Abstract Whole mount clearing is a routine method in morphological study, which allows observation of plant internal structure without section. Using high refractive index materials as medium, clearing techniques reduce light scattering, acquire enhancive light quantity and increase depth of field and vertical planes in a particular focal plane, to facilitate the samples transparency for observation. Nevertheless, clearing materials may disturb the osmosis and pH of sample medium, which is adverse to cells morphology. So far, the effective clearing techniques have been widely used in several studies with ovule and leaf. However, the current protocol is not reliable enough for root tip clearing, because the thin cell wall is vulnerable under the treatment of clearing solutions, resulting in abnormal root tips and cells plasmolysis. To achieve a stable and optimized clearing method for root tip, we established a standard protocol via evaluation of root tip morphology, plasmolysis and cells clarity in Arabidopsis thaliana. With these improved clearing methods, we developed an optimized clearing observation system (including clearing time, pH and composition) for root tip, which could provide a reliable technique for vulnerable tissues clearing. Keywords:morphogenesis;whole mount clearing;root tip;Arabidopsis thaliana
PDF (9273KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 马龙, 李桂林, 李师鹏, 蒋苏. 根尖整体透明技术改良. 植物学报, 2020, 55(5): 596-604 doi:10.11983/CBB20016 Ma Long, Li Guilin, Li Shipeng, Jiang Su. An Improved Protocol for Whole Mount Clearing of Plant Root Tip. Chinese Bulletin of Botany, 2020, 55(5): 596-604 doi:10.11983/CBB20016
Table 1 表1 表1原始pH、拟南芥培养基常用pH、中性pH透明液HCG-1和HCG-2 Table 1Solution HCG-1 and HCG-2 in original pH, the pH used for Arabidopsis thaliana culture and neutral pH
No.
HCG-1
Composition
pH
1
H2O
9 mL
1.2
Chloral hydrate
24 g
Glycerol
3 mL
2
H2O
9 mL
5.8
Chloral hydrate
24 g
Glycerol
3 mL
3
H2O
9 mL
7.2
Chloral hydrate
24 g
Glycerol
3 mL
HCG-2
Composition
pH
4
H2O
9 mL
1.8
Chloral hydrate
12 g
Glycerol
3 mL
5
H2O
9 mL
5.8
Chloral hydrate
12 g
Glycerol
3 mL
6
H2O
9 mL
7.2
Chloral hydrate
12 g
Glycerol
3 mL
HCG: Clearing solution that consists of H2O, chloral hydrate and glycerol. HCG: 由水、水合三氯乙醛和甘油组成的透明液。
分别采用透明液HCG-1 (A)和HCG-2 (B)对拟南芥幼苗透明5、15和25分钟后QC细胞清晰的根尖所占比例。 Figure 3The effects of clearing times on clarity of quiescent center (QC) cells of Arabidopsis thaliana root tips
Proportion of root tips with clear QC cells of Arabidopsis thaliana seedlings cleared by HCG-1 (A) and HCG-2 (B) for 5, 15, 25 min, respectively.
分别采用透明液HCG-1 (A)和HCG-2 (B)对拟南芥幼苗透明5、15和25分钟后根尖伸长区表皮细胞质壁分离比例(* P<0.05, ** P<0.01, *** P<0.001)。 Figure 4The effects of clearing times on plasmolysis of Arabidopsis thaliana root tip cells
Proportion of plasmolytic cells of Arabidopsis thaliana seedlings cleared by HCG-1 (A) and HCG-2 (B) for 5, 15, 25 min, respectively (* P<0.05, ** P<0.01, *** P<0.001).
分别采用透明液HCG-1 (A)和HCG-2 (B)在pH1.2/1.8、pH5.8和pH7.2条件下, 对拟南芥幼苗透明5分钟后, 根尖伸长区表皮细胞质壁分离比例和静止中心(QC)细胞清晰的根尖所占比例。不同大写字母表示差异极显著(P<0.01)。Bars=50 μm Figure 5The effects of pH values on Arabidopsis thaliana root tip clearing
Proportion of plasmolytic cells and proportion of root tips with clear quiescent center (QC) cells of Arabidopsis thaliana seedlings cleared by HCG-1 (A) and HCG-2 (B) for 5 min under pH1.2/1.8, pH5.8, pH7.2, respectively. Different uppercase letters indicate extremely significant differences (P<0.01). Bars=50 μm
分别采用透明液HCG-1和HCG-2在pH7.2条件下, 对拟南芥幼苗透明5分钟后, 根尖伸长区表皮细胞质壁分离比例和QC细胞清晰的根尖所占比例(** P<0.01, *** P<0.001)。Bars=50 μm Figure 6The comparison of solution HCG-1 with HCG-2 on Arabidopsis thaliana root tip clearing
Proportion of plasmolytic cells and proportion of root tips with clear QC cells of Arabidopsis thaliana seedlings cleared by HCG-1 and HCG-2 for 5 min under pH7.2 (** P<0.01, *** P< 0.001). Bars=50 μm
分别采用饱和三氯乙醛溶液、透明液HCG-1、透明液HCG-2和25%甘油溶液对拟南芥幼苗透明5分钟后的根尖形态。Bars= 50 μm Figure 7The comparison of root tips cleared by gradient concentrations of chloral solutions
Morphology of root tips of Arabidopsis thaliana seedlings cleared by saturated chloral, HCG-1, HCG-2 and 25% glycerol solutions for 5 min, respectively. Bars=50 μm
采用透明液HCG-2 (pH7.2)对拟南芥3、9和15日龄幼苗根尖透明5分钟。Bars=50 μm Figure 8Arabidopsis thaliana seedlings in different growth stages were used to identify the optimized protocol
3-day-old, 9-day-old and 15-day-old seedlings of Arabidopsis thaliana were cleared by HCG-2 (pH7.2) for 5 min. Bars=50 μm
BeemsterGTS, BaskinTI ( 1998). Analysis of cell division and elongation underlying the developmental acceleration of root growth in Arabidopsis thaliana Plant Physiol 116, 1515-1526. DOI:10.1104/pp.116.4.1515URLPMID:9536070 [本文引用: 1] To investigate the relation between cell division and expansion in the regulation of organ growth rate, we used Arabidopsis thaliana primary roots grown vertically at 20 degreesC with an elongation rate that increased steadily during the first 14 d after germination. We measured spatial profiles of longitudinal velocity and cell length and calculated parameters of cell expansion and division, including rates of local cell production (cells mm-1 h-1) and cell division (cells cell-1 h-1). Data were obtained for the root cortex and also for the two types of epidermal cell, trichoblasts and atrichoblasts. Accelerating root elongation was caused by an increasingly longer growth zone, while maximal strain rates remained unchanged. The enlargement of the growth zone and, hence, the accelerating root elongation rate, were accompanied by a nearly proportionally increased cell production. This increased production was caused by increasingly numerous dividing cells, whereas their rates of division remained approximately constant. Additionally, the spatial profile of cell division rate was essentially constant. The meristem was longer than generally assumed, extending well into the region where cells elongated rapidly. In the two epidermal cell types, meristem length and cell division rate were both very similar to that of cortical cells, and differences in cell length between the two epidermal cell types originated at the apex of the meristem. These results highlight the importance of controlling the number of dividing cells, both to generate tissues with different cell lengths and to regulate the rate of organ enlargement.
BougourdS, MarrisonJ, HaseloffJ ( 2000). An aniline blue staining procedure for confocal microscopy and 3D imaging of normal and perturbed cellular phenotypes in mature Arabidopsis embryos Plant J 24, 543-550. URLPMID:11115135 [本文引用: 2]
BruzzeseE, HasanS ( 1983). A whole leaf clearing and staining technique for host specificity studies of rust fungi Plant Pathol 32, 335-338. DOI:10.1111/ppa.1983.32.issue-3URL [本文引用: 1]
CraneCF ( 1978). Apomixis and Crossing Incompatibilities in Some Zephyrantheae Ph.D. thesis. Austin: University of Texas. [本文引用: 1]
DerbyshireP, FindlayK, McCannMC, RobertsK ( 2007). Cell elongation in Arabidopsis hypocotyls involves dynamic changes in cell wall thickness J Exp Bot 58, 2079-2089. DOI:10.1093/jxb/erm074URLPMID:17470442 [本文引用: 1] Field-emission scanning electron microscopy was used to measure wall thicknesses of different cell types in freeze-fractured hypocotyls of Arabidopsis thaliana. Measurements of uronic acid content, wall mass, and wall volume suggest that cell wall biosynthesis in this organ does not always keep pace with, and is not always tightly coupled to, elongation. In light-grown hypocotyls, walls thicken, maintain a constant thickness, or become thinner during elongation, depending upon the cell type and the stage of growth. In light-grown hypocotyls, exogenous gibberellic acid represses the extent of thickening and promotes cell elongation by both wall thinning and increased anisotropy during the early stages of hypocotyl elongation, and by increased wall deposition in the latter stages. Dark-grown hypocotyls, in the 48 h period between cold imbibition and seedling emergence, deposit very thick walls that subsequently thin in a narrow developmental window as the hypocotyl rapidly elongates. The rate of wall deposition is then maintained and keeps pace with cell elongation. The outer epidermal wall is always the thickest ( approximately 1 mum) whereas the thinnest walls, about 50 nm, are found in inner cell layers. It is concluded that control of wall thickness in different cell types is tightly regulated during hypocotyl development, and that wall deposition and cell elongation are not invariably coupled.
HagerA ( 2003). Role of the plasma membrane H+-ATPase in auxin-induced elongation growth: historical and new aspects J Plant Res 116, 483-505. DOI:10.1007/s10265-003-0110-xURLPMID:12937999 [本文引用: 1] This article will cover historical and recent aspects of reactions and mechanisms involved in the auxin-induced signalling cascade that terminates in the dramatic elongation growth of cells and plant organs. Massive evidence has accumulated that the final target of auxin action is the plasma membrane H(+)-ATPase, which excretes H(+) ions into the cell wall compartment and, in an antiport, takes up K(+) ions through an inwardly rectifying K(+) channel. The auxin-enhanced H(+) pumping lowers the cell wall pH, activates pH-sensitive enzymes and proteins within the wall, and initiates cell-wall loosening and extension growth. These processes, induced by auxin or by the
HerrJMJ ( 1971). A new clearing-squash technique for the study of ovule development in angiosperms Am J Bot 58, 785-790. DOI:10.1002/ajb2.1971.58.issue-8URL [本文引用: 1]
HerrJMJ (1993). Clearing techniques for the study of vascular plant tissues in whole structures and thick sections In: Tested Studies for Laboratory Teaching. Proceedings of the Fifth Workshop/Conference of the Association for Biology Laboratory Education (ABLE). Toronto: ABLE. pp. 63-84. [本文引用: 2]
ItohK, NakamuraY, KawataH, YamadaT, OhtaE, SakataM ( 1987). Effect of osmotic stress on turgor pressure in mung bean root cells Plant Cell Physiol 28, 987-994. [本文引用: 1]
IvanovVB, DubrovskyJG ( 2013). Longitudinal zonation pattern in plant roots: conflicts and solutions Trends Plant Sci 18, 237-243. DOI:10.1016/j.tplants.2012.10.002URLPMID:23123304 [本文引用: 1] Despite the relative simplicity of Arabidopsis root organization, there is no general agreement regarding the terminology used to describe the longitudinal zonation pattern (LZP) of this model system. In this opinion article, we examine inconsistencies in the terminology and provide a conceptual framework for the LZP that may be applied to all angiosperms. We propose that the root apical meristem (RAM) consists of the cell-proliferation domain where cells maintain a high probability to divide and the transition domain with a low probability of cell division; in both domains cells grow at the same, relatively low, rate. Owing to stochastic termination of cell proliferation in the RAM, the border between the domains is 'fuzzy'. Molecular markers analyzed together with quantitative growth and cell analyses could help to identify developmental zones along the root and lead to a better understanding of the LZP in angiosperms.
Janicka-RussakM (2011). Plant plasma membrane H+- ATPase in adaptation of plants to abiotic stresses. In: Shanker A, ed. Abiotic Stress Response in Plants—Physiological, Biochemical and Genetic Perspectives. Rijeka: Intech Open Press. pp. 197-218. [本文引用: 1]
KimYX, StumpfB, SungJ, LeeSJ ( 2018). The relationship between turgor pressure change and cell hydraulics of midrib parenchyma cells in the leaves of Zea mays Cells 7, 180. DOI:10.3390/cells7100180URL [本文引用: 1]
KuriharaD, MizutaY, SatoY, HigashiyamaT ( 2015). ClearSee: a rapid optical clearing reagent for whole-plant fluorescence imaging Development 142, 4168-4179. DOI:10.1242/dev.127613URLPMID:26493404 [本文引用: 1] Imaging techniques for visualizing and analyzing precise morphology and gene expression patterns are essential for understanding biological processes during development in all organisms. With the aid of chemical screening, we developed a clearing method using chemical solutions, termed ClearSee, for deep imaging of morphology and gene expression in plant tissues. ClearSee rapidly diminishes chlorophyll autofluorescence while maintaining fluorescent protein stability. By adjusting the refractive index mismatch, whole-organ and whole-plant imaging can be performed by both confocal and two-photon excitation microscopy in ClearSee-treated samples. Moreover, ClearSee is applicable to multicolor imaging of fluorescent proteins to allow structural analysis of multiple gene expression. Given that ClearSee is compatible with staining by chemical dyes, the technique is useful for deep imaging in conjunction with genetic markers and for plant species not amenable to transgenic approaches. This method is useful for whole imaging for intact morphology and will help to accelerate the discovery of new phenomena in plant biological research.
LangI, SassmannS, SchmidtB, KomisG ( 2014). Plasmolysis: loss of turgor and beyond Plants (Basel) 3, 583-593. DOI:10.3390/plants3040583URLPMID:27135521 [本文引用: 1] Plasmolysis is a typical response of plant cells exposed to hyperosmotic stress. The loss of turgor causes the violent detachment of the living protoplast from the cell wall. The plasmolytic process is mainly driven by the vacuole. Plasmolysis is reversible (deplasmolysis) and characteristic to living plant cells. Obviously, dramatic structural changes are required to fulfill a plasmolytic cycle. In the present paper, the fate of cortical microtubules and actin microfilaments is documented throughout a plasmolytic cycle in living cells of green fluorescent protein (GFP) tagged Arabidopsis lines. While the microtubules became wavy and highly bundled during plasmolysis, cortical filamentous actin remained in close vicinity to the plasma membrane lining the sites of concave plasmolysis and adjusting readily to the diminished size of the protoplast. During deplasmolysis, cortical microtubule re-organization progressed slowly and required up to 24 h to complete the restoration of the original pre-plasmolytic pattern. Actin microfilaments, again, recovered faster and organelle movement remained intact throughout the whole process. In summary, the hydrostatic skeleton resulting from the osmotic state of the plant vacuole
LerstenNR ( 1986). Modified clearing method to show sieve tubes in minor veins of leaves Stain Technol 61, 231-234. DOI:10.3109/10520298609109942URLPMID:3750353 [本文引用: 1] Leaf samples of Glycine max and numerous other dicotyledonous species were cleared by a common, well established procedure modified by using more concentrated (10% w/v) aqueous NaOH, and by leaving samples in NaOH for 2-4 weeks and in chloral hydrate for 3 days, all at room temperature. A single dye, chlorazol black E (1 g/100 ml absolute ethanol), is used to stain for 3-6 min. Samples are mounted with the lower epidermis upward. Sieve tubes in favorable material can be seen in minor veins and vein endings.
LiSP, ChenM, YuDL, RenSC, SunSF, LiuLD, KetelaarT, EmonsAMC, LiuCM ( 2013). EXO70A1-mediated vesicle trafficking is critical for tracheary element development in Arabidopsis Plant Cell 25, 1774-1786. URLPMID:23709627 [本文引用: 3]
LiSP, van OsGMA, RenSC, YuDL, KetelaarT, EmonsAMC, LiuCM ( 2010). Expression and functional analyses of EXO70 genes in Arabidopsis implicate their roles in regulating cell type-specific exocytosis Plant Physiol 154, 1819-1830. DOI:10.1104/pp.110.164178URLPMID:20943851 [本文引用: 3] During exocytosis, Golgi-derived vesicles are tethered to the target plasma membrane by a conserved octameric complex called the exocyst. In contrast to a single gene in yeast and most animals, plants have greatly increased number of EXO70 genes in their genomes, with functions very much unknown. Reverse transcription-polymerase chain reactions were performed on all 23 EXO70 genes in Arabidopsis (Arabidopsis thaliana) to examine their expression at the organ level. Cell-level expression analyses were performed using transgenic plants carrying beta-glucuronidase reporter constructs, showing that EXO70 genes are primarily expressed in potential exocytosis-active cells such as tip-growing and elongating cells, developing xylem elements, and guard cells, whereas no expression was observed in cells of mature organs such as well-developed leaves, stems, sepals, and petals. Six EXO70 genes are expressed in distinct but partially overlapping stages during microspore development and pollen germination. A mutation in one of these genes, EXO70C1 (At5g13150), led to retarded pollen tube growth and compromised male transmission. This study implies that multiplications of EXO70 genes may allow plants to acquire cell type- and/or cargo-specific regulatory machinery for exocytosis.
LiberatoJR, BarretoRW, ShivasRG ( 2005). Leaf-clearing and staining techniques for the observation of conidiophores in the Phyllactinioideae (Erysiphaceae) Australas Plant Pathol 34, 401-404. DOI:10.1071/AP05027URL [本文引用: 1]
PavelescuI, Vilarrasa-BlasiJ, Planas-RiverolaA, González-GarcíaMP, Ca?o-DelgadoAI, Iba?esM ( 2018). A Sizer model for cell differentiation in Arabidopsis thaliana root growth Mol Syst Biol 14, e7687. DOI:10.15252/msb.20177687URLPMID:29321184 [本文引用: 1] Plant roots grow due to cell division in the meristem and subsequent cell elongation and differentiation, a tightly coordinated process that ensures growth and adaptation to the changing environment. How the newly formed cells decide to stop elongating becoming fully differentiated is not yet understood. To address this question, we established a novel approach that combines the quantitative phenotypic variability of wild-type Arabidopsis roots with computational data from mathematical models. Our analyses reveal that primary root growth is consistent with a Sizer mechanism, in which cells sense their length and stop elongating when reaching a threshold value. The local expression of brassinosteroid receptors only in the meristem is sufficient to set this value. Analysis of roots insensitive to BR signaling and of roots with gibberellin biosynthesis inhibited suggests distinct roles of these hormones on cell expansion termination. Overall, our study underscores the value of using computational modeling together with quantitative data to understand root growth.
RichmondPA, MétrauxJP, TaizL ( 1980). Cell expansion patterns and directionality of wall mechanical properties in Nitella Plant Physiol 65, 211-217. DOI:10.1104/pp.65.2.211URLPMID:16661162 [本文引用: 1] As a means of assessing the extent to which deformation of isolated walls relates to in vivo cell expansion, the directionality of wall mechanical properties was examined in Nitella. Measurements were made of plastic and elastic deformation and creep under both uniaxial and multiaxial stress conditions. Walls of different structural characteristics were obtained from control, isopropyl N-phenylcarbamate (IPC)-treated and IPC recovery cells. Although microfibrils in the inner portion of the wall were transverse for control and recovery cells but random for IPC cells, all walls had similar over-all microfibrillar orientations. Consequently, differences in wall mechanical properties should reflect structural differences in the inner wall. It is the action of the prevailing stress pattern on the inner, not overall, wall microfibrillar organization which dictates the directionality of growth in Nitella. The results indicate that the directional character of expansion is preserved to a large extent in the mechanical properties of isolated walls, and that most, but not all, of the deformation is determined by the inner wall. In addition, directional differences in the threshold for acid-induced extension varied in accord with the pattern of inner wall microfibrils.
ShabalaS, BabourinaO, NewmanI ( 2000). Ion-specific mechanisms of osmoregulation in bean mesophyll cells J Exp Bot 51, 1243-1253. URLPMID:10937700 [本文引用: 1] Transient kinetics of net H+, K+, Ca2+, and Cl- fluxes were measured non-invasively, using an ion-selective microelectrode technique, for bean (Vicia faba L.) leaf mesophyll in response to 150 mM mannitol treatment. In a parallel set of experiments, changes in the plasma membrane potential and the total proline content in leaves were monitored. Regardless of the ionic composition of the bath solution, hyperosmotic stress caused a significant increase in the K+ and Cl- uptake into mesophyll cells. At the same time, no significant proline changes were observed for at least 16 h after the onset of stress. Experiments with inhibitors suggested that potassium inward rectifier (KIR) channels, exhibiting mechanosensitive properties and acting as primary receptors of osmotic stress, are likely to be involved. Due to the coupling by membrane potential, changes in K+ and Cl- transport may modify activity of the plasma membrane H+-pump. Such coupling may also be responsible for the mannitol-induced oscillations (period of about 4 min) in net ion fluxes observed in 90% of plants. Calculations show that influx of K+ and Cl- observed in response to hyperosmotic treatment may provide an adequate osmotic adjustment in bean mesophyll, which suggests that the activity of the plasma membrane transporters for these ions should be targeted to improve osmotolerance, at least in this crop.
ShabalaSN, LewRR ( 2002). Turgor regulation in osmotically stressed Arabidopsis epidermal root cells. Direct support for the role of inorganic ion uptake as revealed by concurrent flux and cell turgor measurements Plant Physiol 129, 290-299. DOI:10.1104/pp.020005URLPMID:12011359 [本文引用: 1] Hyperosmotic stress is known to significantly enhance net uptake of inorganic ions into plant cells. Direct evidence for cell turgor recovery via such a mechanism, however, is still lacking. In the present study, we performed concurrent measurements of net ion fluxes (with the noninvasive microelectrode ion flux estimation technique) and cell turgor changes (with the pressure-probe technique) to provide direct evidence that inorganic ion uptake regulates turgor in osmotically stressed Arabidopsis epidermal root cells. Immediately after onset of hyperosmotic stress (100/100 mM mannitol/sorbitol treatment), the cell turgor dropped from 0.65 to about 0.25 MPa. Turgor recovery started within 2 to 10 min after the treatment and was accompanied by a significant (30-80 nmol m-2 s-1) increase in uptake of K+, Cl-, and Na+ by root cells. In most cells, almost complete (>90% of initial values) recovery of the cell turgor was observed within 40 to 50 min after stress onset. In another set of experiments, we combined the voltage-clamp and the microelectrode ion flux estimation techniques to show that this process is, in part, mediated by voltage-gated K+ transporters at the cell plasma membrane. The possible physiological significance of these findings is discussed.
TakatsukaH, UmedaM ( 2014). Hormonal control of cell division and elongation along differentiation trajectories in roots J Exp Bot 65, 2633-2643. DOI:10.1093/jxb/ert485URLPMID:24474807 [本文引用: 1] The continuous development of roots is supported by a sustainable system for cell production and growth at the root tip. In the stem cell niche that consists of a quiescent centre and surrounding stem cells, an undifferentiated state and low mitotic activity are preserved by the action of auxin and abscisic acid. Stem cell daughters divide several times in the proximal meristem, where auxin and gibberellin mainly promote cell proliferation. Cells then elongate with the help of gibberellin, and become finally differentiated as a constituent of a cell file in the elongation/differentiation zone. In the model plant Arabidopsis thaliana, the transition zone is located between the proximal meristem and the elongation/differentiation zone, and plays an important role in switching from mitosis to the endoreplication that causes DNA polyploidization. Recent studies have shown that cytokinins are essentially required for this transition by antagonizing auxin signalling and promoting degradation of mitotic regulators. In each root zone, different phytohormones interact with one another and coordinately control cell proliferation, cell elongation, cell differentiation, and endoreplication. Such hormonal networks maintain the elaborate structure of the root tip under various environmental conditions. In this review, we summarize and discuss key issues related to hormonal regulation of root growth, and describe how phytohormones are associated with the control of cell cycle machinery.
UrsacheR, AndersenTG, MarhavyP, GeldnerN ( 2018). A protocol for combining fluorescent proteins with histological stains for diverse cell wall components Plant J 93, 399-412. URLPMID:29171896 [本文引用: 1]
VerbelenJP, de CnodderT, LeJ, VissenbergK, Balu?kaF ( 2006). The root apex of Arabidopsis thaliana consists of four distinct zones of growth activities Plant Signal Behav 1, 296-304. DOI:10.4161/psb.1.6.3511URLPMID:19517000 [本文引用: 1] In the growing apex of Arabidopsis thaliana primary roots, cells proceed through four distinct phases of cellular activities. These zones and their boundaries can be well defined based on their characteristic cellular activities. The meristematic zone comprises, and is limited to, all cells that undergo mitotic divisions. Detailed in vivo analysis of transgenic lines reveals that, in the Columbia-0 ecotype, the meristem stretches up to 200 microm away from the junction between root and root cap (RCJ). In the transition zone, 200 to about 520 microm away from the RCJ, cells undergo physiological changes as they prepare for their fast elongation. Upon entering the transition zone, they progressively develop a central vacuole, polarize the cytoskeleton and remodel their cell walls. Cells grow slowly during this transition: it takes ten hours to triplicate cell length from 8.5 to about 35 microm in the trichoblast cell files. In the fast elongation zone, which covers the zone from 520 to about 850 microm from the RCJ, cell length quadruplicates to about 140 microm in only two hours. This is accompanied by drastic and specific cell wall alterations. Finally, root hairs fully develop in the growth terminating zone, where root cells undergo a minor elongation to reach their mature lengths.
VillaniTS, KorochAR, SimonJE ( 2013). An improved clearing and mounting solution to replace chloral hydrate in microscopic applications Appl Plant Sci 1, apps. 1300016. DOI:10.3732/apps.1300016URL [本文引用: 5]
Leukamenin E调节拟南芥幼苗生长发育的模式及其机制 2 2017
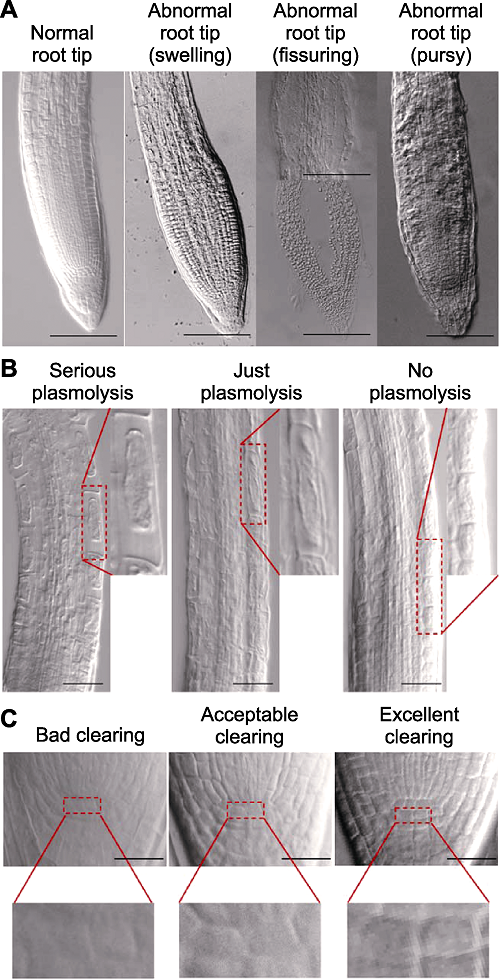
... 整体透明技术是研究植物内部形态结构的重要手段, 可以在不破坏整体结构的情况下对植物样品进行原位观察, 该技术已广泛应用于植物科学研究的诸多领域.植物整体样品之所以不透明, 是由于其组织中的水分、蛋白质、糖类和脂质等多种不同物质的折光系数不同, 当光线通过这些组分的时候会发生散射, 组织样品越厚, 散射就越强, 透射越少, 从而使光学观察受阻(Kurihara et al., 2015; 王培新等, 2016).整体透明技术的原理是利用某些具有高折射率的试剂处理待测样品, 使透射光通过样品时的散射减少, 光通量增加, 并降低球面色差, 以获得较高分辨率的图像(Haseloff, 2003); 此外, 高折射率的介质能够增加视野深度, 使更多层面上的图像进入聚焦平面, 从而达到组织样品透明的效果(Villani et al., 2013).虽然提高透明剂的浓度可以改善透明效果, 但由于透明剂(如水合三氯乙醛)能够改变透明液的渗透势和pH值, 从而对样品细胞形态的保持产生不利影响.因此, 根据材料性质选择适宜的透明液和透明方法是整体透明技术的关键.虽然目前已有多种透明液应用于植物组织透明研究, 但大多适用于叶片或胚珠等细胞壁较厚或液泡较少的组织材料(Lersten, 1986; 杨弘远, 1986, 1988; Liberato et al., 2005; 郝建华和强胜, 2007; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017).根尖作为一种不断保持分裂生长的组织, 细胞壁较薄, 使用现有的透明方法较难维持细胞形态, 造成透明效果的稳定性差(Richmond et al., 1980; Derbyshire et al., 2007; Li et al., 2010, 2013; Villani et al., 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 不仅影响观察对象的清晰度, 还会造成根尖形态不正常和细胞发生质壁分离.虽然整体透明技术已广泛应用于模式植物根尖发育研究(Bougourd et al., 2000; Li et al., 2010, 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 但如何在不影响根尖形态和细胞渗透势的前提下达到对根尖组织最优的透明观察效果, 仍需深入探索实践.本研究以拟南芥(Arabidopsis thaliana)为实验材料, 建立了根尖透明观察体系的评价标准, 即根尖形态是否正常、伸长区细胞是否发生质壁分离及静止中心细胞(QC细胞)的清晰度; 并利用这3个指标, 对透明时间、透明液成分配比和pH值3个要素进行改良, 优化出最适合拟南芥根尖的整体透明观察方法. ...
... 整体透明技术是研究植物内部形态结构的重要手段, 可以在不破坏整体结构的情况下对植物样品进行原位观察, 该技术已广泛应用于植物科学研究的诸多领域.植物整体样品之所以不透明, 是由于其组织中的水分、蛋白质、糖类和脂质等多种不同物质的折光系数不同, 当光线通过这些组分的时候会发生散射, 组织样品越厚, 散射就越强, 透射越少, 从而使光学观察受阻(Kurihara et al., 2015; 王培新等, 2016).整体透明技术的原理是利用某些具有高折射率的试剂处理待测样品, 使透射光通过样品时的散射减少, 光通量增加, 并降低球面色差, 以获得较高分辨率的图像(Haseloff, 2003); 此外, 高折射率的介质能够增加视野深度, 使更多层面上的图像进入聚焦平面, 从而达到组织样品透明的效果(Villani et al., 2013).虽然提高透明剂的浓度可以改善透明效果, 但由于透明剂(如水合三氯乙醛)能够改变透明液的渗透势和pH值, 从而对样品细胞形态的保持产生不利影响.因此, 根据材料性质选择适宜的透明液和透明方法是整体透明技术的关键.虽然目前已有多种透明液应用于植物组织透明研究, 但大多适用于叶片或胚珠等细胞壁较厚或液泡较少的组织材料(Lersten, 1986; 杨弘远, 1986, 1988; Liberato et al., 2005; 郝建华和强胜, 2007; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017).根尖作为一种不断保持分裂生长的组织, 细胞壁较薄, 使用现有的透明方法较难维持细胞形态, 造成透明效果的稳定性差(Richmond et al., 1980; Derbyshire et al., 2007; Li et al., 2010, 2013; Villani et al., 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 不仅影响观察对象的清晰度, 还会造成根尖形态不正常和细胞发生质壁分离.虽然整体透明技术已广泛应用于模式植物根尖发育研究(Bougourd et al., 2000; Li et al., 2010, 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 但如何在不影响根尖形态和细胞渗透势的前提下达到对根尖组织最优的透明观察效果, 仍需深入探索实践.本研究以拟南芥(Arabidopsis thaliana)为实验材料, 建立了根尖透明观察体系的评价标准, 即根尖形态是否正常、伸长区细胞是否发生质壁分离及静止中心细胞(QC细胞)的清晰度; 并利用这3个指标, 对透明时间、透明液成分配比和pH值3个要素进行改良, 优化出最适合拟南芥根尖的整体透明观察方法. ...
... 三氯乙醛和甘油是透明液中的常用成分.三氯乙醛可以提高溶液的折射率, 其在甘油溶液中的折射率达1.428, 接近于玻璃的折射率1.550 (Haseloff, 2003; Villani et al., 2013).这种较高的折射率一方面有助于增大观察体系的数值孔径, 提高分辨率; 另一方面使样品内不同组分的折光率趋于统一, 降低了光线的散射, 提高了光通量.在透明液中加入甘油能够减缓临时装片中水分的蒸发, 防止透明液结晶化(Villani et al., 2013).虽然各种透明液的主要成分相似, 但是不同植物组织样品的整体透明方法不尽相同.目前, 针对胚珠和叶片的整体透明方法较为成熟, 已有Herr 41/2透明、Hoyer氏溶液透明和水杨酸甲酯透明等多种透明体系(Hoyer, 1882; Herr, 1971, 1993; Crane, 1978; Bruzzese and Hasan, 1983; 杨弘远, 1986, 1988; 于明明等, 2009; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017), 但在这些透明体系中并未提出标准化的透明效果评价方案.之前对于胚珠和叶片透明体系的研究是以能够清晰地观察到细胞轮廓作为唯一的评价标准(李彦坤等, 2011; 赵林姝等, 2014; Ursache et al., 2018).相比于叶片, 根尖细胞在胞外渗透压升高时, 胞内膨压降低幅度较大, 使细胞更容易发生质壁分离, 甚至引起根尖整体形态异常(Itoh et al., 1987; Shabala et al., 2000; Shabala and Lew, 2002; Lang et al., 2014; Kim et al., 2018).透明液恰恰形成了一个高渗的胞外环境, 这在以往的研究中经常被忽视(李彦坤等, 2011; Villani et al., 2013; 赵林姝等, 2014).因此, 本文针对透明后的根尖形态、细胞质壁分离情况和透明清晰度, 首次提出了评价拟南芥根尖透明体系的3个量化指标: 形态正常的根尖所占比例、伸长区表皮细胞质壁分离比例及QC细胞清晰的根尖所占比例.综合这三方面评价结果可以更加严谨、完善地对根尖整体透明方法进行分析和改良, 为脆弱组织材料的整体透明研究提供改进依据. ...
... ; 赵林姝等, 2014; Ursache et al., 2018).相比于叶片, 根尖细胞在胞外渗透压升高时, 胞内膨压降低幅度较大, 使细胞更容易发生质壁分离, 甚至引起根尖整体形态异常(Itoh et al., 1987; Shabala et al., 2000; Shabala and Lew, 2002; Lang et al., 2014; Kim et al., 2018).透明液恰恰形成了一个高渗的胞外环境, 这在以往的研究中经常被忽视(李彦坤等, 2011; Villani et al., 2013; 赵林姝等, 2014).因此, 本文针对透明后的根尖形态、细胞质壁分离情况和透明清晰度, 首次提出了评价拟南芥根尖透明体系的3个量化指标: 形态正常的根尖所占比例、伸长区表皮细胞质壁分离比例及QC细胞清晰的根尖所占比例.综合这三方面评价结果可以更加严谨、完善地对根尖整体透明方法进行分析和改良, 为脆弱组织材料的整体透明研究提供改进依据. ...
Analysis of cell division and elongation underlying the developmental acceleration of root growth in Arabidopsis thaliana 1 1998
... 在植物的发育过程研究中, 经常需要对不同苗龄大小的根尖进行透明处理.理论上, 幼苗期的生长发育形成了拟南芥的初生结构, 在这一时期不同苗龄的根尖形态一样(Beemster and Baskin, 1998; Verbelen et al., 2006; Ivanov and Dubrovsky, 2013; Takatsuka and Umeda, 2014; Pavelescu et al., 2018).为了科学严谨地验证本文所筛选出的优选透明方案是否适合整体透明实验中常用的苗龄材料(如3至15日龄幼苗), 我们利用筛选到的透明液HCG-2 (pH7.2)对拟南芥3、9和15日龄幼苗根尖进行透明.结果(图8)表明, 3、9和15日龄幼苗在根尖形态和透明效果上无明显差异.因此, 对于根尖这一特定组织材料, 苗龄大小不会对透明产生不确定性影响.我们改良所得透明液HCG-2 (pH7.2)、透明5分钟的透明方案适用于不同苗龄拟南芥根尖的整体透明. ...
An aniline blue staining procedure for confocal microscopy and 3D imaging of normal and perturbed cellular phenotypes in mature Arabidopsis embryos 2 2000
... 整体透明技术是研究植物内部形态结构的重要手段, 可以在不破坏整体结构的情况下对植物样品进行原位观察, 该技术已广泛应用于植物科学研究的诸多领域.植物整体样品之所以不透明, 是由于其组织中的水分、蛋白质、糖类和脂质等多种不同物质的折光系数不同, 当光线通过这些组分的时候会发生散射, 组织样品越厚, 散射就越强, 透射越少, 从而使光学观察受阻(Kurihara et al., 2015; 王培新等, 2016).整体透明技术的原理是利用某些具有高折射率的试剂处理待测样品, 使透射光通过样品时的散射减少, 光通量增加, 并降低球面色差, 以获得较高分辨率的图像(Haseloff, 2003); 此外, 高折射率的介质能够增加视野深度, 使更多层面上的图像进入聚焦平面, 从而达到组织样品透明的效果(Villani et al., 2013).虽然提高透明剂的浓度可以改善透明效果, 但由于透明剂(如水合三氯乙醛)能够改变透明液的渗透势和pH值, 从而对样品细胞形态的保持产生不利影响.因此, 根据材料性质选择适宜的透明液和透明方法是整体透明技术的关键.虽然目前已有多种透明液应用于植物组织透明研究, 但大多适用于叶片或胚珠等细胞壁较厚或液泡较少的组织材料(Lersten, 1986; 杨弘远, 1986, 1988; Liberato et al., 2005; 郝建华和强胜, 2007; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017).根尖作为一种不断保持分裂生长的组织, 细胞壁较薄, 使用现有的透明方法较难维持细胞形态, 造成透明效果的稳定性差(Richmond et al., 1980; Derbyshire et al., 2007; Li et al., 2010, 2013; Villani et al., 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 不仅影响观察对象的清晰度, 还会造成根尖形态不正常和细胞发生质壁分离.虽然整体透明技术已广泛应用于模式植物根尖发育研究(Bougourd et al., 2000; Li et al., 2010, 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 但如何在不影响根尖形态和细胞渗透势的前提下达到对根尖组织最优的透明观察效果, 仍需深入探索实践.本研究以拟南芥(Arabidopsis thaliana)为实验材料, 建立了根尖透明观察体系的评价标准, 即根尖形态是否正常、伸长区细胞是否发生质壁分离及静止中心细胞(QC细胞)的清晰度; 并利用这3个指标, 对透明时间、透明液成分配比和pH值3个要素进行改良, 优化出最适合拟南芥根尖的整体透明观察方法. ...
Leaf-clearing and staining techniques for the observation of conidiophores in the Phyllactinioideae (Erysiphaceae) 1 2005
... 整体透明技术是研究植物内部形态结构的重要手段, 可以在不破坏整体结构的情况下对植物样品进行原位观察, 该技术已广泛应用于植物科学研究的诸多领域.植物整体样品之所以不透明, 是由于其组织中的水分、蛋白质、糖类和脂质等多种不同物质的折光系数不同, 当光线通过这些组分的时候会发生散射, 组织样品越厚, 散射就越强, 透射越少, 从而使光学观察受阻(Kurihara et al., 2015; 王培新等, 2016).整体透明技术的原理是利用某些具有高折射率的试剂处理待测样品, 使透射光通过样品时的散射减少, 光通量增加, 并降低球面色差, 以获得较高分辨率的图像(Haseloff, 2003); 此外, 高折射率的介质能够增加视野深度, 使更多层面上的图像进入聚焦平面, 从而达到组织样品透明的效果(Villani et al., 2013).虽然提高透明剂的浓度可以改善透明效果, 但由于透明剂(如水合三氯乙醛)能够改变透明液的渗透势和pH值, 从而对样品细胞形态的保持产生不利影响.因此, 根据材料性质选择适宜的透明液和透明方法是整体透明技术的关键.虽然目前已有多种透明液应用于植物组织透明研究, 但大多适用于叶片或胚珠等细胞壁较厚或液泡较少的组织材料(Lersten, 1986; 杨弘远, 1986, 1988; Liberato et al., 2005; 郝建华和强胜, 2007; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017).根尖作为一种不断保持分裂生长的组织, 细胞壁较薄, 使用现有的透明方法较难维持细胞形态, 造成透明效果的稳定性差(Richmond et al., 1980; Derbyshire et al., 2007; Li et al., 2010, 2013; Villani et al., 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 不仅影响观察对象的清晰度, 还会造成根尖形态不正常和细胞发生质壁分离.虽然整体透明技术已广泛应用于模式植物根尖发育研究(Bougourd et al., 2000; Li et al., 2010, 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 但如何在不影响根尖形态和细胞渗透势的前提下达到对根尖组织最优的透明观察效果, 仍需深入探索实践.本研究以拟南芥(Arabidopsis thaliana)为实验材料, 建立了根尖透明观察体系的评价标准, 即根尖形态是否正常、伸长区细胞是否发生质壁分离及静止中心细胞(QC细胞)的清晰度; 并利用这3个指标, 对透明时间、透明液成分配比和pH值3个要素进行改良, 优化出最适合拟南芥根尖的整体透明观察方法. ...
A Sizer model for cell differentiation in Arabidopsis thaliana root growth 1 2018
... 在植物的发育过程研究中, 经常需要对不同苗龄大小的根尖进行透明处理.理论上, 幼苗期的生长发育形成了拟南芥的初生结构, 在这一时期不同苗龄的根尖形态一样(Beemster and Baskin, 1998; Verbelen et al., 2006; Ivanov and Dubrovsky, 2013; Takatsuka and Umeda, 2014; Pavelescu et al., 2018).为了科学严谨地验证本文所筛选出的优选透明方案是否适合整体透明实验中常用的苗龄材料(如3至15日龄幼苗), 我们利用筛选到的透明液HCG-2 (pH7.2)对拟南芥3、9和15日龄幼苗根尖进行透明.结果(图8)表明, 3、9和15日龄幼苗在根尖形态和透明效果上无明显差异.因此, 对于根尖这一特定组织材料, 苗龄大小不会对透明产生不确定性影响.我们改良所得透明液HCG-2 (pH7.2)、透明5分钟的透明方案适用于不同苗龄拟南芥根尖的整体透明. ...
Cell expansion patterns and directionality of wall mechanical properties in Nitella 1 1980
... 整体透明技术是研究植物内部形态结构的重要手段, 可以在不破坏整体结构的情况下对植物样品进行原位观察, 该技术已广泛应用于植物科学研究的诸多领域.植物整体样品之所以不透明, 是由于其组织中的水分、蛋白质、糖类和脂质等多种不同物质的折光系数不同, 当光线通过这些组分的时候会发生散射, 组织样品越厚, 散射就越强, 透射越少, 从而使光学观察受阻(Kurihara et al., 2015; 王培新等, 2016).整体透明技术的原理是利用某些具有高折射率的试剂处理待测样品, 使透射光通过样品时的散射减少, 光通量增加, 并降低球面色差, 以获得较高分辨率的图像(Haseloff, 2003); 此外, 高折射率的介质能够增加视野深度, 使更多层面上的图像进入聚焦平面, 从而达到组织样品透明的效果(Villani et al., 2013).虽然提高透明剂的浓度可以改善透明效果, 但由于透明剂(如水合三氯乙醛)能够改变透明液的渗透势和pH值, 从而对样品细胞形态的保持产生不利影响.因此, 根据材料性质选择适宜的透明液和透明方法是整体透明技术的关键.虽然目前已有多种透明液应用于植物组织透明研究, 但大多适用于叶片或胚珠等细胞壁较厚或液泡较少的组织材料(Lersten, 1986; 杨弘远, 1986, 1988; Liberato et al., 2005; 郝建华和强胜, 2007; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017).根尖作为一种不断保持分裂生长的组织, 细胞壁较薄, 使用现有的透明方法较难维持细胞形态, 造成透明效果的稳定性差(Richmond et al., 1980; Derbyshire et al., 2007; Li et al., 2010, 2013; Villani et al., 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 不仅影响观察对象的清晰度, 还会造成根尖形态不正常和细胞发生质壁分离.虽然整体透明技术已广泛应用于模式植物根尖发育研究(Bougourd et al., 2000; Li et al., 2010, 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 但如何在不影响根尖形态和细胞渗透势的前提下达到对根尖组织最优的透明观察效果, 仍需深入探索实践.本研究以拟南芥(Arabidopsis thaliana)为实验材料, 建立了根尖透明观察体系的评价标准, 即根尖形态是否正常、伸长区细胞是否发生质壁分离及静止中心细胞(QC细胞)的清晰度; 并利用这3个指标, 对透明时间、透明液成分配比和pH值3个要素进行改良, 优化出最适合拟南芥根尖的整体透明观察方法. ...
Ion-specific mechanisms of osmoregulation in bean mesophyll cells 1 2000
... 三氯乙醛和甘油是透明液中的常用成分.三氯乙醛可以提高溶液的折射率, 其在甘油溶液中的折射率达1.428, 接近于玻璃的折射率1.550 (Haseloff, 2003; Villani et al., 2013).这种较高的折射率一方面有助于增大观察体系的数值孔径, 提高分辨率; 另一方面使样品内不同组分的折光率趋于统一, 降低了光线的散射, 提高了光通量.在透明液中加入甘油能够减缓临时装片中水分的蒸发, 防止透明液结晶化(Villani et al., 2013).虽然各种透明液的主要成分相似, 但是不同植物组织样品的整体透明方法不尽相同.目前, 针对胚珠和叶片的整体透明方法较为成熟, 已有Herr 41/2透明、Hoyer氏溶液透明和水杨酸甲酯透明等多种透明体系(Hoyer, 1882; Herr, 1971, 1993; Crane, 1978; Bruzzese and Hasan, 1983; 杨弘远, 1986, 1988; 于明明等, 2009; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017), 但在这些透明体系中并未提出标准化的透明效果评价方案.之前对于胚珠和叶片透明体系的研究是以能够清晰地观察到细胞轮廓作为唯一的评价标准(李彦坤等, 2011; 赵林姝等, 2014; Ursache et al., 2018).相比于叶片, 根尖细胞在胞外渗透压升高时, 胞内膨压降低幅度较大, 使细胞更容易发生质壁分离, 甚至引起根尖整体形态异常(Itoh et al., 1987; Shabala et al., 2000; Shabala and Lew, 2002; Lang et al., 2014; Kim et al., 2018).透明液恰恰形成了一个高渗的胞外环境, 这在以往的研究中经常被忽视(李彦坤等, 2011; Villani et al., 2013; 赵林姝等, 2014).因此, 本文针对透明后的根尖形态、细胞质壁分离情况和透明清晰度, 首次提出了评价拟南芥根尖透明体系的3个量化指标: 形态正常的根尖所占比例、伸长区表皮细胞质壁分离比例及QC细胞清晰的根尖所占比例.综合这三方面评价结果可以更加严谨、完善地对根尖整体透明方法进行分析和改良, 为脆弱组织材料的整体透明研究提供改进依据. ...
Turgor regulation in osmotically stressed Arabidopsis epidermal root cells. Direct support for the role of inorganic ion uptake as revealed by concurrent flux and cell turgor measurements 1 2002
... 三氯乙醛和甘油是透明液中的常用成分.三氯乙醛可以提高溶液的折射率, 其在甘油溶液中的折射率达1.428, 接近于玻璃的折射率1.550 (Haseloff, 2003; Villani et al., 2013).这种较高的折射率一方面有助于增大观察体系的数值孔径, 提高分辨率; 另一方面使样品内不同组分的折光率趋于统一, 降低了光线的散射, 提高了光通量.在透明液中加入甘油能够减缓临时装片中水分的蒸发, 防止透明液结晶化(Villani et al., 2013).虽然各种透明液的主要成分相似, 但是不同植物组织样品的整体透明方法不尽相同.目前, 针对胚珠和叶片的整体透明方法较为成熟, 已有Herr 41/2透明、Hoyer氏溶液透明和水杨酸甲酯透明等多种透明体系(Hoyer, 1882; Herr, 1971, 1993; Crane, 1978; Bruzzese and Hasan, 1983; 杨弘远, 1986, 1988; 于明明等, 2009; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017), 但在这些透明体系中并未提出标准化的透明效果评价方案.之前对于胚珠和叶片透明体系的研究是以能够清晰地观察到细胞轮廓作为唯一的评价标准(李彦坤等, 2011; 赵林姝等, 2014; Ursache et al., 2018).相比于叶片, 根尖细胞在胞外渗透压升高时, 胞内膨压降低幅度较大, 使细胞更容易发生质壁分离, 甚至引起根尖整体形态异常(Itoh et al., 1987; Shabala et al., 2000; Shabala and Lew, 2002; Lang et al., 2014; Kim et al., 2018).透明液恰恰形成了一个高渗的胞外环境, 这在以往的研究中经常被忽视(李彦坤等, 2011; Villani et al., 2013; 赵林姝等, 2014).因此, 本文针对透明后的根尖形态、细胞质壁分离情况和透明清晰度, 首次提出了评价拟南芥根尖透明体系的3个量化指标: 形态正常的根尖所占比例、伸长区表皮细胞质壁分离比例及QC细胞清晰的根尖所占比例.综合这三方面评价结果可以更加严谨、完善地对根尖整体透明方法进行分析和改良, 为脆弱组织材料的整体透明研究提供改进依据. ...
Hormonal control of cell division and elongation along differentiation trajectories in roots 1 2014
... 在植物的发育过程研究中, 经常需要对不同苗龄大小的根尖进行透明处理.理论上, 幼苗期的生长发育形成了拟南芥的初生结构, 在这一时期不同苗龄的根尖形态一样(Beemster and Baskin, 1998; Verbelen et al., 2006; Ivanov and Dubrovsky, 2013; Takatsuka and Umeda, 2014; Pavelescu et al., 2018).为了科学严谨地验证本文所筛选出的优选透明方案是否适合整体透明实验中常用的苗龄材料(如3至15日龄幼苗), 我们利用筛选到的透明液HCG-2 (pH7.2)对拟南芥3、9和15日龄幼苗根尖进行透明.结果(图8)表明, 3、9和15日龄幼苗在根尖形态和透明效果上无明显差异.因此, 对于根尖这一特定组织材料, 苗龄大小不会对透明产生不确定性影响.我们改良所得透明液HCG-2 (pH7.2)、透明5分钟的透明方案适用于不同苗龄拟南芥根尖的整体透明. ...
A protocol for combining fluorescent proteins with histological stains for diverse cell wall components 1 2018
... 三氯乙醛和甘油是透明液中的常用成分.三氯乙醛可以提高溶液的折射率, 其在甘油溶液中的折射率达1.428, 接近于玻璃的折射率1.550 (Haseloff, 2003; Villani et al., 2013).这种较高的折射率一方面有助于增大观察体系的数值孔径, 提高分辨率; 另一方面使样品内不同组分的折光率趋于统一, 降低了光线的散射, 提高了光通量.在透明液中加入甘油能够减缓临时装片中水分的蒸发, 防止透明液结晶化(Villani et al., 2013).虽然各种透明液的主要成分相似, 但是不同植物组织样品的整体透明方法不尽相同.目前, 针对胚珠和叶片的整体透明方法较为成熟, 已有Herr 41/2透明、Hoyer氏溶液透明和水杨酸甲酯透明等多种透明体系(Hoyer, 1882; Herr, 1971, 1993; Crane, 1978; Bruzzese and Hasan, 1983; 杨弘远, 1986, 1988; 于明明等, 2009; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017), 但在这些透明体系中并未提出标准化的透明效果评价方案.之前对于胚珠和叶片透明体系的研究是以能够清晰地观察到细胞轮廓作为唯一的评价标准(李彦坤等, 2011; 赵林姝等, 2014; Ursache et al., 2018).相比于叶片, 根尖细胞在胞外渗透压升高时, 胞内膨压降低幅度较大, 使细胞更容易发生质壁分离, 甚至引起根尖整体形态异常(Itoh et al., 1987; Shabala et al., 2000; Shabala and Lew, 2002; Lang et al., 2014; Kim et al., 2018).透明液恰恰形成了一个高渗的胞外环境, 这在以往的研究中经常被忽视(李彦坤等, 2011; Villani et al., 2013; 赵林姝等, 2014).因此, 本文针对透明后的根尖形态、细胞质壁分离情况和透明清晰度, 首次提出了评价拟南芥根尖透明体系的3个量化指标: 形态正常的根尖所占比例、伸长区表皮细胞质壁分离比例及QC细胞清晰的根尖所占比例.综合这三方面评价结果可以更加严谨、完善地对根尖整体透明方法进行分析和改良, 为脆弱组织材料的整体透明研究提供改进依据. ...
The root apex of Arabidopsis thaliana consists of four distinct zones of growth activities 1 2006
... 在植物的发育过程研究中, 经常需要对不同苗龄大小的根尖进行透明处理.理论上, 幼苗期的生长发育形成了拟南芥的初生结构, 在这一时期不同苗龄的根尖形态一样(Beemster and Baskin, 1998; Verbelen et al., 2006; Ivanov and Dubrovsky, 2013; Takatsuka and Umeda, 2014; Pavelescu et al., 2018).为了科学严谨地验证本文所筛选出的优选透明方案是否适合整体透明实验中常用的苗龄材料(如3至15日龄幼苗), 我们利用筛选到的透明液HCG-2 (pH7.2)对拟南芥3、9和15日龄幼苗根尖进行透明.结果(图8)表明, 3、9和15日龄幼苗在根尖形态和透明效果上无明显差异.因此, 对于根尖这一特定组织材料, 苗龄大小不会对透明产生不确定性影响.我们改良所得透明液HCG-2 (pH7.2)、透明5分钟的透明方案适用于不同苗龄拟南芥根尖的整体透明. ...
An improved clearing and mounting solution to replace chloral hydrate in microscopic applications 5 2013
... 整体透明技术是研究植物内部形态结构的重要手段, 可以在不破坏整体结构的情况下对植物样品进行原位观察, 该技术已广泛应用于植物科学研究的诸多领域.植物整体样品之所以不透明, 是由于其组织中的水分、蛋白质、糖类和脂质等多种不同物质的折光系数不同, 当光线通过这些组分的时候会发生散射, 组织样品越厚, 散射就越强, 透射越少, 从而使光学观察受阻(Kurihara et al., 2015; 王培新等, 2016).整体透明技术的原理是利用某些具有高折射率的试剂处理待测样品, 使透射光通过样品时的散射减少, 光通量增加, 并降低球面色差, 以获得较高分辨率的图像(Haseloff, 2003); 此外, 高折射率的介质能够增加视野深度, 使更多层面上的图像进入聚焦平面, 从而达到组织样品透明的效果(Villani et al., 2013).虽然提高透明剂的浓度可以改善透明效果, 但由于透明剂(如水合三氯乙醛)能够改变透明液的渗透势和pH值, 从而对样品细胞形态的保持产生不利影响.因此, 根据材料性质选择适宜的透明液和透明方法是整体透明技术的关键.虽然目前已有多种透明液应用于植物组织透明研究, 但大多适用于叶片或胚珠等细胞壁较厚或液泡较少的组织材料(Lersten, 1986; 杨弘远, 1986, 1988; Liberato et al., 2005; 郝建华和强胜, 2007; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017).根尖作为一种不断保持分裂生长的组织, 细胞壁较薄, 使用现有的透明方法较难维持细胞形态, 造成透明效果的稳定性差(Richmond et al., 1980; Derbyshire et al., 2007; Li et al., 2010, 2013; Villani et al., 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 不仅影响观察对象的清晰度, 还会造成根尖形态不正常和细胞发生质壁分离.虽然整体透明技术已广泛应用于模式植物根尖发育研究(Bougourd et al., 2000; Li et al., 2010, 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 但如何在不影响根尖形态和细胞渗透势的前提下达到对根尖组织最优的透明观察效果, 仍需深入探索实践.本研究以拟南芥(Arabidopsis thaliana)为实验材料, 建立了根尖透明观察体系的评价标准, 即根尖形态是否正常、伸长区细胞是否发生质壁分离及静止中心细胞(QC细胞)的清晰度; 并利用这3个指标, 对透明时间、透明液成分配比和pH值3个要素进行改良, 优化出最适合拟南芥根尖的整体透明观察方法. ...
... ; Villani et al., 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 不仅影响观察对象的清晰度, 还会造成根尖形态不正常和细胞发生质壁分离.虽然整体透明技术已广泛应用于模式植物根尖发育研究(Bougourd et al., 2000; Li et al., 2010, 2013; 程蔚玲等, 2017; 李芳芳等, 2018), 但如何在不影响根尖形态和细胞渗透势的前提下达到对根尖组织最优的透明观察效果, 仍需深入探索实践.本研究以拟南芥(Arabidopsis thaliana)为实验材料, 建立了根尖透明观察体系的评价标准, 即根尖形态是否正常、伸长区细胞是否发生质壁分离及静止中心细胞(QC细胞)的清晰度; 并利用这3个指标, 对透明时间、透明液成分配比和pH值3个要素进行改良, 优化出最适合拟南芥根尖的整体透明观察方法. ...
... 三氯乙醛和甘油是透明液中的常用成分.三氯乙醛可以提高溶液的折射率, 其在甘油溶液中的折射率达1.428, 接近于玻璃的折射率1.550 (Haseloff, 2003; Villani et al., 2013).这种较高的折射率一方面有助于增大观察体系的数值孔径, 提高分辨率; 另一方面使样品内不同组分的折光率趋于统一, 降低了光线的散射, 提高了光通量.在透明液中加入甘油能够减缓临时装片中水分的蒸发, 防止透明液结晶化(Villani et al., 2013).虽然各种透明液的主要成分相似, 但是不同植物组织样品的整体透明方法不尽相同.目前, 针对胚珠和叶片的整体透明方法较为成熟, 已有Herr 41/2透明、Hoyer氏溶液透明和水杨酸甲酯透明等多种透明体系(Hoyer, 1882; Herr, 1971, 1993; Crane, 1978; Bruzzese and Hasan, 1983; 杨弘远, 1986, 1988; 于明明等, 2009; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017), 但在这些透明体系中并未提出标准化的透明效果评价方案.之前对于胚珠和叶片透明体系的研究是以能够清晰地观察到细胞轮廓作为唯一的评价标准(李彦坤等, 2011; 赵林姝等, 2014; Ursache et al., 2018).相比于叶片, 根尖细胞在胞外渗透压升高时, 胞内膨压降低幅度较大, 使细胞更容易发生质壁分离, 甚至引起根尖整体形态异常(Itoh et al., 1987; Shabala et al., 2000; Shabala and Lew, 2002; Lang et al., 2014; Kim et al., 2018).透明液恰恰形成了一个高渗的胞外环境, 这在以往的研究中经常被忽视(李彦坤等, 2011; Villani et al., 2013; 赵林姝等, 2014).因此, 本文针对透明后的根尖形态、细胞质壁分离情况和透明清晰度, 首次提出了评价拟南芥根尖透明体系的3个量化指标: 形态正常的根尖所占比例、伸长区表皮细胞质壁分离比例及QC细胞清晰的根尖所占比例.综合这三方面评价结果可以更加严谨、完善地对根尖整体透明方法进行分析和改良, 为脆弱组织材料的整体透明研究提供改进依据. ...
... ).这种较高的折射率一方面有助于增大观察体系的数值孔径, 提高分辨率; 另一方面使样品内不同组分的折光率趋于统一, 降低了光线的散射, 提高了光通量.在透明液中加入甘油能够减缓临时装片中水分的蒸发, 防止透明液结晶化(Villani et al., 2013).虽然各种透明液的主要成分相似, 但是不同植物组织样品的整体透明方法不尽相同.目前, 针对胚珠和叶片的整体透明方法较为成熟, 已有Herr 41/2透明、Hoyer氏溶液透明和水杨酸甲酯透明等多种透明体系(Hoyer, 1882; Herr, 1971, 1993; Crane, 1978; Bruzzese and Hasan, 1983; 杨弘远, 1986, 1988; 于明明等, 2009; 王文婧等, 2011; 赵林姝等, 2014; 任媛媛和朱炎, 2017), 但在这些透明体系中并未提出标准化的透明效果评价方案.之前对于胚珠和叶片透明体系的研究是以能够清晰地观察到细胞轮廓作为唯一的评价标准(李彦坤等, 2011; 赵林姝等, 2014; Ursache et al., 2018).相比于叶片, 根尖细胞在胞外渗透压升高时, 胞内膨压降低幅度较大, 使细胞更容易发生质壁分离, 甚至引起根尖整体形态异常(Itoh et al., 1987; Shabala et al., 2000; Shabala and Lew, 2002; Lang et al., 2014; Kim et al., 2018).透明液恰恰形成了一个高渗的胞外环境, 这在以往的研究中经常被忽视(李彦坤等, 2011; Villani et al., 2013; 赵林姝等, 2014).因此, 本文针对透明后的根尖形态、细胞质壁分离情况和透明清晰度, 首次提出了评价拟南芥根尖透明体系的3个量化指标: 形态正常的根尖所占比例、伸长区表皮细胞质壁分离比例及QC细胞清晰的根尖所占比例.综合这三方面评价结果可以更加严谨、完善地对根尖整体透明方法进行分析和改良, 为脆弱组织材料的整体透明研究提供改进依据. ...
 ,*齐鲁师范学院, 济南 250200
,*齐鲁师范学院, 济南 250200

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT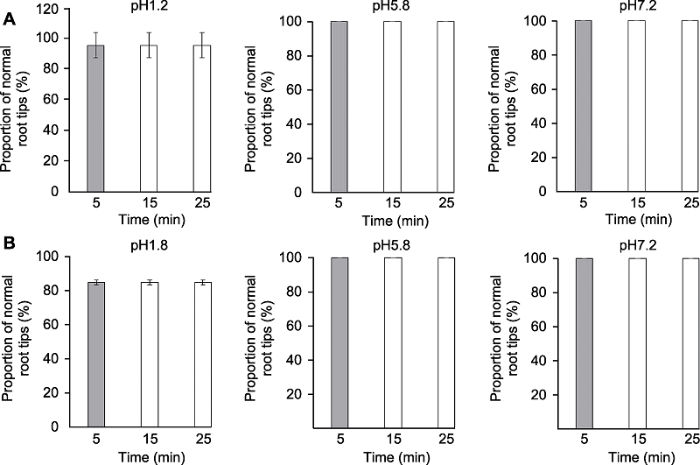 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT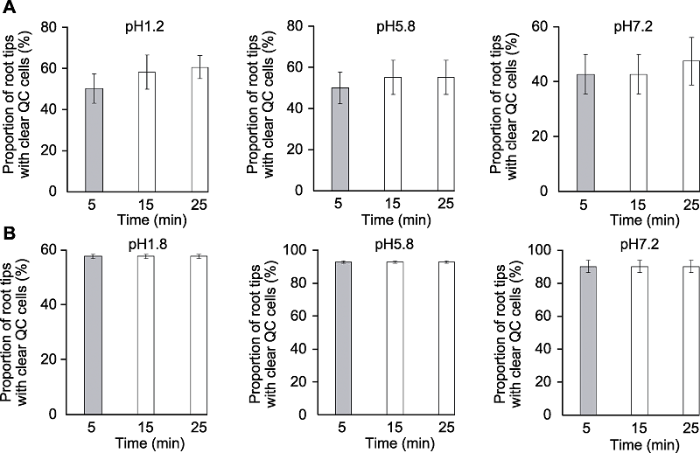 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT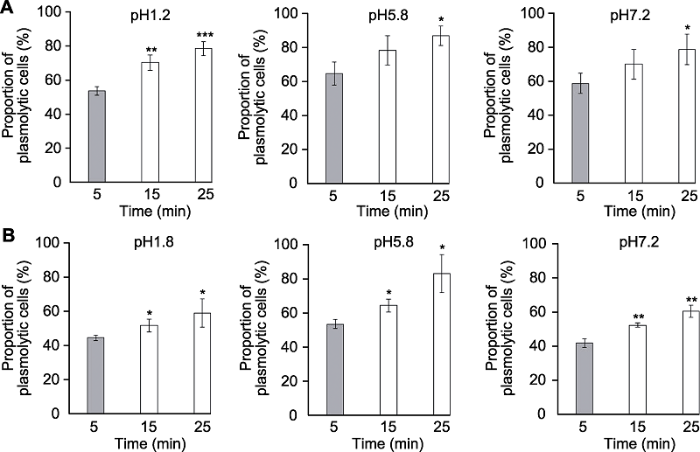 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT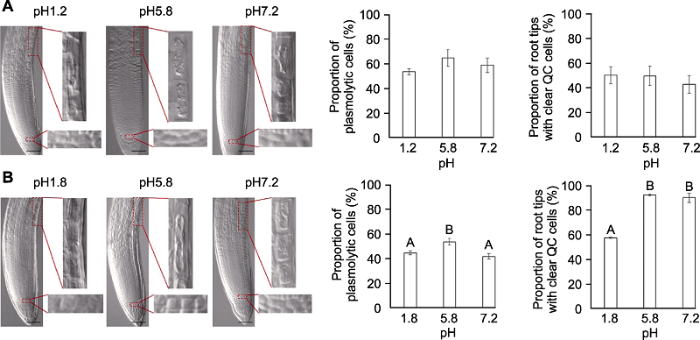 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT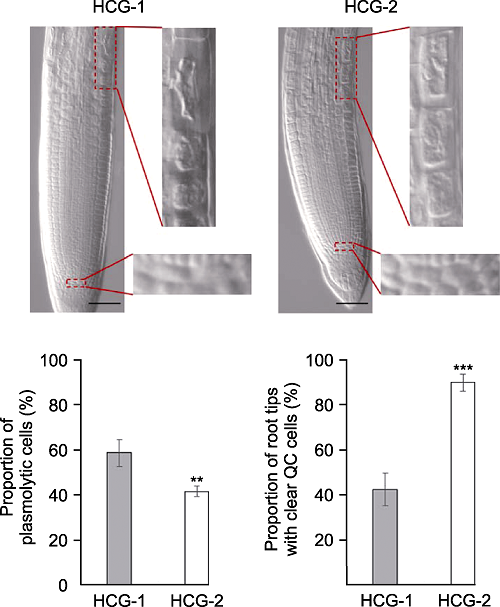 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT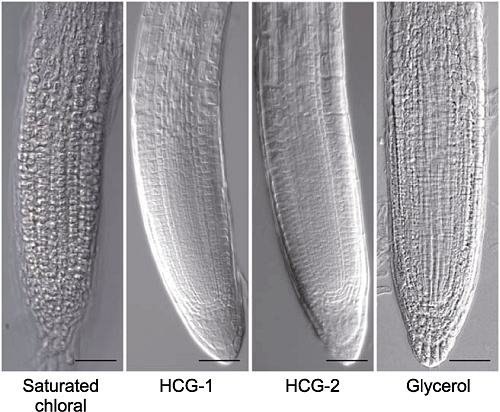 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT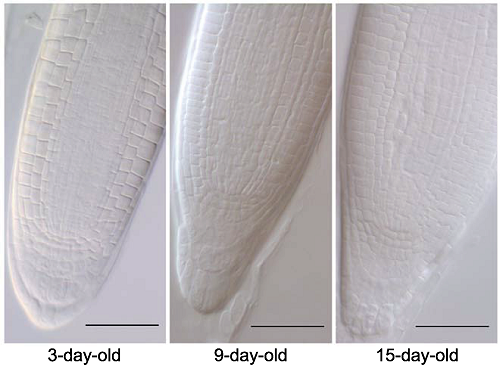 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}