Abstract: This study aimed to reveal the characteristics of photosynthetic carbon assimilation and the change regulation in different internodes of Phyllostachys edulis stems during their rapid growth stage. We analyzed the photosynthetic pigment content and activities of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco), phosphoenolpyruvate carboxylase (PEPC), NADP-malate dehydrogenase (NADP-MDH), NADP-malic enzyme (NADP-ME), phosphoenolpyruvate carboxykinase (PEPCK) and pyruvate, and orthophosphate dikinase (PPDK) in stems with different internodes. With increasing internode number of P. edulis stems, the content of chlorophyll a, chlorophyll b and carotenoid was all significantly reduced and the ratio of chlorophyll a/b showed a gradual upward trend. The activities of Rubisco, PEPC and PPDK in stems of P. edulis decreased significantly from the 1st to the 10th internode, then the rate of decrease gradually slowed down. The activities of NADP-ME declined significantly from the 1st to the 13th internode; after the 13th internode, the enzyme activities tended to be stable. The activities of NADP-MDH decreased markedly from 1st to the 25th internode of stems. The ratio of PEPC/Rubisco increased gradually and the range was between 18.37 and 65.09, which was significantly larger than that in typical C3 plants. The research showed that the assimilating capacity with carbon differed in different internodes, and the middle and lower stems grew relatively fast. The result of many C4 enzymes and activities being significantly high provided strong evidence for the existence of a C4 photosynthetic pathway in the stems of P. edulis.
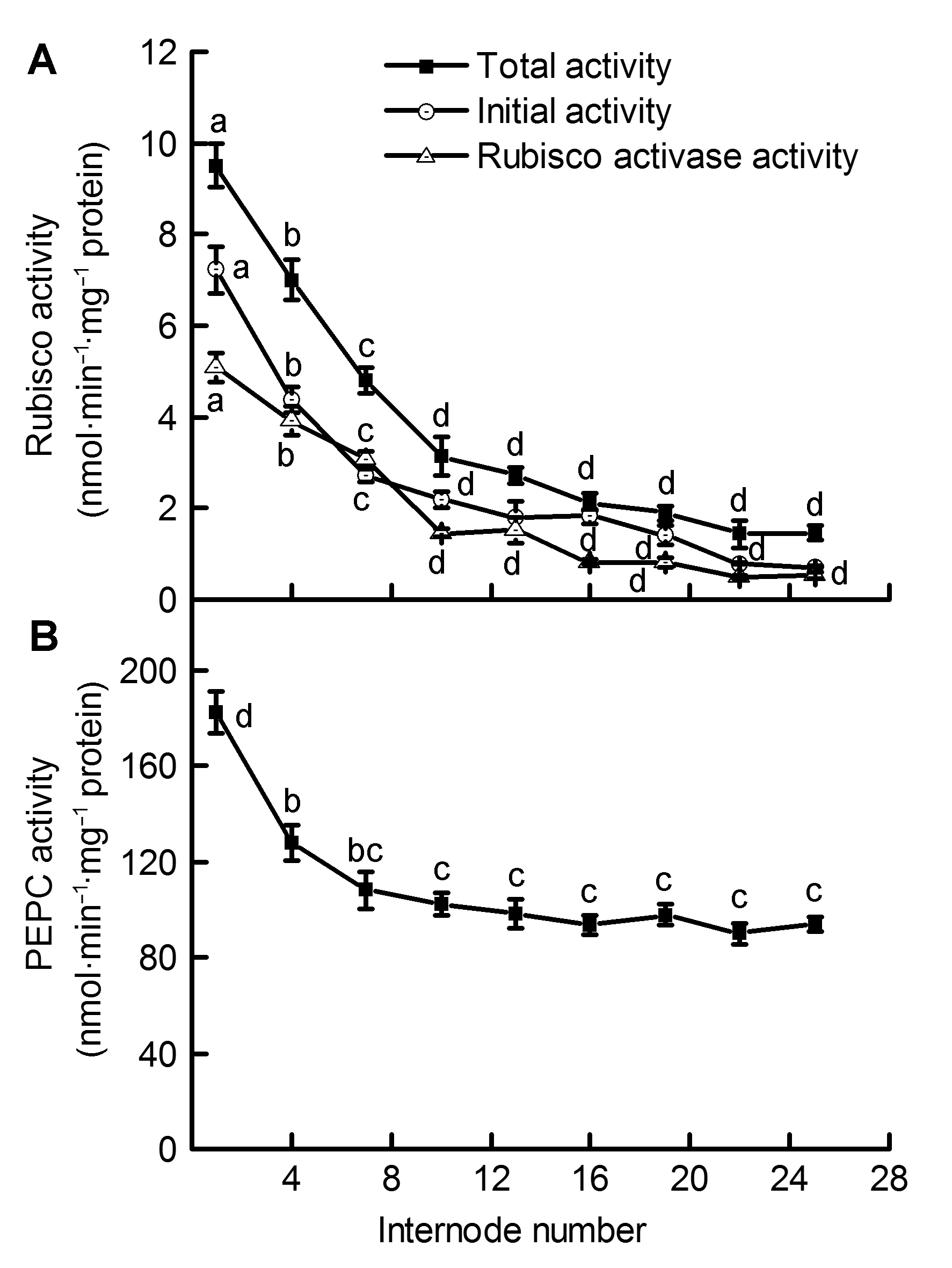
图 1 毛竹茎秆不同节间内核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)活性(A)和磷酸烯醇式丙酮酸羧化酶(PEPC)活性(B)的差异(平均值±标准差) Figure 1 Differences of Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) activity (A) and Phosphoenolpyruvate carboxylase (PEPC) activity (B) in the Phyllostachys edulis stems of different internodes (means±SD)
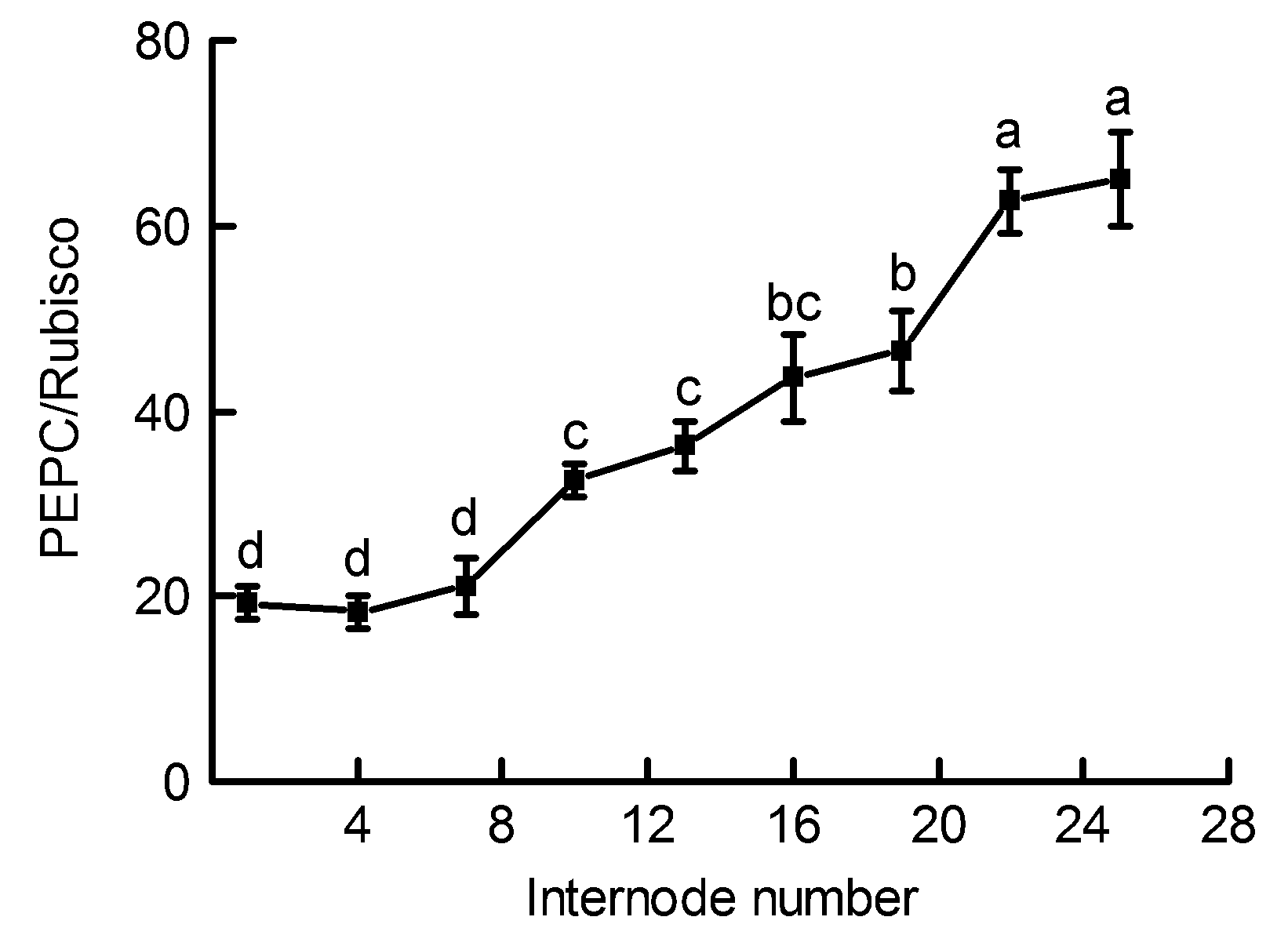
2.7 不同节间PEPC/Rubisco活性比值的差异毛竹茎秆不同节间内PEPC/Rubisco比值是评价2种光合酶在茎秆不同部位表达情况的重要指标。由图3可以看出, 随着毛竹茎秆节间的升高, 其呈逐渐上升的变化趋势。第1-7节间内PEPC/Rubisco比值基本维持在19.70左右, 之后逐渐升高, 第25节间的活性比值与第7节间相比升高了2.09倍(P<0.05)。 图 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-6-773/img_3.png<b>图 3</b> 毛竹茎秆不同节间内磷酸烯醇式丙酮酸羧化酶和核酮糖-1,5-二磷酸羧化酶/加氧酶(PEPC/Rubisco)比值的差异(平均值± 标准差)<br/><b>Figure 3</b> Changes of the ratios of Phosphoenolpyruvate carboxylase/Ribulose-1,5-bisphosphate carboxylase/oxygen- ase (PEPC/Rubisco) in the <i>Phyllostachys edulis </i>stems of different internodes (means±SD) Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-6-773/img_3.png<b>图 3</b> 毛竹茎秆不同节间内磷酸烯醇式丙酮酸羧化酶和核酮糖-1,5-二磷酸羧化酶/加氧酶(PEPC/Rubisco)比值的差异(平均值± 标准差)<br/><b>Figure 3</b> Changes of the ratios of Phosphoenolpyruvate carboxylase/Ribulose-1,5-bisphosphate carboxylase/oxygen- ase (PEPC/Rubisco) in the <i>Phyllostachys edulis </i>stems of different internodes (means±SD)
图 3 毛竹茎秆不同节间内磷酸烯醇式丙酮酸羧化酶和核酮糖-1,5-二磷酸羧化酶/加氧酶(PEPC/Rubisco)比值的差异(平均值± 标准差) Figure 3 Changes of the ratios of Phosphoenolpyruvate carboxylase/Ribulose-1,5-bisphosphate carboxylase/oxygen- ase (PEPC/Rubisco) in the Phyllostachys edulis stems of different internodes (means±SD)
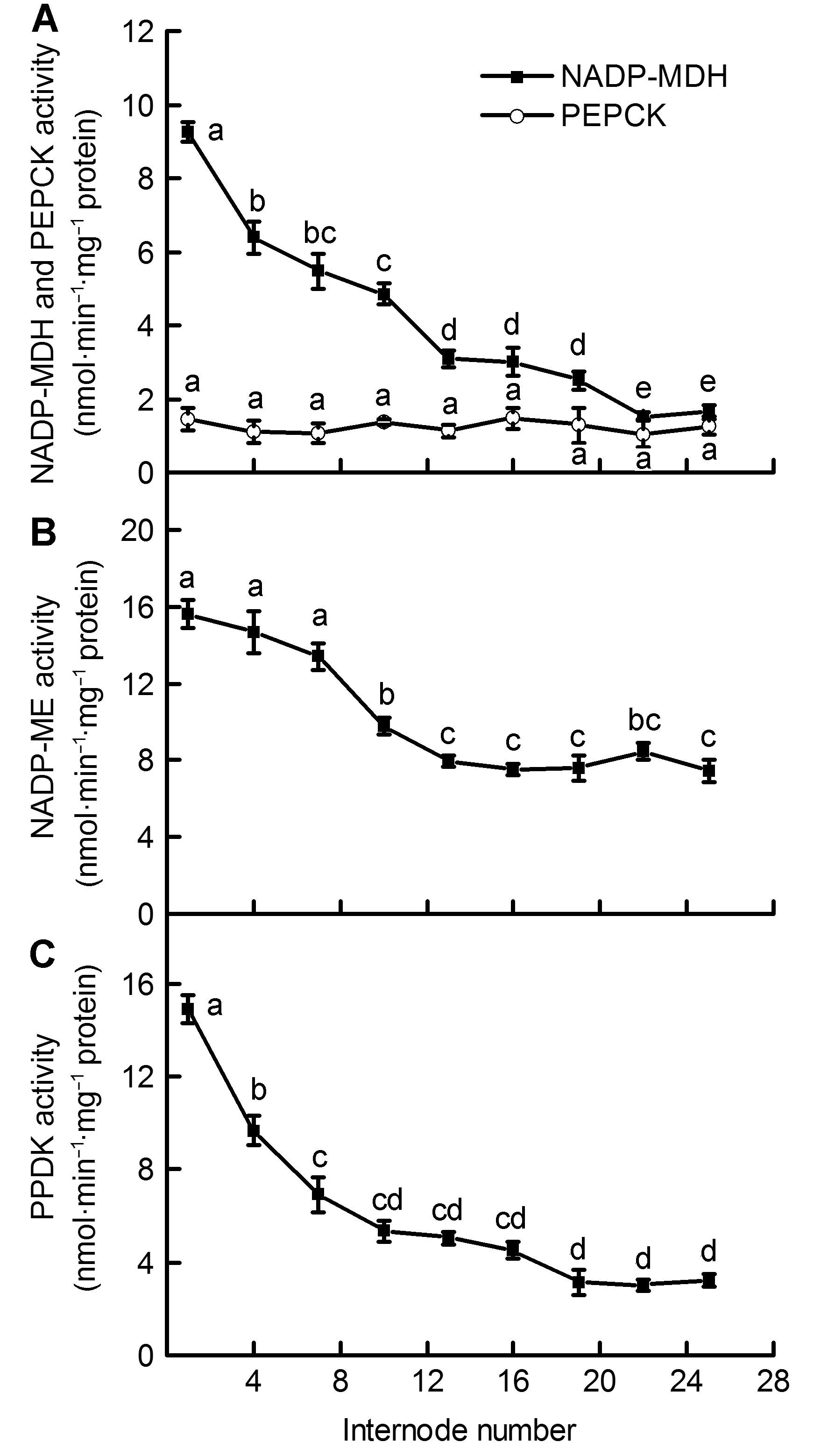
2.8 讨论许多植物的非叶器官包括茎秆、枝条和根系等绿色组织中含有高浓度的叶绿素。Dima等(2006)测定了20种植物枝条内的叶绿素含量, 得出其皮层叶绿素含量的数值范围在19-94 mg·m-2之间。研究表明, 葡萄(Vitis vinifera)绿色茎秆中叶绿素含量在58-303 μg·g-1 (FW)之间(Tikhonov et al., 2017); 旱柳(Salix ma- tsudana)皮层中叶绿素含量是其叶片的10%-13.7% (张金尧等, 2014)。植物茎秆中的叶绿素含量不仅与其植株的生长季节和生长年限有关(Damesin, 2003; Bloemen et al., 2016), 还与其接受的光照时间、光照面积和光质有关(Berveiller and Damesin, 2008; 高苏娟等, 2009; 徐超华等, 2013)。对绿豆(Vigna radiata)幼苗脱黄化初期叶片的研究表明, 2种主要光合色素(叶绿素a和叶绿素b)含量随光照时间的增加均逐渐上升, 叶绿素a/b比值则逐渐降低(张汝民, 2005)。本研究中, 毛竹茎秆中部及下部发育较为成熟, 其表面包被的竹箨已逐渐脱落, 茎秆外层由上至下接受的光照面积和光照时间逐渐增多, 进而导致茎秆皮层绿色组织中光合色素含量逐渐升高, 叶绿素a/b逐渐降低(表1)。以上结果表明, 毛竹在未萌发枝叶的快速生长期内, 茎秆中、下部较高含量的叶绿素及其较强的光能捕获能力可能是维持毛竹笋竹快速生长的原因之一。 Rubisco是植物碳同化过程中的关键酶, 其活性的高低对植物光合速率的快慢起决定性作用(Suzuki and Makino, 2012; Sheth and Thaker, 2014; Sun et al., 2014)。Hrstka等(2012)对欧洲山毛榉和挪威云杉(Picea abies)的研究表明, CO2浓度变化对Rubisco含量和活性存在较大影响。Cen和Sage (2005)发现温度升高能显著提高番薯(Ipomoea batatas)叶片中的Rubisco活性, 但较高或较低温度下活化状态的Rubisco所占比例较小。对黄瓜(Cucumis sativus)植株的研究表明, 弱光处理不仅能使其叶片中Rubisco活性下降, 也会在一定程度上影响相关基因的表达水平(Kosvancova et al., 2009; 孙建磊等, 2017)。本研究表明, 毛竹茎秆中、下部节间表面接受光照较多, 导致Rubisco活性明显高于上部节间, 这些部位的生长发育则相对较快; 茎秆上部节间接受光照较少, Rubisco活性维持在较低水平, 这些节间的生长则相对较为缓慢(图1)。在茎秆外层包被的竹箨脱落前后, Rubisco的活性变化明显加快, 这进一步说明光照面积的增加不仅影响光合色素的合成, 还促进Rubisco活性的升高, 进而促使毛竹茎秆快速生长。这与Wang等(2014)对不同光强下番薯叶片中Rubisco活性变化的研究结果相一致。植物中的RCA能够起到活化Rubisco的作用, 处于活化状态的Rubisco才具有较强的CO2同化能力(Portis, 2003; 姜振升等, 2010; Parry et al., 2013)。在毛竹的快速生长过程中, RCA活性与Rubisco活性随节间上升而表现出相似的变化趋势, 这反映出茎秆中RCA可能对Rubisco发挥一定的调控作用, 促使Rubisco活性保持在最佳状态, 从而促进茎秆快速生长和成熟。 在C4植物CO2浓缩过程的初始阶段, PEPC能够催化CO2固定的最初反应。研究表明, PEPC在很多C3植物的茎秆和枝条等非同化器官中具有较高活性(Berveiller and Damesin, 2008; 王莹等, 2011)。植物茎秆中的PEPC能够固定其自身呼吸作用产生的部分CO2, 并将其运输到C3循环中进行羧化反应(郝迺斌等, 1991)。植物C4途径中, NADP-MDH和NADP- ME能够经多重反应将草酰乙酸(OAA)催化形成C4酸, 并将其水解释放CO2, 进入C3循环中被Rubisco所固定(Carnal et al., 1993; Sage et al., 2012)。PEPCK是1种典型的C4酶, 它在维管束鞘细胞中能够催化OAA脱羧并释放CO2, 从而对许多植物的光合碳同化过程起到重要调节作用(Leegood and Osmond, 1990; Chao et al., 2014)。PPDK是C4途径的专一性限速酶, 其活性受光照、PH及二价阳离子浓度等因素影响(Chastain et al., 2011), 它能催化ATP和丙酮酸等物质形成CO2同化过程中的最初受体PEP, 从而对C4途径中CO2初始固定过程进行调节(Leegood, 2013; Chen et al., 2014)。C3植物中PPDK表达量较低, 目前对其各项功能的研究相对较少(Burnell and Chastain, 2006)。PEPC、NADP-MDH、PEPCK、NADP-ME和PPDK等一系列C4途径酶广泛存在于C3植物非同化器官茎秆、枝条和叶柄等组织中(Hibberd and Quick, 2002; Kocurek and Pilarski, 2011)。在C4植物中, 根据CO2固定过程中所用脱羧酶的不同, 人们将其划分为NADP-ME、NAD-ME和PEPCK三种亚型(Carnal et al., 1993; Muhaidat et al., 2007)。本研究表明, 毛竹茎秆内PEPCK活性较低且前后无明显变化, 而NADP-ME和NADP-MDH亚型活性较高且在不同节间存在规律性变化, 说明毛竹茎秆内主要存在的可能为NADP-ME亚型; 茎秆内存在PEPC、NADP- MDH、NADP-ME和PPDK等多种C4酶, 且这些酶的活性均随茎秆节间升高而逐渐下降, 这与Rubisco活性变化之间具有较为明显的一致性, 也表明此时期茎秆不同节间内可能存在C4光合酶相关的生化过程。 目前, 对植物非同化器官中碳同化类型的研究仍在进行, 但已有较多研究表明, 部分植物的非同化器官如茎秆和枝条等可能存在C4光合途径(Berveiller et al., 2007; 王文杰等, 2010)。对C3植物芹菜(Apium graveolens)和烟草的研究表明, 茎秆和叶柄中存在多种活性较高的C4酶, 且其根系能将呼吸作用产生的CO2初步固定后转运到木质部周围的鞘细胞中, 发生类似C4途径中的CO2羧化与还原反应, 并产生C4光合作用的四碳产物(Hibberd and Quick, 2002)。前人对C4植物的大量解剖实验表明, 其维管束周围的细胞能够排列成不同种类的“花环结构”(Sage et al., 2007; Ocampo and Columbus, 2010), 其中含有叶绿素并具有累积CO2的功能。王星星等(2012)对毛竹茎秆结构的观察表明, 其维管束周围具有类似C4植物的“花环结构”。Cui等(2012)对毛竹茎秆的解剖分析发现, 下部的发育和成熟要早于中和上部。我们对毛竹茎秆不同节间内Rubisco和多种C4酶的研究是对以往研究的重要补充: 茎秆不同节间内多种光合酶活性存在显著差异, 中部和下部节间接受光照较多, Rubisco和多种C4酶活性较高; PEPC/Rubisco比值随茎秆节间的逐渐升高呈不断上升趋势, 其比值范围在18.37-65.09之间(图3), 这远高于典型C3植物中的活性比值。以上结果表明, 毛竹茎秆内可能存在C4光合途径, 在光照条件下呼吸作用产生的大量CO2被细胞内较高活性的PEPC所固定, 进入卡尔文循环进行光合作用, 为茎秆的快速生长提供碳源。值得注意的是, C4光合途径中PEPC只起到固定CO2的作用, 而最终合成有机物以供植物利用的过程是由C3途径中的Rubisco来完成(龚春梅等, 2009)。本研究中, 毛竹内PEPC/Rubisco比值在上部各节间较大, 而Rubisco活性在中和下部节间内较高, 这表明上部茎秆中的PEPC固定CO2后的部分产物可能贮存在上部节间, 并未被直接同化利用; 也可能被运输到茎秆中部或下部, 在中、下部茎秆内经较高活性的Rubisco羧化和还原来实现对CO2的同化, 从而维持中、下部节间的快速生长和发育。 综上所述, 处于快速生长阶段的毛竹茎秆内存在较高活性的Rubisco和多种C4酶, 这为此时期茎秆中存在C4光合途径提供了有力证据。叶绿素含量、Rubisco和多种C4酶活性随节间的升高而呈现规律性变化, 这表明毛竹茎秆不同节间的生长发育存在明显差异, 茎秆中、下部节间生长较快, 上部节间生长相对缓慢; 茎秆中具有较为完善的C4途径酶系统, 各光合酶相互协作, 共同调节茎秆中的光合碳同化过程。
The authors have declared that no competing interests exist.
陈登举, 高培军, 吴兴波, 高岩, 温国胜, 王玉魁, 高荣孚, 张汝民 (2013). 毛竹茎秆叶绿体超微结构及其发射荧光光谱特征. 植物学报 48, 635-642. DOI:10.3724/SP.J.1259.2013.00635URLTo reveal the photosynthetic characteristics of stems of P. pubescens may adapt to far-red light and coordinate the allocation of excitation energy between PSII and PSI by increasing the chlorophyll b relative content and the number of lamella as well as reducing the content of LHCI. [本文引用: 1]
王星星, 刘琳, 张洁, 王玉魁, 温国胜, 高荣孚, 高岩, 张汝民 (2012). 毛竹出笋后快速生长期内茎秆中光合色素和光合酶活性的变化. 植物生态学报 36, 456-462. DOI:10.3724/SP.J.1258.2012.00456URLAimsMethodsImportant findings plants. The content of chlorophyll a, chlorophyll b and carotenoid was significantly ( plants. These results indicated that there was C photosynthetic pathway in the stems, and the pathway may play an important role in the efficient photosynthesis and rapid growth of P. pubescens. [本文引用: 2]
徐超华, 李军营, 崔明昆, 马二登, 黄国宾, 龚明 (2013). 延长光照时间对烟草叶片生长发育及光合特性的影响. 西北植物学报 33, 763-770. DOI:10.3969/j.issn.1000-4025.2013.04.018URLUsing the tobacco variety ‘Yunyan 87’ as experimental material and natural lighting for control,the effects of supplemental lighting on leaf growth,chlorophyll content,photosynthesis,chlorophyll fluorescence,and light response curves in tobacco plants was investigated by additional night artificial lighting for 1,2 and 3 h in this study.The results showed that,as compared with the natural lighting,the treatments with supplemental lighting for 2 h obviously promoted leaf length and width,and plant height,the treatments for 1 h and 3 h showed insignificant effect.These supplemental lighting treatments from 1 to 3 h remarkably decreased specific leaf area (SLA) and increased contents of chlorophyll a,b,chlorophyll a+b and carotenoids,and these changes for 1 h and 3 h treatments were less than 2 h treatment.The treatments with supplemental lighting for 1 h and 2 h obviously raised net photosynthetic rate (),stomatal conductance(),intercellular CO concentration(),3 h treatment had little effects.The supplemental lighting treatments significantly enhanced optimal quantum yield of Φ(/),actual photochemical efficiency of PSⅡ(Φ),and photochemical quenching (),lowered non photochemical quenching (NPQ),but showed little effect on initial fluorescence intensity Fc.Maximum net photosynthetic rate () and light saturation points () were both raised,and light compensation point () was not affected obviously under these supplemental lighting treatments.These results indicated that proper supplemental lighting time could enhance the growth of tobacco leaves,accumulation of dry matter,chlorophyll content and photosynthesis,relieve the photoinhibition phenomenon,make full use of light energy,and improve the efficiency of photosynthesis in tobacco leaves. [本文引用: 1]
祖元刚, 张衷华, 王文杰, 杨逢建, 贺海升 (2006). 薇甘菊叶和茎的光合特性. 植物生态学报 30, 998-1004. URLMikania micranthKey Results) and actual photochemical efficiency of system Ⅱ (PSⅡ) of stems and leaves were positively correlated (0 .97), suggesting similar photosynthetic structure in stems and leaves. The gas exchange rate under constant conditions was stable for leaves but fluctuating for stems, possibly because of the pore factor of stems. Under the same area and saturated light (PPFD=2 000μmol·m·s), the of leaves was 42 .44 and the of stems was 30.32μmol·m·s. However, under thesame area and low light (PPFD=10μmol·m·s), the PSⅡ of leaveswas 0.69 and thePSⅡ of stems was 0.57. The electron transport rate in stems was 4.24 per unit SPAD, 2.3 times that of leaves. Actual photochemical efficiency of system Ⅱ was 0.08 per unit SPAD, 3 times that of leaves. Our research also indicated that light adaptive ability was better in stems than in leaves. Chlorophyll existed mainly in two tissues: cortex and around vascular bundles. It is possible that there are different chlorophyll functions in different tissues. Conclusions This study indicates that photosynthesis occurred in younggreen stems of M. micrantha and the instantaneous efficiency of light utilization in stems was higher than in leaves. Results were unclear regarding the function of chlorophyll in different tissues. [本文引用: 1]
[17]
AschanG, PfanzH (2003). Non-foliar photosynthesis—a strategy of additional carbon acquisition.Flora 198, 81-97. DOI:10.1078/0367-2530-00080URLIn addition to the green leaves, commonly considered as the primary sources of photosynthate production, higher plants can potentially use almost all vegetative and reproductive structures to perform photosynthetic CO 2 assimilation. Green leaves, stems and green sterile flower organs, optimized for light harvesting and photosynthetic performance, are characterized by net photosynthetic assimilation utilizing mainly the atmospheric carbon dioxide. In contrast, chlorophyll-containing bark and wood tissue, most fruit, root and fertile flower organs are principally sub-ordinated to non-photosynthetic functions, but typically perform an effective internal CO 2 recycling using the respiratory released CO 2. Non-foliar photosynthesis, either manifested as net photosynthesis or internal CO 2 refixation is regarded as an important strategy of additional carbon-acquisition. While chlorophyllous stems or aerial roots even can serve as primary photosynthetic organs, reproductive structures could derive up to 60%; of their total carbon requirement from own CO 2 fixation. In the review, the main strategies of additional carbon acquisition by non-foliar photosynthetic organs are illustrated, presenting an extensive compilation of published data completed with relevant own studies. [本文引用: 1]
ávilaE, HerreraA, TezaraW (2014). Contribution of stem CO2 fixation to whole-plant carbon balance in nonsucculent species.Photosynthetica 52, 3-15. DOI:10.1007/s11099-014-0004-2URLIn many plant species that remain leafless part of the year, CO 2 fixation occurring in green stems represents an important carbon gain. Traditionally, a distinction has been made between stem photosynthesis and corticular photosynthesis. All stem photosynthesis is, sensu stricto , cortical, since it is carried out largely by the stem cortex. We proposed the following nomenclature: stem net photosynthesis (SNP), which includes net CO 2 fixation by stems with stomata in the epidermis and net corticular CO 2 fixation in suberized stems, and stem recycling photosynthesis (SRP), which defines CO 2 ling in suberized stems. The proposed terms should reflect differences in anatomical and physiological traits. SNP takes place in the chlorenchyma below the epidermis with stomata, where the net CO 2 uptake occurs, and it resembles leaf photosynthesis in many characteristics. SRP is found in species where the chlorenchyma is beneath a well-developed stomata-free periderm and where reassimilation of internally respired CO 2 occurs. SNP is common in plants from desert ecosystems, rates reaching up to 60% of the leaf photosynthetic rate. SRP has been demonstrated in trees from temperate forests and it offsets partially a carbon loss by respiration of stem nonphotosynthetic tissues. Reassimilation can vary between 7 and 123% of respired CO 2 , the latter figure implying net CO 2 uptake from the atmosphere. Both types of stem photosynthesis contribute positively to the carbon economy of the species, in which they occur; they are advantageous to the plant because they allow the maintenance of physiological activity during stress, an increase of integrated water use efficiency, and they provide the carbon source used in the production of new organs. [本文引用: 1]
[20]
BerveillerD, DamesinC (2008). Carbon assimilation by tree stems: potential involvement of phosphoenolpyruvate car- boxylase.Trees 22, 149-157. DOI:10.1007/s00468-007-0193-4URLIn woody species, the photosynthesis of stems, especially young branches, occurs by refixing part of the internal respiratory CO 2 . The present study aims to improve the physiological characterization of stem photosynthesis by examining enzymatic characteristics. During an entire growing season, three enzymatic activities that are linked to C 3 and C 4 metabolism were investigated in relation to the CO 2 efflux and chlorophyll content of current year stems of European beech and were compared to the corresponding characteristics of leaves. High activities of phospho enol pyruvate carboxylase (PEPC) and NADP malic enzyme were detected in stems (up to 13 times and 30 times higher in stems than in leaves, respectively), whereas Rubisco activity remained low in comparison with leaves. Stem maximal Rubisco and PEPC activities occurred at the beginning of the season when the total chlorophyll content and the CO 2 assimilation rate were also maximal. Stems were characterized by a PEPC: Rubisco ratio that was equal to 2.5 [an intermediate value between that of C 3 -plants (about 0.1) and that of C 4 -plants (about 10)], whereas it was equal to 0.1 in leaves. Eight other tree species were also measured and the PEPC: Rubisco ratio was, on average, equal to 3.6. The potential role of PEPC in stem carbon assimilation is discussed in relation to its known involvement in the anaplerotic function of C 3 plants and in the carbon metabolism of the C 4 pathway. [本文引用: 4]
[21]
BerveillerD, KierzkowskiD, DamesinC (2007). Interspecific variability of stem photosynthesis among tree species.Tree Physiol 27, 53-61. DOI:10.1093/treephys/27.1.53PMID:17169906URLAbstract The photosynthetic characteristics of current-year stems of six deciduous tree species, two evergreen tree species and ginkgo (Ginkgo biloba L.) were compared. Gas exchange, chlorophyll concentration, nitrogen concentration and maximum quantum yield of PSII were measured in stems in summer and winter. A light-induced decrease in stem CO(2) efflux was observed in all species. The apparent gross photosynthetic rate in saturating light ranged from 0.72 micromol m(-2 )s(-1) (ginkgo, in winter) to 3.73 micromol m(-2) s(-1) (Alnus glutinosa (L.) Gaertn., in summer). Despite this variability, a unique correlation (slope = 0.75), based on our results and those reported in the literature, was found between gross photosynthetic rate and dark respiration rate. Mass-based gross photosynthetic rate decreased with stem mass per area and correlated to chlorophyll concentration and nitrogen concentration, both in summer and winter. The radial distribution of stem chlorophyll differed among species, but all species except ginkgo had chlorophyll as deep as the pith. In summer, the maximum quantum yield of stem PSII (estimated from the ratio of variable to maximal fluorescence; F (v)/F (m)) of all species was near the optimal value found for leaves. By contrast, the values were highly variable in winter, suggesting large differences in sensitivity to low-temperature photoinhibition. The winter values of Fv/Fm were only 31-60% of summer values for the deciduous species, whereas the evergreen conifer species maintained high F (v)/F (m) in winter. The results highlight the interspecific variability of gross photosynthesis in the stem and its correlation with structural traits like those found for leaves. The structural correlations suggest that the selection of photosynthetic traits has operated under similar constraints in stems and leaves. [本文引用: 2]
[22]
BloemenJ, VergeynstLL, Overlaet-MichielsL, SteppeK (2016). How important is woody tissue photosynthesis in poplar during drought stress? Trees 30, 63-72. DOI:10.1007/s00468-014-1132-9URLWithin trees, a portion of respired CO2 is assimilated by bark and woody tissue photosynthesis, but its physiological role remains unclear, in particular under unfavour able conditions like drought stress. We hypothesised that woody tissue photosynthesis will contribute to overall tree carbon gain both under sufficient water supply and during drought, and plays a role in maintaining the hydraulic function. We subjected half of the trees to a stem and branch light-exclusion treatment to prevent bark and woody tissue photosynthesis. Then, we measured leaf gas exchange and stem growth in Populus deltoides x nigra 'Monviso' trees both under well-watered and dry conditions. We additionally measured cavitation using acoustic emission in detached control and light-excluded branches to illustrate the role of woody tissue photosynthesis in xylem embolism repair. Under well-watered conditions, light exclusion resulted in reduced stem growth relative to control trees by 30 %. In response to drought, stem shrinkage of light-excluded trees was more pronounced as compared to control trees. During drought stress also maximum photosynthesis and transpiration rate tended to decrease more rapidly in light-excluded trees compared to control trees. Leaf fall in light-excluded branches together with the larger number of acoustic emissions in control branches indicates that in the latter more xylem vessels were still hydraulically functional under drought. Therefore, our study highlights that photosynthesis at branch and stem level might be a key factor in the resilience of trees to drought stress by maintaining both the plant carbon economy and hydraulic function. [本文引用: 2]
[23]
BradfordMM (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding.Anal Biochem 72, 248-254. DOI:10.1016/0003-2697(76)90527-3PMID:202019762020722484URLA protein determination method which involves the binding of Coomassie Brilliant Blue G-250 to protein is described. The binding of the dye to protein causes a shift in the absorption maximum of the dye from 465 to 595 nm, and it is the increase in absorption at 595 nm which is monitored. This assay is very reproducible and rapid with the dye binding process virtually complete in approximately 2 min with good color stability for 1 hr. There is little or no interference from cations such as sodium or potassium nor from carbohydrates such as sucrose. A small amount of color is developed in the presence of strongly alkaline buffering agents, but the assay may be run accurately by the use of proper buffer controls. The only components found to give excessive interfering color in the assay are relatively large amounts of detergents such as sodium dodecyl sulfate. Triton X-100, and commercial glassware detergents. Interference by small amounts of detergent may be eliminated by the use of proper controls. [本文引用: 1]
[24]
BurnellJN (1986). Purification and properties of phosphoenolpyruvate carboxykinase from C4 plants.Aust J Plant Physiol 13, 577-587. DOI:10.1071/pp9860577URLPhosphoenolpyruvate (PEP) carboxykinase (EC 4.1.1.49) from the leaves of Urochloa panicoides, Chloris gayana and Panicum maximum has been purified to homogeneity and its properties determined. The enzyme from all three PEP carboxykinase-type C4 plants have similar physical and kinetic properties. The native enzyme has a molecular weight of 380 000 and its monomeric molecular weight is about 64 000, suggesting the enzyme is hexameric. It is active over a wide pH range and has a pH optimum between 7.4 and 8.2, has a wide nucleotide specificity, has an absolute requirement for Mn2+ and is stimulated by C-. The enzyme is inhibited by 3-phosphoglyceric acid, fructose 6-phosphate and fructose 1,6-bisphosphate; the mechanism of inhibition is discussed. The purified PEP carboxykinase is unable to catalyse the conversion of oxaloacetate to pyruvate, nor does it possess pyruvate kinase activity. These findings are discussed in relation to the C4 photosynthetic pathway operating in PEP carboxykinase-type C4 plants. [本文引用: 1]
[25]
BurnellJN, ChastainCJ (2006). Cloning and expression of maize-leaf pyruvate, Pi dikinase regulatory protein gene.Biochem Biophys Res Commun 345, 675-680. DOI:10.1016/j.bbrc.2006.04.150PMID:16696949URLPyruvate, orthophosphate dikinase (PPDK; E.C. 2.7.9.1) catalyzes the synthesis of the primary inorganic carbon acceptor, phosphoenolpyruvate in the C 4 photosynthetic pathway and is reversibly regulated by light. PPDK regulatory protein (RP), a bifunctional serine/threonine kinase-phosphatase, catalyzes both the ADP-dependent inactivation and the Pi-dependent activation of PPDK. Attempts to clone the RP have to date proven unsuccessful. A bioinformatics approach was taken to identify the nucleotide and amino acid sequence of the protein. Based on previously established characteristics including molecular mass, known inter- and intracellular location, functionality, and low level of expression, available databases were interrogated to ultimately identify a single candidate gene. In this paper, we describe the nucleotide and deduced amino acid sequence of this gene and establish its identity as maize PPDK RP by in vitro analysis of its catalytic properties via the cloning and expression of the recombinant protein. [本文引用: 1]
[26]
CarnalNW, AgostinoA, HatchMD (1993). Photosynthesis in phosphoenolpyruvate carboxykinase-type C4 plants: mechanism and regulation of C4 acid decarboxylation in bundle sheath cells.Arch Biochem Biophys 306, 360-367. DOI:10.1006/abbi.1993.1524URL [本文引用: 2]
[27]
CenYP, SageRF (2005). The regulation of rubisco activity in response to variation in temperature and atmospheric CO2 partial pressure in sweet potato.Plant Physiol 139, 979-990. DOI:10.1104/pp.105.066233PMID:16183840URLThe temperature response of net CO2assimilation rate (A), the rate of whole-chain electron transport, the activity and activation state of Rubisco, and the pool sizes of ribulose-1,5-bisphosphate (RuBP) and 3-phosphoglyceric acid (PGA) were assessed in sweet potato (Ipomoea batatas) grown under greenhouse conditions. Above the thermal optimum of photosynthesis, the activation state of Rubisco declined with increasing temperature. Doubling CO2above 370 bar further reduced the activation state, while reducing CO2by one-half increased it. At cool temperature (<16 C), the activation state of Rubisco declined at CO2levels where photosynthesis was unaffected by a 90% reduction in O2content. Reduction of the partial pressure of CO2at cool temperature also enhanced the activation state of Rubisco. The rate of electron transport showed a pronounced temperature response with the same temperature optimum as A at elevated CO2. RuBP pool size and the RuBP-to-PGA ratio declined with increasing temperature. Increasing CO2also reduced the RuBP pool size. These results are consistent with the hypothesis that the reduction in the activation state of Rubisco at high and low temperature is a regulated response to a limitation in one of the processes contributing to the rate of RuBP regeneration. To further evaluate this possibility, we used measured estimates of Rubisco capacity, electron transport capacity, and the inorganic phosphate regeneration capacity to model the response of A to temperature. At elevated CO2, the activation state of Rubisco declined at high temperatures where electron transport capacity was predicted to be limiting, and at cooler temperatures where the inorganic phosphate regeneration capacity was limiting. At low CO2, where Rubisco capacity was predicted to limit photosynthesis, full activation of Rubisco was observed at all measurement temperatures. [本文引用: 1]
[28]
ChaoQ, LiuXY, MeiYC, GaoZF, ChenYB, QianCR, HaoYB, WanBC (2014). Light-regulated phosphorylation of maize phosphoenolpyruvate carboxykinase plays a vital role in its activity.Plant Mol Biol 85, 95-105. DOI:10.1007/s11103-014-0171-3PMID:24435212URLPhosphoenolpyruvate carboxykinase (PEPCK)—the major decarboxylase in PEPCK-type C 4 plants—is also present in appreciable amounts in the bundle sheath cells of NADP-malic enzyme-type C 4 plants, such as maize ( Zea mays ), where it plays an apparent crucial role during photosynthesis (Wingler et al., in Plant Physiol 120(2):539–546, 1999 ; Furumoto et al., in Plant Mol Biol 41(3):301–311, 1999 ). Herein, we describe the use of mass spectrometry to demonstrate phosphorylation of maize PEPCK residues Ser55, Thr58, Thr59, and Thr120. Western blotting indicated that the extent of Ser55 phosphorylation dramatically increases in the leaves of maize seedlings when the seedlings are transferred from darkness to light, and decreases in the leaves of seedlings transferred from light to darkness. The effect of light on phosphorylation of this residue is opposite that of the effect of light on PEPCK activity, with the decarboxylase activity of PEPCK being less in illuminated leaves than in leaves left in the dark. This inverse relationship between PEPCK activity and the extent of phosphorylation suggests that the suppressive effect of light on PEPCK decarboxylation activity might be mediated by reversible phosphorylation of Ser55. [本文引用: 1]
[29]
ChastainCJ, FailingCJ, ManandharL, ZimmermanMA, LaknerMM, NguyenTHT (2011). Functional evolution of C4 pyruvate, orthophosphate dikinase.J Exp Bot 62, 3083-3091. DOI:10.1093/jxb/err058PMID:21414960URLPyruvate,orthophosphate dikinase (PPDK) plays a controlling role in the PEP-regeneration phase of the C(4) photosynthetic pathway. Earlier studies have fully documented its biochemical properties and its post-translational regulation by the PPDK regulatory protein (PDRP). However, the question of its evolution into the C(4) pathway has, until recently, received little attention. One assumption concerning this evolution is that changes in catalytic and regulatory properties of PPDK were necessary for the enzyme to fulfil its role in the C(4) pathway. In this study, the functional evolution of PPDK from its ancient origins in the Archaea to its ascension as a photosynthetic enzyme in modern C(4) angiosperms is reviewed. This analysis is accompanied by a comparative investigation into key catalytic and regulatory properties of a C(3) PPDK isoform from Arabidopsis and the C(4) PPDK isoform from Zea mays. From these analyses, it is proposed that PPDK first became functionally seated in C(3) plants as an ancillary glycolytic enzyme and that its transition into a C(4) pathway enzyme involved only minor changes in enzyme properties per se. [本文引用: 1]
[30]
ChenYB, LuTC, WangHX, ShenJ, BuTT, ChaoQ, GaoZF, ZhuXG, WangYF, WangBC (2014). Posttranslational modification of maize chloroplast pyruvate orthophosphate dikinase reveals the precise regulatory mechanism of its enzymatic activity.Plant Physiol 165, 534-549. DOI:10.1104/pp.113.231993PMID:24710069URLIn C4 plants, pyruvate orthophosphate dikinase (PPDK) activity is tightly dark/light regulated by reversible phosphorylation of an active-site threonine (Thr) residue; this process is catalyzed by PPDK regulatory protein (PDRP). Phosphorylation and dephosphorylation of PPDK lead to its inactivation and activation, respectively. Here, we show that light intensity rather than the light/dark transition regulates PPDK activity by modulating the reversible phosphorylation at Thr-527 (previously termed Thr-456) of PPDK in maize (Zea mays). The amount of PPDK (unphosphorylated) involved in C4 photosynthesis is indeed strictly controlled by light intensity, despite the high levels of PPDK protein that accumulate in mesophyll chloroplasts. In addition, we identified a transit peptide cleavage site, uncovered partial amino-terminal acetylation, and detected phosphorylation at four serine (Ser)/Thr residues, two of which were previously unknown in maize. In vitro experiments indicated that Thr-527 and Ser-528, but not Thr-309 and Ser-506, are targets of PDRP. Modeling suggests that the two hydrogen bonds between the highly conserved residues Ser-528 and glycine-525 are required for PDRP-mediated phosphorylation of the active-site Thr-527 of PPDK. Taken together, our results suggest that the regulation of maize plastid PPDK isoform (C4PPDK) activity is much more complex than previously reported. These diverse regulatory pathways may work alone or in combination to fine-tune C4PPDK activity in response to changes in lighting. [本文引用: 1]
[31]
CuiK, HeCY, ZhangJG, DuanAG, ZengYF (2012). Temporal and spatial profiling of internode elongation-associated protein expression in rapidly growing culms of bamboo.J Proteome Res 11, 2492-2507. DOI:10.1021/pr2011878PMID:22397471URLIn natural conditions, culms of developing Moso bamboo, Phyllostachys heterocycla var. pubescens, reach their final height of more than ten meters within a short period of two to four months. To study this phenomenon, bamboo culm material collected from different developmental stages and internodes was analyzed. Histological observations indicated that the development of culm was dominated by cell division in the initial stages and by cell elongation in the middle and late stages. Development, maturation, and aging in different regions of the culm were studied systematically from the basal to the top internode. The four major endogenous hormones, indole acetic acid, gibberellic acid, zeatin riboside, and abscisic acid appeared to strongly influence the cell elongation phase. A total of 258 spots were differentially expressed in culm development. Of these, 213 spots were identified by MALDI-TOF/TOF MS and were involved in many physiological and metabolic processes including carbohydrate metabolism, cell di... [本文引用: 1]
[32]
DamesinC (2003). Respiration and photosynthesis characteristics of current-year stems of Fagus sylvatica: from the seasonal pattern to an annual balance. New Phytol 158, 465-475. DOI:10.1046/j.1469-8137.2003.00756.xURL61 Temperature and light responses of CO2efflux of Fagus sylvatica (beech) current-year stems were measured for 1 yr to estimate their annual carbon balance. 61 Gas exchanges were determined using infrared gas analysis. Seasonal patterns of a fluorescence parameter ((Fv/Fm)max), nitrogen and chlorophyll contents were also assessed in stems and leaves, using standard techniques. 61 Basal respiration rates at 20°C ( R20) were very high during the growing season, reaching a maximum of 17 170 μ mol m-3s-1. Light-saturated assimilation followed the same seasonal pattern as R20. During the winter, chlorophyll content was undiminished compared with the summer, N content was slightly increased, and despite low (Fv/Fm)maxvalues, instantaneous maximum assimilation could account for 80-110% of the respiration. 61 For an average-size stem (4 mm diameter), the estimated annual respiration was 0.5 g carbon with 55% of this amount attributed to maintenance respiration. The potential assimilation contributed 0.2 g carbon and approximately compensated for the growth respiration. Information on older branches and trunks is now needed for estimations at the tree and stand levels. [本文引用: 1]
[33]
DimaE, ManetasY, PsarasGK (2006). Chlorophyll distribution pattern in inner stem tissues: evidence from epifluorescence microscopy and reflectance measurements in 20 woody species.Trees 20, 515-521. DOI:10.1007/s00468-006-0067-1URL [本文引用: 1]
[34]
HatchMD, SlackCR (1975). Pyruvate, Pi dikinase from leaves.Methods Enzymol 42, 212-219. DOI:10.1016/0076-6879(75)42117-6URL [本文引用: 1]
[35]
HibberdJM, QuickWP (2002). Characteristics of C4 photosynthesis in stems and petioles of C3 flowering plants.Nature 415, 451-454. DOI:10.1038/415451aURL [本文引用: 3]
[36]
HrstkaM, UrbanO, BabákL (2012). Seasonal changes of rubisco content and activity in Fagus sylvatica and Picea abies affected by elevated CO2 concentration. Chem Pap 66, 836-841. [本文引用: 1]
[37]
JohnsonHS, HatchMD (1970). Properties and regulation of leaf nicotinamide-adenine dinucleotide phosphate-malate dehydrogenase and ‘malic’ enzyme in plants with the C4-dicarboxylic acid pathway of photosynthesis.Biochem J 119, 273-280. DOI:10.1042/bj1190273URL [本文引用: 1]
[38]
KocurekM, PilarskiJ (2011). Activity of C4 enzymes in C3-type herbaceous plants.Photosynthetica 49, 473-477. DOI:10.1007/s11099-011-0053-8URLThe activity of enzymes characteristic for C 4 -type photosynthesis was determined in different organs of two herbaceous plants: Reynoutria japonica Houtt. and Helianthus tuberosus L. The activity of phospho enol pyruvate carboxylase (PEPC) was usually higher in the roots, some of the stem tissues and petioles in comparison to the leaf blades. The highest activity of malic enzymes (NAD-ME, NADP-ME) and phospho enol pyruvate carboxykinase (PEPCK) was in the petioles and stem tissues of both plants and the lowest in the leaf blades and the pith of Helianthus tuberosus L. [本文引用: 1]
[39]
KosvancováM, UrbanO, ŠprtováM, HrstkaM, KalinaJ, TomáskovaI, ŠpundaV, MarekMV (2009). Photosynthetic induction in broadleaved Fagus sylvatica and conife- rous Picea abies cultivated under ambient and elevated CO2 concentrations. Plant Sci 177, 123-130. [本文引用: 1]
[40]
LeegoodRC (2013). Strategies for engineering C4 photosynthesis.J Plant Physiol 170, 378-388. DOI:10.1016/j.jplph.2012.10.011PMID:23245935URLC3 photosynthesis is an inefficient process, because the enzyme that lies at the heart of the Benson–Calvin cycle, ribulose 1,5-bisphosphate carboxylase-oxygenase (Rubisco) is itself a very inefficient enzyme. The oxygenase activity of Rubisco is an unavoidable side reaction that is a consequence of its reaction mechanism. The product of oxygenation, glycollate 2-P, has to be retrieved by photorespiration, a process which results in the loss of a quarter of the carbon that was originally present in glycollate 2-P. Photorespiration therefore reduces carbon gain. Purely in terms of carbon economy, there is, therefore, a strong selection pressure on plants to reduce the rate of photorespiration so as to increase carbon gain, but it also improves water- and nitrogen-use efficiency. Possibilities for the manipulation of plants to decrease the amount of photorespiration include the introduction of improved Rubisco from other species, reconfiguring photorespiration, or introducing carbon-concentrating mechanisms, such as inorganic carbon transporters, carboxysomes or pyrenoids, or engineering a full C4 Kranz pathway using the existing evolutionary progression in C3–C4 intermediates as a blueprint. Possible routes and progress to suppressing photorespiration by introducing C4 photosynthesis in C3 crop plants will be discussed, including whether single cell C4 photosynthesis is feasible, how the evolution of C3–C4 intermediates can be used as a blueprint for engineering C4 photosynthesis, which pathway for the C4 cycle might be introduced and the extent to which processes and structures in C3 plant might require optimisation. [本文引用: 1]
[41]
LeegoodRC, OsmondCB (1990). The flux of metabolites in C4 and CAM plants. In: Dennis DT, Turpin DH, eds. Plant Physiology, Biochemistry and Molecular Biology. Essex: Longman Scientific & Technical. pp. 274-298. [本文引用: 1]
[42]
LichtenthalerHK (1987). Chlorophylls and carotenoids: pigments of photosynthetic biomembranes.Methods Enzymol 148, 350-382. DOI:10.1016/0076-6879(87)48036-1URL [本文引用: 1]
[43]
MuhaidatR, SageRF, DenglerNG (2007). Diversity of Kranz anatomy and biochemistry in C4 eudicots.Am J Bot 94, 362-381. DOI:10.3732/ajb.94.3.362PMID:21636407URLC4 photosynthesis and Kranz anatomy occur in 16 eudicot families, a striking example of convergent evolution. Biochemical subtyping for 13 previously undiagnosed C4 eudicot species indicated that 10 were NADP-malic enzyme (ME) and three were NAD-ME. A total of 33 C4 species, encompassing four Kranz anatomical types (atriplicoid, kochioid, salsoloid, and suaedioid), and 21 closely related C3 species were included in a quantitative anatomical study in which we found that, unlike similar studies in grasses and sedges, anatomical type had no predictive value for the biochemical subtype. In a multivariate canonical discriminant analysis, C4 species were distinguished from C3 species by the mesophyll to bundle sheath ratio and exposure of the bundle sheath surface to intercellular space. Discrimination between NADP-ME and NAD-ME was not significant, although in a Mantel test grouping by biochemical subtype was significant, while grouping by family was not. This comprehensive survey of C4 anatomy and biochemistry unequivocally demonstrated that atriplicoid anatomy and NADP-ME biochemistry predominate in many evolutionary lineages. In addition to a main decarboxylating enzyme, high activity of a second decarboxylating enzyme was often observed. Notably, PEP-carboxykinase activity was significant in a number of species, demonstrating that this enzyme could also serve as a secondary pathway for C4 metabolism in eudicots. [本文引用: 1]
[44]
OcampoG, ColumbusJT (2010). Molecular phylogenetics of suborder cactineae (caryophyllales), including insights into photosynthetic diversification and historical biogeography.Am J Bot 97, 1827-1847. DOI:10.3732/ajb.1000227PMID:21616822URLPremise of the study: Phylogenetic relationships were investigated among the eight families (Anacampserotaceae, Basellaceae, Cactaceae, Didiereaceae, Halophytaceae, Montiaceae, Portulacaceae, Talinaceae) that form suborder Cactineae (= Portulacineae) of the Caryophyllales. In addition, photosynthesis diversification and historical biogeography were addressed. Methods: Chloroplast DNA sequences, mostly noncoding, were used to estimate the phylogeny. Divergence times were calibrated using two Hawaiian Portulaca species, due to the lack of an unequivocal fossil record for Cactineae. Photosynthetic pathways were determined from carbon isotope ratio (δ0106C) and leaf anatomy. Key results: Maximum likelihood and Bayesian analyses were consistent with previous studies in that the suborder, almost all families, and the ACPT clade (Anacampserotaceae, Cactaceae, Portulacaceae, Talinaceae) were strongly supported as monophyletic; however, relationships among families remain uncertain. The age of Cactineae was estimated to be 18.8 Myr. Leaf anatomy and δ0106C and were congruent in most cases, and inconsistencies between these pointed to photosynthetic intermediates. Reconstruction of photosynthesis diversification showed C61 to be the ancestral pathway, a shift to C62 in Portulacaceae, and five independent origins of Crassulacean acid metabolism (CAM). Cactineae were inferred to have originated in the New World. Conclusions: Although the C61 pathway is inferred as the ancestral state in Cactineae, some CAM activity has been reported in the literature in almost every family of the suborder, leaving open the possibility that CAM may have one origin in the group. Incongruence among loci could be due to internal short branches, which possibly represent rapid radiations in response to increasing aridity in the Miocene. [本文引用: 1]
[45]
ParryMAJ, AndralojcPJ, ScalesJC, SalvucciME, Carmo-SilvaAE, AlonsoH, WhitneySM (2013). Rubisco activity and regulation as targets for crop improvement.J Exp Bot 64, 717-730. DOI:10.1093/jxb/ers336PMID:23162118URLRubisco (ribulose-1,5-bisphosphate (RuBP) carboxylase/oxygenase) enables net carbon fixation through the carboxylation of RuBP. However, some characteristics of Rubisco make it surprisingly inefficient and compromise photosynthetic productivity. For example, Rubisco catalyses a wasteful reaction with oxygen that leads to the release of previously fixed CO2 and NH3 and the consumption of energy during photorespiration. Furthermore, Rubisco is slow and large amounts are needed to support adequate photosynthetic rates. Consequently, Rubisco has been studied intensively as a prime target for manipulations to 0900supercharge0964 photosynthesis and improve both productivity and resource use efficiency. The catalytic properties of Rubiscos from diverse sources vary considerably, suggesting that changes in turnover rate, affinity, or specificity for CO2 can be introduced to improve Rubisco performance in specific crops and environments. While attempts to manipulate plant Rubisco by nuclear transformation have had limited success, modifying its catalysis by targeted changes to its catalytic large subunit via chloroplast transformation have been much more successful. However, this technique is still in need of development for most major food crops including maize, wheat, and rice. Other bioengineering approaches for improving Rubisco performance include improving the activity of its ancillary protein, Rubisco activase, in addition to modulating the synthesis and degradation of Rubisco0964s inhibitory sugar phosphate ligands. As the rate-limiting step in carbon assimilation, even modest improvements in the overall performance of Rubisco pose a viable pathway for obtaining significant gains in plant yield, particularly under stressful environmental conditions. [本文引用: 1]
RanganP, FurtadoA, HenryRJ (2016). New evidence for grain specific C4 photosynthesis in wheat.Sci Rep 6, 31721. DOI:10.1038/srep31721URL [本文引用: 1]
[50]
SageRF, SageTL, KocacinarF (2012). Photorespiration and the evolution of C4 photosynthesis.Annu Rev Plant Biol 63, 19-47. DOI:10.1146/annurev-arplant-042811-105511URL [本文引用: 1]
[51]
SageRF, SageTL, PearcyRW, BorschT (2007). The taxonomic distribution of C4 photosynthesis in Amaranthaceae sensu stricto.Am J Bot 94, 1992-2003. DOI:10.3732/ajb.94.12.1992URLC4 photosynthesis evolved multiple times in the Amaranthaceae s.s., but the C4 evolutionary lineages are unclear because the photosynthetic pathway is unknown for most species of the family. To clarify the distribution of C4 photosynthesis in the Amaranthaceae, we determined carbon isotope ratios of 607 species and mapped these onto a phylogeny determined from matK/trnK sequences. Approximately 28% of the Amaranthaceae species use the C4 pathway. C4 species occur in 10 genera erva, Amaranthus, Blutaparon, Alternanthera, Froelichia, Lithophila, Guilleminea, Gomphrena, Gossypianthus, and Tidestromia. Aerva, Alternanthera, and Gomphrena contain both C3 and C4 species. In Aerva, 25% of the sampled species are C4. In Alternanthera, 19.5% are C4, while 89% of the Gomphrena species are C4. Integration of isotope and matK/trnK data indicated C4 photosynthesis evolved five times in the Amaranthaceae, specifically in Aerva, Alternanthera, Amaranthus, Tidestromia, and a lineage containing Froelichia, Blutaparon, Guilleminea, Gomphrena pro parte, and Lithophila. Aerva and Gomphrena are both polyphyletic with C3 and C4 species belonging to distinct clades. Alternanthera appears to be monophyletic with C4 photosynthesis originating in a terminal sublineage of procumbent herbs. Alpine C4 species were also identified in Alternanthera, Amaranthus, and Gomphrena, including one species (Gomphrena meyeniana) from 4600 m a.s.l. [本文引用: 1]
[52]
SaveynA, SteppeK, UbiernaN, DawsonTE (2010). Woody tissue photosynthesis and its contribution to trunk growth and bud development in young plants.Plant Cell Environ 33, 1949-1958. DOI:10.1111/j.1365-3040.2010.02197.xPMID:20561249URLStem photosynthesis can contribute significantly to woody plant carbon balance, particularly in times when leaves are absent or in pen crowns with sufficient light penetration. We explored the significance of woody tissue (stem) photosynthesis for the carbon income in three California native plant species via measurements of chlorophyll concentrations, radial stem growth, bud biomass and stable carbon isotope composition of sugars in different plant organs. Young plants of Prunus ilicifolia, Umbellularia californica and Arctostaphylos manzanita were measured and subjected to manipulations at two levels: trunk light exclusion (100 and 50%) and complete defoliation. We found that long-term light exclusion resulted in a reduction in chlorophyll concentration and radial growth, demonstrating that trunk assimilates contributed to trunk carbon income. In addition, bud biomass was lower in covered plants compared to uncovered plants. Excluding 100% of the ambient light from trunks on defoliated plants led to an enrichment in 13C of trunk phloem sugars. We attributed this effect to a reduction in photosynthetic carbon isotope discrimination against 13C that in turn resulted in an enrichment in 13C of bud sugars. Taken together our results reveal that stem photosynthesis contributes to the total carbon income of all species including the buds in defoliated plants. [本文引用: 1]
[53]
ShethBP, ThakerVS (2014). In silico analyses of Rubisco enzymes from different classes of Algae. Int Res J Biol Sci 3, 11-17. URLRubisco (Ribulose 1, 5 Bisphosphate Carboxylase Oxygenase) is the most predominant enzyme of one of the few carbon assimilatory processes in nature i.e. Photosynthesis. The rbcL and rbcS genes code for the large and small subunits of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) respectively. In this study the rbcL protein sequences selected from various classes of algae were phylogenetically analyzed. Expasy's Prot-param server and Cys_rec tool were used for physico-chemical and functional characterization of these proteins. For comparative structural analysis, experimental structures (X-ray and NMR) of rubisco proteins of representative species of Rhodophyta (Galderia sp. PDBID 1IWA) and Chlorophyta (Chlamydomonas sp. 1GK8) were used. Also, as no experimental structure of rubisco from any member of phaeophyta group is available, homology modeling approach was employed in order to derive structure of the same from Lessonia vadosa, a representative species of phaeophyta group. The validity of the modeled protein was further checked by RAMPAGE, Procheck, WHATIF, Errat, and Verify-3d servers. Studies of secondary structure of these proteins were carried out by the SSCP server. The in silico analysis, confirmed the close correlation between the rhodophyte and the phaeophyte rubico proteins at the functional level due to similarity in adaptability of the enzyme. [本文引用: 1]
[54]
SunJL, SuiXL, WangSH, WeiYX, HuangHY, HuLP, ZhangZX (2014). The response of rbcL, rbcS and rca genes in cucumber(Cucumis sativus L.) to growth and induction light intensity. Acta Physiol Plant 36, 2779-2791. [本文引用: 1]
[55]
SuzukiY, MakinoA (2012). Availability of Rubisco small subunit up-regulates the transcript levels of large subunit for stoichiometric assembly of its holoenzyme in rice.Plant Physiol 160, 533-540. DOI:10.1104/pp.112.201459PMID:22811433URLRubisco is composed of eight small subunits coded for by the nuclear RBCS multigene family and eight large subunits coded for by the rbcL gene in the plastome. For synthesis of the Rubisco holoenzyme, both genes need to be expressed coordinately. To investigate this molecular mechanism, the protein synthesis of two subunits of Rubisco was characterized in transgenic rice (Oryza sativa) plants with overexpression or antisense suppression of the RBCS gene. Total RBCS and rbcL messenger RNA (mRNA) levels and RBCS and RbcL synthesis simultaneously increased in RBCS-sense plants, although the increase in total RBCS mRNA level was greater. In RBCS-antisense plants, the levels of these mRNAs and the synthesis of the corresponding proteins declined to a similar extent. The amount of RBCS synthesized was tightly correlated with rbcL mRNA level among genotypes but not associated with changes in mRNA levels of other major chloroplast-encoded photosynthetic genes. The level of rbcL mRNA, in turn, was tightly correlated with the amount of RbcL synthesized, the molar ratio of RBCS synthesis to RbcL synthesis being identical irrespective of genotype. Polysome loading of rbcL mRNA was not changed. These results demonstrate that the availability of RBCS protein up-regulates the gene expression of rbcL primarily at the transcript level in a quantitative manner for stoichiometric assembly of Rubisco holoenzyme. [本文引用: 1]
[56]
TeskeyRO, SaveynA, SteppeK, McGuireMA (2008). Origin, fate and significance of CO2 in tree stems.New Phytol 177, 17-32. DOI:10.1111/j.1469-8137.2007.02286.xURLAlthough some CO2 released by respiring cells in tree stems diffuses directly to the atmosphere, on a daily basis 15-55% can remain within the tree. High concentrations of CO2 build up in stems because of barriers to diffusion in the inner bark and xylem. In contrast with atmospheric [CO2] of c. 0.04%, the [CO2] in tree stems is often between 3 and 10%, and sometimes exceeds 20%. The [CO2] in stems varies diurnally and seasonally. Some respired CO2 remaining in the stem dissolves in xylem sap and is transported toward the leaves. A portion can be fixed by photosynthetic cells in woody tissues, and a portion diffuses out of the stem into the atmosphere remote from the site of origin. It is now evident that measurements of CO2 efflux to the atmosphere, which have been commonly used to estimate the rate of woody tissue respiration, do not adequately account for the internal fluxes of CO2. New approaches to quantify both internal and external fluxes of CO2 have been developed to estimate the rate of woody tissue respiration. A more complete assessment of internal fluxes of CO2 in stems will improve our understanding of the carbon balance of trees. [本文引用: 1]
[57]
TikhonovKG, KhristinMS, KlimovVV, SundirevaMA, KreslavskiVD, SidorovRA, TsidendambayevVD, SavchenkoTV (2017). Structural and functional characteristics of photosynthetic apparatus of chlorophyllcon- taining grape vine tissue.Russian J Plant Physiol 64, 73-82. DOI:10.1134/S102144371606011XURLPhotosynthesis in tissues under periderm of woody stems and shoots of perennial plants occurs in environment that is very different from the internal environment of leaf chloroplasts. These tissues are characterized by high CO 2 and low O 2 concentrations, more acidic surroundings, besides that only light which have passed through periderm reaches photosynthetic antennas. In contrast to leaves of deciduous plants chlorenchyma tissues of wintering plant organs are exposed to temperature fluctuations during all seasons, that is why the photosynthetic apparatus of woody stems has to be able to adapt to a wide range of environmental temperatures. In order to reveal unique features, which enable photosynthetic apparatus of chlorenchyma cells in woody plant organs to implement biological functions under different light and temperature conditions, we studied photosynthetic tissues of stem cortex in grapevine ( Vitis vinifera L.) under normal conditions and after exposure to suboptimal temperatures and high light intensity. Comparative analysis of photosynthetic pigment composition and low-temperature chlorophyll fluorescence emission spectrum of leaves, young shoots and chlorenchyma of lignified shoots revealed relatively high level of chlorophyll b and carotenoids, and high photosystem II (PSII) to photosystem I (PSI) ratio in woody shoots. Analysis of parameters of variable chlorophyll fluorescence revealed high PSII activity in grapevine shoot cortex and demonstrated improved freeze tolerance and higher sensitivity to light of photosynthetic apparatus in grape vine in comparison to leaves. It was shown for the first time that photosynthetic apparatus in chlorenchyma cells of vine undergoes so-called tate-transition ast rearrangements leading to redistribution of energy between photosystems. Analysis of fatty acid (FA) compositions of lipids in examined tissues showed that the FA unsaturation index in green tissue of vine is lower than in leaves. A distinct feature of FA compositions of lipids in vine cortex was relatively high level of linoleic acid. [本文引用: 1]
[58]
WangQM, HouFY, DongSX, XieBT, LiAX, ZhangHY, ZhangLM (2014). Effects of shading on the photosynthetic capacity, endogenous hormones and root yield in purple-fleshed sweetpotato ( Ipomoea batatas (L.) Lam). Plant Growth Regul 72, 113-122. DOI:10.1007/s10725-013-9842-3URLA field experiment was conducted to examine the effects of shading on the photosynthetic capacity, endogenous hormones and root yield in purple-fleshed sweetpotato [ Ipomoea batatas L. cv. Jishu18 and Ayamuraski (Aya)]. Sweetpotato plants were treated with two shading levels, 40 and 70% shading, with full radiation used as a control. The results showed that the photosynthetic rate, adenosine triphosphatase activity, Ribulose 1,5-bisphosphate carboxylase activity and soluble sugar content decreased under both shading treatments. Leaf indole-3-acetic acid (IAA) and abscisic acid content increased, whereas leaf gibberellic acid content, zeatin riboside (ZR) content, root IAA, and ZR content decreased in the plants under both shading treatments. Shading also altered the production of sweetpotato storage root, including reductions in the root yield and dry matter accumulation, increase in the top/root (T/R) ratio, and the difference between the treatments and control for the T/R value and storage root yield was significant. Therefore, the responses of the photosynthetic parameters and endogenous hormones to shading were closely correlated with the variation in the storage root yield of the different cultivars. In response to shading, the reduction of root ZR contents, the fresh dry weight of the above-ground parts and the root yield for Jishu18 were higher than that for cv. Aya, indicating that cv. Jishu18 might be more sensitive to weak light than cv. Aya. [本文引用: 1]
[59]
WittmannC, PfanzH (2014). Bark and woody tissue photosynthesis: a means to avoid hypoxia or anoxia in deve- loping stem tissues.Funct Plant Biol 41, 940-953. DOI:10.1071/FP14046URLIn woody plants, oxygen transport and delivery via the xylem sap are well described, but the contribution of bark and woody tissue photosynthesis to oxygen delivery in stems is poorly understood. Here, we combined stem chlorophyll fluorescence measurements with microsensor quantifications of bark O2 levels and oxygen gas exchange measurements of isolated current-year stem tissues of beech (Fagus sylvatica L.) and pedunculate oak (Quercus robur L.) to investigate how bark and woody tissue photosynthesis impairs the oxygen status of stems. Measurements were made before bud break, when the axial path of oxygen supply via the xylem sap is impeded. At that time, bark O2 levels showed O2 concentrations below the atmospheric concentration, indicating hypoxic conditions or O2 deficiency within the inner bark, but the values were always far away from anoxic. Under illumination bark and woody tissue photosynthesis rapidly increased internal oxygen concentrations compared with plants in the dark, and thereby counteracted against localised hypoxia. The highest photosynthetic activity and oxygen release rates were found in the outermost cortex tissues. By contrast, rates of woody tissue photosynthesis were considerably lower, due to the high light attenuation of the bark and cortex tissues, as well as resistances in radial oxygen diffusion. Therefore, our results confirm that bark and woody tissue photosynthesis not only play a role in plant carbon economy, but may also be important for preventing low oxygen-limitations of respiration in these dense and metabolically active tissues. [本文引用: 1]
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}