a short HL.
在长亲水环上还含有多个保守的磷酸化位点, 它们是调节PIN转运和亚细胞极性的某些激酶的靶点(图1)。对拟南芥的研究表明, 在3个高度保守的TPRXS基序中, 丝氨酸残基S1-S3 (S231、S252和S290)是PIDs (PINOID)激酶和D6蛋白激酶(D6PKs) 的靶点, 与S1-S3相邻的3个苏氨酸残基(T227、T248和T286)是促分裂素原活化激酶(mitogen activated protein kinases, MAPK)的靶点(Huang et al.2010; Zourelidou et al.2014; Dory et al.2018)。此外, 在长亲水环上不保守的丝氨酸残基S337和苏氨酸残基T340也被证明参与PIN的磷酸化调控。MAPK能通过磷酸化S337来调节拟南芥初生茎中生长素的极性运输, 但靶向T340位点的激酶目前未知(Ganguly et al.2012; Jia et al.2016)。
参与PIN磷酸化过程的激酶家族主要有AGC、MAPK蛋白和钙调蛋白激酶(Ca2+/calmodulin-dependent protein kinases, CDPK) 3类。拟南芥基因组编码的39个AGC激酶隶属AGCVI、AGCVII、AGCVIII、PDK1及其它AGC激酶5个亚族, AGCVIII亚族的23个激酶进一步细分为AGC1、AGC2、AGC3和AGC4四类, PIDs属于AGCVIII激酶的AGC3类, D6PKs属于AGC1类(B?greet al.2003; Galván-Ampudia and Offringa,2007)。PIDs通过磷酸化S1-S3位点调节PIN的极性定位, 过表达PID促进PIN蛋白从细胞基部定位到顶部, 导致一定范围内的生长素不具有浓度梯度; pid突变体茎分生组织分化过程必需的PIN基部到顶部的极性转换受阻, 导致器官发育缺陷(Friml et al.2004)。有研究表明, 磷脂酸(phosphatidic acid, PA)能增强PID介导的PIN2磷酸化, 从而激活生长素外排, 促进盐胁迫下拟南芥的根系生长, 说明AGCVIII激酶可从多方面调控PIN的功能(Wang et al.2019)。D6PKs在大部分细胞中与PIN蛋白共定位于细胞基部, 其磷酸化位点除了S1-S3外, 还包括丝氨酸残基S4和S5 (S271和S215), 这2个位点在PIN3、PIN4和PIN7中保守, 但在PIN1和PIN2中不完全保守。d6pk功能缺失突变体中PIN蛋白的磷酸化和生长素运输速率均降低, 然而与PIDs不同的是, 过表达D6PK并不改变细胞中PIN的极性。PAX是一种D6PKL (D6PK-LIKE)蛋白激酶, 通过磷酸化S1位点激活PIN蛋白活性, 促进生长素流出。在初生韧皮部中, PAX与BRX因子互作调节生长素的含量, 从而在时间上精确控制原生韧皮分子(protophloem sieve elements, PPSE)的分化(Marhava et al.2018)。D6PKs与PIDs共享PIN上的磷酸化靶点, 但它们对这些靶点有不同的选择偏好, 因此其在调控PIN极性上的作用不同(Zourelidou et al.2014; Barbosa et al.2018)。其它AGCVIII激酶也被证明能够在体内或体外通过磷酸化PIN蛋白参与植物生长的调控, 但具体机理尚不清楚(Galván-Ampudia and Offringa,2007; Haga et al.2018)。
拟南芥基因组共编码20个MAPK和10个MAPK 激酶(MKKs), MKK-MAPK信号级联参与多种植物发育进程的调控。MKK7-MAPK3/6级联通过磷酸化S337位点使PIN在下胚轴和茎中去极化, 从而影响茎中生长素的极性运输(Jia et al.2016)。MAPK4和MAPK6能够磷酸化T227、T248和T286, 这3个位点与S1-S3相邻, 说明AGCVIII和MAPK蛋白激酶很可能通过磷酸化PIN蛋白上的丝氨酸和苏氨酸位点介导PIN的极性(Dory et al.2018)。
植物固醇、生长素和其它植物激素等也参与PIN的胞内循环调控, 其中有些植物激素通过反馈调节影响PIN的循环运输。生长素处理可在短时间内维持PIN定位在膜上, 依赖TIR1介导的信号途径抑制PIN进入内吞小泡; 长时间处理则会导致PIN蛋白降解(Pan et al.2009)。植物固醇合成途径的关键基因CPI1突变后, 突变体中的胞吞作用被抑制, 使PIN2的极性定位改变(Men et al.2008)。低浓度茉莉酸甲酯(MeJA)处理抑制PIN2的内吞也依赖TIR1信号途径; 细胞分裂素则促进PIN1从质膜到液泡的降解; 独脚金内酯(SLs)干扰生长素对PIN极性的反馈调节过程(包括对CME途径的干扰), 从而影响维管组织的形成和再生(Sun et al.2011; Marhavyet al.2011; Zhang et al.2020)。
在转录水平上, 有些上游转录因子作用于PIN基因的启动子区, 调节其表达以响应生长素内源以及外源信号变化。在生长素响应因子ARF (AUXIN RE-SPONSE FACTOR)家族中, ARF7与受其调控的FLP转录因子共同形成前馈基序以调控PIN3的转录, 介导侧根形成的早期阶段(Chen et al.2015); 细胞分裂素响应因子CRF2和CRF6作用于PIN1和PIN7启动子上的PIN特异性细胞分裂素响应元件(PCRE), 激活PIN1和PIN7的表达, 并且CRF2受ARF5的调控, 该研究表明生长素的极性运输受生长素和细胞分裂素的交叉信号调控(?imá?kováet al.2015)。参与脂肪酸生物合成的AP2转录因子WRI1 (WRINK-LED1)直接靶向PIN4和PIN5的启动子, 并导致wri1-1突变体中某些PIN基因(如PIN1、PIN3和PIN5-6)的表达水平降低, 为阐明脂肪酸和PIN蛋白的关系提供了思路(Kong et al.2017)。此外, 近期研究发现, NAC转录因子AtNAC2 (ANAC092)与ARF8和PIN4的启动子结合, 通过生长素途径负调控拟南芥根的生长(Xi et al.2019)。此外, IBR5也影响PIN的转录活性, 表现为ibr5-3突变体中, PIN1在表皮细胞质膜和叶脉周边细胞中的分布受到影响, 叶片锯齿形成受阻(Kong et al.2019)。MAKR2 (MEMBRANE ASSOCIATED KINASE REGULATOR2)作为跨膜激酶TMK1 (TRAN SMEMBRANE KINASE1)的负调控因子, 影响PIN2在根两侧的动态分布, 进而参与根的向重力性调控(Marquès-Buenoet al.2021)。表观遗传方面, SWI/SNF的染色质重塑ATP酶BRM (BRAHMA)直接靶向根部“长”PIN蛋白(PIN1-PIN4和PIN7)的染色质, 功能丧失突变体brm表现为根干细胞生态位维持缺陷, 根生长发育不良, 干细胞中一些PIN基因的局部表达量降低, 从而影响生长素的分布(Yang et al.2015)。生长素转运蛋白ABCB家族与PIN蛋白的互作也会调控其表达活性。有研究表明, ABCB1、ABCB19可与PIN1、PIN2互作, 共表达PIN1-ABCB1和PIN1- ABCB19的植株生长素输出速率升高(Blakeslee et al.2007; 贺祯媚等, 2019)。
PIN基因的表达还受多种植物激素的影响(包括生长素自身的诱导)。除了PIN5的表达受生长素负调节外, 其余PIN的表达均由生长素通过Aux/IAA信号转导途径正反馈调控(Mravec et al.2009)。乙烯合成前体ACC处理可使PIN1、PIN2和PIN4的表达上调; 细胞分裂素则通过抑制PIN1-PIN3的表达调节根分生区活性; 外施油菜素内酯(BL)可提高PIN1和PIN2的转录水平, BL合成突变体中PIN1和PIN2的转录水平下降; 茉莉酸(JA)处理可使PIN7的表达水平下调, 并由此控制生长素的极性运输, 调节根中木质部的发育(Li et al.2005; Ruzicka et al.2007, 2009; Jang et al.2019)。上述结果表明, 各植物激素信号对植物生长发育的调控之间存在很重要的调控网络, 但这些网络互作的具体机理尚需进一步探索。
4 禾谷类作物中PIN家族研究进展
单子叶植物中的禾谷类作物(包括水稻、小麦、玉米和高粱等)是人类最重要的粮食作物。虽然禾谷类作物中PIN蛋白的结构与拟南芥具有相似性, 但其在形态和生长习性方面有很大差别(Wang et al.2009)。因此, 单/双子叶植物中PIN介导的生长素极性运输调节植株发育可能具有不同的调控机制。阐明这些机理及深入研究禾谷类作物中的PIN家族, 对分子设计育种和粮食增产均具有重要意义。
AdamowskiM, NarasimhanM, KaniaU, GlancM, DeJaeger G, FrimlJ (2018). A functional study of AUXILIN-LIKE1 and 2, two putative clathrin uncoating factors in Arabidopsis Plant Cell30,700-716. [本文引用: 1]
BarbosaICR, HammesUZ, SchwechheimerC (2018). Activation and polarity control of PIN-FORMED auxin transporters by phosphorylation Trends Plant Sci23,523- 538. [本文引用: 1]
BellCJ, MaherEP (1990). Mutants of Arabidopsis thaliana with abnormal gravitropic responses Mol Gen Genet220,289-293. DOI:10.1007/BF00260496URL [本文引用: 1]
BenkováE, MichniewiczM, SauerM, TeichmannT, SeifertováD, JürgensG, FrimlJ (2003). Local, efflux-dependent auxin gradients as a common module for plant organ formation Cell115,591-602. DOI:10.1016/S0092-8674(03)00924-3URL [本文引用: 2]
BennettT, BrockingtonSF, RothfelsC, GrahamSW, StevensonD, KutchanT, RolfM, ThomasP, WongGKS, LeyserO, GloverBJ, HarrisonCJ (2014). Para- logous radiations of PIN proteins with multiple origins of noncanonical PIN structure Mol Biol Evol31,2042-2060. DOI:10.1093/molbev/msu147URL [本文引用: 1] The plant hormone auxin is a conserved regulator of development which has been implicated in the generation of morphological novelty. PIN-FORMED1 (PIN) auxin efflux carriers are central to auxin function by regulating its distribution. PIN family members have divergent structures and cellular localizations, but the origin and evolutionary significance of this variation is unresolved. To characterize PIN family evolution, we have undertaken phylogenetic and structural analyses with a massive increase in taxon sampling over previous studies. Our phylogeny shows that following the divergence of the bryophyte and lycophyte lineages, two deep duplication events gave rise to three distinct lineages of PIN proteins in euphyllophytes. Subsequent independent radiations within each of these lineages were taxonomically asymmetric, giving rise to at least 21 clades of PIN proteins, of which 15 are revealed here for the first time. Although most PIN protein clades share a conserved canonical structure with a modular central loop domain, a small number of noncanonical clades dispersed across the phylogeny have highly divergent protein structure. We propose that PIN proteins underwent sub- and neofunctionalization with substantial modification to protein structure throughout plant evolution. Our results have important implications for plant evolution as they suggest that structurally divergent PIN proteins that arose in paralogous radiations contributed to the convergent evolution of organ systems in different land plant lineages.
BlakesleeJJ, BandyopadhyayA, LeeOR, MravecJ, TitapiwatanakunB, SauerM, MakamSN, ChengY, BouchardR, AdamecJ, GeislerM, NagashimaA, SakaiT, MartinoiaE, FrimlJ, PeerWA, MurphyAS (2007). Interactions among PIN-FORMED and P-Glycoprotein auxin transporters in Arabidopsis Plant Cell19,131-147. PMID:17237354 [本文引用: 1] Directional transport of the phytohormone auxin is established primarily at the point of cellular efflux and is required for the establishment and maintenance of plant polarity. Studies in whole plants and heterologous systems indicate that PIN-FORMED (PIN) and P-glycoprotein (PGP) transport proteins mediate the cellular efflux of natural and synthetic auxins. However, aromatic anion transport resulting from PGP and PIN expression in nonplant systems was also found to lack the high level of substrate specificity seen in planta. Furthermore, previous reports that PGP19 stabilizes PIN1 on the plasma membrane suggested that PIN-PGP interactions might regulate polar auxin efflux. Here, we show that PGP1 and PGP19 colocalized with PIN1 in the shoot apex in Arabidopsis thaliana and with PIN1 and PIN2 in root tissues. Specific PGP-PIN interactions were seen in yeast two-hybrid and coimmunoprecipitation assays. PIN-PGP interactions appeared to enhance transport activity and, to a greater extent, substrate/inhibitor specificities when coexpressed in heterologous systems. By contrast, no interactions between PGPs and the AUXIN1 influx carrier were observed. Phenotypes of pin and pgp mutants suggest discrete functional roles in auxin transport, but pin pgp mutants exhibited phenotypes that are both additive and synergistic. These results suggest that PINs and PGPs characterize coordinated, independent auxin transport mechanisms but also function interactively in a tissue-specific manner.
BlilouI, XuJ, WildwaterM, WillemsenV, PaponovI, FrimlJ, HeidstraR, AidaM, PalmeK, ScheresB (2005). The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots Nature433,39-44. DOI:10.1038/nature03184URL [本文引用: 1]
B?greL, OkrészL, HenriquesR, AnthonyRG (2003). Growth signaling pathways in Arabidopsis and the AGC protein kinases Trends Plant Sci8,424-431. DOI:10.1016/S1360-1385(03)00188-2URL [本文引用: 1]
CarraroN, ForestanC, CanovaS, TraasJ, VarottoS (2006). ZmPIN1a and ZmPIN1b encode two novel putative candidates for polar auxin transport and plant architecture determination of maize Plant Physiol142,254- 264. PMID:16844839 [本文引用: 1] Shoot apical meristems produce organs in a highly stereotypic pattern that involves auxin. Auxin is supposed to be actively transported from cell to cell by influx (AUXIN/LIKE AUXIN proteins) and efflux (PIN-FORMED proteins) membrane carriers. Current hypotheses propose that, at the meristem surface, PIN proteins create patterns of auxin gradients that, in turn, create patterns of gene expression and morphogenesis. These hypotheses are entirely based on work in Arabidopsis (Arabidopsis thaliana). To verify whether these models also apply to other species, we studied the behavior of PIN proteins during maize (Zea mays) development. We identified two novel putative orthologs of AtPIN1 in maize and analyzed their expression pattern during development. The expression studies were complemented by immunolocalization studies using an anti-AtPIN1 antibody. Interestingly, the maize proteins visualized by this antibody are almost exclusively localized in subepidermal meristematic layers. Both tassel and ear were characterized by a compact group of cells, just below the surface, carrying PIN. In contrast to or to complement what was shown in Arabidopsis, these results point to the importance of internally localized cells in the patterning process. We chose the barren inflorescence2 (bif2) maize mutant to study the role of auxin polar fluxes in inflorescence development. In severe alleles of bif2, the tassel and the ear present altered ZmPIN1a and ZmPIN1b protein expression and localization patterns. In particular, the compact groups of cells in the tassel and ear of the mutant were missing. We conclude that BIF2 is important for PIN organization and could play a role in the establishment of polar auxin fluxes in maize inflorescence, indirectly modulating the process of axillary meristem formation and development.
ChenQ, LiuY, MaereS, LeeE, VanIsterdael G, XieZD, XuanW, LucasJ, VassilevaV, KitakuraS, MarhavyP, WabnikK, GeldnerN, BenkováE, LeJ, FukakiH, GrotewoldE, LiCY, FrimlJ, SackF, BeeckmanT, VannesteS (2015). A coherent transcriptional feed-forward motif model for mediating auxin-sensitive PIN3 expression during lateral root development Nat Commun6,8821. DOI:10.1038/ncomms9821URL [本文引用: 2]
ChenR, HilsonP, SedbrookJ, RosenE, CasparT, MassonPH (1998). The Arabidopsis thaliana AGRAVITROPIC 1 gene encodes a component of the polar-auxin- transport efflux carrier Proc Natl Acad Sci USA95,15112- 15117. DOI:10.1073/pnas.95.25.15112URL [本文引用: 2]
ChenYN, FanXR, SongWJ, ZhangYL, XuGH (2012). Over-expression of OsPIN2 leads to increased tiller num- bers, angle and shorter plant height through suppression of OsLAZY1 Plant Biotechnol J10,139-149. DOI:10.1111/j.1467-7652.2011.00637.xURL [本文引用: 2] Crop architecture parameters such as tiller number, angle and plant height are important agronomic traits that have been considered for breeding programmes. Auxin distribution within the plant has long been recognized to alter architecture. The rice (Oryza sativa L.) genome contains 12 putative PIN genes encoding auxin efflux transporters, including four PIN1 and one PIN2 genes. Here, we report that over-expression of OsPIN2 through a transgenic approach in rice (Japonica cv. Nipponbare) led to a shorter plant height, more tillers and a larger tiller angle when compared with wild type (WT). The expression patterns of the auxin reporter DR5::GUS and quantification of auxin distribution showed that OsPIN2 over-expression increased auxin transport from the shoot to the rootshoot junction, resulting in a non-tissue-specific accumulation of more free auxin at the rootshoot junction relative to WT. Over-expression of OsPIN2 enhanced auxin transport from shoots to roots, but did not alter the polar auxin pattern in the roots. Transgenic plants were less sensitive to N-1-naphthylphthalamic acid, an auxin transport inhibitor, than WT in their root growth. OsPIN2-over-expressing plants had suppressed the expression of a gravitropism-related gene OsLazy1 in the shoots, but unaltered expression of OsPIN1b and OsTAC1, which were reported as tiller angle controllers in rice. The data suggest that OsPIN2 has a distinct auxin-dependent regulation pathway together with OsPIN1b and OsTAC1 controlling rice shoot architecture. Altering OsPIN2 expression by genetic transformation can be directly used for modifying rice architecture.
DitengouFA, GomesD, NzienguiH, KocherspergerP, LasokH, MedeirosV, PaponovIA, NagySK, NádaiTV, MészárosT, BarnabásB, DitengouBI, RappK, QiLL, LiXG, BeckerC, LiCY, DócziR, PalmeK (2018). Characterization of auxin transporter PIN6 plasma membrane targeting reveals a function for PIN6 in plant bolting New Phytol217,1610-1624. DOI:10.1111/nph.14923URL [本文引用: 2]
DoryM, HatzimasouraE, KállaiBM, NagySK, J?gerK, DarulaZ, NádaiTV, MészárosT, López-JuezE, BarnabásB, PalmeK, B?greL, DitengouFA, DócziR (2018). Coevolving MAPK and PID phosphosites indicate an ancient environmental control of PIN auxin transpor- ters in land plants FEBS Lett592,89-102. DOI:10.1002/feb2.2018.592.issue-1URL [本文引用: 2]
FeraruE, FeraruMI, Kleine-VehnJ, MartinièreA, MouilleG, VannesteS, VernhettesS, RunionsJ, FrimlJ (2011). PIN polarity maintenance by the cell wall in Arabidopsis Curr Biol21,338-343. DOI:10.1016/j.cub.2011.01.036URL [本文引用: 1] A central question in developmental biology concerns the mechanism of generation and maintenance of cell polarity, because these processes are essential for many cellular functions and multicellular development [1]. In plants, cell polarity has an additional role in mediating directional transport of the plant hormone auxin that is crucial for multiple developmental processes [2-4]. In addition, plant cells have a complex extracellular matrix, the cell wall [5, 6], whose role in regulating cellular processes, including cell polarity, is unexplored. We have found that polar distribution of PIN auxin transporters [7] in plant cells is maintained by connections between polar domains at the plasma membrane and the cell wall. Genetic and pharmacological interference with cellulose, the major component of the cell wall, or mechanical interference with the cell wall disrupts these connections and leads to increased lateral diffusion and loss of polar distribution of PIN transporters for the phytohormone auxin. Our results reveal a plant-specific mechanism for cell polarity maintenance and provide a conceptual framework for modulating cell polarity and plant development via endogenous and environmental manipulations of the cellulose-based extracellular matrix.
ForestanC, FarinatiS, VarottoS (2012). The maize PIN gene family of auxin transporters Front Plant Sci3,16. [本文引用: 1]
ForestanC, VarottoS (2012). The role of PIN auxin efflux carriers in polar auxin transport and accumulation and their effect on shaping maize development Mol Plant5,787-798. DOI:10.1093/mp/ssr103URL [本文引用: 1] In plants, proper seed development and the continuing post-embryonic organogenesis both require that different cell types are correctly differentiated in response to internal and external stimuli. Among internal stimuli, plant hormones and particularly auxin and its polar transport (PAT) have been shown to regulate a multitude of plant physiological processes during vegetative and reproductive development. Although our current auxin knowledge is almost based on the results from researches on the eudicot Arabidopsis thaliana, during the last few years, many studies tried to transfer this knowledge from model to crop species, maize in particular. Applications of auxin transport inhibitors, mutant characterization, and molecular and cell biology approaches, facilitated by the sequencing of the maize genome, allowed the identification of genes involved in auxin metabolism, signaling, and particularly in polar auxin transport. PIN auxin efflux carriers have been shown to play an essential role in regulating PAT during both seed and post-embryonic development in maize. In this review, we provide a summary of the recent findings on PIN-mediated polar auxin transport during maize development. Similarities and differences between maize and Arabidopsis are analyzed and discussed, also considering that their different plant architecture depends on the differentiation of structures whose development is controlled by auxins.
FrimlJ (2010). Subcellular trafficking of PIN auxin efflux carriers in auxin transport Eur J Cell Biol89,231-235. DOI:10.1016/j.ejcb.2009.11.003URL [本文引用: 1]
G?lweilerL, GuanCH, MüllerA, WismanE, MendgenK, YephremovA, PalmeK (1998). Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue S- cience282,2226-2230. [本文引用: 3]
GangulyA, ParkM, KesawatMS, ChoHT (2014). Functional analysis of the hydrophilic loop in intracellular trafficking of Arabidopsis PIN-FORMED proteins Plant Cell26,1570-1585. DOI:10.1105/tpc.113.118422URL [本文引用: 1] Different PIN-FORMED proteins (PINs) contribute to intercellular and intracellular auxin transport, depending on their distinctive subcellular localizations. Arabidopsis thaliana PINs with a long hydrophilic loop (HL) (PIN1 to PIN4 and PIN7; long PINs) localize predominantly to the plasma membrane (PM), whereas short PINs (PIN5 and PIN8) localize predominantly to internal compartments. However, the subcellular localization of the short PINs has been observed mostly for PINs ectopically expressed in different cell types, and the role of the HL in PIN trafficking remains unclear. Here, we tested whether a long PIN-HL can provide its original molecular cues to a short PIN by transplanting the HL. The transplanted long PIN2-HL was sufficient for phosphorylation and PM trafficking of the chimeric PIN5: PIN2-HL but failed to provide the characteristic polarity of PIN2. Unlike previous observations, PIN5 showed clear PM localization in diverse cell types where PIN5 is natively or ectopically expressed and even polar PM localization in one cell type. Furthermore, in the root epidermis, the subcellular localization of PIN5 switched from PM to internal compartments according to the developmental stage. Our results suggest that the long PIN-HL is partially modular for the trafficking behavior of PINs and that the intracellular trafficking of PIN is plastic depending on cell type and developmental stage.
GangulyA, SasayamaD, ChoHT (2012). Regulation of the polarity of protein trafficking by phosphorylation Mol Cells33,423-430. DOI:10.1007/s10059-012-0039-9URL [本文引用: 1] The asymmetry of environmental stimuli and the execution of developmental programs at the organism level require a corresponding polarity at the cellular level, in both unicellular and multicellular organisms. In plants, cell polarity is important in major developmental processes such as cell division, cell enlargement, cell morphogenesis, embryogenesis, axis formation, organ development, and defense. One of the most important factors controlling cell polarity is the asymmetric distribution of polarity determinants. In particular, phosphorylation is implicated in the polar distribution of the determinant protein factors, a mechanism conserved in both prokaryotes and eukaryotes. In plants, formation of local gradients of auxin, the morphogenic hormone, is critical for plant developmental processes exhibiting polarity. The auxin efflux carriers PIN-FORMEDs (PINs) localize asymmetrically in the plasma membrane and cause the formation of local auxin gradients throughout the plant. The asymmetry of PIN distribution in the plasma membrane is determined by phosphorylationmediated polar trafficking of PIN proteins. This review discusses recent studies on the role of phosphorylation in polar PIN trafficking.
GeislerM, AryalB, DiDonato M, HaoPC (2017). A critical view on ABC transporters and their interacting partners in auxin transport Plant Cell Physiol58,1601-1614. DOI:10.1093/pcp/pcx104URL [本文引用: 1]
GeldnerN, RichterS, VietenA, MarquardtS, Torres-RuizRA, MayerU, JürgensG (2004). Partial loss-of-function alleles reveal a role for GNOM in auxin transport-related, post-embryonic development of Arabidopsis Development131,389-400. [本文引用: 1]
GotoN, StarkeM, KranzAR (1987). Effect of gibberellins on flower development of the pin-formed mutant of Ara- bidopsis thaliana Arabidopsis Inf Serv23,66-71. [本文引用: 1]
HagaK, FrankL, KimuraT, SchwechheimerC, SakaiT (2018). Roles of AGCVIII kinases in the hypocotyl phototropism of Arabidopsis seedlings Plant Cell Physiol59,1060-1071. [本文引用: 1]
HouMM, LuoFF, WuDX, ZhangXH, LouMM, ShenDF, YanM, MaoCZ, FanXR, XuGH, ZhangYL (2021). OsPIN9, an auxin efflux carrier, is required for the regulation of rice tiller bud outgrowth by ammonium New Phytol229,935-949. DOI:10.1111/nph.v229.2URL [本文引用: 3]
HuangF, ZagoMK, AbasL, VanMarion A, Galván-Am-pudia CS, OffringaR (2010). Phosphorylation of conserved PIN motifs directs Arabidopsis PIN1 polarity and auxin transport Plant Cell22,1129-1142. DOI:10.1105/tpc.109.072678URL [本文引用: 1]
HuangX, BaiXH, GuoTY, XieZL, LaimerM, DuDX, GbokieT Jr, ZhangZR, HeCP, LuY, WuWH, YiK (2020). Genome-wide analysis of the PIN auxin efflux carrier gene family in coffee Plants9,1061. DOI:10.3390/plants9091061URL [本文引用: 1]
JangG, YoonY, ChoiYD (2019). Jasmonic acid modulates xylem development by controlling expression of PIN-FOR- MED 7 Plant Signal Behav14,1637664. DOI:10.1080/15592324.2019.1637664URL [本文引用: 1]
JiaWY, LiBH, LiSJ, LiangY, WuXW, MaM, WangJY, GaoJ, CaiYY, ZhangYY, WangYC, LiJY, WangYH (2016). Mitogen-activated protein kinase cascade MKK7- MPK6 plays important roles in plant development and regulates shoot branching by phosphorylating PIN1 in Arabidopsis PLoS Biol14,e1002550. DOI:10.1371/journal.pbio.1002550URL [本文引用: 2]
KeuskampDH, PollmannS, VoesenekLACJ, PeetersAJ, PierikR (2010). Auxin transport through PIN-FORMED 3 (PIN3) controls shade avoidance and fitness during com- petition Proc Natl Acad Sci USA107,22740-22744. DOI:10.1073/pnas.1013457108URL [本文引用: 1]
KitakuraS, VannesteS, RobertS, L?fkeC, TeichmannT, TanakaH, FrimlJ (2011). Clathrin mediates endocytosis and polar distribution of PIN auxin transporters in Arabidopsis Plant Cell23, 1920-1931. DOI:10.1105/tpc.111.083030URL [本文引用: 1] Endocytosis is a crucial mechanism by which eukaryotic cells internalize extracellular and plasma membrane material, and it is required for a multitude of cellular and developmental processes in unicellular and multicellular organisms. In animals and yeast, the best characterized pathway for endocytosis depends on the function of the vesicle coat protein clathrin. Clathrin-mediated endocytosis has recently been demonstrated also in plant cells, but its physiological and developmental roles remain unclear. Here, we assessed the roles of the clathrin-mediated mechanism of endocytosis in plants by genetic means. We interfered with clathrin heavy chain (CHC) function through mutants and dominant-negative approaches in Arabidopsis thaliana and established tools to manipulate clathrin function in a cell type-specific manner. The chc2 single mutants and dominant-negative CHC1 (HUB) transgenic lines were defective in bulk endocytosis as well as in internalization of prominent plasma membrane proteins. Interference with clathrin-mediated endocytosis led to defects in constitutive endocytic recycling of PIN auxin transporters and their polar distribution in embryos and roots. Consistent with this, these lines had altered auxin distribution patterns and associated auxin transport-related phenotypes, such as aberrant embryo patterning, imperfect cotyledon specification, agravitropic growth, and impaired lateral root organogenesis. Together, these data demonstrate a fundamental role for clathrin function in cell polarity, growth, patterning, and organogenesis in plants.
Kleine-VehnJ, WabnikK, MartinièreA, ?angowski?, WilligK, NaramotoS, LeitnerJ, TanakaH, JakobsS, RobertS, LuschnigC, GovaertsW, HellSW, RunionsJ, FrimlJ (2011). Recycling, clustering, and endocytosis jointly maintain PIN auxin carrier polarity at the plasma membrane Mol Syst Biol7,540. DOI:10.1038/msb.2011.72PMID:22027551 [本文引用: 2] Cell polarity reflected by asymmetric distribution of proteins at the plasma membrane is a fundamental feature of unicellular and multicellular organisms. It remains conceptually unclear how cell polarity is kept in cell wall-encapsulated plant cells. We have used super-resolution and semi-quantitative live-cell imaging in combination with pharmacological, genetic, and computational approaches to reveal insights into the mechanism of cell polarity maintenance in Arabidopsis thaliana. We show that polar-competent PIN transporters for the phytohormone auxin are delivered to the center of polar domains by super-polar recycling. Within the plasma membrane, PINs are recruited into non-mobile membrane clusters and their lateral diffusion is dramatically reduced, which ensures longer polar retention. At the circumventing edges of the polar domain, spatially defined internalization of escaped cargos occurs by clathrin-dependent endocytosis. Computer simulations confirm that the combination of these processes provides a robust mechanism for polarity maintenance in plant cells. Moreover, our study suggests that the regulation of lateral diffusion and spatially defined endocytosis, but not super-polar exocytosis have primary importance for PIN polarity maintenance. Molecular Systems Biology 7: 540; published online 25 October 2011; doi:10.1038/msb.2011.72
KongXZ, HuangGQ, XiongYL, ZhaoCY, WangJ, SongXY, GiriJ, ZuoKJ (2019). IBR5 regulates leaf serrations development via modulation of the expression of PIN1 Int J Mol Sci20,4429. DOI:10.3390/ijms20184429URL [本文引用: 1]
K?e?ekP, Sk?paP, LibusJ, NaramotoS, TejosR, FrimlJ, Za?ímalováE (2009). The PIN-FORMED (PIN) protein family of auxin transporters Genome Biol10,249. DOI:10.1186/gb-2009-10-12-249URL [本文引用: 4]
KroukG, LacombeB, BielachA, Perrine-WalkerF, MalinskaK, MounierE, HoyerovaK, TillardP, LeonS, LjungK, ZazimalovaE, BenkovaE, NacryP, GojonA (2010). Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants Dev Cell18,927-937. DOI:10.1016/j.devcel.2010.05.008URL [本文引用: 1]
LiG, LiangWQ, ZhangXQ, RenHY, HuJP, BennettMJ, ZhangDB (2014). Rice actin-binding protein RMD is a key link in the auxin-actin regulatory loop that controls cell growth Proc Natl Acad Sci USA111,10377-10382. DOI:10.1073/pnas.1401680111URL [本文引用: 1]
LiHJ, LinDS, DhonuksheP, NagawaS, ChenDD, FrimlJ, ScheresB, GuoHW, YangZB (2011). Phosphorylation switch modulates the interdigitated pattern of PIN1 localization and cell expansion in Arabidopsis leaf epidermis Cell Res21,970-978. DOI:10.1038/cr.2011.49URL [本文引用: 1] Within a multicellular tissue cells may coordinately form a singular or multiple polar axes, but it is unclear whether a common mechanism governs different types of polar axis formation. The phosphorylation status of PIN proteins, which is directly affected by the PINOID (PID) protein kinase and the PP2A protein phosphatase, is known to regulate the apical-basal polarity of PIN localization in bipolar cells of roots and shoot apices. Here, we provide evidence that the phosphorylation status-mediated PIN polarity switch is widely used to modulate cellular processes in Arabidopsis including multipolar pavement cells (PC) with interdigitated lobes and indentations. The degree of PC interdigitation was greatly reduced either when the FYPP1 gene, which encodes a PP2A called phytochrome-associated serine/threonine protein phosphatase, was knocked out or when the PID gene was overexpressed (35S::PID). These genetic modifications caused PIN1 localization to switch from lobe to indentation regions. The PP2A and PID mediated switching of PIN1 localization is strikingly similar to their regulation of the apical-basal polarity switch of PIN proteins in other cells. Our findings suggest a common mechanism for the regulation of PIN1 polarity formation, a fundamental cellular process that is crucial for pattern formation both at the tissue/organ and cellular levels.
LiL, XuJ, XuZH, XueHW (2005). Brassinosteroids stimulate plant tropisms through modulation of polar auxin transport in Brassica and Arabidopsis Plant Cell17, 2738- 2753. PMID:16141452 [本文引用: 1] Brassinosteroids (BRs) are important plant growth regulators in multiple developmental processes. Previous studies have indicated that BR treatment enhanced auxin-related responses, but the underlying mechanisms remain unknown. Using (14)C-labeled indole-3-acetic acid and Arabidopsis thaliana plants harboring an auxin-responsive reporter construct, we show that the BR brassinolide (BL) stimulates polar auxin transport capacities and modifies the distribution of endogenous auxin. In plants treated with BL or defective in BR biosynthesis or signaling, the transcription of PIN genes, which facilitate functional auxin transport in plants, was differentially regulated. In addition, BL enhanced plant tropistic responses by promoting the accumulation of the PIN2 protein from the root tip to the elongation zone and stimulating the expression and dispersed localization of ROP2 during tropistic responses. Constitutive overexpression of ROP2 results in enhanced polar accumulation of PIN2 protein in the root elongation region and increased gravitropism, which is significantly affected by latrunculin B, an inhibitor of F-actin assembly. The ROP2 dominant negative mutants (35S-ROP2-DA/DN) show delayed tropistic responses, and this delay cannot be reversed by BL addition, strongly supporting the idea that ROP2 modulates the functional localization of PIN2 through regulation of the assembly/reassembly of F-actins, thereby mediating the BR effects on polar auxin transport and tropistic responses.
LiT, YanA, BhatiaN, AltinokA, AfikE, Durand-SmetP, TarrPT, SchroederJI, HeislerMG, MeyerowitzEM (2019a). Calcium signals are necessary to establish auxin transporter polarity in a plant stem cell niche Nat Commun10,726. DOI:10.1038/s41467-019-08575-6URL [本文引用: 1]
LinDL, YaoHY, JiaLH, TanJF, XuZH, ZhengWM, XueHW (2020). Phospholipase D-derived phosphatidic acid promotes root hair development under phosphorus deficiency by suppressing vacuolar degradation of PINFOR- MED2 New Phytol226,142-155. DOI:10.1111/nph.v226.1URL [本文引用: 1]
LiuSP, WangJR, WangL, WangXF, XueYH, WuP, ShouHX (2009). Adventitious root formation in rice requires OsGNOM1 and is mediated by the OsPINs family Cell Res19,1110-1119. DOI:10.1038/cr.2009.70URL [本文引用: 1]
L?fkeC, LuschnigC, Kleine-VehnJ (2013). Posttranslational modification and trafficking of PIN auxin efflux carriers Mech Dev130,82-94. DOI:10.1016/j.mod.2012.02.003URL [本文引用: 1]
LuGW, ConevaV, CasarettoJA, YingS, MahmoodK, LiuF, NambaraE, BiYM, RothsteinSJ (2015). OsPIN5b modulates rice ( Oryza sativa) plant architecture and yield by changing auxin homeostasis, transport and distribution Plant J83,913-925. DOI:10.1111/tpj.2015.83.issue-5URL [本文引用: 2]
LuschnigC, GaxiolaRA, GrisafiP, FinkGR (1998). EIR1, a root-specific protein involved in auxin transport, is required for gravitropism in Arabidopsis thaliana Genes Dev12, 2175-2187. DOI:10.1101/gad.12.14.2175URL [本文引用: 1]
MattssonJ, CkurshumovaW, BerlethT (2003). Auxin signaling in Arabidopsis leaf vascular development Plant Physiol131,1327-1339. PMID:12644682 [本文引用: 1] A number of observations have implicated auxin in the formation of vascular tissues in plant organs. These include vascular strand formation in response to local auxin application, the effects of impaired auxin transport on vascular patterns and suggestive phenotypes of Arabidopsis auxin response mutants. In this study, we have used molecular markers to visualize auxin response patterns in developing Arabidopsis leaves as well as Arabidopsis mutants and transgenic plants to trace pathways of auxin signal transduction controlling the expression of early procambial genes. We show that in young Arabidopsis leaf primordia, molecular auxin response patterns presage sites of procambial differentiation. This is the case not only in normal development but also upon experimental manipulation of auxin transport suggesting that local auxin signals are instrumental in patterning Arabidopsis leaf vasculature. We further found that the activity of the Arabidopsis gene MONOPTEROS, which is required for proper vascular differentiation, is also essential in a spectrum of auxin responses, which include the regulation of rapidly auxin-inducible AUX/IAA genes, and discovered the tissue-specific vascular expression profile of the class I homeodomain-leucine zipper gene, AtHB20. Interestingly, MONOPTEROS activity is a limiting factor in the expression of AtHB8 and AtHB20, two genes encoding transcriptional regulators expressed early in procambial development. Our observations connect general auxin signaling with early controls of vascular differentiation and suggest molecular mechanisms for auxin signaling in patterned cell differentiation.
MravecJ, Sk?paP, BaillyA, HoyerováK, K?e?ekP, BielachA, Petrá?ekJ, ZhangJ, GaykovaV, StierhofYD, DobrevPI, SchwarzerováK, Rol?íkJ, SeifertováD, LuschnigC, BenkováE, Za?ímalováE, GeislerM, FrimlJ (2009). Subcellular homeostasis of phytohormone auxin is mediated by the ER-localized PIN5 transporter Nature459,1136-1140. DOI:10.1038/nature08066URL [本文引用: 3]
MüllerA, GuanC, G?lweilerL, T?nzlerP, HuijserP, MarchantA, ParryG, BennettM, WismanE, PalmeK (1998). AtPIN2 defines a locus of Arabidopsis for root gravitropism control EMBO J17,6903-6911. DOI:10.1093/emboj/17.23.6903URL [本文引用: 1]
OkadaK, ShimuraY (1990). Reversible root tip rotation in Arabidopsis seedlings induced by obstacle-touching stimu- lus Science250,274-276. [本文引用: 1]
OkadaK, UedaJ, KomakiMK, BellCJ, ShimuraY (1991). Requirement of the auxin polar transport system in early stages of Arabidopsis floral bud formation Plant Cell3,677-684. [本文引用: 1]
PanJW, FujiokaS, PengJL, ChenJH, LiGM, ChenRJ (2009). The E3 ubiquitin ligase SCFTIR1/AFB and membrane sterols play key roles in auxin regulation of endocytosis, recycling, and plasma membrane accumulation of the auxin efflux transporter PIN2 in Arabidopsis thaliana Plant Cell21,568-580. DOI:10.1105/tpc.108.061465URL [本文引用: 1]
PaponovIA, BudnykV, PaponovM, TealeW, PalmeK (2020). Butylated Hydroxytoluene (BHT) inhibits PIN1 exocytosis from BFA compartments in Arabidopsis roots Front Plant Sci11,393. DOI:10.3389/fpls.2020.00393PMID:32322261 [本文引用: 1] The activity of polarly localized PIN-FORMED (PIN) auxin efflux carriers contributes to the formation of auxin gradients which guide plant growth, development, and tropic responses. Both the localization and abundance of PIN proteins in the plasma membrane depend on the regulation of PIN trafficking through endocytosis and exocytosis and are influenced by many external and internal stimuli, such as reactive oxygen species, auxin transport inhibitors, flavonoids and plant hormones. Here, we investigated the regulation of endosomal PIN cycling by using a Brefeldin A (BFA) assay to study the effect of a phenolic antioxidant ionol, butylated hydroxytoluene (BHT), on the endocytosis and exocytosis of PIN1 and PIN2. BHT is one of the most widely used antioxidants in the food and feed industries, and as such is commonly released into the environment; however, the effect of BHT on plants remains poorly characterized. Preincubation of Arabidopsis seedlings with BHT before BFA treatment strongly enhanced the internalization of PIN1 into BFA compartments. After the simultaneous application of BHT and NAA, the NAA effect dominated PIN internalization suggesting the BHT effect occurred downstream to that of NAA. Washing seedlings with BHT after BFA treatment prevented the release of PIN1 from BFA compartments back to the plasma membrane, indicating that BHT application inhibited PIN1 exocytosis. Overall rates of PIN2 internalization were less pronounced than those of PIN1 in seedlings pre-incubated with BHT before BFA treatment, and PIN2 exocytosis was not inhibited by BHT, indicating a specific activity of BHT on PIN1 exocytosis. Comparison of BHT activity with other potential stimuli of PIN1 and PIN2 trafficking [e.g., HO (ROS), salt stress, reduced glutathione (GSH), dithiothreitol (DTT), and flavonoids] showed that BHT has a new activity distinct from the activities of other regulators of PIN trafficking. The findings support BHT as a potentially interesting pharmacological tool for dissecting PIN trafficking and auxin transport.Copyright ? 2020 Paponov, Budnyk, Paponov, Teale and Palme.
PeerWA, BlakesleeJJ, YangHB, MurphyAS (2011). Seven things we think we know about auxin transport Mol Plant4,487-504. DOI:10.1093/mp/ssr034URL [本文引用: 1] Polar transport of the phytohormone auxin and the establishment of localized auxin maxima regulate embryonic development, stem cell maintenance, root and shoot architecture, and tropic growth responses. The past decade has been marked by dramatic progress in efforts to elucidate the complex mechanisms by which auxin transport regulates plant growth. As the understanding of auxin transport regulation has been increasingly elaborated, it has become clear that this process is involved in almost all plant growth and environmental responses in some way. However, we still lack information about some basic aspects of this fundamental regulatory mechanism. In this review, we present what we know (or what we think we know) and what we do not know about seven auxin-regulated processes. We discuss the role of auxin transport in gravitropism in primary and lateral roots, phototropism, shoot branching, leaf expansion, and venation. We also discuss the auxin reflux/fountain model at the root tip, flavonoid modulation of auxin transport processes, and outstanding aspects of post-translational regulation of auxin transporters. This discussion is not meant to be exhaustive, but highlights areas in which generally held assumptions require more substantive validation.
PéretB, SwarupK, FergusonA, SethM, YangYD, DhondtS, JamesN, CasimiroI, PerryP, SyedA, YangHB, ReemmerJ, VenisonE, HowellsC, Perez-AmadorMA, YunJ, AlonsoJ, BeemsterGTS, LaplazeL, MurphyA, BennettMJ, NielsenE, SwarupR (2012). AUX/ LAX genes encode a family of auxin influx transporters that perform distinct functions during Arabidopsis development Plant Cell24,2874-2885. DOI:10.1105/tpc.112.097766URL [本文引用: 1] Auxin transport, which is mediated by specialized influx and efflux carriers, plays a major role in many aspects of plant growth and development. AUXIN1 (AUX1) has been demonstrated to encode a high-affinity auxin influx carrier. In Arabidopsis thaliana, AUX1 belongs to a small multigene family comprising four highly conserved genes (i.e., AUX1 and LIKE AUX1 [LAX] genes LAX1, LAX2, and LAX3). We report that all four members of this AUX/LAX family display auxin uptake functions. Despite the conservation of their biochemical function, AUX1, LAX1, and LAX3 have been described to regulate distinct auxin-dependent developmental processes. Here, we report that LAX2 regulates vascular patterning in cotyledons. We also describe how regulatory and coding sequences of AUX/LAX genes have undergone subfunctionalization based on their distinct patterns of spatial expression and the inability of LAX sequences to rescue aux1 mutant phenotypes, respectively. Despite their high sequence similarity at the protein level, transgenic studies reveal that LAX proteins are not correctly targeted in the AUX1 expression domain. Domain swapping studies suggest that the N-terminal half of AUX1 is essential for correct LAX localization. We conclude that Arabidopsis AUX/LAX genes encode a family of auxin influx transporters that perform distinct developmental functions and have evolved distinct regulatory mechanisms.
Petrá?ekJ, FrimlJ (2009). Auxin transport routes in plant development Development136,2675-2688. DOI:10.1242/dev.030353URL [本文引用: 1]
RanochaP, DimaO, NagyR, FeltenJ, Corratgé-FaillieC, NovákO, MorreelK, LacombeB, MartinezY, PfrunderS, JinX, RenouJP, ThibaudJB, LjungK, FischerU, MartinoiaE, BoerjanW, GoffnerD (2013). Arabidopsis WAT1 is a vacuolar auxin transport facilitator required for auxin homoeostasis Nat Commun4,2625. DOI:10.1038/ncomms3625URL [本文引用: 1]
RigóG, AyaydinF, TietzO, ZsigmondL, KovácsH, PáyA, SalchertK, DarulaZ, MedzihradszkyKF, SzabadosL, PalmeK, KonczC, Csépl?á (2013). Inactivation of plasma membrane-localized CDPK-RELATED KINASE5 decelerates PIN2 exocytosis and root gravitropic response in Arabidopsis Plant Cell25, 1592-1608. DOI:10.1105/tpc.113.110452URL [本文引用: 1] CRK5 is a member of the Arabidopsis thaliana Ca2+/calmodulin-dependent kinase-related kinase family. Here, we show that inactivation of CRK5 inhibits primary root elongation and delays gravitropic bending of shoots and roots. Reduced activity of the auxin-induced DR5-green fluorescent protein reporter suggests that auxin is depleted from crk5 root tips. However, no tip collapse is observed and the transcription of genes for auxin biosynthesis, AUXIN TRANSPORTER/AUXIN TRANSPORTER-LIKE PROTEIN (AUX/LAX) auxin influx, and PIN-FORMED (PIN) efflux carriers is unaffected by the crk5 mutation. Whereas AUX1, PIN1, PIN3, PIN4, and PIN7 display normal localization, PIN2 is depleted from apical membranes of epidermal cells and shows basal to apical relocalization in the cortex of the crk5 root transition zone. This, together with an increase in the number of crk5 lateral root primordia, suggests facilitated auxin efflux through the cortex toward the elongation zone. CRK5 is a plasma membrane-associated kinase that forms U-shaped patterns facing outer lateral walls of epidermis and cortex cells. Brefeldin inhibition of exocytosis stimulates CRK5 internalization into brefeldin bodies. CRK5 phosphorylates the hydrophilic loop of PIN2 in vitro, and PIN2 shows accelerated accumulation in brefeldin bodies in the crk5 mutant. Delayed gravitropic response of the crk5 mutant thus likely reflects defective phosphorylation of PIN2 and deceleration of its brefeldin-sensitive membrane recycling.
RomanG, LubarskyB, KieberJJ, RothenbergM, EckerJR (1995). Genetic analysis of ethylene signal transduction in Arabidopsis thaliana: five novel mutant loci integrated into a stress response pathway Genetics139,1393-1409. DOI:10.1093/genetics/139.3.1393URL [本文引用: 1]
RosqueteMR, WaidmannS, Kleine-VehnJ,(2018). PIN7 auxin carrier has a preferential role in terminating radial root expansion in Arabidopsis thaliana Int J Mol Sci19, 1238. DOI:10.3390/ijms19041238URL [本文引用: 1]
R??i?kaK, LjungK, VannesteS, PodhorskáR, BeeckmanT, FrimlJ, BenkováE (2007). Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution Plant Cell19, 2197- 2212. DOI:10.1105/tpc.107.052126URL [本文引用: 1]
R??i?kaK, ?imá?kováM, DuclercqJ, Petrá?ekJ, Za?ímalováE, SimonS, FrimlJ, VanMontagu MCE, BenkováE (2009). Cytokinin regulates root meristem activity via modulation of the polar auxin transport Proc Natl Acad Sci USA106,4284-4289. DOI:10.1073/pnas.0900060106URL [本文引用: 1]
SabatiniS, BeisD, WolkenfeltH, MurfettJ, GuilfoyleT, MalamyJ, BenfeyP, LeyserO, BechtoldN, WeisbeekP, ScheresB (1999). An auxin-dependent distal organi- zer of pattern and polarity in the Arabidopsis root Cell99,463-472. DOI:10.1016/S0092-8674(00)81535-4URL [本文引用: 1]
Sancho-AndrésG, Soriano-OrtegaE, GaoCJ, Berna- bé-OrtsJM, NarasimhanM, MüllerAO, TejosR, JiangL, FrimlJ, AnientoF, MarcoteMJ (2016). Sorting motifs involved in the trafficking and localization of the PIN1 auxin efflux carrier Plant Physiol171,1965-1982. DOI:10.1104/pp.16.00373URL [本文引用: 1]
SantinF, BhogaleS, FantinoE, GrandellisC, BanerjeeAK, UlloaRM (2017). Solanum tuberosum StCDPK1 is regulated by miR390 at the posttranscriptional level and phosphorylates the auxin efflux carrier StPIN4 in vitro, a potential downstream target in potato development Phy- siol Plant159,244-261. [本文引用: 1]
ShenCJ, BaiYH, WangSK, ZhangSN, WuYR, ChenM, JiangDA, QiYH (2010). Expression profile of PIN, AUX/LAX and PGP auxin transporter gene families in Sorghum bicolor under phytohormone and abiotic stress FEBS J277,2954-2969. DOI:10.1111/j.1742-4658.2010.07706.xURL [本文引用: 3]
SidlerM, HassaP, HasanS, RingliC, DudlerR (1998). Involvement of an ABC transporter in a developmental pathway regulating hypocotyl cell elongation in the light Plant Cell10,1623-1636. DOI:10.1105/tpc.10.10.1623URL [本文引用: 1]
?imá?kováM, O’BrienJA, KhanM, VanNoorden G, ?tv?sK, VietenA, DeClercq I, VanHaperen JMA, CuestaC, HoyerováK, VannesteS, MarhavyP, WabnikK, VanBreusegem F, NowackM, MurphyA, FrimlJ, WeijersD, BeeckmanT, BenkováE (2015). Cytokinin response factors regulate PIN-FORMED auxin transpor- ters Nat Commun6,8717. DOI:10.1038/ncomms9717PMID:26541513 [本文引用: 1] Simaskova, Maria; O'Brien, Jose Antonio; Van Noorden, Giel; De Clercq, Inge; Vanneste, Steffen; Van Breusegem, Frank; Nowack, Moritz; Beeckman, Tom VIB, Dept Plant Syst Biol, B-9052 Ghent, Belgium. Simaskova, Maria; O'Brien, Jose Antonio; Van Noorden, Giel; De Clercq, Inge; Vanneste, Steffen; Van Breusegem, Frank; Nowack, Moritz; Beeckman, Tom Univ Ghent, Dept Plant Biotechnol & Bioinformat, B-9052 Ghent, Belgium. Khan, Mamoona; Oetvoes, Krisztina; Cuesta, Candela; Marhavy, Peter; Wabnik, Krzysztof; Friml, Jiri; Benkova, Eva IST Austria, A-3400 Klosterneuburg, Austria. Vieten, Anne Univ Tubingen, Ctr Mol Biol Plants, Dept Dev Genet, D-72076 Tubingen, Germany. Van Haperen, Johanna Maria Adriana; Weijers, Dolf Wageningen Univ, Biochem Lab, NL-6703 HA Wageningen, Netherlands. Hoyerova, Klara ASCR, Inst Expt Bot, Prague 16502, Czech Republic. Murphy, Angus Univ Maryland, Dept Plant Sci & Landscape Architecture, College Pk, MD 20742 USA.
SunHW, TaoJY, BiY, HouMM, LouJJ, ChenXN, ZhangXH, LuoL, XieXN, YoneyamaK, ZhaoQZ, XuGH, ZhangYL (2018). OsPIN1b is involved in rice seminal root elongation by regulating root apical meristem activity in response to low nitrogen and phosphate Sci Rep8,13014. DOI:10.1038/s41598-018-29784-xURL [本文引用: 1]
SunJQ, ChenQ, QiLL, JiangHL, LiSY, XuYX, LiuF, ZhouWK, PanJW, LiXG, PalmeK, LiCY (2011). Jasmonate modulates endocytosis and plasma memb- rane accumulation of the Arabidopsis PIN2 protein New Phytol191,360-375. DOI:10.1111/nph.2011.191.issue-2URL [本文引用: 1]
TejosR, SauerM, VannesteS, Palacios-GomezM, LiHJ, HeilmannM, VanWijk R, VermeerJEM, HeilmannI, MunnikT, FrimlJ (2014). Bipolar plasma membrane distribution of phosphoinositides and their requirement for auxin-mediated cell polarity and patterning in Arabidopsis Plant Cell26, 2114-2128. DOI:10.1105/tpc.114.126185URL [本文引用: 1] Cell polarity manifested by asymmetric distribution of cargoes, such as receptors and transporters, within the plasma membrane (PM) is crucial for essential functions in multicellular organisms. In plants, cell polarity (re) establishment is intimately linked to patterning processes. Despite the importance of cell polarity, its underlying mechanisms are still largely unknown, including the definition and distinctiveness of the polar domains within the PM. Here, we show in Arabidopsis thaliana that the signaling membrane components, the phosphoinositides phosphatidylinositol 4-phosphate (PtdIns4P) and phosphatidylinositol 4,5-bisphosphate [PtdIns(4,5)P-2] as well as PtdIns4P 5-kinases mediating their interconversion, are specifically enriched at apical and basal polar plasma membrane domains. The PtdIns4P 5-kinases PIP5K1 and PIP5K2 are redundantly required for polar localization of specifically apical and basal cargoes, such as PIN-FORMED transporters for the plant hormone auxin. As a consequence of the polarity defects, instructive auxin gradients as well as embryonic and postembryonic patterning are severely compromised. Furthermore, auxin itself regulates PIP5K transcription and PtdIns4P and PtdIns(4,5)P-2 levels, in particular their association with polar PM domains. Our results provide insight into the polar domain-delineating mechanisms in plant cells that depend on apical and basal distribution of membrane lipids and are essential for embryonic and postembryonic patterning.
UtsunoK, ShikanaiT, YamadaY, HashimotoT (1998). AGR, an Agravitropic locus of Arabidopsis thaliana, encodes a novel membrane-protein family member Plant Cell Physiol39,1111-1118. PMID:9871369 [本文引用: 1] Mutations in the Agr locus of Arabidopsis thaliana impair the root gravitropic response. Root growth of agr mutants is moderately resistant to ethylene and to an auxin transport inhibitor. Vertically placed agr roots grow into agar medium containing IAA or naphthalene-1-acetic acid, but not into medium containing 2,4-D. Positional cloning showed that AGR encodes a root-specific member of a novel membrane-protein family with limited homology to bacterial transporters.
VietenA, SauerM, BrewerPB, FrimlJ (2007). Molecular and cellular aspects of auxin-transport-mediated deve- lopment Trends Plant Sci12,160-168. DOI:10.1016/j.tplants.2007.03.006URL [本文引用: 1]
VietenA, VannesteS, WisniewskaJ, BenkováE, BenjaminsR, BeeckmanT, LuschnigC, FrimlJ (2005). Functional redundancy of PIN proteins is accompanied by auxin-dependent cross-regulation of PIN expression De- velopment132,4521-4531. [本文引用: 1]
WangJR, HuH, WangGH, LiJ, ChenJY, WuP (2009). Expression of PIN genes in rice ( Oryza sativa L.): tissue specificity and regulation by hormones Mol Plant2,823- 831. DOI:10.1093/mp/ssp023URL [本文引用: 2]
WangPP, ShenL, GuoJH, JingW, QuYN, LiWY, BiRR, XuanW, ZhangQ, ZhangWH (2019). Phosphatidic acid directly regulates PINOID-dependent phosphorylation and activation of the PIN-FORMED2 auxin efflux transporter in response to salt stress Plant Cell31,250-271. DOI:10.1105/tpc.18.00528URL [本文引用: 1]
WangSK, ShenCJ, ZhangSN, XuYX, JiangDA (2011). Analysis of subcellular localization of auxin carriers PIN, AUX/LAX and PGP in Sorghum bicolor Plant Signal Behav6, 2023-2025. DOI:10.4161/psb.6.12.17968URL [本文引用: 1]
WuHM, XieDJ, TangZS, ShiDQ, YangWC (2020). PINOID regulates floral organ development by modulating auxin transport and interacts with MADS16 in rice Plant Biotechnol J18,1778-1795. DOI:10.1111/pbi.v18.8URL [本文引用: 1]
XiDD, ChenX, WangYX, ZhongRL, HeJM, ShenJB, MingF (2019). Arabidopsis ANAC092 regulates auxin- mediated root development by binding to the ARF8 and PIN4 promoters J Integr Plant Biol61,1015-1031. DOI:10.1111/jipb.12735URL [本文引用: 1] Auxin is an important plant hormone that is essential for growth and development due to its effects on organogenesis, morphogenesis, tropisms, and apical dominance. The functional diversity of auxin highlights the importance of its biosynthesis, transport, and associated responses. In this study, we show that a NAC transcription factor, ANAC092 (also named AtNAC2 and ORESARA1), known to positively regulate leaf senescence and contribute to abiotic stress responses, also affects primary root development. Plants overexpressing <italic>ANAC092</italic> had altered root meristem lengths and shorter primary roots compared with the wild‐type control. Additionally, expression of the <italic>proANAC092::GUS</italic> was strongly induced by indole‐3‐acetic acid. Quantitative real‐time RT‐PCR (qRT‐PCR) analysis revealed that the <italic>YUCCA2, PIN</italic>, and <italic>ARF</italic> expression levels were downregulated in <italic>ANAC092</italic>‐overexpressing plants. Moreover, yeast one‐hybrid and chromatin immunoprecipitation assays confirmed that ANAC092 binds to the promoters of <italic>AUXIN RESPONSE FACTOR 8</italic> (<italic>ARF8</italic>) and <italic>PIN‐FORMED</italic> 4 (<italic>PIN4</italic>). Furthermore, a dual‐luciferase assay indicated that ANAC092 decreases <italic>ARF8</italic> and <italic>PIN4</italic> promoter activities. We also applied a CRISPR/Cas9 system to mutate <italic>ANAC092</italic>. The roots of three of the analyzed mutants were longer than normal. Collectively, our findings indicate that ANAC092 negatively affects root development by controlling the auxin pathway.
XuHW, MoYW, WangW, WangH, WangZ (2014). OsPIN1a gene participates in regulating negative phototropism of rice roots Rice Sci21,83-89. DOI:10.1016/S1672-6308(13)60168-7URL [本文引用: 2] The complete open reading frame of OsPIN1a was amplified through reverse transcriptase- polymerase chain reaction (RT-PCR) based on the sequence deposited in GenBank to explore the relationship between the auxin efflux protein OsPIN1a and the negative phototropism of rice roots. Sequencing results showed that the GC content of OsPIN1a was 65.49%. The fusion expression vector pCAMBIA-1301-OsPIN1a::GFP containing the OsPIN1a gene and a coding green fluorescent protein (gfp) gene was constructed. The fusion vector was transferred into onion epidermal cells by Agrobacterium tumefaciens transformation. The transient expression of OsPIN1a-GFP was mainly located in the nucleus and cell membrane. Moreover, the transgenic plants were obtained by Agrobacterium-mediated genetic transformation. Molecular detection performed by using PCR and β-glucuronidase staining showed that the target construct was integrated into the genome of rice. The negative phototropic curvatures of the transgenic rice roots were higher than those of the wild type. Similarly, the expression levels of OsPIN1a in the transgenic plants were considerably higher than those in the wild-type plants. These results suggest that OsPIN1a is crucial in the negative phototropic curvature of rice roots.
XuM, ZhuL, ShouHX, WuP (2005). A PIN1 family gene, OsPIN1, involved in auxin-dependent adventitious root emergence and tillering in rice Plant Cell Physiol46,1674-1681. DOI:10.1093/pcp/pci183URL [本文引用: 2]
XuWW, HuangWH (2017). Calcium-dependent protein kinases in phytohormone signaling pathways Int J Mol Sci18,2436. DOI:10.3390/ijms18112436URL [本文引用: 1]
YangHB, MurphyAS (2009). Functional expression and characterization of Arabidopsis ABCB, AUX1 and PIN auxin transporters in Schizosaccharomyces pombe Plant J59,179-191. DOI:10.1111/tpj.2009.59.issue-1URL [本文引用: 1]
YangSG, LiCL, ZhaoLM, GaoSJ, LuJX, ZhaoML, ChenCY, LiuXC, LuoM, CuiYH, YangCW, WuKQ (2015). The Arabidopsis SWI2/SNF2 chromatin remodeling ATPase BRAHMA targets directly to PINs and is required for root stem cell niche maintenance Plant Cell27, 1670- 1680. DOI:10.1105/tpc.15.00091URL [本文引用: 1]
YueRQ, TieSG, SunT, ZhangL, YangYJ, QiJS, YanSF, HanXH, WangHZ, ShenCJ (2015). Genome-wide identification and expression profiling analysis of ZmPIN, ZmPILS, ZmLAX and ZmABCB auxin transporter gene families in maize (Zea mays L.) under various abiotic stresses PLoS One10,e0118751. DOI:10.1371/journal.pone.0118751URL [本文引用: 3]
ZengYF, WenJY, ZhaoWB, WangQ, HuangWC (2020). Rational improvement of rice yield and cold tolerance by editing the three genes OsPIN5b, GS3, and OsMYB30 with the CRISPR-Cas9 system Front Plant Sci10,1663. DOI:10.3389/fpls.2019.01663URL [本文引用: 2]
ZhangJ, MazurE, BallaJ, GalleiM, KalousekP, Medve?ováZ, LiY, WangYP, PrátT, VasilevaM, Rein?hlV, ProcházkaS, HalouzkaR, TarkowskiP, LuschnigC, BrewerPB, FrimlJ (2020). Strigolactones inhibit auxin feedback on PIN-dependent auxin transport canalization Nat Commun11,3508. DOI:10.1038/s41467-020-17252-yPMID:32665554 [本文引用: 1] Directional transport of the phytohormone auxin is a versatile, plant-specific mechanism regulating many aspects of plant development. The recently identified plant hormones, strigolactones (SLs), are implicated in many plant traits; among others, they modify the phenotypic output of PIN-FORMED (PIN) auxin transporters for fine-tuning of growth and developmental responses. Here, we show in pea and Arabidopsis that SLs target processes dependent on the canalization of auxin flow, which involves auxin feedback on PIN subcellular distribution. D14 receptor- and MAX2 F-box-mediated SL signaling inhibits the formation of auxin-conducting channels after wounding or from artificial auxin sources, during vasculature de novo formation and regeneration. At the cellular level, SLs interfere with auxin effects on PIN polar targeting, constitutive PIN trafficking as well as clathrin-mediated endocytosis. Our results identify a non-transcriptional mechanism of SL action, uncoupling auxin feedback on PIN polarity and trafficking, thereby regulating vascular tissue formation and regeneration.
ZhangQ, LiJJ, ZhangWJ, YanSN, WangR, ZhaoJF, LiYJ, QiZG, SunZX, ZhuZG (2012). The putative auxin efflux carrier OsPIN3t is involved in the drought stress response and drought tolerance Plant J72,805-816. DOI:10.1111/j.1365-313X.2012.05121.xURL [本文引用: 2] The phytohormone auxin plays a critical role in plant growth and development, and its spatial distribution largely depends on the polar localization of the PIN-FORMED (PIN) auxin efflux carrier family members. In this study, we identify a putative auxin efflux carrier gene in rice, OsPIN3t, which acts in auxin polar transport but is also involved in the drought stress response in rice. We show that OsPIN3tGFP fusion proteins are localized in plasma membranes, and this subcellular localization changes under 1-N-naphthylphthalamic acid (NPA) treatment. The tissue-specific expression patterns of OsPIN3t were also investigated using a beta-glucuronidase (GUS) reporter, which showed that OsPIN3t was mainly expressed in vascular tissue. The GUS activity in OsPIN3tpro::GUS plants increased by NAA treatment and decreased by NPA treatment. Moreover, knockdown of OsPIN3t caused crown root abnormalities in the seedling stage that could be phenocopied by treatment of wild-type plants with NPA, which indicated that OsPIN3t is involved in the control of polar auxin transport. Overexpression of OsPIN3t led to improved drought tolerance, and GUS activity significantly increased when OsPIN3tpro::GUS plants were subjected to 20% polyethylene glycol stress. Taken together, these results suggest that OsPIN3t is involved in auxin transport and the drought stress response, which suggests that a polar auxin transport pathway is involved in the regulation of the response to water stress in plants.
ZhuZX, LiuY, LiuSJ, MaoCZ, WuYR, WuP (2012). A gain-of-function mutation in OsIAA11 affects lateral root development in rice Mol Plant5,154-161. DOI:10.1093/mp/ssr074URL [本文引用: 1] Lateral roots are important to plants for the uptake of nutrients and water. Several members of the Aux/IAA family have been shown to play crucial roles in lateral root development. Here, a member of the rice Aux/IAA family genes, OsIAA11 (LOC_Os03g43400), was isolated from a rice mutant defective in lateral root development. The gain-of-function mutation in OsIAA11 strictly blocks the initiation of lateral root primordia, but it does not affect crown root development. The expression of OsIAA11 is defined in root tips, lateral root caps, steles, and lateral root primordia. The auxin reporter DR5-GUS (beta-glucuronidase) was expressed at lower levels in the mutant than in wild-type, indicating that OsIAA11 is involved in auxin signaling in root caps. The transcript abundance of both OsPIN1b and OsPIN10a was diminished in root tips of the Osiaa11 mutant. Taken together, the results indicate that the gain-of-function mutation in OsIAA11 caused the inhibition of lateral root development in rice.
ZourelidouM, AbsmannerB, WellerB, BarbosaIC, WilligeBC, FastnerA, StreitV, PortSA, ColcombetJ, De LaFuente, Van BentemS, HirtH, KusterB, SchulzeWX, HammesUZ, SchwechheimerC (2014). Auxin efflux by PIN-FORMED proteins is activated by two different protein kinases, D6 PROTEIN KINASE and PINOID eLife3,e02860. DOI:10.7554/eLife.02860URL [本文引用: 2]
植物ABCB亚家族生物学功能研究进展 1 2019
... 在转录水平上, 有些上游转录因子作用于PIN基因的启动子区, 调节其表达以响应生长素内源以及外源信号变化.在生长素响应因子ARF (AUXIN RE-SPONSE FACTOR)家族中, ARF7与受其调控的FLP转录因子共同形成前馈基序以调控PIN3的转录, 介导侧根形成的早期阶段(Chen et al.2015); 细胞分裂素响应因子CRF2和CRF6作用于PIN1和PIN7启动子上的PIN特异性细胞分裂素响应元件(PCRE), 激活PIN1和PIN7的表达, 并且CRF2受ARF5的调控, 该研究表明生长素的极性运输受生长素和细胞分裂素的交叉信号调控(?imá?kováet al.2015).参与脂肪酸生物合成的AP2转录因子WRI1 (WRINK-LED1)直接靶向PIN4和PIN5的启动子, 并导致wri1-1突变体中某些PIN基因(如PIN1、PIN3和PIN5-6)的表达水平降低, 为阐明脂肪酸和PIN蛋白的关系提供了思路(Kong et al.2017).此外, 近期研究发现, NAC转录因子AtNAC2 (ANAC092)与ARF8和PIN4的启动子结合, 通过生长素途径负调控拟南芥根的生长(Xi et al.2019).此外, IBR5也影响PIN的转录活性, 表现为ibr5-3突变体中, PIN1在表皮细胞质膜和叶脉周边细胞中的分布受到影响, 叶片锯齿形成受阻(Kong et al.2019).MAKR2 (MEMBRANE ASSOCIATED KINASE REGULATOR2)作为跨膜激酶TMK1 (TRAN SMEMBRANE KINASE1)的负调控因子, 影响PIN2在根两侧的动态分布, 进而参与根的向重力性调控(Marquès-Buenoet al.2021).表观遗传方面, SWI/SNF的染色质重塑ATP酶BRM (BRAHMA)直接靶向根部“长”PIN蛋白(PIN1-PIN4和PIN7)的染色质, 功能丧失突变体brm表现为根干细胞生态位维持缺陷, 根生长发育不良, 干细胞中一些PIN基因的局部表达量降低, 从而影响生长素的分布(Yang et al.2015).生长素转运蛋白ABCB家族与PIN蛋白的互作也会调控其表达活性.有研究表明, ABCB1、ABCB19可与PIN1、PIN2互作, 共表达PIN1-ABCB1和PIN1- ABCB19的植株生长素输出速率升高(Blakeslee et al.2007; 贺祯媚等, 2019). ...
... 拟南芥PIN家族在植物整个生长发育和形态建成中发挥重要作用已有大量报道, 但对PIN1的报道最多. PIN1在整个维管组织和发育的器官中均有表达, 且是唯一能在茎尖生长点检测到的PIN家族成员.G?lweiler等(1998)和Blilou等(2005)研究发现, PIN1在根中柱表达, 将地上部合成的生长素输送至根尖, 并参与茎尖分生组织环流、器官分化、叶和花芽的形成、茎的重力响应和维管发育.PIN2主要在根尖伸长区的皮层和胚胎发生过程中表达, 参与根中生长素的向基运输和调节根的向重力性(Chen at al., 1998).PIN3主要在根冠和维管组织中表达, 调节根尖生长素的再分配, 参与侧根的早期形成、根尖钩的形成和维持, 以及调控向重力性和向光性, 参与植物的向光生长和荫蔽反应(Keuskamp et al.2010; Chen et al.2015).与PIN3类似, PIN4也在植物的向光反应和根尖钩发育中起作用, PIN4在成熟或发育中的根分生组织表达, 定位于根尖静止中心(quiescent center, QC)区域, 参与生长素的向QC运输(Friml et al.2002).PIN5定位于内质网, 主要在下胚轴伸长部位、子叶维管组织和保卫细胞中表达, 在植物发育后期表达较弱, 它通过调控细胞内生长素的动态平衡, 参与一系列发育过程(包括侧根的发生、子叶扩张、下胚轴和根的生长等)(Mravec et al.2009).PIN6定位于质膜和内质网膜, 在生长素信号介导的发育过程(如侧根和不定根的发生、根毛生长和根尖优势)中起作用.组织定位结果显示, PIN6在侧根发生原基的边缘和根茎结合处表达(Ditengou et al.2018).PIN7在根尖中的定位与PIN3有部分重叠, 且也参与根的重力响应过程(Rosquete et al.2018).PIN8定位于内质网, 在花药中特异性表达, 控制花粉、雄配子体和孢子体的发育.Ding等(2012)和Lee等(2020)研究表明, 在侧根发生后期可检测到PIN8在根尖分生组织基部和伸长/成熟区的韧皮部中呈点状表达, 表明其也参与侧根的发生, 但与质膜定位的PIN引导胞间生长素回流的机制不同, PIN8很可能通过调节细胞内生长素的转运来影响侧根的发育过程.虽然PIN家族成员均有各自功能, 但它们在植物生理过程中相互协调、分工合作来调节生长素的分布与流向(Leyser, 2006; K?e?eket al.2009). ...
Coevolving MAPK and PID phosphosites indicate an ancient environmental control of PIN auxin transpor- ters in land plants 2 2018
... 在长亲水环上还含有多个保守的磷酸化位点, 它们是调节PIN转运和亚细胞极性的某些激酶的靶点(图1).对拟南芥的研究表明, 在3个高度保守的TPRXS基序中, 丝氨酸残基S1-S3 (S231、S252和S290)是PIDs (PINOID)激酶和D6蛋白激酶(D6PKs) 的靶点, 与S1-S3相邻的3个苏氨酸残基(T227、T248和T286)是促分裂素原活化激酶(mitogen activated protein kinases, MAPK)的靶点(Huang et al.2010; Zourelidou et al.2014; Dory et al.2018).此外, 在长亲水环上不保守的丝氨酸残基S337和苏氨酸残基T340也被证明参与PIN的磷酸化调控.MAPK能通过磷酸化S337来调节拟南芥初生茎中生长素的极性运输, 但靶向T340位点的激酶目前未知(Ganguly et al.2012; Jia et al.2016). ...
... 拟南芥基因组共编码20个MAPK和10个MAPK 激酶(MKKs), MKK-MAPK信号级联参与多种植物发育进程的调控.MKK7-MAPK3/6级联通过磷酸化S337位点使PIN在下胚轴和茎中去极化, 从而影响茎中生长素的极性运输(Jia et al.2016).MAPK4和MAPK6能够磷酸化T227、T248和T286, 这3个位点与S1-S3相邻, 说明AGCVIII和MAPK蛋白激酶很可能通过磷酸化PIN蛋白上的丝氨酸和苏氨酸位点介导PIN的极性(Dory et al.2018). ...
Auxin activity: past, present, and future 1 2015
... 生长素是最早被发现的植物激素, 参与植物生长发育的各个阶段(Enders and Strader, 2015).生长素在植物组织和器官中建立的不对称分布是其行使功能的重要基础.例如, 调控根分生组织的活性、器官的形成和维管组织分化等, 是长期适应的结果(Sabatini et al.1999; Benkováet al.2003; Mattsson et al.2003).生长素从合成部位(如幼叶和顶芽)运输到作用部位主要有2条途径: 一是在整个植株范围内, 以不固定方向被动运输的方式, 通过韧皮部完成长距离运输; 二是需要载体参与的细胞与细胞之间短距离极性运输, 包括从茎尖向根的单向运输和组织间的局部单向运输( Petrá?ek and Friml,2009).极性运输对建立和维持生长素的浓度梯度至关重要, 进而特异性调节植物的生长发育及其对外界环境变化的响应. ...
PIN polarity maintenance by the cell wall in Arabidopsis 1 2011
Regulation of the polarity of protein trafficking by phosphorylation 1 2012
... 在长亲水环上还含有多个保守的磷酸化位点, 它们是调节PIN转运和亚细胞极性的某些激酶的靶点(图1).对拟南芥的研究表明, 在3个高度保守的TPRXS基序中, 丝氨酸残基S1-S3 (S231、S252和S290)是PIDs (PINOID)激酶和D6蛋白激酶(D6PKs) 的靶点, 与S1-S3相邻的3个苏氨酸残基(T227、T248和T286)是促分裂素原活化激酶(mitogen activated protein kinases, MAPK)的靶点(Huang et al.2010; Zourelidou et al.2014; Dory et al.2018).此外, 在长亲水环上不保守的丝氨酸残基S337和苏氨酸残基T340也被证明参与PIN的磷酸化调控.MAPK能通过磷酸化S337来调节拟南芥初生茎中生长素的极性运输, 但靶向T340位点的激酶目前未知(Ganguly et al.2012; Jia et al.2016). ...
A critical view on ABC transporters and their interacting partners in auxin transport 1 2017
... 生长素的极性运输需要载体, 目前已证实有家族蛋白具有生长素转运功能(Rubery and Sheldrake, 1974; Bennett et al.1996; G?lweileret al.1998; Sidler et al.1998).生长素在细胞间的运输主要依赖3种膜定位蛋白, 分别为生长素输入载体AUX/LAX (AUXIN1/LIKE-AUX1)、生长素输出载体PIN (PIN- FORMED)以及ABCB/PGP (ATP-Binding Cassette subfamily B/P-glycoprotein)转运体(Péretet al.2012; Geisler et al.2017; Zhou and Luo, 2018).此外, 在拟南芥(Arabidopsis thaliana)中, 硝酸盐转运体NRT1.1也具有转运生长素的能力, 低氮条件下其通过促进生长素的向基运输抑制侧根的生长(Krouk et al.2010).生长素在细胞内的运输则依赖定位于胞内的PILS (PIN-Like)蛋白家族、液泡生长素转运蛋白WAT1 (WALLS ARE THIN 1)以及具有短亲水环结构的部分PIN蛋白(Barbez et al.2012; Ranocha et al.2013). ...
The Arabidopsis GNOM ARF-GEF mediates endosomal recycling, auxin transport, and auxin-dependent plant growth 1 2003
OsPIN1与AtPIN1类似, 主要在维管组织和不定根原基中表达, 对水稻不定根发育、分蘖、地下部和地上部的比例具有重要调控作用, 下调OsPIN1的表达明显抑制水稻的分蘖数和不定根发生(Xu et al.2005).对OsPIN1的4个基因研究发现, OsPIN1a-d在根中的表达量相对高于其它组织, 它们的单突变体较野生型不具有明显表型, 而pin1a/pin1b双突变体表现出地上部和初生根变短、根毛变长, 以及重力性缺失表型, pin1c/pin1d双突变植株则表现出类似拟南芥pin1突变体的针状花序表型, 说明OsPIN1a和OsPIN1b主要参与水稻根和茎的发育, 而OsPIN1c和OsPIN1d参与穗的形成, 并且它们之间存在功能冗余(Li et al.2019b).也有研究表明, OsPIN1a在水稻根的负趋光性调控中起作用(Xu et al.2014).OsPIN2主要在根中分布, 极性分布于根尖的表皮和皮层区, 过表达OsPIN2可增强生长素从地上部向根茎结合部的转运, 从而促进水稻分蘖, 并使分蘖角度增大、株高降低(Chen et al.2012).OsPIN5b在多种组织中均有表达, 亚细胞定位于内质网, 参与生长素的胞内转运和平衡, 其过表达株系的分蘖数、株高、穗长和产量均降低, 而ospin5b突变体的穗长变长, 说明OsPIN5b通过生长素信号调控水稻的株型和产量(Lu et al.2015; Zeng et al.2020).OsPIN9主要在分蘖芽及其维管组织中表达,其亲水环长度介于“长”PIN蛋白与“短”PIN蛋白之间.ospin9突变体的分蘖数减少, 过表达株系的表型则相反.大田试验结果显示, 低氮条件下OsPIN9过表达株系的分蘖数和产量均高于高氮条件(Hou et al.2021).OsPIN10a (OsPIN3t/OsPIN3a)主要在维管组织中表达, 亚细胞定位于质膜, 其过表达株系的抗旱能力增强, 说明OsPIN10a所介导的生长素极性运输与植株抗旱响应有关(Miyashita et al.2010; Zhang et al.2012).OsPIN8基因的功能尚未见报道, 系统发育分析表明其与拟南芥的AtPIN8相近, 蛋白结构上缺少中心亲水环, 推测与拟南芥的PIN8一样定位于内质网, 介导生长素的胞内运输, 但尚需实验验证.对比水稻和拟南芥PIN家族的生物学功能, 发现PIN在水稻地上部株型的相关性状(如分蘖数、分蘖角和株高)调控中发挥重要作用, 并且影响作物的产量性状, 继续推进水稻等禾谷类作物中PIN蛋白的研究具有重要意义. ...
... OsPIN1与AtPIN1类似, 主要在维管组织和不定根原基中表达, 对水稻不定根发育、分蘖、地下部和地上部的比例具有重要调控作用, 下调OsPIN1的表达明显抑制水稻的分蘖数和不定根发生(Xu et al.2005).对OsPIN1的4个基因研究发现, OsPIN1a-d在根中的表达量相对高于其它组织, 它们的单突变体较野生型不具有明显表型, 而pin1a/pin1b双突变体表现出地上部和初生根变短、根毛变长, 以及重力性缺失表型, pin1c/pin1d双突变植株则表现出类似拟南芥pin1突变体的针状花序表型, 说明OsPIN1a和OsPIN1b主要参与水稻根和茎的发育, 而OsPIN1c和OsPIN1d参与穗的形成, 并且它们之间存在功能冗余(Li et al.2019b).也有研究表明, OsPIN1a在水稻根的负趋光性调控中起作用(Xu et al.2014).OsPIN2主要在根中分布, 极性分布于根尖的表皮和皮层区, 过表达OsPIN2可增强生长素从地上部向根茎结合部的转运, 从而促进水稻分蘖, 并使分蘖角度增大、株高降低(Chen et al.2012).OsPIN5b在多种组织中均有表达, 亚细胞定位于内质网, 参与生长素的胞内转运和平衡, 其过表达株系的分蘖数、株高、穗长和产量均降低, 而ospin5b突变体的穗长变长, 说明OsPIN5b通过生长素信号调控水稻的株型和产量(Lu et al.2015; Zeng et al.2020).OsPIN9主要在分蘖芽及其维管组织中表达,其亲水环长度介于“长”PIN蛋白与“短”PIN蛋白之间.ospin9突变体的分蘖数减少, 过表达株系的表型则相反.大田试验结果显示, 低氮条件下OsPIN9过表达株系的分蘖数和产量均高于高氮条件(Hou et al.2021).OsPIN10a (OsPIN3t/OsPIN3a)主要在维管组织中表达, 亚细胞定位于质膜, 其过表达株系的抗旱能力增强, 说明OsPIN10a所介导的生长素极性运输与植株抗旱响应有关(Miyashita et al.2010; Zhang et al.2012).OsPIN8基因的功能尚未见报道, 系统发育分析表明其与拟南芥的AtPIN8相近, 蛋白结构上缺少中心亲水环, 推测与拟南芥的PIN8一样定位于内质网, 介导生长素的胞内运输, 但尚需实验验证.对比水稻和拟南芥PIN家族的生物学功能, 发现PIN在水稻地上部株型的相关性状(如分蘖数、分蘖角和株高)调控中发挥重要作用, 并且影响作物的产量性状, 继续推进水稻等禾谷类作物中PIN蛋白的研究具有重要意义. ...
... 水稻PIN基因的表达受多种途径调控, 尽管目前已有一些研究报道, 但大部分并未阐明具体调控机制.Sun等(2018)研究发现, 低磷、低氮和GR24 (类似于SLs)等处理下, OsPIN1b的表达明显下调.进一步研究发现, OsPIN1b作为NO和SLs的下游介质, 在低磷和低氮条件下通过调节生长素的运输来激活水稻根尖分生区的活性.此外, 铵态氮(NH4+)在RNA和蛋白水平上诱导OsPIN9的表达来影响分蘖数量, 说明水稻分蘖以及根尖生长受生长素运输和氮磷等营养元素供应的协同调控(Hou et al.2021).osiaa11突变体的侧根原基发育受阻, 且OsPIN1b和OsPIN- 10a的表达下调, 暗示OsIAA11可能通过调节生长素运输载体的表达来影响植株的发育(Zhu et al.2012). ...
Phosphorylation of conserved PIN motifs directs Arabidopsis PIN1 polarity and auxin transport 1 2010
... 在长亲水环上还含有多个保守的磷酸化位点, 它们是调节PIN转运和亚细胞极性的某些激酶的靶点(图1).对拟南芥的研究表明, 在3个高度保守的TPRXS基序中, 丝氨酸残基S1-S3 (S231、S252和S290)是PIDs (PINOID)激酶和D6蛋白激酶(D6PKs) 的靶点, 与S1-S3相邻的3个苏氨酸残基(T227、T248和T286)是促分裂素原活化激酶(mitogen activated protein kinases, MAPK)的靶点(Huang et al.2010; Zourelidou et al.2014; Dory et al.2018).此外, 在长亲水环上不保守的丝氨酸残基S337和苏氨酸残基T340也被证明参与PIN的磷酸化调控.MAPK能通过磷酸化S337来调节拟南芥初生茎中生长素的极性运输, 但靶向T340位点的激酶目前未知(Ganguly et al.2012; Jia et al.2016). ...
Genome-wide analysis of the PIN auxin efflux carrier gene family in coffee 1 2020
... 根据化学渗透偶联学说, 生长素输出载体在细胞膜上的不对称分布是生长素极性运输的关键(Friml, 2010; Peer et al.2011).已有研究表明, PIN蛋白在质膜上的极性定位关系到生长素的运输方向, 是生长素在植物体内不对称分布的重要原因(Benkováet al.2003; Wi?niewska,2006; Vieten et al.2007).运用全基因组关联分析(genome wide association study, GWAS)方法, 目前已在30多种植物中鉴定出不同数量的PIN基因(Zhou and Luo, 2018).例如, 在咖啡(Coffea arabica)、玉米(Zea mays)以及高粱(Sorghum bicolor)中分别发现了17、15和11个PIN基因(Shen et al.2010; Yue et al.2015; Huang et al.2020).模式植物拟南芥中有8个PIN蛋白成员, 其生物学功能已有相关报道. 然而对水稻(Oryza sativa)、小麦(Triticum aestivum)和番茄(Solanum lycopersicum)等作物以及裸子植物中的PIN基因功能还知之甚少, 尚待进一步研究和探索(K?e?eket al.2009).本文主要综述了PIN蛋白的分子结构、极性定位的分子调控机理及其在拟南芥和禾谷类作物中的研究进展. ...
Jasmonic acid modulates xylem development by controlling expression of PIN-FOR- MED 7 1 2019
... PIN基因的表达还受多种植物激素的影响(包括生长素自身的诱导).除了PIN5的表达受生长素负调节外, 其余PIN的表达均由生长素通过Aux/IAA信号转导途径正反馈调控(Mravec et al.2009).乙烯合成前体ACC处理可使PIN1、PIN2和PIN4的表达上调; 细胞分裂素则通过抑制PIN1-PIN3的表达调节根分生区活性; 外施油菜素内酯(BL)可提高PIN1和PIN2的转录水平, BL合成突变体中PIN1和PIN2的转录水平下降; 茉莉酸(JA)处理可使PIN7的表达水平下调, 并由此控制生长素的极性运输, 调节根中木质部的发育(Li et al.2005; Ruzicka et al.2007, 2009; Jang et al.2019).上述结果表明, 各植物激素信号对植物生长发育的调控之间存在很重要的调控网络, 但这些网络互作的具体机理尚需进一步探索. ...
Mitogen-activated protein kinase cascade MKK7- MPK6 plays important roles in plant development and regulates shoot branching by phosphorylating PIN1 in Arabidopsis 2 2016
... 在长亲水环上还含有多个保守的磷酸化位点, 它们是调节PIN转运和亚细胞极性的某些激酶的靶点(图1).对拟南芥的研究表明, 在3个高度保守的TPRXS基序中, 丝氨酸残基S1-S3 (S231、S252和S290)是PIDs (PINOID)激酶和D6蛋白激酶(D6PKs) 的靶点, 与S1-S3相邻的3个苏氨酸残基(T227、T248和T286)是促分裂素原活化激酶(mitogen activated protein kinases, MAPK)的靶点(Huang et al.2010; Zourelidou et al.2014; Dory et al.2018).此外, 在长亲水环上不保守的丝氨酸残基S337和苏氨酸残基T340也被证明参与PIN的磷酸化调控.MAPK能通过磷酸化S337来调节拟南芥初生茎中生长素的极性运输, 但靶向T340位点的激酶目前未知(Ganguly et al.2012; Jia et al.2016). ...
... 拟南芥基因组共编码20个MAPK和10个MAPK 激酶(MKKs), MKK-MAPK信号级联参与多种植物发育进程的调控.MKK7-MAPK3/6级联通过磷酸化S337位点使PIN在下胚轴和茎中去极化, 从而影响茎中生长素的极性运输(Jia et al.2016).MAPK4和MAPK6能够磷酸化T227、T248和T286, 这3个位点与S1-S3相邻, 说明AGCVIII和MAPK蛋白激酶很可能通过磷酸化PIN蛋白上的丝氨酸和苏氨酸位点介导PIN的极性(Dory et al.2018). ...
Auxin transport through PIN-FORMED 3 (PIN3) controls shade avoidance and fitness during com- petition 1 2010
... 拟南芥PIN家族在植物整个生长发育和形态建成中发挥重要作用已有大量报道, 但对PIN1的报道最多. PIN1在整个维管组织和发育的器官中均有表达, 且是唯一能在茎尖生长点检测到的PIN家族成员.G?lweiler等(1998)和Blilou等(2005)研究发现, PIN1在根中柱表达, 将地上部合成的生长素输送至根尖, 并参与茎尖分生组织环流、器官分化、叶和花芽的形成、茎的重力响应和维管发育.PIN2主要在根尖伸长区的皮层和胚胎发生过程中表达, 参与根中生长素的向基运输和调节根的向重力性(Chen at al., 1998).PIN3主要在根冠和维管组织中表达, 调节根尖生长素的再分配, 参与侧根的早期形成、根尖钩的形成和维持, 以及调控向重力性和向光性, 参与植物的向光生长和荫蔽反应(Keuskamp et al.2010; Chen et al.2015).与PIN3类似, PIN4也在植物的向光反应和根尖钩发育中起作用, PIN4在成熟或发育中的根分生组织表达, 定位于根尖静止中心(quiescent center, QC)区域, 参与生长素的向QC运输(Friml et al.2002).PIN5定位于内质网, 主要在下胚轴伸长部位、子叶维管组织和保卫细胞中表达, 在植物发育后期表达较弱, 它通过调控细胞内生长素的动态平衡, 参与一系列发育过程(包括侧根的发生、子叶扩张、下胚轴和根的生长等)(Mravec et al.2009).PIN6定位于质膜和内质网膜, 在生长素信号介导的发育过程(如侧根和不定根的发生、根毛生长和根尖优势)中起作用.组织定位结果显示, PIN6在侧根发生原基的边缘和根茎结合处表达(Ditengou et al.2018).PIN7在根尖中的定位与PIN3有部分重叠, 且也参与根的重力响应过程(Rosquete et al.2018).PIN8定位于内质网, 在花药中特异性表达, 控制花粉、雄配子体和孢子体的发育.Ding等(2012)和Lee等(2020)研究表明, 在侧根发生后期可检测到PIN8在根尖分生组织基部和伸长/成熟区的韧皮部中呈点状表达, 表明其也参与侧根的发生, 但与质膜定位的PIN引导胞间生长素回流的机制不同, PIN8很可能通过调节细胞内生长素的转运来影响侧根的发育过程.虽然PIN家族成员均有各自功能, 但它们在植物生理过程中相互协调、分工合作来调节生长素的分布与流向(Leyser, 2006; K?e?eket al.2009). ...
Clathrin mediates endocytosis and polar distribution of PIN auxin transporters in Arabidopsis 1 2011
Brassinosteroids stimulate plant tropisms through modulation of polar auxin transport in Brassica and Arabidopsis 1 2005
... PIN基因的表达还受多种植物激素的影响(包括生长素自身的诱导).除了PIN5的表达受生长素负调节外, 其余PIN的表达均由生长素通过Aux/IAA信号转导途径正反馈调控(Mravec et al.2009).乙烯合成前体ACC处理可使PIN1、PIN2和PIN4的表达上调; 细胞分裂素则通过抑制PIN1-PIN3的表达调节根分生区活性; 外施油菜素内酯(BL)可提高PIN1和PIN2的转录水平, BL合成突变体中PIN1和PIN2的转录水平下降; 茉莉酸(JA)处理可使PIN7的表达水平下调, 并由此控制生长素的极性运输, 调节根中木质部的发育(Li et al.2005; Ruzicka et al.2007, 2009; Jang et al.2019).上述结果表明, 各植物激素信号对植物生长发育的调控之间存在很重要的调控网络, 但这些网络互作的具体机理尚需进一步探索. ...
Calcium signals are necessary to establish auxin transporter polarity in a plant stem cell niche 1 2019a
Phospholipase D-derived phosphatidic acid promotes root hair development under phosphorus deficiency by suppressing vacuolar degradation of PINFOR- MED2 1 2020
... 拟南芥PIN家族在植物整个生长发育和形态建成中发挥重要作用已有大量报道, 但对PIN1的报道最多. PIN1在整个维管组织和发育的器官中均有表达, 且是唯一能在茎尖生长点检测到的PIN家族成员.G?lweiler等(1998)和Blilou等(2005)研究发现, PIN1在根中柱表达, 将地上部合成的生长素输送至根尖, 并参与茎尖分生组织环流、器官分化、叶和花芽的形成、茎的重力响应和维管发育.PIN2主要在根尖伸长区的皮层和胚胎发生过程中表达, 参与根中生长素的向基运输和调节根的向重力性(Chen at al., 1998).PIN3主要在根冠和维管组织中表达, 调节根尖生长素的再分配, 参与侧根的早期形成、根尖钩的形成和维持, 以及调控向重力性和向光性, 参与植物的向光生长和荫蔽反应(Keuskamp et al.2010; Chen et al.2015).与PIN3类似, PIN4也在植物的向光反应和根尖钩发育中起作用, PIN4在成熟或发育中的根分生组织表达, 定位于根尖静止中心(quiescent center, QC)区域, 参与生长素的向QC运输(Friml et al.2002).PIN5定位于内质网, 主要在下胚轴伸长部位、子叶维管组织和保卫细胞中表达, 在植物发育后期表达较弱, 它通过调控细胞内生长素的动态平衡, 参与一系列发育过程(包括侧根的发生、子叶扩张、下胚轴和根的生长等)(Mravec et al.2009).PIN6定位于质膜和内质网膜, 在生长素信号介导的发育过程(如侧根和不定根的发生、根毛生长和根尖优势)中起作用.组织定位结果显示, PIN6在侧根发生原基的边缘和根茎结合处表达(Ditengou et al.2018).PIN7在根尖中的定位与PIN3有部分重叠, 且也参与根的重力响应过程(Rosquete et al.2018).PIN8定位于内质网, 在花药中特异性表达, 控制花粉、雄配子体和孢子体的发育.Ding等(2012)和Lee等(2020)研究表明, 在侧根发生后期可检测到PIN8在根尖分生组织基部和伸长/成熟区的韧皮部中呈点状表达, 表明其也参与侧根的发生, 但与质膜定位的PIN引导胞间生长素回流的机制不同, PIN8很可能通过调节细胞内生长素的转运来影响侧根的发育过程.虽然PIN家族成员均有各自功能, 但它们在植物生理过程中相互协调、分工合作来调节生长素的分布与流向(Leyser, 2006; K?e?eket al.2009). ...
... PIN基因的表达还受多种植物激素的影响(包括生长素自身的诱导).除了PIN5的表达受生长素负调节外, 其余PIN的表达均由生长素通过Aux/IAA信号转导途径正反馈调控(Mravec et al.2009).乙烯合成前体ACC处理可使PIN1、PIN2和PIN4的表达上调; 细胞分裂素则通过抑制PIN1-PIN3的表达调节根分生区活性; 外施油菜素内酯(BL)可提高PIN1和PIN2的转录水平, BL合成突变体中PIN1和PIN2的转录水平下降; 茉莉酸(JA)处理可使PIN7的表达水平下调, 并由此控制生长素的极性运输, 调节根中木质部的发育(Li et al.2005; Ruzicka et al.2007, 2009; Jang et al.2019).上述结果表明, 各植物激素信号对植物生长发育的调控之间存在很重要的调控网络, 但这些网络互作的具体机理尚需进一步探索. ...
AtPIN2 defines a locus of Arabidopsis for root gravitropism control 1 1998
... EIR1基因的克隆早于PIN1, Luschnig等(1998)研究发现, 表达EIR1蛋白的酵母(Saccharomyces cerevisiae)能将有毒的IAA类似物5-氟-吲哚运出细胞, 从而表现出对该化合物的抗性, 表明EIR1蛋白具有生长素输出功能.在此之前, 通过不同途径鉴定到的根向地性缺失突变体agr1和wavy6、根特异的乙烯不敏感突变体eir1的突变基因, 以及同年由其它3个实验室从根向地性缺失突变体中分离克隆的基因, 经序列分析表明为同一基因, 该基因与PIN1同源, 命名为PIN2(Bell and Maher, 1990; Okada and Shimura, 1990; Roman et al.1995; Chen et al.1998; Mülleret al.1998; Utsuno et al.1998).不同角度的研究结果共同证实了PIN的生长素输出功能, 最直接的证据是当在植物细胞(如烟草(Nicotiana tabacum) BY-2细胞和拟南芥悬浮细胞)和异源表达系统(如爪蟾(Xenopus laevis)卵母细胞和酵母)中表达PIN蛋白时, 其均能表现出外排生长素的能力(Petráseket al.2006; Yang and Murphy, 2009). ...
Reversible root tip rotation in Arabidopsis seedlings induced by obstacle-touching stimu- lus 1 1990
... EIR1基因的克隆早于PIN1, Luschnig等(1998)研究发现, 表达EIR1蛋白的酵母(Saccharomyces cerevisiae)能将有毒的IAA类似物5-氟-吲哚运出细胞, 从而表现出对该化合物的抗性, 表明EIR1蛋白具有生长素输出功能.在此之前, 通过不同途径鉴定到的根向地性缺失突变体agr1和wavy6、根特异的乙烯不敏感突变体eir1的突变基因, 以及同年由其它3个实验室从根向地性缺失突变体中分离克隆的基因, 经序列分析表明为同一基因, 该基因与PIN1同源, 命名为PIN2(Bell and Maher, 1990; Okada and Shimura, 1990; Roman et al.1995; Chen et al.1998; Mülleret al.1998; Utsuno et al.1998).不同角度的研究结果共同证实了PIN的生长素输出功能, 最直接的证据是当在植物细胞(如烟草(Nicotiana tabacum) BY-2细胞和拟南芥悬浮细胞)和异源表达系统(如爪蟾(Xenopus laevis)卵母细胞和酵母)中表达PIN蛋白时, 其均能表现出外排生长素的能力(Petráseket al.2006; Yang and Murphy, 2009). ...
Requirement of the auxin polar transport system in early stages of Arabidopsis floral bud formation 1 1991
... PIN家族最早被发现的基因是PIN1.该突变体的特征是花序茎上几乎无叶和花等器官, 形状类似大头针, 因此命名为pin1 (pin-formed1)突变体(Goto et al.1987).研究发现, 在添加生长素极性运输抑制剂培养基上生长的野生型拟南芥会表现出与pin1突变体相似的表型, 且pin1突变体中生长素的极性运输活性降低, 因此推测PIN1参与花序发育中生长素的极性运输(Okada et al.1991).1998年, PIN1基因被克隆出来, 进一步研究表明, PIN1的蛋白结构类似于细菌和真核生物中的载体蛋白, 且极性定位于维管组织细胞的细胞膜基部, 与化学渗透假说推测的生长素输出载体相符(G?lweileret al.1998). ...
The E3 ubiquitin ligase SCFTIR1/AFB and membrane sterols play key roles in auxin regulation of endocytosis, recycling, and plasma membrane accumulation of the auxin efflux transporter PIN2 in Arabidopsis thaliana 1 2009
... 植物固醇、生长素和其它植物激素等也参与PIN的胞内循环调控, 其中有些植物激素通过反馈调节影响PIN的循环运输.生长素处理可在短时间内维持PIN定位在膜上, 依赖TIR1介导的信号途径抑制PIN进入内吞小泡; 长时间处理则会导致PIN蛋白降解(Pan et al.2009).植物固醇合成途径的关键基因CPI1突变后, 突变体中的胞吞作用被抑制, 使PIN2的极性定位改变(Men et al.2008).低浓度茉莉酸甲酯(MeJA)处理抑制PIN2的内吞也依赖TIR1信号途径; 细胞分裂素则促进PIN1从质膜到液泡的降解; 独脚金内酯(SLs)干扰生长素对PIN极性的反馈调节过程(包括对CME途径的干扰), 从而影响维管组织的形成和再生(Sun et al.2011; Marhavyet al.2011; Zhang et al.2020). ...
Butylated Hydroxytoluene (BHT) inhibits PIN1 exocytosis from BFA compartments in Arabidopsis roots 1 2020
Genetic analysis of ethylene signal transduction in Arabidopsis thaliana: five novel mutant loci integrated into a stress response pathway 1 1995
... EIR1基因的克隆早于PIN1, Luschnig等(1998)研究发现, 表达EIR1蛋白的酵母(Saccharomyces cerevisiae)能将有毒的IAA类似物5-氟-吲哚运出细胞, 从而表现出对该化合物的抗性, 表明EIR1蛋白具有生长素输出功能.在此之前, 通过不同途径鉴定到的根向地性缺失突变体agr1和wavy6、根特异的乙烯不敏感突变体eir1的突变基因, 以及同年由其它3个实验室从根向地性缺失突变体中分离克隆的基因, 经序列分析表明为同一基因, 该基因与PIN1同源, 命名为PIN2(Bell and Maher, 1990; Okada and Shimura, 1990; Roman et al.1995; Chen et al.1998; Mülleret al.1998; Utsuno et al.1998).不同角度的研究结果共同证实了PIN的生长素输出功能, 最直接的证据是当在植物细胞(如烟草(Nicotiana tabacum) BY-2细胞和拟南芥悬浮细胞)和异源表达系统(如爪蟾(Xenopus laevis)卵母细胞和酵母)中表达PIN蛋白时, 其均能表现出外排生长素的能力(Petráseket al.2006; Yang and Murphy, 2009). ...
PIN7 auxin carrier has a preferential role in terminating radial root expansion in Arabidopsis thaliana 1 2018
... 拟南芥PIN家族在植物整个生长发育和形态建成中发挥重要作用已有大量报道, 但对PIN1的报道最多. PIN1在整个维管组织和发育的器官中均有表达, 且是唯一能在茎尖生长点检测到的PIN家族成员.G?lweiler等(1998)和Blilou等(2005)研究发现, PIN1在根中柱表达, 将地上部合成的生长素输送至根尖, 并参与茎尖分生组织环流、器官分化、叶和花芽的形成、茎的重力响应和维管发育.PIN2主要在根尖伸长区的皮层和胚胎发生过程中表达, 参与根中生长素的向基运输和调节根的向重力性(Chen at al., 1998).PIN3主要在根冠和维管组织中表达, 调节根尖生长素的再分配, 参与侧根的早期形成、根尖钩的形成和维持, 以及调控向重力性和向光性, 参与植物的向光生长和荫蔽反应(Keuskamp et al.2010; Chen et al.2015).与PIN3类似, PIN4也在植物的向光反应和根尖钩发育中起作用, PIN4在成熟或发育中的根分生组织表达, 定位于根尖静止中心(quiescent center, QC)区域, 参与生长素的向QC运输(Friml et al.2002).PIN5定位于内质网, 主要在下胚轴伸长部位、子叶维管组织和保卫细胞中表达, 在植物发育后期表达较弱, 它通过调控细胞内生长素的动态平衡, 参与一系列发育过程(包括侧根的发生、子叶扩张、下胚轴和根的生长等)(Mravec et al.2009).PIN6定位于质膜和内质网膜, 在生长素信号介导的发育过程(如侧根和不定根的发生、根毛生长和根尖优势)中起作用.组织定位结果显示, PIN6在侧根发生原基的边缘和根茎结合处表达(Ditengou et al.2018).PIN7在根尖中的定位与PIN3有部分重叠, 且也参与根的重力响应过程(Rosquete et al.2018).PIN8定位于内质网, 在花药中特异性表达, 控制花粉、雄配子体和孢子体的发育.Ding等(2012)和Lee等(2020)研究表明, 在侧根发生后期可检测到PIN8在根尖分生组织基部和伸长/成熟区的韧皮部中呈点状表达, 表明其也参与侧根的发生, 但与质膜定位的PIN引导胞间生长素回流的机制不同, PIN8很可能通过调节细胞内生长素的转运来影响侧根的发育过程.虽然PIN家族成员均有各自功能, 但它们在植物生理过程中相互协调、分工合作来调节生长素的分布与流向(Leyser, 2006; K?e?eket al.2009). ...
Carrier-mediated auxin transport 1 1974
... 生长素的极性运输需要载体, 目前已证实有家族蛋白具有生长素转运功能(Rubery and Sheldrake, 1974; Bennett et al.1996; G?lweileret al.1998; Sidler et al.1998).生长素在细胞间的运输主要依赖3种膜定位蛋白, 分别为生长素输入载体AUX/LAX (AUXIN1/LIKE-AUX1)、生长素输出载体PIN (PIN- FORMED)以及ABCB/PGP (ATP-Binding Cassette subfamily B/P-glycoprotein)转运体(Péretet al.2012; Geisler et al.2017; Zhou and Luo, 2018).此外, 在拟南芥(Arabidopsis thaliana)中, 硝酸盐转运体NRT1.1也具有转运生长素的能力, 低氮条件下其通过促进生长素的向基运输抑制侧根的生长(Krouk et al.2010).生长素在细胞内的运输则依赖定位于胞内的PILS (PIN-Like)蛋白家族、液泡生长素转运蛋白WAT1 (WALLS ARE THIN 1)以及具有短亲水环结构的部分PIN蛋白(Barbez et al.2012; Ranocha et al.2013). ...
Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution 1 2007
... PIN基因的表达还受多种植物激素的影响(包括生长素自身的诱导).除了PIN5的表达受生长素负调节外, 其余PIN的表达均由生长素通过Aux/IAA信号转导途径正反馈调控(Mravec et al.2009).乙烯合成前体ACC处理可使PIN1、PIN2和PIN4的表达上调; 细胞分裂素则通过抑制PIN1-PIN3的表达调节根分生区活性; 外施油菜素内酯(BL)可提高PIN1和PIN2的转录水平, BL合成突变体中PIN1和PIN2的转录水平下降; 茉莉酸(JA)处理可使PIN7的表达水平下调, 并由此控制生长素的极性运输, 调节根中木质部的发育(Li et al.2005; Ruzicka et al.2007, 2009; Jang et al.2019).上述结果表明, 各植物激素信号对植物生长发育的调控之间存在很重要的调控网络, 但这些网络互作的具体机理尚需进一步探索. ...
Cytokinin regulates root meristem activity via modulation of the polar auxin transport 1 2009
... PIN基因的表达还受多种植物激素的影响(包括生长素自身的诱导).除了PIN5的表达受生长素负调节外, 其余PIN的表达均由生长素通过Aux/IAA信号转导途径正反馈调控(Mravec et al.2009).乙烯合成前体ACC处理可使PIN1、PIN2和PIN4的表达上调; 细胞分裂素则通过抑制PIN1-PIN3的表达调节根分生区活性; 外施油菜素内酯(BL)可提高PIN1和PIN2的转录水平, BL合成突变体中PIN1和PIN2的转录水平下降; 茉莉酸(JA)处理可使PIN7的表达水平下调, 并由此控制生长素的极性运输, 调节根中木质部的发育(Li et al.2005; Ruzicka et al.2007, 2009; Jang et al.2019).上述结果表明, 各植物激素信号对植物生长发育的调控之间存在很重要的调控网络, 但这些网络互作的具体机理尚需进一步探索. ...
An auxin-dependent distal organi- zer of pattern and polarity in the Arabidopsis root 1 1999
... 生长素是最早被发现的植物激素, 参与植物生长发育的各个阶段(Enders and Strader, 2015).生长素在植物组织和器官中建立的不对称分布是其行使功能的重要基础.例如, 调控根分生组织的活性、器官的形成和维管组织分化等, 是长期适应的结果(Sabatini et al.1999; Benkováet al.2003; Mattsson et al.2003).生长素从合成部位(如幼叶和顶芽)运输到作用部位主要有2条途径: 一是在整个植株范围内, 以不固定方向被动运输的方式, 通过韧皮部完成长距离运输; 二是需要载体参与的细胞与细胞之间短距离极性运输, 包括从茎尖向根的单向运输和组织间的局部单向运输( Petrá?ek and Friml,2009).极性运输对建立和维持生长素的浓度梯度至关重要, 进而特异性调节植物的生长发育及其对外界环境变化的响应. ...
Sorting motifs involved in the trafficking and localization of the PIN1 auxin efflux carrier 1 2016
Solanum tuberosum StCDPK1 is regulated by miR390 at the posttranscriptional level and phosphorylates the auxin efflux carrier StPIN4 in vitro, a potential downstream target in potato development 1 2017
Phosphatidic acid directly regulates PINOID-dependent phosphorylation and activation of the PIN-FORMED2 auxin efflux transporter in response to salt stress 1 2019
... 参与PIN磷酸化过程的激酶家族主要有AGC、MAPK蛋白和钙调蛋白激酶(Ca2+/calmodulin-dependent protein kinases, CDPK) 3类.拟南芥基因组编码的39个AGC激酶隶属AGCVI、AGCVII、AGCVIII、PDK1及其它AGC激酶5个亚族, AGCVIII亚族的23个激酶进一步细分为AGC1、AGC2、AGC3和AGC4四类, PIDs属于AGCVIII激酶的AGC3类, D6PKs属于AGC1类(B?greet al.2003; Galván-Ampudia and Offringa,2007).PIDs通过磷酸化S1-S3位点调节PIN的极性定位, 过表达PID促进PIN蛋白从细胞基部定位到顶部, 导致一定范围内的生长素不具有浓度梯度; pid突变体茎分生组织分化过程必需的PIN基部到顶部的极性转换受阻, 导致器官发育缺陷(Friml et al.2004).有研究表明, 磷脂酸(phosphatidic acid, PA)能增强PID介导的PIN2磷酸化, 从而激活生长素外排, 促进盐胁迫下拟南芥的根系生长, 说明AGCVIII激酶可从多方面调控PIN的功能(Wang et al.2019).D6PKs在大部分细胞中与PIN蛋白共定位于细胞基部, 其磷酸化位点除了S1-S3外, 还包括丝氨酸残基S4和S5 (S271和S215), 这2个位点在PIN3、PIN4和PIN7中保守, 但在PIN1和PIN2中不完全保守.d6pk功能缺失突变体中PIN蛋白的磷酸化和生长素运输速率均降低, 然而与PIDs不同的是, 过表达D6PK并不改变细胞中PIN的极性.PAX是一种D6PKL (D6PK-LIKE)蛋白激酶, 通过磷酸化S1位点激活PIN蛋白活性, 促进生长素流出.在初生韧皮部中, PAX与BRX因子互作调节生长素的含量, 从而在时间上精确控制原生韧皮分子(protophloem sieve elements, PPSE)的分化(Marhava et al.2018).D6PKs与PIDs共享PIN上的磷酸化靶点, 但它们对这些靶点有不同的选择偏好, 因此其在调控PIN极性上的作用不同(Zourelidou et al.2014; Barbosa et al.2018).其它AGCVIII激酶也被证明能够在体内或体外通过磷酸化PIN蛋白参与植物生长的调控, 但具体机理尚不清楚(Galván-Ampudia and Offringa,2007; Haga et al.2018). ...
Analysis of subcellular localization of auxin carriers PIN, AUX/LAX and PGP in Sorghum bicolor 1 2011
Functional expression and characterization of Arabidopsis ABCB, AUX1 and PIN auxin transporters in Schizosaccharomyces pombe 1 2009
... EIR1基因的克隆早于PIN1, Luschnig等(1998)研究发现, 表达EIR1蛋白的酵母(Saccharomyces cerevisiae)能将有毒的IAA类似物5-氟-吲哚运出细胞, 从而表现出对该化合物的抗性, 表明EIR1蛋白具有生长素输出功能.在此之前, 通过不同途径鉴定到的根向地性缺失突变体agr1和wavy6、根特异的乙烯不敏感突变体eir1的突变基因, 以及同年由其它3个实验室从根向地性缺失突变体中分离克隆的基因, 经序列分析表明为同一基因, 该基因与PIN1同源, 命名为PIN2(Bell and Maher, 1990; Okada and Shimura, 1990; Roman et al.1995; Chen et al.1998; Mülleret al.1998; Utsuno et al.1998).不同角度的研究结果共同证实了PIN的生长素输出功能, 最直接的证据是当在植物细胞(如烟草(Nicotiana tabacum) BY-2细胞和拟南芥悬浮细胞)和异源表达系统(如爪蟾(Xenopus laevis)卵母细胞和酵母)中表达PIN蛋白时, 其均能表现出外排生长素的能力(Petráseket al.2006; Yang and Murphy, 2009). ...
The Arabidopsis SWI2/SNF2 chromatin remodeling ATPase BRAHMA targets directly to PINs and is required for root stem cell niche maintenance 1 2015
... 在转录水平上, 有些上游转录因子作用于PIN基因的启动子区, 调节其表达以响应生长素内源以及外源信号变化.在生长素响应因子ARF (AUXIN RE-SPONSE FACTOR)家族中, ARF7与受其调控的FLP转录因子共同形成前馈基序以调控PIN3的转录, 介导侧根形成的早期阶段(Chen et al.2015); 细胞分裂素响应因子CRF2和CRF6作用于PIN1和PIN7启动子上的PIN特异性细胞分裂素响应元件(PCRE), 激活PIN1和PIN7的表达, 并且CRF2受ARF5的调控, 该研究表明生长素的极性运输受生长素和细胞分裂素的交叉信号调控(?imá?kováet al.2015).参与脂肪酸生物合成的AP2转录因子WRI1 (WRINK-LED1)直接靶向PIN4和PIN5的启动子, 并导致wri1-1突变体中某些PIN基因(如PIN1、PIN3和PIN5-6)的表达水平降低, 为阐明脂肪酸和PIN蛋白的关系提供了思路(Kong et al.2017).此外, 近期研究发现, NAC转录因子AtNAC2 (ANAC092)与ARF8和PIN4的启动子结合, 通过生长素途径负调控拟南芥根的生长(Xi et al.2019).此外, IBR5也影响PIN的转录活性, 表现为ibr5-3突变体中, PIN1在表皮细胞质膜和叶脉周边细胞中的分布受到影响, 叶片锯齿形成受阻(Kong et al.2019).MAKR2 (MEMBRANE ASSOCIATED KINASE REGULATOR2)作为跨膜激酶TMK1 (TRAN SMEMBRANE KINASE1)的负调控因子, 影响PIN2在根两侧的动态分布, 进而参与根的向重力性调控(Marquès-Buenoet al.2021).表观遗传方面, SWI/SNF的染色质重塑ATP酶BRM (BRAHMA)直接靶向根部“长”PIN蛋白(PIN1-PIN4和PIN7)的染色质, 功能丧失突变体brm表现为根干细胞生态位维持缺陷, 根生长发育不良, 干细胞中一些PIN基因的局部表达量降低, 从而影响生长素的分布(Yang et al.2015).生长素转运蛋白ABCB家族与PIN蛋白的互作也会调控其表达活性.有研究表明, ABCB1、ABCB19可与PIN1、PIN2互作, 共表达PIN1-ABCB1和PIN1- ABCB19的植株生长素输出速率升高(Blakeslee et al.2007; 贺祯媚等, 2019). ...
Genome-wide identification and expression profiling analysis of ZmPIN, ZmPILS, ZmLAX and ZmABCB auxin transporter gene families in maize (Zea mays L.) under various abiotic stresses 3 2015
... 根据化学渗透偶联学说, 生长素输出载体在细胞膜上的不对称分布是生长素极性运输的关键(Friml, 2010; Peer et al.2011).已有研究表明, PIN蛋白在质膜上的极性定位关系到生长素的运输方向, 是生长素在植物体内不对称分布的重要原因(Benkováet al.2003; Wi?niewska,2006; Vieten et al.2007).运用全基因组关联分析(genome wide association study, GWAS)方法, 目前已在30多种植物中鉴定出不同数量的PIN基因(Zhou and Luo, 2018).例如, 在咖啡(Coffea arabica)、玉米(Zea mays)以及高粱(Sorghum bicolor)中分别发现了17、15和11个PIN基因(Shen et al.2010; Yue et al.2015; Huang et al.2020).模式植物拟南芥中有8个PIN蛋白成员, 其生物学功能已有相关报道. 然而对水稻(Oryza sativa)、小麦(Triticum aestivum)和番茄(Solanum lycopersicum)等作物以及裸子植物中的PIN基因功能还知之甚少, 尚待进一步研究和探索(K?e?eket al.2009).本文主要综述了PIN蛋白的分子结构、极性定位的分子调控机理及其在拟南芥和禾谷类作物中的研究进展. ...
... 目前, 玉米中发现有15个PIN家族成员, 其可分为3个亚族.第I亚族为: ZmPin1 (ZmPIN1a-d)、ZmPIN8和ZmPIN10 (ZmPIN10a-b); 第II亚族为ZmPIN5(ZmPIN 5a-d); 第III亚族为: ZmPIN9、ZmPIN13、ZmPIN14和ZmPIN15.大部分ZmPIN基因与水稻PIN家族基因同源, 而ZmPIN13和ZmPIN15的分支为玉米所特有(Yue et al.2015).ZmPIN优先在地上部表达, ZmPIN1d在雄蕊、穗和成株的第5节中特异表达, ZmPIN9在根内皮层和中柱鞘特异表达, 系统发育分析和ZmPIN基因家族表达模式表明, 玉米某些PIN蛋白的亚功能化可能与单子叶植物的特定器官、组织分化和发育有关(Forestan et al.2012).尽管全基因组测序使我们对玉米PIN家族有了宏观的了解, 但还缺乏对单个基因的生物学功能以及表达调控等的认识.已有研究表明, ZmPIN1a和ZmPIN1b可能在玉米分生组织中起基础性作用, ZmPIN1以极性和非极性两种方式定位于某些组织的细胞质膜, 或聚集在胚乳的一种特殊组织细胞的内膜上, 是玉米胚胎发生、胚乳发育以及生殖发育过程中细胞和组织分化所必需的蛋白(Carraro et al.2006; Forestan and Varotto, 2012). ...
OsPIN1与AtPIN1类似, 主要在维管组织和不定根原基中表达, 对水稻不定根发育、分蘖、地下部和地上部的比例具有重要调控作用, 下调OsPIN1的表达明显抑制水稻的分蘖数和不定根发生(Xu et al.2005).对OsPIN1的4个基因研究发现, OsPIN1a-d在根中的表达量相对高于其它组织, 它们的单突变体较野生型不具有明显表型, 而pin1a/pin1b双突变体表现出地上部和初生根变短、根毛变长, 以及重力性缺失表型, pin1c/pin1d双突变植株则表现出类似拟南芥pin1突变体的针状花序表型, 说明OsPIN1a和OsPIN1b主要参与水稻根和茎的发育, 而OsPIN1c和OsPIN1d参与穗的形成, 并且它们之间存在功能冗余(Li et al.2019b).也有研究表明, OsPIN1a在水稻根的负趋光性调控中起作用(Xu et al.2014).OsPIN2主要在根中分布, 极性分布于根尖的表皮和皮层区, 过表达OsPIN2可增强生长素从地上部向根茎结合部的转运, 从而促进水稻分蘖, 并使分蘖角度增大、株高降低(Chen et al.2012).OsPIN5b在多种组织中均有表达, 亚细胞定位于内质网, 参与生长素的胞内转运和平衡, 其过表达株系的分蘖数、株高、穗长和产量均降低, 而ospin5b突变体的穗长变长, 说明OsPIN5b通过生长素信号调控水稻的株型和产量(Lu et al.2015; Zeng et al.2020).OsPIN9主要在分蘖芽及其维管组织中表达,其亲水环长度介于“长”PIN蛋白与“短”PIN蛋白之间.ospin9突变体的分蘖数减少, 过表达株系的表型则相反.大田试验结果显示, 低氮条件下OsPIN9过表达株系的分蘖数和产量均高于高氮条件(Hou et al.2021).OsPIN10a (OsPIN3t/OsPIN3a)主要在维管组织中表达, 亚细胞定位于质膜, 其过表达株系的抗旱能力增强, 说明OsPIN10a所介导的生长素极性运输与植株抗旱响应有关(Miyashita et al.2010; Zhang et al.2012).OsPIN8基因的功能尚未见报道, 系统发育分析表明其与拟南芥的AtPIN8相近, 蛋白结构上缺少中心亲水环, 推测与拟南芥的PIN8一样定位于内质网, 介导生长素的胞内运输, 但尚需实验验证.对比水稻和拟南芥PIN家族的生物学功能, 发现PIN在水稻地上部株型的相关性状(如分蘖数、分蘖角和株高)调控中发挥重要作用, 并且影响作物的产量性状, 继续推进水稻等禾谷类作物中PIN蛋白的研究具有重要意义. ...
... OsPIN1与AtPIN1类似, 主要在维管组织和不定根原基中表达, 对水稻不定根发育、分蘖、地下部和地上部的比例具有重要调控作用, 下调OsPIN1的表达明显抑制水稻的分蘖数和不定根发生(Xu et al.2005).对OsPIN1的4个基因研究发现, OsPIN1a-d在根中的表达量相对高于其它组织, 它们的单突变体较野生型不具有明显表型, 而pin1a/pin1b双突变体表现出地上部和初生根变短、根毛变长, 以及重力性缺失表型, pin1c/pin1d双突变植株则表现出类似拟南芥pin1突变体的针状花序表型, 说明OsPIN1a和OsPIN1b主要参与水稻根和茎的发育, 而OsPIN1c和OsPIN1d参与穗的形成, 并且它们之间存在功能冗余(Li et al.2019b).也有研究表明, OsPIN1a在水稻根的负趋光性调控中起作用(Xu et al.2014).OsPIN2主要在根中分布, 极性分布于根尖的表皮和皮层区, 过表达OsPIN2可增强生长素从地上部向根茎结合部的转运, 从而促进水稻分蘖, 并使分蘖角度增大、株高降低(Chen et al.2012).OsPIN5b在多种组织中均有表达, 亚细胞定位于内质网, 参与生长素的胞内转运和平衡, 其过表达株系的分蘖数、株高、穗长和产量均降低, 而ospin5b突变体的穗长变长, 说明OsPIN5b通过生长素信号调控水稻的株型和产量(Lu et al.2015; Zeng et al.2020).OsPIN9主要在分蘖芽及其维管组织中表达,其亲水环长度介于“长”PIN蛋白与“短”PIN蛋白之间.ospin9突变体的分蘖数减少, 过表达株系的表型则相反.大田试验结果显示, 低氮条件下OsPIN9过表达株系的分蘖数和产量均高于高氮条件(Hou et al.2021).OsPIN10a (OsPIN3t/OsPIN3a)主要在维管组织中表达, 亚细胞定位于质膜, 其过表达株系的抗旱能力增强, 说明OsPIN10a所介导的生长素极性运输与植株抗旱响应有关(Miyashita et al.2010; Zhang et al.2012).OsPIN8基因的功能尚未见报道, 系统发育分析表明其与拟南芥的AtPIN8相近, 蛋白结构上缺少中心亲水环, 推测与拟南芥的PIN8一样定位于内质网, 介导生长素的胞内运输, 但尚需实验验证.对比水稻和拟南芥PIN家族的生物学功能, 发现PIN在水稻地上部株型的相关性状(如分蘖数、分蘖角和株高)调控中发挥重要作用, 并且影响作物的产量性状, 继续推进水稻等禾谷类作物中PIN蛋白的研究具有重要意义. ...
The PIN-FORMED auxin efflux carriers in plants 2 2018
... 生长素的极性运输需要载体, 目前已证实有家族蛋白具有生长素转运功能(Rubery and Sheldrake, 1974; Bennett et al.1996; G?lweileret al.1998; Sidler et al.1998).生长素在细胞间的运输主要依赖3种膜定位蛋白, 分别为生长素输入载体AUX/LAX (AUXIN1/LIKE-AUX1)、生长素输出载体PIN (PIN- FORMED)以及ABCB/PGP (ATP-Binding Cassette subfamily B/P-glycoprotein)转运体(Péretet al.2012; Geisler et al.2017; Zhou and Luo, 2018).此外, 在拟南芥(Arabidopsis thaliana)中, 硝酸盐转运体NRT1.1也具有转运生长素的能力, 低氮条件下其通过促进生长素的向基运输抑制侧根的生长(Krouk et al.2010).生长素在细胞内的运输则依赖定位于胞内的PILS (PIN-Like)蛋白家族、液泡生长素转运蛋白WAT1 (WALLS ARE THIN 1)以及具有短亲水环结构的部分PIN蛋白(Barbez et al.2012; Ranocha et al.2013). ...
... 根据化学渗透偶联学说, 生长素输出载体在细胞膜上的不对称分布是生长素极性运输的关键(Friml, 2010; Peer et al.2011).已有研究表明, PIN蛋白在质膜上的极性定位关系到生长素的运输方向, 是生长素在植物体内不对称分布的重要原因(Benkováet al.2003; Wi?niewska,2006; Vieten et al.2007).运用全基因组关联分析(genome wide association study, GWAS)方法, 目前已在30多种植物中鉴定出不同数量的PIN基因(Zhou and Luo, 2018).例如, 在咖啡(Coffea arabica)、玉米(Zea mays)以及高粱(Sorghum bicolor)中分别发现了17、15和11个PIN基因(Shen et al.2010; Yue et al.2015; Huang et al.2020).模式植物拟南芥中有8个PIN蛋白成员, 其生物学功能已有相关报道. 然而对水稻(Oryza sativa)、小麦(Triticum aestivum)和番茄(Solanum lycopersicum)等作物以及裸子植物中的PIN基因功能还知之甚少, 尚待进一步研究和探索(K?e?eket al.2009).本文主要综述了PIN蛋白的分子结构、极性定位的分子调控机理及其在拟南芥和禾谷类作物中的研究进展. ...
A gain-of-function mutation in OsIAA11 affects lateral root development in rice 1 2012
... 水稻PIN基因的表达受多种途径调控, 尽管目前已有一些研究报道, 但大部分并未阐明具体调控机制.Sun等(2018)研究发现, 低磷、低氮和GR24 (类似于SLs)等处理下, OsPIN1b的表达明显下调.进一步研究发现, OsPIN1b作为NO和SLs的下游介质, 在低磷和低氮条件下通过调节生长素的运输来激活水稻根尖分生区的活性.此外, 铵态氮(NH4+)在RNA和蛋白水平上诱导OsPIN9的表达来影响分蘖数量, 说明水稻分蘖以及根尖生长受生长素运输和氮磷等营养元素供应的协同调控(Hou et al.2021).osiaa11突变体的侧根原基发育受阻, 且OsPIN1b和OsPIN- 10a的表达下调, 暗示OsIAA11可能通过调节生长素运输载体的表达来影响植株的发育(Zhu et al.2012). ...
Auxin efflux by PIN-FORMED proteins is activated by two different protein kinases, D6 PROTEIN KINASE and PINOID 2 2014
... 在长亲水环上还含有多个保守的磷酸化位点, 它们是调节PIN转运和亚细胞极性的某些激酶的靶点(图1).对拟南芥的研究表明, 在3个高度保守的TPRXS基序中, 丝氨酸残基S1-S3 (S231、S252和S290)是PIDs (PINOID)激酶和D6蛋白激酶(D6PKs) 的靶点, 与S1-S3相邻的3个苏氨酸残基(T227、T248和T286)是促分裂素原活化激酶(mitogen activated protein kinases, MAPK)的靶点(Huang et al.2010; Zourelidou et al.2014; Dory et al.2018).此外, 在长亲水环上不保守的丝氨酸残基S337和苏氨酸残基T340也被证明参与PIN的磷酸化调控.MAPK能通过磷酸化S337来调节拟南芥初生茎中生长素的极性运输, 但靶向T340位点的激酶目前未知(Ganguly et al.2012; Jia et al.2016). ...
... 参与PIN磷酸化过程的激酶家族主要有AGC、MAPK蛋白和钙调蛋白激酶(Ca2+/calmodulin-dependent protein kinases, CDPK) 3类.拟南芥基因组编码的39个AGC激酶隶属AGCVI、AGCVII、AGCVIII、PDK1及其它AGC激酶5个亚族, AGCVIII亚族的23个激酶进一步细分为AGC1、AGC2、AGC3和AGC4四类, PIDs属于AGCVIII激酶的AGC3类, D6PKs属于AGC1类(B?greet al.2003; Galván-Ampudia and Offringa,2007).PIDs通过磷酸化S1-S3位点调节PIN的极性定位, 过表达PID促进PIN蛋白从细胞基部定位到顶部, 导致一定范围内的生长素不具有浓度梯度; pid突变体茎分生组织分化过程必需的PIN基部到顶部的极性转换受阻, 导致器官发育缺陷(Friml et al.2004).有研究表明, 磷脂酸(phosphatidic acid, PA)能增强PID介导的PIN2磷酸化, 从而激活生长素外排, 促进盐胁迫下拟南芥的根系生长, 说明AGCVIII激酶可从多方面调控PIN的功能(Wang et al.2019).D6PKs在大部分细胞中与PIN蛋白共定位于细胞基部, 其磷酸化位点除了S1-S3外, 还包括丝氨酸残基S4和S5 (S271和S215), 这2个位点在PIN3、PIN4和PIN7中保守, 但在PIN1和PIN2中不完全保守.d6pk功能缺失突变体中PIN蛋白的磷酸化和生长素运输速率均降低, 然而与PIDs不同的是, 过表达D6PK并不改变细胞中PIN的极性.PAX是一种D6PKL (D6PK-LIKE)蛋白激酶, 通过磷酸化S1位点激活PIN蛋白活性, 促进生长素流出.在初生韧皮部中, PAX与BRX因子互作调节生长素的含量, 从而在时间上精确控制原生韧皮分子(protophloem sieve elements, PPSE)的分化(Marhava et al.2018).D6PKs与PIDs共享PIN上的磷酸化靶点, 但它们对这些靶点有不同的选择偏好, 因此其在调控PIN极性上的作用不同(Zourelidou et al.2014; Barbosa et al.2018).其它AGCVIII激酶也被证明能够在体内或体外通过磷酸化PIN蛋白参与植物生长的调控, 但具体机理尚不清楚(Galván-Ampudia and Offringa,2007; Haga et al.2018). ...
 ,1,2,*
,1,2,*
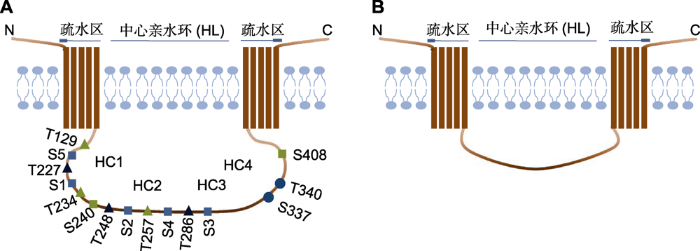 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT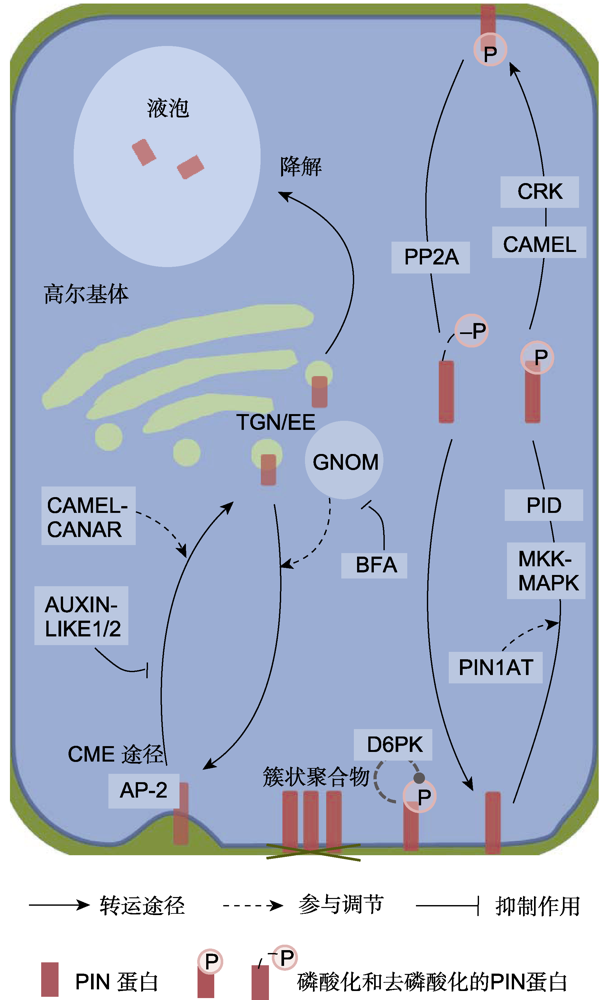 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}