Antifungal Activity and Mechanisms of Natamycin Against Colletotrichum gloeosporioides in Postharvest Mango Fruit
Jiayi Liu1, Jiaxin Wang1, Haichao Song2, Zhengke Zhang1, Xiangbin Xu1, Xuncong Ji,3,*, Xuequn Shi,1,*1 College of Food Science and Technology, Hainan University, Haikou 570228, China 2 Institute of Tropical Agriculture and Forestry, Hainan University, Haikou 570228, China 3 Institute of Plant Protection, Hainan Academy of Agricultural Sciences, Haikou 571100, China
Abstract In this study, we examined the inhibitory effects of natamycin at different concentrations on the conidial germination and mycelial growth of Colletotrichum gloeosporioides in vitro as well as the controlled effect of natamycin on postharvest anthracnose of mango (Mangifera indica) fruit inoculated with C. gloeosporioides. To further explore the underlying antifungal mechanism, we analyzed the membrane permeability, soluble protein content, changes in cell membrane integrity, intracellular reactive oxygen species (ROS) level and mitochondrial distribution in C. gloeosporioides after natamycin treatment. Natamycin at 3 mg?L -1 effectively suppressed the conidial germination, germ tube elongation and mycelial growth of C. gloeosporioides. Also, 80 mg?L -1natamycin significantly inhibited the expansion of anthracnose lesions in mango fruit during storage. Furthermore, natamycin treatment increased the relative permeability and soluble protein content in the cell membrane of C. gloeosporioides. After 8h treatment with natamycin 2 mg?L -1, the staining rate of damaged cell membranes in C. gloeosporioides was 33.6% and 13.9% in the control. The staining rate of intracellular ROS reached 46.9% in treated conidia, which was 39.7% higher than that of the control. Natamycin treatment caused heterogeneous distribution of intracellular mitochondria along with weaker fluorescence as compared with the control. In summary, natamycin can destroy the cell membrane of C. gloeosporioides, induce ROS accumulation and reduce mitochondrial activity, thus interfering in the normal physiological activity of C. gloeosporioides and affecting its metabolic activities. Keywords:natamycin;mango;Colletotrichum gloeosporioides;antifungal mechanism
PDF (5925KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 刘佳怡, 王嘉欣, 宋海超, 张正科, 徐祥彬, 吉训聪, 史学群. 纳他霉素对芒果采后胶孢炭疽菌的抑菌效果及机理. 植物学报, 2019, 54(4): 455-463 doi:10.11983/CBB19044 Liu Jiayi, Wang Jiaxin, Song Haichao, Zhang Zhengke, Xu Xiangbin, Ji Xuncong, Shi Xuequn. Antifungal Activity and Mechanisms of Natamycin Against Colletotrichum gloeosporioides in Postharvest Mango Fruit. Chinese Bulletin of Botany, 2019, 54(4): 455-463 doi:10.11983/CBB19044
芒果(Mangifera indica)是热带和亚热带地区重要的经济作物, 其口味醇香甘甜, 营养丰富, 深受消费者喜爱(Sivakumar et al., 2011)。但它作为一种呼吸跃变型果实, 后熟期间极易受到各种病原菌的侵染, 引起炭疽病和蒂腐病等采后病害, 使芒果在运输和贮藏期间发生组织腐烂, 造成果实品质下降, 常带来严重的经济损失(Jongsri et al., 2017; Perumal et al., 2017)。胶孢炭疽菌(Colletotrichum gloeosporioides)是引起芒果炭疽病的主要病原菌, 可潜伏侵染果实多个部位, 常在果实成熟期引发症状(Kefialew and Ayalew, 2008)。目前, 我国采后病害的防治主要依赖化学杀菌剂, 如多菌灵、噻苯达唑和苯菌灵, 但化学杀菌剂的大量施用会使病原菌产生耐药性, 减弱杀菌效果(Hu et al., 2014)。随着人们对农药残留、食品安全及环境污染等问题的日渐关注, 开发低毒高效的环保型抑菌剂成为防治采后病害的重要途径。
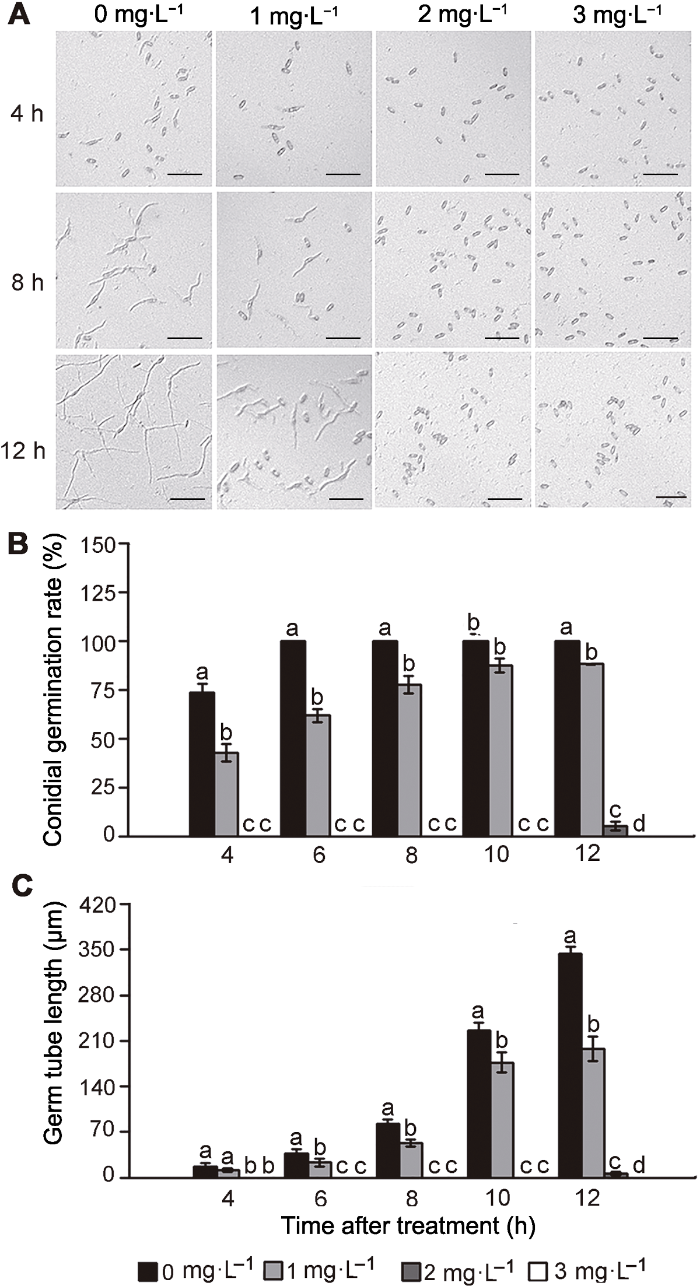
(A) 孢子萌发显微图(Bars=50 μm); (B) 孢子萌发率; (C) 芽管长度。不同小写字母表示各处理组间差异显著(P<0.05)。 Figure 1Effect of natamycin on conidial germination and germ tube elongation of Colletotrichum gloeosporioides
(A) Micrograph of conidial germination (Bars=50 μm); (B) Conidial germination rate; (C) Germ tube length. Treatments followed by different lowercase letters are statistically different by the Duncan’s multiple range test (P<0.05).
2.2 纳他霉素对胶孢炭疽菌菌丝生长的影响
由图2可知, 纳他霉素对胶孢炭疽菌的菌丝生长有显著抑制作用, 各浓度间差异显著(P<0.05), 且抑菌效果随浓度的增加而增强。培养至第5天, 对照组菌落直径达67.17 mm, 1和2 mg∙L-1纳他霉素处理后的菌落直径分别为52.5和38.33 mm, 3 mg∙L-1处理组菌落直径仅为27.83 mm。
(A) 菌落生长图(Bar=15 mm); (B) 菌落直径。不同小写字母表示各处理组间差异显著(P<0.05)。 Figure 2Effect of natamycin on colony growth of Colletotrichum gloeosporioides
(A) Colony morphology (Bar=15 mm); (B) Colony diameter. Treatments followed by different lowercase letters are statistically different by the Duncan’s multiple range test (P<0.05).
2.3 纳他霉素对芒果炭疽病斑扩展的影响
由图3可知, 随着贮藏时间的延长, 各组芒果果实病斑直径均逐渐增大。但经80和160 mg∙L-1纳他霉素处理的果实, 炭疽病病斑直径明显小于对照组, 且各浓度之间差异显著(P<0.05), 抑菌效果随浓度增加而增强。贮存至第8天, 对照组病斑直径达16.10 mm, 而80和160 mg∙L-1纳他霉素处理的果实病斑直径仅分别为12.5和7.18 mm, 病斑扩展分别减少22.36%和55.40%。
(A) 炭疽病发病情况(Bar=8 mm); (B) 炭疽病斑直径。不同小写字母表示各处理组间差异显著(P<0.05)。 Figure 3Effect of natamycin on anthracnose in mango fruit
(A) Symptoms of anthracnose in fruit (Bar=8 mm); (B) Lesion diameter of anthracnose. Treatments followed by different lowercase letters are statistically different by the Duncan’s multiple range test (P<0.05).
(A) 荧光显微镜下碘化丙啶(PI)染色情况, 细胞膜受损的孢子呈红色荧光(Bars=50 μm); (B) PI染色率。不同小写字母表示各处理组间差异显著(P<0.05)。 Figure 5Effect of natamycin on membrane integrity of Colletotrichum gloeosporioides conidia
(A) Propidium iodide (PI) staining under fluorescence microscope, conidia with damaged plasma membranes showed red fluorescence (Bars=50 μm); (B) PI staining rate. Treatments followed by different lowercase letters are statistically different by the Duncan’s multiple range test (P<0.05).
(A) 荧光显微镜下DCFH-DA染色情况, 诱导活性氧产生的孢子呈绿色荧光(Bars=50 μm); (B) DCFH-DA染色率。不同小写字母表示各处理组间差异显著(P<0.05)。 Figure 6Effect of natamycin on reactive oxygen species (ROS) content of Colletotrichum gloeosporioides
(A) DCFH-DA staining under fluorescence microscope, conidia with intracellular ROS induction exhibit green fluorescence (Bars=50 μm); (B) DCFH-DA staining rate. Treatments followed by different lowercase letters are statistically different by the Duncan’s multiple range test (P<0.05).
AngelovaMB, PashovaSB, SpasovaBK, VassilevSV, SlokoskaLS ( 2005). Oxidative stress response of filamentous fungi induced by hydrogen peroxide and paraquat Mycol Res 109, 150-158. [本文引用: 1]
AparicioJF, BarrealesEG, PayeroTD, VicenteCM, de PedroA, Santos-AberturasJ ( 2016). Biotechnological production and application of the antibiotic pimaricin: biosynthesis and its regulation Appl Microbiol Biotechnol 100, 61-78. [本文引用: 1]
ChanDC ( 2006). Mitochondria: dynamic organelles in disease, aging, and development Cell 125, 1241-1252. [本文引用: 1]
CircuML, AwTY ( 2010). Reactive oxygen species, cellular redox systems, and apoptosis Free Radic Biol Med 48, 749-762. [本文引用: 1]
FajardoP, MartinsJT, Fuci?osC, PastranaL, TeixeiraJA, VicenteAA ( 2010). Evaluation of a chitosan-based edible film as carrier of natamycin to improve the storability of saloio cheese J Food Eng 101, 349-356. [本文引用: 1]
HaackSE, IvorsKL, HolmesGJ, F?rsterH, AdaskavegJE ( 2018). Natamycin, a new biofungicide for managing crown rot of strawberry caused by QoI-resistant Colletotrichum acutatum Plant Dis 102, 1687-1695. [本文引用: 1]
HeC, ZhangZQ, LiBQ, XuY, TianSP ( 2019). Effect of natamycin on Botrytis cinerea and Penicillium expansum-postharvest pathogens of grape berries and jujube fruit Postharvest Biol Technol 151, 134-141. [本文引用: 1]
HondrodimouO, KourkoutasY, PanagouEZ ( 2011). Efficacy of natamycin to control fungal growth in natural black olive fermentation Food Microbiol 28, 621-627. [本文引用: 1]
HuMJ, YangDP, HuberDJ, JiangYM, LiM, GaoZY, ZhangZK ( 2014). Reduction of postharvest anthracnose and enhancement of disease resistance in ripening mango fruit by nitric oxide treatment Postharvest Biol Technol 97, 115-122. [本文引用: 1]
IndoHP, DavidsonM, YenHC, SuenagaS, TomitaK, NishiiT, HiguchiM, KogaY, OzawaT, MajimaHJ ( 2007). Evidence of ROS generation by mitochondria in cells with impaired electron transport chain and mitochondrial DNA damage Mitochondrion 7, 106-118. [本文引用: 1]
JongsriP, RojsitthisakP, WangsomboondeeT, SeraypheapK ( 2017). Influence of chitosan coating combined with spermidine on anthracnose disease and qualities of ‘Nam Dok Mai’ mango after harvest Sci Hortic 224, 180-187. [本文引用: 1]
KefialewY, AyalewA ( 2008). Postharvest biological control of anthracnose (Colletotrichum gloeosporioides) on mango( Mangifera indica) Postharvest Biol Technol 50, 8-11. [本文引用: 1]
LaiTF, LiBQ, QinGZ, TianSP ( 2011). Oxidative damage involves in the inhibitory effect of nitric oxide on spore germination of Penicillium expansum Curr Microbiol 62, 229-234. [本文引用: 1]
MarchiS, GiorgiC, SuskiJM, AgnolettoC, BononiA, BonoraM, De MarchiE, MissiroliS, PatergnaniS, PolettiF, RimessiA, DuszynskiJ, WieckowskiMR, PintonP ( 2012). Mitochondria-ros crosstalk in the control of cell death and aging J Signal Transduction 2012, 329635. [本文引用: 1]
MehyarGF, Al NabulsiAA, SalehM, OlaimatAN, HolleyRA ( 2017). Effects of chitosan coating containingly- sozyme or natamycin on shelf-life, microbial quality, and sensory properties of Halloumi cheese brined in normal and reduced salt solutions J Food Process Pres 42, e13324. [本文引用: 1]
PerumalAB, SellamuthuPS, NambiarRB, SadikuER ( 2018). Effects of essential oil vapour treatment on the postharvest disease control and different defence responses in two mango (Mangifera indica L.) cultivars Food Bioprocess Tech 10, 1131-1141. [本文引用: 1]
PipekP, RohlíkBA, LojkováA, StaruchL ( 2010). Suppression of mould growth on dry sausages Czech Food Sci 28, 258-263. [本文引用: 1]
ShiXQ, LiBQ, QinGZ, TianSP ( 2012). Mechanism of antifungal action of borate against Colletotrichum gloeosporioides related to mitochondrial degradation in spores Postharvest Biol Technol 67, 138-143. [本文引用: 1]
SivakumarD, JiangY, YahiaEM ( 2011). Maintaining mango (Mangifera indica L.) fruit quality during the export chain Food Res Int 44, 1254-1263. [本文引用: 1]
te WelscherYM, JonesL, van LeeuwenMR, DijksterhuisJ, de KruijffB, EitzenG, BreukinkE ( 2010). Natamycin inhibits vacuole fusion at the priming phase via a specific interaction with ergosterol Antimicrob Agents Chemother 54, 2618-2625. [本文引用: 1]
Van LeeuwenMR, GolovinaEA, DijksterhuisJ ( 2009). The polyene antimycotics nystatin and filipin disrupt the plasma membrane, whereas natamycin inhibits endocytosis in germinating conidia of Penicillium discolor J Appl Microbiol 106, 1908-1918. [本文引用: 1]
Reactive oxygen species, cellular redox systems, and apoptosis 1 2010
... 活性氧是呼吸作用常见的副产物, 但高浓度活性氧能够与脂类、蛋白质和DNA反应, 产生蛋白羰基化、脂质过氧化或mtDNA损伤, 使细胞发生功能障碍, 导致膜破损或细胞死亡(Circu and Aw, 2010; Marchi et al., 2012).已有研究表明, 许多外源性刺激都会使真菌病原体产生诱导活性氧大量生成的应激反应(Xu and Tian, 2008; Lai et al., 2011).本研究中胶孢炭疽菌经纳他霉素处理后, 孢子内活性氧含量显著升高, 说明纳他霉素可以通过诱导活性氧大量产生而对菌体造成伤害. ...
Evaluation of a chitosan-based edible film as carrier of natamycin to improve the storability of saloio cheese 1 2010
... 纳他霉素是纳他链霉菌(Streptomyces natalensis)发酵过程中产生的天然多烯大环内酯类物质, 对霉菌和酵母有较强的抑制作用, 被公认为安全有效的食品防腐剂之一(Hondrodimou et al., 2011).纳他霉素可与真菌细胞膜上的麦角甾醇或其它甾醇类物质结合, 阻遏麦角甾醇的生物合成, 使细胞膜畸变, 从而引起细胞死亡(Aparicio et al., 2016).纳他霉素具有低毒高效、抑菌谱广、无污染和无抗药性等优点(Arroyo-López et al., 2012), 包括美国在内的许多国家已将纳他霉素纳入食品添加剂目录, 广泛用于肉类、乳制品防腐(Pipek et al., 2010; Mehyar et al., 2017)和果蔬采后保鲜(Fajardo et al., 2010).在草莓(Fragaria ananassa) (呼玉侠等, 2006)、柑橘(Citrus reticulata) (孙远功等, 2006)和樱桃(Prunus avium) (姜爱丽等, 2009)等水果中添加不同浓度的纳他霉素, 对防止果实采后腐烂、提高贮藏品质有显著效果.研究表明, 纳他霉素可以有效抑制灰葡萄孢(Botrytis cinerea)、扩展青霉(Penicillium expansum)和尖孢炭疽菌(Colletotrichum acutatum)的孢子萌发和菌丝生长, 从而控制采后病害的发生(郭萌萌等, 2013; Haack et al., 2018; He et al., 2019).目前, 纳他霉素对芒果采后胶孢炭疽菌的抑菌效果和作用机理研究较少. ...
Natamycin, a new biofungicide for managing crown rot of strawberry caused by QoI-resistant Colletotrichum acutatum 1 2018
... 纳他霉素是纳他链霉菌(Streptomyces natalensis)发酵过程中产生的天然多烯大环内酯类物质, 对霉菌和酵母有较强的抑制作用, 被公认为安全有效的食品防腐剂之一(Hondrodimou et al., 2011).纳他霉素可与真菌细胞膜上的麦角甾醇或其它甾醇类物质结合, 阻遏麦角甾醇的生物合成, 使细胞膜畸变, 从而引起细胞死亡(Aparicio et al., 2016).纳他霉素具有低毒高效、抑菌谱广、无污染和无抗药性等优点(Arroyo-López et al., 2012), 包括美国在内的许多国家已将纳他霉素纳入食品添加剂目录, 广泛用于肉类、乳制品防腐(Pipek et al., 2010; Mehyar et al., 2017)和果蔬采后保鲜(Fajardo et al., 2010).在草莓(Fragaria ananassa) (呼玉侠等, 2006)、柑橘(Citrus reticulata) (孙远功等, 2006)和樱桃(Prunus avium) (姜爱丽等, 2009)等水果中添加不同浓度的纳他霉素, 对防止果实采后腐烂、提高贮藏品质有显著效果.研究表明, 纳他霉素可以有效抑制灰葡萄孢(Botrytis cinerea)、扩展青霉(Penicillium expansum)和尖孢炭疽菌(Colletotrichum acutatum)的孢子萌发和菌丝生长, 从而控制采后病害的发生(郭萌萌等, 2013; Haack et al., 2018; He et al., 2019).目前, 纳他霉素对芒果采后胶孢炭疽菌的抑菌效果和作用机理研究较少. ...
Effect of natamycin on Botrytis cinerea and Penicillium expansum-postharvest pathogens of grape berries and jujube fruit 1 2019
... 纳他霉素是纳他链霉菌(Streptomyces natalensis)发酵过程中产生的天然多烯大环内酯类物质, 对霉菌和酵母有较强的抑制作用, 被公认为安全有效的食品防腐剂之一(Hondrodimou et al., 2011).纳他霉素可与真菌细胞膜上的麦角甾醇或其它甾醇类物质结合, 阻遏麦角甾醇的生物合成, 使细胞膜畸变, 从而引起细胞死亡(Aparicio et al., 2016).纳他霉素具有低毒高效、抑菌谱广、无污染和无抗药性等优点(Arroyo-López et al., 2012), 包括美国在内的许多国家已将纳他霉素纳入食品添加剂目录, 广泛用于肉类、乳制品防腐(Pipek et al., 2010; Mehyar et al., 2017)和果蔬采后保鲜(Fajardo et al., 2010).在草莓(Fragaria ananassa) (呼玉侠等, 2006)、柑橘(Citrus reticulata) (孙远功等, 2006)和樱桃(Prunus avium) (姜爱丽等, 2009)等水果中添加不同浓度的纳他霉素, 对防止果实采后腐烂、提高贮藏品质有显著效果.研究表明, 纳他霉素可以有效抑制灰葡萄孢(Botrytis cinerea)、扩展青霉(Penicillium expansum)和尖孢炭疽菌(Colletotrichum acutatum)的孢子萌发和菌丝生长, 从而控制采后病害的发生(郭萌萌等, 2013; Haack et al., 2018; He et al., 2019).目前, 纳他霉素对芒果采后胶孢炭疽菌的抑菌效果和作用机理研究较少. ...
Efficacy of natamycin to control fungal growth in natural black olive fermentation 1 2011
... 纳他霉素是纳他链霉菌(Streptomyces natalensis)发酵过程中产生的天然多烯大环内酯类物质, 对霉菌和酵母有较强的抑制作用, 被公认为安全有效的食品防腐剂之一(Hondrodimou et al., 2011).纳他霉素可与真菌细胞膜上的麦角甾醇或其它甾醇类物质结合, 阻遏麦角甾醇的生物合成, 使细胞膜畸变, 从而引起细胞死亡(Aparicio et al., 2016).纳他霉素具有低毒高效、抑菌谱广、无污染和无抗药性等优点(Arroyo-López et al., 2012), 包括美国在内的许多国家已将纳他霉素纳入食品添加剂目录, 广泛用于肉类、乳制品防腐(Pipek et al., 2010; Mehyar et al., 2017)和果蔬采后保鲜(Fajardo et al., 2010).在草莓(Fragaria ananassa) (呼玉侠等, 2006)、柑橘(Citrus reticulata) (孙远功等, 2006)和樱桃(Prunus avium) (姜爱丽等, 2009)等水果中添加不同浓度的纳他霉素, 对防止果实采后腐烂、提高贮藏品质有显著效果.研究表明, 纳他霉素可以有效抑制灰葡萄孢(Botrytis cinerea)、扩展青霉(Penicillium expansum)和尖孢炭疽菌(Colletotrichum acutatum)的孢子萌发和菌丝生长, 从而控制采后病害的发生(郭萌萌等, 2013; Haack et al., 2018; He et al., 2019).目前, 纳他霉素对芒果采后胶孢炭疽菌的抑菌效果和作用机理研究较少. ...
Reduction of postharvest anthracnose and enhancement of disease resistance in ripening mango fruit by nitric oxide treatment 1 2014
... 芒果(Mangifera indica)是热带和亚热带地区重要的经济作物, 其口味醇香甘甜, 营养丰富, 深受消费者喜爱(Sivakumar et al., 2011).但它作为一种呼吸跃变型果实, 后熟期间极易受到各种病原菌的侵染, 引起炭疽病和蒂腐病等采后病害, 使芒果在运输和贮藏期间发生组织腐烂, 造成果实品质下降, 常带来严重的经济损失(Jongsri et al., 2017; Perumal et al., 2017).胶孢炭疽菌(Colletotrichum gloeosporioides)是引起芒果炭疽病的主要病原菌, 可潜伏侵染果实多个部位, 常在果实成熟期引发症状(Kefialew and Ayalew, 2008).目前, 我国采后病害的防治主要依赖化学杀菌剂, 如多菌灵、噻苯达唑和苯菌灵, 但化学杀菌剂的大量施用会使病原菌产生耐药性, 减弱杀菌效果(Hu et al., 2014).随着人们对农药残留、食品安全及环境污染等问题的日渐关注, 开发低毒高效的环保型抑菌剂成为防治采后病害的重要途径. ...
Evidence of ROS generation by mitochondria in cells with impaired electron transport chain and mitochondrial DNA damage 1 2007
Influence of chitosan coating combined with spermidine on anthracnose disease and qualities of ‘Nam Dok Mai’ mango after harvest 1 2017
... 芒果(Mangifera indica)是热带和亚热带地区重要的经济作物, 其口味醇香甘甜, 营养丰富, 深受消费者喜爱(Sivakumar et al., 2011).但它作为一种呼吸跃变型果实, 后熟期间极易受到各种病原菌的侵染, 引起炭疽病和蒂腐病等采后病害, 使芒果在运输和贮藏期间发生组织腐烂, 造成果实品质下降, 常带来严重的经济损失(Jongsri et al., 2017; Perumal et al., 2017).胶孢炭疽菌(Colletotrichum gloeosporioides)是引起芒果炭疽病的主要病原菌, 可潜伏侵染果实多个部位, 常在果实成熟期引发症状(Kefialew and Ayalew, 2008).目前, 我国采后病害的防治主要依赖化学杀菌剂, 如多菌灵、噻苯达唑和苯菌灵, 但化学杀菌剂的大量施用会使病原菌产生耐药性, 减弱杀菌效果(Hu et al., 2014).随着人们对农药残留、食品安全及环境污染等问题的日渐关注, 开发低毒高效的环保型抑菌剂成为防治采后病害的重要途径. ...
Postharvest biological control of anthracnose (Colletotrichum gloeosporioides) on mango( Mangifera indica) 1 2008
... 芒果(Mangifera indica)是热带和亚热带地区重要的经济作物, 其口味醇香甘甜, 营养丰富, 深受消费者喜爱(Sivakumar et al., 2011).但它作为一种呼吸跃变型果实, 后熟期间极易受到各种病原菌的侵染, 引起炭疽病和蒂腐病等采后病害, 使芒果在运输和贮藏期间发生组织腐烂, 造成果实品质下降, 常带来严重的经济损失(Jongsri et al., 2017; Perumal et al., 2017).胶孢炭疽菌(Colletotrichum gloeosporioides)是引起芒果炭疽病的主要病原菌, 可潜伏侵染果实多个部位, 常在果实成熟期引发症状(Kefialew and Ayalew, 2008).目前, 我国采后病害的防治主要依赖化学杀菌剂, 如多菌灵、噻苯达唑和苯菌灵, 但化学杀菌剂的大量施用会使病原菌产生耐药性, 减弱杀菌效果(Hu et al., 2014).随着人们对农药残留、食品安全及环境污染等问题的日渐关注, 开发低毒高效的环保型抑菌剂成为防治采后病害的重要途径. ...
Oxidative damage involves in the inhibitory effect of nitric oxide on spore germination of Penicillium expansum 1 2011
... 活性氧是呼吸作用常见的副产物, 但高浓度活性氧能够与脂类、蛋白质和DNA反应, 产生蛋白羰基化、脂质过氧化或mtDNA损伤, 使细胞发生功能障碍, 导致膜破损或细胞死亡(Circu and Aw, 2010; Marchi et al., 2012).已有研究表明, 许多外源性刺激都会使真菌病原体产生诱导活性氧大量生成的应激反应(Xu and Tian, 2008; Lai et al., 2011).本研究中胶孢炭疽菌经纳他霉素处理后, 孢子内活性氧含量显著升高, 说明纳他霉素可以通过诱导活性氧大量产生而对菌体造成伤害. ...
Mitochondria-ros crosstalk in the control of cell death and aging 1 2012
... 活性氧是呼吸作用常见的副产物, 但高浓度活性氧能够与脂类、蛋白质和DNA反应, 产生蛋白羰基化、脂质过氧化或mtDNA损伤, 使细胞发生功能障碍, 导致膜破损或细胞死亡(Circu and Aw, 2010; Marchi et al., 2012).已有研究表明, 许多外源性刺激都会使真菌病原体产生诱导活性氧大量生成的应激反应(Xu and Tian, 2008; Lai et al., 2011).本研究中胶孢炭疽菌经纳他霉素处理后, 孢子内活性氧含量显著升高, 说明纳他霉素可以通过诱导活性氧大量产生而对菌体造成伤害. ...
Effects of chitosan coating containingly- sozyme or natamycin on shelf-life, microbial quality, and sensory properties of Halloumi cheese brined in normal and reduced salt solutions 1 2017
... 纳他霉素是纳他链霉菌(Streptomyces natalensis)发酵过程中产生的天然多烯大环内酯类物质, 对霉菌和酵母有较强的抑制作用, 被公认为安全有效的食品防腐剂之一(Hondrodimou et al., 2011).纳他霉素可与真菌细胞膜上的麦角甾醇或其它甾醇类物质结合, 阻遏麦角甾醇的生物合成, 使细胞膜畸变, 从而引起细胞死亡(Aparicio et al., 2016).纳他霉素具有低毒高效、抑菌谱广、无污染和无抗药性等优点(Arroyo-López et al., 2012), 包括美国在内的许多国家已将纳他霉素纳入食品添加剂目录, 广泛用于肉类、乳制品防腐(Pipek et al., 2010; Mehyar et al., 2017)和果蔬采后保鲜(Fajardo et al., 2010).在草莓(Fragaria ananassa) (呼玉侠等, 2006)、柑橘(Citrus reticulata) (孙远功等, 2006)和樱桃(Prunus avium) (姜爱丽等, 2009)等水果中添加不同浓度的纳他霉素, 对防止果实采后腐烂、提高贮藏品质有显著效果.研究表明, 纳他霉素可以有效抑制灰葡萄孢(Botrytis cinerea)、扩展青霉(Penicillium expansum)和尖孢炭疽菌(Colletotrichum acutatum)的孢子萌发和菌丝生长, 从而控制采后病害的发生(郭萌萌等, 2013; Haack et al., 2018; He et al., 2019).目前, 纳他霉素对芒果采后胶孢炭疽菌的抑菌效果和作用机理研究较少. ...
Effects of essential oil vapour treatment on the postharvest disease control and different defence responses in two mango (Mangifera indica L.) cultivars 1 2018
... 芒果(Mangifera indica)是热带和亚热带地区重要的经济作物, 其口味醇香甘甜, 营养丰富, 深受消费者喜爱(Sivakumar et al., 2011).但它作为一种呼吸跃变型果实, 后熟期间极易受到各种病原菌的侵染, 引起炭疽病和蒂腐病等采后病害, 使芒果在运输和贮藏期间发生组织腐烂, 造成果实品质下降, 常带来严重的经济损失(Jongsri et al., 2017; Perumal et al., 2017).胶孢炭疽菌(Colletotrichum gloeosporioides)是引起芒果炭疽病的主要病原菌, 可潜伏侵染果实多个部位, 常在果实成熟期引发症状(Kefialew and Ayalew, 2008).目前, 我国采后病害的防治主要依赖化学杀菌剂, 如多菌灵、噻苯达唑和苯菌灵, 但化学杀菌剂的大量施用会使病原菌产生耐药性, 减弱杀菌效果(Hu et al., 2014).随着人们对农药残留、食品安全及环境污染等问题的日渐关注, 开发低毒高效的环保型抑菌剂成为防治采后病害的重要途径. ...
Suppression of mould growth on dry sausages 1 2010
... 纳他霉素是纳他链霉菌(Streptomyces natalensis)发酵过程中产生的天然多烯大环内酯类物质, 对霉菌和酵母有较强的抑制作用, 被公认为安全有效的食品防腐剂之一(Hondrodimou et al., 2011).纳他霉素可与真菌细胞膜上的麦角甾醇或其它甾醇类物质结合, 阻遏麦角甾醇的生物合成, 使细胞膜畸变, 从而引起细胞死亡(Aparicio et al., 2016).纳他霉素具有低毒高效、抑菌谱广、无污染和无抗药性等优点(Arroyo-López et al., 2012), 包括美国在内的许多国家已将纳他霉素纳入食品添加剂目录, 广泛用于肉类、乳制品防腐(Pipek et al., 2010; Mehyar et al., 2017)和果蔬采后保鲜(Fajardo et al., 2010).在草莓(Fragaria ananassa) (呼玉侠等, 2006)、柑橘(Citrus reticulata) (孙远功等, 2006)和樱桃(Prunus avium) (姜爱丽等, 2009)等水果中添加不同浓度的纳他霉素, 对防止果实采后腐烂、提高贮藏品质有显著效果.研究表明, 纳他霉素可以有效抑制灰葡萄孢(Botrytis cinerea)、扩展青霉(Penicillium expansum)和尖孢炭疽菌(Colletotrichum acutatum)的孢子萌发和菌丝生长, 从而控制采后病害的发生(郭萌萌等, 2013; Haack et al., 2018; He et al., 2019).目前, 纳他霉素对芒果采后胶孢炭疽菌的抑菌效果和作用机理研究较少. ...
Mechanism of antifungal action of borate against Colletotrichum gloeosporioides related to mitochondrial degradation in spores 1 2012
Maintaining mango (Mangifera indica L.) fruit quality during the export chain 1 2011
... 芒果(Mangifera indica)是热带和亚热带地区重要的经济作物, 其口味醇香甘甜, 营养丰富, 深受消费者喜爱(Sivakumar et al., 2011).但它作为一种呼吸跃变型果实, 后熟期间极易受到各种病原菌的侵染, 引起炭疽病和蒂腐病等采后病害, 使芒果在运输和贮藏期间发生组织腐烂, 造成果实品质下降, 常带来严重的经济损失(Jongsri et al., 2017; Perumal et al., 2017).胶孢炭疽菌(Colletotrichum gloeosporioides)是引起芒果炭疽病的主要病原菌, 可潜伏侵染果实多个部位, 常在果实成熟期引发症状(Kefialew and Ayalew, 2008).目前, 我国采后病害的防治主要依赖化学杀菌剂, 如多菌灵、噻苯达唑和苯菌灵, 但化学杀菌剂的大量施用会使病原菌产生耐药性, 减弱杀菌效果(Hu et al., 2014).随着人们对农药残留、食品安全及环境污染等问题的日渐关注, 开发低毒高效的环保型抑菌剂成为防治采后病害的重要途径. ...
Natamycin inhibits vacuole fusion at the priming phase via a specific interaction with ergosterol 1 2010
... 纳他霉素的抑菌机理主要是其可与真菌和酵母细胞膜上的麦角甾醇结合, 改变细胞膜透性.此外, 纳他霉素可以干扰酵母液泡融合和膜融合前麦角甾醇依赖性启动反应, 影响营养物质运输, 对麦角甾醇依赖性相关蛋白产生一定影响(te Welscher et al., 2010).为探究纳他霉素对胶孢炭疽菌的抑制机理, 我们测定了纳他霉素处理后菌丝细胞膜相对渗透率, 结果表明纳他霉素可以增加细胞膜透性, 使胞内物质渗出而导致细胞死亡; 同时发现纳他霉素处理后菌丝可溶性蛋白含量增加.黄勤知等(2013)用抗菌蛋白处理芒果炭疽菌以及唐群勇(2011)用抗菌肽处理桃(Amygdalus persica)软腐病菌后, 均发现菌丝内可溶性蛋白含量增加, 这可能是菌体受到化学胁迫后的一种调节反应, 或是药物处理使细胞功能紊乱, 抑制了蛋白质的分解代谢.PI染色结果表明细胞膜是纳他霉素的作用靶点, 其可引起真菌细胞膜功能紊乱, 破坏细胞膜结构, 导致细胞死亡(Aparicio et al., 2004; Van Leeuwen et al., 2009). ...
The polyene antimycotics nystatin and filipin disrupt the plasma membrane, whereas natamycin inhibits endocytosis in germinating conidia of Penicillium discolor 1 2009
... 纳他霉素的抑菌机理主要是其可与真菌和酵母细胞膜上的麦角甾醇结合, 改变细胞膜透性.此外, 纳他霉素可以干扰酵母液泡融合和膜融合前麦角甾醇依赖性启动反应, 影响营养物质运输, 对麦角甾醇依赖性相关蛋白产生一定影响(te Welscher et al., 2010).为探究纳他霉素对胶孢炭疽菌的抑制机理, 我们测定了纳他霉素处理后菌丝细胞膜相对渗透率, 结果表明纳他霉素可以增加细胞膜透性, 使胞内物质渗出而导致细胞死亡; 同时发现纳他霉素处理后菌丝可溶性蛋白含量增加.黄勤知等(2013)用抗菌蛋白处理芒果炭疽菌以及唐群勇(2011)用抗菌肽处理桃(Amygdalus persica)软腐病菌后, 均发现菌丝内可溶性蛋白含量增加, 这可能是菌体受到化学胁迫后的一种调节反应, 或是药物处理使细胞功能紊乱, 抑制了蛋白质的分解代谢.PI染色结果表明细胞膜是纳他霉素的作用靶点, 其可引起真菌细胞膜功能紊乱, 破坏细胞膜结构, 导致细胞死亡(Aparicio et al., 2004; Van Leeuwen et al., 2009). ...
Salicylic acid alleviated pathogen-induced oxidative stress in harvested sweet cherry fruit 1 2008
... 活性氧是呼吸作用常见的副产物, 但高浓度活性氧能够与脂类、蛋白质和DNA反应, 产生蛋白羰基化、脂质过氧化或mtDNA损伤, 使细胞发生功能障碍, 导致膜破损或细胞死亡(Circu and Aw, 2010; Marchi et al., 2012).已有研究表明, 许多外源性刺激都会使真菌病原体产生诱导活性氧大量生成的应激反应(Xu and Tian, 2008; Lai et al., 2011).本研究中胶孢炭疽菌经纳他霉素处理后, 孢子内活性氧含量显著升高, 说明纳他霉素可以通过诱导活性氧大量产生而对菌体造成伤害. ...
 ,3,*, 史学群
,3,*, 史学群
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT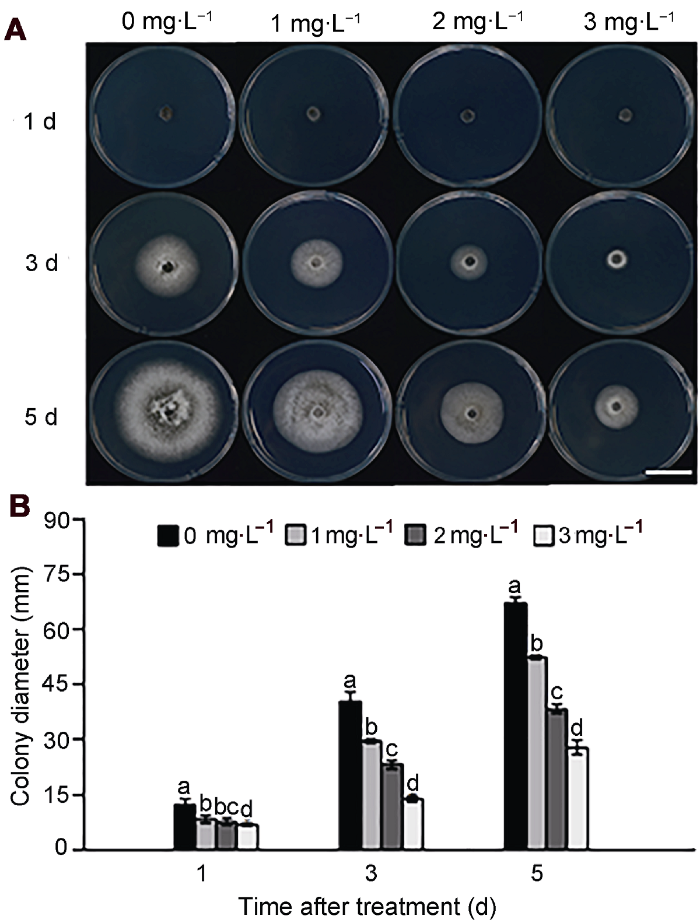 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT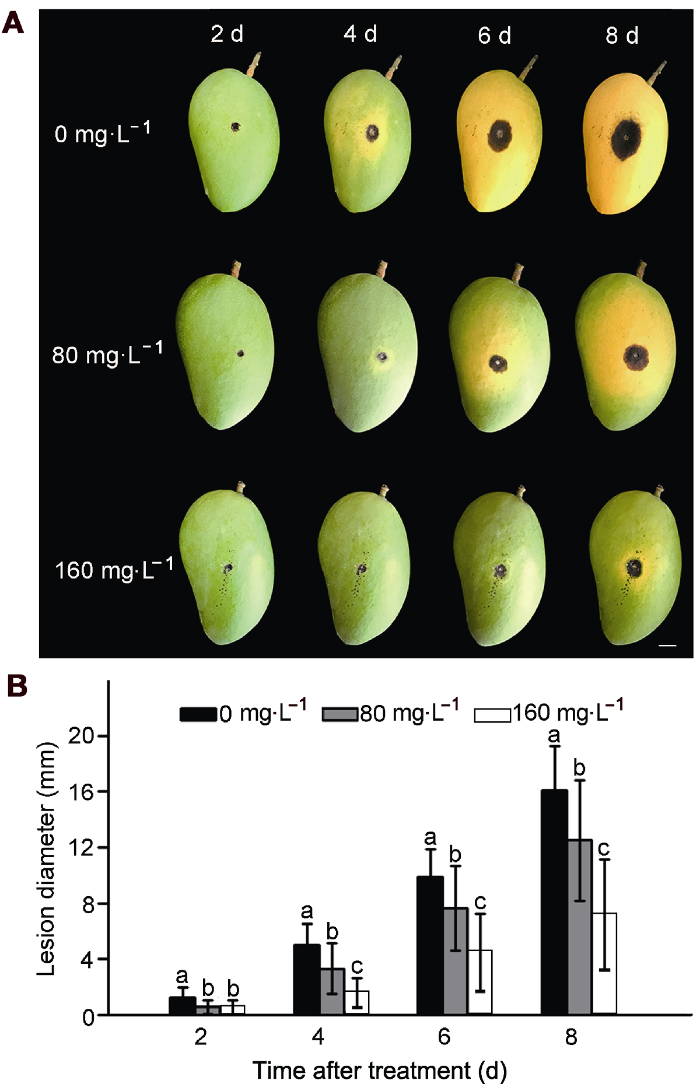 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT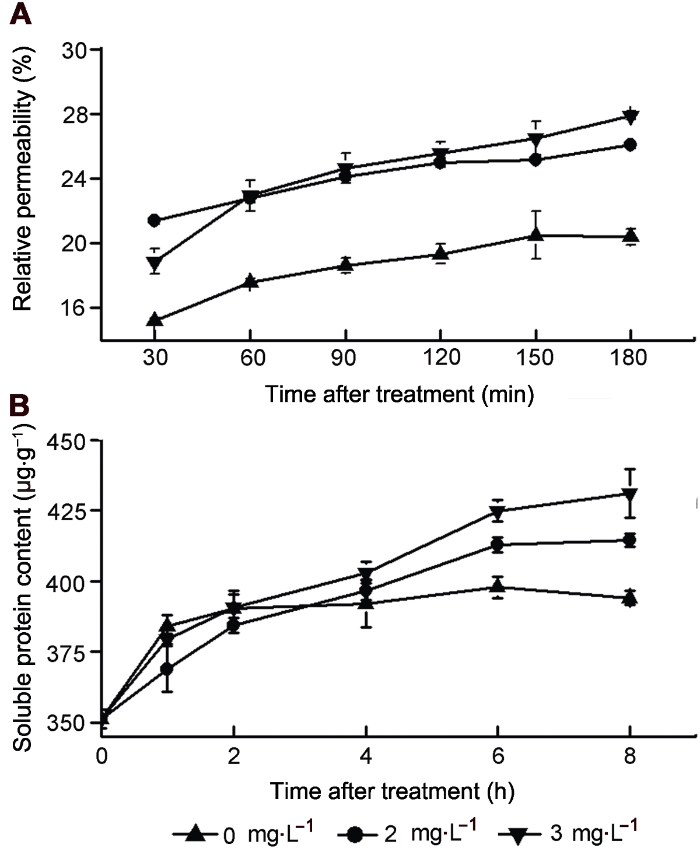 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT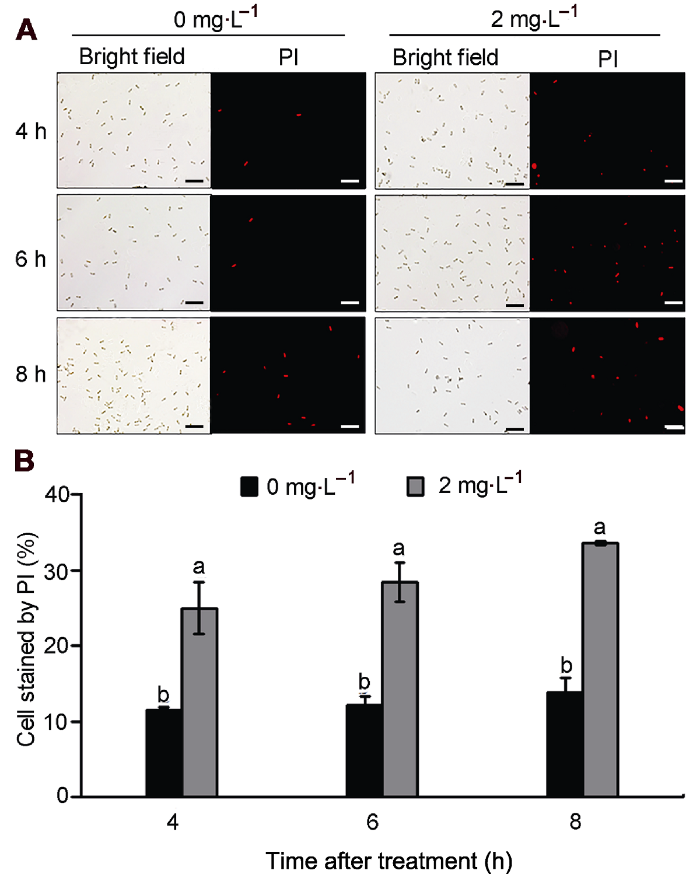 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT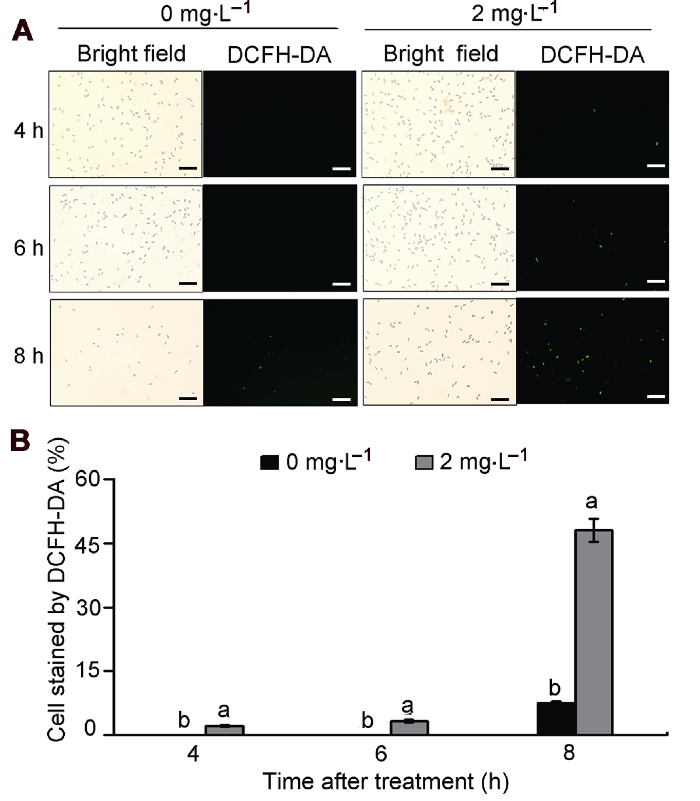 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT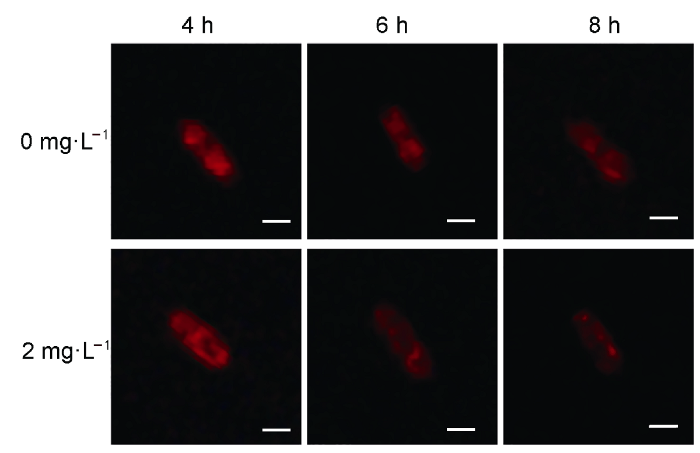 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}