Abstract Peach (Amygdalus persica) seedlings were used to analyze the effect of ammonium molybdate on the expression of molybdenum cofactor sulfurase gene (LOS5/ABA3), ABA content and physiological indexes related to drought resistance under drought stress. Water, chlorophyll and proline contents of peach seedling leaves were significantly higher with ammonium molybdate, preferably 0.04%, than control treatment, and electrolyte leakage was lower. Under drought stress, the expression of LOS5/ABA3 in leaves treated with 0.04% ammonium molybdate was significantly increased and ABA content, water use efficiency, and net photosynthetic rate was higher than with control treatment. Transpiration rate was lower, antioxidant enzyme activity was higher, and malondialdehyde content was lower in leaves; the quality loss of leaves in vitro was reduced. Thus, ammonium molybdate treatment of peach seedlings can increase the content of ABA and proline, increase the activity of antioxidant enzymes, relieve cell membrane oxidation damage, decrease the rate of water loss of leaves, and alleviate drought stress damage by regulating the expression of drought- resistant genes, then increase the drought resistance of plants. Keywords:peach seedlings;ammonium molybdate;drought stress;LOS5/ABA3;drought resistance
PDF (615KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 郜怀峰, 张亚飞, 王国栋, 孙希武, 贺月, 彭福田, 肖元松. 钼在桃树干旱胁迫响应中的作用解析. 植物学报, 2019, 54(2): 227-236 doi:10.11983/CBB18142 Gao Huaifeng, Zhang Yafei, Wang Guodong, Sun Xiwu, He Yue, Peng Futian, Xiao Yuansong. The Effect of Molybdenum on Drought Stress Response in Peach. Chinese Bulletin of Botany, 2019, 54(2): 227-236 doi:10.11983/CBB18142
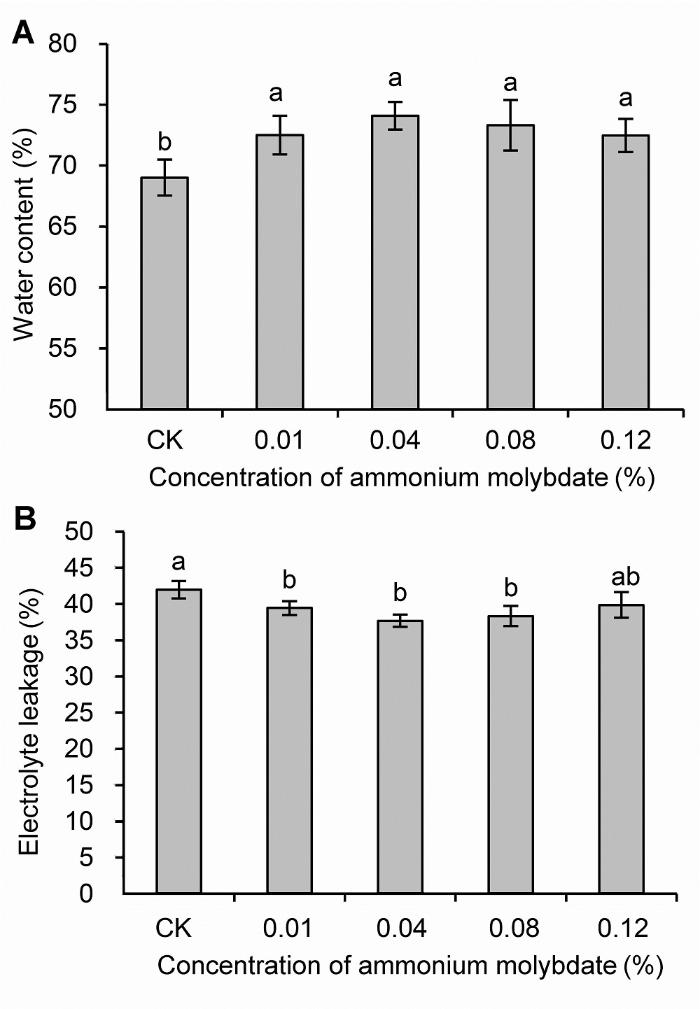
不同小写字母表示处理间差异达0.05显著水平。 Figure 1Water content (A) and electrolyte leakage (B) of peach seedlings treated with different concentrations of ammonium molybdate under drought stress
Different lowercase letters above the bars indicate significant differences at 0.05 level.
不同小写字母表示处理间差异达0.05显著水平。 Figure 2Chlorophyll (A) and proline (B) content of peach seedlings treated with different concentrations of ammonium molybdate under drought stress
Different lowercase letters above the bars indicate significant differences at 0.05 level.
不同小写字母表示处理间差异达0.05显著水平。 Figure 3The SPAD (A), net photosynthetic rate (B), transportation rate (C) and water use efficiency (D) of peach seedlings treated with ammonium molybdate under drought stress
Different lowercase letters above the bars indicate significant differences at 0.05 level.
CK: 正常条件+清水; T1: 正常条件+0.04%钼酸铵; T2: 中度干旱胁迫+清水; T3: 中度干旱胁迫+0.04%钼酸铵; T4: 重度干旱胁迫+清水; T5: 重度干旱胁迫+0.04%钼酸铵。不同小写字母表示处理间差异达0.05显著水平。 Figure 4The changes of the quality percentage of detached leaves of peach
CK: Normal conditions+water; T1: Normal conditions+0.04% ammonium molybdate; T2: Medium drought stress+water; T3: Medium drought stress+0.04% ammonium molybdate; T4: Severe drought stress+water; T5: Severe drought stress+ 0.04% ammonium molybdate. Different lowercase letters above the bars indicate significant differences at 0.05 level.
MDA: 丙二醛; CAT: 过氧化氢酶; POD: 过氧化物酶; SOD: 超氧化物歧化酶。不同小写字母表示处理间差异达0.05显著水平。 Figure 5The effect of ammonium molybdate on the MDA content (A) and the activity of CAT (B), POD (C) and SOD (D) of peach seedlings
MDA: Malondialdehyde; CAT: Catalase; POD: Peroxidase; SOD: Superoxide dismutase. Different lowercase letters above the bars indicate significant differences at 0.05 level.
(A) 正常条件; (B) 中度干旱胁迫; (C) 重度干旱胁迫。不同小写字母表示各处理间差异达0.05显著水平 Figure 7The expression of LOS5/ABA3 in leaves of peach seedlings under different drought stresses
(A) Normal conditions; (B) Medium drought stress; (C) Severe drought stress. Different lowercase letters above the bars indicate significant differences at 0.05 level.
2.8 讨论
钼作为植物体必需的微量元素, 能够调控植物的碳氮代谢及激素合成, 但钼只有结合钼辅因子后才能发挥其生物有效性。LOS5/ABA3作为钼辅因子硫化酶的编码基因, 不仅参与调控AO的活性及激素合成, 还进一步调控了植物的抗氧化防御能力及其它抗性基因的表达, 是ABA生物合成、胁迫诱导基因表达和胁迫耐受性的关键调节因子(Xiong and Zhu, 2001)。植株敲除LOS5/ABA3基因后对逆境胁迫的敏感性增强, 再次证实该基因与逆境胁迫有关(宋哲等, 2008)。本研究表明, 干旱胁迫下, 0.04%钼酸铵处理上调了LOS5/ABA3的表达水平, 且随着干旱胁迫程度的增强, 上调幅度更加显著。这可能是由于在干旱等非生物胁迫下, 植物启动了一系列复杂的信号网络调控抗旱基因及蛋白质的表达, 进而影响植物体内各种代谢过程, 最终增强了植物的抗逆性(武松伟等, 2016)。有研究表明, LOS5/ABA3能够提高脯氨酸合成酶基因(P5CS1)在转基因植物中的表达量(Yue et al., 2011; Li et al., 2013), 通过间接调控脯氨酸合成增强植物的抗旱能力。我们进一步研究发现, 干旱胁迫下0.04%钼酸铵处理提高了叶片中的脯氨酸含量, 而脯氨酸含量升高可增强细胞渗透势, 有利于植株在胁迫下维持细胞正常的生理活动, 是植物对逆境条件的一种适应性反应。
BarkerR, DaweD, TuongTP, BhuiyanSI, GuerraLC ( 1998). The outlook for water resources in the Year 2020: challenges for research on water management in rice production International Rice Commission Newsletter 49, 7-21. [本文引用: 1]
GarberoM, PedranzaniH, ZirulnikF, MolinaA, Pérez- ChacaMV, ViglioccoA, AbdalaG ( 2011). Short-term cold stress in two cultivars of Digitaria eriantha : effects on stress-related hormones and antioxidant defense system Acta Physiol Plant 33, 497-507.
HanksRJ ( 1983). Yield and water-use relationships: an overview. In: Taylor HM, Jordan WR, Sinclair TR, eds. Limitations to Efficient Water Use in Crop Production Ma- dison: American Society of Agronomy. pp. 393-411. [本文引用: 2]
MahajanS, TutejaN ( 2005). Cold, salinity and drought stres- ses: an overview Arch Biochem Biophys 444, 139-158. DOI:10.1016/j.abb.2005.10.018URL [本文引用: 1]
PatnaitkD, KhuranaP ( 2002). Wheat biotechnology: a minireview Electron J Biotechnol 4, 74-102. [本文引用: 1]
SunXC, HuCX, TanQL, LiuJS, LiuHE ( 2009). Effects of molybdenum on expression of cold-responsive genes in abscisic acid (ABA)-dependent and ABA-independent pathways in winter wheat under low-temperature stress Ann Bot 104, 345-356. DOI:10.1093/aob/mcp133URL [本文引用: 1]
WilkinsonS, DaviesWJ ( 2002). ABA-based chemical signaling: the co-ordination of responses to stress in plants Plant Cell Environ 25, 195-210. DOI:10.1046/j.0016-8025.2001.00824.xURL [本文引用: 1]
WuSW, HuCX, TanQL, NieZJ, SunXC ( 2014). Effects of molybdenum on water utilization, antioxidative defense system and osmotic-adjustment ability in winter wheat (Triticum aestivum) under drought stress Plant Physiol Biochem 83, 365-374. [本文引用: 1]
XiongLM, IshitaniM, LeeH, ZhuJK ( 2001). The Arabidopsis LOS5/ABA3 locus encodes a molybdenum cof- actor sulfurase and modulates cold stress- and osmotic stress-responsive gene expression Plant Cell 13, 2063-2084. [本文引用: 1]
Short-term cold stress in two cultivars of Digitaria eriantha : effects on stress-related hormones and antioxidant defense system 0 2011
Yield and water-use relationships: an overview. In: Taylor HM, Jordan WR, Sinclair TR, eds. Limitations to Efficient Water Use in Crop Production 2 1983
... 钼是高等植物正常生长发育和生理代谢必需的微量元素.前人研究表明, 施钼能够提高小麦(Triti- cum aestivum)叶片脱落酸(ABA)的含量, 影响植物的抗逆性(Sun et al., 2009), 但钼是否影响桃树的抗逆性尚未见报道.钼能通过含钼酶调控植物体内的碳、氮、激素和活性氧代谢等多种生理过程(武松伟等, 2016), 对植物的生长发育不可或缺.缺钼会使植物的叶绿素含量明显减少且结构异常, 导致叶片发黄, 光合速率降低.钼也与植物的非生物胁迫(如低温(孙学成, 2006)、干旱(Wu et al., 2014)和盐害(Zhang et al., 2012)等)抗性有关.钼不仅能通过含钼酶调控ABA和IAA的合成, 还能调节钼酶(孙学成, 2006)和抗氧化酶活性(Ma et al., 2012)及渗透调节物质的含量(Mahajan and Tuteja, 2005), 从而影响植物的抗性.在生物体内, 钼单独存在时并不能发挥作用, 只有形成钼辅因子(molybdenum cofactor, Moco)结合到钼酶上其才具有生物活性(Hille et al., 2014; Atwal and Scaglia, 2016).拟南芥(Arabidopsis thaliana)中, LOS5/ABA3编码钼辅因子硫化酶.硫化酶催化MoCo硫酸化, MoCo是植物中ABA生物合成途径最后一步起关键作用的醛氧化酶(aldehydeoxidases, AO)所必需的辅助因子(Li et al., 2013).ABA在干旱胁迫下的营养组织中产生, 是植物干旱胁迫响应的必须信使(Wilkinson and Davies, 2002). ...
Expression of an Arabidopsis molybdenum cofactor sulphurase gene in soybean enhances drought tolerance and increases yield under field conditions 2 2013
... 近年干旱天气时有发生, 对桃生产造成了不利影响, 因此降低干旱伤害是提高桃产量的重要因素.研究表明, 钼可增强玉米(Zea mays) (Lu et al., 2013)、大豆(Glycine max) (Li et al., 2013)以及野棉花(Anemone vitifolia) (Yue et al., 2012)等作物的抗旱性, 从而提高作物产量.但钼能否调控钼辅因子硫化酶编码基因的表达进而提高桃树的抗旱性, 缓解干旱胁迫伤害, 至今未见报道.因此, 本研究探讨了干旱胁迫下, 钼酸铵对桃叶片钼辅因子硫化酶编码基因表达, 及叶片含水量、光合特性、细胞膜氧化伤害、脱落酸与脯氨酸含量的影响, 以期为生产中桃树抗干旱胁迫提供参考. ...
... 钼是高等植物正常生长发育和生理代谢必需的微量元素.前人研究表明, 施钼能够提高小麦(Triti- cum aestivum)叶片脱落酸(ABA)的含量, 影响植物的抗逆性(Sun et al., 2009), 但钼是否影响桃树的抗逆性尚未见报道.钼能通过含钼酶调控植物体内的碳、氮、激素和活性氧代谢等多种生理过程(武松伟等, 2016), 对植物的生长发育不可或缺.缺钼会使植物的叶绿素含量明显减少且结构异常, 导致叶片发黄, 光合速率降低.钼也与植物的非生物胁迫(如低温(孙学成, 2006)、干旱(Wu et al., 2014)和盐害(Zhang et al., 2012)等)抗性有关.钼不仅能通过含钼酶调控ABA和IAA的合成, 还能调节钼酶(孙学成, 2006)和抗氧化酶活性(Ma et al., 2012)及渗透调节物质的含量(Mahajan and Tuteja, 2005), 从而影响植物的抗性.在生物体内, 钼单独存在时并不能发挥作用, 只有形成钼辅因子(molybdenum cofactor, Moco)结合到钼酶上其才具有生物活性(Hille et al., 2014; Atwal and Scaglia, 2016).拟南芥(Arabidopsis thaliana)中, LOS5/ABA3编码钼辅因子硫化酶.硫化酶催化MoCo硫酸化, MoCo是植物中ABA生物合成途径最后一步起关键作用的醛氧化酶(aldehydeoxidases, AO)所必需的辅助因子(Li et al., 2013).ABA在干旱胁迫下的营养组织中产生, 是植物干旱胁迫响应的必须信使(Wilkinson and Davies, 2002). ...
Effects of molybdenum on expression of cold-responsive genes in abscisic acid (ABA)-dependent and ABA-independent pathways in winter wheat under low-temperature stress 1 2009
... 钼是高等植物正常生长发育和生理代谢必需的微量元素.前人研究表明, 施钼能够提高小麦(Triti- cum aestivum)叶片脱落酸(ABA)的含量, 影响植物的抗逆性(Sun et al., 2009), 但钼是否影响桃树的抗逆性尚未见报道.钼能通过含钼酶调控植物体内的碳、氮、激素和活性氧代谢等多种生理过程(武松伟等, 2016), 对植物的生长发育不可或缺.缺钼会使植物的叶绿素含量明显减少且结构异常, 导致叶片发黄, 光合速率降低.钼也与植物的非生物胁迫(如低温(孙学成, 2006)、干旱(Wu et al., 2014)和盐害(Zhang et al., 2012)等)抗性有关.钼不仅能通过含钼酶调控ABA和IAA的合成, 还能调节钼酶(孙学成, 2006)和抗氧化酶活性(Ma et al., 2012)及渗透调节物质的含量(Mahajan and Tuteja, 2005), 从而影响植物的抗性.在生物体内, 钼单独存在时并不能发挥作用, 只有形成钼辅因子(molybdenum cofactor, Moco)结合到钼酶上其才具有生物活性(Hille et al., 2014; Atwal and Scaglia, 2016).拟南芥(Arabidopsis thaliana)中, LOS5/ABA3编码钼辅因子硫化酶.硫化酶催化MoCo硫酸化, MoCo是植物中ABA生物合成途径最后一步起关键作用的醛氧化酶(aldehydeoxidases, AO)所必需的辅助因子(Li et al., 2013).ABA在干旱胁迫下的营养组织中产生, 是植物干旱胁迫响应的必须信使(Wilkinson and Davies, 2002). ...
ABA-based chemical signaling: the co-ordination of responses to stress in plants 1 2002
... 钼是高等植物正常生长发育和生理代谢必需的微量元素.前人研究表明, 施钼能够提高小麦(Triti- cum aestivum)叶片脱落酸(ABA)的含量, 影响植物的抗逆性(Sun et al., 2009), 但钼是否影响桃树的抗逆性尚未见报道.钼能通过含钼酶调控植物体内的碳、氮、激素和活性氧代谢等多种生理过程(武松伟等, 2016), 对植物的生长发育不可或缺.缺钼会使植物的叶绿素含量明显减少且结构异常, 导致叶片发黄, 光合速率降低.钼也与植物的非生物胁迫(如低温(孙学成, 2006)、干旱(Wu et al., 2014)和盐害(Zhang et al., 2012)等)抗性有关.钼不仅能通过含钼酶调控ABA和IAA的合成, 还能调节钼酶(孙学成, 2006)和抗氧化酶活性(Ma et al., 2012)及渗透调节物质的含量(Mahajan and Tuteja, 2005), 从而影响植物的抗性.在生物体内, 钼单独存在时并不能发挥作用, 只有形成钼辅因子(molybdenum cofactor, Moco)结合到钼酶上其才具有生物活性(Hille et al., 2014; Atwal and Scaglia, 2016).拟南芥(Arabidopsis thaliana)中, LOS5/ABA3编码钼辅因子硫化酶.硫化酶催化MoCo硫酸化, MoCo是植物中ABA生物合成途径最后一步起关键作用的醛氧化酶(aldehydeoxidases, AO)所必需的辅助因子(Li et al., 2013).ABA在干旱胁迫下的营养组织中产生, 是植物干旱胁迫响应的必须信使(Wilkinson and Davies, 2002). ...
Effects of molybdenum on water utilization, antioxidative defense system and osmotic-adjustment ability in winter wheat (Triticum aestivum) under drought stress 1 2014
The Arabidopsis LOS5/ABA3 locus encodes a molybdenum cof- actor sulfurase and modulates cold stress- and osmotic stress-responsive gene expression 1 2001
... 钼作为植物体必需的微量元素, 能够调控植物的碳氮代谢及激素合成, 但钼只有结合钼辅因子后才能发挥其生物有效性.LOS5/ABA3作为钼辅因子硫化酶的编码基因, 不仅参与调控AO的活性及激素合成, 还进一步调控了植物的抗氧化防御能力及其它抗性基因的表达, 是ABA生物合成、胁迫诱导基因表达和胁迫耐受性的关键调节因子(Xiong and Zhu, 2001).植株敲除LOS5/ABA3基因后对逆境胁迫的敏感性增强, 再次证实该基因与逆境胁迫有关(宋哲等, 2008).本研究表明, 干旱胁迫下, 0.04%钼酸铵处理上调了LOS5/ABA3的表达水平, 且随着干旱胁迫程度的增强, 上调幅度更加显著.这可能是由于在干旱等非生物胁迫下, 植物启动了一系列复杂的信号网络调控抗旱基因及蛋白质的表达, 进而影响植物体内各种代谢过程, 最终增强了植物的抗逆性(武松伟等, 2016).有研究表明, LOS5/ABA3能够提高脯氨酸合成酶基因(P5CS1)在转基因植物中的表达量(Yue et al., 2011; Li et al., 2013), 通过间接调控脯氨酸合成增强植物的抗旱能力.我们进一步研究发现, 干旱胁迫下0.04%钼酸铵处理提高了叶片中的脯氨酸含量, 而脯氨酸含量升高可增强细胞渗透势, 有利于植株在胁迫下维持细胞正常的生理活动, 是植物对逆境条件的一种适应性反应. ...
Abiotic stress signal transduction in plants: molecular and genetic perspectives 1 2001
... 近年干旱天气时有发生, 对桃生产造成了不利影响, 因此降低干旱伤害是提高桃产量的重要因素.研究表明, 钼可增强玉米(Zea mays) (Lu et al., 2013)、大豆(Glycine max) (Li et al., 2013)以及野棉花(Anemone vitifolia) (Yue et al., 2012)等作物的抗旱性, 从而提高作物产量.但钼能否调控钼辅因子硫化酶编码基因的表达进而提高桃树的抗旱性, 缓解干旱胁迫伤害, 至今未见报道.因此, 本研究探讨了干旱胁迫下, 钼酸铵对桃叶片钼辅因子硫化酶编码基因表达, 及叶片含水量、光合特性、细胞膜氧化伤害、脱落酸与脯氨酸含量的影响, 以期为生产中桃树抗干旱胁迫提供参考. ...
Overexpression of the AtLOS5 gene increased abscisic acid level and drought tolerance in transgenic cotton 1 2012
... 钼作为植物体必需的微量元素, 能够调控植物的碳氮代谢及激素合成, 但钼只有结合钼辅因子后才能发挥其生物有效性.LOS5/ABA3作为钼辅因子硫化酶的编码基因, 不仅参与调控AO的活性及激素合成, 还进一步调控了植物的抗氧化防御能力及其它抗性基因的表达, 是ABA生物合成、胁迫诱导基因表达和胁迫耐受性的关键调节因子(Xiong and Zhu, 2001).植株敲除LOS5/ABA3基因后对逆境胁迫的敏感性增强, 再次证实该基因与逆境胁迫有关(宋哲等, 2008).本研究表明, 干旱胁迫下, 0.04%钼酸铵处理上调了LOS5/ABA3的表达水平, 且随着干旱胁迫程度的增强, 上调幅度更加显著.这可能是由于在干旱等非生物胁迫下, 植物启动了一系列复杂的信号网络调控抗旱基因及蛋白质的表达, 进而影响植物体内各种代谢过程, 最终增强了植物的抗逆性(武松伟等, 2016).有研究表明, LOS5/ABA3能够提高脯氨酸合成酶基因(P5CS1)在转基因植物中的表达量(Yue et al., 2011; Li et al., 2013), 通过间接调控脯氨酸合成增强植物的抗旱能力.我们进一步研究发现, 干旱胁迫下0.04%钼酸铵处理提高了叶片中的脯氨酸含量, 而脯氨酸含量升高可增强细胞渗透势, 有利于植株在胁迫下维持细胞正常的生理活动, 是植物对逆境条件的一种适应性反应. ...
Arabidopsis LOS5/ABA3 overexpression in transgenic tobacco( Nicotiana tabacum cv. ‘Xanthi-nc’) results in enhanced drought tolerance 1 2011
... 钼是高等植物正常生长发育和生理代谢必需的微量元素.前人研究表明, 施钼能够提高小麦(Triti- cum aestivum)叶片脱落酸(ABA)的含量, 影响植物的抗逆性(Sun et al., 2009), 但钼是否影响桃树的抗逆性尚未见报道.钼能通过含钼酶调控植物体内的碳、氮、激素和活性氧代谢等多种生理过程(武松伟等, 2016), 对植物的生长发育不可或缺.缺钼会使植物的叶绿素含量明显减少且结构异常, 导致叶片发黄, 光合速率降低.钼也与植物的非生物胁迫(如低温(孙学成, 2006)、干旱(Wu et al., 2014)和盐害(Zhang et al., 2012)等)抗性有关.钼不仅能通过含钼酶调控ABA和IAA的合成, 还能调节钼酶(孙学成, 2006)和抗氧化酶活性(Ma et al., 2012)及渗透调节物质的含量(Mahajan and Tuteja, 2005), 从而影响植物的抗性.在生物体内, 钼单独存在时并不能发挥作用, 只有形成钼辅因子(molybdenum cofactor, Moco)结合到钼酶上其才具有生物活性(Hille et al., 2014; Atwal and Scaglia, 2016).拟南芥(Arabidopsis thaliana)中, LOS5/ABA3编码钼辅因子硫化酶.硫化酶催化MoCo硫酸化, MoCo是植物中ABA生物合成途径最后一步起关键作用的醛氧化酶(aldehydeoxidases, AO)所必需的辅助因子(Li et al., 2013).ABA在干旱胁迫下的营养组织中产生, 是植物干旱胁迫响应的必须信使(Wilkinson and Davies, 2002). ...
Molybdenum improves antioxidant and osmotic-adjustment ability against salt stress in Chinese cabbage ( Brassica campestris L. ssp 2 2012
 ,*, 肖元松
,*, 肖元松
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT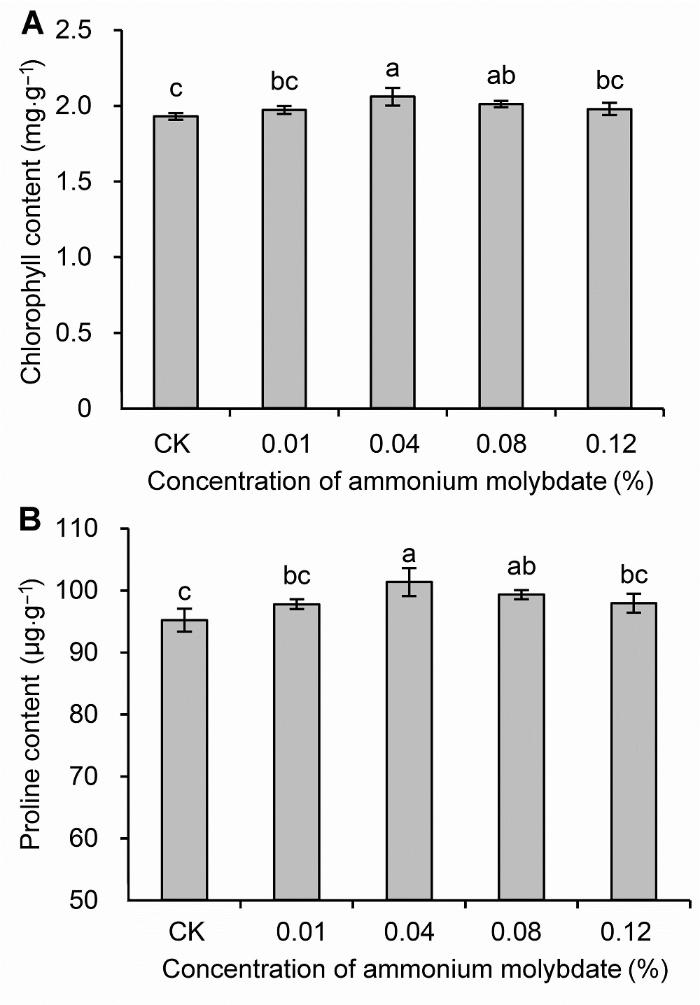 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT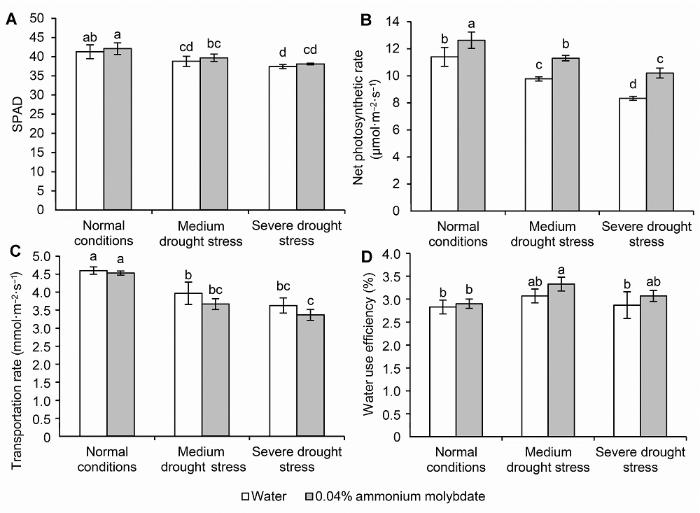 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT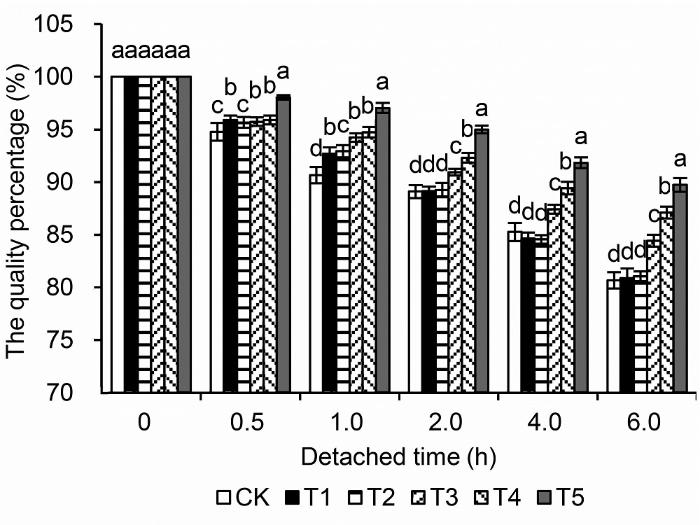 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT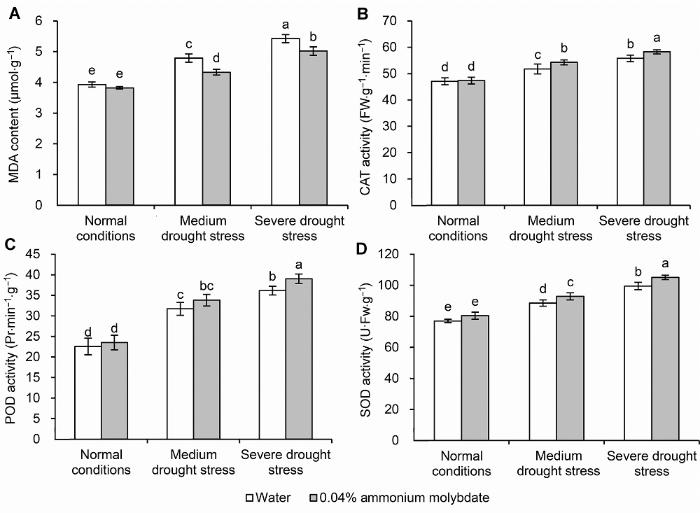 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT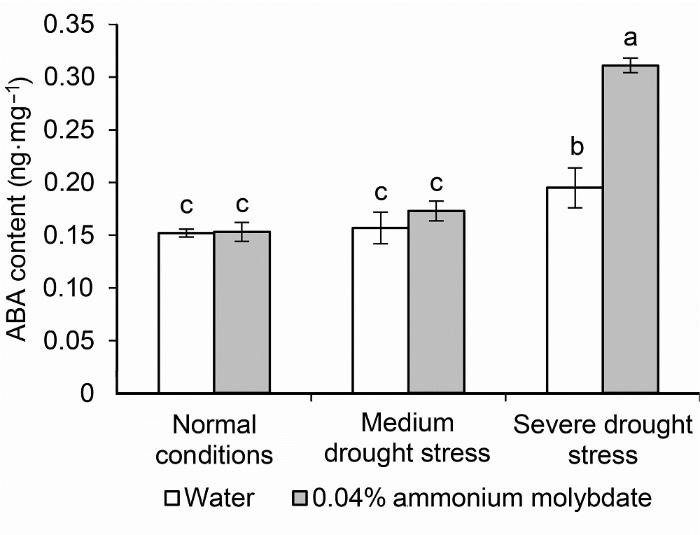 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT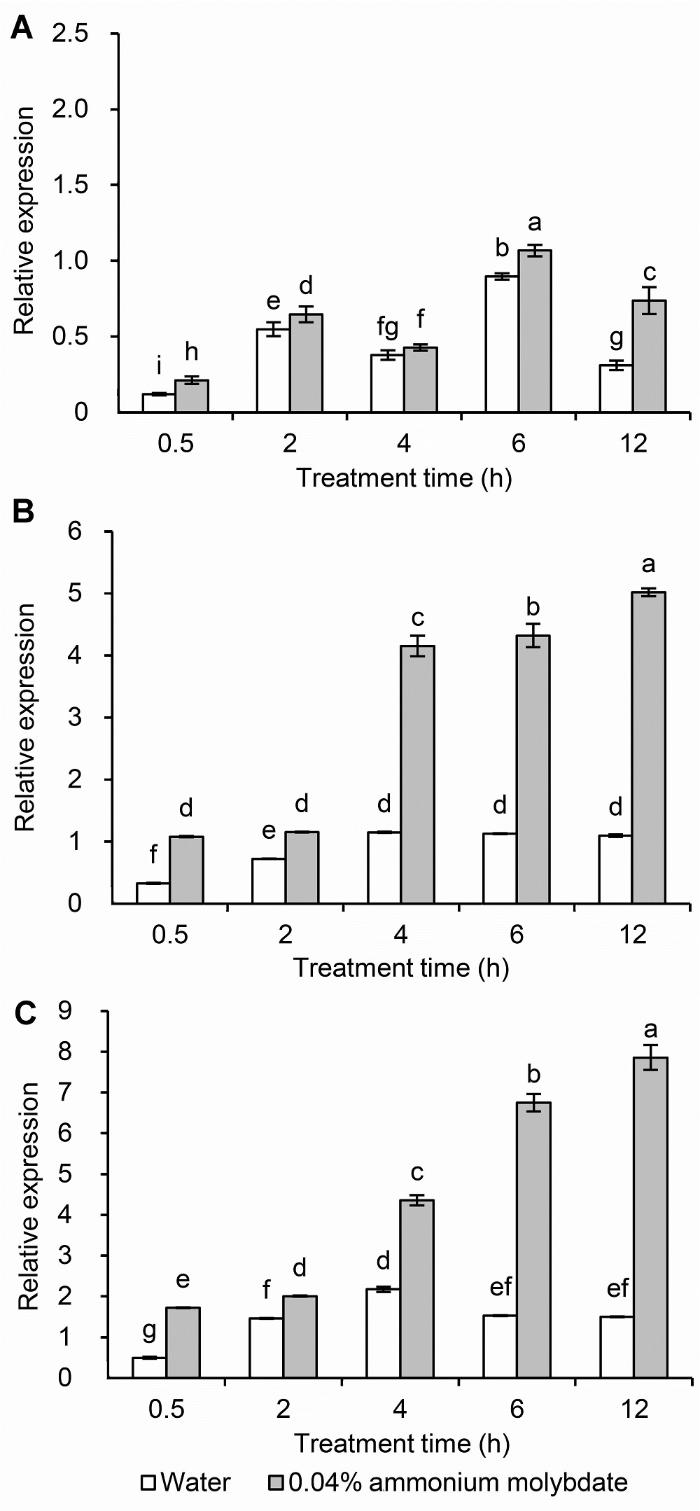 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}