包宇 1 , 2 ,
乔娅楠 3 ,
闫兴富 4 ,
王灵艳 1 , 2 ,
曲文静 1 , 2 ,
唐占辉 1 , 2 , * , 1 国家环境保护湿地生态与植被恢复重点实验室, 长春 1301172 东北师范大学环境学院, 长春 1301173 武警后勤学院学员二旅, 天津 3003094 北方民族大学生物科学与工程学院, 银川 750021Bao Yu 1 , 2 ,
Qiao Yanan 3 ,
Yan Xingfu 4 ,
Wang Lingyan 1 , 2 ,
Qu Wenjing 1 , 2 ,
Tang Zhanhui 1 , 2 , * , 1 State Environmental Protection Key Laboratory of Wetland Ecology and Vegetation Restoration, Changchun 130117, China2 School of Environment Northeast Normal University, Changchun 130117, China3 No.2 Student Brigade, Logistics University of People’s Armed Police Force, Tianjin 300309, China4 College of Biological Science and Engineering, BeiFang University of Nationalities, Yinchuan 750021, China引用本文 包宇 ,
乔娅楠 ,
闫兴富 ,
王灵艳 ,
曲文静 ,
唐占辉 . 浅裂剪秋萝和丝瓣剪秋萝传粉特性的初步研究. 植物学报, 2018, 53(5): 634-642
贡献者
* 通讯作者。E-mail: tangzh789@nenu.edu.cn 接受日期:2017-04-15接受日期:2017-12-27网络出版日期:2018-09-10
-->Copyright
2018《植物学报》编辑部
Contributors
* Author for correspondence. E-mail: tangzh789@nenu.edu.cn History
Received :
Accepted :
Online :
摘要: 为了探究浅裂剪秋萝(Lychnis cognata )和丝瓣剪秋萝(L. wilfordii )的传粉生物学特征, 从花部特征、花粉呈现规律及传粉者访花行为等方面对其开展研究。结果表明, 2种剪秋萝的花期持续时间相近, 但浅裂剪秋萝始花日期较早; 二者花部特征虽有显著差异, 但主要传粉者均为碧翠凤蝶(Papilio bianor )。在开花过程中, 2种剪秋萝的雄蕊均分2批呈现花粉, 第1批雄蕊的花粉生活力在开花后第1天达到最大值, 而第2批雄蕊在第2天达到最大值。浅裂剪秋萝的柱头可授性在开花后第5天最强, 而丝瓣剪秋萝在开花后第4天最强, 花粉生活力和柱头活性的时间差异表明两物种均为雌雄异熟。两物种共同的传粉者碧翠凤蝶对浅裂剪秋萝的访花高峰出现在上午8:00-11:00, 而对丝瓣剪秋萝的访花高峰出现在上午11:00-12:00, 导致访花高峰出现差异的主要影响因素可能是生境和花药开裂时间。 关键词 :
传粉生物学 ;
浅裂剪秋萝 ;
丝瓣剪秋萝 ;
花部特征 Abstract :
The aim of this study was to study the pollination biology of Lychnis cognata and L. wilfordii from the aspects floral syndrome, pollen occurrence regularity and pollinator flower-visiting behavior. The floral syndrome of L. cognata and L. wilfordii differed significantly, but the main pollinator of these two species was the same: Papilio bianor . The stamens of the two species shed their pollen in two batches, and the pollen viability of the two batches peaked at different times. The pollen viability of the first batch of the two species peaked on the first day after flowering but that of the second batch on the second day after flowering. The receptivity of the stigma of L. cognata reached the peak on the fifth day after flowering, 1 day later than that of L. wilfordii . Hence, the two species are dichogamous. The flowering period was earlier for L. cognata than L. wilfordii , but the flowering duration of two species was similar. The highest visiting frequency of pollinators to L. cognata was at 8:00 to 11:00 am and to L. wilfordii was at 11:00 to 12:00 am. The important factors influencing the visiting time of pollinators may be habitat environmental factors or time of anther dehiscence. Key words :
pollination biology ;
Lychnis cognata ;
L. wilfordii ;
floral syndrome 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 )。近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究。此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物。植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程。另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究。基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索。
浅裂剪秋萝和丝瓣剪秋萝是分布在吉林省龙湾自然保护区的石竹科(Caryophyllaceae)剪秋萝属(
Lychnis )多年生草本植物(
傅沛云, 1995 )。前期的野外观察发现, 2种剪秋萝在传粉过程中存在雄蕊分批生长传播花粉现象, 雄蕊在花部的时空配置可能在花部特征进化过程中扮演着重要角色, 且2种剪秋萝野外种群数量近年来呈下降趋势, 故对其保护和研究势在必行。目前, 关于该属植物的研究主要集中在引种栽培管理、园林应用、组织培养及多倍体育种等方面(
刘明财等, 2004 ;
王艳菊, 2006 ;
肖宁等, 2012 ;
冯敏等, 2014 ), 而对传粉生物学方面的研究报道较少。本文试图对这2种剪秋萝的传粉生物学特征进行初步探究, 并对其开花物候、雄蕊和雌蕊呈现规律、花部特征差异与传粉者访花规律进行探索, 一方面为进一步研究剪秋萝的传粉过程提供参考资料, 另一方面为深入认识剪秋萝在引种栽培过程中提高传粉效率等方面的问题提供借鉴与思路。
1 材料与方法1.1 研究地区概况研究地点位于吉林省通化市辉南县龙湾自然保护区金川镇东1.5 km 的样地(126°15'55″-126°32'02″E, 42°16'20″-42°26'57″N)。研究区平均海拔741 m, 年平均气温4.1°C, 年平均降水量704.2 mm, 降水时间分布受气候影响显著, 夏季雨量集中, 占全年降水的61% (
盛连喜等, 2013 )。
1.2 研究材料本研究所用浅裂剪秋萝(
Lychnis cognata Maxim)和丝瓣剪秋萝(
L. wilfordii (Regel) Maxim)野外居群分别位于保护区公路沿路林缘下和河岸边湿地, 二者野外居群相距约1 km。两物种的二歧聚伞花序具数花, 花两性, 雄蕊10枚, 雌蕊5枚合生, 子房卵形; 花期7-8月。前者株高35-90 cm (
图1A ), 叶长圆状披针形或长圆形, 多生于海拔500-1 000 m的林下或灌丛; 后者株高45-100 cm (
图1B ), 叶长圆状披针形或长披针形, 花序稍紧密, 多生于海拔250-1 200 m的湿草甸、河岸低湿地(
傅沛云, 1995 )。本研究于2016年7-9月进行。
1.3 研究方法
1.3.1 开花生物学观测 详细记录浅裂剪秋萝居群(200株)和丝瓣剪秋萝居群(50株)的始花日期、盛花期、花期持续时间及终花期等开花参数(株数不同是由于野外居群数量限制)。花期的判断标准采用
Dafni (1992) 文献所述原则(居群水平上5%的植株开始开花定义为始花, 盛花期指群体中50%以上的植株同时开放所持续的时间, 终花期指5%以下的植株仍在开花。花期持续时间定义为从第1棵植株始花到最后1株花凋谢所持续的时间)。此外, 在野外记录2个物种各10朵单花的开放动态、单花寿命和花药开裂时间, 在盛花期选取2个物种的花各10朵, 于早上9:00用游标卡尺测量花冠长度、花冠宽度、花瓣裂深、花萼长度、子房长度、子房宽度、雄蕊高度和雌蕊长度等表型参数。
图1 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-634/img_1.png <b>图1</b> 浅裂剪秋萝(A)和丝瓣剪秋萝(B)的花部形态<br/><b>Figure 1</b> Floral morphology of <i>Lychnis cognata</i> (A) and<i> L. wilfordii</i> (B) Figure 1 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-634/img_1.png <b>图1</b> 浅裂剪秋萝(A)和丝瓣剪秋萝(B)的花部形态<br/><b>Figure 1</b> Floral morphology of <i>Lychnis cognata</i> (A) and<i> L. wilfordii</i> (B) 在新窗口打开 下载 下载原图ZIP 生成PPT 图1
浅裂剪秋萝(A)和丝瓣剪秋萝(B)的花部形态
Figure 1
Floral morphology of
Lychnis cognata (A) and
L. wilfordii (B)
1.3.2 花粉生活力和柱头可授性变化 采用花粉离体萌发法测定花粉生活力。在花朵即将开放前和开花后第1、2、3天早上8:00随机采集20朵花的花粉(由于2物种的花具2轮雄蕊, 分2批呈现花粉, 分别采集第1批和第2批雄蕊的花粉), 用镊子摘除花药后, 现场将花粉刮进装有液体培养基的离心管中。20朵花的第1批和第2批花粉分别放入不同的离心管, 液体培养基的成分为20%蔗糖和0.01%硼酸(
胡适宜, 1993 )。随后带回实验室在25°C条件下培养2小时以上, 然后在显微镜下检测花粉的萌发率(萌发率=萌发的花粉数/花粉总数) (花粉管长度大于花粉直径视为花粉萌发), 重复计数3次, 最后计算20株植物花的花粉萌发率的平均值。
柱头可授性采用联苯胺-过氧化氢法检测, 在花朵即将开放前和开花后24、48、72、96、120和144小时, 分别随机选取10朵花, 将柱头浸入联苯胺-过氧化氢反应液(1%联苯胺:3%过氧化氢:水=4:11:22, v/v/v)中。若柱头具有可授性, 则柱头周围呈蓝色并有大量的气泡出现; 若柱头不具有可授性, 则柱头周围不会呈蓝色也不会产生气泡。根据气泡多少和颜色的深浅判断柱头可受性的强弱(
胡适宜, 1993 )。用-和+表示观察到的情况。-表示柱头不具有可授性, 即无气泡与变蓝情况; +表示柱头具有可授性, 即轻微变蓝并有气泡产生; ++表示柱头具有较强的可授性, 即有较多的气泡和比较明显的变蓝情况; +++表示柱头具有极强的可授性, 即有大量的气泡和非常明显的变蓝情况。
1.3.3 花粉数量和花粉胚珠比值(P/O)的测定 在每个物种居群内随机选取即将开放的花各30朵, 每朵花取下全部花药。将第1批雄蕊和第2批雄蕊分别置于蒸馏水中, 定容至1 mL。摇匀后, 取20 μL悬浮液放在载玻片上, 在显微镜下记录花粉粒数, 重复计数3次。用手术刀横切子房并记录胚珠数目, 统计各花的单花花粉数和胚珠数, 最后计算花粉/胚珠(P/O)比值(
胡适宜, 1993 )。参照
Cruden (1977) 的P/O值划分繁育系统类型, P/O值在2.7-5.4区间时繁育系统为闭花授粉; 18.1-39.0时为专性自交; 31.9-396.0时为兼性自交; 244.7-2 588.0时为兼性异交; 2 108.0- 195 525.0时为专性异交。P/O值降低意味着近交程度升高, P/O值升高则表明远交程度上升。
1.3.4 异交指数(OCI)的计算 按照Dafni (1992) 所述标准计算异交指数(out-cross- ing index, OCI)。(1) 花朵或花序直径<1 mm记为0; 1-2 mm记为1; 2-6 mm记为2; > 6 mm记为3。(2) 雄蕊成熟与柱头具可授性的时间间隔: 同时或雌蕊先熟记为0; 雄蕊先熟记为1。(3) 柱头与花药处于同一高度记为0; 空间分离记为1。三者累计得OCI值。
1.3.5 传粉者访花行为观察 选择晴天, 设置1.5 m ×1.5 m的样方, 分别随机选取每个物种居群中的10朵花编号, 于8:00-17:00进行不间断观察, 跟踪观察并记录访花者种类、访花行为、访花次数、在每个花朵上的停留时间, 昆虫的访花频率用单花每小时的访问次数表示。在阴雨天观察访花昆虫的访花情况, 晴天和阴雨天各重复观察记录3天。
1.4 数据分析采用SPSS 19.0统计软件进行数据分析。在比较前, 先检验数据是否符合正态分布。对服从正态分布的数据, 使用独立样本
t -检验(independent samples
t test)检验相关变量间是否存在显著性差异; 对于不服从正态分布的数据, 首先进行数据转换, 如果转换后的数据服从正态分布, 使用独立样本
t -检验来检验相关变量间是否存在显著差异。若仍不服从正态分布, 则使用非参数检验中的2个独立样本非参数检验。上述所有显著性检验的显著度均设为0.05, 统计数据用平均值±标准误表示。使用软件Sigmaplot 12.5绘图。
2 结果与讨论2.1 开花生物学特性
2.1.1 开花物候 浅裂剪秋萝单花寿命约为6-7天, 始花日期为7月20日, 5天后进入盛花期, 8月25日进入终花期, 花期持续时间为35天。丝瓣剪秋萝单花寿命约为6-7天, 始花日期为7月27日, 4天后进入盛花期, 8月23日进入终花期, 花期持续时间为27天。浅裂剪秋萝的始花日期比丝瓣剪秋萝的始花日期早7天左右。
2.1.2 花部形态特征 浅裂剪秋萝的花冠长度、花冠宽度、花瓣裂深、花萼长度、子房长度、子房宽度、雄蕊高度和雌蕊长度等所有表型参数均与丝瓣剪秋萝的相应表型参数有显著差异(
表1 )。
表1 Table 1 表1 表1 浅裂剪秋萝与丝瓣剪秋萝的花部特征比较(平均值±标准误,
n =10)
Table 1 Floral morphology of
Lychnis cognata and
L. wilfordii (means±SE,
n =10)
Observation variable Lychnis cognata L. wilfordii P Length of corolla (mm) 40.52±1.04 32.80±0.65 P <0.05Width of corolla (mm) 6.14±0.16 3.96±0.09 P <0.05Depth of crack (mm) 8.71±0.26 9.67±0.23 P <0.01Length of sepal (mm) 26.45±0.32 16.21±0.17 P <0.05Length of ovary (mm) 7.03±0.14 5.82±0.11 P <0.05Width of ovary (mm) 3.21±0.08 2.51±0.07 P <0.05Height of stamens A (mm) 19.61±0.24 12.30±0.24 P <0.05Height of stamens B (mm) 19.76±0.28 11.56±0.22 P <0.05Length of pistil (mm) 13.74±0.40 7.68±0.25 P <0.05
Stamens A: The stamen that shed pollen in the first batch; Stamens B: The stamen that shed pollen in the second batch 雄蕊A: 第1批散粉的雄蕊; 雄蕊B: 第2批散粉的雄蕊 表1
浅裂剪秋萝与丝瓣剪秋萝的花部特征比较(平均值±标准误,
n =10)
Table 1
Floral morphology of
Lychnis cognata and
L. wilfordii (means±SE,
n =10)
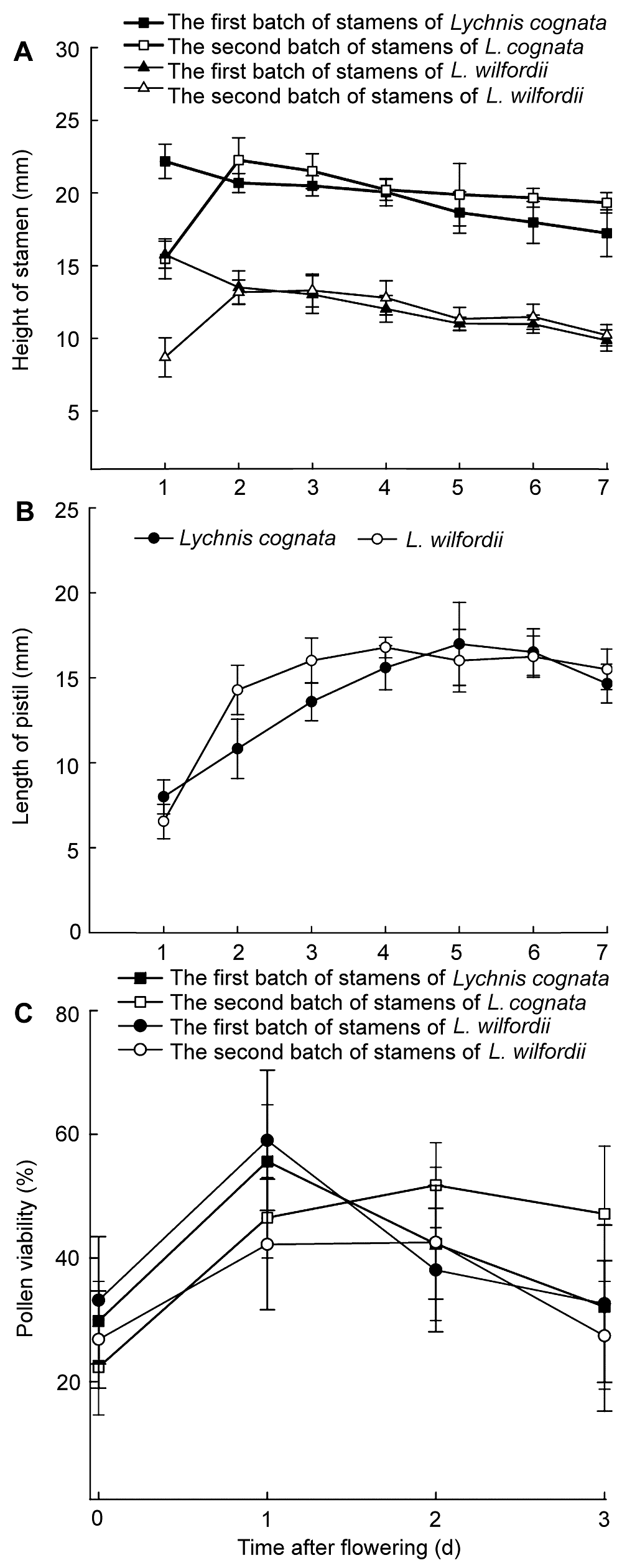
2.1.3 雄蕊和雌蕊的呈现规律 浅裂剪秋萝和丝瓣剪秋萝开花时间多集中在8:00- 12:00。开花时花瓣逐一展开直至5片花瓣分离, 花即开放。浅裂剪秋萝雌蕊长度在花蕾期小于雄蕊长度, 柱头在花药下方, 开花第0-24小时第1批5枚雄蕊继续生长并高于花筒, 花粉囊开裂进行散粉, 花粉囊开裂方式为纵裂, 花粉囊开裂时间一般为8:00-9:00, 此时第2批5枚雄蕊位于花筒下方, 花粉囊未开裂; 第24-48小时第1批散粉的5枚雄蕊平卧在花瓣之上, 第2批5枚雄蕊继续伸长并高于花筒, 花粉囊开裂进行散粉, 花粉囊开裂时间也为8:00-9:00, 两批雄蕊花药开裂时间相差24小时, 散粉期为3-4天; 第48-72小时所有雄蕊均平卧于花瓣之上, 雄蕊不再生长, 柱头伸长位于花筒上方占据最佳授粉位置, 此时柱头可授性才开始逐渐升高; 第96小时雌蕊继续生长; 第120小时花瓣开始萎蔫, 雌蕊长度基本不变(
图2A , B)。丝瓣剪秋萝的开花过程与浅裂剪秋萝类似, 但是其两批雄蕊花粉囊开裂时间为9:00-10:00, 稍晚于浅裂剪秋萝(
图2A , B)。
图2 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-634/img_2.png <b>图2</b> 2种剪秋萝的两批雄蕊高度(A)、雌蕊长度(B)和花粉生活力(C)随时间的变化<br/><b>Figure 2</b> Changes of the height of two batches of stamens (A), the pistil length (B), and pollen viability (C) of <i>Lychnis cognata</i> and<i> L. wilfordii</i> during anthesis Figure 2 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-634/img_2.png <b>图2</b> 2种剪秋萝的两批雄蕊高度(A)、雌蕊长度(B)和花粉生活力(C)随时间的变化<br/><b>Figure 2</b> Changes of the height of two batches of stamens (A), the pistil length (B), and pollen viability (C) of <i>Lychnis cognata</i> and<i> L. wilfordii</i> during anthesis 在新窗口打开 下载 下载原图ZIP 生成PPT 图2
2种剪秋萝的两批雄蕊高度(A)、雌蕊长度(B)和花粉生活力(C)随时间的变化
Figure 2
Changes of the height of two batches of stamens (A), the pistil length (B), and pollen viability (C) of
Lychnis cognata and
L. wilfordii during anthesis
2.2 花粉生活力与柱头可授性变化浅裂剪秋萝和丝瓣剪秋萝的花粉在开花前均有生活力。浅裂剪秋萝第1批散粉雄蕊的花粉生活力在开花第1天达到峰值, 为55.63%, 随后急剧下降; 第2批散粉雄蕊的花粉生活力在第2天达到峰值, 为49.88%, 随后缓慢下降, 第4天已很难从花药中收集到花粉。丝瓣剪秋萝第1批散粉雄蕊的花粉生活力在开花第1天达到峰值, 为54.89%, 随后急剧下降; 第2批散粉雄蕊的花粉生活力在第2天达到峰值, 为42.54%, 第3天开始下降, 从第4天开始, 基本不能从花药中收集到花粉。在开花第1天, 浅裂剪秋萝和丝瓣剪秋萝的第1批散粉雄蕊的花粉生活力均大于第2批散粉雄蕊的花粉生活力; 在第2天, 第2批散粉雄蕊的花粉生活力均大于第1批散粉雄蕊的花粉生活力, 雄蕊的这种分批呈现拉开了花粉输出的时间跨度(
图2C )。
浅裂剪秋萝和丝瓣剪秋萝在开花前柱头均无可授性, 浅裂剪秋萝在开花第2天部分柱头才具有可授性; 开花后第3天柱头可授性逐渐增强, 滴入联苯胺后, 柱头迅速变蓝并产生大量的气泡; 开花后第5天可授性最强, 随后可授性逐渐减弱; 开花后第7天, 仅部分柱头具有可授性。丝瓣剪秋萝在开花第3天柱头才具可授性, 随即可授性逐渐增强, 开花后第4天柱头可授性最强, 然后逐渐减弱, 开花后第7天, 柱头便完全检测不到可授性(
表2 )。
表2 Table 2 表2 表2 浅裂剪秋萝和丝瓣剪秋萝柱头可授性随时间的变化
Table 2 Changes in stigma receptivity of
Lychnis cognata and
L. wilfordii during anthesis
Flowering Stigma receptivity ofLychnis cognata Stigma receptivity L. wilfordii 0 - - 24 - - 48 + - 72 ++ ++ 96 ++ +++ 120 +++ ++ 144 ++ ++ 168 + -
-: Stigma have no receptivity; +: Stigma have receptivity. The sample size n =10. -: 柱头不具可授性;+: 柱头具可授性。柱头可授性检测的样本量n =10。 表2
浅裂剪秋萝和丝瓣剪秋萝柱头可授性随时间的变化
Table 2
Changes in stigma receptivity of
Lychnis cognata and
L. wilfordii during anthesis
2.3 单花花粉量、花粉胚珠比值及异交指数浅裂剪秋萝和丝瓣剪秋萝单花全部雄蕊的花粉量分别为143 910±2 439.77和77 021±2 025.37个。方差分析表明, 两物种单花花粉量有显著差异(
F =1.89,
P < 0.05), 胚珠分别为103±2.60和74±2.34个; P/O值分别为1 426.65±44.19和1 083.96±58.86。方差分析表明, 这2个物种单花P/O值存在显著差异(
F =0.42,
P <0.05)。2种剪秋萝的异交指数OCI为5, 均大于或等于4。根据P/O值和异交指数OCI, 判断2个物种的繁育系统均属于兼性异交类型(
表3 )。
表3 Table 3 表3 表3 浅裂剪秋萝和丝瓣剪秋萝的异交指数
Table 3 The out-crossing index of
Lychnis cognata and
L. wilfordii Species Diameter of Temporal separation of anther Spatial positioning of OCI Type of breeding L. cognata 3 1 1 5 Xenogamy L. wilfordii 3 1 1 5 Xenogamy
OCI: Out-crossing index OCI: 异交指数 表3
浅裂剪秋萝和丝瓣剪秋萝的异交指数
Table 3
The out-crossing index of
Lychnis cognata and
L. wilfordii 2种剪秋萝除单花花粉量具显著差异外, 浅裂剪秋萝第1批雄蕊的花粉量为86 266±2 070.47, 显著大于第2批雄蕊的花粉量57 644±1 928.45 (
F =0.46,
P <0.05); 丝瓣剪秋萝第1批雄蕊的花粉量为42 533± 1 275.84, 也显著大于第2批雄蕊的花粉量35 555± 1 275.84 (
F =0.43,
P <0.05)。
2.4 昆虫访花行为、频率和时间浅裂剪秋萝和丝瓣剪秋萝的主要传粉者均为碧翠凤蝶(
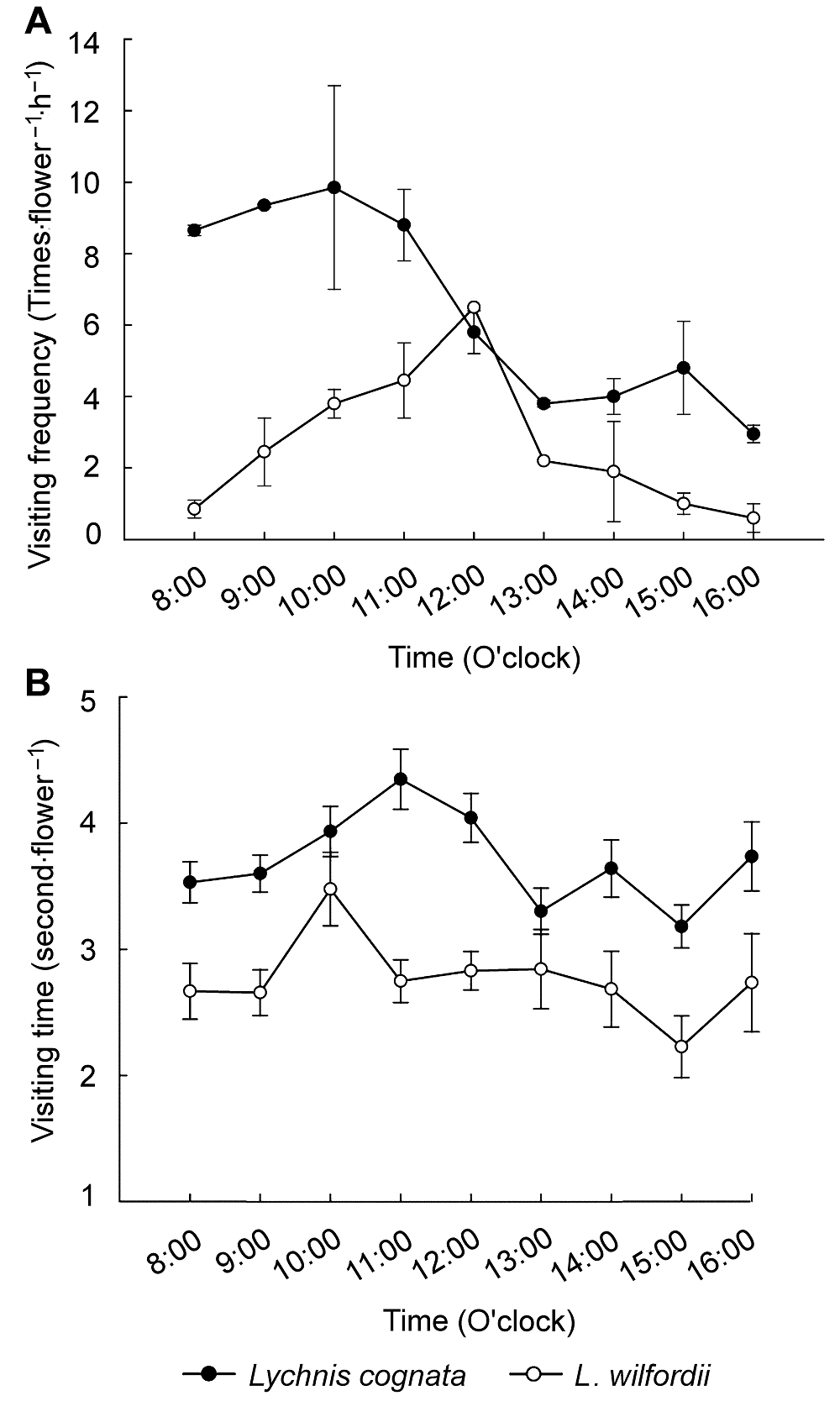
Papilio bianor )。此外, 还有一些蜂类访花, 但出现的频率极低。在晴朗的天气条件下, 碧翠凤蝶从8:00-17:00均访花, 但上午访花较频繁。浅裂剪秋萝日访花频率高峰出现在8:00-11:00, 在14:00-15:00又有1个小高峰; 随着花粉量和气温的下降, 16:00以后, 传粉者访花频率逐渐降低, 17:00以后几乎无传粉者访花。丝瓣剪秋萝日访花高峰出现在11:00-12:00, 14:00之后传粉者访花频率逐渐降低(
图5 )。在阴雨等不良天气状况下, 两物种几乎无传粉者。浅裂剪秋萝的单花访问时间高峰为11:00, 约为(3.16±0.24) S/次; 而丝瓣剪秋萝的单花访问时间高峰为10:00, 约为(2.55±0.29) S/次; 2个物种单花访问时间最低点均为15:00 (
图3A , B)。
图3 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-634/img_3.png <b>图3</b> 浅裂剪秋萝和丝瓣剪秋萝的传粉者访花频率(A)和访花时间(B)<br/><b>Figure 3</b> Visiting frequency (A) and visiting time (B) of pollinators to flowers of <i>Lychnis cognata</i> and <i>L. wilfordii</i> Figure 3 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-634/img_3.png <b>图3</b> 浅裂剪秋萝和丝瓣剪秋萝的传粉者访花频率(A)和访花时间(B)<br/><b>Figure 3</b> Visiting frequency (A) and visiting time (B) of pollinators to flowers of <i>Lychnis cognata</i> and <i>L. wilfordii</i> 在新窗口打开 下载 下载原图ZIP 生成PPT 图3
浅裂剪秋萝和丝瓣剪秋萝的传粉者访花频率(A)和访花时间(B)
Figure 3
Visiting frequency (A) and visiting time (B) of pollinators to flowers of
Lychnis cognata and
L. wilfordii 2.5 讨论
2.5.1 花部特征差异 花部特征的演化受到包括传粉者在内的多种因素影响(
黄双全, 2007 ,
2014 )。2种剪秋萝的主要传粉者相同, 但传粉综合征的差异也不显著: 花瓣均为红色且基部形成花冠管, 雄性先熟, 分批呈现雄蕊, 雄蕊散粉后平卧于花瓣上。因此, 传粉者应不是导致2种剪秋萝花部形态特征产生差异的主要因素。2种剪秋萝花部形态差异很大, 丝瓣剪秋萝的花瓣边缘撕裂程度显著大于浅裂剪秋萝, 且花部的各项表型参数值均小于浅裂剪秋萝, 造成这种花部特征差异的重要影响因素之一应是生境。由于丝瓣剪秋萝的生活环境湿度较大, 呈丝状的较小花瓣可以减少水分的附着, 同时较小的花筒直径可以减少外界水分对花粉的影响, 进而减少对传粉过程的不利影响。例如, 茄科山莨菪属铃铛子(
Anisodus luridus )花期花梗弯曲是为了避免高山环境中频繁的降雨和强烈的紫外辐射对花粉的影响, 表明生境对高山植物的花部特征具有可塑性(
Wang et al., 2010 )。因此, 环境因素对2种生境差异较大的剪秋萝花部形态特征的塑造具有重要作用。花部特征在适应性进化过程中对各种因素的折中和妥协远比我们之前了解的复杂得多, 这些问题均需要进一步研究(
杨春锋和郭友好, 2005 )。
2.5.2 雄蕊和雌蕊呈现规律 浅裂剪秋萝和丝瓣剪秋萝的10枚雄蕊在开花过程中均分2批成熟, 且在不同时间开裂散粉。这是花粉分发的一种机制(
Harder and Wilson, 1994 ;
陆婷和谭敦炎, 2007 )。理论认为, 花粉在不同时间成熟, 可以将花粉分发给不同的访花者, 提高花粉的输出量(
Harder and Wilson, 1994 )。例如, 沙生蝇子草(
Silene olgiana )具有典型的雄性先熟、雌雄异位和花粉逐步呈现现象, 这对避免自交、雌雄干扰及增加后代适合度、保证该物种繁殖成功具有重要意义(
刘建华, 2009 )。
从花粉呈现空间位置看, 初花期雄蕊处于花冠口上方, 与柱头存在着明显的雌雄异位。这种空间分配有利于异交授粉和异交传粉的各自进行, 降低雌雄功能的相互干扰(
Barrett, 2002 )。同时本研究中的2种剪秋萝的两批雄蕊通过空间位置的变化在不同时间占据最佳传粉位置, 降低两批雄蕊之间的相互干扰, 使两批雄蕊能各自与访花者充分地接触, 进而促进花粉输出。这可能表明在两性花内部, 不仅存在雌雄之间的性别干扰, 在雄蕊之间也可能存在着雄雄之间的性别内干扰, 这种性别内干扰可能成为塑造花部特征的影响因素之一(
任明迅, 2010 )。2种剪秋萝的这种雌雄蕊呈现规律既降低了雌雄功能的干扰, 又降低了雄雄功能的干扰, 是提高花粉输出率的一种适应机制。
从花粉呈现的时间角度看, 两批雄蕊散粉间隔24小时, 这与传粉者数量和效率密切相关(
Harder and Thomson, 1989 )。在传粉者数量稀少的情况下, 植物偏好同时呈现花粉, 尽量避免最后1次访花后仍存留大量花粉造成雄性资源浪费; 在传粉者数量较多的情况下, 植物通过花粉时序性呈现可以使更多的传粉者参与传粉过程, 让成功到达柱头上的花粉数量增加, 进而提高雄性适合度(
Harder and Thomson, 1989 ;
Thomason and Thomson, 1992 ;
李新蓉, 2006 ;
陆婷和谭敦炎, 2007 )。
Thomson等(2000) 提出了简化数学模型, 该模型认为增加单个传粉者花粉移出量对总的雄性繁殖力不利, 因为1次移出大量的花粉会降低使胚珠受精的花粉比例。所以在传粉者数量充足的条件下, 限制单个传粉者1次访花转移的花粉数量和吸引更多的传粉者参与传粉是提高繁殖力的有效途径(
Thomson et al., 2000 )。本研究中2种剪秋萝的生境虽然不同, 但是各自生境中的传粉者数量众多, 花粉分批呈现的规律符合对传粉者数量的适应。然而并不是所有植物的传粉模式都符合花粉呈现理论。例如, 早春短命植物黑鳞顶冰花(
Gagea nigra )的访花者稀少, 但花粉逐步呈现, 这种呈现策略降低了花粉损失的风险, 有利于应对新疆早春的极端天气(
郑荣倩等, 2016 )。
2.5.3 花粉数量和繁育系统 浅裂剪秋萝和丝瓣剪秋萝第1批散粉雄蕊的花粉量显著大于第2批雄蕊的花粉量, 环境和传粉者是重要的影响因素。天气变化会导致花粉损失, 如果开花第2天遇到不利天气, 第2批雄蕊的花粉基本不能输出, 造成雄性资源浪费(
Richardson and Stephenson, 1989 ;
陆婷和谭敦炎, 2007 )。开花第1天, 花朵颜色鲜艳, 对传粉者的吸引较大, 第1批散粉雄蕊的花粉量大于第2批雄蕊的花粉量有助于花粉尽可能的散发出去。所以剪秋萝两批雄蕊花粉量的差异是一种与环境和传粉者适应性进化的结果, 同时也是风险分摊机制的一种表现(
Henning and Weigend, 2012 )。
Cruden等(1977) 认为大量的花粉和较高的P/O值促进异交。浅裂剪秋萝和丝瓣剪秋萝花粉数量较大, P/O值分别为1 426.65和1 083.96, 且雄蕊先于雌蕊成熟; 开花第3天, 花粉生活力虽然大幅下降, 但仍有活性。如果部分花粉被带到自身柱头, 也存在自交的可能, 但柱头可授性是逐渐上升而不是开花第3天就达到峰值。开花4天后已基本采集不到花粉, 此时柱头可授性达到峰值, 这种雌雄功能时间上的分离可有效避免自花授粉。本研究结果与OCI方法检测到的剪秋萝繁育系统结论一致, 为兼性异交类型。
2.5.4 昆虫访花规律 对传粉者的观察表明, 访花时间从早晨8:00左右开始。浅裂剪秋萝8:00-11:00访花活动最频繁, 丝瓣剪秋萝11:00-12:00访花活动最频繁。对花药开裂时间的观察发现2种剪秋萝也有差异, 但多数花药上午开裂, 说明昆虫访花与花药的散粉具同步性, 散粉高峰与访花高峰重叠。对在相同时间开花并在同一区域分布的植物来说, 当它们的传粉者一致时, 就可能存在对传粉者的竞争。相同时间开花可供给更大的传粉者群落, 并能提高传粉昆虫对每种植物的访问频率(
朱兴福, 2009 )。本研究中, 2种剪秋萝花药开裂的时间不同, 使传粉者在不同时间访问不同的物种, 提高了传粉效率, 促进了植物传粉成功。
2个物种的访花高峰均出现在一天当中花粉囊开裂后温度较高且湿度较低的时间段。浅裂剪秋萝的访花高峰比丝瓣剪秋萝出现的早, 可能是生境不同所致。温度和湿度是影响花粉生活力的重要因素(
Aronne et al., 2006 )。花药在温暖干燥的气候条件下开裂快, 花粉活力较高; 而高湿度会减缓花药的开裂速度, 花粉生活力也会降低。故2种剪秋萝的访花高峰均出现在一天当中温度较高且湿度较低的时段。温度是影响蝴蝶访花的重要因素(
唐宇翀, 2015 ), 浅裂剪秋萝的生境为林缘, 相比生活在河流岸边或湿地中的丝瓣剪秋萝来说, 生境比较干燥。日出后植株周围环境温度迅速上升湿度减小, 访花者(碧翠凤蝶)访花频率升高, 生境湿度较大的丝瓣剪秋萝需稍长时间来降低植株周围环境湿度提高温度, 因而访花高峰出现的时间比浅裂剪秋萝稍晚。
The authors have declared that no competing interests exist. 参考文献 文献选项 原文顺序 文献年度倒序 文中引用次数倒序 被引期刊影响因子 [1] 邓园艺 , 喻勋林 , 雷瑞虎 , 黄娟 , 徐永福 , 杨文 , 向剑锋 (2009 ). 油茶的传粉生物学特性. 经济林研究 27, 72 -75 .[2] 段友爱 , 李庆军 (2008 ). 少花柊叶传粉生物学的研究. 植物分类学报 46, 545 -553 .[3] 冯敏 , 顾德峰 , 董然 , 王阳 (2014 ). 大花剪秋萝离体快繁技术. 东北林业大学学报 42, 90 -93, 103 .[4] 傅沛云 (1995 ). 东北植物检索表. 北京: 科学出版社. pp. 141 -160 .[5] 高江云 , 任盘宇 , 李庆军 (2005 ). 姜科、闭鞘姜科植物繁育系统与传粉生物学的研究进展. 植物分类学报 43, 574 -585 .[6] 胡适宜 (1993 ). 植物胚胎学实验方法(一)花粉生活力的测定. 植物学通报 10(2 ), 60 -62 .[7] 黄双全 (2007 ). 植物与传粉者相互作用的研究及其意义. 生物多样性 15, 569 -575 .[8] 黄双全 (2014 ). 花部特征演化的最有效传粉者原则: 证据与疑问. 生命科学 26, 118 -124 .[9] 李新蓉 (2006 ). 沙冬青属植物繁殖生物学研究. 博士论文. 乌鲁木齐: 新疆农业大学 . pp. 73 -74 .[10] 刘建华 (2009 ). 沙生蝇子草雄性先熟及花粉序次呈现的生态适应意义. 硕士论文. 乌鲁木齐: 新疆农业大学 . pp. 29 -31 .[11] 刘明财 , 崔凯峰 , 郑明艳 (2004 ). 长白山野生观赏植物引种与栽培试验. 东北林业大学学报 32(4 ), 22 -28 .[12] 陆时万 , 徐样生 , 沈敏健 (1991 ). 植物学(上册)(第2版) . 北京: 高等教育出版社. pp. 224 -225 .[13] 陆婷 , 谭敦炎 (2007 ). 动物传粉植物花粉呈现时序的进化意义. 生物多样性 15, 673 -679 .[14] 萝毅波 , 裴颜龙 , 潘开玉 , 洪德元 (1998 ). 矮牡丹传粉生物学的初步研究. 植物分类学报 36, 134 -144 .[15] 吕文 , 刘文哲 (2010 ). 雄全异株植物瘿椒树(省沽油科)的传粉生物学. 植物学报 45, 713 -722 .[16] 任明迅 (2010 ). 两性花的雄蕊运动: 多样性和适应意义. 植物生态学报 34, 867 -875 .[17] 盛连喜 , 张吉顺 (2013 ). 吉林龙湾国家级自然保护区综合科学考察报告. 长春: 吉林科学技术出版社. pp. 1 -2 .[18] 唐欣 , 李新蓉 (2017 ). 荒漠孑遗植物裸果木的开花物候特征. 植物学报 52, 487 -495 .[19] 唐宇翀 (2015 ). 金斑蝶成虫访花行为及其访花活动规律. 安徽农业科学 43, 18 -19, 48 .[20] 王艳菊 (2006 ). 剪秋萝离体培养再生体系建立及多倍体诱导研究. 硕士论文. 杭州: 浙江大学 . pp. 14 -16 .[21] 王英强 , 张奠湘 , 陈忠毅 (2005 ). 草豆蔻传粉生物学的研究. 植物分类学报 43, 37 -49 .[22] 肖宁 , 李旦 , 武永明 , 曹芹 , 夏昶 , 张金凤 (2012 ). 剪秋萝种子催芽及秋水仙碱诱导多倍体研究. 中国农学通报 16, 208 -214 .[23] 杨春锋 , 郭友好 (2005 ). 被子植物花部进化: 传粉选择作用的客观评价. 科学通报 50, 2575 -2582 .[24] 庾晓红 , 萝毅波 , 董鸣 (2008 ). 春兰(兰科)传粉生物学的研究. 植物分类学报 46, 163 -174 .[25] 张文标 , 金则新 (2008 ). 濒危植物夏蜡梅(Sinocalycanthus chinensis )的开花物候与传粉成功. 生态学报 28, 4037 -4046 .[26] 郑荣倩 , 候真珍 , 张爱勤 (2016 ). 早春短命植物黑鳞顶冰花的花粉呈现时序及其适应性. 植物学报 51, 594 -600 .[27] 朱兴福 (2009 ). 同域分布的偏花报春和海仙报春的传粉生态学研究. 硕士论文. 兰州: 兰州大学 . pp. 21 -23 .[28] Aronne G De Micco V Scala M 2006 ). Effects of relative humidity and temperature conditions on pollen fluorochromatic reaction of Rosmarinus officinalis L.(Lamia- ceae). Protoplasma 127 -130 .[29] Barrett SCH 2002 ). Sexual interference of the floral kind.Heredity 88, 154 -159 .[30] Cruden RW 1977 ). Pollen-ovule ratios: a conservative indicator of breeding systems in flowering plants.Evolution 31, 32 -46 .[31] Dafni A 1992 ). Pollination Ecology: A Practical Approach. Oxford: Oxford University Press. pp. 1 -57 .[32] Harder LD Thomson JD 1989 ). Evolutionary options for maximizing pollen dispersal of animal-pollinated plants.Am Nat 133, 323 -344 .[33] Harder LD Wilson WG 1994 ). Floral evolution and male reproductive success: optimal dispensing schedules for pollen dispersal by animal-pollinated plants.Evol Ecol 8, 542 -559 .[34] Henning T Weigend M 2012 ). Total control-pollen presentation and floral longevity in Loasaceae (Blazing Star Family) are modulated by light, temperature and pollinator visitation rates.PLoS One 7, e41121 .[35] Li QJ Xu ZF Kress WJ Xia YM Zhang L Deng XB Gao JY Bai ZL 2001 ). Pollination: flexible style that encourages outcrossing.Nature 410, 432 -432 .[36] Richardson TE Stephenson AG 1989 ). Pollen removal and pollen deposition affect the duration of the staminate and pistillate phases in Campanula rapunculoides . Am J Bot 532 -538 .[37] Thomson JD Thomson BA 1992 ). Pollen presentation and viability schedules in animal-pollinated plants: consequen- ces for reproductive success. In: Wyatt R, ed. Ecology and Evolution of Plant Reproduction, New York: Chapman & Hall Press. pp. 1 -24 .[38] Thomson JD Wilson P Valenzuela M Malzone M 2000 ). Pollen presentation and pollination syndromes, with special reference to Penstemon . Plant Spec Biol 11 -29 .[39] Wang Y Meng LH Yang YP Duan YW 2010 ). Change in floral orientation in Anisodus luridus(Solanaceae) protects pollen grains and facilitates development of fertilized ovu- les. Am J Bot 1618 -1624 .
油茶的传粉生物学特性
1
2009
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
少花柊叶传粉生物学的研究
1
2008
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
大花剪秋萝离体快繁技术
1
2014
... 浅裂剪秋萝和丝瓣剪秋萝是分布在吉林省龙湾自然保护区的石竹科(Caryophyllaceae)剪秋萝属(
Lychnis )多年生草本植物(
傅沛云, 1995 ).前期的野外观察发现, 2种剪秋萝在传粉过程中存在雄蕊分批生长传播花粉现象, 雄蕊在花部的时空配置可能在花部特征进化过程中扮演着重要角色, 且2种剪秋萝野外种群数量近年来呈下降趋势, 故对其保护和研究势在必行.目前, 关于该属植物的研究主要集中在引种栽培管理、园林应用、组织培养及多倍体育种等方面(
刘明财等, 2004 ;
王艳菊, 2006 ;
肖宁等, 2012 ;
冯敏等, 2014 ), 而对传粉生物学方面的研究报道较少.本文试图对这2种剪秋萝的传粉生物学特征进行初步探究, 并对其开花物候、雄蕊和雌蕊呈现规律、花部特征差异与传粉者访花规律进行探索, 一方面为进一步研究剪秋萝的传粉过程提供参考资料, 另一方面为深入认识剪秋萝在引种栽培过程中提高传粉效率等方面的问题提供借鉴与思路. ...
东北植物检索表
2
1995
... 浅裂剪秋萝和丝瓣剪秋萝是分布在吉林省龙湾自然保护区的石竹科(Caryophyllaceae)剪秋萝属(
Lychnis )多年生草本植物(
傅沛云, 1995 ).前期的野外观察发现, 2种剪秋萝在传粉过程中存在雄蕊分批生长传播花粉现象, 雄蕊在花部的时空配置可能在花部特征进化过程中扮演着重要角色, 且2种剪秋萝野外种群数量近年来呈下降趋势, 故对其保护和研究势在必行.目前, 关于该属植物的研究主要集中在引种栽培管理、园林应用、组织培养及多倍体育种等方面(
刘明财等, 2004 ;
王艳菊, 2006 ;
肖宁等, 2012 ;
冯敏等, 2014 ), 而对传粉生物学方面的研究报道较少.本文试图对这2种剪秋萝的传粉生物学特征进行初步探究, 并对其开花物候、雄蕊和雌蕊呈现规律、花部特征差异与传粉者访花规律进行探索, 一方面为进一步研究剪秋萝的传粉过程提供参考资料, 另一方面为深入认识剪秋萝在引种栽培过程中提高传粉效率等方面的问题提供借鉴与思路. ...
... 本研究所用浅裂剪秋萝(
Lychnis cognata Maxim)和丝瓣剪秋萝(
L. wilfordii (Regel) Maxim)野外居群分别位于保护区公路沿路林缘下和河岸边湿地, 二者野外居群相距约1 km.两物种的二歧聚伞花序具数花, 花两性, 雄蕊10枚, 雌蕊5枚合生, 子房卵形; 花期7-8月.前者株高35-90 cm (
图1A ), 叶长圆状披针形或长圆形, 多生于海拔500-1 000 m的林下或灌丛; 后者株高45-100 cm (
图1B ), 叶长圆状披针形或长披针形, 花序稍紧密, 多生于海拔250-1 200 m的湿草甸、河岸低湿地(
傅沛云, 1995 ).本研究于2016年7-9月进行. ...
姜科、闭鞘姜科植物繁育系统与传粉生物学的研究进展
1
2005
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
植物胚胎学实验方法(一)花粉生活力的测定
3
1993
... 采用花粉离体萌发法测定花粉生活力.在花朵即将开放前和开花后第1、2、3天早上8:00随机采集20朵花的花粉(由于2物种的花具2轮雄蕊, 分2批呈现花粉, 分别采集第1批和第2批雄蕊的花粉), 用镊子摘除花药后, 现场将花粉刮进装有液体培养基的离心管中.20朵花的第1批和第2批花粉分别放入不同的离心管, 液体培养基的成分为20%蔗糖和0.01%硼酸(
胡适宜, 1993 ).随后带回实验室在25°C条件下培养2小时以上, 然后在显微镜下检测花粉的萌发率(萌发率=萌发的花粉数/花粉总数) (花粉管长度大于花粉直径视为花粉萌发), 重复计数3次, 最后计算20株植物花的花粉萌发率的平均值. ...
... 柱头可授性采用联苯胺-过氧化氢法检测, 在花朵即将开放前和开花后24、48、72、96、120和144小时, 分别随机选取10朵花, 将柱头浸入联苯胺-过氧化氢反应液(1%联苯胺:3%过氧化氢:水=4:11:22, v/v/v)中.若柱头具有可授性, 则柱头周围呈蓝色并有大量的气泡出现; 若柱头不具有可授性, 则柱头周围不会呈蓝色也不会产生气泡.根据气泡多少和颜色的深浅判断柱头可受性的强弱(
胡适宜, 1993 ).用-和+表示观察到的情况.-表示柱头不具有可授性, 即无气泡与变蓝情况; +表示柱头具有可授性, 即轻微变蓝并有气泡产生; ++表示柱头具有较强的可授性, 即有较多的气泡和比较明显的变蓝情况; +++表示柱头具有极强的可授性, 即有大量的气泡和非常明显的变蓝情况. ...
... 在每个物种居群内随机选取即将开放的花各30朵, 每朵花取下全部花药.将第1批雄蕊和第2批雄蕊分别置于蒸馏水中, 定容至1 mL.摇匀后, 取20 μL悬浮液放在载玻片上, 在显微镜下记录花粉粒数, 重复计数3次.用手术刀横切子房并记录胚珠数目, 统计各花的单花花粉数和胚珠数, 最后计算花粉/胚珠(P/O)比值(
胡适宜, 1993 ).参照
Cruden (1977) 的P/O值划分繁育系统类型, P/O值在2.7-5.4区间时繁育系统为闭花授粉; 18.1-39.0时为专性自交; 31.9-396.0时为兼性自交; 244.7-2 588.0时为兼性异交; 2 108.0- 195 525.0时为专性异交.P/O值降低意味着近交程度升高, P/O值升高则表明远交程度上升. ...
植物与传粉者相互作用的研究及其意义
1
2007
... 花部特征的演化受到包括传粉者在内的多种因素影响(
黄双全, 2007 ,
2014 ).2种剪秋萝的主要传粉者相同, 但传粉综合征的差异也不显著: 花瓣均为红色且基部形成花冠管, 雄性先熟, 分批呈现雄蕊, 雄蕊散粉后平卧于花瓣上.因此, 传粉者应不是导致2种剪秋萝花部形态特征产生差异的主要因素.2种剪秋萝花部形态差异很大, 丝瓣剪秋萝的花瓣边缘撕裂程度显著大于浅裂剪秋萝, 且花部的各项表型参数值均小于浅裂剪秋萝, 造成这种花部特征差异的重要影响因素之一应是生境.由于丝瓣剪秋萝的生活环境湿度较大, 呈丝状的较小花瓣可以减少水分的附着, 同时较小的花筒直径可以减少外界水分对花粉的影响, 进而减少对传粉过程的不利影响.例如, 茄科山莨菪属铃铛子(
Anisodus luridus )花期花梗弯曲是为了避免高山环境中频繁的降雨和强烈的紫外辐射对花粉的影响, 表明生境对高山植物的花部特征具有可塑性(
Wang et al., 2010 ).因此, 环境因素对2种生境差异较大的剪秋萝花部形态特征的塑造具有重要作用.花部特征在适应性进化过程中对各种因素的折中和妥协远比我们之前了解的复杂得多, 这些问题均需要进一步研究(
杨春锋和郭友好, 2005 ). ...
花部特征演化的最有效传粉者原则: 证据与疑问
1
2014
... 花部特征的演化受到包括传粉者在内的多种因素影响(
黄双全, 2007 ,
2014 ).2种剪秋萝的主要传粉者相同, 但传粉综合征的差异也不显著: 花瓣均为红色且基部形成花冠管, 雄性先熟, 分批呈现雄蕊, 雄蕊散粉后平卧于花瓣上.因此, 传粉者应不是导致2种剪秋萝花部形态特征产生差异的主要因素.2种剪秋萝花部形态差异很大, 丝瓣剪秋萝的花瓣边缘撕裂程度显著大于浅裂剪秋萝, 且花部的各项表型参数值均小于浅裂剪秋萝, 造成这种花部特征差异的重要影响因素之一应是生境.由于丝瓣剪秋萝的生活环境湿度较大, 呈丝状的较小花瓣可以减少水分的附着, 同时较小的花筒直径可以减少外界水分对花粉的影响, 进而减少对传粉过程的不利影响.例如, 茄科山莨菪属铃铛子(
Anisodus luridus )花期花梗弯曲是为了避免高山环境中频繁的降雨和强烈的紫外辐射对花粉的影响, 表明生境对高山植物的花部特征具有可塑性(
Wang et al., 2010 ).因此, 环境因素对2种生境差异较大的剪秋萝花部形态特征的塑造具有重要作用.花部特征在适应性进化过程中对各种因素的折中和妥协远比我们之前了解的复杂得多, 这些问题均需要进一步研究(
杨春锋和郭友好, 2005 ). ...
沙冬青属植物繁殖生物学研究
1
2006
... 从花粉呈现的时间角度看, 两批雄蕊散粉间隔24小时, 这与传粉者数量和效率密切相关(
Harder and Thomson, 1989 ).在传粉者数量稀少的情况下, 植物偏好同时呈现花粉, 尽量避免最后1次访花后仍存留大量花粉造成雄性资源浪费; 在传粉者数量较多的情况下, 植物通过花粉时序性呈现可以使更多的传粉者参与传粉过程, 让成功到达柱头上的花粉数量增加, 进而提高雄性适合度(
Harder and Thomson, 1989 ;
Thomason and Thomson, 1992 ;
李新蓉, 2006 ;
陆婷和谭敦炎, 2007 ).
Thomson等(2000) 提出了简化数学模型, 该模型认为增加单个传粉者花粉移出量对总的雄性繁殖力不利, 因为1次移出大量的花粉会降低使胚珠受精的花粉比例.所以在传粉者数量充足的条件下, 限制单个传粉者1次访花转移的花粉数量和吸引更多的传粉者参与传粉是提高繁殖力的有效途径(
Thomson et al., 2000 ).本研究中2种剪秋萝的生境虽然不同, 但是各自生境中的传粉者数量众多, 花粉分批呈现的规律符合对传粉者数量的适应.然而并不是所有植物的传粉模式都符合花粉呈现理论.例如, 早春短命植物黑鳞顶冰花(
Gagea nigra )的访花者稀少, 但花粉逐步呈现, 这种呈现策略降低了花粉损失的风险, 有利于应对新疆早春的极端天气(
郑荣倩等, 2016 ). ...
沙生蝇子草雄性先熟及花粉序次呈现的生态适应意义
1
2009
... 浅裂剪秋萝和丝瓣剪秋萝的10枚雄蕊在开花过程中均分2批成熟, 且在不同时间开裂散粉.这是花粉分发的一种机制(
Harder and Wilson, 1994 ;
陆婷和谭敦炎, 2007 ).理论认为, 花粉在不同时间成熟, 可以将花粉分发给不同的访花者, 提高花粉的输出量(
Harder and Wilson, 1994 ).例如, 沙生蝇子草(
Silene olgiana )具有典型的雄性先熟、雌雄异位和花粉逐步呈现现象, 这对避免自交、雌雄干扰及增加后代适合度、保证该物种繁殖成功具有重要意义(
刘建华, 2009 ). ...
长白山野生观赏植物引种与栽培试验
1
2004
... 浅裂剪秋萝和丝瓣剪秋萝是分布在吉林省龙湾自然保护区的石竹科(Caryophyllaceae)剪秋萝属(
Lychnis )多年生草本植物(
傅沛云, 1995 ).前期的野外观察发现, 2种剪秋萝在传粉过程中存在雄蕊分批生长传播花粉现象, 雄蕊在花部的时空配置可能在花部特征进化过程中扮演着重要角色, 且2种剪秋萝野外种群数量近年来呈下降趋势, 故对其保护和研究势在必行.目前, 关于该属植物的研究主要集中在引种栽培管理、园林应用、组织培养及多倍体育种等方面(
刘明财等, 2004 ;
王艳菊, 2006 ;
肖宁等, 2012 ;
冯敏等, 2014 ), 而对传粉生物学方面的研究报道较少.本文试图对这2种剪秋萝的传粉生物学特征进行初步探究, 并对其开花物候、雄蕊和雌蕊呈现规律、花部特征差异与传粉者访花规律进行探索, 一方面为进一步研究剪秋萝的传粉过程提供参考资料, 另一方面为深入认识剪秋萝在引种栽培过程中提高传粉效率等方面的问题提供借鉴与思路. ...
1
1991
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
动物传粉植物花粉呈现时序的进化意义
3
2007
... 浅裂剪秋萝和丝瓣剪秋萝的10枚雄蕊在开花过程中均分2批成熟, 且在不同时间开裂散粉.这是花粉分发的一种机制(
Harder and Wilson, 1994 ;
陆婷和谭敦炎, 2007 ).理论认为, 花粉在不同时间成熟, 可以将花粉分发给不同的访花者, 提高花粉的输出量(
Harder and Wilson, 1994 ).例如, 沙生蝇子草(
Silene olgiana )具有典型的雄性先熟、雌雄异位和花粉逐步呈现现象, 这对避免自交、雌雄干扰及增加后代适合度、保证该物种繁殖成功具有重要意义(
刘建华, 2009 ). ...
... 从花粉呈现的时间角度看, 两批雄蕊散粉间隔24小时, 这与传粉者数量和效率密切相关(
Harder and Thomson, 1989 ).在传粉者数量稀少的情况下, 植物偏好同时呈现花粉, 尽量避免最后1次访花后仍存留大量花粉造成雄性资源浪费; 在传粉者数量较多的情况下, 植物通过花粉时序性呈现可以使更多的传粉者参与传粉过程, 让成功到达柱头上的花粉数量增加, 进而提高雄性适合度(
Harder and Thomson, 1989 ;
Thomason and Thomson, 1992 ;
李新蓉, 2006 ;
陆婷和谭敦炎, 2007 ).
Thomson等(2000) 提出了简化数学模型, 该模型认为增加单个传粉者花粉移出量对总的雄性繁殖力不利, 因为1次移出大量的花粉会降低使胚珠受精的花粉比例.所以在传粉者数量充足的条件下, 限制单个传粉者1次访花转移的花粉数量和吸引更多的传粉者参与传粉是提高繁殖力的有效途径(
Thomson et al., 2000 ).本研究中2种剪秋萝的生境虽然不同, 但是各自生境中的传粉者数量众多, 花粉分批呈现的规律符合对传粉者数量的适应.然而并不是所有植物的传粉模式都符合花粉呈现理论.例如, 早春短命植物黑鳞顶冰花(
Gagea nigra )的访花者稀少, 但花粉逐步呈现, 这种呈现策略降低了花粉损失的风险, 有利于应对新疆早春的极端天气(
郑荣倩等, 2016 ). ...
... 浅裂剪秋萝和丝瓣剪秋萝第1批散粉雄蕊的花粉量显著大于第2批雄蕊的花粉量, 环境和传粉者是重要的影响因素.天气变化会导致花粉损失, 如果开花第2天遇到不利天气, 第2批雄蕊的花粉基本不能输出, 造成雄性资源浪费(
Richardson and Stephenson, 1989 ;
陆婷和谭敦炎, 2007 ).开花第1天, 花朵颜色鲜艳, 对传粉者的吸引较大, 第1批散粉雄蕊的花粉量大于第2批雄蕊的花粉量有助于花粉尽可能的散发出去.所以剪秋萝两批雄蕊花粉量的差异是一种与环境和传粉者适应性进化的结果, 同时也是风险分摊机制的一种表现(
Henning and Weigend, 2012 ). ...
矮牡丹传粉生物学的初步研究
1
1998
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
雄全异株植物瘿椒树(省沽油科)的传粉生物学
1
2010
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
两性花的雄蕊运动: 多样性和适应意义
1
2010
... 从花粉呈现空间位置看, 初花期雄蕊处于花冠口上方, 与柱头存在着明显的雌雄异位.这种空间分配有利于异交授粉和异交传粉的各自进行, 降低雌雄功能的相互干扰(
Barrett, 2002 ).同时本研究中的2种剪秋萝的两批雄蕊通过空间位置的变化在不同时间占据最佳传粉位置, 降低两批雄蕊之间的相互干扰, 使两批雄蕊能各自与访花者充分地接触, 进而促进花粉输出.这可能表明在两性花内部, 不仅存在雌雄之间的性别干扰, 在雄蕊之间也可能存在着雄雄之间的性别内干扰, 这种性别内干扰可能成为塑造花部特征的影响因素之一(
任明迅, 2010 ).2种剪秋萝的这种雌雄蕊呈现规律既降低了雌雄功能的干扰, 又降低了雄雄功能的干扰, 是提高花粉输出率的一种适应机制. ...
吉林龙湾国家级自然保护区综合科学考察报告
1
2013
... 研究地点位于吉林省通化市辉南县龙湾自然保护区金川镇东1.5 km 的样地(126°15'55″-126°32'02″E, 42°16'20″-42°26'57″N).研究区平均海拔741 m, 年平均气温4.1°C, 年平均降水量704.2 mm, 降水时间分布受气候影响显著, 夏季雨量集中, 占全年降水的61% (
盛连喜等, 2013 ). ...
荒漠孑遗植物裸果木的开花物候特征
1
2017
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
金斑蝶成虫访花行为及其访花活动规律
1
2015
... 2个物种的访花高峰均出现在一天当中花粉囊开裂后温度较高且湿度较低的时间段.浅裂剪秋萝的访花高峰比丝瓣剪秋萝出现的早, 可能是生境不同所致.温度和湿度是影响花粉生活力的重要因素(
Aronne et al., 2006 ).花药在温暖干燥的气候条件下开裂快, 花粉活力较高; 而高湿度会减缓花药的开裂速度, 花粉生活力也会降低.故2种剪秋萝的访花高峰均出现在一天当中温度较高且湿度较低的时段.温度是影响蝴蝶访花的重要因素(
唐宇翀, 2015 ), 浅裂剪秋萝的生境为林缘, 相比生活在河流岸边或湿地中的丝瓣剪秋萝来说, 生境比较干燥.日出后植株周围环境温度迅速上升湿度减小, 访花者(碧翠凤蝶)访花频率升高, 生境湿度较大的丝瓣剪秋萝需稍长时间来降低植株周围环境湿度提高温度, 因而访花高峰出现的时间比浅裂剪秋萝稍晚. ...
剪秋萝离体培养再生体系建立及多倍体诱导研究
1
2006
... 浅裂剪秋萝和丝瓣剪秋萝是分布在吉林省龙湾自然保护区的石竹科(Caryophyllaceae)剪秋萝属(
Lychnis )多年生草本植物(
傅沛云, 1995 ).前期的野外观察发现, 2种剪秋萝在传粉过程中存在雄蕊分批生长传播花粉现象, 雄蕊在花部的时空配置可能在花部特征进化过程中扮演着重要角色, 且2种剪秋萝野外种群数量近年来呈下降趋势, 故对其保护和研究势在必行.目前, 关于该属植物的研究主要集中在引种栽培管理、园林应用、组织培养及多倍体育种等方面(
刘明财等, 2004 ;
王艳菊, 2006 ;
肖宁等, 2012 ;
冯敏等, 2014 ), 而对传粉生物学方面的研究报道较少.本文试图对这2种剪秋萝的传粉生物学特征进行初步探究, 并对其开花物候、雄蕊和雌蕊呈现规律、花部特征差异与传粉者访花规律进行探索, 一方面为进一步研究剪秋萝的传粉过程提供参考资料, 另一方面为深入认识剪秋萝在引种栽培过程中提高传粉效率等方面的问题提供借鉴与思路. ...
草豆蔻传粉生物学的研究
1
2005
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
剪秋萝种子催芽及秋水仙碱诱导多倍体研究
1
2012
... 浅裂剪秋萝和丝瓣剪秋萝是分布在吉林省龙湾自然保护区的石竹科(Caryophyllaceae)剪秋萝属(
Lychnis )多年生草本植物(
傅沛云, 1995 ).前期的野外观察发现, 2种剪秋萝在传粉过程中存在雄蕊分批生长传播花粉现象, 雄蕊在花部的时空配置可能在花部特征进化过程中扮演着重要角色, 且2种剪秋萝野外种群数量近年来呈下降趋势, 故对其保护和研究势在必行.目前, 关于该属植物的研究主要集中在引种栽培管理、园林应用、组织培养及多倍体育种等方面(
刘明财等, 2004 ;
王艳菊, 2006 ;
肖宁等, 2012 ;
冯敏等, 2014 ), 而对传粉生物学方面的研究报道较少.本文试图对这2种剪秋萝的传粉生物学特征进行初步探究, 并对其开花物候、雄蕊和雌蕊呈现规律、花部特征差异与传粉者访花规律进行探索, 一方面为进一步研究剪秋萝的传粉过程提供参考资料, 另一方面为深入认识剪秋萝在引种栽培过程中提高传粉效率等方面的问题提供借鉴与思路. ...
被子植物花部进化: 传粉选择作用的客观评价
1
2005
... 花部特征的演化受到包括传粉者在内的多种因素影响(
黄双全, 2007 ,
2014 ).2种剪秋萝的主要传粉者相同, 但传粉综合征的差异也不显著: 花瓣均为红色且基部形成花冠管, 雄性先熟, 分批呈现雄蕊, 雄蕊散粉后平卧于花瓣上.因此, 传粉者应不是导致2种剪秋萝花部形态特征产生差异的主要因素.2种剪秋萝花部形态差异很大, 丝瓣剪秋萝的花瓣边缘撕裂程度显著大于浅裂剪秋萝, 且花部的各项表型参数值均小于浅裂剪秋萝, 造成这种花部特征差异的重要影响因素之一应是生境.由于丝瓣剪秋萝的生活环境湿度较大, 呈丝状的较小花瓣可以减少水分的附着, 同时较小的花筒直径可以减少外界水分对花粉的影响, 进而减少对传粉过程的不利影响.例如, 茄科山莨菪属铃铛子(
Anisodus luridus )花期花梗弯曲是为了避免高山环境中频繁的降雨和强烈的紫外辐射对花粉的影响, 表明生境对高山植物的花部特征具有可塑性(
Wang et al., 2010 ).因此, 环境因素对2种生境差异较大的剪秋萝花部形态特征的塑造具有重要作用.花部特征在适应性进化过程中对各种因素的折中和妥协远比我们之前了解的复杂得多, 这些问题均需要进一步研究(
杨春锋和郭友好, 2005 ). ...
春兰(兰科)传粉生物学的研究
1
2008
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
濒危植物夏蜡梅(
Sinocalycanthus chinensis )的开花物候与传粉成功
1
2008
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
早春短命植物黑鳞顶冰花的花粉呈现时序及其适应性
1
2016
... 从花粉呈现的时间角度看, 两批雄蕊散粉间隔24小时, 这与传粉者数量和效率密切相关(
Harder and Thomson, 1989 ).在传粉者数量稀少的情况下, 植物偏好同时呈现花粉, 尽量避免最后1次访花后仍存留大量花粉造成雄性资源浪费; 在传粉者数量较多的情况下, 植物通过花粉时序性呈现可以使更多的传粉者参与传粉过程, 让成功到达柱头上的花粉数量增加, 进而提高雄性适合度(
Harder and Thomson, 1989 ;
Thomason and Thomson, 1992 ;
李新蓉, 2006 ;
陆婷和谭敦炎, 2007 ).
Thomson等(2000) 提出了简化数学模型, 该模型认为增加单个传粉者花粉移出量对总的雄性繁殖力不利, 因为1次移出大量的花粉会降低使胚珠受精的花粉比例.所以在传粉者数量充足的条件下, 限制单个传粉者1次访花转移的花粉数量和吸引更多的传粉者参与传粉是提高繁殖力的有效途径(
Thomson et al., 2000 ).本研究中2种剪秋萝的生境虽然不同, 但是各自生境中的传粉者数量众多, 花粉分批呈现的规律符合对传粉者数量的适应.然而并不是所有植物的传粉模式都符合花粉呈现理论.例如, 早春短命植物黑鳞顶冰花(
Gagea nigra )的访花者稀少, 但花粉逐步呈现, 这种呈现策略降低了花粉损失的风险, 有利于应对新疆早春的极端天气(
郑荣倩等, 2016 ). ...
同域分布的偏花报春和海仙报春的传粉生态学研究
1
2009
... 对传粉者的观察表明, 访花时间从早晨8:00左右开始.浅裂剪秋萝8:00-11:00访花活动最频繁, 丝瓣剪秋萝11:00-12:00访花活动最频繁.对花药开裂时间的观察发现2种剪秋萝也有差异, 但多数花药上午开裂, 说明昆虫访花与花药的散粉具同步性, 散粉高峰与访花高峰重叠.对在相同时间开花并在同一区域分布的植物来说, 当它们的传粉者一致时, 就可能存在对传粉者的竞争.相同时间开花可供给更大的传粉者群落, 并能提高传粉昆虫对每种植物的访问频率(
朱兴福, 2009 ).本研究中, 2种剪秋萝花药开裂的时间不同, 使传粉者在不同时间访问不同的物种, 提高了传粉效率, 促进了植物传粉成功. ...
1
2006
... 2个物种的访花高峰均出现在一天当中花粉囊开裂后温度较高且湿度较低的时间段.浅裂剪秋萝的访花高峰比丝瓣剪秋萝出现的早, 可能是生境不同所致.温度和湿度是影响花粉生活力的重要因素(
Aronne et al., 2006 ).花药在温暖干燥的气候条件下开裂快, 花粉活力较高; 而高湿度会减缓花药的开裂速度, 花粉生活力也会降低.故2种剪秋萝的访花高峰均出现在一天当中温度较高且湿度较低的时段.温度是影响蝴蝶访花的重要因素(
唐宇翀, 2015 ), 浅裂剪秋萝的生境为林缘, 相比生活在河流岸边或湿地中的丝瓣剪秋萝来说, 生境比较干燥.日出后植株周围环境温度迅速上升湿度减小, 访花者(碧翠凤蝶)访花频率升高, 生境湿度较大的丝瓣剪秋萝需稍长时间来降低植株周围环境湿度提高温度, 因而访花高峰出现的时间比浅裂剪秋萝稍晚. ...
1
2002
... 从花粉呈现空间位置看, 初花期雄蕊处于花冠口上方, 与柱头存在着明显的雌雄异位.这种空间分配有利于异交授粉和异交传粉的各自进行, 降低雌雄功能的相互干扰(
Barrett, 2002 ).同时本研究中的2种剪秋萝的两批雄蕊通过空间位置的变化在不同时间占据最佳传粉位置, 降低两批雄蕊之间的相互干扰, 使两批雄蕊能各自与访花者充分地接触, 进而促进花粉输出.这可能表明在两性花内部, 不仅存在雌雄之间的性别干扰, 在雄蕊之间也可能存在着雄雄之间的性别内干扰, 这种性别内干扰可能成为塑造花部特征的影响因素之一(
任明迅, 2010 ).2种剪秋萝的这种雌雄蕊呈现规律既降低了雌雄功能的干扰, 又降低了雄雄功能的干扰, 是提高花粉输出率的一种适应机制. ...
2
1977
... 在每个物种居群内随机选取即将开放的花各30朵, 每朵花取下全部花药.将第1批雄蕊和第2批雄蕊分别置于蒸馏水中, 定容至1 mL.摇匀后, 取20 μL悬浮液放在载玻片上, 在显微镜下记录花粉粒数, 重复计数3次.用手术刀横切子房并记录胚珠数目, 统计各花的单花花粉数和胚珠数, 最后计算花粉/胚珠(P/O)比值(
胡适宜, 1993 ).参照
Cruden (1977) 的P/O值划分繁育系统类型, P/O值在2.7-5.4区间时繁育系统为闭花授粉; 18.1-39.0时为专性自交; 31.9-396.0时为兼性自交; 244.7-2 588.0时为兼性异交; 2 108.0- 195 525.0时为专性异交.P/O值降低意味着近交程度升高, P/O值升高则表明远交程度上升. ...
...
Cruden等(1977) 认为大量的花粉和较高的P/O值促进异交.浅裂剪秋萝和丝瓣剪秋萝花粉数量较大, P/O值分别为1 426.65和1 083.96, 且雄蕊先于雌蕊成熟; 开花第3天, 花粉生活力虽然大幅下降, 但仍有活性.如果部分花粉被带到自身柱头, 也存在自交的可能, 但柱头可授性是逐渐上升而不是开花第3天就达到峰值.开花4天后已基本采集不到花粉, 此时柱头可授性达到峰值, 这种雌雄功能时间上的分离可有效避免自花授粉.本研究结果与OCI方法检测到的剪秋萝繁育系统结论一致, 为兼性异交类型. ...
2
1992
... 详细记录浅裂剪秋萝居群(200株)和丝瓣剪秋萝居群(50株)的始花日期、盛花期、花期持续时间及终花期等开花参数(株数不同是由于野外居群数量限制).花期的判断标准采用
Dafni (1992) 文献所述原则(居群水平上5%的植株开始开花定义为始花, 盛花期指群体中50%以上的植株同时开放所持续的时间, 终花期指5%以下的植株仍在开花.花期持续时间定义为从第1棵植株始花到最后1株花凋谢所持续的时间).此外, 在野外记录2个物种各10朵单花的开放动态、单花寿命和花药开裂时间, 在盛花期选取2个物种的花各10朵, 于早上9:00用游标卡尺测量花冠长度、花冠宽度、花瓣裂深、花萼长度、子房长度、子房宽度、雄蕊高度和雌蕊长度等表型参数. ...
...
按照Dafni (1992) 所述标准计算异交指数(out-cross- ing index, OCI).(1) 花朵或花序直径<1 mm记为0; 1-2 mm记为1; 2-6 mm记为2; > 6 mm记为3.(2) 雄蕊成熟与柱头具可授性的时间间隔: 同时或雌蕊先熟记为0; 雄蕊先熟记为1.(3) 柱头与花药处于同一高度记为0; 空间分离记为1.三者累计得OCI值. ...
2
1989
... 从花粉呈现的时间角度看, 两批雄蕊散粉间隔24小时, 这与传粉者数量和效率密切相关(
Harder and Thomson, 1989 ).在传粉者数量稀少的情况下, 植物偏好同时呈现花粉, 尽量避免最后1次访花后仍存留大量花粉造成雄性资源浪费; 在传粉者数量较多的情况下, 植物通过花粉时序性呈现可以使更多的传粉者参与传粉过程, 让成功到达柱头上的花粉数量增加, 进而提高雄性适合度(
Harder and Thomson, 1989 ;
Thomason and Thomson, 1992 ;
李新蓉, 2006 ;
陆婷和谭敦炎, 2007 ).
Thomson等(2000) 提出了简化数学模型, 该模型认为增加单个传粉者花粉移出量对总的雄性繁殖力不利, 因为1次移出大量的花粉会降低使胚珠受精的花粉比例.所以在传粉者数量充足的条件下, 限制单个传粉者1次访花转移的花粉数量和吸引更多的传粉者参与传粉是提高繁殖力的有效途径(
Thomson et al., 2000 ).本研究中2种剪秋萝的生境虽然不同, 但是各自生境中的传粉者数量众多, 花粉分批呈现的规律符合对传粉者数量的适应.然而并不是所有植物的传粉模式都符合花粉呈现理论.例如, 早春短命植物黑鳞顶冰花(
Gagea nigra )的访花者稀少, 但花粉逐步呈现, 这种呈现策略降低了花粉损失的风险, 有利于应对新疆早春的极端天气(
郑荣倩等, 2016 ). ...
... ).在传粉者数量稀少的情况下, 植物偏好同时呈现花粉, 尽量避免最后1次访花后仍存留大量花粉造成雄性资源浪费; 在传粉者数量较多的情况下, 植物通过花粉时序性呈现可以使更多的传粉者参与传粉过程, 让成功到达柱头上的花粉数量增加, 进而提高雄性适合度(
Harder and Thomson, 1989 ;
Thomason and Thomson, 1992 ;
李新蓉, 2006 ;
陆婷和谭敦炎, 2007 ).
Thomson等(2000) 提出了简化数学模型, 该模型认为增加单个传粉者花粉移出量对总的雄性繁殖力不利, 因为1次移出大量的花粉会降低使胚珠受精的花粉比例.所以在传粉者数量充足的条件下, 限制单个传粉者1次访花转移的花粉数量和吸引更多的传粉者参与传粉是提高繁殖力的有效途径(
Thomson et al., 2000 ).本研究中2种剪秋萝的生境虽然不同, 但是各自生境中的传粉者数量众多, 花粉分批呈现的规律符合对传粉者数量的适应.然而并不是所有植物的传粉模式都符合花粉呈现理论.例如, 早春短命植物黑鳞顶冰花(
Gagea nigra )的访花者稀少, 但花粉逐步呈现, 这种呈现策略降低了花粉损失的风险, 有利于应对新疆早春的极端天气(
郑荣倩等, 2016 ). ...
2
1994
... 浅裂剪秋萝和丝瓣剪秋萝的10枚雄蕊在开花过程中均分2批成熟, 且在不同时间开裂散粉.这是花粉分发的一种机制(
Harder and Wilson, 1994 ;
陆婷和谭敦炎, 2007 ).理论认为, 花粉在不同时间成熟, 可以将花粉分发给不同的访花者, 提高花粉的输出量(
Harder and Wilson, 1994 ).例如, 沙生蝇子草(
Silene olgiana )具有典型的雄性先熟、雌雄异位和花粉逐步呈现现象, 这对避免自交、雌雄干扰及增加后代适合度、保证该物种繁殖成功具有重要意义(
刘建华, 2009 ). ...
... ).理论认为, 花粉在不同时间成熟, 可以将花粉分发给不同的访花者, 提高花粉的输出量(
Harder and Wilson, 1994 ).例如, 沙生蝇子草(
Silene olgiana )具有典型的雄性先熟、雌雄异位和花粉逐步呈现现象, 这对避免自交、雌雄干扰及增加后代适合度、保证该物种繁殖成功具有重要意义(
刘建华, 2009 ). ...
1
2012
... 浅裂剪秋萝和丝瓣剪秋萝第1批散粉雄蕊的花粉量显著大于第2批雄蕊的花粉量, 环境和传粉者是重要的影响因素.天气变化会导致花粉损失, 如果开花第2天遇到不利天气, 第2批雄蕊的花粉基本不能输出, 造成雄性资源浪费(
Richardson and Stephenson, 1989 ;
陆婷和谭敦炎, 2007 ).开花第1天, 花朵颜色鲜艳, 对传粉者的吸引较大, 第1批散粉雄蕊的花粉量大于第2批雄蕊的花粉量有助于花粉尽可能的散发出去.所以剪秋萝两批雄蕊花粉量的差异是一种与环境和传粉者适应性进化的结果, 同时也是风险分摊机制的一种表现(
Henning and Weigend, 2012 ). ...
1
2001
... 传粉是植物繁殖过程中的一个重要环节, 是成熟花粉从雄蕊花药中散出后, 传送到雌蕊柱头上的过程(
陆时万等, 1991 ).近年来, 国内****对传粉生物学的研究主要集中在以下两方面: (1) 对濒危物种(如夏蜡梅(
Sinocalycanthus chinensis )和矮牡丹(
Paeonia suffruticosa subsp
. spontanea ))的传粉过程及花粉散布规律的研究(
萝毅波等, 1998 ;
张文标和金则新, 2008 ;
吕文和刘文哲, 2010 ;
唐欣和李新蓉, 2017 ); (2) 对一些经济作物和观赏植物(如油茶(
Camellia oleifera )和春兰(
Cymbidium goeringii )) (
庾晓红等, 2008 ;
邓园艺等, 2009 )从应用方面展开一系列的传粉过程研究.此外, ****们也发现一些在传粉过程中具有独特性表达方式和散粉策略的植物, 如具有花柱卷曲式(
Li et al., 2001 ;
王英强等, 2005 ;
高江云等, 2005 )和爆发式运动(
段友爱和李庆军, 2008 )等传粉方式的植物.植物的传粉生物学特征在其种群生活史进化过程中是一个十分重要的选择因素, 其表型特征会受到生物和非生物因素的长期选择, 因而从传粉生物学特征入手, 可以深入认识植物的繁殖特征进化及适应过程.另外, 以往的传粉生物学研究多针对单一植物进行, 很少有物种间的比较, 特别是同域分布的同属植物传粉特性的比较研究.基于以上分析, 我们对前期野外调查中发现的具有较为独特传粉特性的2种剪秋萝属植物(浅裂剪秋萝(
Lychnis cognata )和丝瓣剪秋萝(
L. wilfordii ))进行了详细探索. ...
1
1989
... 浅裂剪秋萝和丝瓣剪秋萝第1批散粉雄蕊的花粉量显著大于第2批雄蕊的花粉量, 环境和传粉者是重要的影响因素.天气变化会导致花粉损失, 如果开花第2天遇到不利天气, 第2批雄蕊的花粉基本不能输出, 造成雄性资源浪费(
Richardson and Stephenson, 1989 ;
陆婷和谭敦炎, 2007 ).开花第1天, 花朵颜色鲜艳, 对传粉者的吸引较大, 第1批散粉雄蕊的花粉量大于第2批雄蕊的花粉量有助于花粉尽可能的散发出去.所以剪秋萝两批雄蕊花粉量的差异是一种与环境和传粉者适应性进化的结果, 同时也是风险分摊机制的一种表现(
Henning and Weigend, 2012 ). ...
1
1992
... 从花粉呈现的时间角度看, 两批雄蕊散粉间隔24小时, 这与传粉者数量和效率密切相关(
Harder and Thomson, 1989 ).在传粉者数量稀少的情况下, 植物偏好同时呈现花粉, 尽量避免最后1次访花后仍存留大量花粉造成雄性资源浪费; 在传粉者数量较多的情况下, 植物通过花粉时序性呈现可以使更多的传粉者参与传粉过程, 让成功到达柱头上的花粉数量增加, 进而提高雄性适合度(
Harder and Thomson, 1989 ;
Thomason and Thomson, 1992 ;
李新蓉, 2006 ;
陆婷和谭敦炎, 2007 ).
Thomson等(2000) 提出了简化数学模型, 该模型认为增加单个传粉者花粉移出量对总的雄性繁殖力不利, 因为1次移出大量的花粉会降低使胚珠受精的花粉比例.所以在传粉者数量充足的条件下, 限制单个传粉者1次访花转移的花粉数量和吸引更多的传粉者参与传粉是提高繁殖力的有效途径(
Thomson et al., 2000 ).本研究中2种剪秋萝的生境虽然不同, 但是各自生境中的传粉者数量众多, 花粉分批呈现的规律符合对传粉者数量的适应.然而并不是所有植物的传粉模式都符合花粉呈现理论.例如, 早春短命植物黑鳞顶冰花(
Gagea nigra )的访花者稀少, 但花粉逐步呈现, 这种呈现策略降低了花粉损失的风险, 有利于应对新疆早春的极端天气(
郑荣倩等, 2016 ). ...
2
2000
... 从花粉呈现的时间角度看, 两批雄蕊散粉间隔24小时, 这与传粉者数量和效率密切相关(
Harder and Thomson, 1989 ).在传粉者数量稀少的情况下, 植物偏好同时呈现花粉, 尽量避免最后1次访花后仍存留大量花粉造成雄性资源浪费; 在传粉者数量较多的情况下, 植物通过花粉时序性呈现可以使更多的传粉者参与传粉过程, 让成功到达柱头上的花粉数量增加, 进而提高雄性适合度(
Harder and Thomson, 1989 ;
Thomason and Thomson, 1992 ;
李新蓉, 2006 ;
陆婷和谭敦炎, 2007 ).
Thomson等(2000) 提出了简化数学模型, 该模型认为增加单个传粉者花粉移出量对总的雄性繁殖力不利, 因为1次移出大量的花粉会降低使胚珠受精的花粉比例.所以在传粉者数量充足的条件下, 限制单个传粉者1次访花转移的花粉数量和吸引更多的传粉者参与传粉是提高繁殖力的有效途径(
Thomson et al., 2000 ).本研究中2种剪秋萝的生境虽然不同, 但是各自生境中的传粉者数量众多, 花粉分批呈现的规律符合对传粉者数量的适应.然而并不是所有植物的传粉模式都符合花粉呈现理论.例如, 早春短命植物黑鳞顶冰花(
Gagea nigra )的访花者稀少, 但花粉逐步呈现, 这种呈现策略降低了花粉损失的风险, 有利于应对新疆早春的极端天气(
郑荣倩等, 2016 ). ...
... 提出了简化数学模型, 该模型认为增加单个传粉者花粉移出量对总的雄性繁殖力不利, 因为1次移出大量的花粉会降低使胚珠受精的花粉比例.所以在传粉者数量充足的条件下, 限制单个传粉者1次访花转移的花粉数量和吸引更多的传粉者参与传粉是提高繁殖力的有效途径(
Thomson et al., 2000 ).本研究中2种剪秋萝的生境虽然不同, 但是各自生境中的传粉者数量众多, 花粉分批呈现的规律符合对传粉者数量的适应.然而并不是所有植物的传粉模式都符合花粉呈现理论.例如, 早春短命植物黑鳞顶冰花(
Gagea nigra )的访花者稀少, 但花粉逐步呈现, 这种呈现策略降低了花粉损失的风险, 有利于应对新疆早春的极端天气(
郑荣倩等, 2016 ). ...
1
2010
... 花部特征的演化受到包括传粉者在内的多种因素影响(
黄双全, 2007 ,
2014 ).2种剪秋萝的主要传粉者相同, 但传粉综合征的差异也不显著: 花瓣均为红色且基部形成花冠管, 雄性先熟, 分批呈现雄蕊, 雄蕊散粉后平卧于花瓣上.因此, 传粉者应不是导致2种剪秋萝花部形态特征产生差异的主要因素.2种剪秋萝花部形态差异很大, 丝瓣剪秋萝的花瓣边缘撕裂程度显著大于浅裂剪秋萝, 且花部的各项表型参数值均小于浅裂剪秋萝, 造成这种花部特征差异的重要影响因素之一应是生境.由于丝瓣剪秋萝的生活环境湿度较大, 呈丝状的较小花瓣可以减少水分的附着, 同时较小的花筒直径可以减少外界水分对花粉的影响, 进而减少对传粉过程的不利影响.例如, 茄科山莨菪属铃铛子(
Anisodus luridus )花期花梗弯曲是为了避免高山环境中频繁的降雨和强烈的紫外辐射对花粉的影响, 表明生境对高山植物的花部特征具有可塑性(
Wang et al., 2010 ).因此, 环境因素对2种生境差异较大的剪秋萝花部形态特征的塑造具有重要作用.花部特征在适应性进化过程中对各种因素的折中和妥协远比我们之前了解的复杂得多, 这些问题均需要进一步研究(
杨春锋和郭友好, 2005 ). ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}