Effect of SmGGPPS2 Expression on Tanshinones Biosynthesis in Salvia miltiorrhiza
Wenping Hua1,2, Chen Chen3,4, Yuan Zhi2, Li Liu1, Zhezhi Wang2, Cuiqin Li,2,*1 Department of Life Sciences and Food Technology, Shaanxi Xueqian Normal University, Xi’an 710061, China 2 Key Laboratory of Medicinal Resources and Natural Pharmaceutical Chemistry, Ministry of Education, National Engineering Laboratory for Resource Developing of Endangered Chinese Crude Drugs in Northwest of China, College of Life Sciences, Shaanxi Normal University, Xi’an 710062, China 3 Xi’an Botanical Garden, Xi’an 710068, China 4 Shaanxi Engineering Research Centre for Conservation and Utilization of Botanical Resources, Xi’an 710068, China
Abstract Geranylgerany pyrophosphate synthase (GGPPS) is an important regulator in the plant diterpenoid biosynthesis pathway. The GGPPS gene family plays a critical role in the development of the medicinal model plant Salvia miltiorrhiza. However, the biological function of SmGGPPS2, especially in the biosynthesis of tanshinone or other active ingredients, is still unclear. For functional investigation, SmGGPPS2 expression was up- or down-regulated in S. miltiorrhiza plants via overexpression or RNA interference, respectively. Then we detected the content of tanshinones, the expression of genes related to tanshinone biosynthesis, and the physiological indexes of transgenic S. miltiorrhiza plants. The content of fat-soluble components, such as tanshinone IIA and ferruginol, was increased significantly in SmGG- PPS2-overexpressed lines compared with the wild type, and the content of fat soluble-components was lower in SmGGPPS2-RNAi lines than those in the wild type lines. With the regulation of SmGGPPS2, the expression of key enzyme genes related to tanshinone biosynthesis in S. miltiorrhiza, such as SmHMGR1 and SmCPS1, was changed. In addition, the regulation of SmGGPPS2 expression also affected the resistance of S. miltiorrhiza. Our results indicate that SmGGPPS2 plays an important regulatory role in tanshinone biosynthesis. Keywords:Salvia miltiorrhiza;GGPPS;tanshinone;gene expression
PDF (2775KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 化文平, 陈尘, 智媛, 刘莉, 王喆之, 李翠芹. SmGGPPS2对丹参酮合成的影响. 植物学报, 2019, 54(2): 217-226 doi:10.11983/CBB18222 Hua Wenping, Chen Chen, Zhi Yuan, Liu Li, Wang Zhezhi, Li Cuiqin. Effect of SmGGPPS2 Expression on Tanshinones Biosynthesis in Salvia miltiorrhiza. Chinese Bulletin of Botany, 2019, 54(2): 217-226 doi:10.11983/CBB18222
香叶基香叶基焦磷酸合酶(Geranylgerany Pyrophosphate Synthase, GGPPS) (EC: 2.5.1.29)催化法尼基焦磷酸(Farnesyl Diphosphate, FPP)和异戊烯基焦磷酸(Isopentenyl Diphosphate, IPP)缩合反应, 生成20碳的香叶基香叶基焦磷酸(Geranylgerany Pyrophosphate, GGPP)。GGPP则是合成叶绿素、类胡萝卜素、赤霉素、香叶基化蛋白和维生素E侧链等二萜类物质的前体物质(Chang et al., 2006)。因而催化合成GGPP的GGPPS在调控二萜类物质代谢途径中发挥重要作用(Aharoni et al., 2003; Han et al., 2006; Chen et al., 2015)。近年来, GGPPS基因在植物次生代谢物合成中的调控功能和生理功能备受关注。例如, 在拟南芥(Arabidopsis thaliana)中过表达甘薯(Ipomoea batatas) IbGGPPS基因不仅增加其类胡萝卜素含量, 同时增强了植株抗渗透胁迫能力(Chen et al., 2015)。在烟草(Nicotiana tabacum)中异源表达向日葵(Helianthus annuus) HaGGPPS基因使其赤霉素含量升高, 促进植物生长(Tata et al., 2016)。
丹参是我国大宗中药材之一, 其药源植物丹参(Salvia miltiorrhiza)是次生代谢研究的药用模式植物。近年来, 有关丹参次生代谢物合成中相关基因的研究及调控已成为国内药用植物的研究热点。探明丹参GGPPS基因功能, 可为揭示丹参酮类药用成分(二萜类衍生物)的合成调控机制奠定基础。近年来, Sm- GGPPS1、SmGGPPS2和SmGGPPS3三个丹参GGPPS家族基因被克隆(张蕾, 2009; Kai et al., 2010; Hua et al., 2012; Ma et al., 2012; 化文平等, 2014)。其中SmGGPPS1和SmGGPPS3都具有GGP- PS合酶活性。SmGGPPS1主要参与叶绿素等物质的合成, 还参与调控丹参酮的合成(Kai et al., 2010; Hua et al., 2012); SmGGPPS3在丹参各器官中均有表达, 但在根中表达量最高, 推测其与根中萜类物质的合成相关(化文平等, 2014)。而SmGGPS2主要在丹参成熟植株的根和花中表达, 推测其可能参与丹参花中类胡萝卜素的合成, 且不具有GGPPS合酶活性(张蕾, 2009; 化文平等, 2014)。SmGGPPS2基因的生物学功能尚不确定, 其在丹参酮等有效成分合成过程中的作用也不明确。本研究通过探讨SmGGPPS2表达对丹参中丹参酮含量、丹参酮合成相关基因表达和转基因丹参的抗性生理指标等的影响, 旨在阐明SmGGPPS2在丹参酮合成调控中的作用及其生物学功能。
A–K同图3。WT: 野生型。*** 差异显著(P<0.001) Figure 4Contents of different ingredients from fat-soluble extract in transgenic lines (A) and contents oftanshinone IIA in different transgenic lines (B) of Salvia miltiorrhiza
GGPPS在植物二萜类产物的合成中起重要调控作用(Chen et al., 2015; Tata et al., 2016)。前期研究已从丹参中克隆得到3个GGPPS基因。其中SmGGPPS1存在于质体类细胞器中, 主要在丹参叶中表达; 在丹参中过表达SmGGPPS1, 丹参酮含量明显升高(Kai et al., 2010; Hua et al., 2012; 化文平等, 2014)。Sm- GGPPS3在根部的表达最高, 可能定位于线粒体并参与丹参根中丹参酮类物质合成(化文平等, 2014)。SmGGPPS2主要在丹参成熟植株的根和花中表达, 推测其可能主要参与花中类胡萝卜素的合成; 但在酵母(Saccharomyces cerevisiae)中进行全长SmGG- PPS2基因功能互补验证, 显示其无法发挥GGPPS的功能(张蕾, 2009)。我们在丹参中过表达Sm- GGPPS2后, 发现丹参酮IIA等多种萜类产物的含量显著升高; 在RNA干涉SmGGPPS2的丹参株系中, 多种萜类成分含量都明显降低。上述结果表明, SmG- GPPS2在丹参植株中有调节萜类物质合成的功能。其在酵母中异源表达不显示功能, 可能是由于在酵母细胞中缺乏SmGGPPS2蛋白加工步骤或成分。
在过表达SmGGPPS1的发根丹参株系中, 总丹参酮含量增高近1.5倍(Shi et al., 2016)。在丹参中过表达SmGGPPS2后, 丹参酮IIA含量是对照株系的2倍(图4B)。铁锈醇是从GGPP到丹参酮类化合物合成的重要中间产物(Ma et al., 2015)。本研究中的GC- MS检测结果表明, 在过表达SmGGPPS2丹参中丹参酮IIA和铁锈醇的含量显著高于野生型对照; RNA干涉SmGGPPS2的丹参株系丹参酮IIA和铁锈醇等萜类物质含量明显低于野生型对照。这表明SmGGP- PS2的表达量与丹参酮IIA和铁锈醇等物质的含量呈一定程度的正相关。
已有研究表明, SmDXS2、SmHMGR1、SmHM- GR2和SmCPS1等基因在丹参酮的合成中发挥重要调控作用(Kai et al., 2011; Cheng et al., 2014; Shi et al., 2014; Cui et al., 2015)。在过表达SmGGPPS1的发根丹参株系中, SmCPS1基因的表达明显上调(Shi et al., 2016)。本研究表明, 在过表达SmGGPPS2的丹参株系中, SmHMGR1、SmHMGR2、SmDXS2、SmDXS3、SmFPS和SmCPS1等关键酶基因的表达均明显上调; 在RNA干涉SmGGPPS2的丹参株系中, SmDXS2、SmHMGR1和SmCPS1等基因呈现下调趋势。同时, 在过表达SmGGPPS2时, SmGGPPS1的表达也呈上调趋势; 而RNA干涉SmGGPPS2时, SmGGPPS3有补偿性上调趋势。由此推测SmGG- PPS2的表达可能通过影响SmDXS2、SmHMGR1和SmCPS1等基因进一步影响丹参酮的合成。
调节GGPPS的表达不仅影响植物次生代谢物的合成, 也可能影响植物的抗性。在拟南芥中过表达马铃薯(Solanum tuberosum) GGPPS基因, 不仅改变其类胡萝卜素等萜类物质的含量, 还影响植物对渗透胁迫的抗性, 导致转基因株系中SOD活性增强, MDA含量降低(Chen et al., 2015)。类胡萝卜素等二萜类物质是GGPPS的下游产物, 具有明显的抗氧化活性(Young and Lowe, 2001)。丹参中丹参酮等有效成分也具有明显的抗氧化活性(Lin and Hsieh, 2010)。在本研究中过表达SmGGPPS2虽然引起丹参中丹参酮等物质含量升高, 但可能由于类胡萝卜素含量呈下降趋势(变化不显著, 数据未显示), 影响转基因株系的脂膜抗氧化能力; 在RNA干涉SmGGPPS2株系中, POD活性、脯氨酸含量和脂膜抗氧化能力有所提升, 这可能是由于GGPPS家族其它成员的补偿表达或其它相关代谢通路的增强所致, 具体机制尚需进一步研究。
AharoniA, GiriAP, DeuerleinS, GriepinkF, de KogelWJ, VerstappenFWA, VerhoevenHA, JongsmaMA, SchwabW, BouwmeesterHJ ( 2003). Terpenoid metabolism in wild-type and transgenic Arabidopsis plants Plant Cell 15, 2866-2884. DOI:10.1105/tpc.016253URL [本文引用: 1]
ChangTH, GuoRT, KoTP, WangAHJ, LiangPH ( 2006). Crystal structure of type-III geranylgeranyl pyrophosphate synthase from Saccharomyces cerevisiae and the mecha- nism of product chain length determination J Biol Chem 281, 14991-15000. [本文引用: 1]
ChenC, ZhangY, QiakefuK, ZhangX, HanLM, HuaWP, YanYP, WangZZ ( 2016). Overexpression of tomato Prosystemin (LePS) enhances pest resistance and the production of tanshinones in Salvia miltiorrhiza Bunge J Agric Food Chem 64, 7760-7769. [本文引用: 1]
ChenW, HeSZ, LiuDG, PatilGB, ZhaiH, WangFB, StephensonTJ, WangYN, WangB, ValliyodanB, NguyenHT, LiuQC ( 2015). A sweetpotato geranylgeranyl pyrophosphate synthase gene, IbGGPS, increases carotenoid content and enhances osmotic stress tolerance in Arabidopsis thaliana PLoS One 10, e0137623. [本文引用: 5]
ChengQQ, SuP, HuYT, HeYF, GaoW, HuangLQ ( 2014). RNA interference-mediated repression of SmCPS (Copalyldiphosphate synthase) expression in hairy roots of Salvia miltiorrhiza causes a decrease of tanshinones and sheds light on the functional role of SmCPS Biotechnol Lett 36, 363-369. [本文引用: 1]
CuiGH, DuanLX, JinBL, QianJ, XueZY, ShenGA, SnyderJH, SongJY, ChenSL, HuangLQ, PetersRJ, QiXQ ( 2015). Functional divergence of diterpene syntheses in the medicinal plant Salvia miltiorrhiza Plant Physiol 169, 1607-1618. [本文引用: 1]
HanJL, LiuBY, YeHC, WangH, LiZQ, LiGF ( 2006). Effects of overexpression of the endogenous farnesyl diphosphate synthase on the artemisinin content in Artemisia annua L J Integr Plant Biol 48, 482-487. [本文引用: 1]
HuaWP, SongJ, LiCQ, WangZZ ( 2012). Molecular cloning and characterization of the promoter of SmGGPPs and its expression pattern in Salvia miltiorrhiza Mol Biol Rep 39, 5775-5783. [本文引用: 3]
HuaWP, ZhangY, SongJ, ZhaoLJ, WangZZ ( 2011). De novo transcriptome sequencing in Salvia miltiorrhiza to identify genes involved in the biosynthesis of active ingredients Genomics 98, 272-279. [本文引用: 2]
KaiGY, LiaoP, ZhangT, ZhouW, WangJ, XuH, LiuYY, ZhangL ( 2010). Characterization, expression profiling, and functional identification of a gene encoding geranylgeranyl diphosphate synthase from Salvia miltiorrhiza Bio- technol Bioproc Eng 15, 236-245. [本文引用: 3]
KaiGY, XuH, ZhouCC, LiaoP, XiaoJB, LuoXQ, YouLJ, ZhangL ( 2011). Metabolic engineering tanshinone biosynthetic pathway in Salvia miltiorrhiza hairy root cultures Metab Eng 13, 319-327. [本文引用: 1]
LinTH, HsiehCL ( 2010). Pharmacological effects of Salvia miltiorrhiza( Danshen) on cerebral infarction Chin Med 5, 22. [本文引用: 2]
MaXH, MaY, TangJF, HeYL, LiuYC, MaXJ, ShenY, CuiGH, LinHX, RongQX, GuoJ, HuangLQ ( 2015). The biosynthetic pathways of tanshinones and phenolic acids in Salvia miltiorrhiza Molecules 20, 16235-16254. [本文引用: 1]
MaYM, YuanLC, WuB, LiXE, ChenSL, LuSF ( 2012). Genome-wide identification and characterization of novel genes involved in terpenoid biosynthesis in Salvia miltiorrhiza J Exp Bot 63, 2809-2823. [本文引用: 1]
ShiM, LuoXQ, JuGH, LiLL, HuangSX, ZhangT, WangHZ, KaiGY ( 2016). Enhanced diterpene tanshinone accumulation and bioactivity of transgenic Salvia miltiorrhiza hairy roots by pathway engineering J Agric Food Chem 64, 2523-2530. [本文引用: 2]
ShiM, LuoXQ, JuGH, YuXH, HaoXL, HuangQ, XiaoJB, CuiLJ, KaiGY ( 2014). Increased accumulation of the cardio-cerebrovascular disease treatment drug tanshinone in Salvia miltiorrhiza hairy roots by the enzymes 3-hy- droxy-3-methylglutaryl CoA reductase and 1-deoxy-Dxylulose 5-phosphate reductoisomerase Funct Integr Genom 14, 603-615. [本文引用: 1]
TataSK, JungJ, KimYH, ChoiJY, JungJY, LeeIJ, ShinJS, RyuSB ( 2016). Heterologous expression of chloroplast-localized geranylgeranyl pyrophosphate synthase confers fast plant growth, early flowering and increased seed yield Plant Biotechnol J 14, 29-39. DOI:10.1111/pbi.12333URL [本文引用: 2]
VandesompeleJ, De PreterK, PattynF, PoppeB, Van RoyN, De PaepeA, SpelemanF ( 2002). Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes Genome Biol 3, research0034. [本文引用: 1]
YanYP, WangZZ ( 2007). Genetic transformation of the medicinal plant Salvia miltiorrhiza by Agrobacterium tumefaciens-mediated method Plant Cell Tiss Org 88, 175-184. [本文引用: 1]
YoungAJ, LoweGM ( 2001). Antioxidant and prooxidant properties of carotenoids Arch Biochem Biophys 385, 20-27. DOI:10.1006/abbi.2000.2149URL [本文引用: 1]
... GGPPS在植物二萜类产物的合成中起重要调控作用(Chen et al., 2015; Tata et al., 2016).前期研究已从丹参中克隆得到3个GGPPS基因.其中SmGGPPS1存在于质体类细胞器中, 主要在丹参叶中表达; 在丹参中过表达SmGGPPS1, 丹参酮含量明显升高(Kai et al., 2010; Hua et al., 2012; 化文平等, 2014).Sm- GGPPS3在根部的表达最高, 可能定位于线粒体并参与丹参根中丹参酮类物质合成(化文平等, 2014).SmGGPPS2主要在丹参成熟植株的根和花中表达, 推测其可能主要参与花中类胡萝卜素的合成; 但在酵母(Saccharomyces cerevisiae)中进行全长SmGG- PPS2基因功能互补验证, 显示其无法发挥GGPPS的功能(张蕾, 2009).我们在丹参中过表达Sm- GGPPS2后, 发现丹参酮IIA等多种萜类产物的含量显著升高; 在RNA干涉SmGGPPS2的丹参株系中, 多种萜类成分含量都明显降低.上述结果表明, SmG- GPPS2在丹参植株中有调节萜类物质合成的功能.其在酵母中异源表达不显示功能, 可能是由于在酵母细胞中缺乏SmGGPPS2蛋白加工步骤或成分. ...
Terpenoid metabolism in wild-type and transgenic Arabidopsis plants 1 2003
... 香叶基香叶基焦磷酸合酶(Geranylgerany Pyrophosphate Synthase, GGPPS) (EC: 2.5.1.29)催化法尼基焦磷酸(Farnesyl Diphosphate, FPP)和异戊烯基焦磷酸(Isopentenyl Diphosphate, IPP)缩合反应, 生成20碳的香叶基香叶基焦磷酸(Geranylgerany Pyrophosphate, GGPP).GGPP则是合成叶绿素、类胡萝卜素、赤霉素、香叶基化蛋白和维生素E侧链等二萜类物质的前体物质(Chang et al., 2006).因而催化合成GGPP的GGPPS在调控二萜类物质代谢途径中发挥重要作用(Aharoni et al., 2003; Han et al., 2006; Chen et al., 2015).近年来, GGPPS基因在植物次生代谢物合成中的调控功能和生理功能备受关注.例如, 在拟南芥(Arabidopsis thaliana)中过表达甘薯(Ipomoea batatas) IbGGPPS基因不仅增加其类胡萝卜素含量, 同时增强了植株抗渗透胁迫能力(Chen et al., 2015).在烟草(Nicotiana tabacum)中异源表达向日葵(Helianthus annuus) HaGGPPS基因使其赤霉素含量升高, 促进植物生长(Tata et al., 2016). ...
Crystal structure of type-III geranylgeranyl pyrophosphate synthase from Saccharomyces cerevisiae and the mecha- nism of product chain length determination 1 2006
... 香叶基香叶基焦磷酸合酶(Geranylgerany Pyrophosphate Synthase, GGPPS) (EC: 2.5.1.29)催化法尼基焦磷酸(Farnesyl Diphosphate, FPP)和异戊烯基焦磷酸(Isopentenyl Diphosphate, IPP)缩合反应, 生成20碳的香叶基香叶基焦磷酸(Geranylgerany Pyrophosphate, GGPP).GGPP则是合成叶绿素、类胡萝卜素、赤霉素、香叶基化蛋白和维生素E侧链等二萜类物质的前体物质(Chang et al., 2006).因而催化合成GGPP的GGPPS在调控二萜类物质代谢途径中发挥重要作用(Aharoni et al., 2003; Han et al., 2006; Chen et al., 2015).近年来, GGPPS基因在植物次生代谢物合成中的调控功能和生理功能备受关注.例如, 在拟南芥(Arabidopsis thaliana)中过表达甘薯(Ipomoea batatas) IbGGPPS基因不仅增加其类胡萝卜素含量, 同时增强了植株抗渗透胁迫能力(Chen et al., 2015).在烟草(Nicotiana tabacum)中异源表达向日葵(Helianthus annuus) HaGGPPS基因使其赤霉素含量升高, 促进植物生长(Tata et al., 2016). ...
Overexpression of tomato Prosystemin (LePS) enhances pest resistance and the production of tanshinones in Salvia miltiorrhiza Bunge 1 2016
... GGPPS在植物二萜类产物的合成中起重要调控作用(Chen et al., 2015; Tata et al., 2016).前期研究已从丹参中克隆得到3个GGPPS基因.其中SmGGPPS1存在于质体类细胞器中, 主要在丹参叶中表达; 在丹参中过表达SmGGPPS1, 丹参酮含量明显升高(Kai et al., 2010; Hua et al., 2012; 化文平等, 2014).Sm- GGPPS3在根部的表达最高, 可能定位于线粒体并参与丹参根中丹参酮类物质合成(化文平等, 2014).SmGGPPS2主要在丹参成熟植株的根和花中表达, 推测其可能主要参与花中类胡萝卜素的合成; 但在酵母(Saccharomyces cerevisiae)中进行全长SmGG- PPS2基因功能互补验证, 显示其无法发挥GGPPS的功能(张蕾, 2009).我们在丹参中过表达Sm- GGPPS2后, 发现丹参酮IIA等多种萜类产物的含量显著升高; 在RNA干涉SmGGPPS2的丹参株系中, 多种萜类成分含量都明显降低.上述结果表明, SmG- GPPS2在丹参植株中有调节萜类物质合成的功能.其在酵母中异源表达不显示功能, 可能是由于在酵母细胞中缺乏SmGGPPS2蛋白加工步骤或成分. ...
... 调节GGPPS的表达不仅影响植物次生代谢物的合成, 也可能影响植物的抗性.在拟南芥中过表达马铃薯(Solanum tuberosum) GGPPS基因, 不仅改变其类胡萝卜素等萜类物质的含量, 还影响植物对渗透胁迫的抗性, 导致转基因株系中SOD活性增强, MDA含量降低(Chen et al., 2015).类胡萝卜素等二萜类物质是GGPPS的下游产物, 具有明显的抗氧化活性(Young and Lowe, 2001).丹参中丹参酮等有效成分也具有明显的抗氧化活性(Lin and Hsieh, 2010).在本研究中过表达SmGGPPS2虽然引起丹参中丹参酮等物质含量升高, 但可能由于类胡萝卜素含量呈下降趋势(变化不显著, 数据未显示), 影响转基因株系的脂膜抗氧化能力; 在RNA干涉SmGGPPS2株系中, POD活性、脯氨酸含量和脂膜抗氧化能力有所提升, 这可能是由于GGPPS家族其它成员的补偿表达或其它相关代谢通路的增强所致, 具体机制尚需进一步研究. ...
RNA interference-mediated repression of SmCPS (Copalyldiphosphate synthase) expression in hairy roots of Salvia miltiorrhiza causes a decrease of tanshinones and sheds light on the functional role of SmCPS 1 2014
... 已有研究表明, SmDXS2、SmHMGR1、SmHM- GR2和SmCPS1等基因在丹参酮的合成中发挥重要调控作用(Kai et al., 2011; Cheng et al., 2014; Shi et al., 2014; Cui et al., 2015).在过表达SmGGPPS1的发根丹参株系中, SmCPS1基因的表达明显上调(Shi et al., 2016).本研究表明, 在过表达SmGGPPS2的丹参株系中, SmHMGR1、SmHMGR2、SmDXS2、SmDXS3、SmFPS和SmCPS1等关键酶基因的表达均明显上调; 在RNA干涉SmGGPPS2的丹参株系中, SmDXS2、SmHMGR1和SmCPS1等基因呈现下调趋势.同时, 在过表达SmGGPPS2时, SmGGPPS1的表达也呈上调趋势; 而RNA干涉SmGGPPS2时, SmGGPPS3有补偿性上调趋势.由此推测SmGG- PPS2的表达可能通过影响SmDXS2、SmHMGR1和SmCPS1等基因进一步影响丹参酮的合成. ...
Functional divergence of diterpene syntheses in the medicinal plant Salvia miltiorrhiza 1 2015
... 已有研究表明, SmDXS2、SmHMGR1、SmHM- GR2和SmCPS1等基因在丹参酮的合成中发挥重要调控作用(Kai et al., 2011; Cheng et al., 2014; Shi et al., 2014; Cui et al., 2015).在过表达SmGGPPS1的发根丹参株系中, SmCPS1基因的表达明显上调(Shi et al., 2016).本研究表明, 在过表达SmGGPPS2的丹参株系中, SmHMGR1、SmHMGR2、SmDXS2、SmDXS3、SmFPS和SmCPS1等关键酶基因的表达均明显上调; 在RNA干涉SmGGPPS2的丹参株系中, SmDXS2、SmHMGR1和SmCPS1等基因呈现下调趋势.同时, 在过表达SmGGPPS2时, SmGGPPS1的表达也呈上调趋势; 而RNA干涉SmGGPPS2时, SmGGPPS3有补偿性上调趋势.由此推测SmGG- PPS2的表达可能通过影响SmDXS2、SmHMGR1和SmCPS1等基因进一步影响丹参酮的合成. ...
Effects of overexpression of the endogenous farnesyl diphosphate synthase on the artemisinin content in Artemisia annua L 1 2006
... 香叶基香叶基焦磷酸合酶(Geranylgerany Pyrophosphate Synthase, GGPPS) (EC: 2.5.1.29)催化法尼基焦磷酸(Farnesyl Diphosphate, FPP)和异戊烯基焦磷酸(Isopentenyl Diphosphate, IPP)缩合反应, 生成20碳的香叶基香叶基焦磷酸(Geranylgerany Pyrophosphate, GGPP).GGPP则是合成叶绿素、类胡萝卜素、赤霉素、香叶基化蛋白和维生素E侧链等二萜类物质的前体物质(Chang et al., 2006).因而催化合成GGPP的GGPPS在调控二萜类物质代谢途径中发挥重要作用(Aharoni et al., 2003; Han et al., 2006; Chen et al., 2015).近年来, GGPPS基因在植物次生代谢物合成中的调控功能和生理功能备受关注.例如, 在拟南芥(Arabidopsis thaliana)中过表达甘薯(Ipomoea batatas) IbGGPPS基因不仅增加其类胡萝卜素含量, 同时增强了植株抗渗透胁迫能力(Chen et al., 2015).在烟草(Nicotiana tabacum)中异源表达向日葵(Helianthus annuus) HaGGPPS基因使其赤霉素含量升高, 促进植物生长(Tata et al., 2016). ...
Molecular cloning and characterization of the promoter of SmGGPPs and its expression pattern in Salvia miltiorrhiza 3 2012
... 丹参是我国大宗中药材之一, 其药源植物丹参(Salvia miltiorrhiza)是次生代谢研究的药用模式植物.近年来, 有关丹参次生代谢物合成中相关基因的研究及调控已成为国内药用植物的研究热点.探明丹参GGPPS基因功能, 可为揭示丹参酮类药用成分(二萜类衍生物)的合成调控机制奠定基础.近年来, Sm- GGPPS1、SmGGPPS2和SmGGPPS3三个丹参GGPPS家族基因被克隆(张蕾, 2009; Kai et al., 2010; Hua et al., 2012; Ma et al., 2012; 化文平等, 2014).其中SmGGPPS1和SmGGPPS3都具有GGP- PS合酶活性.SmGGPPS1主要参与叶绿素等物质的合成, 还参与调控丹参酮的合成(Kai et al., 2010; Hua et al., 2012); SmGGPPS3在丹参各器官中均有表达, 但在根中表达量最高, 推测其与根中萜类物质的合成相关(化文平等, 2014).而SmGGPS2主要在丹参成熟植株的根和花中表达, 推测其可能参与丹参花中类胡萝卜素的合成, 且不具有GGPPS合酶活性(张蕾, 2009; 化文平等, 2014).SmGGPPS2基因的生物学功能尚不确定, 其在丹参酮等有效成分合成过程中的作用也不明确.本研究通过探讨SmGGPPS2表达对丹参中丹参酮含量、丹参酮合成相关基因表达和转基因丹参的抗性生理指标等的影响, 旨在阐明SmGGPPS2在丹参酮合成调控中的作用及其生物学功能. ...
Characterization, expression profiling, and functional identification of a gene encoding geranylgeranyl diphosphate synthase from Salvia miltiorrhiza 3 2010
... 丹参是我国大宗中药材之一, 其药源植物丹参(Salvia miltiorrhiza)是次生代谢研究的药用模式植物.近年来, 有关丹参次生代谢物合成中相关基因的研究及调控已成为国内药用植物的研究热点.探明丹参GGPPS基因功能, 可为揭示丹参酮类药用成分(二萜类衍生物)的合成调控机制奠定基础.近年来, Sm- GGPPS1、SmGGPPS2和SmGGPPS3三个丹参GGPPS家族基因被克隆(张蕾, 2009; Kai et al., 2010; Hua et al., 2012; Ma et al., 2012; 化文平等, 2014).其中SmGGPPS1和SmGGPPS3都具有GGP- PS合酶活性.SmGGPPS1主要参与叶绿素等物质的合成, 还参与调控丹参酮的合成(Kai et al., 2010; Hua et al., 2012); SmGGPPS3在丹参各器官中均有表达, 但在根中表达量最高, 推测其与根中萜类物质的合成相关(化文平等, 2014).而SmGGPS2主要在丹参成熟植株的根和花中表达, 推测其可能参与丹参花中类胡萝卜素的合成, 且不具有GGPPS合酶活性(张蕾, 2009; 化文平等, 2014).SmGGPPS2基因的生物学功能尚不确定, 其在丹参酮等有效成分合成过程中的作用也不明确.本研究通过探讨SmGGPPS2表达对丹参中丹参酮含量、丹参酮合成相关基因表达和转基因丹参的抗性生理指标等的影响, 旨在阐明SmGGPPS2在丹参酮合成调控中的作用及其生物学功能. ...
... GGPPS在植物二萜类产物的合成中起重要调控作用(Chen et al., 2015; Tata et al., 2016).前期研究已从丹参中克隆得到3个GGPPS基因.其中SmGGPPS1存在于质体类细胞器中, 主要在丹参叶中表达; 在丹参中过表达SmGGPPS1, 丹参酮含量明显升高(Kai et al., 2010; Hua et al., 2012; 化文平等, 2014).Sm- GGPPS3在根部的表达最高, 可能定位于线粒体并参与丹参根中丹参酮类物质合成(化文平等, 2014).SmGGPPS2主要在丹参成熟植株的根和花中表达, 推测其可能主要参与花中类胡萝卜素的合成; 但在酵母(Saccharomyces cerevisiae)中进行全长SmGG- PPS2基因功能互补验证, 显示其无法发挥GGPPS的功能(张蕾, 2009).我们在丹参中过表达Sm- GGPPS2后, 发现丹参酮IIA等多种萜类产物的含量显著升高; 在RNA干涉SmGGPPS2的丹参株系中, 多种萜类成分含量都明显降低.上述结果表明, SmG- GPPS2在丹参植株中有调节萜类物质合成的功能.其在酵母中异源表达不显示功能, 可能是由于在酵母细胞中缺乏SmGGPPS2蛋白加工步骤或成分. ...
Metabolic engineering tanshinone biosynthetic pathway in Salvia miltiorrhiza hairy root cultures 1 2011
... 已有研究表明, SmDXS2、SmHMGR1、SmHM- GR2和SmCPS1等基因在丹参酮的合成中发挥重要调控作用(Kai et al., 2011; Cheng et al., 2014; Shi et al., 2014; Cui et al., 2015).在过表达SmGGPPS1的发根丹参株系中, SmCPS1基因的表达明显上调(Shi et al., 2016).本研究表明, 在过表达SmGGPPS2的丹参株系中, SmHMGR1、SmHMGR2、SmDXS2、SmDXS3、SmFPS和SmCPS1等关键酶基因的表达均明显上调; 在RNA干涉SmGGPPS2的丹参株系中, SmDXS2、SmHMGR1和SmCPS1等基因呈现下调趋势.同时, 在过表达SmGGPPS2时, SmGGPPS1的表达也呈上调趋势; 而RNA干涉SmGGPPS2时, SmGGPPS3有补偿性上调趋势.由此推测SmGG- PPS2的表达可能通过影响SmDXS2、SmHMGR1和SmCPS1等基因进一步影响丹参酮的合成. ...
Pharmacological effects of Salvia miltiorrhiza( Danshen) on cerebral infarction 2 2010
... 调节GGPPS的表达不仅影响植物次生代谢物的合成, 也可能影响植物的抗性.在拟南芥中过表达马铃薯(Solanum tuberosum) GGPPS基因, 不仅改变其类胡萝卜素等萜类物质的含量, 还影响植物对渗透胁迫的抗性, 导致转基因株系中SOD活性增强, MDA含量降低(Chen et al., 2015).类胡萝卜素等二萜类物质是GGPPS的下游产物, 具有明显的抗氧化活性(Young and Lowe, 2001).丹参中丹参酮等有效成分也具有明显的抗氧化活性(Lin and Hsieh, 2010).在本研究中过表达SmGGPPS2虽然引起丹参中丹参酮等物质含量升高, 但可能由于类胡萝卜素含量呈下降趋势(变化不显著, 数据未显示), 影响转基因株系的脂膜抗氧化能力; 在RNA干涉SmGGPPS2株系中, POD活性、脯氨酸含量和脂膜抗氧化能力有所提升, 这可能是由于GGPPS家族其它成员的补偿表达或其它相关代谢通路的增强所致, 具体机制尚需进一步研究. ...
The biosynthetic pathways of tanshinones and phenolic acids in Salvia miltiorrhiza 1 2015
... 在过表达SmGGPPS1的发根丹参株系中, 总丹参酮含量增高近1.5倍(Shi et al., 2016).在丹参中过表达SmGGPPS2后, 丹参酮IIA含量是对照株系的2倍(图4B).铁锈醇是从GGPP到丹参酮类化合物合成的重要中间产物(Ma et al., 2015).本研究中的GC- MS检测结果表明, 在过表达SmGGPPS2丹参中丹参酮IIA和铁锈醇的含量显著高于野生型对照; RNA干涉SmGGPPS2的丹参株系丹参酮IIA和铁锈醇等萜类物质含量明显低于野生型对照.这表明SmGGP- PS2的表达量与丹参酮IIA和铁锈醇等物质的含量呈一定程度的正相关. ...
Genome-wide identification and characterization of novel genes involved in terpenoid biosynthesis in Salvia miltiorrhiza 1 2012
... 丹参是我国大宗中药材之一, 其药源植物丹参(Salvia miltiorrhiza)是次生代谢研究的药用模式植物.近年来, 有关丹参次生代谢物合成中相关基因的研究及调控已成为国内药用植物的研究热点.探明丹参GGPPS基因功能, 可为揭示丹参酮类药用成分(二萜类衍生物)的合成调控机制奠定基础.近年来, Sm- GGPPS1、SmGGPPS2和SmGGPPS3三个丹参GGPPS家族基因被克隆(张蕾, 2009; Kai et al., 2010; Hua et al., 2012; Ma et al., 2012; 化文平等, 2014).其中SmGGPPS1和SmGGPPS3都具有GGP- PS合酶活性.SmGGPPS1主要参与叶绿素等物质的合成, 还参与调控丹参酮的合成(Kai et al., 2010; Hua et al., 2012); SmGGPPS3在丹参各器官中均有表达, 但在根中表达量最高, 推测其与根中萜类物质的合成相关(化文平等, 2014).而SmGGPS2主要在丹参成熟植株的根和花中表达, 推测其可能参与丹参花中类胡萝卜素的合成, 且不具有GGPPS合酶活性(张蕾, 2009; 化文平等, 2014).SmGGPPS2基因的生物学功能尚不确定, 其在丹参酮等有效成分合成过程中的作用也不明确.本研究通过探讨SmGGPPS2表达对丹参中丹参酮含量、丹参酮合成相关基因表达和转基因丹参的抗性生理指标等的影响, 旨在阐明SmGGPPS2在丹参酮合成调控中的作用及其生物学功能. ...
Enhanced diterpene tanshinone accumulation and bioactivity of transgenic Salvia miltiorrhiza hairy roots by pathway engineering 2 2016
... 在过表达SmGGPPS1的发根丹参株系中, 总丹参酮含量增高近1.5倍(Shi et al., 2016).在丹参中过表达SmGGPPS2后, 丹参酮IIA含量是对照株系的2倍(图4B).铁锈醇是从GGPP到丹参酮类化合物合成的重要中间产物(Ma et al., 2015).本研究中的GC- MS检测结果表明, 在过表达SmGGPPS2丹参中丹参酮IIA和铁锈醇的含量显著高于野生型对照; RNA干涉SmGGPPS2的丹参株系丹参酮IIA和铁锈醇等萜类物质含量明显低于野生型对照.这表明SmGGP- PS2的表达量与丹参酮IIA和铁锈醇等物质的含量呈一定程度的正相关. ...
... 已有研究表明, SmDXS2、SmHMGR1、SmHM- GR2和SmCPS1等基因在丹参酮的合成中发挥重要调控作用(Kai et al., 2011; Cheng et al., 2014; Shi et al., 2014; Cui et al., 2015).在过表达SmGGPPS1的发根丹参株系中, SmCPS1基因的表达明显上调(Shi et al., 2016).本研究表明, 在过表达SmGGPPS2的丹参株系中, SmHMGR1、SmHMGR2、SmDXS2、SmDXS3、SmFPS和SmCPS1等关键酶基因的表达均明显上调; 在RNA干涉SmGGPPS2的丹参株系中, SmDXS2、SmHMGR1和SmCPS1等基因呈现下调趋势.同时, 在过表达SmGGPPS2时, SmGGPPS1的表达也呈上调趋势; 而RNA干涉SmGGPPS2时, SmGGPPS3有补偿性上调趋势.由此推测SmGG- PPS2的表达可能通过影响SmDXS2、SmHMGR1和SmCPS1等基因进一步影响丹参酮的合成. ...
Increased accumulation of the cardio-cerebrovascular disease treatment drug tanshinone in Salvia miltiorrhiza hairy roots by the enzymes 3-hy- droxy-3-methylglutaryl CoA reductase and 1-deoxy-Dxylulose 5-phosphate reductoisomerase 1 2014
... 已有研究表明, SmDXS2、SmHMGR1、SmHM- GR2和SmCPS1等基因在丹参酮的合成中发挥重要调控作用(Kai et al., 2011; Cheng et al., 2014; Shi et al., 2014; Cui et al., 2015).在过表达SmGGPPS1的发根丹参株系中, SmCPS1基因的表达明显上调(Shi et al., 2016).本研究表明, 在过表达SmGGPPS2的丹参株系中, SmHMGR1、SmHMGR2、SmDXS2、SmDXS3、SmFPS和SmCPS1等关键酶基因的表达均明显上调; 在RNA干涉SmGGPPS2的丹参株系中, SmDXS2、SmHMGR1和SmCPS1等基因呈现下调趋势.同时, 在过表达SmGGPPS2时, SmGGPPS1的表达也呈上调趋势; 而RNA干涉SmGGPPS2时, SmGGPPS3有补偿性上调趋势.由此推测SmGG- PPS2的表达可能通过影响SmDXS2、SmHMGR1和SmCPS1等基因进一步影响丹参酮的合成. ...
Heterologous expression of chloroplast-localized geranylgeranyl pyrophosphate synthase confers fast plant growth, early flowering and increased seed yield 2 2016
... 香叶基香叶基焦磷酸合酶(Geranylgerany Pyrophosphate Synthase, GGPPS) (EC: 2.5.1.29)催化法尼基焦磷酸(Farnesyl Diphosphate, FPP)和异戊烯基焦磷酸(Isopentenyl Diphosphate, IPP)缩合反应, 生成20碳的香叶基香叶基焦磷酸(Geranylgerany Pyrophosphate, GGPP).GGPP则是合成叶绿素、类胡萝卜素、赤霉素、香叶基化蛋白和维生素E侧链等二萜类物质的前体物质(Chang et al., 2006).因而催化合成GGPP的GGPPS在调控二萜类物质代谢途径中发挥重要作用(Aharoni et al., 2003; Han et al., 2006; Chen et al., 2015).近年来, GGPPS基因在植物次生代谢物合成中的调控功能和生理功能备受关注.例如, 在拟南芥(Arabidopsis thaliana)中过表达甘薯(Ipomoea batatas) IbGGPPS基因不仅增加其类胡萝卜素含量, 同时增强了植株抗渗透胁迫能力(Chen et al., 2015).在烟草(Nicotiana tabacum)中异源表达向日葵(Helianthus annuus) HaGGPPS基因使其赤霉素含量升高, 促进植物生长(Tata et al., 2016). ...
... GGPPS在植物二萜类产物的合成中起重要调控作用(Chen et al., 2015; Tata et al., 2016).前期研究已从丹参中克隆得到3个GGPPS基因.其中SmGGPPS1存在于质体类细胞器中, 主要在丹参叶中表达; 在丹参中过表达SmGGPPS1, 丹参酮含量明显升高(Kai et al., 2010; Hua et al., 2012; 化文平等, 2014).Sm- GGPPS3在根部的表达最高, 可能定位于线粒体并参与丹参根中丹参酮类物质合成(化文平等, 2014).SmGGPPS2主要在丹参成熟植株的根和花中表达, 推测其可能主要参与花中类胡萝卜素的合成; 但在酵母(Saccharomyces cerevisiae)中进行全长SmGG- PPS2基因功能互补验证, 显示其无法发挥GGPPS的功能(张蕾, 2009).我们在丹参中过表达Sm- GGPPS2后, 发现丹参酮IIA等多种萜类产物的含量显著升高; 在RNA干涉SmGGPPS2的丹参株系中, 多种萜类成分含量都明显降低.上述结果表明, SmG- GPPS2在丹参植株中有调节萜类物质合成的功能.其在酵母中异源表达不显示功能, 可能是由于在酵母细胞中缺乏SmGGPPS2蛋白加工步骤或成分. ...
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes 1 2002
 ,2,*1
,2,*1 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT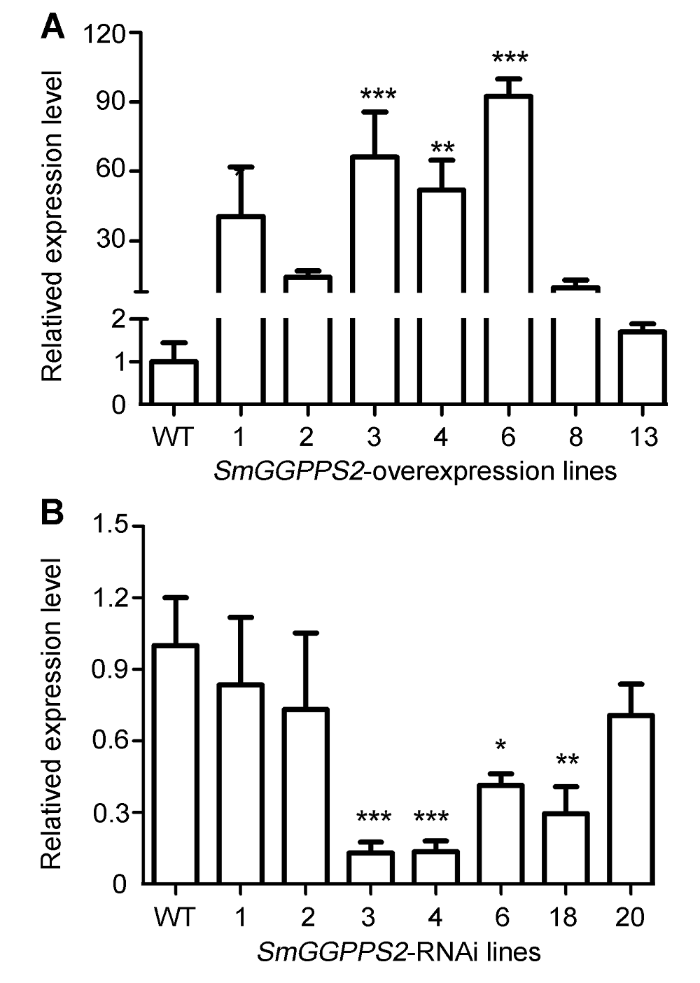 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT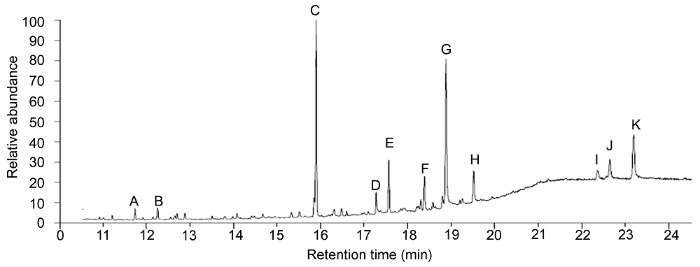 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT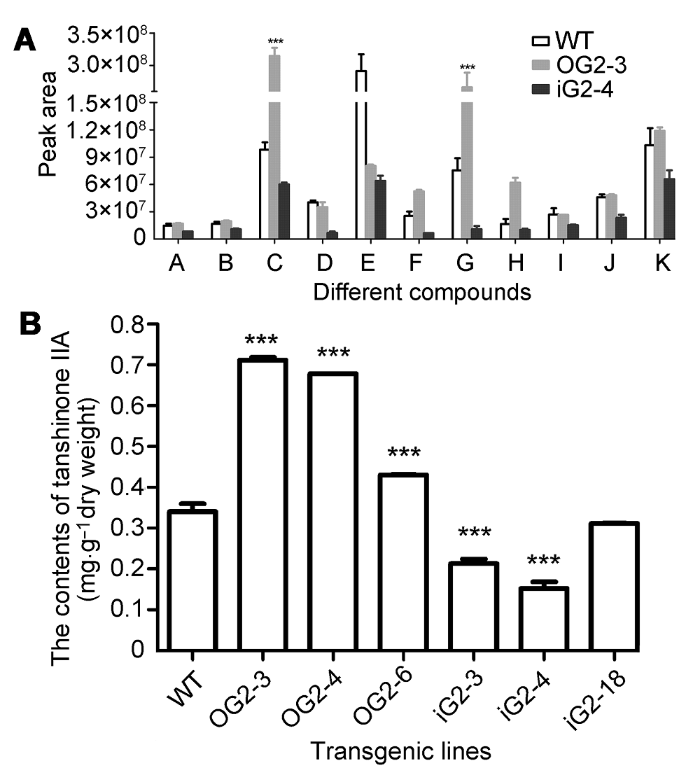 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT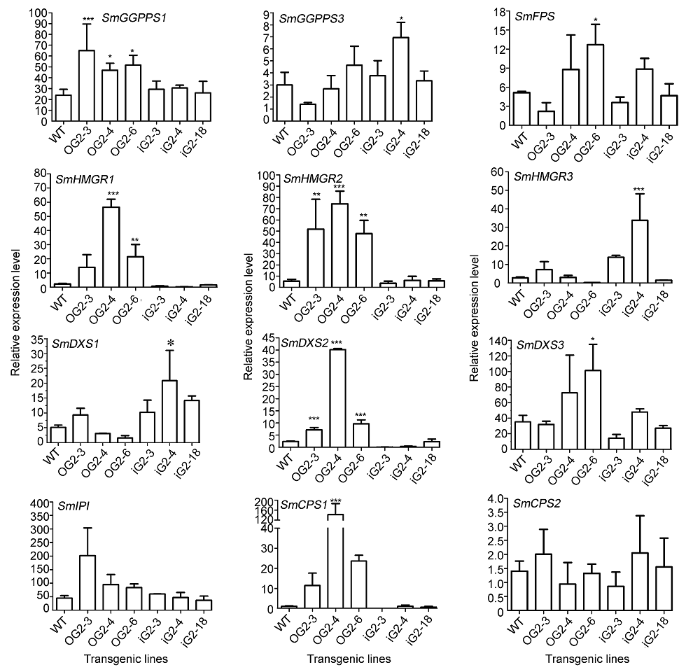 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT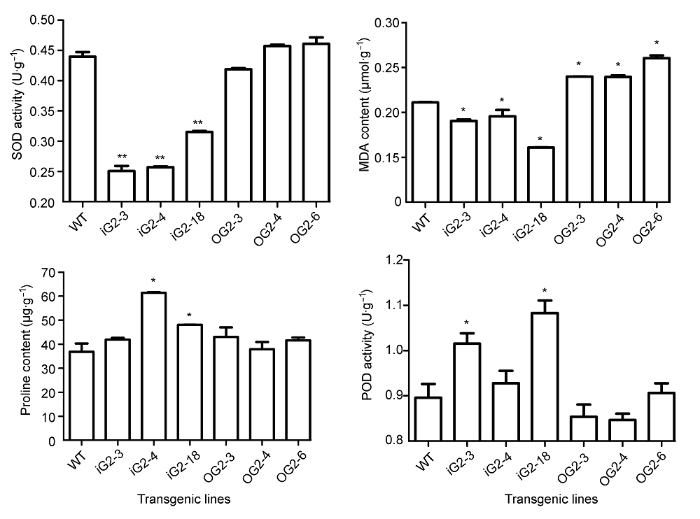 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}