Functional Analysis of Brassinosteroids in Salt Stress Responses in Rice
Lulu Li, Wenchao Yin, Mei Niu, Wenjing Meng, Xiaoxing Zhang, Hongning Tong,*Institute of Crop Sciences, Chinese Academy of Agricultural Sciences/National Key Facility for Crop Gene Resources and Genetic Improvement, Beijing 100081, China
Abstract Brassinosteroids (BRs) are a class of steroid phytohormones that play diverse roles in plant growth and development and stress responses. Rapid progresses have been made in how BRs regulate plant growth and development in recent years. However, the roles of BRs in stress response in Oryza sativa remain unclear. Here, we investigated the relation between salinity stress and BR synthesis in rice. Both salt stress and abscisic acid, the well-known stress hormone, strongly inhibited the expression of two BR-synthetic genes, D2 and D11. In addition, both d2-2, the BR synthetic mutant, and d61-1, the BR receptor mutant, showed impaired tolerance to salt stress. Moreover, by using transgenic plants overexpressing OsBZR1, the key BR signaling transcriptional factor, we found that BRs strongly induced dephosphorylation of OsBZR1, but high concentrations of salt suppressed OsBZR1 protein accumulation as well as its dephosphorylation. Furthermore, transcriptome analyses revealed that 38.4% of BR-regulated genes were also regulated by high concentrations of salt, and importantly, 91.5% of the co-regulated genes are consistently up- or downregulated by both BR and salt. Gene Ontology analyses revealed that these overlapping genes were highly enriched in the biological process “response to stimulus”. Taken together, our results suggest that BRs contribute to salt stress tolerance, and salt stress suppresses BR synthesis to restrict rice growth. Keywords:brassinosteroid;rice;salt stress;abscisic acid;OsBZR1
PDF (526KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 栗露露, 殷文超, 牛梅, 孟文静, 张晓星, 童红宁. 油菜素甾醇调控水稻盐胁迫应答的作用研究. 植物学报, 2019, 54(2): 185-193 doi:10.11983/CBB19013 Li Lulu, Yin Wenchao, Niu Mei, Meng Wenjing, Zhang Xiaoxing, Tong Hongning. Functional Analysis of Brassinosteroids in Salt Stress Responses in Rice. Chinese Bulletin of Botany, 2019, 54(2): 185-193 doi:10.11983/CBB19013
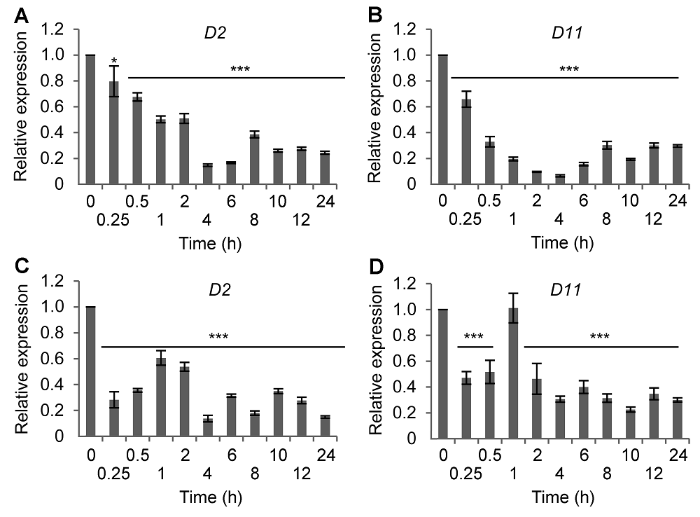
(A) 盐处理后D2基因的表达; (B) 盐处理后D11基因的表达; (C) ABA处理后D2基因的表达; (D) ABA处理后D11基因的表达。* P< 0.05; *** P<0.001 Figure 1Time-course expression of BR synthetic genes in rice following salt or ABA treatment
(A) D2 expression after NaCl treatment; (B) D11 expression after NaCl treatment; (C) D2 expression after ABA treatment; (D) D11 expression after ABA treatment. * P<0.05; *** P<0.001
(A) 盐处理后d2-2突变体及其野生型的生长情况; (B) 盐处理后d2-2突变体及其野生型的存活率统计; (C) 盐处理后d61-1突变体及其野生型的生长情况; (D) 盐处理后d61-1突变体及其野生型的存活率统计。** P<0.01 Figure 2Survival rate of rice BR defective mutants and the wild type under salt stress
(A) Growth status of d2-2 mutant and the wild type after salt treatment; (B) Statistic data of the survival rate of d2-2 and the wild type after salt treatment; (C) Growth status of d61-1 mutant and the wild type after salt treatment; (D) Statistic data of the survival rate of d61-1 and the wild type after salt treatment. ** P<0.01
(A) BR和ABA与盐差异调控基因的共调控分析; (B) BR上调(BR-UP)和下调(BR-DN)以及NaCl上调(NaCl-UP)和下调(NaCl-DN)基因的共调控分析。图中显示差异基因的分布及数目。 Figure 4Co-regulation analyses of BR-, ABA- and NaCl-regulated genes of rice
(A) Co-regulated gene numbers between BR-, ABA- and NaCl-regulated different expression genes (DEGs); (B) Co-regulation analyses among BR-upregulated (BR-UP), BR-downregulated (BR-DN), NaCl-upregulated (NaCl-UP) and NaCl-downregulated (NaCl-DN) genes. Distribution of the gene numbers was indicated.
HeJX, GendronJM, SunY, GampalaSSL, GendronN, SunCQ, WangZY ( 2005). BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses Science 307, 1634-1638. DOI:10.1126/science.1107580URL [本文引用: 1]
HeJX, GendronJM, YangYL, LiJM, WangZY ( 2002). The GSK3-like kinase BIN2 phosphorylates and destabilizes BZR1, a positive regulator of the brassinosteroid signaling pathway in Arabidopsis Proc Natl Acad Sci USA 99, 10185-10190. DOI:10.1073/pnas.152342599URL [本文引用: 1]
HongZ, Ueguchi-TanakaM, UmemuraK, UozuS, FujiokaS, TakatsutoS, YoshidaS, AshikariM, KitanoH, MatsuokaM ( 2003). A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P450 Plant Cell 15, 2900-2910. [本文引用: 1]
KhripachV, ZhabinskiiV, De GrootA ( 2000). Twenty years of brassinosteroids: steroidal plant hormones warrant better crops for the XXI century Ann Bot 86, 441-447. DOI:10.1006/anbo.2000.1227URL [本文引用: 2]
KrishnaP, PrasadBD, RahmanT ( 2017). Brassinosteroid action in plant abiotic stress tolerance. In: Russinova E, Caño-Delgado AI, eds. Brassinosteroids. New York: Hum- ana Press. pp. 193-202. [本文引用: 1]
MorinakaY, SakamotoT, InukaiY, AgetsumaM, KitanoH, AshikariM, MatsuokaM ( 2006). Morphological alteration caused by brassinosteroid insensitivity increases the biomass and grain production of rice Plant Physiol 141, 924-931. DOI:10.1104/pp.106.077081URL [本文引用: 1]
NakashimaK, Yamaguchi-ShinozakiK ( 2013). ABA signaling in stress-response and seed development Plant Cell Rep 32, 959-970. DOI:10.1007/s00299-013-1418-1URL [本文引用: 1]
NolanTM, BrennanB, YangMR, ChenJN, ZhangMC, LiZH, WangXL, BasshamDC, WalleyJ, YinYH ( 2017). Selective autophagy of BES1 mediated by DSK2 balances plant growth and survival Dev Cell 41, 33-46.e7. [本文引用: 1]
SunY, FanXY, CaoDM, TangWQ, HeK, ZhuJY, HeJX, BaiMY, ZhuSW, OhE, PatilS, KimTW, JiHK, WongWH, RheeSY, WangZY ( 2010). Integration of bras- sinosteroid signal transduction with the transcription network for plant growth regulation in Arabidopsis Dev Cell 19, 765-777. DOI:10.1016/j.devcel.2010.10.010URL [本文引用: 1]
TongHN, ChuCC ( 2016). Reply: brassinosteroid regulates gibberellin synthesis to promote cell elongation in rice: critical comments on ross and quittenden's letter Plant Cell 28, 833-835. [本文引用: 1]
TongHN, ChuCC ( 2018). Functional specificities of bras- sinosteroid and potential utilization for crop improvement Trends Plant Sci 23, 1016-1028. [本文引用: 4]
TongHN, LiuLC, JinY, DuL, YinYH, QianQ, ZhuLH, ChuCC ( 2012). DWARF AND LOW-TILLERING acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses in rice Plant Cell 24, 2562-2577. DOI:10.1105/tpc.112.097394URL [本文引用: 1]
YamamuroC, IharaY, WuX, NoguchiT, FujiokaS, TakatsutoS, AshikariM, KitanoH, MatsuokaM ( 2000). Loss of function of a rice brassinosteroid insensitive 1 homolog prevents internode elongation and bending of the lamina joint Plant Cell 12, 1591-1606. [本文引用: 1]
YangMR, LiCX, CaiZY, HuYM, NolanT, YuFF, YinYH, XieQ, TangGL, WangXL ( 2017). SINAT E3 ligases control the light-mediated stability of the brassinosteroid- activated transcription factor BES1 in Arabidopsis Dev Cell 41, 47-58.e4. [本文引用: 1]
YinWC, DongNN, NiuM, ZhangXX, LiLL, LiuJ, LiuB, TongHN ( 2018). Brassinosteroid-regulated plant growth and development and gene expression in soybean Crop J. DOI: 10.1016/j.cj.2018.10.003. [本文引用: 1]
YinYH, WangZY, Mora-GarciaS, LiJM, YoshidaS, AsamiT, ChoryJ ( 2002). BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation Cell 109, 181-191. DOI:10.1016/S0092-8674(02)00721-3URL [本文引用: 1]
YuXF, LiL, ZolaJ, AluruM, YeHX, FoudreeA, GuoHQ, AndersonS, AluruS, LiuP, RodermelS, YinYH ( 2011). A brassinosteroid transcriptional network revealed by genome-wide identification of BESI target genes in Ara- bidopsis thaliana Plant J 65, 634-646. [本文引用: 1]
ZhaoX, DouLR, GongZZ, WangXF, MaoTL ( 2019). BES1 hinders ABSCISIC ACID INSENSITIVE5 and promotes seed germination in Arabidopsis New Phytol 221, 908-918. DOI:10.1111/nph.15437URL [本文引用: 1]
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
Brassinosteroid biosynthesis and inactiva- tion 1 2006
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
Brassinosteroid: a biotechno- logical target for enhancing crop yield and stress tole- rance 1 2009
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
Down-regulation of BdBRI1, a putative brassinosteroid receptor gene produces a dwarf phenotype with enhanced drought tolerance in Brachy- podium distachyon 1 2015
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen 1 1979
... BR作为较晚发现的一类植物激素(Grove et al., 1979), 广泛参与调控植物的生长发育以及耐逆性, 在农业生产及农业生物技术上具有重要的应用价值.一方面BR同源类似物可作为植物生长调节剂直接应用于田间生产(Khripach et al., 2000); 另一方面可利用BR的生物功能通过农业生物技术途径对作物进行改良(Tong and Chu, 2018).例如, 科学家通过降低BR合成(或信号)基因的表达, 对水稻叶夹角进行调节使其更加直立, 从而使得植物适合密植, 增加产量(Morinaka et al., 2006; Sakamoto et al., 2006).再者也可通过增加BR功能促进水稻灌浆来增加产量(Wu et al., 2008).然而, 目前尚缺乏水稻中BR对抗逆作用的研究, 这些策略是否可引起水稻耐逆性的改变尚不清楚.因此, 开展水稻BR耐逆性研究对其最终应用十分重要. ...
Brassinosteroids modu- late ABA-induced stomatal closure in Arabidopsis 1 2016
BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses 1 2005
... 本研究表明, BR缺陷突变体对高盐处理超敏感, 说明BR参与水稻的耐盐反应并具有正调控作用.转录组分析表明, BR调控的基因很大比例同时受到盐胁迫的调控, 并且绝大部分受到两者一致调控.由于盐胁迫可显著抑制BR合成基因的表达, 这些结果暗示高盐处理对水稻部分基因的调控可能通过抑制BR合成来完成.此外, 我们发现, OsBZR1去磷酸化形式的蛋白水平受到盐胁迫的明显抑制.OsBZR1为BR信号转导过程的核心转录因子, 对拟南芥的研究表明这类转录因子可直接结合下游基因的启动子, 进而调控大量BR下游基因的表达(Sun et al., 2010; Yu et al., 2011).另外, 作为反馈调控机制, BZR1类转录因子可直接结合BR合成基因, 抑制其表达(Wang et al., 2002; He et al., 2005).本研究创制了OsBZR1融合双标签的转基因株系, 并通过BR处理证实这一材料非常适用于对OsBZR1进行蛋白水平的研究.我们也发现OsBZR1在盐处理2-4小时显著积累, 而此时D2和D11基因表达也达到最低水平, 暗示在盐胁迫过程中植物可对内源激素平衡进行精细调控. ...
The GSK3-like kinase BIN2 phosphorylates and destabilizes BZR1, a positive regulator of the brassinosteroid signaling pathway in Arabidopsis 1 2002
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
Bras- sinosteroids and rice architecture 1 2004
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P450 1 2003
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
Twenty years of brassinosteroids: steroidal plant hormones warrant better crops for the XXI century 2 2000
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
... BR作为较晚发现的一类植物激素(Grove et al., 1979), 广泛参与调控植物的生长发育以及耐逆性, 在农业生产及农业生物技术上具有重要的应用价值.一方面BR同源类似物可作为植物生长调节剂直接应用于田间生产(Khripach et al., 2000); 另一方面可利用BR的生物功能通过农业生物技术途径对作物进行改良(Tong and Chu, 2018).例如, 科学家通过降低BR合成(或信号)基因的表达, 对水稻叶夹角进行调节使其更加直立, 从而使得植物适合密植, 增加产量(Morinaka et al., 2006; Sakamoto et al., 2006).再者也可通过增加BR功能促进水稻灌浆来增加产量(Wu et al., 2008).然而, 目前尚缺乏水稻中BR对抗逆作用的研究, 这些策略是否可引起水稻耐逆性的改变尚不清楚.因此, 开展水稻BR耐逆性研究对其最终应用十分重要. ...
Brassinosteroid signal transduc- tion from receptor kinases to transcription factors 2 2010
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
... ).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
1 2017
... 越来越多的研究表明, BR在不同物种、器官、组织、浓度以及环境下的功能可能完全不同(Singh and Savaldi-Goldstein, 2015; Tong and Chu, 2018).例如, 在黑暗条件下BR促进水稻生长, 但却抑制大豆(Glycine max)生长(Yin et al., 2018); 低浓度BR促进植物不同组织生长, 而高浓度BR显著抑制水稻株高以及根的生长, 但却促进胚芽鞘生长及叶夹角增大(Tong and Chu, 2016).此外, BR还可调节植物的耐逆反应, 且针对不同逆境BR的调控功能不尽相同.例如, 在拟南芥中, BR缺陷突变体对盐更加敏感, 但却更为耐旱(Krishna et al., 2017).这些结果说明, 需根据不同物种和不同环境具体分析BR的作用. ...
Morphological alteration caused by brassinosteroid insensitivity increases the biomass and grain production of rice 1 2006
... BR作为较晚发现的一类植物激素(Grove et al., 1979), 广泛参与调控植物的生长发育以及耐逆性, 在农业生产及农业生物技术上具有重要的应用价值.一方面BR同源类似物可作为植物生长调节剂直接应用于田间生产(Khripach et al., 2000); 另一方面可利用BR的生物功能通过农业生物技术途径对作物进行改良(Tong and Chu, 2018).例如, 科学家通过降低BR合成(或信号)基因的表达, 对水稻叶夹角进行调节使其更加直立, 从而使得植物适合密植, 增加产量(Morinaka et al., 2006; Sakamoto et al., 2006).再者也可通过增加BR功能促进水稻灌浆来增加产量(Wu et al., 2008).然而, 目前尚缺乏水稻中BR对抗逆作用的研究, 这些策略是否可引起水稻耐逆性的改变尚不清楚.因此, 开展水稻BR耐逆性研究对其最终应用十分重要. ...
ABA signaling in stress-response and seed development 1 2013
Selective autophagy of BES1 mediated by DSK2 balances plant growth and survival 1 2017
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
Hormone balance and abiotic stress tolerance in crop plants 1 2011
Nucleocytoplasmic shuttling of BZR1 mediated by phos- phorylation is essential in Arabidopsis brassinosteroid signaling 1 2007
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
Erect leaves caused by brassinosteroid deficiency inc- rease biomass production and grain yield in rice 1 2006
... BR作为较晚发现的一类植物激素(Grove et al., 1979), 广泛参与调控植物的生长发育以及耐逆性, 在农业生产及农业生物技术上具有重要的应用价值.一方面BR同源类似物可作为植物生长调节剂直接应用于田间生产(Khripach et al., 2000); 另一方面可利用BR的生物功能通过农业生物技术途径对作物进行改良(Tong and Chu, 2018).例如, 科学家通过降低BR合成(或信号)基因的表达, 对水稻叶夹角进行调节使其更加直立, 从而使得植物适合密植, 增加产量(Morinaka et al., 2006; Sakamoto et al., 2006).再者也可通过增加BR功能促进水稻灌浆来增加产量(Wu et al., 2008).然而, 目前尚缺乏水稻中BR对抗逆作用的研究, 这些策略是否可引起水稻耐逆性的改变尚不清楚.因此, 开展水稻BR耐逆性研究对其最终应用十分重要. ...
... 越来越多的研究表明, BR在不同物种、器官、组织、浓度以及环境下的功能可能完全不同(Singh and Savaldi-Goldstein, 2015; Tong and Chu, 2018).例如, 在黑暗条件下BR促进水稻生长, 但却抑制大豆(Glycine max)生长(Yin et al., 2018); 低浓度BR促进植物不同组织生长, 而高浓度BR显著抑制水稻株高以及根的生长, 但却促进胚芽鞘生长及叶夹角增大(Tong and Chu, 2016).此外, BR还可调节植物的耐逆反应, 且针对不同逆境BR的调控功能不尽相同.例如, 在拟南芥中, BR缺陷突变体对盐更加敏感, 但却更为耐旱(Krishna et al., 2017).这些结果说明, 需根据不同物种和不同环境具体分析BR的作用. ...
Integration of bras- sinosteroid signal transduction with the transcription network for plant growth regulation in Arabidopsis 1 2010
... 本研究表明, BR缺陷突变体对高盐处理超敏感, 说明BR参与水稻的耐盐反应并具有正调控作用.转录组分析表明, BR调控的基因很大比例同时受到盐胁迫的调控, 并且绝大部分受到两者一致调控.由于盐胁迫可显著抑制BR合成基因的表达, 这些结果暗示高盐处理对水稻部分基因的调控可能通过抑制BR合成来完成.此外, 我们发现, OsBZR1去磷酸化形式的蛋白水平受到盐胁迫的明显抑制.OsBZR1为BR信号转导过程的核心转录因子, 对拟南芥的研究表明这类转录因子可直接结合下游基因的启动子, 进而调控大量BR下游基因的表达(Sun et al., 2010; Yu et al., 2011).另外, 作为反馈调控机制, BZR1类转录因子可直接结合BR合成基因, 抑制其表达(Wang et al., 2002; He et al., 2005).本研究创制了OsBZR1融合双标签的转基因株系, 并通过BR处理证实这一材料非常适用于对OsBZR1进行蛋白水平的研究.我们也发现OsBZR1在盐处理2-4小时显著积累, 而此时D2和D11基因表达也达到最低水平, 暗示在盐胁迫过程中植物可对内源激素平衡进行精细调控. ...
Reply: brassinosteroid regulates gibberellin synthesis to promote cell elongation in rice: critical comments on ross and quittenden's letter 1 2016
... 越来越多的研究表明, BR在不同物种、器官、组织、浓度以及环境下的功能可能完全不同(Singh and Savaldi-Goldstein, 2015; Tong and Chu, 2018).例如, 在黑暗条件下BR促进水稻生长, 但却抑制大豆(Glycine max)生长(Yin et al., 2018); 低浓度BR促进植物不同组织生长, 而高浓度BR显著抑制水稻株高以及根的生长, 但却促进胚芽鞘生长及叶夹角增大(Tong and Chu, 2016).此外, BR还可调节植物的耐逆反应, 且针对不同逆境BR的调控功能不尽相同.例如, 在拟南芥中, BR缺陷突变体对盐更加敏感, 但却更为耐旱(Krishna et al., 2017).这些结果说明, 需根据不同物种和不同环境具体分析BR的作用. ...
Functional specificities of bras- sinosteroid and potential utilization for crop improvement 4 2018
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
... ; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
... BR作为较晚发现的一类植物激素(Grove et al., 1979), 广泛参与调控植物的生长发育以及耐逆性, 在农业生产及农业生物技术上具有重要的应用价值.一方面BR同源类似物可作为植物生长调节剂直接应用于田间生产(Khripach et al., 2000); 另一方面可利用BR的生物功能通过农业生物技术途径对作物进行改良(Tong and Chu, 2018).例如, 科学家通过降低BR合成(或信号)基因的表达, 对水稻叶夹角进行调节使其更加直立, 从而使得植物适合密植, 增加产量(Morinaka et al., 2006; Sakamoto et al., 2006).再者也可通过增加BR功能促进水稻灌浆来增加产量(Wu et al., 2008).然而, 目前尚缺乏水稻中BR对抗逆作用的研究, 这些策略是否可引起水稻耐逆性的改变尚不清楚.因此, 开展水稻BR耐逆性研究对其最终应用十分重要. ...
... 越来越多的研究表明, BR在不同物种、器官、组织、浓度以及环境下的功能可能完全不同(Singh and Savaldi-Goldstein, 2015; Tong and Chu, 2018).例如, 在黑暗条件下BR促进水稻生长, 但却抑制大豆(Glycine max)生长(Yin et al., 2018); 低浓度BR促进植物不同组织生长, 而高浓度BR显著抑制水稻株高以及根的生长, 但却促进胚芽鞘生长及叶夹角增大(Tong and Chu, 2016).此外, BR还可调节植物的耐逆反应, 且针对不同逆境BR的调控功能不尽相同.例如, 在拟南芥中, BR缺陷突变体对盐更加敏感, 但却更为耐旱(Krishna et al., 2017).这些结果说明, 需根据不同物种和不同环境具体分析BR的作用. ...
DWARF AND LOW-TILLERING acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses in rice 1 2012
Brassinosteroid regulates cell elongation by modulating gibberellin metabolism in rice 1 2014
... 植物材料包括粳稻中花11 (ZH11)、日本晴(NIP)、Shiokari (SHI)、Taichung65 (TC65)、突变体d2-2 (SHI背景)、d61-1 (TC65背景)和OsBZR1过表达材料GM-OsBZR1 (融合GFP和MYC双重标签, ZH11背景).其中, 野生型、d2-2和d61-1均为本实验室保存(Tong et al., 2014).GM-OsBZR1为本实验室创制.选取37°C浸水2天, 且萌发一致的水稻(Oryza sativa L.)种子, 单粒胚向上置于去除底部的96孔PCR板中, 使用1/2 MS (Murashige and Skoog medium, Duch- efa)液体培养基, 于光照培养箱(宁波江南)中培养.白日光照14小时, 温度30°C; 夜晚10小时, 温度28°C. ...
Nuclear-localized BZR1 mediates bras- sinosteroid-induced growth and feedback suppression of brassinosteroid biosynthesis 1 2002
... 本研究表明, BR缺陷突变体对高盐处理超敏感, 说明BR参与水稻的耐盐反应并具有正调控作用.转录组分析表明, BR调控的基因很大比例同时受到盐胁迫的调控, 并且绝大部分受到两者一致调控.由于盐胁迫可显著抑制BR合成基因的表达, 这些结果暗示高盐处理对水稻部分基因的调控可能通过抑制BR合成来完成.此外, 我们发现, OsBZR1去磷酸化形式的蛋白水平受到盐胁迫的明显抑制.OsBZR1为BR信号转导过程的核心转录因子, 对拟南芥的研究表明这类转录因子可直接结合下游基因的启动子, 进而调控大量BR下游基因的表达(Sun et al., 2010; Yu et al., 2011).另外, 作为反馈调控机制, BZR1类转录因子可直接结合BR合成基因, 抑制其表达(Wang et al., 2002; He et al., 2005).本研究创制了OsBZR1融合双标签的转基因株系, 并通过BR处理证实这一材料非常适用于对OsBZR1进行蛋白水平的研究.我们也发现OsBZR1在盐处理2-4小时显著积累, 而此时D2和D11基因表达也达到最低水平, 暗示在盐胁迫过程中植物可对内源激素平衡进行精细调控. ...
Brassinosteroids regulate grain filling in rice 1 2008
... BR作为较晚发现的一类植物激素(Grove et al., 1979), 广泛参与调控植物的生长发育以及耐逆性, 在农业生产及农业生物技术上具有重要的应用价值.一方面BR同源类似物可作为植物生长调节剂直接应用于田间生产(Khripach et al., 2000); 另一方面可利用BR的生物功能通过农业生物技术途径对作物进行改良(Tong and Chu, 2018).例如, 科学家通过降低BR合成(或信号)基因的表达, 对水稻叶夹角进行调节使其更加直立, 从而使得植物适合密植, 增加产量(Morinaka et al., 2006; Sakamoto et al., 2006).再者也可通过增加BR功能促进水稻灌浆来增加产量(Wu et al., 2008).然而, 目前尚缺乏水稻中BR对抗逆作用的研究, 这些策略是否可引起水稻耐逆性的改变尚不清楚.因此, 开展水稻BR耐逆性研究对其最终应用十分重要. ...
Loss of function of a rice brassinosteroid insensitive 1 homolog prevents internode elongation and bending of the lamina joint 1 2000
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
SINAT E3 ligases control the light-mediated stability of the brassinosteroid- activated transcription factor BES1 in Arabidopsis 1 2017
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
Brassinosteroid-regulated plant growth and development and gene expression in soybean 1 2018
... 越来越多的研究表明, BR在不同物种、器官、组织、浓度以及环境下的功能可能完全不同(Singh and Savaldi-Goldstein, 2015; Tong and Chu, 2018).例如, 在黑暗条件下BR促进水稻生长, 但却抑制大豆(Glycine max)生长(Yin et al., 2018); 低浓度BR促进植物不同组织生长, 而高浓度BR显著抑制水稻株高以及根的生长, 但却促进胚芽鞘生长及叶夹角增大(Tong and Chu, 2016).此外, BR还可调节植物的耐逆反应, 且针对不同逆境BR的调控功能不尽相同.例如, 在拟南芥中, BR缺陷突变体对盐更加敏感, 但却更为耐旱(Krishna et al., 2017).这些结果说明, 需根据不同物种和不同环境具体分析BR的作用. ...
BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation 1 2002
... 油菜素甾醇(brassinosteroids, BRs)调控着植物生长发育的各个方面, 包括植物的耐逆性.例如, 在二穗短柄草(Brachypodium distachyum)中, 下调BR受体基因BRI1表达可提高其抗旱性(Feng et al., 2015), 而在杨树中过表达BR合成基因可提高杨树的抗旱性(王沛雅等, 2014).因此, BR被认为在提高植物产量及抗性等方面具有重要的应用潜力(Khripach et al., 2000; Divi and Krishna, 2009).模式植物拟南芥(Arabidopsis thaliana)中BR合成及信号转导途径的研究已相对清楚(Choe, 2006; Kim and Wang, 2010; Tong and Chu, 2018).BR由膜受体BRI1识别后, 通过一系列磷酸化和去磷酸化事件, 将信号传递给BZR1/BES1转录因子, 进而调控下游基因的表达(Kim and Wang, 2010).拟南芥中, BZR1或BES1作为BR信号下游的转录因子, 在蛋白水平受到磷酸化、核质穿梭和蛋白降解等多方面调控.因此, 其通常作为BR信号输出的指示蛋白(He et al., 2002; Yin et al., 2002; Ryu et al., 2007; Nolan et al., 2017; Yang et al., 2017).在水稻中, BR调控着诸多重要农艺性状, 包括株高、叶夹角和籽粒大小等(Hong et al., 2004; Tong and Chu, 2018).BR合成突变体(如d2)以及信号突变体(d61等)通常表现出植株矮化、叶片直立和籽粒变小等表型(Yamamuro et al., 2000; Hong et al., 2003).目前, 在水稻中关于BR的研究主要集中在对株型和粒型的调控上, 而对于其在逆境胁迫过程中的作用报道尚少. ...
A brassinosteroid transcriptional network revealed by genome-wide identification of BESI target genes in Ara- bidopsis thaliana 1 2011
... 本研究表明, BR缺陷突变体对高盐处理超敏感, 说明BR参与水稻的耐盐反应并具有正调控作用.转录组分析表明, BR调控的基因很大比例同时受到盐胁迫的调控, 并且绝大部分受到两者一致调控.由于盐胁迫可显著抑制BR合成基因的表达, 这些结果暗示高盐处理对水稻部分基因的调控可能通过抑制BR合成来完成.此外, 我们发现, OsBZR1去磷酸化形式的蛋白水平受到盐胁迫的明显抑制.OsBZR1为BR信号转导过程的核心转录因子, 对拟南芥的研究表明这类转录因子可直接结合下游基因的启动子, 进而调控大量BR下游基因的表达(Sun et al., 2010; Yu et al., 2011).另外, 作为反馈调控机制, BZR1类转录因子可直接结合BR合成基因, 抑制其表达(Wang et al., 2002; He et al., 2005).本研究创制了OsBZR1融合双标签的转基因株系, 并通过BR处理证实这一材料非常适用于对OsBZR1进行蛋白水平的研究.我们也发现OsBZR1在盐处理2-4小时显著积累, 而此时D2和D11基因表达也达到最低水平, 暗示在盐胁迫过程中植物可对内源激素平衡进行精细调控. ...
BES1 hinders ABSCISIC ACID INSENSITIVE5 and promotes seed germination in Arabidopsis 1 2019
 ,*农作物基因资源与基因改良国家重大科学工程/中国农业科学院作物科学研究所, 北京 100081
,*农作物基因资源与基因改良国家重大科学工程/中国农业科学院作物科学研究所, 北京 100081
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT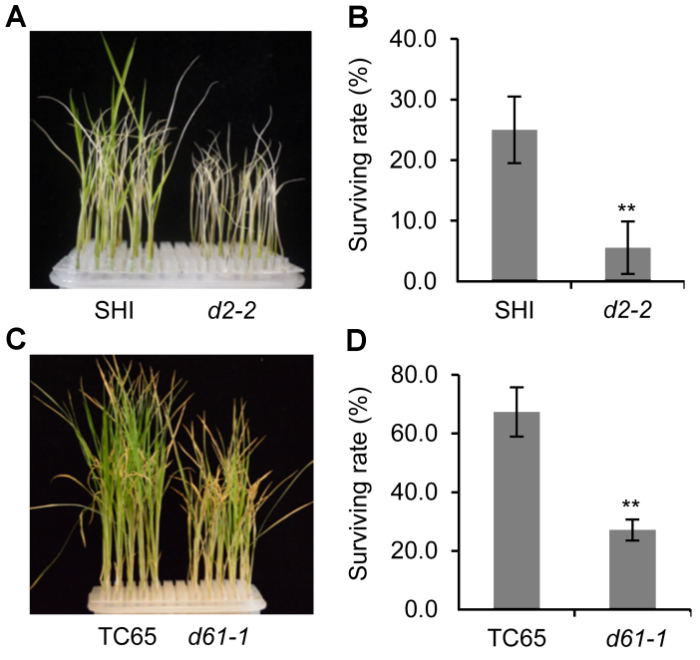 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT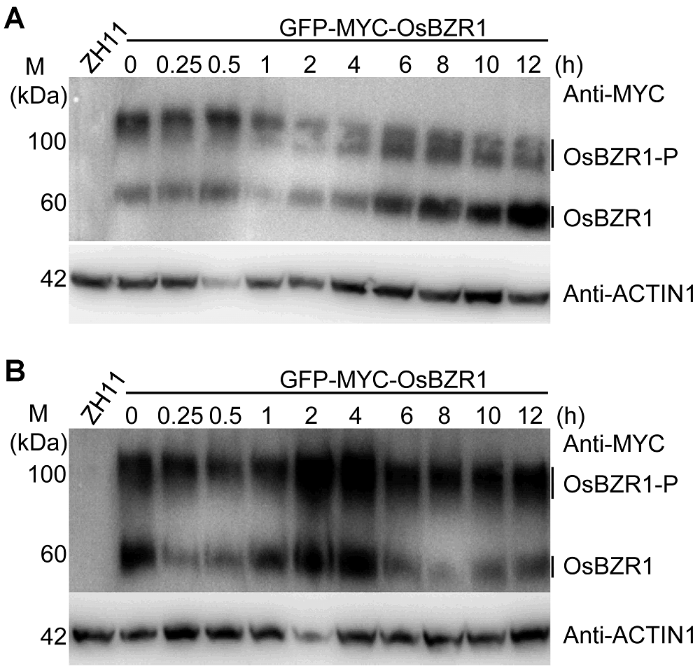 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT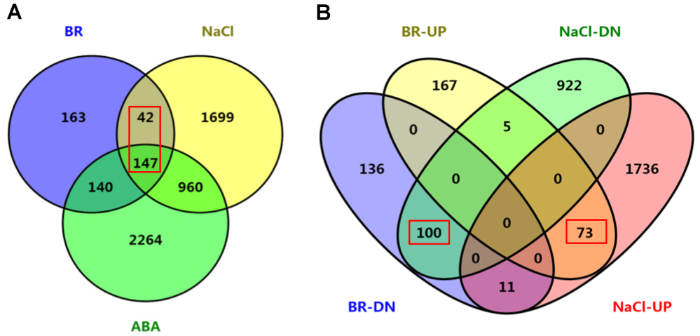 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT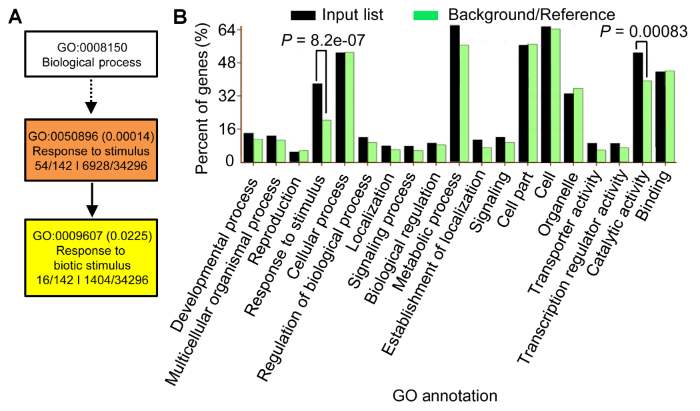 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT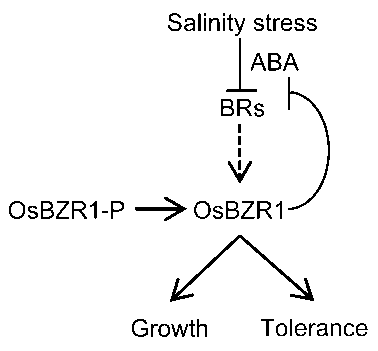 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}