
引用本文
贡献者
基金资助
接受日期:2017-09-24网络出版日期:2018-11-1
-->Copyright
2018《植物学报》编辑部
Contributors
History
Received:Online:
摘要:
Abstract:
Key words:
脂质不仅是微藻细胞中各种生物膜的骨架成分, 还是一种能量储存形式, 参与许多重要的生物学过程, 如叶绿体的光合作用、线粒体的电子传递及信号转导(Li-Beisson et al., 2016)。甘油酯包括极性脂和中性脂, 作为微藻脂质的主要组分, 其化学多样性与藻细胞的多种功能密切相关(Horn and Benning, 2016)。在正常生长条件下, 微藻甘油酯以极性甘油酯为主, 广泛分布于叶绿体中, 一些极性甘油酯还分布于叶绿体外的多种膜结构如线粒体、内质网及细胞质膜中, 对藻细胞的生长繁殖具有重要作用(Li et al., 2016)。在多种胁迫(光照、盐度、温度和pH等)条件下, 微藻能大量积累中性脂甘油三酯(TAG) (Zienki- ewicz et al., 2016)。极性脂与TAG在藻细胞的不同亚细胞区室内发生合成、分解与转化等多种生物化学 反应, 共同调控微藻的脂质代谢, 从而维持藻细胞的脂质内稳态(Li-Beisson et al., 2015; Allen et al., 2017)。因此, 极性甘油酯在微藻应对环境胁迫诱导的脂质代谢中发挥重要作用, 系统完整地解析微藻极性甘油酯组将有助于揭示微藻脂质代谢调控及生物学功能。
基于液相色谱及质谱的脂质组学是对脂质进行定性定量分析并在分子水平上研究脂质代谢的一种强有力的工具(Han and Gross, 2003; Li et al., 2014)。已有利用液质联用技术研究微藻脂质代谢的报道, 研究对象包括莱茵衣藻(Chlamydomonas reinhardtii) (Yang et al., 2015; Légeret et al., 2016)、三角褐指藻(Phaeodactylum tricornutum) (Abida et al., 2015)、微拟球藻(Nannochloropsis sp.)及小球藻(Chlorella sp.) (Martin et al., 2014)等, 但这些报道多局限于微藻脂质组的定性及相对定量或各脂质种类总含量的定量研究, 关于微藻各甘油酯分子的绝对定量研究较少(Popko et al., 2016; Han et al., 2017)。本研究以模式微藻莱茵衣藻为研究对象, 旨在建立一种基于超高效液相色谱-四级杆线性离子阱高性能质谱(UPLC-Q-Trap/MS)及超高效液相色谱-二维线性离子阱静电场轨道阱组合式高分辨质谱(UPLC- Orbitrap/MS2)技术的极性甘油酯定性定量分析方法, 为获得完整的莱茵衣藻极性甘油酯分子库及其分布信息提供技术平台, 进而为开展微藻脂质代谢, 尤其是极性甘油酯的动态变化及其在逆境胁迫中的响应机制研究奠定基础。
1 材料与方法1.1 藻种培养莱茵衣藻(Chlamydomonas reinhardtii L.)缺壁株CC4326购自莱茵衣藻中心(http://www.chlamycol- lection.org/)。于鼓泡式柱状光生物反应器(内径4.5 cm, 高45 cm, 培养体积为600 mL)中进行通气培养, 通气量为120 mL·min-1, CO2通入比例为2%, 单面光照强度为50 μmol·m-2·s-1, 起始接种密度为1×104个·mL-1。连续光照培养48小时后将光周期调整为12小时光照/12小时黑暗, 再继续培养48小时, 待进入光照期4小时后藻细胞密度达1×107个·mL-1, 离心收获呈指数生长的藻细胞(3 000 ×g, 4°C, 5分钟)。藻细胞经冷冻干燥后磨成粉状, 保存于-80°C冰箱中, 用于后续脂质分析。
1.2 脂质提取按照Bligh和Dyer (1959)的方法提取莱茵衣藻的脂质。用分析天平(MSE125P-1CEDI, Sartorius, Germany)准确称取5 mg上述藻粉, 加入950 μL氯仿:甲醇:水(1:2:0.8, v/v/v), 超声15分钟后振荡30秒; 加入250 μL氯仿后振荡30秒; 加入250 μL水后再振荡30秒; 离心(15 520 ×g, 2分钟)分层。将下层有机相转移至2 mL进样瓶中, 再重复提取2次, 合并有机相后用氮气吹干, 用氯仿:甲醇(1:4, v/v)复溶并定容至1 mL, 经尼龙膜(孔径0.22 μm)过滤后待用。
1.3 液相色谱分离取上述脂质提取物利用反相液相色谱(reversed phase liquid chromatography, RPLC, Waters ACQUITY UPLC)在Ascentis? Express C8色谱柱(粒径2.7 μm, 15 cm×2.1 mm I.D., Sigma-Aldrich, USA)上进行分离。二元泵系统包括流动相A及B, A为乙腈:水(60:40, v/v), B为异丙醇:乙腈(90:10, v/v), 二者均含有10 mmol·L-1甲酸铵。流速设置为0.26 mL·min-1, 柱温及样品盘温度分别设置为55°C及20°C。洗脱梯度程序设置为33分钟, 初始流动相为30%B并保持1.5分钟, 在2.5分钟内线性增加至40%, 随后在11分钟内线性增加至60%, 然后在3分钟内增加至70%, 接着在4分钟内增加至75%, 随后在4分钟内增加至97%并维持3分钟, 然后在1分钟内恢复至初始梯度30%B, 平衡3分钟, 等待下次进样。
1.4 基于UPLC-ESI-Q-Trap/MS及UPLC-ESI-Or- bitrap/MS2的莱茵衣藻甘油酯组定性分析利用UPLC-Q-Trap/MS系统(Waters ACQUITY UP- LC, AB SCIEX Q-Trap 5500)在正离子模式下对莱茵衣藻极性甘油酯种类及分子进行辨认。其中糖脂(单半乳糖甘油二酯(MGDG)、双半乳糖甘油二酯(DGDG)及硫代异鼠李糖甘油二酯(SQDG))及磷脂(磷脂酰乙醇胺(PE)、磷脂酰甘油(PG)及磷脂酰肌醇(PI))以中性丢失扫描(neutral loss scan, NL)模式进行扫描, 甜菜碱脂(N,N,N-三甲基高丝氨酸甘油二酯(DGTS))以母离子扫描(precursor ion scan, PIS)模式进行扫描(Anesi and Guella, 2015)。此外, MGDG、DGDG、SQDG、PG及PI以[M+NH4]+形式检测, PE及DGTS以[M+H]+形式检测。以上7种甘油酯的碰撞能(collision energy, CE)经优化分别设置为20、20、32、15、27、28及43 eV (表1), 每种极性甘油酯的特征碎片信息(Anesi and Guella, 2015)见表1。离子源为Turbo VTM电喷雾离子源(ESI), 离子源电压及温度分别为5 500 eV及500°C, 去簇电压100 V, 气帘气压力10 psi。利用Analyst 1.4.2 (AB SCIEX)软件采集并处理数据, 最终得到具有不同质荷比(m/z)的所有极性甘油酯分子。
表 1
Table 1
表 1
| Lipids | Classes | Adduct ion | Scanning mode | Mass of characteristic fragment (m/z) | Chemical formula of characteristic fragment | Collision energy (eV) |
|---|---|---|---|---|---|---|
| Glycolipids | MGDG | [M+NH4]+ | NL | 179 | [C6H13NO5] | 20 |
| DGDG | [M+NH4]+ | NL | 341 | [C12H23NO10] | 20 | |
| SQDG | [M+NH4]+ | NL | 261 | [C6H15NO8S] | 32 | |
| Phospholipids | PE | [M+H]+ | NL | 141 | [C2H8NO4P] | 28 |
| PG | [M+NH4]+ | NL | 189 | [C3H12NO6P] | 15 | |
| PI | [M+NH4]+ | NL | 277 | [C6H16NO9P] | 27 | |
| Betaine lipids | DGTS | [M+H]+ | PIS | 236 | [C10H22NO5]+ | 43 |
表 1
莱茵衣藻各类甘油酯中性丢失或母离子扫描参数
Table 1
Parameters of neutral loss or precursor ion scanning for the glycerolipids in Chlamydomonas reinhardtii
基于以上鉴定出的极性甘油酯分子, 利用UPLC系统经ESI与LTQ-Orbitrap XL质谱(Thermo Fisher Scientific, CA, USA)相连, 分别在正、负离子模式下通过二级碎片信息扫描鉴定具有相同质荷比但不同酰基链组成的分子种类。在正离子模式下, MGDG、DGDG及SQDG以[M+NH4]+形式检测, DGTS以[M+H]+形式检测; 在负离子模式下, PE、PG及PI以[M-H]-形式检测。质量扫描范围设置为(140-1 000) m/z。在样品检测之前, 需提前计算或确认每个极性甘油酯分子的精确质量(精确至小数点后四位)以确保高分辨率。该精确质量的查询可在LIPIDMAPS (http://www.lipidmaps.org/)数据库中进行, 同时需确定每个甘油酯分子的保留时间以设置二级碎片扫描信息。一级质谱扫描采用傅里叶变换(FTMS)模式, 二级质谱扫描采用离子阱(ion trap, IT)模式。碰撞诱导解离(collision-induced dissociation, CID)碎裂由氦气碰撞产生。MGDG及SQDG的碰撞能设置为50 eV, DGDG为70 V, PE、PG、PI及DGTS为35 eV。活化Q值(Activation Q)设为0.18。隔离宽度(isolation width)分别设置为1.0 m/z (PE、PG、PI及DGTS)及3.0 m/z (MGDG、DGDG及SQDG)。利用Xcalibur 2.1 (Ther- mo Fisher Scientific, CA, USA)软件进行数据采集。各极性甘油酯分子的脂酰基链位置分布(sn-1及sn-2位)按已发表文献中的检测结果确定(Giroud et al., 1988; Légeret et al., 2016)。
1.5 基于UPLC-ESI-Q-Trap/MS的莱茵衣藻甘油酯分子定量分析通过联合超高效液相色谱(Waters ACQUITY UPLC)及串联质谱(AB SCIEX Q-TRAP 5500), 在多级反应监测模式(multiple reaction monitoring, MRM)下对上述已鉴定出的各极性甘油酯分子进行靶向定量分析。设置的所有母离子-子离子对信息如表2所示。制备的样品浓度为5 mg·mL-1, 进样量为2 μL, 碰撞能设置同上述部分(表1)。结合每种极性甘油酯分子的保留时间, 利用Analyst 1.4.2 (AB SCIEX)软件的定量功能对采集的数据进行自动整合, 得到每种分子的峰面积积分值。利用外标法对各极性甘油酯分子含量进行绝对定量测定, 这些标准品分别为购自Matreya LLC (Pleasant Gap, PA, USA)的MGDG 18:0/18:0和DGDG 18:0/18:0以及购自Avanti Polar Lipids (Alabaster, AL, USA)的SQDG 16:0/18:3、PE 17:0/17:0、PG 17:0/17:0、PI 16:0/16:0和DGTS 16:0/16:0。藻细胞各极性甘油酯分子的绝对含量以每毫克藻细胞干重所含有分子的纳摩尔数表征(nmol·mg-1), 该绝对含量先通过相应标准品浓度(nmol·mL-1)及其峰面积之间的线性回归方程计算出目标分子浓度(nmol·mL-1), 再通过藻细胞油脂提取液的生物质浓度(mg·mL-1)计算得到。
表 2
Table 2
表 2
| Lipid species (C:N) | Theoretical mass (m/z) | Chemical formula | No. | Molecular species (sn-1/sn-2) | Precursor ion (m/z) | Product ion (m/z) |
|---|---|---|---|---|---|---|
| MGDG 34:7 | 762.5156 | C43H72NO10 | 1 | MGDG 18:3/16:4 | 762.5 | 335.2 |
| MGDG 34:6 | 764.5312 | C43H74NO10 | 2 | MGDG 18:3/16:3 | 764.5 | 335.2 |
| 3 | MGDG 18:2/16:4 | 764.5 | 337.2 | |||
| 4 | MGDG 18:4/16:2 | 764.5 | 309.2 | |||
| MGDG 34:5 | 766.5469 | C43H76NO10 | 5 | MGDG 18:3/16:2 | 766.5 | 335.2 |
| 6 | MGDG 18:2/16:3 | 766.5 | 337.2 | |||
| 7 | MGDG 18:1/16:4 | 766.5 | 339.2 | |||
| 8 | MGDG 18:4/16:1 | 766.5 | 333.2 | |||
| MGDG 34:4 | 768.5625 | C43H78NO10 | 9 | MGDG 18:1/16:3 | 768.5 | 339.2 |
| 10 | MGDG 18:2/16:2 | 768.5 | 337.2 | |||
| 11 | MGDG 18:3/16:1 | 768.5 | 311.2 | |||
| 12 | MGDG 18:4/16:0 | 768.5 | 313.2 | |||
| 13 | MGDG 18:0/16:4 | 768.5 | 341.2 | |||
| MGDG 34:3 | 770.5781 | C43H80NO10 | 14 | MGDG 18:1/16:2 | 770.5 | 339.2 |
| 15 | MGDG 18:2/16:1 | 770.5 | 337.2 | |||
| 16 | MGDG 18:3/16:0 | 770.5 | 335.2 | |||
| MGDG 34:2 | 772.5938 | C43H82NO10 | 17 | MGDG 18:1/16:1 | 772.5 | 339.2 |
| 18 | MGDG 18:0/16:2 | 772.5 | 341.2 | |||
| 19 | MGDG 18:2/16:0 | 772.5 | 313.2 | |||
| MGDG 34:1 | 774.6094 | C43H84NO10 | 20 | MGDG 18:1/16:0 | 774.6 | 339.2 |
| 21 | MGDG 18:0/16:1 | 774.6 | 341.2 | |||
| DGDG 32:3 | 904.5997 | C47H86NO15 | 22 | DGDG 16:0/16:3 | 904.5 | 313.2 |
| 23 | DGDG 16:1/16:2 | 904.5 | 311.2 | |||
| DGDG 34:7 | 924.5684 | C49H82NO15 | 24 | DGDG 18:3/16:4 | 924.5 | 335.2 |
| DGDG 34:6 | 926.5840 | C49H84NO15 | 25 | DGDG 18:3/16:3 | 926.5 | 335.2 |
| 26 | DGDG 18:2/16:4 | 926.5 | 337.2 | |||
| 27 | DGDG 18:4/16:2 | 926.5 | 333.2 | |||
| DGDG 34:5 | 928.5997 | C49H86NO15 | 28 | DGDG 18:2/16:3 | 928.5 | 337.2 |
| 29 | DGDG 18:3/16:2 | 928.5 | 335.2 | |||
| DGDG 34:4 | 930.6153 | C49H88NO15 | 30 | DGDG 18:3/16:1 | 930.6 | 335.2 |
| 31 | DGDG 18:2/16:2 | 930.6 | 337.2 | |||
| 32 | DGDG 18:1/16:3 | 930.6 | 339.2 | |||
| DGDG 34:3 | 932.6309 | C49H90NO15 | 33 | DGDG 18:1/16:2 | 932.6 | 339.2 |
| 34 | DGDG 18:3/16:0 | 932.6 | 313.2 | |||
| 35 | DGDG 18:2/16:1 | 932.6 | 337.2 | |||
| DGDG 34:2 | 934.6466 | C49H92NO15 | 36 | DGDG 18:1/16:1 | 934.6 | 339.2 |
| 37 | DGDG 18:2/16:0 | 934.6 | 337.2 | |||
| DGDG 34:1 | 936.6622 | C49H94NO15 | 38 | DGDG 18:1/16:0 | 936.6 | 339.2 |
| DGDG 34:0 | 938.6779 | C49H96NO15 | 39 | DGDG 18:0/16:0 | 938.6 | 341.2 |
| DGDG 36:4 | 958.6466 | C51H92NO15 | 40 | DGDG 18:1/18:3 | 958.6 | 339.2 |
| 41 | DGDG 18:2/18:2 | 958.6 | 337.2 | |||
| DGDG 36:3 | 960.6622 | C51H94NO15 | 42 | DGDG 18:0/18:3 | 960.6 | 341.2 |
| SQDG 32:1 | 810.5400 | C41H80NO12S | 43 | SQDG 16:0/16:1 | 810.5 | 313.2 |
| SQDG 32:0 | 812.5557 | C41H82NO12S | 44 | SQDG 16:0/16:0 | 812.5 | 313.2 |
| SQDG 34:3 | 834.5400 | C43H80NO12S | 45 | SQDG 18:3/16:0 | 834.5 | 313.2 |
| Lipid species (C:N) | Theoretical mass (m/z) | Chemical formula | No. | Molecular species (sn-1/sn-2) | Precursor ion (m/z) | Product ion (m/z) |
| SQDG 34:2 | 836.5557 | C43H82NO12S | 46 | SQDG 18:2/16:0 | 836.5 | 313.2 |
| SQDG 34:1 | 838.5713 | C43H84NO12S | 47 | SQDG 18:1/16:0 | 838.5 | 313.2 |
| SQDG 34:0 | 840.587 | C43H86NO12S | 48 | SQDG 18:0/16:0 | 840.5 | 341.2 |
| PE 32:2 | 688.4916 | C37H71NO8P | 49 | PE 16:1/16:1 | 688.4 | 547.4 |
| PE 32:1 | 690.5073 | C37H73NO8P | 50 | PE 16:0/16:1 | 690.5 | 549.5 |
| PE 32:0 | 692.5229 | C37H75NO8P | 51 | PE 16:0/16:0 | 692.5 | 551.5 |
| PE 34:4 | 712.4916 | C39H71NO8P | 52 | PE 16:0/18:3 | 714.5 | 573.5 |
| PE 34:2 | 716.5229 | C39H75NO8P | 53 | PE 34:2 | 716.5 | 575.5 |
| PE 34:1 | 718.5386 | C39H77NO8P | 54 | PE 16:0/18:1 | 718.5 | 577.5 |
| PE 36:6 | 736.4916 | C41H71NO8P | 55 | PE 18:3/18:3 | 736.4 | 595.4 |
| PE 36:5 | 738.5073 | C41H73NO8P | 56 | PE 18:2/18:3 | 738.5 | 597.5 |
| PE 36:4 | 740.5229 | C41H75NO8P | 57 | PE 36:4 | 740.5 | 599.5 |
| PE 36:3 | 742.5386 | C41H77NO8P | 58 | PE 18:0/18:3 | 742.5 | 601.5 |
| PE 36:2 | 744.5543 | C41H79NO8P | 59 | PE 18:1/18:1 | 744.5 | 603.5 |
| PG 32:2 | 736.5128 | C38H75NO10P | 60 | PG 16:1/16:1 | 736.5 | 547.5 |
| PG 32:1 | 738.5284 | C38H77NO10P | 61 | PG 16:0/16:1 | 738.5 | 549.5 |
| PG 32:0 | 740.5441 | C38H79NO10P | 62 | PG 16:0/16:0 | 740.5 | 551.5 |
| PG 34:4 | 760.5128 | C40H75NO10P | 63 | PG 18:3/16:1 | 760.5 | 571.5 |
| PG 34:3 | 762.5284 | C40H77NO10P | 64 | PG 34:3 | 762.5 | 573.5 |
| PG 34:2 | 764.5441 | C40H79NO10P | 65 | PG 34:2 | 764.5 | 575.5 |
| PG 34:1 | 766.5598 | C40H81NO10P | 66 | PG 18:1/16:0 | 766.5 | 577.5 |
| PG 34:0 | 768.5754 | C40H83NO10P | 67 | PG 18:0/16:0 | 768.5 | 579.5 |
| PI 34:3 | 850.5445 | C43H81NO13P | 68 | PI 18:3/16:0 | 850.5 | 573.5 |
| PI 34:2 | 852.5601 | C43H83NO13P | 69 | PI 34:2 | 852.5 | 575.5 |
| PI 34:1 | 854.5757 | C43H85NO13P | 70 | PI 18:1/16:0 | 854.5 | 577.5 |
| PI 34:0 | 856.5914 | C43H87NO13P | 71 | PI 18:0/16:0 | 856.5 | 579.5 |
| DGTS 32:4 | 704.5464 | C42H74NO7 | 72 | DGTS 16:0/16:4 | 704.5 | 474.4 |
| 73 | DGTS 16:3/16:1 | 704.5 | 472.4 | |||
| 74 | DGTS 16:2/16:2 | 704.5 | 470.4 | |||
| DGTS 32:3 | 706.5621 | C42H76NO7 | 75 | DGTS 16:0/16:3 | 706.5 | 474.4 |
| 76 | DGTS 16:2/16:1 | 706.5 | 470.4 | |||
| DGTS 32:2 | 708.5778 | C42H78NO7 | 77 | DGTS 16:0/16:2 | 708.5 | 474.4 |
| DGTS 32:1 | 710.5934 | C42H80NO7 | 78 | DGTS 16:0/16:1 | 710.5 | 474.4 |
| DGTS 32:0 | 712.6091 | C42H82NO7 | 79 | DGTS 16:0/16:0 | 712.6 | 474.4 |
| DGTS 34:7 | 726.5308 | C44H72NO7 | 80 | DGTS 16:4/18:3 | 726.5 | 466.4 |
| 81 | DGTS 16:3/18:4 | 726.5 | 468.4 | |||
| DGTS 34:6 | 728.5464 | C44H74NO7 | 82 | DGTS 16:3/18:3 | 728.5 | 468.4 |
| 83 | DGTS 16:4/18:2 | 728.5 | 466.4 | |||
| 84 | DGTS 16:2/18:4 | 728.5 | 494.4 | |||
| DGTS 34:5 | 730.5621 | C44H76NO7 | 85 | DGTS 16:4/18:1 | 730.5 | 500.4 |
| 86 | DGTS 16:1/18:4 | 730.5 | 472.4 | |||
| 87 | DGTS 16:3/18:2 | 730.5 | 498.4 | |||
| 88 | DGTS 16:2/18:3 | 730.5 | 470.4 | |||
| DGTS 34:4 | 732.5778 | C44H78NO7 | 89 | DGTS 16:1/18:3 | 732.5 | 496.4 |
| 90 | DGTS 16:2/18:2 | 732.5 | 498.4 | |||
| 91 | DGTS 16:3/18:1 | 732.5 | 500.4 | |||
| 92 | DGTS 16:0/18:4 | 732.5 | 474.4 | |||
| Lipid species (C:N) | Theoretical mass (m/z) | Chemical formula | No. | Molecular species (sn-1/sn-2) | Precursor ion (m/z) | Product ion (m/z) |
| 93 | DGTS 16:4/18:0 | 732.5 | 502.4 | |||
| DGTS 34:3 | 734.5934 | C44H80NO7 | 94 | DGTS 16:0/18:3 | 734.5 | 474.4 |
| DGTS 34:2 | 736.6091 | C44H82NO7 | 95 | DGTS 16:0/18:2 | 736.6 | 474.4 |
| DGTS 34:1 | 738.6247 | C44H84NO7 | 96 | DGTS 16:0/18:1 | 738.6 | 474.4 |
| 97 | DGTS 16:1/18:0 | 738.6 | 502.4 | |||
| DGTS 36:7 | 754.5621 | C46H76NO7 | 98 | DGTS 18:3/18:4 | 754.5 | 496.4 |
| DGTS 36:6 | 756.5778 | C46H78NO7 | 99 | DGTS 18:3/18:3 | 756.5 | 496.4 |
| 100 | DGTS 18:4/18:2 | 756.5 | 498.4 | |||
| DGTS 36:5 | 758.5934 | C46H80NO7 | 101 | DGTS 18:2/18:3 | 758.5 | 498.4 |
| 102 | DGTS 18:1/18:4 | 758.5 | 500.4 | |||
| DGTS 36:4 | 760.6091 | C46H82NO7 | 103 | DGTS 18:2/18:2 | 760.6 | 498.4 |
| 104 | DGTS 18:1/18:3 | 760.6 | 500.4 | |||
| 105 | DGTS 18:0/18:4 | 760.6 | 502.4 | |||
| DGTS 36:3 | 762.6247 | C46H84NO7 | 106 | DGTS 18:0/18:3 | 762.6 | 502.4 |
| 107 | DGTS 18:1/18:2 | 762.6 | 500.4 | |||
| DGTS 36:2 | 764.6404 | C46H86NO7 | 108 | DGTS 18:1/18:1 | 764.6 | 500.4 |
| 109 | DGTS 18:0/18:2 | 764.6 | 502.4 | |||
表 2
基于UPLC-Q-Trap/MS和UPLC-Orbitrap/MS2的莱茵衣藻极性甘油酯分子种类及MRM模式下的109个离子对信息
Table 2
The molecular species of polar glycerolipids in Chlamydomonas reinhardtii identified by UPLC-Q-Trap/MS and UPLC-Orbitrap/MS2 and the information of 109 ion pairs of polar lipids in MRM mode
2 结果与讨论2.1 基于UPLC-Q-Trap/MS的莱茵衣藻甘油酯分子鉴定通过UPLC-Q-Trap/MS在正离子模式下对极性脂进行一级质谱全扫描, 包括中性丢失或母离子扫描, 共得到65种具有不同质荷比的分子, 其中糖脂MGDG、DGDG及SQDG分别检出7、11和6种, 磷脂PE、PG及PI分别检出11、8和4种, 甜菜碱脂DGTS检出18种, 每种甘油酯分子种类的质荷比及分子式如表2所示。根据每种极性甘油酯分子的质荷比, 在LIPIDMAPS数据库中进行比对, 确认每种分子的碳原子及不饱和键数目, 表示为C:N (表2)。结果表明, 莱茵衣藻极性脂分子包括C32、C34及C36共3类, 其sn-1及sn-2位酰基链组合形式分别为C16/C16、C16/C18及C18/ C18 (该组合形式不区分sn-1及sn-2位置)。
2.2 基于UPLC-Orbitrap/MS2的莱茵衣藻极性甘油酯分子的酰基链解析根据上述鉴定出的65种具有不同质荷比的分子种类组成, 先利用UPLC-Orbitrap/MS在FTMS模式及正负离子模式下获得莱茵衣藻所有甘油酯分子的高分辨质谱全扫描图, 对Q-Trap/MS获得的质荷比不同的所有甘油酯分子进行再次确认及鉴定; 再利用UPLC- Orbitrap/MS2在IT模式下对上述各极性甘油酯分子进行二级碎片信息扫描。通过对每类极性甘油酯分子二级质谱(MS/MS)信息进行解析, 获得各类极性甘油酯的碎裂方式以及碰撞诱导解离(CID)碎片(表3) (Han, 2016a)。
表 3
Table 3
表 3
| Lipid class | Adduct ion | Ion mode | CE (eV) | CID fragments |
|---|---|---|---|---|
| MGDG | [M+NH4]+ | + | 50 | [M+NH4-17]+, [M+NH4-35]+, [M+NH4-179]+, [M+NH4-197]+, [M+NH4-179-RxCOOH]+ |
| DGDG | [M+NH4]+ | + | 70 | [M+NH4-17]+, [M+NH4-35]+, [M+NH4-180]+, [M+NH4-341]+, [M+NH4- 359]+, [M+NH4-341-RxCOOH]+ |
| SQDG | [M+NH4]+ | + | 50 | [M+NH4-17]+, [M+NH4-35]+, [M+NH4-261]+, [M+NH4-261-RxCO+H]+ |
| PE | [M-H]- | - | 35 | [RxCOO]-, [M-H-RxCOOH]-, [M-H-RxCO+H]- |
| PG | [M-H]- | - | 35 | [RxCOO]-, [M-H-RxCOOH]-, [M-H-RxCO+H]-, [PA-H-RxCOOH]- |
| PI | [M-H]- | - | 35 | [RxCOO]-, [M-H-RxCOOH]-, [M-H-RCxO+H]-, [M-H-R1COOH-R2- COOH]-, [PA-H-RxCOOH]-, 241 |
| DGTS | [M+H]+ | + | 35 | [M+H-RxCOOH]+, [M+H-RxCO+H]+, 236 |
表 3
莱茵衣藻甘油酯的碰接诱导解离(CID)碎片
Table 3
The collision-induced dissociation (CID) fragments of glycerolipids in Chlamydomonas reinhardtii
图1-图3为莱茵衣藻7种极性甘油酯各典型分子的二级质谱图。在所有质谱图中, 除PE外均未检测到各甘油酯分子的母离子质谱峰, 可能是由于在Orbi- trap/MS下所设置的碰撞能偏高所致。其中糖脂MGDG、DGDG及SQDG在正离子模式下分别有5、6和4种碎片离子, 我们以MGDG 18:3/16:4、DGDG 18:1/16:0及SQDG 16:0/16:0为例进行说明(图1)。首先, 3种糖脂有4种相同的碎裂方式, 分别为丢失NH3 (碎片离子m/z分别为745.51、918.83和795.50)、NH3+H2O (碎片离子m/z分别为727.48、901.78和777.55)、加NH3极性基团(碎片离子m/z分别为583.47、595.50和551.56)和同时丢失加NH3极性基团及sn-1或sn-2位脂肪酸形成的碎片离子([M+NH4- 179-RxCOOH]+、[M+NH4-341-RxCOOH]+和[M+NH4- 261-RxCO+H]+)。其次, MGDG及DGDG还有另外1种相同的碎裂方式, 即同时丢失加NH3极性基团及H2O形成的碎片离子, 其m/z分别为565.48和577.52。最后, 值得注意的是, DGDG还能同时丢失极性基团-单半乳糖基及H2O, 形成碎片离子[M+NH4-180]+。
图 1
Figure 1
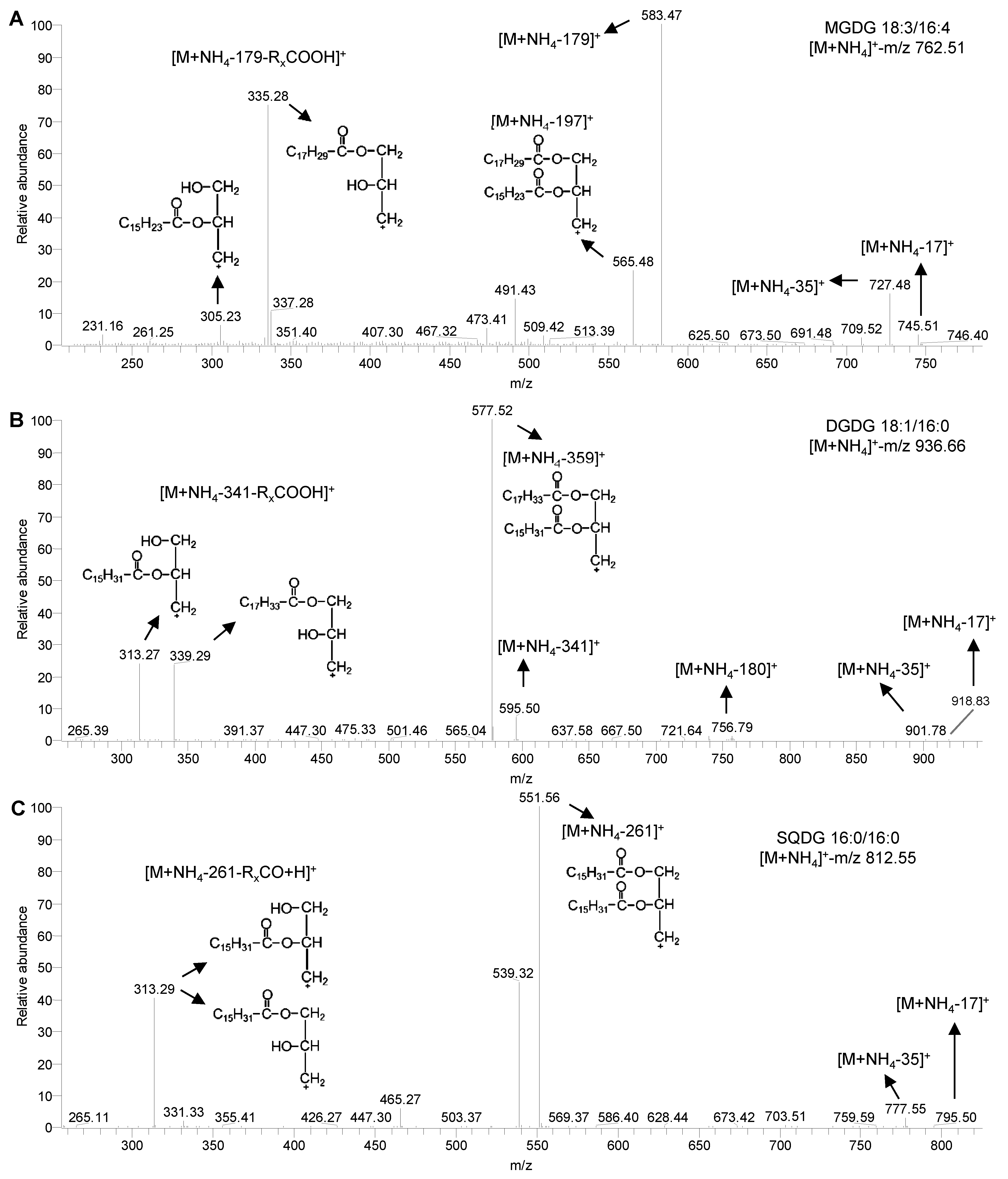
下载原图ZIP
生成PPT
图 1
莱茵衣藻的糖脂在ESI-MS/MS下的碎裂方式
(A) MGDG 18:3/16:4的二级碎裂方式; (B) DGDG 18:1/16:0的二级碎裂方式; (C) SQDG 16:0/16:0的二级碎裂方式。MGDG、DGDG和SQDG同
Figure 1
ESI-MS/MS spectra of glycolipids in Chlamydomonas reinhardtii
(A) MS/MS spectra of MGDG 18:3/16:4; (B) MS/MS spectra of DGDG 18:1/16:0; (C) MS/MS spectra of SQDG 16:0/16:0. MGDG, DGDG and SQDG see
图 1
Figure 2
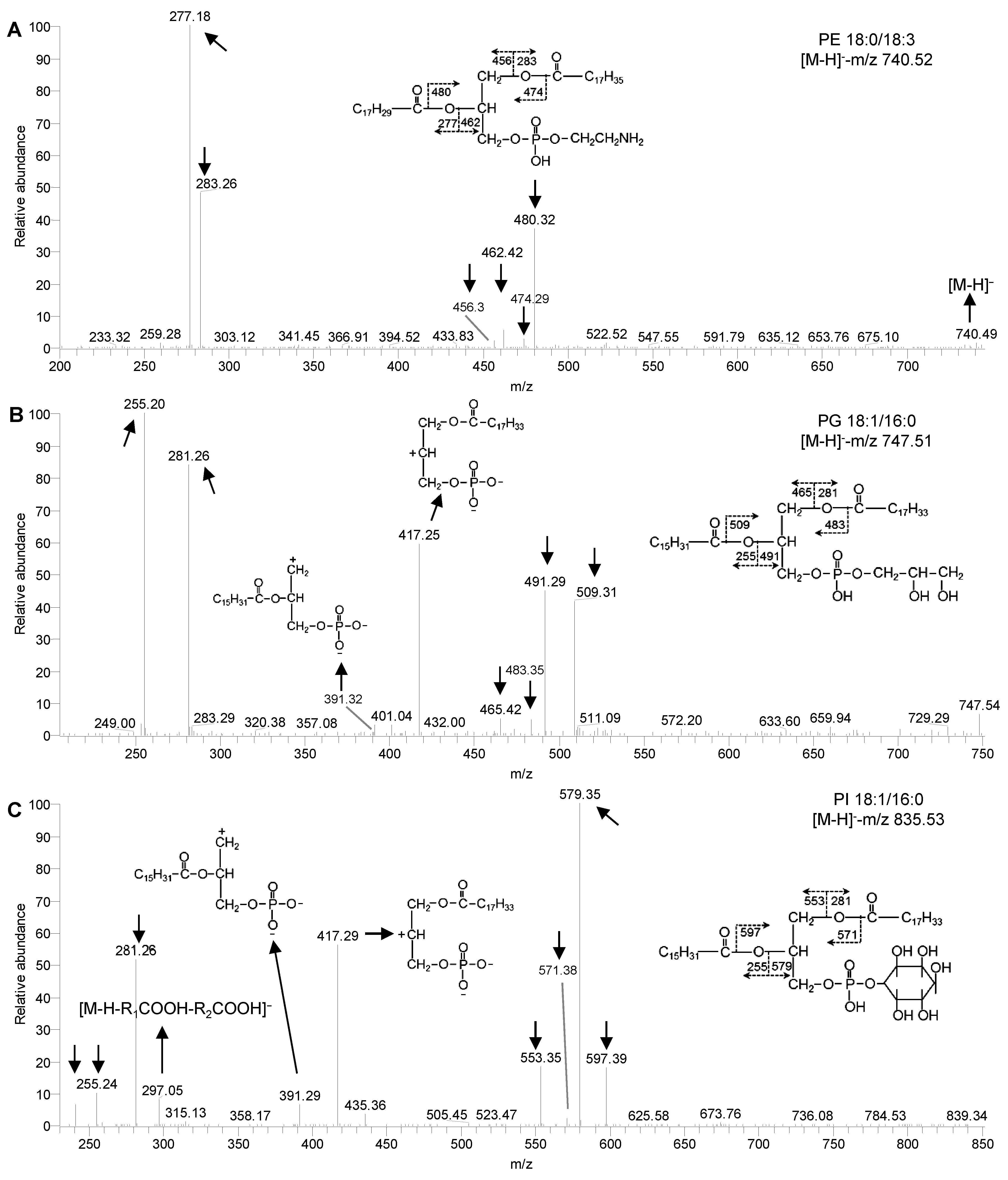
下载原图ZIP
生成PPT
图 1
莱茵衣藻的磷脂在ESI-MS/MS下的碎裂方式
(A) PE 18:0/18:3的二级碎裂方式; (B) PG 18:1/16:0的二级碎裂方式; (C) PI 18:1/16:0的二级碎裂方式。PE、PG和PI同
Figure 2
ESI-MS/MS spectra of phospholipids in Chlamydomonas reinhardtii
(A) MS/MS spectra of PE 18:0/18:3; (B) MS/MS spectra of PG 18:1/16:0; (C) MS/MS spectra of PI 18:1/16:0. PE, PG and PI see
图 3
Figure 3
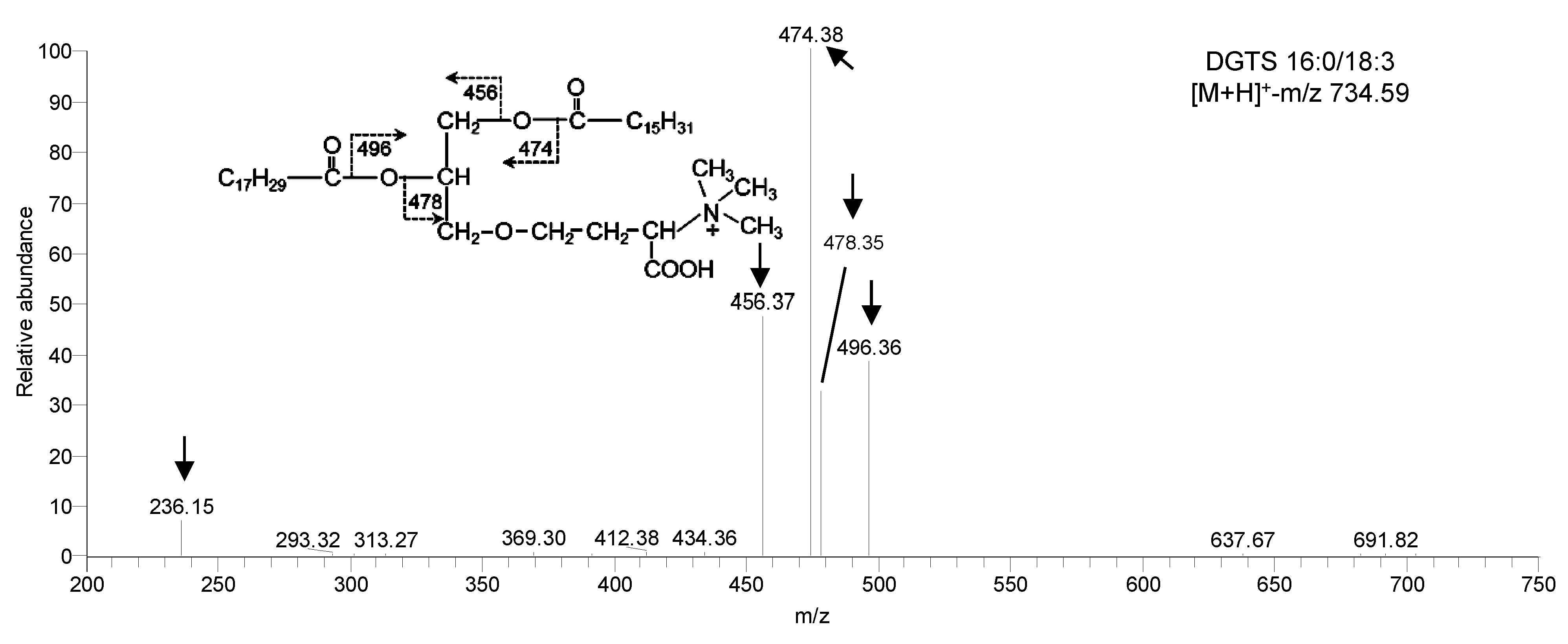
下载原图ZIP
生成PPT
图 3
莱茵衣藻的甜菜碱脂在ESI-MS/MS下的碎裂方式
DGTS同
Figure 3
ESI-MS/MS spectra of betaine lipid in Chlamydomonas reinhardtii
DGTS see
在负离子模式下, 磷脂PE、PG及PI的碎裂形式(图2)各有特点, 分别能形成3、4和6种碎片离子, 我们以PE 18:0/18:3、PG 18:1/16:0及PI 18:1/16:0为例, 其母离子均为[M-H]-。首先, 这3种磷脂均有相同的碎片离子[RxCOO]-、[M-H-RxCOOH]-及[M-H-RxCO+ H]-。其中, [RxCOO]-如图2A (-m/z 277.18及283.26)、图2B (-m/z 255.20及281.26)和图2C (-m/z 255.24及281.26)所示; [M-H-RxCOOH]-如图2A (-m/z 456.3及462.42)、图2B (-m/z 465.42及491.29)和图2C (-m/z 553.35及579.35)所示; [M-H-RxCO+H]-如图2A (-m/z 474.29及480.32)、图2B (-m/z 483.35及509.31)和图2C (-m/z 571.38及597.39)所示。其次, PG与PI还具有另外1种相同的碎片离子[PA-H-RxCOOH]-, 通过丢失1个酰基链及极性基团(甘油或肌醇基团)获得, 如图2B (-m/z 391.32及417.25)和图2C (-m/z 391.29及417.29)所示。最后, PI还有特征碎片-m/z 241, 通过丢失2个酰基链(R1CO+及R2CO+)及甘油基获得, 由磷酰基及肌醇基团组成。
在正离子模式下对甜菜碱脂DGTS进行碎裂(图3), 其碎片离子共3种, 包括丢失1个酰基链的碎片离子[M+H-RxCOOH]+及[M+H-RxCO+H]+。其中[M+H- RxCOOH]+如图3中的456.37及478.35, [M+H-RxCO+H]+如图3中的474.38及496.36; 此外, DGTS的碎片离子还包括特征碎片离子[C10H22NO5]+-m/z 236, 通过丢失2个酰基链(R1CO+及R2CO+)获得, 由甘油基及甜菜碱基组成。
通过对莱茵衣藻所有极性甘油酯种类及分子进行辨认(基于UPLC-Q-Trap/MS), 同时对极性甘油酯分子进行二级碎片信息扫描和结构解析(基于UPLC- Orbitrap/MS2), 结合相关数据库搜索及文献比对, 最终获得莱茵衣藻的极性甘油酯组整体轮廓, 其中包括7个甘油酯亚类, 即3种糖脂(MGDG、DGDG及SQ- DG)、3种磷脂(PE、PG及PI)及1种甜菜碱脂(DGTS)。通过以上分析, 共检出109种极性甘油酯分子, 包括21种MGDG、21种DGDG、6种SQDG、11种PE、8种PG、4种PI及38种DGTS (表2)。
2.3 基于UPLC-Q-Trap/MS的莱茵衣藻极性甘油酯分子的定量分析利用UPLC-Q-Trap/MS在MRM模式下采用外标法获得每一种极性甘油酯分子的含量, 每1种极性甘油酯分子的母离子-子离子对信息及提取离子色谱图(XIC)(表2; 图4)。糖脂的子离子为同时丢失加NH3极性基团及sn-1或sn-2脂肪酸后的碎片离子, 磷脂PE、PG及PI的子离子分别为中性丢失极性基团(m/z分别为141、189和277)后的碎片离子, 甜菜碱脂DGTS的子离子为丢失1个酰基链后的碎片离子([M+H-RxCO+ H]+)。我们在MRM模式下同时扫描了甘油一酯(MAG)、甘油二酯(DAG)、TAG及Lyso-甘油酯分子(图4), 由于是针对莱茵衣藻极性甘油酯的定性定量分析, 故不再提及上述分子。每种标准品浓度及其峰面积之间的线性回归方程和各自的线性范围如表4所示。藻细胞各种甘油酯分子的绝对含量通过相应标准品的线性回归方程及油脂提取液中其生物质浓度计算得出。基于Q-Trap/MS的检测灵敏度及检出限, 共准确定量检测出45种极性甘油酯分子, 包括4种MGDG、12种DGDG、4种SQDG、4种PE、6种PG、4种PI及11种DGTS。表2中其余64种离子对并未在Q-Trap/MS的MRM模式下扫描到或其含量低于定量的检测下限。结果表明, 莱茵衣藻的主要极性甘油酯为糖脂MGDG及DGDG, 其含量分别为69及37 nmol·mg-1, 二者占总极性脂含量的58%, 还包括15%的甜菜碱脂DGTS, 其含量为27 nmol·mg-1; 此外, 光合膜脂SQDG及PG含量均占总极性脂含量的9%, 磷脂PE及PI含量最低, 分别占总极性脂含量的5%及4% (图5)。
图 4
Figure 4
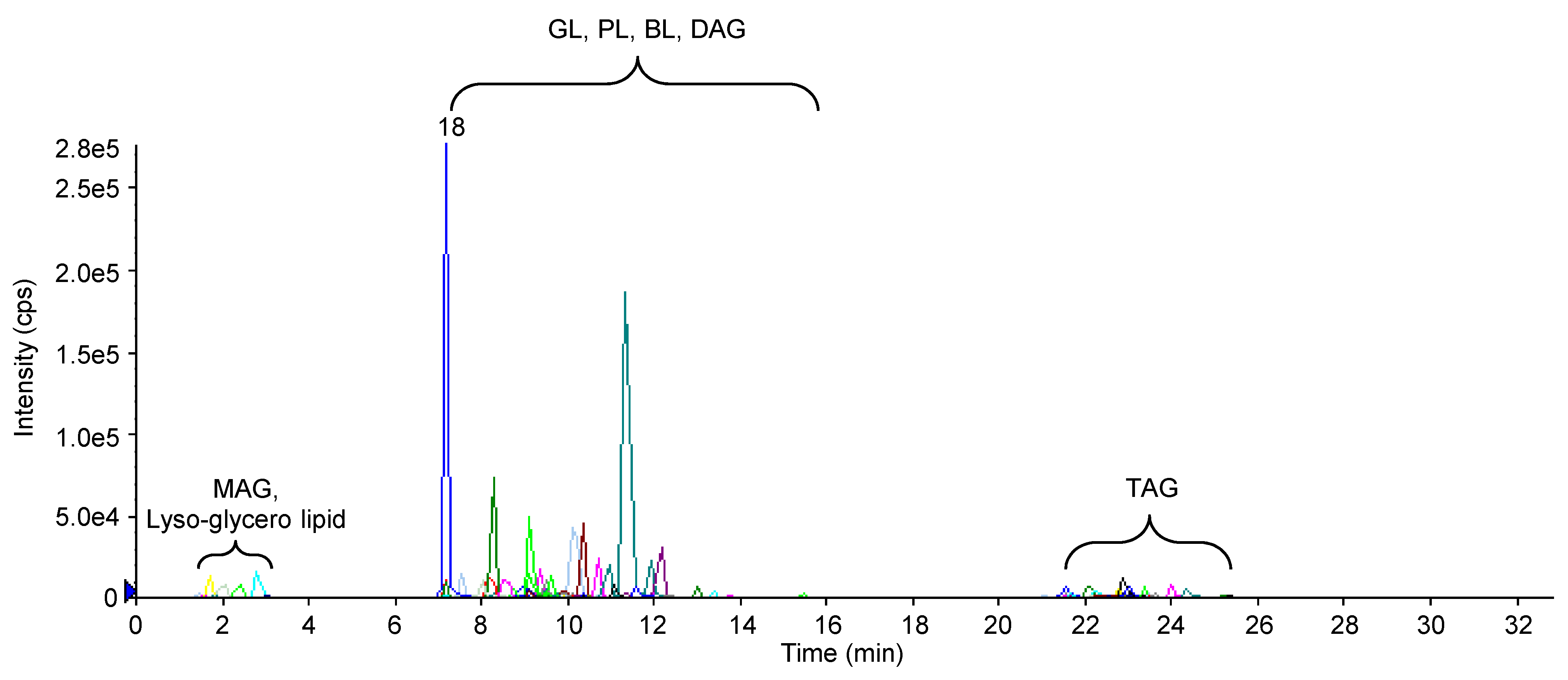
下载原图ZIP
生成PPT
图 4
正离子及MRM扫描模式下莱茵衣藻极性脂的提取离子色谱图(XIC)
MAG: 甘油一酯; GL: 糖脂; PL: 磷脂; BL: 甜菜碱脂; DAG: 甘油二酯; TAG: 甘油三酯
Figure 4
The extracted ion chromatography (XIC) of polar lipids of Chlamydomonas reinhardtii in positive and MRM scanning mode
MAG: Monoacylglycerol; GL: Glycolipid; PL: Phospholipid; BL: Betaine lipid; DAG: Diacylglycerol; TAG: Triacylglycerol
图 5
Figure 5
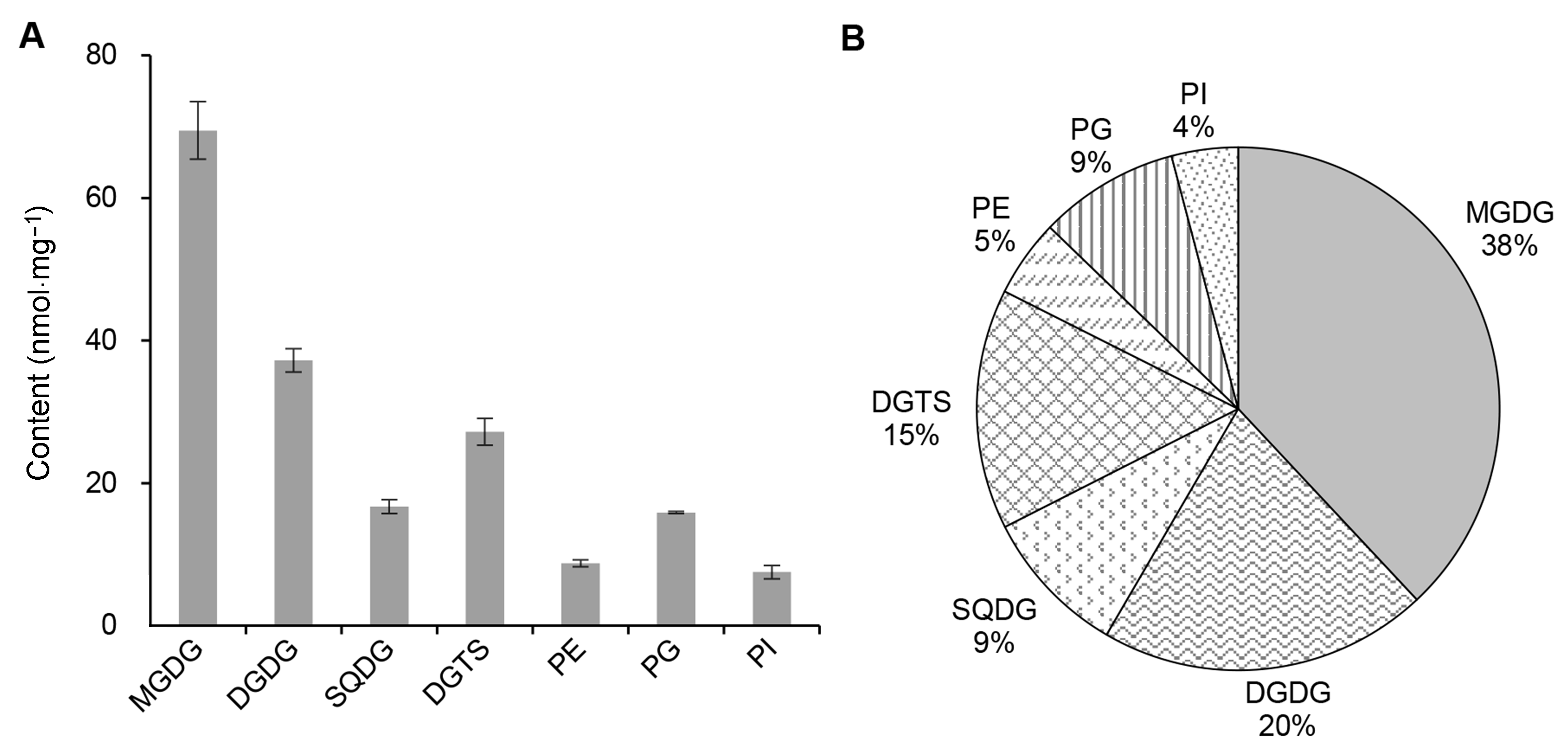
下载原图ZIP
生成PPT
图 5
莱茵衣藻极性甘油酯的组成
(A) 莱茵衣藻7种极性甘油酯的含量(nmol·mg-1); (B) 莱茵衣藻7种极性甘油酯的相对百分含量(%)。MGDG、DGDG、SQDG、PE、PG、PI和DGTS同
Figure 5
The glycerolipid components of Chlamydomonas reinhardtii
(A) The content of the individual polar glycerolipid of C. reinhardtii (nmol·mg-1); (B) The relative abundance (%) of the individual polar glycerolipid of C. reinhardtii. MGDG, DGDG, SQDG, PE, PG, PI and DGTS see
表 4
Table 4
表 4
| Lipid standard | Linear equation | Correlation coefficient (R2) | Linear range (nmol·mL-1) |
|---|---|---|---|
| MGDG 18:0/18:0 | y=6.00E-05x+2.50E-01 | 0.9981 | 0.37-39.88 |
| DGDG 18:0/18:0 | y=6.45E-04x+7.98E-01 | 0.9989 | 1.26-41.38 |
| SQDG 18:3/16:0 | y=2.18E-04x-3.30E-01 | 0.9919 | 0.02-35.95 |
| PE 17:0/17:0 | y=2.39E-04x+4.26E-01 | 0.9994 | 1.02-27.76 |
| PG 17:0/17:0 | y=9.25E-05x+6.39E-01 | 0.9971 | 0.68-26.02 |
| PI 16:0/16:0 | y=1.49E-04x+1.65E-01 | 0.9993 | 0.23-22.61 |
| DGTS 16:0/16:0 | y=1.37E-05x+8.63E-02 | 0.9998 | 1.00-28.11 |
表 4
脂质标准品浓度及其峰面积之间的线性关系
Table 4
Linear correlations between the concentrations and the peak areas of lipid standards
由图6可知, MGDG的主要分子为18:3/16:4, 其含量在MGDG所有分子含量中的比例为69%, 还包括18:2/16:3、18:3/16:3及18:2/16:4, MGDG的sn-1及sn-2位脂肪酸几乎全部为PUFA。DGDG的主要分子为18:1/16:0、18:2/16:0及18:3/16:0, 三者总含量为DGDG所有分子总含量的59%。SQDG的主要分子为16:0/16:0, 占其总分子含量的80%, sn-1及sn-2位全部为饱和脂肪酸C16:0, 此外还包括少量的18:1/ 16:0、18:2/16:0及18:3/16:0, 与DGDG具有相同的DAG碳骨架。PG的主要分子为18:2/16:1及18:3/16:1, 二者占其总分子含量的65%。
图 6
Figure 6
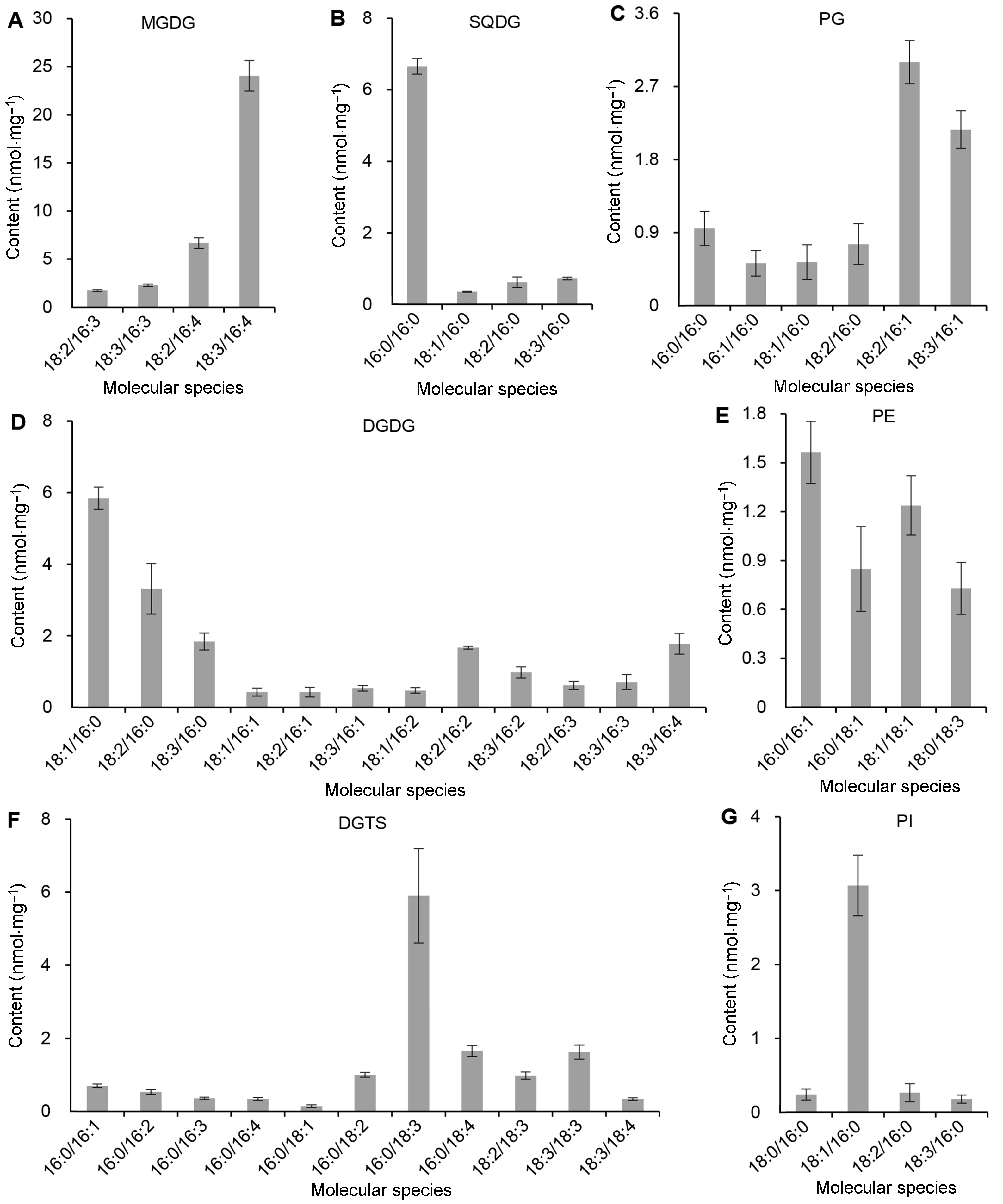
下载原图ZIP
生成PPT
图 6
莱茵衣藻各极性甘油酯分子的主要组分及其含量
(A) MGDG的各分子含量; (B) SQDG的各分子含量; (C) PG的各分子含量; (D) DGDG的各分子含量; (E) PE的各分子含量; (F) DGTS的各分子含量; (G) PI的各分子含量。MGDG、DGDG、SQDG、PE、PG、PI和DGTS同
Figure 6
The constituents and contents of each molecular species of polar lipids in Chlamydomonas reinhardtii
(A) The content of the individual molecular species of MGDG; (B) The content of the individual molecular species of SQDG; (C) The content of the individual molecular species of PG; (D) The content of the individual molecular species of DGDG; (E) The content of the individual molecular species of PE; (F) The content of the individual molecular species of DGTS; (G) The content of the individual molecular species of PI. MGDG, DGDG, SQDG, PE, PG, PI and DGTS see
莱茵衣藻的磷脂PE主要包括16:0/18:1、18:1/ 18:1及18:0/18:3, 还包括35%的16:0/16:1。PI分子几乎全部由18:1/16:0组成, 该分子占其总分子含量的82%, 此外还包括少量的18:0/16:0、18:2/16:0及18:3/16:0。甜菜碱脂DGTS的分子种类多, 包括C32、C34及C36, 其主要分子为16:0/18:3, 该分子占其总分子含量的43%, 其它还包括16:0/18:1、16:0/18:2及16:0/18:4分子。
2.4 讨论脂质组是微藻科学领域中一种新兴的重要工具(Ten- enboim et al., 2016), 在微藻中应用脂质组技术能够对微藻脂质组轮廓及其生物合成、调控、响应、重组、功能及相互作用有深入认识, 因此建立微藻脂质组学研究方法尤为重要。
脂质组学研究的核心是分析技术平台, LC-ESI- MS具有快速及高灵敏度等特点, 已广泛应用于检测识别低含量并具有重要生物功能的各类脂质分子(王涛等, 2010), 其分析流程主要包括脂质提取、基于液相色谱的脂质分离、脂质结构解析及鉴定和脂质分子的定量分析。RPLC根据脂质的脂酰基链长短和不饱和度的差异进行分离, RPLC-MS (/MS)能与绝大多数亲脂性的HPLC流动相兼容, 由于流动相含一定比例的水, 故质谱分析时雾化效果好, 对包括极性及非极性在内的所有脂质组分均能良好地保留。本研究采用RPLC-MS (/MS)对莱茵衣藻的极性甘油酯组进行了定性定量分析, 极大地丰富了微藻脂质组学的研究内容(t’Kindt et al., 2012)。此外, 脂质结构“组成模块”(building block)的特点是进行脂质结构鉴定的基础。特定的MS/MS信息(表征极性头基和酰基链等组成模块) (Murphy and Axelsen, 2011)、保留时间规律(Brouwers, 2011)及精确质量是脂质鉴定需要综合考虑的因素。脂质鉴定包括一级质谱分子离子鉴定及二级质谱碎片离子解析。目前基于组学的定量包括2种, 即相对定量及绝对定量。在脂质组学中, 相对定量测定脂质组中各脂质分子的类型变化及相对丰度变化, 能表征脂质组在刺激物作用下的响应变化, 同时有利于生物标志物的筛选; 绝对定量是基于外标(用与目标化合物相同的标准品进行平行测定)或内标(加入样品中不存在的标准品进行同时测定)测定每种脂质分子、脂质亚类及整个脂质组的质量含量变化, 而脂质分子质量含量的动态变化有利于代谢途径的构建或生物化学机制的解析(Han, 2016b)。本研究采用外标法对莱茵衣藻各甘油酯分子进行了绝对定量, 有助于微藻脂质代谢的深入研究。
本研究利用UPLC-Q-Trap/MS进行一级质谱分子离子鉴定及基于MRM模式的甘油酯分子靶向定量分析。基于QQQ的脂质分析包括母离子扫描、子离子扫描及中性丢失扫描, 是“自下而上”(Bottom-up)的脂质组学分析典型方法。利用微藻各类甘油酯特定的中性丢失或母离子扫描模式不仅可以准确辨认出具有不同质荷比的各类甘油酯分子(表1), 即一级结构鉴定; 而且还可通过非靶向方法获取MRM离子对, 进行复杂样品中目标化合物的靶向定量分析, 即“拟靶标”代谢组学方法(Chen et al., 2013; Zhang et al., 2016), 其优势在于定量分析的灵敏度、稳定性、重复性较好及较宽的线性范围(104-105), 同时数据处理及定量分析过程简单(Chen et al., 2013)。目前, 基于Q-Trap/MS的脂质组学研究集中于哺乳动物(Overgaard et al., 2016; Slatter et al., 2016)及高等植物(Tarazona et al., 2015), 在微藻中的应用还很少(Meng et al., 2017)。本研究利用Q-Trap/MS的MRM模式能快速靶向定量分析莱茵衣藻各极性甘油酯分子的质量含量, 从而为微藻脂质代谢的动态变化及新途径的构建奠定基础。
Orbitrap/MS (Zubarev and Makarov, 2013)是近年来新开发出的一种具有高分辨率、高灵敏度、高准确度的质谱, 与普通质谱相比, 它在扫描碎片离子、分析鉴别各种脂质同分异构体方面具有明显优势(Hu et al., 2005; Taguchi and Ishikawa, 2010)。本研究所用的LTQ Orbitrap/MS质量分辨率高达240 000 FWHM (full width half maximum, 半峰处全峰宽), 能进行精确的质量测定(精度为1-3 mg·L-1), 同时快速采集高分辨率的质谱全扫描谱图, 并基于不同脂质类别的特定元素组成进行脂质鉴定, 是一种高通量的“自上而下”(Top-down)的脂质组学分析模式(Schuhmann et al., 2011)。此外, LTQ Orbitrap/MS作为一种高分辨率质谱, 同时具备FTMS和IT模式。这2种检测模式可进行快速切换, 并具备多级质谱的扫描功能(Eliuk and Makarov, 2015), 目前已广泛应用于蛋白质(Geiger et al., 2010)、多糖(Rohmer et al., 2011)、类胡萝卜素(Bijttebier et al., 2013)及番茄碱(Caprioli et al., 2015)等多类复杂化合物的二级及多级结构鉴定。而基于Orbitrap/MS的甘油酯组二级及多级结构鉴定研究还较少。本研究利用Orbitrap/ MS2采集各甘油酯分子二级质谱的碎片离子信息, 确定了莱茵衣藻各极性甘油酯分子的二级结构, 为其定量分析提供了准确可靠的信息。
综上所述, 在脂质组的定性定量分析中, Q-Trap/ MS兼具一级结构鉴定(母离子及中性丢失扫描)及靶向定量分析(MRM模式)的优势, 而Orbitrap/MS2的高质量分辨率性能使其更适于进行脂质分子二级结构的准确鉴定。本研究中莱茵衣藻脂质经UPLC的反相色谱分离、ESI离子化、Q-Trap/MS及Orbitrap/MS2检测, 确定了109种极性甘油酯分子种类及结构谱图。基于以上极性甘油酯组轮廓, 又通过UPLC-Q- Trap/MS的MRM模式定量分析出莱茵衣藻的45种极性甘油酯分子含量。虽然Q-Trap/MS并未全部扫描并定量检出基于Orbitrap/MS2定性得到的所有极性脂分子, 但研究所获得的定性定量信息可以完善目前现有的莱茵衣藻极性甘油酯组(Yang et al., 2015; Légeret et al., 2016), 从而为进一步研究其合成机制及生物学功能奠定技术基础。
根据以上定性定量分析结果可知, 莱茵衣藻的极性脂以糖脂MGDG、DGDG及甜菜碱脂DGTS为主, 还包括光合膜脂SQDG、PG及磷脂PE、PI。这些极性脂的组成比例与Li-Beisson等(2015)的研究结果一致, 同时各类极性脂的分子种类及酰基组成各有特点, 这也表明其合成途径及生物学功能各不相同。在高等植物中, 甘油酯通过真核或原核途径合成(Roughan and Slack, 1982)。在真核途径中, 脂肪酸由叶绿体合成后被转运至叶绿体外区室形成酰基辅酶A, 然后在内质网中进一步合成膜脂。在内质网中合成的膜脂再返回至叶绿体并转化成DAG, 随后DAG在叶绿体被膜中被进一步糖基化合成MGDG及DGDG, 高等植物通过真核途径合成甘油酯的sn-2位脂肪酸为C18脂肪酸; 相比之下其原核途径几乎全部发生于叶绿体中, 叶绿体合成的脂肪酸先与酰基载体蛋白(ACP)结合, 再合成脂质前体物质磷脂酸(PA)和DAG, 最后合成膜脂, 这些通过原核途径合成甘油酯的sn-2位脂肪酸为C16脂肪酸(Warakanont et al., 2015)。本研究中, 莱茵衣藻的MGDG、DGDG、SQDG、PG及PI的sn-2位脂肪酸全部为C16脂肪酸, 这表明莱茵衣藻的光合膜脂及磷脂PI全部通过原核途径合成; 与此相反, 莱茵衣藻DGTS与PE的sn-2位脂肪酸同时含有C16及C18脂肪酸, 表明这2种膜脂的合成既包括原核途径又包括真核途径(Giroud et al., 1988)。莱茵衣藻甘油酯合成途径与高等植物不同, 这也表明了植物在进化上的多样性。
此外, DGDG、SQDG及PI这3种甘油酯均含有相同的DAG碳骨架分子18:1/16:0、18:2/16:0及18:3/ 16:0, 表明其作为sn-1位C18脂肪酸去饱和的载体; 而DGTS中16:0/18:1、16:0/18:2、16:0/18:3及16:0/ 18:4分子的存在表明其作为sn-2位C18脂肪酸去饱和的载体, 这与Schlapfer和Eichenberger (1983)的研究结果相一致。虽然这4种甘油酯均为C18脂肪酸去饱和的载体, 在莱茵衣藻的生长代谢过程中具有重要的生理功能, 但是DGTS与DGDG、SQDG和PI的具体作用机制可能因脂肪酸sn位的差异而不同。研究表明, 在微拟球藻及高等植物中, PC是C18脂肪酸去饱和的载体, 与其它极性脂的合成密切相关(Botella et al., 2017; Meng et al., 2017), 而DGTS与PC的结构及生物物理性质类似, 因此二者在脂质代谢过程中的生理功能可能也相似(Liu and Benning, 2013)。这些结果也进一步丰富了微藻的脂质代谢信息。
3 结论本研究利用液质联用技术鉴定了莱茵衣藻的极性甘油酯组成, 并对其进行了定量分析。通过分析莱茵衣藻极性脂组分的特征碎片离子与裂解规律, 共获得109种具有不同酰基链组成的极性脂分子。此外, 通过MRM模式对各极性甘油酯分子进行靶向定量分析, 确定其主要组分为糖脂MGDG、DGDG及甜菜碱脂DGTS。莱茵衣藻各极性脂分子的酰基链组成特点表明, 其极性脂通过专一性原核途径或混合型原核及真核途径合成, 同时多种极性脂包括DGDG、SQDG、PI及DGTS, 它们均具有C18脂肪酸去饱和载体的功能。综上所述, 本研究所建立的莱茵衣藻极性甘油酯组定性定量分析方法, 可为获得完整的极性甘油酯分子库及其分布信息提供技术平台, 并为进一步开展微藻脂质代谢, 尤其是极性甘油酯的动态变化及其在逆境胁迫中的响应机制研究奠定基础。
参考文献
文献选项
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | DOI:10.3969/j.issn.1674-3466.2010.02.015URL脂质不仅是生物膜的骨架成分和能量贮存物质,越来越多的证据表明,脂质也参与细胞的许多重要功能。脂质组学是代谢组学的一个重要分支,主要研究生物体内所有的脂质分子的特性以及它们在蛋白质表达和基因调控过程中的作用。脂质组学是依赖技术驱动的科学。近年来,随着人们对脂质研究的重视,脂质组学研究方法和策略有了突破性进展,在动物上开发出的脂质组学分析方法已经扩展应用到植物上。该文重点介绍脂质组学的研究方法及其应用,以期推动脂质组学,特别是植物脂质组学的进一步发展。 [本文引用: 1] |
| [2] | [本文引用: 1] |
| [3] | DOI:10.1074/jbc.M116.760843PMID:5217694URLDeriving biofuels and other lipoid products from algae is a promising future technology directly addressing global issues of atmospheric CO 2 balance. To better understand the metabolism of triglyceride synthesis in algae, we examined their metabolic origins in the model species, Coccomyxa subellipsoidea C169, using stable isotopic labeling. Labeling patterns arising from [U- 13 C]glucose, 13 CO 2 , or D 2 O supplementation were analyzed by GC-MS and/or LC-MS over time courses during nitrogen starvation to address the roles of catabolic carbon recycling, acyl chain redistribution, and de novo fatty acid (FA) synthesis during the expansion of the lipid bodies. The metabolic origin of stress-induced triglyceride was found to be a continuous 8:2 ratio between de novo synthesized FA and acyl chain transfer from pre-stressed membrane lipids with little input from lipid remodeling. Membrane lipids were continually synthesized with associated acyl chain editing during nitrogen stress, in contrast to an overall decrease in total membrane lipid. The incorporation rates of de novo synthesized FA into lipid classes were measured over a time course of nitrogen starvation. The synthesis of triglycerides, phospholipids, and galactolipids followed a two-stage pattern where nitrogen starvation resulted in a 2.5-fold increase followed by a gradual decline. Acyl chain flux into membrane lipids was dominant in the first stage followed by triglycerides. These data indicate that the level of metabolic control that determines acyl chain flux between membrane lipids and triglycerides during nitrogen stress relies primarily on the Kennedy pathway and de novo FA synthesis with limited, defined input from acyl editing reactions. [本文引用: 1] |
| [4] | DOI:10.1016/j.chroma.2015.01.035PMID:25655586URLIn this paper, we report the development of a new method based on HILIC-ESI-MS for the separation of several different membrane lipid classes and their detection on a triple quadrupole mass spectrometer using Precursor Ion (PIS) and Neutral Loss (NL) scanning in positive ion mode. Four different columns were tested for their ability to separate, under different conditions, a mixture of 14 lipid standards containing 7 glycerophospholipids (GPL), 2 glycosphyngolipids (GSL), 3 glycolipids (GL) and 2 betaine lipids (BL). The best separation was obtained using a Lichrosphere DIOL column as stationary phase and water (10mm ammonium acetate)/acetonitrile gradient elution as mobile phase which allows the separation of the 14 lipid classes within 35min runtime. Our method was successfully tested for the separation and analysis of crude lipid extracts obtained from a green alga (Jaoa bullata), a dinoflagellate (Peridinium cinctum) and a plant (Vitis vinifera cv. Corvina) [本文引用: 2] |
| [5] | DOI:10.1002/jms.3203PMID:23722965URLVegetables are a major source of carotenoids and carotenoids are identified as potentially important natural antioxidants that may aid in the prevention of several human chronic degenerative diseases. Characterization of carotenoids in organic biological matrices is a crucial step in any research valorization trajectory. This study reports for the first time the use of high mass resolution and exact mass orbitrap technology for the elucidation of carotenoid fragmentation pathways. This contributes to the generation of new tools for identifying unknown carotenoids based on fragmentation patterns. Two different chromatographic methods making use of different mobile phases resulted in the generation of different ion species because of the large influence of the mobile phase solvent composition on ionization. It was shown that depending on the molecular ion species that are generated (protonated ions or radical molecular ions), different fragments are formed when applying higher energy collisional dissociation. Fragmentation and the abundance of fragments provide valuable structural information on the type of functional groups, the polyene backbone and the location of double bonds in ring structures of carotenoids. Furthermore, coherence between specific substructures in the molecules and characteristic fragmentation patterns was observed allowing the assignment of fragmentation patterns for carotenoid substructures that can theoretically be extrapolated to carotenoids with similar (sub)structures. Differentiation between isomeric carotenoids by compound specific fragments could however not be made for all the isomeric groups under study. As a wide variety of isomeric forms of carotenoids exist in nature, the combination of good chromatographic separation with high resolution mass spectrometry and other complementary qualitative structure elucidation techniques such as a photo diode array detector and/or nuclear magnetic resonance spectroscopy are indispensable for unambiguous identification of unknown carotenoids. Copyright 2013 John Wiley & Sons, Ltd. [本文引用: 1] |
| [6] | DOI:10.1139/y59-099URL [本文引用: 1] |
| [7] | DOI:10.1016/j.plipres.2016.11.001PMID:27871883URLIn plant cells, phosphatidylcholine (PC) is a major glycerolipid of most membranes but practically lacking from the plastid internal membranes. In chloroplasts, PC is absent from the thylakoids and the inner envelope membrane. It is however the main component of the outer envelope membrane, where it exclusively distributes in the outer monolayer. This unique distribution is likely related with operational compartmentalization of plant lipid metabolism. In this review, we summarize the different mechanisms involved in homeostasis of PC in plant cells. The specific origin of chloroplast PC is examined and the involvement of the P4-ATPase family of phospholipid flippases (ALA) is considered with a special attention to the recently reported effect of the endoplasmic reticulum-localized ALA10 on modification of chloroplast PC desaturation. The different possible roles of chloroplast PC are then discussed and analyzed in consideration of plant physiology. [本文引用: 1] |
| [8] | DOI:10.1016/j.bbalip.2011.08.001PMID:21851861URL78 Advantages of liquid chromatography in lipidomic analysis. 78 Added value of other ionization techniques than ESI. 78 Normal- and reverse phase separations. 78 How to deal with ion suppression in quantification. [本文引用: 1] |
| [9] | [本文引用: 1] |
| [10] | DOI:10.1021/ac4016787PMID:23889541URLUntargeted analysis performed using full-scan mass spectrometry (MS) coupled with liquid chromatography (LC) is commonly used in metabolomics. Although they are commonly employed, full-scan MS methods such as quadrupole-time-of-flight (Q-TOF) MS have been restricted by various factors including their limited linear range and complicated data processing. LC coupled with triple quadrupole (QQQ) MS operated in the multiple reaction monitoring (MRM) mode is the gold standard for metabolite quantification; however, only known metabolites are generally quantified, limiting its applications in metabolomic analysis. In this study, a pseudotargeted approach was proposed to perform serum metabolomic analysis using an ultra high-performance liquid chromatography (UHPLC)/QQQ MS system operated in the MRM mode, for which the MRM ion pairs were acquired from the serum samples through untargeted tandem MS using UHPLC/Q-TOF MS. The UHPLC/QQQ MRM MS-based pseudotargeted method displayed better repeatability and wider linear range than the traditional UHPLC/Q-TOF MS-based untargeted metabolomics method, and no complicated peak alignment was required. The developed method was applied to discover serum biomarkers for patients with hepatocellular carcinoma (HCC). Patients with HCC had decreased lysophosphatidylcholine, increased long-chain and decreased medium-chain acylcarnitines, and increased aromatic and decreased branched-chain amino acid levels compared to healthy controls. The novelty of this work is that it provides an approach to acquire MRM ion pairs from real samples, is not limited to metabolite standards, and it provides a foundation to achieve pseudotargeted metabolomic analysis on the widely used LC/QQQ MS platform. [本文引用: 2] |
| [11] | DOI:10.1146/annurev-anchem-071114-040325PMID:26161972URLAbstract We discuss the evolution of Orbitrap mass spectrometry (MS) from its birth in the late 1990s to its current role as one of the most prominent techniques for MS. The Orbitrap mass analyzer is the first high-performance mass analyzer that employs trapping of ions in electrostatic fields. Tight integration with the ion injection process enables the high-resolution, mass accuracy, and sensitivity that have become essential for addressing analytical needs in numerous areas of research, as well as in routine analysis. We examine three major families of instruments (related to the LTQ Orbitrap, Q Exactive, and Orbitrap Fusion mass spectrometers) in the context of their historical development over the past ten eventful years. We discuss as well future trends and perspectives of Orbitrap MS. We illustrate the compelling potential of Orbitrap-based mass spectrometers as (ultra) high-resolution platforms, not only for high-end proteomic applications, but also for routine targeted analysis. [本文引用: 1] |
| [12] | DOI:10.1074/mcp.M110.001537PMID:20610777URLThe orbitrap mass analyzer combines high sensitivity, high resolution, and high mass accuracy in a compact format. In proteomics applications, it is used in a hybrid configuration with a linear ion trap (LTQ-Orbitrap) where the linear trap quadrupole (LTQ) accumulates, isolates, and fragments peptide ions. Alternatively, isolated ions can be fragmented by higher energy collisional dissociation. A recently introduced stand-alone orbitrap analyzer (Exactive) also features a higher energy collisional dissociation cell but cannot isolate ions. Here we report that this instrument can efficiently characterize protein mixtures by alternating MS and "all-ion fragmentation" (AIF) MS/MS scans in a manner similar to that previously described for quadrupole time-of-flight instruments. We applied the peak recognition algorithms of the MaxQuant software at both the precursor and product ion levels. Assignment of fragment ions to co-eluting precursor ions was facilitated by high resolution (100,000 at m/z 200) and high mass accuracy. For efficient fragmentation of different mass precursors, we implemented a stepped collision energy procedure with cumulative MS readout. AIF on the Exactive identified 45 of 48 proteins in an equimolar protein standard mixture and all of them when using a small database. The technique also identified proteins with more than 100-fold abundance differences in a high dynamic range standard. When applied to protein identification in gel slices, AIF unambiguously characterized an immunoprecipitated protein that was barely visible by Coomassie staining and quantified it relative to contaminating proteins. AIF on a benchtop orbitrap instrument is therefore an attractive technology for a wide range of proteomics analyses. [本文引用: 1] |
| [13] | [本文引用: 2] |
| [14] | [本文引用: 1] |
| [15] | |
| [16] | [本文引用: 2] |
| [17] | DOI:10.1194/jlr.R300004-JLR200URL [本文引用: 1] |
| [18] | DOI:10.1126/science.aaf6206PMID:27634522URLLipids and oils derived from plant and algal photosynthesis constitute much of human daily caloric intake and provide the basis for high-energy bioproducts, chemical feedstocks for countless applications, and even fossil fuels over geological time scales. Sustainable production of high-energy compounds from plants is essential to preserving fossil fuel sources and ensuring the well-being of future generations. As a result of progress in basic research on plant and algal lipid metabolism, in combination with advances in synthetic biology, we can now tailor plant lipids for desirable biological, physical, and chemical properties. We highlight recent advances in plant lipid translational biology and discuss untapped areas of research that might expand the application of plant lipids. [本文引用: 1] |
| [19] | DOI:10.1002/jms.856PMID:15838939URLAbstract Research areas such as proteomics and metabolomics are driving the demand for mass spectrometers that have high performance but modest power requirements, size, and cost. This paper describes such an instrument, the Orbitrap, based on a new type of mass analyzer invented by Makarov. The Orbitrap operates by radially trapping ions about a central spindle electrode. An outer barrel-like electrode is coaxial with the inner spindlelike electrode and mass/charge values are measured from the frequency of harmonic ion oscillations, along the axis of the electric field, undergone by the orbitally trapped ions. This axial frequency is independent of the energy and spatial spread of the ions. Ion frequencies are measured non-destructively by acquisition of time-domain image current transients, with subsequent fast Fourier transforms (FFTs) being used to obtain the mass spectra. In addition to describing the Orbitrap mass analyzer, this paper also describes a complete Orbitrap-based mass spectrometer, equipped with an electrospray ionization source (ESI). Ions are transferred from the ESI source through three stages of differential pumping using RF guide quadrupoles. The third quadrupole, pressurized to less than 10(-3) Torr with collision gas, acts as an ion accumulator; ion/neutral collisions slow the ions and cause them to pool in an axial potential well at the end of the quadrupole. Ion bunches are injected from this pool into the Orbitrap analyzer for mass analysis. The ion injection process is described in a simplified way, including a description of electrodynamic squeezing, field compensation for the effects of the ion injection slit, and criteria for orbital stability. Features of the Orbitrap at its present stage of development include high mass resolution (up to 150,000), large space charge capacity, high mass accuracy (2-5 ppm), a mass/charge range of at least 6000, and dynamic range greater than 10(3). Applications based on electrospray ionization are described, including characterization of transition-metal complexes, oligosaccharides, peptides, and proteins. Use is also made of the high-resolution capabilities of the Orbitrap to confirm the presence of metaclusters of serine octamers in ESI mass spectra and to perform H/D exchange experiments on these ions in the storage quadrupole. Copyright 2005 John Wiley & Sons, Ltd. [本文引用: 1] |
| [20] | DOI:10.1111/pce.12656PMID:26477535URLAbstract Studying how photosynthetic cells modify membrane lipids in response to heat stress is important to understand how plants and microalgae adapt to daily fluctuations in temperature and to investigate new lipid pathways. Here, we investigate changes occurring in lipid molecular species and lipid metabolism genes during early response to heat stress in the model photosynthetic microorganism Chlamydomonas reinhardtii . Lipid molecular species analyses revealed that, after 6065min at 4265°C, a strong decrease in specific polyunsaturated membrane lipids was observed together with an increase in polyunsaturated triacylglycerols (TAGs) and diacylglycerols (DAGs). The fact that decrease in the major chloroplastic monogalactosyldiacylglycerol sn1 -18:3/ sn2 -16:4 was mirrored by an accumulation of DAG sn1 -18:3/ sn2 -16:4 and TAG sn1 -18:3/ sn2 -16:4/ sn3 -18:3 indicated that newly accumulated TAGs were formed via direct conversion of monogalactosyldiacylglycerols to DAGs then TAGs. Lipidomic analyses showed that the third fatty acid of a TAG likely originated from a phosphatidylethanolamine or a diacylglyceryl- O -4′-(N,N,N,-trimethyl)-homoserine betaine lipid species. Candidate genes for this TAG synthesis pathway were provided through comparative transcriptomic analysis and included a phospholipase A2 homolog and the DAG acyltransferase DGTT1. This study gives insights into the molecular events underlying changes in membrane lipids during heat stress and reveals an alternative route for TAG synthesis. [本文引用: 3] |
| [21] | [本文引用: 1] |
| [22] | DOI:10.1016/j.tplants.2015.10.011PMID:26616197URLFAX1, a novel membrane protein in the inner envelope of chloroplasts, mediates FA export. The discovery and analysis of membrane transporters allows future development of models for transport mechanisms of lipophilic compounds. The flow of FAs from plastids to their final cellular destination in lipid molecules is controlled by membrane-intrinsic and membrane-attached proteins. Plant membrane transporters for FAs and lipid derivatives represent essential components in growth, development, and plant performance. [本文引用: 1] |
| [23] | [本文引用: 1] |
| [24] | [本文引用: 1] |
| [25] | DOI:10.1016/j.copbio.2012.08.008PMID:22981869URLMicroalgae are attracting renewed interest from both the scientific and public communities owing to their potential applications as sustainable feed stocks for the production of biofuels and high value compounds, and environmental remediation. Recent advances in molecular and biochemical analyses of microalgae point toward interesting differences in lipid metabolism between algal species and in comparison to plants. These differences range from distinct acyl groups present in algal lipids, to a possible more direct role of plastids in the assembly of TAGs with consequences for the overall subcellular organization of glycerolipid metabolism. Thus, studying lipid metabolism in microalgae points to new possible avenues of genetic engineering of lipid metabolism in this organism group, and may also inform studies of lipid metabolism in plants. [本文引用: 1] |
| [26] | [本文引用: 1] |
| [27] | [本文引用: 2] |
| [28] | DOI:10.1002/mas.20284PMID:3117083URLElectrospray and matrix assisted laser desorption ionization generate abundant molecular ion species from all known lipids that have long chain fatty acyl groups esterified or amidated to many different polar headgroup features. Molecular ion species include both positive ions from proton addition [M+H]+ and negative ions from proton abstraction [M61H]61 as well as positive ions from alkali metal attachment and negative ions from acetate or chloride attachment. Collisional activation of both MALDI and ESI behave very similarly in that generated molecular species yield product ions that reveal many structural features of the fatty acyl lipids that can be detected in tandem mass spectrometric experiments. For many lipid species, collision induced dissociation of the positive [M+H]+ reveals information about the polar headgroup, while collision induced dissociation of the negative [M61H]61 provides information about the fatty acyl chain. The mechanisms of formation of many of these lipid product ions have been studied in detail and many established pathways are reviewed here. Specific examples of mass spectrometric behavior of several molecular species are presented, including fatty acids, triacylglycerol, phosphatidic acid, phosphatidylcholine, phosphatidylethanolamine, phosphatidylinositol, phosphatidylserine, phosphatidylglycerol, ceramide, and sphingomeylin. 08 2010 Wiley Periodicals, Inc., Mass Spec Rev 30:579–599, 2011 [本文引用: 1] |
| [29] | DOI:10.1007/s11306-015-0889-1PMID:26612984URLThe early mechanisms regulating progression towards beta cell failure in type 1 diabetes (T1D) are poorly understood, but it is generally acknowledged that genetic and environmental components are involved. The metabolomic phenotype is sensitive to minor variations in both, and accordingly reflects changes that may lead to the development of T1D. We used two different extraction methods in combination with both liquid- and gas chromatographic techniques coupled to mass spectrometry to profile the metabolites in a transgenic non-diabetes prone C57BL/6 mouse expressing CD154 under the control of the rat insulin promoter (RIP) crossed into the immuno-deficient recombination-activating gene (RAG) knockout (-/-) C57BL/6 mouse, resembling the early stages of human T1D. We hypothesized that alterations in the metabolomic phenotype would characterize the early pathogenesis of T1D, thus metabolomic profiling could provide new insight to the development of T1D. Comparison of the metabolome of the RIP CD154 脳 RAG-/- mice to RAG-/- mice and C57BL/6 mice revealed alterations of >100 different lipids and metabolites in serum. Low lysophosphatidylcholine levels, accumulation of ceramides as well as methionine deficits were detected in the pre-type 1 diabetic mice. Additionally higher lysophosphatidylinositol levels and low phosphatidylglycerol levels where novel findings in the pre-type 1 diabetic mice. These observations suggest that metabolomic disturbances precede the onset of T1D. [本文引用: 1] |
| [30] | DOI:10.1371/journal.pone.0164673URLOleaginous microalgae are considered as a promising resource for the production of biofuels. Especially diatoms arouse interest as biofuel producers since they are most productive in carbon fixation and very flexible to environmental changes in the nature. Naturally, triacylglycerol (TAG) accumulation in algae only occurs under stress conditions like nitrogen-limitation. We focused on Phaeodactylum strain Pt4 (UTEX 646), because of its ability to grow in medium with low salinity and therefore being suited when saline water is less available or for wastewater cultivation strategies. Our data show an increase in neutral lipids during nitrogen-depletion and predominantly 16:0 and 16:1(n-7) accumulated in the TAG fraction. The molecular species composition of TAG suggests a remodeling primarily from the betaine lipid diacylglyceroltrimethylhomoserine (DGTS), but a contribution of the chloroplast galactolipid monogalactosyldiacylglycerol (MGDG) cannot be excluded. Interestingly, the acyl-CoA pool is rich in 20:5(n-3) and 22:6(n-3) in all analyzed conditions, but these fatty acids are almost excluded from TAG. Other metabolites most obviously depleted under nitrogen-starvation were amino acids, lyso-phospholipids and tricarboxylic acid (TCA) cycle intermediates, whereas sulfur-containing metabolites as dimethylsulfoniopropionate, dimethylsulfoniobutyrate and methylsulfate as well as short acyl chain carnitines, propanoyl-carnitine and butanoyl-carnitine increased upon nitrogen-starvation. Moreover, the Calvin cycle may be de-regulated since sedoheptulose accumulated after nitrogen-depletion. Together the data provide now the basis for new strategies to improve lipid production and storage in Phaeodactylum strain Pt4. [本文引用: 1] |
| [31] | DOI:10.1016/j.ijms.2010.11.008URLMatrix-assisted laser desorption/ionization time-of-flight mass spectrometry is a highly appreciated method in oligosaccharide analysis due to its high sensitivity and ease of use. As underivatized oligosaccharides suffer from low ionization efficiency, derivatization is a widespread tool. However, subsequent sample purification and toxic or otherwise dangerous reagents complicate the oligosaccharides’ analysis. On-target derivatization performed by the matrix 3-aminoquinoline does not require such purification and yields Schiff bases which can be measured in positive and negative ion modes from one single spot. In this article, this simple and convenient method is applied for structural characterization of oligosaccharides using a MALDI LTQ Orbitrap. Information received by the different fragmentation techniques collision-induced dissociation (CID), pulsed-Q dissociation (PQD) and higher energy C-trap dissociation (HCD) are compared. By combining the information received from fragmentation of [M+H] + ions (in positive ion mode) and [M+NO 3] 61 ions (in negative ion mode) of 3-AQ-derivatized species by all three fragmentation techniques, a complete structural characterization in terms of linkage, branching and anomeric configuration of glycosidic bonds of oligosaccharides could be achieved. The analysis of isomeric human milk oligosaccharides derivatized with 3-AQ yielded comprehensive information on the isomers’ structures. Finally, a simple combination of peak lists obtained by the different fragmentation techniques and automatic measurements enhance and facilitate oligosaccharide analysis. [本文引用: 1] |
| [32] | DOI:10.1146/annurev.pp.33.060182.000525URLUsing a small substrate (thymidine 5'-(p-nitrophenyl phosphate) 3'-phosphate), the kinetics of staphylococcal nuclease insolubilized on CNBr-activated Sepharoses 4B and 6B are affected by internal diffusional limitations. Since we demonstrate that we are working under conditions in which external mass-transfer resistances do not influence the reaction rate, we propose a simple theoretical model that considers only the case of mixed enzymic reaction-internal diffusion kinetics. In the Eadie-Hofstee plots we find very good agreement between theory and experiment. The model accounts very well for the results obtained by changing support texture, reaction conditions, and/or enzyme concentration in the insoluble derivatives, variables that modify the diffusional restrictions of the system. [本文引用: 1] |
| [33] | |
| [34] | DOI:10.1021/ac102505fPMID:21634439URLAbstract Higher energy collision dissociation (HCD) is a complementary fragmentation tool that has recently become available on mass spectrometers of the LTQ Orbitrap family. We report on a shotgun bottom-up lipidomics approach that relies on HCD of the isolated lipid precursors. HCD, together with the high mass resolution and mass accuracy of the Orbitrap analyzer, improved the confidence of molecular species assignment and accuracy of their quantification in total lipid extracts. These capabilities were particularly important for accounting for biologically interesting lipid species comprising polyunsaturated and odd numbered fatty acid moieties. We argue that now both bottom-up and top-down shotgun lipidomics could be performed on the same instrumentation platform. [本文引用: 1] |
| [35] | DOI:10.1016/j.cmet.2016.04.001PMID:27133131URLSlatter et al. characterize the lipidomic network of human platelets. Besides the characterization of nearly 200 oxidized species, the resource shows that remodeling of the membrane via phospholipase activity provides energy substrates for respiration. The findings demonstrate a direct link between innate immunity and mitochondrial bioenergetics in human platelets. [本文引用: 1] |
| [36] | DOI:10.1021/ac202646vPMID:22111752URLAn LC-MS based method for the profiling and characterization of ceramide species in the upper layer of human skin is described. Ceramide samples, collected by tape stripping of human skin, were analyzed by reversed-phase liquid chromatography coupled to high-resolution quadrupole time-of-flight mass spectrometry operated in both positive and negative electrospray ionization mode. All known classes of ceramides could be measured in a repeatable manner. Furthermore, the data set showed several undiscovered ceramides, including a class with four hydroxyl functionalities in its sphingoid base. High-resolution MS/MS fragmentation spectra revealed that each identified ceramide species is composed of several skeletal isomers due to variation in carbon length of the respective sphingoid bases and fatty acyl building blocks. The resulting variety in skeletal isomers has not been previously demonstrated. It is estimated that over 1000 unique ceramide structures could be elucidated in human stratum corneum. Ceramide... [本文引用: 1] |
| [37] | DOI:10.1016/j.chroma.2010.04.034PMID:20452604URLIn the present research, we have established a new lipidomics approach for the comprehensive and precise identification of molecular species in a crude lipid mixture using a LTQ Orbitrap mass spectrometer (MS) and reverse-phase liquid chromatography (RPLC) combination with our newly developed lipid search engine “Lipid Search”. LTQ Orbitrap provides high mass accuracy MS spectra by Fourier-transform (FT) mass spectrometer mode and can perform rapid MS n by ion trap (IT) mass spectrometer mode. In this study, the negative ion mode was selected to detect fragment ions from phospholipids, such as fatty acid anions, by MS2 or MS3. We selected the specific detection approach by neutral loss survey-dependent MS3, for the identification of molecular species of phosphatidylcholine, sphingomyelin and phosphatidylserine. Identification of molecular species was performed by using both the high mass accuracy of the mass spectrometric data obtained from FT mode and structural data obtained from fragments in IT mode. Some alkylacyl and alkenylacyl species have the same m/ z value as molecular-related ions and fragment ions, thus, direct acid hydrolysis analysis was performed to identify alkylacyl and alkenylacyl species, and then the RPLC–LTQ Orbitrap method was applied. As a result, 290 species from mouse liver and 248 species from mouse brain were identified within six different classes of phospholipid, only those in manually detected and confirmed. Most of all manually detected mass peaks were also automatically detected by “Lipid Search”. Adding to differences in molecular species in different classes of phospholipids, many characteristic differences in molecular species were detected in mouse liver and brain. More variable number of saturated and monounsaturated fatty acid-containing molecular species were detected in mouse brain than liver. [本文引用: 1] |
| [38] | DOI:10.1111/tpj.2015.84.issue-3URL [本文引用: 1] |
| [39] | DOI:10.1016/j.biochi.2016.06.004URL61Lipid biosynthesis, structure and regulation are highly complex.61Lipidomics allows analysis, detection and quantification of lipids.61Many cellular, organismal and ecological phenomena were elucidated using lipidomics. [本文引用: 1] |
| [40] | [本文引用: 1] |
| [41] | [本文引用: 2] |
| [42] | DOI:10.1002/jssc.201500858PMID:26517975URLAbstract Rice is one of the most important food crops in the world. Metabolite composition in rice seeds varies significantly depending on genetic variety, climatic alternation and agricultural practice. Metabolomics is a powerful tool to reveal the metabolic response of rice to various conditions. In this work, a rice seed sample-directed pseudotargeted metabolomics method was first established and validated based on ultra high performance liquid chromatography with triple quadrupole mass spectrometry in the multiple reaction monitoring mode. A total of 749 and 617 ion pairs in positive and negative modes were achieved, respectively. Among them, about 200 metabolites were identified or tentatively identified. The developed method showed better linearity and repeatability than those of non-targeted metabolomics method. Good intra-day and inter-day precisions, recoveries and wide linear range were also obtained. Furthermore, the method was applied for the investigation of metabolic variation of rice seeds with two wild cultivars and their transgenic lines that were grown in two locations. Principal component analysis indicated that the effects of cultivar and location on metabolic variations were far more than those of gene modification. The nonparametric Mann hitney U test revealed that most metabolites were influenced by cultivar, location and gene modifications together. [本文引用: 1] |
| [43] | [本文引用: 1] |
| [44] | DOI:10.1021/ac4001223URL [本文引用: 1] |
脂质组学研究方法及其应用
1
2010
... 脂质组学研究的核心是分析技术平台, LC-ESI- MS具有快速及高灵敏度等特点, 已广泛应用于检测识别低含量并具有重要生物功能的各类脂质分子(
1
2015
... 基于液相色谱及质谱的脂质组学是对脂质进行定性定量分析并在分子水平上研究脂质代谢的一种强有力的工具(
1
2017
... 脂质不仅是微藻细胞中各种生物膜的骨架成分, 还是一种能量储存形式, 参与许多重要的生物学过程, 如叶绿体的光合作用、线粒体的电子传递及信号转导(
2
2015
... 利用UPLC-Q-Trap/MS系统(Waters ACQUITY UP- LC, AB SCIEX Q-Trap 5500)在正离子模式下对莱茵衣藻极性甘油酯种类及分子进行辨认.其中糖脂(单半乳糖甘油二酯(MGDG)、双半乳糖甘油二酯(DGDG)及硫代异鼠李糖甘油二酯(SQDG))及磷脂(磷脂酰乙醇胺(PE)、磷脂酰甘油(PG)及磷脂酰肌醇(PI))以中性丢失扫描(neutral loss scan, NL)模式进行扫描, 甜菜碱脂(N,N,N-三甲基高丝氨酸甘油二酯(DGTS))以母离子扫描(precursor ion scan, PIS)模式进行扫描(
... ), 每种极性甘油酯的特征碎片信息(
1
2013
... Orbitrap/MS (
1
1959
...
1
2017
... 此外, DGDG、SQDG及PI这3种甘油酯均含有相同的DAG碳骨架分子18:1/16:0、18:2/16:0及18:3/ 16:0, 表明其作为sn-1位C18脂肪酸去饱和的载体; 而DGTS中16:0/18:1、16:0/18:2、16:0/18:3及16:0/ 18:4分子的存在表明其作为sn-2位C18脂肪酸去饱和的载体, 这与Schlapfer和Eichenberger (1983)的研究结果相一致.虽然这4种甘油酯均为C18脂肪酸去饱和的载体, 在莱茵衣藻的生长代谢过程中具有重要的生理功能, 但是DGTS与DGDG、SQDG和PI的具体作用机制可能因脂肪酸sn位的差异而不同.研究表明, 在微拟球藻及高等植物中, PC是C18脂肪酸去饱和的载体, 与其它极性脂的合成密切相关(
1
2011
... 脂质组学研究的核心是分析技术平台, LC-ESI- MS具有快速及高灵敏度等特点, 已广泛应用于检测识别低含量并具有重要生物功能的各类脂质分子(
1
2015
... Orbitrap/MS (
2
2013
... 本研究利用UPLC-Q-Trap/MS进行一级质谱分子离子鉴定及基于MRM模式的甘油酯分子靶向定量分析.基于QQQ的脂质分析包括母离子扫描、子离子扫描及中性丢失扫描, 是“自下而上”(Bottom-up)的脂质组学分析典型方法.利用微藻各类甘油酯特定的中性丢失或母离子扫描模式不仅可以准确辨认出具有不同质荷比的各类甘油酯分子(
... ), 同时数据处理及定量分析过程简单(
1
2015
... Orbitrap/MS (
1
2010
... Orbitrap/MS (
2
1988
... 基于以上鉴定出的极性甘油酯分子, 利用UPLC系统经ESI与LTQ-Orbitrap XL质谱(Thermo Fisher Scientific, CA, USA)相连, 分别在正、负离子模式下通过二级碎片信息扫描鉴定具有相同质荷比但不同酰基链组成的分子种类.在正离子模式下, MGDG、DGDG及SQDG以[M+NH4]+形式检测, DGTS以[M+H]+形式检测; 在负离子模式下, PE、PG及PI以[M-H]-形式检测.质量扫描范围设置为(140-1 000) m/z.在样品检测之前, 需提前计算或确认每个极性甘油酯分子的精确质量(精确至小数点后四位)以确保高分辨率.该精确质量的查询可在LIPIDMAPS (http://www.lipidmaps.org/)数据库中进行, 同时需确定每个甘油酯分子的保留时间以设置二级碎片扫描信息.一级质谱扫描采用傅里叶变换(FTMS)模式, 二级质谱扫描采用离子阱(ion trap, IT)模式.碰撞诱导解离(collision-induced dissociation, CID)碎裂由氦气碰撞产生.MGDG及SQDG的碰撞能设置为50 eV, DGDG为70 V, PE、PG、PI及DGTS为35 eV.活化Q值(Activation Q)设为0.18.隔离宽度(isolation width)分别设置为1.0 m/z (PE、PG、PI及DGTS)及3.0 m/z (MGDG、DGDG及SQDG).利用Xcalibur 2.1 (Ther- mo Fisher Scientific, CA, USA)软件进行数据采集.各极性甘油酯分子的脂酰基链位置分布(sn-1及sn-2位)按已发表文献中的检测结果确定(
... 根据以上定性定量分析结果可知, 莱茵衣藻的极性脂以糖脂MGDG、DGDG及甜菜碱脂DGTS为主, 还包括光合膜脂SQDG、PG及磷脂PE、PI.这些极性脂的组成比例与Li-Beisson等(2015)的研究结果一致, 同时各类极性脂的分子种类及酰基组成各有特点, 这也表明其合成途径及生物学功能各不相同.在高等植物中, 甘油酯通过真核或原核途径合成(
1
2017
... 基于液相色谱及质谱的脂质组学是对脂质进行定性定量分析并在分子水平上研究脂质代谢的一种强有力的工具(
2016a
2
2016b
... 根据上述鉴定出的65种具有不同质荷比的分子种类组成, 先利用UPLC-Orbitrap/MS在FTMS模式及正负离子模式下获得莱茵衣藻所有甘油酯分子的高分辨质谱全扫描图, 对Q-Trap/MS获得的质荷比不同的所有甘油酯分子进行再次确认及鉴定; 再利用UPLC- Orbitrap/MS2在IT模式下对上述各极性甘油酯分子进行二级碎片信息扫描.通过对每类极性甘油酯分子二级质谱(MS/MS)信息进行解析, 获得各类极性甘油酯的碎裂方式以及碰撞诱导解离(CID)碎片(
... 脂质组学研究的核心是分析技术平台, LC-ESI- MS具有快速及高灵敏度等特点, 已广泛应用于检测识别低含量并具有重要生物功能的各类脂质分子(
1
2003
... 基于液相色谱及质谱的脂质组学是对脂质进行定性定量分析并在分子水平上研究脂质代谢的一种强有力的工具(
1
2016
... 脂质不仅是微藻细胞中各种生物膜的骨架成分, 还是一种能量储存形式, 参与许多重要的生物学过程, 如叶绿体的光合作用、线粒体的电子传递及信号转导(
1
2005
... Orbitrap/MS (
3
2016
... 基于液相色谱及质谱的脂质组学是对脂质进行定性定量分析并在分子水平上研究脂质代谢的一种强有力的工具(
... 基于以上鉴定出的极性甘油酯分子, 利用UPLC系统经ESI与LTQ-Orbitrap XL质谱(Thermo Fisher Scientific, CA, USA)相连, 分别在正、负离子模式下通过二级碎片信息扫描鉴定具有相同质荷比但不同酰基链组成的分子种类.在正离子模式下, MGDG、DGDG及SQDG以[M+NH4]+形式检测, DGTS以[M+H]+形式检测; 在负离子模式下, PE、PG及PI以[M-H]-形式检测.质量扫描范围设置为(140-1 000) m/z.在样品检测之前, 需提前计算或确认每个极性甘油酯分子的精确质量(精确至小数点后四位)以确保高分辨率.该精确质量的查询可在LIPIDMAPS (http://www.lipidmaps.org/)数据库中进行, 同时需确定每个甘油酯分子的保留时间以设置二级碎片扫描信息.一级质谱扫描采用傅里叶变换(FTMS)模式, 二级质谱扫描采用离子阱(ion trap, IT)模式.碰撞诱导解离(collision-induced dissociation, CID)碎裂由氦气碰撞产生.MGDG及SQDG的碰撞能设置为50 eV, DGDG为70 V, PE、PG、PI及DGTS为35 eV.活化Q值(Activation Q)设为0.18.隔离宽度(isolation width)分别设置为1.0 m/z (PE、PG、PI及DGTS)及3.0 m/z (MGDG、DGDG及SQDG).利用Xcalibur 2.1 (Ther- mo Fisher Scientific, CA, USA)软件进行数据采集.各极性甘油酯分子的脂酰基链位置分布(sn-1及sn-2位)按已发表文献中的检测结果确定(
... 综上所述, 在脂质组的定性定量分析中, Q-Trap/ MS兼具一级结构鉴定(母离子及中性丢失扫描)及靶向定量分析(MRM模式)的优势, 而Orbitrap/MS2的高质量分辨率性能使其更适于进行脂质分子二级结构的准确鉴定.本研究中莱茵衣藻脂质经UPLC的反相色谱分离、ESI离子化、Q-Trap/MS及Orbitrap/MS2检测, 确定了109种极性甘油酯分子种类及结构谱图.基于以上极性甘油酯组轮廓, 又通过UPLC-Q- Trap/MS的MRM模式定量分析出莱茵衣藻的45种极性甘油酯分子含量.虽然Q-Trap/MS并未全部扫描并定量检出基于Orbitrap/MS2定性得到的所有极性脂分子, 但研究所获得的定性定量信息可以完善目前现有的莱茵衣藻极性甘油酯组(
1
2014
... 基于液相色谱及质谱的脂质组学是对脂质进行定性定量分析并在分子水平上研究脂质代谢的一种强有力的工具(
1
2016
... 脂质不仅是微藻细胞中各种生物膜的骨架成分, 还是一种能量储存形式, 参与许多重要的生物学过程, 如叶绿体的光合作用、线粒体的电子传递及信号转导(
1
2015
... 脂质不仅是微藻细胞中各种生物膜的骨架成分, 还是一种能量储存形式, 参与许多重要的生物学过程, 如叶绿体的光合作用、线粒体的电子传递及信号转导(
1
2016
... 脂质不仅是微藻细胞中各种生物膜的骨架成分, 还是一种能量储存形式, 参与许多重要的生物学过程, 如叶绿体的光合作用、线粒体的电子传递及信号转导(
1
2013
... 此外, DGDG、SQDG及PI这3种甘油酯均含有相同的DAG碳骨架分子18:1/16:0、18:2/16:0及18:3/ 16:0, 表明其作为sn-1位C18脂肪酸去饱和的载体; 而DGTS中16:0/18:1、16:0/18:2、16:0/18:3及16:0/ 18:4分子的存在表明其作为sn-2位C18脂肪酸去饱和的载体, 这与Schlapfer和Eichenberger (1983)的研究结果相一致.虽然这4种甘油酯均为C18脂肪酸去饱和的载体, 在莱茵衣藻的生长代谢过程中具有重要的生理功能, 但是DGTS与DGDG、SQDG和PI的具体作用机制可能因脂肪酸sn位的差异而不同.研究表明, 在微拟球藻及高等植物中, PC是C18脂肪酸去饱和的载体, 与其它极性脂的合成密切相关(
1
2014
... 基于液相色谱及质谱的脂质组学是对脂质进行定性定量分析并在分子水平上研究脂质代谢的一种强有力的工具(
2
2017
... 本研究利用UPLC-Q-Trap/MS进行一级质谱分子离子鉴定及基于MRM模式的甘油酯分子靶向定量分析.基于QQQ的脂质分析包括母离子扫描、子离子扫描及中性丢失扫描, 是“自下而上”(Bottom-up)的脂质组学分析典型方法.利用微藻各类甘油酯特定的中性丢失或母离子扫描模式不仅可以准确辨认出具有不同质荷比的各类甘油酯分子(
... 此外, DGDG、SQDG及PI这3种甘油酯均含有相同的DAG碳骨架分子18:1/16:0、18:2/16:0及18:3/ 16:0, 表明其作为sn-1位C18脂肪酸去饱和的载体; 而DGTS中16:0/18:1、16:0/18:2、16:0/18:3及16:0/ 18:4分子的存在表明其作为sn-2位C18脂肪酸去饱和的载体, 这与Schlapfer和Eichenberger (1983)的研究结果相一致.虽然这4种甘油酯均为C18脂肪酸去饱和的载体, 在莱茵衣藻的生长代谢过程中具有重要的生理功能, 但是DGTS与DGDG、SQDG和PI的具体作用机制可能因脂肪酸sn位的差异而不同.研究表明, 在微拟球藻及高等植物中, PC是C18脂肪酸去饱和的载体, 与其它极性脂的合成密切相关(
1
2011
... 脂质组学研究的核心是分析技术平台, LC-ESI- MS具有快速及高灵敏度等特点, 已广泛应用于检测识别低含量并具有重要生物功能的各类脂质分子(
1
2016
... 本研究利用UPLC-Q-Trap/MS进行一级质谱分子离子鉴定及基于MRM模式的甘油酯分子靶向定量分析.基于QQQ的脂质分析包括母离子扫描、子离子扫描及中性丢失扫描, 是“自下而上”(Bottom-up)的脂质组学分析典型方法.利用微藻各类甘油酯特定的中性丢失或母离子扫描模式不仅可以准确辨认出具有不同质荷比的各类甘油酯分子(
1
2016
... 基于液相色谱及质谱的脂质组学是对脂质进行定性定量分析并在分子水平上研究脂质代谢的一种强有力的工具(
1
2011
... Orbitrap/MS (
1
1982
... 根据以上定性定量分析结果可知, 莱茵衣藻的极性脂以糖脂MGDG、DGDG及甜菜碱脂DGTS为主, 还包括光合膜脂SQDG、PG及磷脂PE、PI.这些极性脂的组成比例与Li-Beisson等(2015)的研究结果一致, 同时各类极性脂的分子种类及酰基组成各有特点, 这也表明其合成途径及生物学功能各不相同.在高等植物中, 甘油酯通过真核或原核途径合成(
1983
1
2011
... Orbitrap/MS (
1
2016
... 本研究利用UPLC-Q-Trap/MS进行一级质谱分子离子鉴定及基于MRM模式的甘油酯分子靶向定量分析.基于QQQ的脂质分析包括母离子扫描、子离子扫描及中性丢失扫描, 是“自下而上”(Bottom-up)的脂质组学分析典型方法.利用微藻各类甘油酯特定的中性丢失或母离子扫描模式不仅可以准确辨认出具有不同质荷比的各类甘油酯分子(
1
2012
... 脂质组学研究的核心是分析技术平台, LC-ESI- MS具有快速及高灵敏度等特点, 已广泛应用于检测识别低含量并具有重要生物功能的各类脂质分子(
1
2010
... Orbitrap/MS (
1
2015
... 本研究利用UPLC-Q-Trap/MS进行一级质谱分子离子鉴定及基于MRM模式的甘油酯分子靶向定量分析.基于QQQ的脂质分析包括母离子扫描、子离子扫描及中性丢失扫描, 是“自下而上”(Bottom-up)的脂质组学分析典型方法.利用微藻各类甘油酯特定的中性丢失或母离子扫描模式不仅可以准确辨认出具有不同质荷比的各类甘油酯分子(
1
2016
... 脂质组是微藻科学领域中一种新兴的重要工具(
1
2015
... 根据以上定性定量分析结果可知, 莱茵衣藻的极性脂以糖脂MGDG、DGDG及甜菜碱脂DGTS为主, 还包括光合膜脂SQDG、PG及磷脂PE、PI.这些极性脂的组成比例与Li-Beisson等(2015)的研究结果一致, 同时各类极性脂的分子种类及酰基组成各有特点, 这也表明其合成途径及生物学功能各不相同.在高等植物中, 甘油酯通过真核或原核途径合成(
2
2015
... 基于液相色谱及质谱的脂质组学是对脂质进行定性定量分析并在分子水平上研究脂质代谢的一种强有力的工具(
... 综上所述, 在脂质组的定性定量分析中, Q-Trap/ MS兼具一级结构鉴定(母离子及中性丢失扫描)及靶向定量分析(MRM模式)的优势, 而Orbitrap/MS2的高质量分辨率性能使其更适于进行脂质分子二级结构的准确鉴定.本研究中莱茵衣藻脂质经UPLC的反相色谱分离、ESI离子化、Q-Trap/MS及Orbitrap/MS2检测, 确定了109种极性甘油酯分子种类及结构谱图.基于以上极性甘油酯组轮廓, 又通过UPLC-Q- Trap/MS的MRM模式定量分析出莱茵衣藻的45种极性甘油酯分子含量.虽然Q-Trap/MS并未全部扫描并定量检出基于Orbitrap/MS2定性得到的所有极性脂分子, 但研究所获得的定性定量信息可以完善目前现有的莱茵衣藻极性甘油酯组(
1
2016
... 本研究利用UPLC-Q-Trap/MS进行一级质谱分子离子鉴定及基于MRM模式的甘油酯分子靶向定量分析.基于QQQ的脂质分析包括母离子扫描、子离子扫描及中性丢失扫描, 是“自下而上”(Bottom-up)的脂质组学分析典型方法.利用微藻各类甘油酯特定的中性丢失或母离子扫描模式不仅可以准确辨认出具有不同质荷比的各类甘油酯分子(
1
2016
... 脂质不仅是微藻细胞中各种生物膜的骨架成分, 还是一种能量储存形式, 参与许多重要的生物学过程, 如叶绿体的光合作用、线粒体的电子传递及信号转导(
1
2013
... Orbitrap/MS (

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}