Abstract: Palynology is an important method to solve the micro-morphological differentiation of species in some complex groups. With the development of molecular phylogeny, combining the advantages of these two subjects may effectively solve the taxonomic issues in plants. Microlepia (Dennstaedtiaceae) is one of the most difficult genera in ferns in terms of taxonomy. In the present study, based on palynology matched with phylogeny and population sampling, we constructed the phylogeny of 280 samples by using both Maximum Likelihood and Bayesian methods with four plastid markers (rbcL, trnL-F, psbA-trnH and rps4). The spore morphology of 100 samples was observed and analyzed. Our results of comprehensive molecular phylogeny and palynology showed that (1) 15 species widely accepted based on plant morphology were strongly supported, and this complex group of Microlepia has been further clarified; (2) There may be crytic species in the populations of M. marginata; (3) M. yaoshanica Ching, M. lofoushanensis Ching, M. szechuanica Ching and M. subspeluncae Ching, treated as synonyms in previous studies, should be restored as independent species; (4) Many hybridization events of Microlepia were found; and (5) A complete infrageneric taxonomy of Microlepia was proposed.
Key words:crytic species ; hybrids ; palynology ; phylogeny and systematics ; pteridophytes ; taxonomy
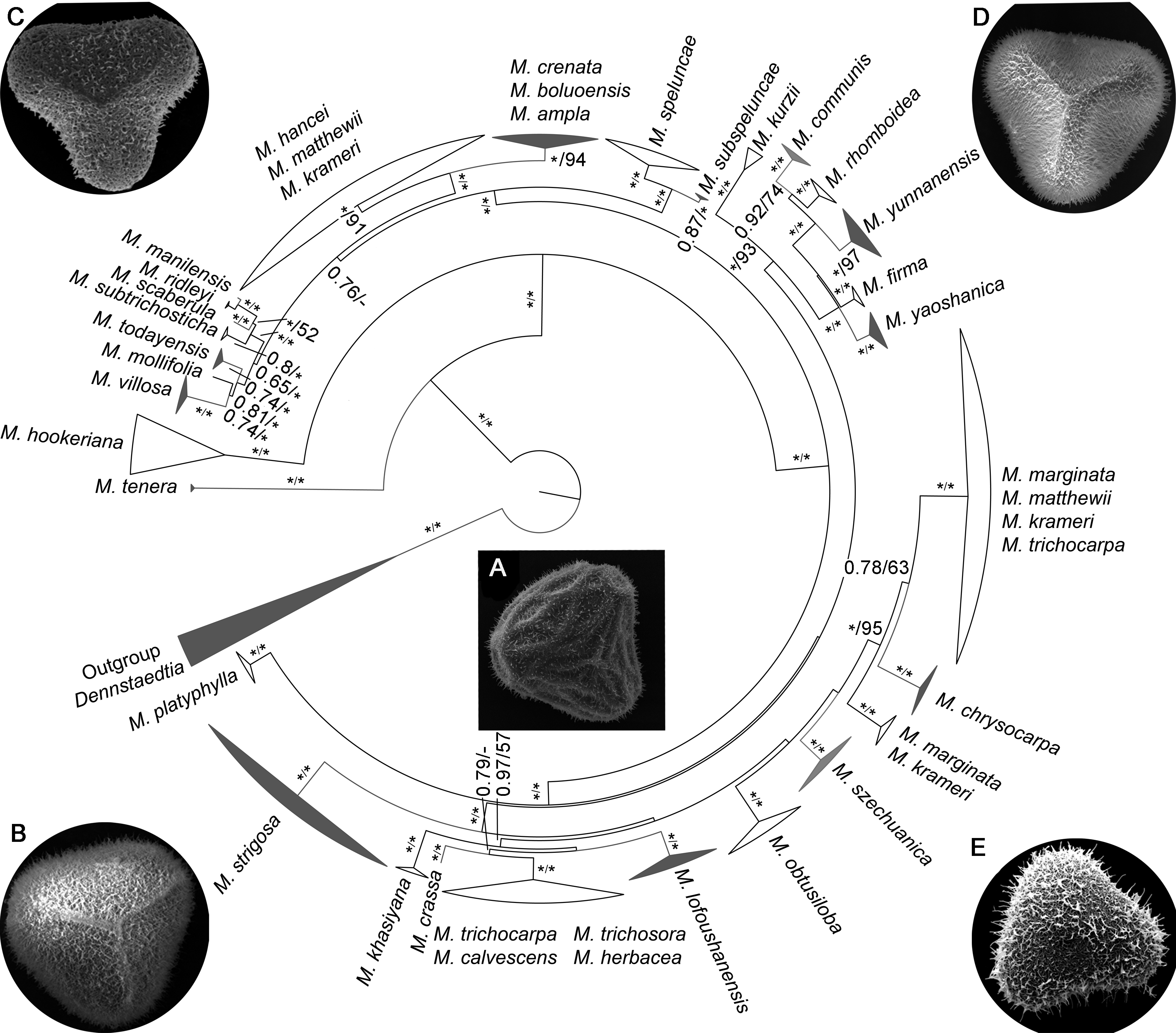
图 1 基于4个叶绿体片段rbcL、trnL-F、psbA-trnH和rps4联合数据构建的鳞盖蕨属贝叶斯系统发育树 分支旁边的数字表示贝叶斯后验概率与ML自展支持率(≥50%); * 表示1或100; - 表示<0.5或50%。 Figure 1 Bayesian inference phylogeny of Microlepia derived from the combined data (rbcL, trnL-F, psbA-trnH and rps4) Values beside each branch represent bootstrap support for parsimony, Bayesian posterior probabilities and maximum likelihood (≥50%); * means 1 or 100; - means <0.5 or 50%.
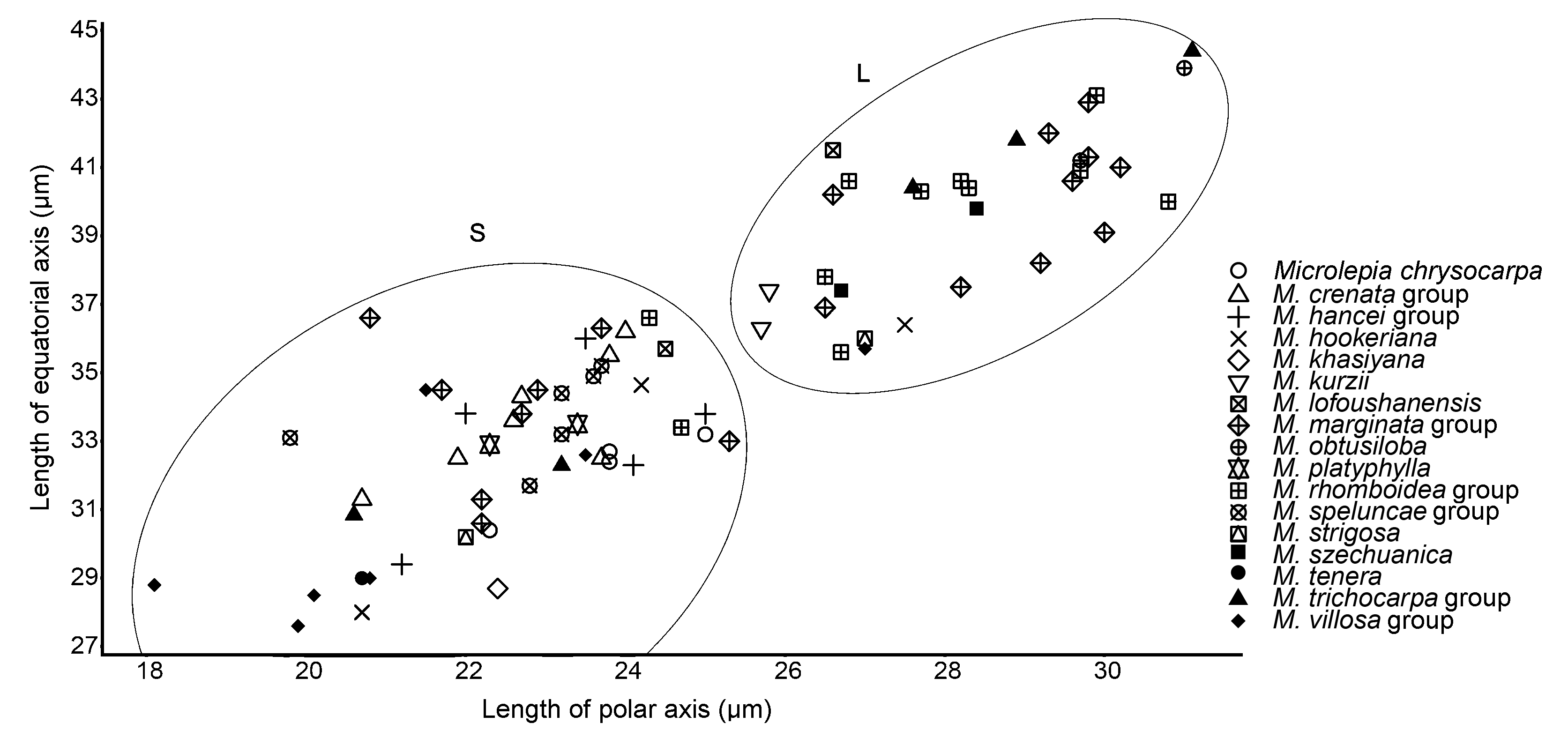
2.2 鳞盖蕨属孢子大小差异与谱系分化的比较本研究所观察的100份材料中, 有15份材料的孢子形态畸形, 85份材料的孢子形态正常。根据谱系分化情况统计了80份材料的孢子大小(图2), 图2显示, 鳞盖蕨属的孢子极轴长度与赤道轴长度呈线性相关。根据孢子大小该属植物较明显地聚为2类, 一类较小(以S表示), 另一类较大(以L表示)。对于同物种而言, 除了毛阔叶鳞盖蕨、团羽鳞盖蕨、阔叶鳞盖蕨及薄叶鳞盖蕨在孢子大小上较为集中外, 其余均较为分散; 但是 在谱系上先分化的薄叶鳞盖蕨、虎克鳞盖蕨以及聚为1个大单系支的热带鳞盖蕨分支、密毛鳞盖蕨分支、华南鳞盖蕨分支和圆齿鳞盖蕨分支等鳞盖蕨属植物孢子较小, 除虎克鳞盖蕨和密毛鳞盖蕨分支各有1个样本位于L的下部区域外, 其余均分布在S区域; 然而后分化的毛阔叶鳞盖蕨和斜方鳞盖蕨分支则孢子较大, 除阔叶鳞盖蕨与斜方鳞盖蕨分支中2个样本分布在S区域外, 其余均分布在L区域, 最后聚为1个大单系支的植物孢子大小较为多变。团羽鳞盖蕨与四川鳞盖蕨只分布在L区域, 金果鳞盖蕨只分布在S区域, 而粗毛鳞盖蕨、罗浮鳞盖蕨、毛果鳞盖蕨分支与边缘鳞盖蕨分支则在2个区域均有分布。 图 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-6-782/img_2.png<b>图 2</b> 孢子大小分布对应鳞盖蕨属的谱系分化<br/><b>Figure 2</b> Distribution of spore size corresponded to phylogeny differentiation of <i>Microlepia</i> Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-6-782/img_2.png<b>图 2</b> 孢子大小分布对应鳞盖蕨属的谱系分化<br/><b>Figure 2</b> Distribution of spore size corresponded to phylogeny differentiation of <i>Microlepia</i>
附图1 基于4个叶绿体片段rbcL、trnL-F、psbA-trnH及rps4联合数据构建的贝叶斯系统发育树(包含rbcL假基因)Appendix figure 1 Bayesian information phylogeny derived from the combined data (rbcL, trnL-F, psbA-trnH and rps4), included rbcL pseudogene http://www.chinbullbotany.com/fileup/PDF/t17258-1.pdf
杨鲁红 (2012). 中国石韦属植物系统分类学研究. 硕士论文. 昆明: 云南大学. pp. 1-142. URL石韦属Pyrrosia Mirbel隶属于水龙骨科Polypodiaceae、石韦亚科Pyrrosioideae,全世界约有70种,是水龙骨科中较大的属。该属植物分布于旧大陆,东亚和东南亚是其主要分布区。该属植物一般附生在树上或岩石上,少数土生,是蕨类植物中比较进化的一类中小型蕨类。石韦属自成立以来,.以其特殊的脉序和星芒状的毛被而成为一个自然的类群,从而与水龙骨科其它属相区别,是研究水龙骨科系统分类的关键类群之一。 本文在文献考证及标本研究的基础上,从鳞片、脉序、毛被、孢粉纹饰、矿质元素等方面,对石韦属植物进行系统分类学研究,提出一个新的属下分类系统,确定中国产石韦属植物共27种、3变种、2变型、1存疑种。9个名称处理为新异名。 1鳞片解剖 对石韦属38种(《中国植物志》资料)代表植物的鳞片类型、形状、颜色、着生方式和边缘特征进行研究。本属植物的鳞片具有3种着生方式:盾状着生、拟盾状着生和基部着生;4种形状:披针形渐尖、披针形尾尖、卵钻形和卵圆形;3种颜色:浅棕色、深棕色及近黑色(颜色之间有一定的过渡,与植物的生长状态和生长阶段有关);4种边缘:全缘、睫状齿、长纤毛和顶端毛发状。 2脉序类型 研究了石韦属10种代表植物和水龙骨科其它5属6种植物的脉序类型。结果表明,水龙骨科植物叶脉分为分离叶脉和结合叶脉,石韦属植物的脉序属于结合脉序,但与被研究的水龙骨科其他属的结合脉序不同。分为两种:弯脉蕨型与拟槲蕨型。前者与特产于美洲的水龙科弯脉蕨属脉序相同,可能二属有一定的亲缘关系。 3毛被特征 主要对石韦属38个种的叶背面毛被进行了显微和扫描电镜观察。石韦属植物毛被由特殊的星状毛组成。星状毛分支臂形状有3种类型:披针形、针状、卷曲绒毛状,分别为浅棕色、深棕色和白色;分支臂类型有2种:一型(分支臂仅1种形状)和二型(分支臂有2种形状,即同轴上长出披针形和长细丝状两种分支臂);这些不同分支臂的星状毛相互组合,将石韦属叶下表面毛被分为以下5种类型:Ⅰ.一种披针形臂毛被;Ⅱ.一种针状臂毛被;Ⅲ.一种分支臂二型毛被;Ⅳ两种针状和披针形毛被;V.两种针状和卷曲绒毛状毛被。这些特征性性状对石韦属部分种类的分类鉴定具有一定的意义。 4孢粉特征 光镜和扫描电镜下观察石韦属19种植物的孢子特征,石韦属孢子黄色,形态均为肾形,两侧对称,单裂缝,说明该属植物是一个自然类群。表面纹饰类型有3种:Pyrrosia rupestris-type、Pyrrosia subfurfuracea-type及Pyrrosia christii-type。孢子表面纹饰特征相对稳定,在种间存在较大差异,可以为属下分类提供重要依据。 5矿质元素 采用原子吸收分光光度法(AAS)测定采自中国云南石韦属9种植物样品的矿物元素含量,分析表明,矿物元素含量在石韦属植物种间存在大幅度的变化,意味着该属每种植物都有一个固定的矿物元素的吸收和积累过程,从而可以控制植物药材及其制剂的质量、扩大天然药物的资源,并从化学分类学的角度进一步研究石韦属植物的系统分类,为研究蕨类植物的自然演化关系和化学分类学提供新的资料。 6属下分类 根据孢粉、叶形、叶脉和毛被等特征将石韦属分为2个亚属和6个组。2亚属:石韦亚属subgen. Pyrrosia和厚毛石韦亚属subgen. Porosae S. G. Lu et L. H. Yang, subgen. nov.;6组:钱币石韦组sect. Nummulariifoliae S. G. Lu et L. H. Yang, sect. nov.,石韦组sect. Pyrrosia,披针石韦组sect. Lanceolatae S. G. Lu et L. H. Yang, sect. nov.,毡毛石韦组sect. Drakeanae (Ching et Shing) S. G. Lu et L. H. Yang, stat. nov.,厚毛石韦组sect. Porosae,下延石韦组sect. Costatae (Ching et Shing) S. G. Lu et L. H. Yang, stat. nov.。综合这些特征列出属下检索表。 7种类修订 通过对国内外各大标本馆(PE, PYU, KUN, IBSC, HITBC, BM, K)馆藏标本和模式标本的研究考证,结合野外及栽培观察,综合分析有关分类学资料,对《中国植物志》上记载的38种中国产石韦属植物进行了新的分类学修订。经本次修订,确认中国产石韦属植物有27种、3变种、2变型,分属于6个组;9个名称处理为新异名:尾叶石韦P. caudifrons和佛海石韦P. fuohaiensis处理为纸质石韦P. heteractis的异名;冯氏石韦P. fengiana、相似石韦P. similis、波氏石韦P. bonii处理为布氏石韦P. boothii的异名;西南石韦P. gralla处理为华北石韦P. davidii的异名,并将其新拟为中国石韦;黄山石韦P. dimorpha处理为相近石韦P. assimilis的异名;钙生石韦P. adnascens f. calcicola处理为贴生石韦P. adnascens的异名;平绒石韦P. porosa var. mollissima处理为柔软石韦P. porosa的异名。3个名称下降为变种:贴生石韦P. adnascens和裸叶石韦P. nuda降为披针叶石韦P. lanceolata的变种;神农石韦P. shennongensis降为布氏石韦P. boothii的变种。2个降为变型:剑叶石韦P. ensata降为石韦P. lingua的变型;拟毡毛石韦P. pseudodrakeana Shing降为毡毛石韦P. drakeana的变型,另有1个种暂时存疑。 [本文引用: 1]
[15]
张宪春, 卫然, 刘红梅, 何丽娟, 王丽, 张钢民 (2013). 中国现代石松类和蕨类的系统发育与分类系统. 植物学报 48, 119-137. DOI:10.3724/SP.J.1259.2013.00119URLLycophytes and ferns are spore-bearing vascular plants, and are important groups in the evolution history of land plants. With the progress of molecular systematic, the relationships of the major groups have been resolved, and the traditional concepts have been revised. Meanwhile, new systematic arrangements have been proposed and are continuously being revised and updated. In the present review, the most recent phylogenetic studies were discussed and a new classification system of the extant lycophytes and ferns from China was also proposed. The system comprises five subclasses, 14 orders, 39 families with 12 subfamilies, and about 140 genera of lycophytes and ferns distributed in China. [本文引用: 1]
AkaikeH (1974). A new look at the statistical model identification.IEEE Trans Automat Contr 19, 716-723. DOI:10.1109/TAC.1974.1100705URL [本文引用: 1]
[18]
BurlandTG (2000). Dnastar’s lasergene sequence analysis software. In: Misener S, Krawetz SA, eds. Bioinformatics Methods and Protocols. Methods in Molecular BiologyTM, Vol. 132. Totowa, NJ: Humana Press. pp. 71-91. [本文引用: 1]
[19]
DenkT, GrimmGW (2009). Significance of pollen characteristics for infrageneric classification and phylogeny in Quercus(Fagaceae). Int J Plant Sci 170, 926-940. DOI:10.1086/600134URLPatterns of tectum ornamentation in pollen ofQuercus(oak trees, Fagaceae) are investigated using high esolution scanning electron microscopy. Tectum ornamentation is highly consistent at the infrageneric level and can be used to polarize character states withinQuercusby comparison with other genera in Fagaceae. In particular, pollen data strongly suggest the recognition of an infragenericIlexgroup and, for the first time, allow definition of the set of taxa that comprise this group. The infragenericIlexgroup displays a relatively simple tectum ornamentation exclusively found in this group of oaks, inFagus, and in extinct lineages related toTrigonobalanus,Colombobalanus, andFormanodendron.Such a simple type of tectum ornamentation is also known to represent an early developmental stage in infrageneric groups of Quercusthat have otherwise complex ornamentation (QuercusandLobatae). Ornamentation of the tectum in the infragenericCyclobalanopsisgroup can be derived from the plesiomorphicIlextype showing little accumulation of secondary sporopollenin. In the infragenericCerris,Protobalanus,Quercus, andLobataegroups, accumulation of secondary sporopollenin masks the basicIlexpattern. The five distinct types of tectum ornamentation are in perfect agreement with published molecular phylogenies ofQuercus.Thus, pollen ornamentation is a most valuable tool to identify members of the major infrageneric groups in Quercusand provides the basis for a reevaluation of the fossil record of Quercus. [本文引用: 1]
[20]
EbiharaA, NittaJH, ItoM (2010). Molecular species identification with rich floristic sampling: DNA barcoding the pteridophyte flora of Japan.PLoS One 5, e15136. DOI:10.1371/journal.pone.0015136PMID:2999545URLDNA barcoding is expected to be an effective identification tool for organisms with heteromorphic generations such as pteridophytes, which possess a morphologically simple gametophyte generation. Although a reference data set including complete coverage of the target local flora/fauna is necessary for accurate identification, DNA barcode studies including such rich taxonomic sampling on a countrywide scale are lacking. [本文引用: 2]
[21]
HallTA (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT.Nucl Acids Sym Ser 41, 95-98. [本文引用: 1]
[22]
HasebeM, OmoriT, NakazawaM, SanoT, KatoM, IwatsukiK (1994). rbcL gene sequences provide evidence for the evolutionary lineages of leptosporangiate ferns. Proc Natl Acad Sci USA 91, 5730-5734. [本文引用: 1]
[23]
HuangTC (1981). Spore Flora of Taiwan (Pteridophyta). Taiwan: National Taiwan University. pp. 52-53. [本文引用: 1]
[24]
KramerKU (1990). Dennstaedtiaceae. In: Kramer KU, Green PS, eds. The Families and Genera of Vascular Plants, Vol. I. Pteridophytes and Gymnosperms. Berlin: Springer-Verlag. pp. 81-94. [本文引用: 1]
[25]
LehtonenS, WahlbergN, ChristenhuszMJM (2012). Diversification of lindsaeoid ferns and phylogenetic uncertainty of early polypod relationships.Bot J Linn Soc 170, 489-503. DOI:10.1111/j.1095-8339.2012.01312.xURLWe analysed one nuclear gene (18S) and seven plastid markers [five protein coding (atpA, atpB, rbcL, rpoC1, rps4) and two non-coding (trnH-psbA, trnL-trnF] for 31 members of Polypodiales and four outgroup taxa. We focused our sampling on the lindsaeoids and associated ferns in order to obtain a better understanding of the diversification of the early polypods. However, the exact phylogenetic position of Saccoloma and Cystodium remained uncertain. Based on relaxed molecular clock analyses, it appears that the crown group lindsaeoids diversified in the Caenozoic, more or less simultaneously with the main radiation of other Polypodiales, even though the original divergence between the lindsaeoid and non-lindsaeoid polypods occurred before the end of the Jurassic. The current pantropical distribution of lindsaeoids can be explained by either long-distance dispersal across the oceans or vicariance caused by the retreat of previously widely distributed tropical forests from higher to lower latitudes. 0008 2012 The Linnean Society of London, Botanical Journal of the Linnean Society, 2012, 170, 489090009503. [本文引用: 1]
[26]
LiFW, KuoLY, RothfelsCJ, EbiharaA, ChiouWL, WindhamMD, PryerKM (2011). rbcL and matK earn two thumbs up as the core DNA barcode for ferns. PLoS One 6, e26597. DOI:10.1371/journal.pone.0026597PMID:3197659URLDNA barcoding will revolutionize our understanding of fern ecology, most especially because the accurate identification of the independent but cryptic gametophyte phase of the fern's life history—an endeavor previously impossible—will finally be feasible. In this study, we assess the discriminatory power of the core plant DNA barcode (rbcLandmatK), as well as alternatively proposed fern barcodes (trnH-psbAandtrnL-F), across all major fern lineages. We also present plastid barcode data for two genera in the hyperdiverse polypod clade—Deparia(Woodsiaceae) and theCheilanthes marginatagroup (currently being segregated as a new genus of Pteridaceae)—to further evaluate the resolving power of these loci. Our results clearly demonstrate the value ofmatKdata, previously unavailable in ferns because of difficulties in amplification due to a major rearrangement of the plastid genome. With its high sequence variation,matKcomplementsrbcLto provide a two-locus barcode with strong resolving power. With sequence variation comparable tomatK,trnL-Fappears to be a suitable alternative barcode region in ferns, and perhaps should be added to the core barcode region if universal primer development formatKfails. In contrast,trnH-psbAshows dramatically reduced sequence variation for the majority of ferns. This is likely due to the translocation of this segment of the plastid genome into the inverted repeat regions, which are known to have a highly constrained substitution rate. Our study provides the first endorsement of the two-locus barcode (rbcL+matK) in ferns, and favorstrnL-FovertrnH-psbAas a potential back-up locus. Future work should focus on gathering more fernmatKsequence data to facilitate universal primer development.
[27]
MillerMA, PfeifferW, SchwartzT (2010). Creating the cipres science gateway for inference of large phylogenetic trees. In: Gateway Computing Environments Workshop (GCE). New Orleans: IEEE. pp. 1-8. [本文引用: 1]
[28]
MoranRC, HanksJG, RouhanG (2007). Spore morpho- logy in relation to phylogeny in the fern genus Elaphoglossum(Dryopteridaceae). Int J Plant Sci 168, 905-929. [本文引用: 1]
[29]
NakatoN, EbiharaA (2011). Chromosome number of Microlepia hookeriana (Dennstaedtiaceae) and chromosome number evolution in the genus Microlepia. Bull Natl Mus Nat Sci Ser B 37, 75-78. [本文引用: 1]
PryerKM, SchuettpelzE, WolfPG, SchneiderH, SmithAR, CranfillR (2004). Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences.Am J Bot 91, 1582-1598. DOI:10.3732/ajb.91.10.1582PMID:21652310URLThe phylogenetic structure of ferns (= monilophytes) is explored here, with a special focus on the early divergences among leptosporangiate lineages. Despite considerable progress in our understanding of fern relationships, a rigorous and comprehensive analysis of the early leptosporangiate divergences was lacking. Therefore, a data set was designed here to include critical taxa that were not included in earlier studies. More than 5000 bp from the plastid (rbcL, atpB, rps4) and the nuclear (18S rDNA) genomes were sequenced for 62 taxa. Phylogenetic analyses of these data (1) confirm that Osmundaceae are sister to the rest of the leptosporangiates, (2) resolve a diverse set of ferns formerly thought to be a subsequent grade as possibly monophyletic (((Dipteridaceae, Matoniaceae), Gleicheniaceae), Hymenophyllaceae), and (3) place schizaeoid ferns as sister to a large clade of "core leptosporangiates" that includes heterosporous ferns, tree ferns, and polypods. Divergence time estimates for ferns are reported from penalized likelihood analyses of our molecular data, with constraints from a reassessment of the fossil record. [本文引用: 1]
[32]
PryerKM, SmithAR, HuntJS, DubuissonJY (2001). rbcL data reveal two monophyletic groups of filmy ferns(Filicopsida: Hymenophyllaceae). Am J Bot 88, 1118-1130. DOI:10.2307/2657095PMID:11410477URLThe "filmy fern" family, Hymenophyllaceae, is traditionally partitioned into two principal genera, Trichomanes s.l. (sensu lato) and Hymenophyllum s.l., based upon sorus shape characters. This basic split in the family has been widely debated this past century and hence was evaluated here by using rbcL nucleotide sequence data in a phylogenetic study of 26 filmy ferns and nine outgroup taxa. Our results confirm the monophyly of the family and provide robust support for two monophyletic groups that correspond to the two classical genera. In addition, we show that some taxa of uncertain affinity, such as the monotypic genera Cardiomanes and Serpyllopsis, and at least one species of Microtrichomanes, are convincingly included within Hymenophyllum s.l. The tubular- or conical-based sorus that typifies Trichomanes s.l. and Cardiomanes, the most basal member of Hymenophyllum s.l., is a plesiomorphic character state for the family. Tubular-based sori occurring in other members of Hymenophyllum s.l. are most likely derived independently and more than one time. While rbcL data are able to provide a well-supported phylogenetic estimate within Trichomanes s.l., they are inadequate for resolving relationships within Hymenophyllum s.l., which will require data from additional sources. This disparity in resolution reflects differential rates of evolution for rbcL within Hymenophyllaceae.
RonquistF, TeslenkoM, Van der MarkP, AyresDL, DarlingA, HohnaS, LargetB, LiuL, SuchardMA, HuelsenbeckJP (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space.Syst Biol 61, 539-542. DOI:10.1093/sysbio/sys029PMID:22357727URLAbstract Since its introduction in 2001, MrBayes has grown in popularity as a software package for Bayesian phylogenetic inference using Markov chain Monte Carlo (MCMC) methods. With this note, we announce the release of version 3.2, a major upgrade to the latest official release presented in 2003. The new version provides convergence diagnostics and allows multiple analyses to be run in parallel with convergence progress monitored on the fly. The introduction of new proposals and automatic optimization of tuning parameters has improved convergence for many problems. The new version also sports significantly faster likelihood calculations through streaming single-instruction-multiple-data extensions (SSE) and support of the BEAGLE library, allowing likelihood calculations to be delegated to graphics processing units (GPUs) on compatible hardware. Speedup factors range from around 2 with SSE code to more than 50 with BEAGLE for codon problems. Checkpointing across all models allows long runs to be completed even when an analysis is prematurely terminated. New models include relaxed clocks, dating, model averaging across time-reversible substitution models, and support for hard, negative, and partial (backbone) tree constraints. Inference of species trees from gene trees is supported by full incorporation of the Bayesian estimation of species trees (BEST) algorithms. Marginal model likelihoods for Bayes factor tests can be estimated accurately across the entire model space using the stepping stone method. The new version provides more output options than previously, including samples of ancestral states, site rates, site d(N)/d(S) rations, branch rates, and node dates. A wide range of statistics on tree parameters can also be output for visualization in FigTree and compatible software. [本文引用: 1]
[36]
SchuettpelzE, KorallP, PryerKM (2006). Plastid atpA data provide improved support for deep relationships among ferns. Taxon 55, 897-906. URL [本文引用: 3]
[37]
SchuettpelzE, PryerKM (2007). Fern phylogeny inferred from 400 leptosporangiate species and three plastid genes.Taxon 56, 1037-1050. DOI:10.2307/25065903URL [本文引用: 1]
[38]
SchwarzG (1978). Estimating the dimension of a model.Ann Statist 6, 461-464. DOI:10.1214/aos/1176344136URLProject Euclid - mathematics and statistics online
[39]
ShawJ, LickeyEB, BeckJT, FarmerSB, LiuWS, MillerJ, SiripunKC, WinderCT, SchillingEE, SmallRL (2005). The tortoise and the hare II: relative utility of 21 noncoding chloroplast DNA sequences for phylogenetic analysis.Am J Bot 92, 142-166. DOI:10.3732/ajb.92.1.142URL [本文引用: 1]
[40]
SmithAR, PryerKM, SchuettpelzE, KorallP, SchneiderH, WolfPG (2006). A classification for extant ferns.Taxon 55, 705-731. DOI:10.2307/25065646URLWe present a revised classification for extant ferns, with emphasis on ordinal and familial ranks, and a synopsis of included genera. Our classification reflects recently published phylogenetic hypotheses based on both morphological and molecular data. Within our new classification, we recognize four monophyletic classes, 11 monophyletic orders, and 37 families, 32 of which are strongly supported as monophyletic. One new family, Cibotiaceae Korall, is described. The phylogenetic affinities of a few genera in the order Polypodiales are unclear and their familial placements are therefore tentative. Alphabetical lists of accepted genera (including common synonyms), families, orders, and taxa of higher rank are provided.
[41]
Souza-ChiesTT, BittarG, NadotS, CarterL, BesinE, LejeuneB (1997). Phylogenetic analysis of Iridaceae with parsimony and distance methods using the plastid gene rps4. Plant Syst Evol 204, 109-123. [本文引用: 1]
[42]
SwoffordDL (2003). Paup*: Phylogenetic Analysis Using Parsimony, Version 4.0 b10.
[43]
TaberletP, GiellyL, PautouG, BouvetJ (1991). Universal primers for amplification of three non-coding regions of chloroplast DNA.Plant Mol Biol 17, 1105-1109. DOI:10.1007/BF00037152URL [本文引用: 1]
TateJA, SimpsonBB (2003). Paraphyly of Tarasa(Malvaceae) and diverse origins of the polyploid species. Syst Bot 28, 723-737. DOI:10.1043/02-64.1URLA molecular phylogenetic analysis of the Andean genus Tarasa (Malvaceae) and related genera yielded unexpected results regarding generic boundaries, the origins of polyploidy, and the morphological attributes of the polyploid taxa. The polyploid species of Tarasa are particularly unusual because they have life histories and floral morphologies that contradict two traditional polyploid dogmas: they are annuals and have smaller floral features (including pollen) than the diploid species. Typically, polyploids are perennial and larger than their parents. Nuclear (ITS1, 5.8S, and ITS2) and chloroplast (psbA-trnH and trnT-trnL spacers, matK-3' trnK intron) sequence data were used to reconstruct independent phylogenies to test the monophyly of the genus, determine its sister group(s), and investigate the origin of the polyploid species. Neither the nuclear nor the chloroplast phylogeny supports monophyly of Tarasa as currently circumscribed. The high Andean genus Nototriche, the North/South American disjunct genus Sphaeralcea, and Malacothammus chilensis are placed within the Tarasa clade. The polyploid species of Tarasa are not monophyletic and thus have been generated multiple times. These findings suggest that the unusual morphological features of the tetraploids are the result of convergent evolution and not shared ancestry. [本文引用: 2]
[46]
The Pteridophyte PhylogenyGroup (2016). A community-derived classification for extant lycophytes and ferns.J Syst Evol 54, 563-603. DOI:10.1111/jse.12229URLAbstract Phylogeny has long informed pteridophyte classification. As our ability to infer evolutionary trees has improved, classifications aimed at recognizing natural groups have become increasingly predictive and stable. Here, we provide a modern, comprehensive classification for lycophytes and ferns, down to the genus level, utilizing a community-based approach. We use monophyly as the primary criterion for the recognition of taxa, but also aim to preserve existing taxa and circumscriptions that are both widely accepted and consistent with our understanding of pteridophyte phylogeny. In total, this classification treats an estimated 11 916 species in 337 genera, 51 families, 14 orders, and two classes. This classification is not intended as the final word on lycophyte and fern taxonomy, but rather a summary statement of current hypotheses, derived from the best available data and shaped by those most familiar with the plants in question. We hope that it will serve as a resource for those wanting references to the recent literature on pteridophyte phylogeny and classification, a framework for guiding future investigations, and a stimulus to further discourse. [本文引用: 4]
[47]
TryonAF, LugardonB (1991). Spores of the Pteridophyta. Berlin: Springer-Verlag. pp. 1-279. [本文引用: 1]
[48]
WangFG, LiuHM, HeCM, YangDM, XingFW (2015). Taxonomic and evolutionary implications of spore ornamentation in Davalliaceae.J Syst Evol 53, 72-81. DOI:10.1111/jse.12115URLAbstract Davalliaceae is a small family of derived ferns that grow epiphytic or occasionally epilithic in the tropics and subtropics of the Old World. Despite recent taxonomic and phylogenetic achievements, some aspects of its classification remain controversial. In this study, we provided new spore observations on Davalliaceae by using scanning electron microscopy. The newly generated data were integrated together with previously reported spore evidence of this group. The evolution of spore ornamentation and taxonomic implications of spore characters was optimized onto a phylogenetic tree based on plastid sequences. Verrucate colliculate spore ornamentation was identified as plesiomorphic character state within Davalliaceae. The Humata clade was characterized by the transition from verrucate colliculate to fused verrucate ornamentation. Several species complexes subject to taxonomic controversy were found to show informative variation in the spore ornamentation. In conclusion, our observations support the utility of spores as a morphological taxonomic trait in ferns, as long as the evolution of spore morphology is considered. [本文引用: 3]
[49]
WolfPG (1995). Phylogenetic analyses of rbcL and nuclear ribosomal RNA gene sequences in Dennstaedtiaceae. Am Fern J 85, 306-327. DOI:10.2307/1547812URLSixteen new sequences of the chloroplast gene rbcL were used to assess evolutionary relationships in Dennstaedtiaceae. Sequence data from nuclear 18S rRNA genes from several dennstaedtioid ferns were also analyzed and found to support some of the inferences based on rbcL data. Of the 1860 nucleotide sites of the 18S rRNA gene examined, 215 sites (11.6%) were variable and 71 sites (3.8%) were phylogenetically informative. The 18S rRNA gene appeared to be evolving at about one ninth of the rate of rbcL, and therefore 18S data should provide increased resolution for phylogenetic studies of the early branches in fern evolution. In the context of those species sampled, the following phylogenetic patterns were evident from maximum parsimony and maximum likelihood analyses of rbcL: 1) Dennstaedtia was the only genus that was not monophyletic: Leptolepia and Microlepia diverged from within Dennstaedtia; 2) Tapeinidium did not emerge with the lindsaeoid genera, in which it is usually treated, but instead diverged at the base of the dryopteroid clade; 3) Lonchitis diverged at the base of the lindsaeoid clade, as in previous analyses of rbcL, a pattern supported by phylogenetic analysis of 18S rDNA; 4) Saccoloma emerged within the Hypolepis clade; 5) Orthiopteris (a segregate genus of Saccoloma) diverged after Dicksoniaceae but before all other clades of higher indusiate ferns; 6) Coptodipteris (usually treated as Dennstaedtia) emerged as a sister to Saccoloma; 7) Dennstaedtiaceae sensu lato appear to be polyphyletic, with the lindsaeoid genera (plus Lonchitis) emerging as a separate clade to Dennstaedtiaceae sensu stricto; 8) Monachosorum diverged from within Dennstaedtiaceae sensu stricto. [本文引用: 1]
[50]
WolfPG (1997). Evaluation of atpB nucleotide sequences for phylogenetic studies of ferns and other pteridophytes. Am J Bot 84, 1429-1440. DOI:10.2307/2446141PMID:21708550URLInferring basal relationships among vascular plants poses a major challenge to plant systematists. The divergence events that describe these relationships occurred long ago and considerable homoplasy has since accrued for both molecular and morphological characters. A potential solution is to examine phylogenetic analyses from multiple data sets. Here I present a new source of phylogenetic data for ferns and other pteridophytes. I sequenced the chloroplast gene atpB from 23 pteridophyte taxa and used maximum parsimony to infer relationships. A 588-bp region of the gene appeared to contain a statistically significant amount of phylogenetic signal and the resulting trees were largely congruent with similar analyses of nucleotide sequences from rbcL. However, a combined analysis of atpB plus rbcL produced a better resolved tree than did either data set alone. In the shortest trees, leptosporangiate ferns formed a monophyletic group. Also, I detected a wellsupported clade of Psilotaceae (Psilotum and Tmesipteris) plus Ophioglossaceae (Ophioglossum and Botrychium). The demonstrated utility of atpB suggests that sequences from this gene should play a role in phylogenetic analyses that incorporate data from chloroplast genes, nuclear genes, morphology, and fossil data. [本文引用: 1]
[51]
XuY, HuCM, HaoG (2016). Pollen morphology of Androsace(Primulaceae) and its systematic implications. J Syst Evol 54, 48-64. DOI:10.1111/jse.12149URLA close relationship between Androsace and related genera ( Douglasia , Vitaliana , and Pomatosace ) has long been recognized. Recent molecular studies have provided abundant evidence that Douglasia , Vitaliana , and Pomatosace are nested within Androsace and together constitute the monophyletic “ Androsace group”. We investigated the pollen morphology of 80 taxa representing all sections of Androsace s.s. as traditionally construed, as well as Douglasia , Vitaliana , and Pomatosace, to see whether they are congruent with phylogenetic relationships. We uncovered subtle variation in pollen morphology within the group. The shape of pollen grains ranges from spheroidal to perprolate. Pollen size ranges from 9.376508m in Androsace sect. Samuelia to 20.686508m in Douglasia . Exine ornamentation includes microreticulate, microechinate, perforate, and rugulate types. The polar view varies from circular, triangular and planaperturate, to triangular and angulaperturate. Various pollen morphological characters support the monophyly of major clades, including /Septentrionalis , /Pomatosace , /Orthocaulon , and /Megista , which were recognized previously based on molecular evidence.
[52]
YanYH, QiXP, ZhangXC (2013). Dennstaedtiaceae. In: Wu ZY, Raven PH, Hong DY, eds. Flora of China, Vol. 2-3. Pteridophytes. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press. pp. 147-168. [本文引用: 20]
[53]
YanezA, MarquezGJ, MorbelliMA (2016). Palynological analysis of Dennstaedtiaceae taxa from the paranaense phytogeographic province that produce trilete spores II: Microlepia speluncae and Pteridium arachnoideum. An Acad Bras Ciênc 88, 877-890. [本文引用: 3]
[54]
YuanY, FuL, MaCY (2012). Microlepia boluoensis sp. nov.(Dennsteadtiaceae) from Guangdong, China. Nord J Bot 30, 168-173. DOI:10.1111/j.1756-1051.2011.01318.xURLMicrolepia boluoensis Y. Yuan & L. Fu, a new species of the genus Microlepia from Luofu mountain, Boluo county, Guangdong province, China, is described and illustrated. Its gametophytic chromosome number is n =47. Microlepia boluoensis has a similar morphology to M. scyphoformis, M. speluncae and M. hancei , such as creeping rhizomes, bipinnate-tripinnatified laminas, intramarginal sori, cup-shaped indusium and trilete spores, but it has a shorter creeping rhizome, larger frond (up to 351 cm), dry chartaceous, longer stipe (up to 139 cm), 23 pairs of primary pinnae, 113 sori, and spores with a distinct and raised laesurae. [本文引用: 1]
[55]
ZhouXM, ZhangLB (2015). A classification of Selaginella(Selaginellaceae) based on molecular (chloroplast and nuclear), macromorphological, and spore features. Taxon 64, 1117-1140. DOI:10.12705/646.2URLThe largest genus of seed-free vascular plants Selaginella alone constitutes the family Selaginellaceae, the largest of the lycophyte families. The genus is estimated to contain ca. 800 species distributed on all continents except Antarctica, with the highest species diversity in tropical and subtropical regions. The monophyly of Selaginella has rarely been doubted, in contrast its infrageneric classification has been contentious. In the present study, based on chloroplast and nuclear DNA evidence, macromorphology, spore features, and/or distribution information, Selaginella is classified into six subgenera: S. subg. Selaginella, S. subg. Boreoselaginella, S. subg. Pulviniella, S. subg. Ericetorum, S. subg. Heterostachys, and S. subg. Stachygynandrum. The latter three subgenera are further classified into six, five, and seven sections, respectively. All of these infrageneric divisions, identified with molecular data, are supported by non-molecular features. A key to infrageneric taxa is given. Thirty-seven infrageneric taxa published in earlier literature are lectotypified and classified into those infrageneric taxa here recognized. A nomenclatural account of each infrageneric taxon is given.
中国蕨类植物孢子的形态VII 1 2007
... 借助电子显微镜技术的孢粉学所揭示的微形态分化是当前解决植物疑难类群分类的重要方法(张玉龙等, 1976; Tryon and Lugardon, 1991; 王全喜等, 1997; 曹建国等, 2007; 王全喜和戴锡玲, 2010; 杨鲁红, 2012; Yanez et al., 2016).分子系统学也是当前解决复杂类群物种鉴定和分类的重要方法(Hasebe et al., 1994; Wolf, 1995; Schuettpelz and Pryer, 2007; 刘红梅等, 2009).近年来, 已有不少****进行了孢粉学和分子系统学相结合的研究. 例如, 在种子植物中, 栎属(Quercus)花粉顶层纹饰形态具有明显的属下分组识别特征, 并且与分子系统发育研究具有较好的一致性(Denk and Grimm, 2009); 在点地梅属(Androsace)中, 花粉形态结合分子证据可以更好地解决该属的系统发育问题(Xu et al., 2016); 在蕨类植物中, 舌蕨属(Elaphoglossum)孢子周壁纹饰在系统发育研究中具有重要作用(Moran et al., 2007); 在卷柏属(Selaginella)中, 结合分子、形态和孢子特征等证据厘清了属下分类(Zhou and Zhang, 2015); 骨碎补科(Davalliaceae)的复合群内物种的识别也得到了孢子形态和分子证据的支持(Wang et al., 2015).但由于这些研究的样品和分子系统学样品来源各异, 仅在点地梅属和舌蕨属研究中有部分材料来源相同, 使两者的结合缺少同一性, 无法完全避免物种鉴定误差导致的结论偏差.目前鳞盖蕨属尚未开展全面的分子系统学研究, 仅在大尺度研究中涉及10种鳞盖蕨属植物样本(Wolf, 1995, 1997; Pryer et al., 2004; Schuettpelz et al., 2006; Schuettpelz and Pryer, 2007; Ebihara et al., 2010; Lehtonen et al., 2012), 在孢粉学方面也没有展开全面的研究, 仅见31种鳞盖蕨属植物孢子形态的光镜观察资料和14种扫描电镜观察资料(张玉龙等, 1976; Huang, 1981; Tryon and Lugardon, 1991; 王全喜和戴锡玲, 2010; Yuan et al., 2012; Yanez et al., 2016).因此, 该属的系统分类研究需要全面的孢粉学与分子系统学相结合且一一对应. ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}