Abstract: Abscisic acid (ABA) acts as a major phytohormone in regulating seed dormancy and germination. Endogenous ABA level in the seed and embryo sensitivity to ABA jointly regulate the seed dormancy and germination processes, ensuring that the dormant seed can maintain self-reproductive ability under adverse situations and germinate under optimal conditions. A number of key genes are involved in regulating the ABA metabolism and signaling transduction pathway, and these genes accurately control the processes of embryogenesis, seed maturation, dormancy, and germination via a complicated regulation network. In this paper, we review the latest research in the molecular mechanism of ABA-regulated seed dormancy and germination and pinpoint prospects for future research.
种子休眠和萌发是植物生长发育过程中2个紧密相连的阶段, 受到种子发育的内源因素和外界环境因素共同调控。Finch-Savage和Leubner-Metzger (2006)对种子休眠和萌发进行了定义: 休眠是指具有活力的种子在适宜的环境条件下不能萌发的状态; 而萌发则是种子内胚根突破外周的组织结构(胚乳和种皮)而展露在种皮之外的过程。脱落酸(abscisic acid, ABA)是调控种子休眠和萌发过程最重要的植物激素, 具有诱导和维持种子休眠、抑制种子萌发及幼苗生长发育的作用。十几年来, ABA调控种子休眠和萌发的研究取得了重大进展(Holdsworth et al., 2008; Weitbrecht et al., 2011; Shu et al., 2016)。刘一灵等(2014)从ABA代谢与种子发育、穗萌和种子萌发等方面进行了综述。操春燕等(2017)则从ABA与细胞分裂素(cytokinin, CK)和赤霉素(gibberellin, GA)等相互作用调控种子萌发的角度概述了相关的最新研究进展。近年来, 随着参与种子ABA代谢和ABA信号基因相关突变体研究的深入, ABA调控种子休眠和萌发的分子机制已逐渐明晰, 本文参考ABA代谢基因、种子成熟基因以及ABA信号基因的遗传功能和表达调控的最新研究成果(表1), 综述ABA调控种子休眠和萌发的分子机制, 以期为植物激素的作用机理研究和农作物分子设计育种提供重要的理论依据。
参与种子休眠的关键基因
Key genes involved in seed dormancy
基因名称
突变体休眠能力
基本功能
参考文献
NCED6/9
变化不显著
ABA合成途径关键代谢酶
Lefebvre et al., 2006
CYP707A1/2
增强
ABA代谢途径关键代谢酶, 其表达受ABI4调控
Kushiro et al., 2004; Okamoto et al., 2006; Shu et al., 2013
ABI3
降低
促进种子发育, 抑制种子萌发
Raz et al., 2001; Dekkers et al., 2016
FUS3
降低
促进种子发育, 增强NCED6和NCED9的表达
Raz et al., 2001; Tiedemann et al., 2008; Yamamoto et al., 2010
LEC2
降低
促进种子发育, 促进LEC1、FUC3和DOG1的表达
Raz et al., 2001; Braybrook et al., 2006; Stone et al., 2008
LEC1/L1L
降低
促进种子发育, 激活ABI3、FUS3和LEC2的表达
Raz et al., 2001; Kagaya et al., 2005; To et al., 2006
DEP
降低
促进ABI3的表达
Barrero et al., 2010
DOG1
降低
种子休眠正向调控因子
Née et al., 2017
ABI4
降低
促进种子发育, 是ABA/GA的关键调控因子
Shu et al., 2013
SPT
Col背景中增强; Ler背景中降低
调控ABI4表达, 在Col和Ler背景下功能相反
Belmonte et al., 2013; Vaistij et al., 2013
CHO1
降低
促进ABA合成, 抑制ABA降解
Yano et al., 2009
MYB96
降低
促进种子休眠, 抑制种子萌发
Lee et al., 2015a, 2015b
ABI5
变化不显著
不直接参与种子休眠调控, 但负调控种子萌发
Piskurewicz et al., 2008; Kanai et al., 2010
bZIP10/25/53
降低
协同促进种子发育
Alonso et al., 2009
WRKY41
降低
促进ABI3转录
Ding et al., 2014
DAG1
降低
促进ABA而抑制GA积累, 正调控种子休眠
Boccaccini et al., 2014, 2016
DOF6
未提及
与RGL2构成复合体, 共同促进种子初级休眠
Ravindran et al., 2017
KYP/SUVH4
增强
抑制DOG1和ABI3转录, 负调控种子休眠
Zheng et al., 2012
LDL1/LDL2
增强
抑制DOG1表达, 负调控种子休眠
Zhao et al., 2015
1 ABA调控种子成熟和休眠的分子机制ABA在促进种子成熟及诱导和维持种子休眠中发挥重要作用。在种子发育过程中, ABA对种子休眠的诱导和维持与种子内源ABA水平及种胚发育信号密切相关(Holdsworth et al., 2008; Shu et al., 2016)。在种子发育过程中, 内源ABA水平的降低导致成熟种子的休眠能力下降甚至丧失, 而ABA含量的增加则会使种子休眠能力增强及萌发推迟(Okamoto et al., 2006; Nonogaki et al., 2014)。 1.1 种子ABA合成代谢途径的基因表达调控种子内源ABA积累取决于ABA合成与代谢途径的动态平衡。在拟南芥(Arabidopsis thaliana)中, ABA合成初期是在细胞质体中进行, 胡萝卜素前体被AtABA1/ ZEP编码的玉米黄质环氧酶(zeaxanthin epoxidase, ZEP)催化形成玉米黄质, 再经9-顺式-环氧类胡萝卜素双加氧酶(9-cis-epoxycarotenoid dioxygenases, NCEDs)氧化形成黄质醛(xanthoxin), 后者是ABA合成的关键阶段。随后, 黄质醛被转运至细胞溶胶中, 经短链醇脱氢酶(short-chain dehydrogenase, 由At- ABA2编码)和醛氧化酶(aldehyde oxidase 3, AAO3)催化及AtABA3编码的钼辅因子硫化酶(molybdenum cofactor sulfurase)的活化, 最终形成具有生物活性的ABA (Lin et al., 2007)。ABA代谢主要包括2条途径: 羟基化途径以及葡糖基转移酶参与的螯合途径。前者通过细胞色素P450单加氧酶(也称为ABA 8'-羟化酶)催化, 使ABA转变为红花菜豆酸(phaseic acid, PA)和二氢红花菜豆酸(dihydrophaseic acid, DPA), 从而失去生物活性。拟南芥CYP707A家族的4个成员在ABA代谢过程中起重要作用(Kushiro et al., 2004)。螯合途径则由葡糖基转移酶(glucosyltransferases, UGTs)催化, 使游离态ABA上的羧基与葡糖基共价结合形成ABA葡糖基酯(ABA glucosyl ester, ABA-GE), 从而失去活性。 ABA-GE被认为是ABA的贮存和长距离运输的非活性形态(Zhang, 2014)。 高等植物种子发育包括胚和胚乳发育(形态发生)及种子成熟, 在该过程中内源ABA的含量会出现2个高峰。拟南芥种子进入成熟阶段(形成心形胚)前, 首个ABA积累高峰主要来自母体韧皮部中ABA的合成, ABA由母体运输至胚内, 抑制种子成熟前的胎萌; 进入成熟阶段后, 内源ABA积累以胚内合成为主, 种子成熟时达到第2个高峰; 且ABA缺陷型突变体在成熟阶段, 母体合成的ABA仍会向胚内运输(Kanno et al., 2010)。对拟南芥ABA合成途径相关基因突变体的研究表明, 拟南芥ABA1 (与烟草(Nicotiana tabacum) ABA2同源)缺失突变体aba1呈现植株萎蔫、内源ABA缺乏及ABA诱导的基因表达下调等表型。aba2和aba3表型与aba1相同, 且种子休眠能力下降(Finke- lstein et al., 2002)。AtABA2过量表达株系种子中内源ABA含量高于野生型, 萌发延迟(Lin et al., 2007)。AtNCED6和AtNCED9均在种子发育期特异性表达, 前者仅在胚乳中表达, 而后者在胚和胚乳中都表达, 是诱导和维持胚休眠的主要基因(Lefebvre et al., 2006)。AtNCED6和AtNCED9存在功能冗余, 拟南芥nced6/nced9双突变体种子休眠程度减弱而单突变体无相关表型(Lefebvre et al., 2006)。种子成熟后期, 胚内含水量下降, AtNCED5表达上调, 与AtNCED6和AtNCED9协同调控种子休眠, 三突变体nced5/ nced6/nced9休眠水平显著低于三者间的双突变体(Frey et al., 2012)。值得关注的是, 只有胚内合成的ABA能诱导和维持种子的初级休眠, 母体合成或胚发育期间外源施加ABA均不能诱导种子的初级休眠(Karssen et al., 1983; Koornneef and Karssen, 1994)。 在ABA代谢方面, AtCYP707A1主要在种子成熟中期的种皮和胚乳中表达, 导致ABA水平下降, 突变体cyp707a1种子表现出较野生型更强的休眠能力。AtCYP707A2主要在种子成熟晚期的种皮、胚乳和胚中以及吸胀的种子中表达; cyp707a2干种子ABA含量较野生型高5倍, 呈深度休眠表型; 吸胀后AtCYP- 707A2表达量显著增强, 表明CYP707A2是种子萌发过程中的主效基因。AtCYP707A3主要在胚以及成苗莲座叶中表达, 受种子萌发和干旱胁迫的诱导, 但cyp707a3突变体种子休眠程度不如cyp707a1和cyp707a2变化显著, 干种子ABA含量与野生型相近, 说明CYP707A3可能并不调控种子的发育, 而是调控萌发后幼苗生长及胁迫应答过程中ABA的积累。At- CYP707A4主要在种子发育中后期的角果皮中表达, 而在种子中无表达活性, 暗示AtCYP707A4并不调控种子发育过程中的ABA积累(Kushiro et al., 2004; Okamoto et al., 2006)。双突变体表型分析显示, cyp707a1/cyp707a2种子休眠程度高于cyp707a2/ cyp707a3, 而后者又高于cyp707a1/cyp707a3 (Ok- amoto et al., 2006)。
1.2 种子成熟相关转录因子的表达调控种子成熟阶段是休眠诱导的关键时期, 种子在此期间经历一系列的变化, 包括营养物质(淀粉、贮藏蛋白和脂类)的贮藏、脱水耐性的获得、胚发育的停滞以及种子休眠的建立。近年来, 种子成熟和激素合成代谢基因的表达调控机制及其遗传功能方面的研究取得了很大进展。 1.2.1 B3型转录因子和LEC1型NF-YB转录因子 拟南芥中, 调控种胚发育和种子成熟的转录因子主要由B3型转录因子的AFL支线ABI3 (ABA INSENSITIVE 3)、 FUS3 (FUSCA 3)和LEC2 (LEAFY COTY- LEDON 2), 以及2个LEC1型NF-YB (NUCLEAR FA- CTOR Y B subunit)家族成员LEC1和L1L (LEC1- LIKE)组成, 统称为LAFL。B3型转录因子最先是从玉米(Zea mays) vp1 (viviparous1)和拟南芥abi3 (abscisic acid insensitive 3)突变体中被发现, 属于植物特有的B3型DNA结合结构域蛋白家族。该家族蛋白能识别种子特异基因启动子的RY重复或Sph元件的保守序列, 具有促进种胚发育和成熟的功能(Jia et al., 2014)。其中, ABI3是种子休眠与萌发的主要调控因子, 具有调控种皮色素改变、诱导种子休眠和促进贮藏蛋白积累等功能(Dekkers et al., 2016)。在ABI3上游, Barrero等(2010)发现1个RING-finger蛋白(一类E3泛素连接酶)的编码基因DEP (DESPIERTO), 在种子发育期间调控ABI3表达并影响休眠的诱导, 突变体dep-1种子休眠完全丧失。FUS3在种子发育中后期增强NCED6、NCED9和ABA2的表达, 促进种子干旱耐受性的获得, 并决定子叶上皮细胞的一致性(Tiedemann et al., 2008; Yamamoto et al., 2010)。LEC2在胚胎发育早中期表达, 促进贮藏蛋白和油体蛋白的积累, 并参与种子成熟阶段的基因表达调控, 如DOG1 (DELAY OF GERMINATION 1) (Braybr- ook et al., 2006)。拟南芥DOG1蛋白是种子休眠的关键调控因子, 通过抑制特定蛋白磷酸酶2C (protein phosphatase 2Cs, PP2C)活性来调控休眠, 且其蛋白水平决定了休眠释放的时机。dog1突变体种子休眠完全缺失(Née et al., 2017)。LEC1型NF-YB家族转录因子具有结合CCAAT-box的结构域, 通过调控AFL基因促进种子贮藏蛋白(seed storage protein, SSP)基因的表达(Kagaya et al., 2005)。LEC1在胚胎发育早期表达并促进种子胚胎形成(Lotan et al., 1998)。L1L在发育中的胚胎(稍晚于LEC1)以及营养器官中表达, 表明除正向调控种胚发育外, L1L还具有与LEC1不同的调控功能(Kwong et al., 2003)。 LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟。研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008)。种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006)。转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013)。突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007)。但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015)。突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015)。突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015)。 B3型转录因子家族的VAL (VIVIPAROUS1/ ABI3-LIKE)支线VAL1/HSI2 (HIGH-LEVEL EXPRESSION OF SUGAR-INDUCIBLE GENE 2)、VAL2/HSL1 (HSI2-LIKE1)和VAL3/HSL2 (HSI2- LIKE2)参与对LAFL的转录抑制(Suzuki et al., 2007)。VAL蛋白除含有B3结构域外, 还具有与染色质因子相关的PHD-L (plant homeodomain-like)和CW (cys- and trp-containing domain)结构域, 以及EAR (ethy- lene response factor-associated repression)转录抑制结构域(Perry and Zhao, 2003; Bienz, 2006)。在种子萌发过程中, VAL蛋白通过CW结构域, 将组蛋白脱乙酰酶复合物(histone deacetylase complex, HDAC)招募至含有Sph/RY元件的LAFL基因上, 并抑制它们的表达, 从而正调控种子萌发及幼苗生长。有趣的是, 已有研究显示VAL1是FUS3的直接靶标(Wang and Perry, 2013)。 1.2.2 AP2型转录因子 APETALA 2/乙烯应答元件结合因子(AP2/ERF)是植物特有的转录因子家族, 调控植物生长发育及胁迫响应。目前已证实在拟南芥和水稻(Oryza sativa)中, AP2型转录因子通过调节ABA/GA的平衡来调控种子的休眠和萌发。拟南芥ABI4通过结合CYP707A1和CYP707A2基因启动子的CCAC顺式元件抑制其表达, 同时直接促进NCED6和GA2ox7 (GA2 oxidase 7)基因的表达, 且ABI4缺失突变体能恢复ga1-t、cyp707a1和cyp707a2突变体不萌发的表型, 证明ABI4通过正调控ABA合成和负调控GA合成来调控种子的休眠与萌发(Shu et al., 2013, 2018)。ABI4基因的转录受到控制种子休眠的关键转录因子SPT (SPATULA)的调控, 提示SPT-ABI4在种子休眠的诱导和维持方面具有重要作用(Vaistij et al., 2013)。拟南芥CHO1 (CHOTTO1)具有双重的AP2结构, CHO1在ABI4下游, 正调控种子休眠, 通过促进ABA1/ZEP和NCED9的表达, 抑制CYP707A2和CYP707A3的表达, 调控种子萌发过程中ABA介导的GA合成抑制过程(Yano et al., 2009)。拟南芥ORA47 (octadeca- noid-responsive AP2/ERF-domain transcription fac- tor 47)和DREB2C通过正调控NCED3和NCED9的表达, 促进ABA合成, 从而对非生物胁迫作出响应(Je et al., 2014; Chen et al., 2016)。此外, 转录因子MYB96通过调控ABI4和ABA合成基因NCED2与NCED6的表达, 促进种子休眠而抑制萌发(Lee et al., 2015a, 2015b)。 1.2.3 bZIP型转录因子 转录组分析显示, 拟南芥干种子中含有种类丰富的mRNA, 包括种子发育晚期丰富蛋白(late embryogenesis abundant, LEA)和其它贮藏蛋白基因的mRNA。绝大多数高丰度mRNA编码基因的启动子中含有ABA应答元件(ABA response element, ABRE)。拟南芥bZIP型转录因子具有亮氨酸拉链结构(leucine zipper), 能特异识别基因启动子ABRE应答元件中的ACGT核心序列, 调控相应基因的表达。ABI5作为bZIP型转录因子家族中的重要成员, 能够激活种胚内ABA调控基因的表达, 如LEA蛋白的编码基因EM1 (EARLY METHIONINE-LABELED 1)、EM6和LEAD- 34 (Finkelstein and Lynch, 2000)。酵母单杂交实验结果显示, ABI5直接与EM6启动子的ABRE结合从而促进其表达(Nakamura et al., 2001; Carles et al., 2002)。种子休眠的关键转录因子DOG1是通过促进ABI5转录, 从而激活下游LEA和HSP (HEAT SHO- CK PROTEIN)基因的表达(Dekkers et al., 2016)。ABI5不直接参与种子休眠调控, 但负调控种子的萌发, 说明ABA调控种子休眠和萌发是通过2个不同的信号转导途径(Piskurewicz et al., 2008; Kanai et al., 2010)。ABI5能与ABI3进行体外蛋白互作, 且位于ABI3的下游, 激活下游基因表达(Nakamura et al., 2001; Lopez-Molina et al., 2002)。此外, 拟南芥转录因子bZIP53通过结合种子贮藏蛋白基因(如SSP和LEA)启动子G-box元件, 促进相关基因表达, 进而调控种子成熟。体外结合实验显示, 当bZIP53与bZIP10或bZIP25异源二聚化后, 能增强bZIP53与G-box的结合活性(Alonso et al., 2009)。同时, bZIP10和bZIP25也能促进SSP基因的表达(Lara et al., 2003), 表明这3个转录因子在促进种子成熟过程中具有协同作用。 1.2.4 其它转录因子 WRKY转录因子因蛋白N端具有保守的氨基酸序列WRKYGQK而得名, 其蛋白C端具有锌指结构域。拟南芥WRKY41能直接结合在ABI3启动子3个邻近W-box元件, 从而促进ABI3转录。wrky41缺失突变体与abi3表型相似, 突变体种子表现出休眠程度降低和ABA不敏感表型(Ding et al., 2014)。 植物特有的转录因子DOF蛋白(DNA BINDING WITH ONE FINGER)具有1个锌指型DNA结合域, 参与调控植物不同的生理过程。转录因子DAG1 (DOF AFFECTING GERMINATION 1)在种子成熟过程中表达, 影响种子胚胎发育。dag1缺失突变体种子休眠能力减弱, 且胚胎畸形(Boccaccini et al., 2014)。DAG1受表观遗传调控, 直接与GA3ox1 (GA3 oxidase 1)和CYP707A2的启动子结合, 抑制其转录, 从而抑制GA并促进ABA积累, 调控种子休眠。突变体dag1种子内源ABA含量降低, 而GA含量升高(Bocc- accini et al., 2016)。在种子成熟和休眠过程中, DAG1通过结合DAG2的启动子抑制DAG2的表达, 但dag2突变体种胚表型与野生型无差异, 表明DAG2并不影响胚胎发育和种子休眠。进一步研究表明, DAG2在PIL5 (PHYTOCHROME INTERACTING FACTOR3- LIKE 5)的下游, 正调控光促进的种子萌发过程(Santopolo et al., 2015)。最新研究表明, DOF6蛋白能与RGL2 (RGA-LIKE2) (GA信号途径的负调控蛋白)构成复合体, 共同结合在RGL2下游靶标GATA12的DOF相关基序上, 激活其表达, 从而促进种子初级休眠的维持; DOF6表达上调株系种子的休眠能力显著增强(Ravindran et al., 2017)。
1.3 种子成熟的染色质结构调控种子休眠的维持可能与染色质结构相关, 促进种子萌发的基因由于空间位阻不能被转录因子结合而处于静止状态(Shu et al., 2016)。拟南芥KYP/SUVH4编码一种H3 Lys 9甲基转移酶, 为H3K9甲基化所需; 同时KYP/SUVH4负调控种子休眠, 其缺失突变体休眠程度增强。研究表明, ABA可能通过抑制KYP/SUVH4的表达从而促进种子休眠(Zheng et al., 2012)。组蛋白乙酰转移酶(histone acetyltransferases)通过调节染色质活力来调控基因表达。大麦(Hordeum vulgare)种子经外源ABA处理后, 组蛋白乙酰转移酶基因HvGNAT/MYST表达上调(Papaefthimiou et al., 2010)。表观遗传调控因子HUB1 (HISTONE MON- OU-BIQUITINATION 1 )和RDO2 (REDUCED DOR- MANCY 2)在休眠诱导期受到ABA的强烈诱导(Liu et al., 2011)。此外, 组蛋白脱甲基酶LDL1 (LYSINES- PECIFIC DEMETHYLASE LIKE 1)和LDL2通过抑制DOG1、ABA2和ABI3的表达, 负调控种子休眠(Zhao et al., 2015)。KYP/SUVH4和SUVH5在种子成熟期抑制DOG1和ABI3的表达(Zheng et al., 2012), 且DO- G1的染色质结构重塑参与种子的休眠循环过程(Foo- titt et al., 2015), 表明DOG1可能通过2条不同的途 径诱导种子休眠。关于ABA通过影响染色质结构重塑调节种子休眠与萌发的分子机制, 目前尚不完全明确。
2 ABA调控种子萌发的分子机制种子萌发是指种子吸水膨胀, 种胚重新恢复生长, 胚根突破胚乳和种皮的一个连续性事件。种子萌发的启动依赖于种子内源ABA的降解和种胚对ABA的脱敏作用。 2.1 内源ABA代谢对种子萌发的调控种子休眠的维持取决于种子内源ABA与GA含量的比值(ABA/GA), 而休眠解除是由种子内源ABA含量下降和GA含量升高所致(Cadman et al., 2006)。拟南芥休眠型Cvi (cape verde islands)和非休眠型Col (Columbia)种子吸胀2-3小时后, CYP707A2转录均被激活, 导致ABA含量迅速下降, 说明CYP707A2是种子吸胀过程中ABA降解的主要代谢酶(Preston et al., 2009)。研究表明, 休眠型Cvi种子后熟作用的休眠释放(dormancy release)与CYP707A2和GA3ox1基因表达相关。cyp707a2缺失突变体种子中GA4含量和GA3ox1的转录水平均有所下降, 表明种子萌发时ABA通过负调控GA合成来抑制种子萌发(Yano et al., 2009)。同样, GA也能够抑制ABA的合成途径, ABA与GA的拮抗作用主要受AP2型转录因子的调控(Oh et al., 2007; Shu et al., 2013)。种子吸胀过程中, GA合成基因GA3ox和GA20ox高表达, 促使GA在种胚内大量积累(Ogawa et al., 2003)。种子萌发早期, ABA/GA比值下降至原有水平的1/3; 而在萌发后期, ABA/GA比值下降至1/10 (Weitbrecht et al., 2011)。淀粉酶作为种子萌发过程中重要的水解酶, 通过分解胚乳中的营养物质来为萌发中的胚提供能量和营养。Kondhare等(2014)使用外源激素处理小麦(Triticum aestivum)和水稻种子时, 发现ABA抑制而GA促进α-淀粉酶的活性, 可见ABA/GA平衡还能调控种子内部营养物质的改变。 近年来, 转录组测序为研究后熟作用的休眠释放机制提供了重要线索。拟南芥新鲜种子和后熟种子的转录组分析结果显示, 其大部分转录本并无差异, 仅NADPH-氧化酶编码基因RBOHB (RESPIRATORY BURST OXIDASE HOMOLOG B)的表达存在明显差异, 表明后熟作用促进种子休眠释放可能通过蛋白质的氧化作用而非调节细胞代谢功能实现(Müller et al., 2009)。另外, 拟南芥休眠型Cvi干种子与其后熟种子的转录组分析表明, 在Cvi的珠孔胚乳中, ABA合成酶基因NCED2和GA降解酶基因GA2ox6转录本丰度有所升高(Dekkers et al., 2016)。
2.2 ABA信号对种子萌发的调控机制ABA通过特定的信号转导途径抑制种子萌发。ABA信号转导起始于ABA受体对ABA信号的感知。2009年, 2个实验室各自独立发现1个含有START (STeroidogenic Acute Regulatory Related Lipid Transfer)特征区的蛋白质PYR (PYRABACTIN RESISTANCE)/ PYL (PYR1-LIKE)/RCAR (REGULATORY COMPO- NENTS OF ABA RECEPTORS)家族成员能够结合ABA, 是一种细胞内ABA受体。其作用机理是PYR/ PYL/RCAR通过与下游信号调节子PP2C直接相互作用, 抑制PP2C的活性, 解除PP2C对其下游信号调节子SNF1相关蛋白激酶2 (SNF1-related protein kinase 2, SnRK2)的抑制, 从而使SnRK2能够通过磷酸化激活下游转录因子ABI5/ABF4 (ABRE binding factors), 实现ABA生理效应(图1) (Ma et al., 2009; Park et al., 2009; Hubbard et al., 2010; Hauser et al., 2011)。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-542/img_1.png<b>图1</b> 以PYR/PYL/RCAR和ABAR/CHLH为受体的ABA信号转导通路模型<br/>细胞质ABA受体PYR/PYL/RCAR在静息状态下以二聚体形式存在, 当与ABA结合后则以单体形式与PP2Cs结合, 解除对SnRK2s的抑制, 磷酸化ABI5及RAV1等转录因子, 激活下游ABA响应基因。ABAR/CHLH是叶绿体膜上的ABA受体, 其C-端和N-端暴露在细胞质中。低浓度ABA时, 叶绿体内伴侣蛋白CPN20结合ABAR, 抑制ABAR与负调控因子WRKY40的相互作用, 同时WRKY40与WRKY18和WRKY60通过直接或间接作用, 抑制<i>ABF4</i>/<i>ABI4</i>/<i>ABI5</i>的表达。高浓度ABA促使WRKY40向细胞质迁移, ABAR的C-端与WRKY40相互作用, 解除WRKY40对<i>ABF4</i>/<i>ABI4</i>/<i>ABI5</i>的转录抑制, 实现ABA生理效应。RAV1通过抑制<i>ABI3</i>/<i>ABI4</i>/<i>ABI5</i>转录来负调控ABA信号; WRKY6、WRKY41和NF-YC-RGL2分别通过下调RAV1的表达, 促进<i>ABI3</i>和<i>ABI5</i>的表达, 它们均正调控ABA信号。图中(+)和(-)分别表示该基因在ABA信号通路中具有正向和反向调控作用。<br/><b>Figure 1</b> Models of ABA signaling transduction pathway based on ABA receptors PYR/PYL/RCAR and ABAR/CHLH<br/>Intracellular ABA receptor PYR/PYL/RCAR exists as a homodimer in the resting state. Upon binding to ABA, PYR/PYL/RCAR turns into monomer and interacts with PP2Cs to release SnRK2s, which phosphorylates transcription factors like ABI5 and RAV1, to activate downstream ABA responsive genes. ABAR/CHLH is a protein across the chloroplast membrane, whose C- and N- ends are exposed to the cytoplasm. In the absence or low levels of ABA, the interaction of CPN20 with ABAR attenuates the interaction between ABAR and the negative regulator WRKY40, meanwhile WRKY40, WRKY18 and WRKY60 directly or indirectly inhibit the expression of <i>ABF4</i>/<i>ABI4</i>/<i>ABI5</i>. In response to a high level of ABA, WRKY40 is recruited from nucleus to the cytosol. C-terminal part of ABA-bound ABAR interacts with WRKY40 and relieves the suppression of <i>ABF4</i>/<i>ABI4</i>/<i>ABI5</i> by WRKY40, promoting the physiological effects of ABA. RAV1 negatively regulates the ABA signaling by inhibiting the transcription of <i>ABI3</i>/<i>ABI4</i>/<i>ABI5. </i>WRKY6, WRKY41, and NF-YC-RGL2 play the positive roles in the ABA signaling, by down-regulating the expression of <i>RAV1</i>, promoting the expression of <i>ABI3</i> and <i>ABI5</i>, respectively. (+) and (-) in this figure denote the positive regulation and the negative regulation in the ABA signaling pathway, respectively. Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-542/img_1.png<b>图1</b> 以PYR/PYL/RCAR和ABAR/CHLH为受体的ABA信号转导通路模型<br/>细胞质ABA受体PYR/PYL/RCAR在静息状态下以二聚体形式存在, 当与ABA结合后则以单体形式与PP2Cs结合, 解除对SnRK2s的抑制, 磷酸化ABI5及RAV1等转录因子, 激活下游ABA响应基因。ABAR/CHLH是叶绿体膜上的ABA受体, 其C-端和N-端暴露在细胞质中。低浓度ABA时, 叶绿体内伴侣蛋白CPN20结合ABAR, 抑制ABAR与负调控因子WRKY40的相互作用, 同时WRKY40与WRKY18和WRKY60通过直接或间接作用, 抑制<i>ABF4</i>/<i>ABI4</i>/<i>ABI5</i>的表达。高浓度ABA促使WRKY40向细胞质迁移, ABAR的C-端与WRKY40相互作用, 解除WRKY40对<i>ABF4</i>/<i>ABI4</i>/<i>ABI5</i>的转录抑制, 实现ABA生理效应。RAV1通过抑制<i>ABI3</i>/<i>ABI4</i>/<i>ABI5</i>转录来负调控ABA信号; WRKY6、WRKY41和NF-YC-RGL2分别通过下调RAV1的表达, 促进<i>ABI3</i>和<i>ABI5</i>的表达, 它们均正调控ABA信号。图中(+)和(-)分别表示该基因在ABA信号通路中具有正向和反向调控作用。<br/><b>Figure 1</b> Models of ABA signaling transduction pathway based on ABA receptors PYR/PYL/RCAR and ABAR/CHLH<br/>Intracellular ABA receptor PYR/PYL/RCAR exists as a homodimer in the resting state. Upon binding to ABA, PYR/PYL/RCAR turns into monomer and interacts with PP2Cs to release SnRK2s, which phosphorylates transcription factors like ABI5 and RAV1, to activate downstream ABA responsive genes. ABAR/CHLH is a protein across the chloroplast membrane, whose C- and N- ends are exposed to the cytoplasm. In the absence or low levels of ABA, the interaction of CPN20 with ABAR attenuates the interaction between ABAR and the negative regulator WRKY40, meanwhile WRKY40, WRKY18 and WRKY60 directly or indirectly inhibit the expression of <i>ABF4</i>/<i>ABI4</i>/<i>ABI5</i>. In response to a high level of ABA, WRKY40 is recruited from nucleus to the cytosol. C-terminal part of ABA-bound ABAR interacts with WRKY40 and relieves the suppression of <i>ABF4</i>/<i>ABI4</i>/<i>ABI5</i> by WRKY40, promoting the physiological effects of ABA. RAV1 negatively regulates the ABA signaling by inhibiting the transcription of <i>ABI3</i>/<i>ABI4</i>/<i>ABI5. </i>WRKY6, WRKY41, and NF-YC-RGL2 play the positive roles in the ABA signaling, by down-regulating the expression of <i>RAV1</i>, promoting the expression of <i>ABI3</i> and <i>ABI5</i>, respectively. (+) and (-) in this figure denote the positive regulation and the negative regulation in the ABA signaling pathway, respectively.
图1 以PYR/PYL/RCAR和ABAR/CHLH为受体的ABA信号转导通路模型 细胞质ABA受体PYR/PYL/RCAR在静息状态下以二聚体形式存在, 当与ABA结合后则以单体形式与PP2Cs结合, 解除对SnRK2s的抑制, 磷酸化ABI5及RAV1等转录因子, 激活下游ABA响应基因。ABAR/CHLH是叶绿体膜上的ABA受体, 其C-端和N-端暴露在细胞质中。低浓度ABA时, 叶绿体内伴侣蛋白CPN20结合ABAR, 抑制ABAR与负调控因子WRKY40的相互作用, 同时WRKY40与WRKY18和WRKY60通过直接或间接作用, 抑制ABF4/ABI4/ABI5的表达。高浓度ABA促使WRKY40向细胞质迁移, ABAR的C-端与WRKY40相互作用, 解除WRKY40对ABF4/ABI4/ABI5的转录抑制, 实现ABA生理效应。RAV1通过抑制ABI3/ABI4/ABI5转录来负调控ABA信号; WRKY6、WRKY41和NF-YC-RGL2分别通过下调RAV1的表达, 促进ABI3和ABI5的表达, 它们均正调控ABA信号。图中(+)和(-)分别表示该基因在ABA信号通路中具有正向和反向调控作用。 Figure 1 Models of ABA signaling transduction pathway based on ABA receptors PYR/PYL/RCAR and ABAR/CHLH Intracellular ABA receptor PYR/PYL/RCAR exists as a homodimer in the resting state. Upon binding to ABA, PYR/PYL/RCAR turns into monomer and interacts with PP2Cs to release SnRK2s, which phosphorylates transcription factors like ABI5 and RAV1, to activate downstream ABA responsive genes. ABAR/CHLH is a protein across the chloroplast membrane, whose C- and N- ends are exposed to the cytoplasm. In the absence or low levels of ABA, the interaction of CPN20 with ABAR attenuates the interaction between ABAR and the negative regulator WRKY40, meanwhile WRKY40, WRKY18 and WRKY60 directly or indirectly inhibit the expression of ABF4/ABI4/ABI5. In response to a high level of ABA, WRKY40 is recruited from nucleus to the cytosol. C-terminal part of ABA-bound ABAR interacts with WRKY40 and relieves the suppression of ABF4/ABI4/ABI5 by WRKY40, promoting the physiological effects of ABA. RAV1 negatively regulates the ABA signaling by inhibiting the transcription of ABI3/ABI4/ABI5. WRKY6, WRKY41, and NF-YC-RGL2 play the positive roles in the ABA signaling, by down-regulating the expression of RAV1, promoting the expression of ABI3 and ABI5, respectively. (+) and (-) in this figure denote the positive regulation and the negative regulation in the ABA signaling pathway, respectively.
拟南芥pyr1/pyl1/pyl2/prl4四突变体种子萌发表现出对ABA高度不敏感(Park et al., 2009)。PP2C是一类丝氨酸/苏氨酸残基蛋白磷酸酶, 包括ABI1 (ABA INSENSITIVE 1)、ABI2、HAB1 (HOMOLOGY TO ABA INSENSITIVE 1)、HAB2和AHG3/PP2CA (ABA-HYPERSENSITIVE GERMINATION 3), 在ABA信号转导中起着关键的负调控作用(Hauser et al., 2011)。ABI1和ABI2的显性负突变体abi1-1和abi2-1由于蛋白质结构的改变而不能传递ABA信号, 呈现出种子休眠水平下降和对外源ABA不敏感表型(Ma et al., 2009; Park et al., 2009)。hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004)。AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006)。SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014)。其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程。双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007)。三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009)。RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014)。 清华大学张大鹏研究组发现, 定位于叶绿体被膜上的蛋白质镁螯合酶H亚基(Mg2+-chelatase H subunit, CHLH)能够结合ABA, 他们提出1条CHLH/ ABAR (ABA receptor)介导的始于叶绿体的解除转录抑制的ABA信号通路。该模型认为在生理状态或低浓度ABA时, WRKY40抑制一组ABA信号正调节基因(ABF4、ABI4和ABI5)的表达; 高浓度ABA时, ABAR结合ABA后, 刺激WRKY40从细胞核转移到细胞质, 促进ABAR与WRKY40的相互作用, 进而激发一种未知因子(或信号系统), 阻遏WRKY40的表达, 从而解除WRKY40对ABA响应基因转录的抑制, 最终实现ABA的生理效应(Shen et al., 2006; Shang et al., 2010; 张大鹏, 2011)。 研究表明, 拟南芥ABAR/CHLH过表达株系表现出对ABA的高度敏感, 而突变体abar-2和ABAR RN- Ai株系的种子萌发对ABA不敏感。上述结果与ABAR作为ABA受体的结论相符。WRKY40是细胞内关键的ABA信号负调控因子, 在细胞核中通过抑制ABI5的表达而抑制下游的ABA应答反应。WRKY40缺失突变体种子萌发表型对ABA高度敏感(Jiang and Yu, 2009; Shang et al., 2010)。拟南芥种子萌发及之后早期幼苗生长发育阶段, ABA诱导WRKY2的积累需要ABI3、ABI5、ABA2和ABA3的存在。wrky2缺失突变体种子萌发及幼苗生长对ABA超敏感, 说明WRKY2也负调控ABA抑制的种子萌发和幼苗早期生长发育(Jiang and Yu, 2009)。另外, 早期研究表明, 野燕麦(Avena fatua) WRKY蛋白ABF1和ABF2能结合在α-淀粉酶编码基因启动子的box2/W-box元件上, 促进胚乳中淀粉的水解, 从而间接促进种子萌发(Rush- ton et al., 1995)。 在拟南芥WRKY转录因子中, WRKY6、WRKY- 18、WRKY41、WRKY60和WRKY63被认为是ABA信号的正调控因子, 提示WRKY转录因子家族在ABA信号通路中扮演着多重角色。Huang等(2016)发现WRKY6通过直接抑制RAV1表达而促进ABI3、ABI4和ABI5的表达, 调节ABA抑制的种子萌发过程。Ding等(2014)发现, WRKY41通过结合在ABI3基因启动子上而直接促进ABI3的转录; wrky41缺失突变体的种子萌发和幼苗生长对ABA的敏感性降低, 而ABI3过量表达能够恢复wrky41种子对ABA的敏感性。Chen等(2010)发现, WRKY18和WRKY60缺失突变体种子萌发对ABA敏感性降低, 而过量表达株系对ABA高度敏感, 提出WRKY18和WRKY60正调控种子萌发过程中的ABA信号。此外, 由于WRKY18和WRKY40能识别并结合WRKY60启动子的W-box元件, WRKY60可能是WRKY18和WRKY40的直接靶标; 另一方面, WRKY40与WRKY18和WRKY60又可通过复杂的相互作用, 结合ABI4和ABI5的启动子, 抑制它们的表达, 从而负调控ABA信号转导(Liu et al., 2012)。研究表明, WRKY63的T-DNA插入突变体种子萌发对ABA超敏感, WRKY63位于ABI5下游, 参与ABA抑制种子萌发的信号转导。种子萌发时, ABAR/CHLH与ABA的结合促使WRKY40离开细胞核, ABI5恢复转录活性; 另一方面, PYR/PYL/RCAR与ABA结合使下游ABI5蛋白磷酸化, 继而促进WRKY63表达, 最后激活其下游基因的表达(Ren et al., 2010)。 近年来, 一些NF-Y转录因子也被发现通过影响ABA信号调节种子萌发。酵母双杂交分析表明, NF- YB2能与ABF3 (ABA RESPONSE ELEMENT BIND- ING PROTEINS 3/ABA BINDING FACTORS 3)结合, NF-YC9能与ABF2结合; NF-YB2和NF-YB3过量表达时表现出对ABA敏感性的显著改变, 但缺失突变体无相关表型, 提示NF-YB2和NF-YB3并不是影响种子萌发的关键转录因子。三突变体nf-yc3/nf-yc4/ nf-yc9种子萌发对ABA敏感性增强, 而nf-yc4单突变体种子萌发对ABA敏感性下降, 提示NF-YC3、NF-YC4和NF-YC9基因之间存在功能冗余(Kumimo- to et al., 2013)。进一步研究表明, NF-YC (NF-YC3、NF-YC4和NF-YC9)能与DELLA蛋白RGL2 (GA信号的关键负调控蛋白)构成复合物NF-YC-RGL2, 通过识别ABI5启动子的CCAAT元件, 促进ABI5表达从而激活下游的ABA和GA介导的种子萌发过程(Liu et al., 2016)。此外, 研究人员还发现位于叶绿体内的ABA- R/CHLH互作蛋白——伴侣蛋白20 (co-chaperonin 20, CPN20)负调控ABA信号转导通路。cpn20-1突变体中WRKY40表达下调, 且CPN20通过其N-Signa- ture结构域与WRKY40竞争性结合ABAR, 从而减少ABAR-WRKY40互作, 抑制ABA下游信号。相反, ABA又通过直接抑制ABAR-CPN20的互作降低CPN- 20的表达, 同时促进ABAR-WRKY40互作来促进信号传递(图1) (Zhang et al., 2013, 2014)。
AlonsoR, Oñate-SánchezL, WeltmeierF, EhlertA, DiazI, DietrichK, Vicente-CarbajosaJ, Dröge-LaserW (2009). A pivotal role of the basic leucine zipper transcription factor bZIP53 in the regulation of Arabidopsis seed maturation gene expression based on heterodimerization and protein complex formation.Plant Cell 21, 1747-1761. [本文引用: 1]
[6]
BarreroJM, MillarAA, GriffithsJ, CzechowskiT, ScheibleWR, UdvardiM, ReidJB, RossJJ, JacobsenJV, GublerF (2010). Gene expression profiling identifies two re- gulatory genes controlling dormancy and ABA sensitivity in Arabidopsis seeds.Plant J 61, 611-622. [本文引用: 1]
[7]
BelmonteMF, KirkbrideRC, StoneSL, PelletierJM, BuiAQ, YeungEC, HashimotoM, FeiJ, HaradaCM, MunozMD, LeBH, DrewsGN, BradySM, GoldbergRB, HaradaJJ (2013). Comprehensive developmental profiles of gene activity in regions and subregions of the Arabidopsis seed.Proc Natl Acad Sci USA 110, E435-E444.
[8]
BienzM (2006). The PHD finger, a nuclear protein-interac- tion domain.Trends Biochem Sci 31, 35-40. [本文引用: 1]
[9]
BoccacciniA, LorraiR, RutaV, FreyA, Mercey-BoutetS, Marion-PollA, TarkowskáD, StrnadM, CostantinoP, VittoriosoP (2016). The DAG1 transcription factor negatively regulates the seed-to-seedling transition in Arabidopsis acting on ABA and GA levels.BMC Plant Biol 16, 198.
[10]
BoccacciniA, SantopoloS, CapautoD, LorraiR, MinutelloE, BelcramK, PalauquiJC, CostantinoP, VittoriosoP (2014). Independent and interactive effects of DOF affecting germination 1 (DAG1) and the Della proteins GA insensitive (GAI) and repressor of ga1-3 (RGA) in embryo development and seed germination. BMC Plant Biol 14, 200. [本文引用: 2]
[11]
BraybrookSA, StoneSL, ParkS, BuiAQ, LeBH, FischerRL, GoldbergRB, HaradaJJ (2006). Genes directly regulated by LEAFY COTYLEDON 2 provide insight into the control of embryo maturation and somatic embryogenesis.Proc Natl Acad Sci USA 103, 3468-3473. [本文引用: 1]
[12]
CadmanCSC, TooropPE, HilhorstHWM, Finch-SavageWE (2006). Gene expression profiles of Arabidopsis Cvi seeds during dormancy cycling indicate a common underlying dormancy control mechanism.Plant J 46, 805-822. [本文引用: 1]
[13]
CarlesC, Bies-EtheveN, AspartL, Léon-KloosterzielKM, KoornneefM, EcheverriaM, DelsenyM (2002). Regulation of Arabidopsis thaliana Em genes: role of ABI5. Plant J 30, 373-383. [本文引用: 1]
[14]
ChenH, LaiZB, ShiJW, XiaoY, ChenZX, XuXP (2010). Roles of Arabidopsis WRKY18, WRKY40 and WRKY60 transcription factors in plant responses to abscisic acid and abiotic stress.BMC Plant Biol 10, 281. [本文引用: 1]
[15]
ChenHY, HsiehEJ, ChengMC, ChenCY, HwangSY, LinTP (2016). ORA47 (octadecanoid-responsive AP2/ERF- domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through bin- ding to a novel cis-element. New Phytol 211, 599-613. [本文引用: 1]
[16]
DekkersBJW, HeHZ, HansonJ, WillemsLAJ, JamarDCL, CueffG, RajjouL, HilhorstHWM, BentsinkL (2016). The Arabidopsis DELAY OF GERMINATION 1 gene affects ABSCISIC ACID INSENSITIVE 5 (ABI5) expression and genetically interacts with ABI3 during Arabidopsis seed development. Plant J 85, 451-465. [本文引用: 3]
[17]
DingZJ, YanJY, LiGX, WuZC, ZhangSQ, ZhengSJ (2014). WRKY41 controls Arabidopsis seed dormancy via direct regulation of ABI3 transcript levels not downstream of ABA. Plant J 79, 810-823. [本文引用: 2]
[18]
FengCZ, ChenY, WangC, KongYH, WuWH, ChenYF (2014). Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development. Plant J 80, 654-668. [本文引用: 2]
[19]
Finch-SavageWE, Leubner-MetzgerG (2006). Seed dormancy and the control of germination.New Phytol 171, 501-523. [本文引用: 1]
[20]
FinkelsteinRR, GampalaSSL, RockCD (2002). Abscisic acid signaling in seeds and seedlings.Plant Cell 14, S15-S45. [本文引用: 2]
FootittS, MüllerK, KermodeAR, Finch-SavageWE (2015). Seed dormancy cycling in Arabidopsis: chromatin remodelling and regulation of DOG1 in response to seasonal environmental signals.Plant J 81, 413-425. [本文引用: 1]
[23]
FreyA, EffroyD, LefebvreV, SeoM, PerreauF, BergerA, SechetJ, ToA, NorthHM, Marion-PollA (2012). Epoxycarotenoid cleavage by NCED5 fine-tunes ABA accumulation and affects seed dormancy and drought tolerance with other NCED family members.Plant J 70, 501-512. [本文引用: 1]
[24]
FujiiH, VersluesPE, ZhuJK (2007). Identification of two protein kinases required for abscisic acid regulation of seed germination, root growth, and gene expression in Arabidopsis.Plant Cell 19, 485-494. [本文引用: 1]
[25]
FujiiH, ZhuJK (2009). Arabidopsis mutant deficient in 3 abscisic acid-activated protein kinases reveals critical roles in growth, reproduction, and stress.Proc Natl Acad Sci USA 106, 8380-8385. [本文引用: 1]
HuangMK, HuYL, LiuX, LiYG, HouXL (2015). Arabidopsis LEAFY COTYLEDON 1 mediates postembryonic development via interacting with PHYTOCHROME-INTER- ACTING FACTOR 4.Plant Cell 27, 3099-3111. [本文引用: 1]
[30]
HuangY, FengCZ, YeQ, WuWH, ChenYF (2016). Arabidopsis WRKY6 transcription factor acts as a positive regulator of abscisic acid signaling during seed germination and early seedling development.PLoS Genet 12, e1005833. [本文引用: 1]
[31]
HubbardKE, NishimuraN, HitomiK, GetzoffED, Sch- roederJI (2010). Early abscisic acid signal transduction mechanisms: newly discovered components and newly emerging questions.Genes Dev 24, 1695-1708. [本文引用: 1]
[32]
JeJ, ChenH, SongC, LimCO (2014). Arabidopsis DREB2C modulates ABA biosynthesis during germination.Biochem Biophys Res Commun 452, 91-98. [本文引用: 1]
[33]
JiaHY, SuzukiM, McCartyDR (2014). Regulation of the seed to seedling developmental phase transition by the LAFL and VAL transcription factor networks.Wiley Interdiscip Rev Dev Biol 3, 135-145. [本文引用: 1]
[34]
JiangWB, YuDQ (2009). Arabidopsis WRKY2 transcription factor mediates seed germination and postgermination arrest of development by abscisic acid.BMC Plant Biol 9, 96. [本文引用: 2]
[35]
JunkerA, MönkeG, RuttenT, KeilwagenJ, SeifertM, ThiTM, RenouJP, BalzergueS, ViehöverP, HähnelU, Ludwig-MüllerJ, AltschmiedL, ConradU, WeisshaarB, BäumleinH (2012). Elongation-related functions of LEAFY COTYLEDON 1 during the development of Arabidopsis thaliana. Plant J 71, 427-442. [本文引用: 1]
[36]
KagayaY, ToyoshimaR, OkudaR, UsuiH, YamamotoA, HattoriT (2005). LEAFY COTYLEDON 1 controls seed storage protein genes through its regulation of FUSCA3 and ABSCISIC ACID INSENSITIVE 3. Plant Cell Physiol 46, 399-406. [本文引用: 2]
[37]
KanaiM, NishimuraM, HayashiM (2010). A peroxisomal ABC transporter promotes seed germination by inducing pectin degradation under the control of ABI5. Plant J 62, 936-947. [本文引用: 1]
[38]
KannoY, JikumaruY, HanadaA, NambaraE, AbramsSR, KamiyaY, SeoM (2010). Comprehensive hormone profiling in developing Arabidopsis seeds: examination of the site of ABA biosynthesis, ABA transport and hormone interactions.Plant Cell Physiol 51, 1988-2001. [本文引用: 1]
[39]
KarssenCM, Brinkhorst-van der SwanDLC, BreeklandAE, KoornneefM (1983). Induction of dormancy during seed development by endogenous abscisic acid: studies on abscisic acid deficient genotypes of Arabidopsis thaliana (L.) Heynh. Planta 157, 158-165. [本文引用: 1]
[40]
KondhareKR, HeddenP, KettlewellPS, FarrellAD, MonaghanJM (2014). Quantifying the impact of exogenous abscisic acid and gibberellins on pre-maturity α- amylase formation in developing wheat grains.Sci Rep 4, 5355. [本文引用: 1]
[41]
KoornneefM, KarssenCM (1994). Seed dormancy and germination. In: Meyerowitz EM, Somerville CR, eds. Arabidopsis. New York: Cold Spring Harbor Laboratory Press. pp. 313-334. [本文引用: 1]
[42]
KumimotoRW, SiriwardanaCL, GaylerKK, RisingerJR, SiefersN, Holt BF3rd (2013). NUCLEAR FACTOR Y transcription factors have both opposing and additive roles in ABA-mediated seed germination.PLoS One 8, e59481. [本文引用: 1]
[43]
KushiroT, OkamotoM, NakabayashiK, YamagishiK, KitamuraS, AsamiT, HiraiN, KoshibaT, KamiyaY, NambaraE (2004). The Arabidopsis cytochrome P450 CYP707A encodes ABA 8'-hydroxylases: key enzymes in ABA catabolism.EMBO J 23, 1647-1656. [本文引用: 2]
[44]
KwongRW, BuiAQ, LeeH, KwongLW, FischerRL, GoldbergRB, HaradaJJ (2003). LEAFY COTYLEDON1-LIKE defines a class of regulators essential for embryo development.Plant Cell 15, 5-18. [本文引用: 1]
[45]
LaraP, Oñate-SánchezL, AbrahamZ, FerrándizC, DíazI, CarboneroP, Vicente-CarbajosaJ (2003). Synergistic activation of seed storage protein gene expression in Arabidopsis by ABI3 and two bZIPs related to OPAQUE2.J Biol Chem 278, 21003-21011. [本文引用: 1]
[46]
LeeHG, LeeK, SeoPJ (2015a). The Arabidopsis MYB96 transcription factor plays a role in seed dormancy.Plant Mol Biol 87, 371-381. [本文引用: 1]
[47]
LeeK, LeeHG, YoonS, KimHU, SeoPJ (2015b). The Arabidopsis MYB96 transcription factor is a positive regulator of ABSCISIC ACID-INSENSITIVE 4 in the control of seed germination. Plant Physiol 168, 677-689. [本文引用: 1]
[48]
LefebvreV, NorthH, FreyA, SottaB, SeoM, OkamotoM, NambaraE, Marion-PollA (2006). Functional analysis of Arabidopsis NCED6 and NCED9 genes indicates that ABA synthesized in the endosperm is involved in the induction of seed dormancy. Plant J 45, 309-319. [本文引用: 2]
[49]
LinPC, HwangSG, EndoA, OkamotoM, KoshibaT, ChengWH (2007). Ectopic expression of ABSCISIC ACID 2/GLUCOSE INSENSITIVE 1 in Arabidopsis promotes seed dormancy and stress tolerance. Plant Physiol 143, 745-758. [本文引用: 2]
[50]
LiuX, HuP, HuangM, TangY, LiY, LiL, HouX (2016). The NF-YC-RGL2 module integrates GA and ABA signalling to regulate seed germination in Arabidopsis.Nat Com- mun 7, 12768. [本文引用: 1]
[51]
LiuYX, GeyerR, van ZantenM, CarlesA, LiY, HöroldA, van NockerS, SoppeWJJ (2011). Identification of the Arabidopsis REDUCED DORMANCY 2 gene uncovers a role for the polymerase associated factor 1 complex in seed dormancy. PLoS One 6, e22241. [本文引用: 1]
[52]
LiuZQ, YanL, WuZ, MeiC, LuK, YuYT, LiangS, ZhangXF, WangXF, ZhangDP (2012). Cooperation of three WRKY-domain transcription factors WRKY18, WRKY40, and WRKY60 in repressing two ABA-responsive genes ABI4 and ABI5 in Arabidopsis. J Exp Bot 63, 6371-6392. [本文引用: 1]
[53]
Lopez-MolinaL, MongrandS, McLachlinDT, ChaitBT, ChuaNH (2002). ABI5 acts downstream of ABI3 to execute an ABA-dependent growth arrest during germination.Plant J 32, 317-328. [本文引用: 1]
[54]
LotanT, OhtoMA, YeeKM, WestMA, LoR, KwongRW, YamagishiK, FischerRL, GoldbergRB, HaradaJJ (1998). Arabidopsis LEAFY COTYLEDON 1 is sufficient to induce embryo development in vegetative cells.Cell 93, 1195-1205. [本文引用: 1]
[55]
MaY, SzostkiewiczI, KorteA, MoesD, YangY, ChristmannA, GrillE (2009). Regulators of PP2C phosphatase activity function as abscisic acid sensors.Science 324, 1064-1068. [本文引用: 2]
[56]
MidhatU, TingMKY, TeresinskiHJ, SneddenWA (2018). The calmodulin-like protein, CML39, is involved in regulating seed development, germination, and fruit development in Arabidopsis.Plant Mol Biol 96, 375-392. [本文引用: 1]
[57]
MönkeG, SeifertM, KeilwagenJ, MohrM, GrosseI, HähnelU, JunkerA, WeisshaarB, ConradU, BäumleinH, AltschmiedL (2012). Toward the identification and regulation of the Arabidopsis thaliana ABI3 regulon. Nucleic Acids Res 40, 8240-8254. [本文引用: 1]
[58]
MüllerK, CarstensAC, LinkiesA, TorresMA, Leubner-MetzgerG (2009). The NADPH-oxidase AtrbohB plays a role in Arabidopsis seed after-ripening. New Phytol 184, 885-897. [本文引用: 1]
[59]
NakamuraS, LynchTJ, FinkelsteinRR (2001). Physical interactions between ABA response loci of Arabidopsis.Plant J 26, 627-635. [本文引用: 2]
[60]
NakashimaK, FujitaY, KanamoriN, KatagiriT, UmezawaT, KidokoroS, MaruyamaK, YoshidaT, IshiyamaK, KobayashiM, ShinozakiK, Yamaguchi-ShinozakiK (2009). Three Arabidopsis SnRK2 protein kinases, SRK- 2D/SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK- 2.3, involved in ABA signaling are essential for the control of seed development and dormancy.Plant Cell Physiol 50, 1345-1363. [本文引用: 1]
[61]
NéeG, KramerK, NakabayashiK, YuanBJ, XiangY, MiattonE, FinkemeierI, SoppeWJJ (2017). DELAY OF GERMINATION 1 requires PP2C phosphatases of the ABA signaling pathway to control seed dormancy.Nat Commun 8, 72. [本文引用: 1]
[62]
NonogakiM, SallK, NambaraE, NonogakiH (2014). Amplification of ABA biosynthesis and signaling through a positive feedback mechanism in seeds.Plant J 78, 527-539. [本文引用: 1]
[63]
OgawaM, HanadaA, YamauchiY, KuwaharaA, KamiyaY, YamaguchiS (2003). Gibberellin biosynthesis and response during Arabidopsis seed germination.Plant Cell 15, 1591-1604. [本文引用: 1]
[64]
OhE, YamaguchiS, HuJH, YusukeJ, JungB, PaikI, LeeHS, SunTP, KamiyaY, ChoiG (2007). PIL5, a phytochrome-interacting bHLH protein, regulates gibberellin responsiveness by binding directly to the GAI and RGA promoters in Arabidopsis seeds. Plant Cell 19, 1192-1208. [本文引用: 1]
[65]
OkamotoM, KuwaharaA, SeoM, KushiroT, AsamiT, HiraiN, KamiyaY, KoshibaT, NambaraE (2006). CYP707A1 and CYP707A2, which encode abscisic acid 8′-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis.Plant Phy- siol 141, 97-107. [本文引用: 3]
[66]
PapaefthimiouD, LikotrafitiE, KapazoglouA, BladenopoulosK, TsaftarisA (2010). Epigenetic chromatin modifiers in barley: III. Isolation and characterization of the barley GNAT-MYST family of histone acetyltransferases and responses to exogenous ABA.Plant Physiol Biochem 48, 98-107. [本文引用: 1]
[67]
ParkSY, FungP, NishimuraN, JensenDR, FujiiH, ZhaoY, LumbaS, SantiagoJ, RodriguesA, ChowTFF, AlfredSE, BonettaD, FinkelsteinR, ProvartNJ, DesveauxD, RodriguezPL, McCourtP, ZhuJK, SchroederJI, VolkmanBF, CutlerSR (2009). Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins.Science 324, 1068-1071. [本文引用: 3]
[68]
PerryJ, ZhaoYD (2003). The CW domain, a structural module shared amongst vertebrates, vertebrate-infecting parasites and higher plants.Trends Biochem Sci 28, 576-580. [本文引用: 1]
[69]
PiskurewiczU, JikumaruY, KinoshitaN, NambaraE, KamiyaY, Lopez-MolinaL (2008). The gibberellic acid signaling repressor RGL2 inhibits Arabidopsis seed germination by stimulating abscisic acid synthesis and ABI5 activity.Plant Cell 20, 2729-2745. [本文引用: 1]
[70]
PrestonJ, TatematsuK, KannoY, HoboT, KimuraM, JikumaruY, YanoR, KamiyaY, NambaraE (2009). Temporal expression patterns of hormone metabolism genes during imbibition of Arabidopsis thaliana seeds: a comparative study on dormant and non-dormant accessions. Plant Cell Physiol 50, 1786-1800. [本文引用: 1]
[71]
RavindranP, VermaV, StammP, KumarPP (2017). A novel RGL2-DOF6 complex contributes to primary seed dormancy in Arabidopsis thaliana by regulating a GATA transcription factor. Mol Plant 10, 1307-1320. [本文引用: 1]
[72]
RazV, BergervoetJH, KoornneefM (2001). Sequential steps for developmental arrest in Arabidopsis seeds.Development 128, 243-252. [本文引用: 1]
[73]
RenXZ, ChenZZ, LiuY, ZhangHR, ZhangM, LiuQ, HongXH, ZhuJK, GongZZ (2010). ABO3, a WRKY transcription factor, mediates plant responses to abscisic acid and drought tolerance in Arabidopsis.Plant J 63, 417-429. [本文引用: 1]
[74]
RoscoeTT, GuilleminotJ, BessouleJJ, BergerF, DevicM (2015). Complementation of seed maturation phenotypes by ectopic expression of ABSCISIC ACID INSENSITIVE 3, FUSCA 3 and LEAFY COTYLEDON 2 in Arabidopsis. Plant Cell Physiol 56, 1215-1228. [本文引用: 3]
[75]
RushtonPJ, MacdonaldH, HuttlyAK, LazarusCM, HooleyR (1995). Members of a new family of DNA-binding proteins bind to a conserved cis-element in the promoters of α-Amy2 genes. Plant Mol Biol 29, 691-702. [本文引用: 1]
[76]
SaezA, ApostolovaN, Gonzalez-GuzmanM, Gonzalez- GarciaMP, NicolasC, LorenzoO, RodriguezPL (2004). Gain-of-function and loss-of-function phenotypes of the protein phosphatase 2C HAB1 reveal its role as a negative regulator of abscisic acid signaling.Plant J 37, 354-369. [本文引用: 1]
[77]
SantopoloS, BoccacciniA, LorraiR, RutaV, CapautoD, MinutelloE, SerinoG, CostantinoP, VittoriosoP (2015). DOF AFFECTING GERMINATION 2 is a positive regulator of light-mediated seed germination and is repressed by DOF AFFECTING GERMINATION 1.BMC Plant Biol 15, 72. [本文引用: 1]
[78]
ShangY, YanL, LiuZQ, CaoZ, MeiC, XinQ, WuFQ, WangXF, DuSY, JiangT, ZhangXF, ZhaoR, SunHL, LiuR, YuYT, ZhangDP (2010). The Mg-chelatase H subunit of Arabidopsis antagonizes a group of WRKY transcription repressors to relieve ABA-responsive genes of inhibition.Plant Cell 22, 1909-1935. [本文引用: 2]
[79]
ShenYY, WangXF, WuFQ, DuSY, CaoZ, ShangY, WangXL, PengCC, YuXC, ZhuSY, FanRC, XuYH, ZhangDP (2006). The Mg-chelatase H subunit is an abscisic acid receptor.Nature 443, 823-826. [本文引用: 1]
[80]
ShuK, LiuXD, XieQ, HeZH (2016). Two faces of one seed: hormonal regulation of dormancy and germination.Mol Plant 9, 34-45. [本文引用: 3]
[81]
ShuK, ZhangHW, WangSF, ChenML, WuYR, TangSY, LiuCY, FengYQ, CaoXF, XieQ (2013). ABI4 regulates primary seed dormancy by regulating the biogenesis of abscisic acid and gibberellins in Arabidopsis.PLoS Genet 9, e1003577. [本文引用: 2]
SuzukiM, WangHHY, McCartyDR (2007). Repression of the LEAFY COTYLEDON 1/B3 regulatory network in plant embryo development by VP1/ABSCISIC ACID INSENSITIVE 3-LIKE B3 genes. Plant Physiol 143, 902-911. [本文引用: 1]
[85]
TiedemannJ, RuttenT, MönkeG, VorwiegerA, RolletschekH, MeissnerD, MilkowskiC, PetereckS, MockHP, ZankT, BäumleinH (2008). Dissection of a complex seed phenotype: novel insights of FUSCA3 regulated developmental processes. Dev Biol 317, 1-12. [本文引用: 1]
[86]
ToA, ValonC, SavinoG, GuilleminotJ, DevicM, GiraudatJ, ParcyF (2006). A network of local and redundant gene regulation governs Arabidopsis seed maturation.Plant Cell 18, 1642-1651. [本文引用: 2]
[87]
VaistijFE, GanYB, PenfieldS, GildayAD, DaveA, HeZS, JosseEM, ChoiG, HallidayKJ, GrahamIA (2013). Differential control of seed primary dormancy in Arabidopsis ecotypes by the transcription factor SPATULA.Proc Natl Acad Sci USA 110, 10866-10871. [本文引用: 1]
[88]
VirdiAS, SinghS, SinghP (2015). Abiotic stress responses in plants: roles of calmodulin-regulated proteins.Front Plant Sci 6, 809. [本文引用: 1]
[89]
WangFF, PerrySE (2013). Identification of direct targets of FUSCA3, a key regulator of Arabidopsis seed development.Plant Physiol 161, 1251-1264. [本文引用: 2]
[90]
WeitbrechtK, MüllerK, Leubner-MetzgerG (2011). First off the mark: early seed germination.J Exp Bot 62, 3289-3309. [本文引用: 2]
[91]
YamamotoA, KagayaY, UsuiH, HoboT, TakedaS, HattoriT (2010). Diverse roles and mechanisms of gene regu- lation by the Arabidopsis seed maturation master regulator FUS3 revealed by microarray analysis.Plant Cell Physiol 51, 2031-2046. [本文引用: 2]
[92]
YanoR, KannoY, JikumaruY, NakabayashiK, KamiyaY, NambaraE (2009). CHOTTO1, a putative double APETA- LA2 repeat transcription factor, is involved in abscisic acid- mediated repression of gibberellin biosynthesis during seed germination in Arabidopsis.Plant Physiol 151, 641-654. [本文引用: 2]
[93]
YoshidaT, NishimuraN, KitahataN, KuromoriT, ItoT, AsamiT, ShinozakiK, HirayamaT (2006). ABA-hyper- sensitive germination 3 encodes a protein phosphatase 2C (AtPP2CA) that strongly regulates abscisic acid signaling during germination among Arabidopsis protein phospha- tase 2Cs. Plant Physiol 140, 115-126. [本文引用: 1]
[94]
ZhangDP (2014). Abscisic Acid: Metabolism, Transport and Signaling. Dordrecht: Springer. pp. 33-35. [本文引用: 2]
ZhangXF, JiangT, YuYT, WuZ, JiangSC, LuK, FengXJ, LiangS, LuYF, WangXF, ZhangDP (2014). Arabidopsis co-chaperonin CPN20 antagonizes Mg-chelatase H subunit to derepress ABA-responsive WRKY40 transcription repressor.Sci China Life Sci 57, 11-21.
[97]
ZhaoML, YangSG, LiuXC, WuKQ (2015). Arabidopsis histone demethylases LDL1 and LDL2 control primary seed dormancy by regulating DELAY OF GERMINATION 1 and ABA signaling-related genes.Front Plant Sci 6, 159. [本文引用: 1]
[98]
ZhengJ, ChenFY, WangZ, CaoH, LiXY, DengX, SoppeWJJ, LiY, LiuYX (2012). A novel role for histone methyltransferase KYP/SUVH4 in the control of Arabidopsis primary seed dormancy.New Phytol 193, 605-616. [本文引用: 2]
脱落酸调控种子萌发和休眠的研究进展 1 2017
... 种子休眠和萌发是植物生长发育过程中2个紧密相连的阶段, 受到种子发育的内源因素和外界环境因素共同调控.Finch-Savage和Leubner-Metzger (2006)对种子休眠和萌发进行了定义: 休眠是指具有活力的种子在适宜的环境条件下不能萌发的状态; 而萌发则是种子内胚根突破外周的组织结构(胚乳和种皮)而展露在种皮之外的过程.脱落酸(abscisic acid, ABA)是调控种子休眠和萌发过程最重要的植物激素, 具有诱导和维持种子休眠、抑制种子萌发及幼苗生长发育的作用.十几年来, ABA调控种子休眠和萌发的研究取得了重大进展(Holdsworth et al., 2008; Weitbrecht et al., 2011; Shu et al., 2016).刘一灵等(2014)从ABA代谢与种子发育、穗萌和种子萌发等方面进行了综述.操春燕等(2017)则从ABA与细胞分裂素(cytokinin, CK)和赤霉素(gibberellin, GA)等相互作用调控种子萌发的角度概述了相关的最新研究进展.近年来, 随着参与种子ABA代谢和ABA信号基因相关突变体研究的深入, ABA调控种子休眠和萌发的分子机制已逐渐明晰, 本文参考ABA代谢基因、种子成熟基因以及ABA信号基因的遗传功能和表达调控的最新研究成果(表1), 综述ABA调控种子休眠和萌发的分子机制, 以期为植物激素的作用机理研究和农作物分子设计育种提供重要的理论依据. ...
植物激素ABA调控种子发育与萌发的研究进展 1 2014
... 种子休眠和萌发是植物生长发育过程中2个紧密相连的阶段, 受到种子发育的内源因素和外界环境因素共同调控.Finch-Savage和Leubner-Metzger (2006)对种子休眠和萌发进行了定义: 休眠是指具有活力的种子在适宜的环境条件下不能萌发的状态; 而萌发则是种子内胚根突破外周的组织结构(胚乳和种皮)而展露在种皮之外的过程.脱落酸(abscisic acid, ABA)是调控种子休眠和萌发过程最重要的植物激素, 具有诱导和维持种子休眠、抑制种子萌发及幼苗生长发育的作用.十几年来, ABA调控种子休眠和萌发的研究取得了重大进展(Holdsworth et al., 2008; Weitbrecht et al., 2011; Shu et al., 2016).刘一灵等(2014)从ABA代谢与种子发育、穗萌和种子萌发等方面进行了综述.操春燕等(2017)则从ABA与细胞分裂素(cytokinin, CK)和赤霉素(gibberellin, GA)等相互作用调控种子萌发的角度概述了相关的最新研究进展.近年来, 随着参与种子ABA代谢和ABA信号基因相关突变体研究的深入, ABA调控种子休眠和萌发的分子机制已逐渐明晰, 本文参考ABA代谢基因、种子成熟基因以及ABA信号基因的遗传功能和表达调控的最新研究成果(表1), 综述ABA调控种子休眠和萌发的分子机制, 以期为植物激素的作用机理研究和农作物分子设计育种提供重要的理论依据. ...
... 拟南芥pyr1/pyl1/pyl2/prl4四突变体种子萌发表现出对ABA高度不敏感(Park et al., 2009).PP2C是一类丝氨酸/苏氨酸残基蛋白磷酸酶, 包括ABI1 (ABA INSENSITIVE 1)、ABI2、HAB1 (HOMOLOGY TO ABA INSENSITIVE 1)、HAB2和AHG3/PP2CA (ABA-HYPERSENSITIVE GERMINATION 3), 在ABA信号转导中起着关键的负调控作用(Hauser et al., 2011).ABI1和ABI2的显性负突变体abi1-1和abi2-1由于蛋白质结构的改变而不能传递ABA信号, 呈现出种子休眠水平下降和对外源ABA不敏感表型(Ma et al., 2009; Park et al., 2009).hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004).AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006).SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014).其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程.双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007).三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009).RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ... ... 启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ...
1 2006
... 种子休眠和萌发是植物生长发育过程中2个紧密相连的阶段, 受到种子发育的内源因素和外界环境因素共同调控.Finch-Savage和Leubner-Metzger (2006)对种子休眠和萌发进行了定义: 休眠是指具有活力的种子在适宜的环境条件下不能萌发的状态; 而萌发则是种子内胚根突破外周的组织结构(胚乳和种皮)而展露在种皮之外的过程.脱落酸(abscisic acid, ABA)是调控种子休眠和萌发过程最重要的植物激素, 具有诱导和维持种子休眠、抑制种子萌发及幼苗生长发育的作用.十几年来, ABA调控种子休眠和萌发的研究取得了重大进展(Holdsworth et al., 2008; Weitbrecht et al., 2011; Shu et al., 2016).刘一灵等(2014)从ABA代谢与种子发育、穗萌和种子萌发等方面进行了综述.操春燕等(2017)则从ABA与细胞分裂素(cytokinin, CK)和赤霉素(gibberellin, GA)等相互作用调控种子萌发的角度概述了相关的最新研究进展.近年来, 随着参与种子ABA代谢和ABA信号基因相关突变体研究的深入, ABA调控种子休眠和萌发的分子机制已逐渐明晰, 本文参考ABA代谢基因、种子成熟基因以及ABA信号基因的遗传功能和表达调控的最新研究成果(表1), 综述ABA调控种子休眠和萌发的分子机制, 以期为植物激素的作用机理研究和农作物分子设计育种提供重要的理论依据. ...
2 2002
... 高等植物种子发育包括胚和胚乳发育(形态发生)及种子成熟, 在该过程中内源ABA的含量会出现2个高峰.拟南芥种子进入成熟阶段(形成心形胚)前, 首个ABA积累高峰主要来自母体韧皮部中ABA的合成, ABA由母体运输至胚内, 抑制种子成熟前的胎萌; 进入成熟阶段后, 内源ABA积累以胚内合成为主, 种子成熟时达到第2个高峰; 且ABA缺陷型突变体在成熟阶段, 母体合成的ABA仍会向胚内运输(Kanno et al., 2010).对拟南芥ABA合成途径相关基因突变体的研究表明, 拟南芥ABA1 (与烟草(Nicotiana tabacum) ABA2同源)缺失突变体aba1呈现植株萎蔫、内源ABA缺乏及ABA诱导的基因表达下调等表型.aba2和aba3表型与aba1相同, 且种子休眠能力下降(Finke- lstein et al., 2002).AtABA2过量表达株系种子中内源ABA含量高于野生型, 萌发延迟(Lin et al., 2007).AtNCED6和AtNCED9均在种子发育期特异性表达, 前者仅在胚乳中表达, 而后者在胚和胚乳中都表达, 是诱导和维持胚休眠的主要基因(Lefebvre et al., 2006).AtNCED6和AtNCED9存在功能冗余, 拟南芥nced6/nced9双突变体种子休眠程度减弱而单突变体无相关表型(Lefebvre et al., 2006).种子成熟后期, 胚内含水量下降, AtNCED5表达上调, 与AtNCED6和AtNCED9协同调控种子休眠, 三突变体nced5/ nced6/nced9休眠水平显著低于三者间的双突变体(Frey et al., 2012).值得关注的是, 只有胚内合成的ABA能诱导和维持种子的初级休眠, 母体合成或胚发育期间外源施加ABA均不能诱导种子的初级休眠(Karssen et al., 1983; Koornneef and Karssen, 1994). ... ... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
1 2000
... 转录组分析显示, 拟南芥干种子中含有种类丰富的mRNA, 包括种子发育晚期丰富蛋白(late embryogenesis abundant, LEA)和其它贮藏蛋白基因的mRNA.绝大多数高丰度mRNA编码基因的启动子中含有ABA应答元件(ABA response element, ABRE).拟南芥bZIP型转录因子具有亮氨酸拉链结构(leucine zipper), 能特异识别基因启动子ABRE应答元件中的ACGT核心序列, 调控相应基因的表达.ABI5作为bZIP型转录因子家族中的重要成员, 能够激活种胚内ABA调控基因的表达, 如LEA蛋白的编码基因EM1 (EARLY METHIONINE-LABELED 1)、EM6和LEAD- 34 (Finkelstein and Lynch, 2000).酵母单杂交实验结果显示, ABI5直接与EM6启动子的ABRE结合从而促进其表达(Nakamura et al., 2001; Carles et al., 2002).种子休眠的关键转录因子DOG1是通过促进ABI5转录, 从而激活下游LEA和HSP (HEAT SHO- CK PROTEIN)基因的表达(Dekkers et al., 2016).ABI5不直接参与种子休眠调控, 但负调控种子的萌发, 说明ABA调控种子休眠和萌发是通过2个不同的信号转导途径(Piskurewicz et al., 2008; Kanai et al., 2010).ABI5能与ABI3进行体外蛋白互作, 且位于ABI3的下游, 激活下游基因表达(Nakamura et al., 2001; Lopez-Molina et al., 2002).此外, 拟南芥转录因子bZIP53通过结合种子贮藏蛋白基因(如SSP和LEA)启动子G-box元件, 促进相关基因表达, 进而调控种子成熟.体外结合实验显示, 当bZIP53与bZIP10或bZIP25异源二聚化后, 能增强bZIP53与G-box的结合活性(Alonso et al., 2009).同时, bZIP10和bZIP25也能促进SSP基因的表达(Lara et al., 2003), 表明这3个转录因子在促进种子成熟过程中具有协同作用. ...
1 2015
... 种子休眠的维持可能与染色质结构相关, 促进种子萌发的基因由于空间位阻不能被转录因子结合而处于静止状态(Shu et al., 2016).拟南芥KYP/SUVH4编码一种H3 Lys 9甲基转移酶, 为H3K9甲基化所需; 同时KYP/SUVH4负调控种子休眠, 其缺失突变体休眠程度增强.研究表明, ABA可能通过抑制KYP/SUVH4的表达从而促进种子休眠(Zheng et al., 2012).组蛋白乙酰转移酶(histone acetyltransferases)通过调节染色质活力来调控基因表达.大麦(Hordeum vulgare)种子经外源ABA处理后, 组蛋白乙酰转移酶基因HvGNAT/MYST表达上调(Papaefthimiou et al., 2010).表观遗传调控因子HUB1 (HISTONE MON- OU-BIQUITINATION 1 )和RDO2 (REDUCED DOR- MANCY 2)在休眠诱导期受到ABA的强烈诱导(Liu et al., 2011).此外, 组蛋白脱甲基酶LDL1 (LYSINES- PECIFIC DEMETHYLASE LIKE 1)和LDL2通过抑制DOG1、ABA2和ABI3的表达, 负调控种子休眠(Zhao et al., 2015).KYP/SUVH4和SUVH5在种子成熟期抑制DOG1和ABI3的表达(Zheng et al., 2012), 且DO- G1的染色质结构重塑参与种子的休眠循环过程(Foo- titt et al., 2015), 表明DOG1可能通过2条不同的途 径诱导种子休眠.关于ABA通过影响染色质结构重塑调节种子休眠与萌发的分子机制, 目前尚不完全明确. ...
1 2012
... 高等植物种子发育包括胚和胚乳发育(形态发生)及种子成熟, 在该过程中内源ABA的含量会出现2个高峰.拟南芥种子进入成熟阶段(形成心形胚)前, 首个ABA积累高峰主要来自母体韧皮部中ABA的合成, ABA由母体运输至胚内, 抑制种子成熟前的胎萌; 进入成熟阶段后, 内源ABA积累以胚内合成为主, 种子成熟时达到第2个高峰; 且ABA缺陷型突变体在成熟阶段, 母体合成的ABA仍会向胚内运输(Kanno et al., 2010).对拟南芥ABA合成途径相关基因突变体的研究表明, 拟南芥ABA1 (与烟草(Nicotiana tabacum) ABA2同源)缺失突变体aba1呈现植株萎蔫、内源ABA缺乏及ABA诱导的基因表达下调等表型.aba2和aba3表型与aba1相同, 且种子休眠能力下降(Finke- lstein et al., 2002).AtABA2过量表达株系种子中内源ABA含量高于野生型, 萌发延迟(Lin et al., 2007).AtNCED6和AtNCED9均在种子发育期特异性表达, 前者仅在胚乳中表达, 而后者在胚和胚乳中都表达, 是诱导和维持胚休眠的主要基因(Lefebvre et al., 2006).AtNCED6和AtNCED9存在功能冗余, 拟南芥nced6/nced9双突变体种子休眠程度减弱而单突变体无相关表型(Lefebvre et al., 2006).种子成熟后期, 胚内含水量下降, AtNCED5表达上调, 与AtNCED6和AtNCED9协同调控种子休眠, 三突变体nced5/ nced6/nced9休眠水平显著低于三者间的双突变体(Frey et al., 2012).值得关注的是, 只有胚内合成的ABA能诱导和维持种子的初级休眠, 母体合成或胚发育期间外源施加ABA均不能诱导种子的初级休眠(Karssen et al., 1983; Koornneef and Karssen, 1994). ...
1 2007
... 拟南芥pyr1/pyl1/pyl2/prl4四突变体种子萌发表现出对ABA高度不敏感(Park et al., 2009).PP2C是一类丝氨酸/苏氨酸残基蛋白磷酸酶, 包括ABI1 (ABA INSENSITIVE 1)、ABI2、HAB1 (HOMOLOGY TO ABA INSENSITIVE 1)、HAB2和AHG3/PP2CA (ABA-HYPERSENSITIVE GERMINATION 3), 在ABA信号转导中起着关键的负调控作用(Hauser et al., 2011).ABI1和ABI2的显性负突变体abi1-1和abi2-1由于蛋白质结构的改变而不能传递ABA信号, 呈现出种子休眠水平下降和对外源ABA不敏感表型(Ma et al., 2009; Park et al., 2009).hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004).AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006).SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014).其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程.双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007).三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009).RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ...
1 2009
... 拟南芥pyr1/pyl1/pyl2/prl4四突变体种子萌发表现出对ABA高度不敏感(Park et al., 2009).PP2C是一类丝氨酸/苏氨酸残基蛋白磷酸酶, 包括ABI1 (ABA INSENSITIVE 1)、ABI2、HAB1 (HOMOLOGY TO ABA INSENSITIVE 1)、HAB2和AHG3/PP2CA (ABA-HYPERSENSITIVE GERMINATION 3), 在ABA信号转导中起着关键的负调控作用(Hauser et al., 2011).ABI1和ABI2的显性负突变体abi1-1和abi2-1由于蛋白质结构的改变而不能传递ABA信号, 呈现出种子休眠水平下降和对外源ABA不敏感表型(Ma et al., 2009; Park et al., 2009).hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004).AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006).SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014).其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程.双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007).三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009).RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ...
1 2007
... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
2 2011
... ABA通过特定的信号转导途径抑制种子萌发.ABA信号转导起始于ABA受体对ABA信号的感知.2009年, 2个实验室各自独立发现1个含有START (STeroidogenic Acute Regulatory Related Lipid Transfer)特征区的蛋白质PYR (PYRABACTIN RESISTANCE)/ PYL (PYR1-LIKE)/RCAR (REGULATORY COMPO- NENTS OF ABA RECEPTORS)家族成员能够结合ABA, 是一种细胞内ABA受体.其作用机理是PYR/ PYL/RCAR通过与下游信号调节子PP2C直接相互作用, 抑制PP2C的活性, 解除PP2C对其下游信号调节子SNF1相关蛋白激酶2 (SNF1-related protein kinase 2, SnRK2)的抑制, 从而使SnRK2能够通过磷酸化激活下游转录因子ABI5/ABF4 (ABRE binding factors), 实现ABA生理效应(图1) (Ma et al., 2009; Park et al., 2009; Hubbard et al., 2010; Hauser et al., 2011). ... ... 拟南芥pyr1/pyl1/pyl2/prl4四突变体种子萌发表现出对ABA高度不敏感(Park et al., 2009).PP2C是一类丝氨酸/苏氨酸残基蛋白磷酸酶, 包括ABI1 (ABA INSENSITIVE 1)、ABI2、HAB1 (HOMOLOGY TO ABA INSENSITIVE 1)、HAB2和AHG3/PP2CA (ABA-HYPERSENSITIVE GERMINATION 3), 在ABA信号转导中起着关键的负调控作用(Hauser et al., 2011).ABI1和ABI2的显性负突变体abi1-1和abi2-1由于蛋白质结构的改变而不能传递ABA信号, 呈现出种子休眠水平下降和对外源ABA不敏感表型(Ma et al., 2009; Park et al., 2009).hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004).AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006).SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014).其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程.双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007).三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009).RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ...
2 2008
... 种子休眠和萌发是植物生长发育过程中2个紧密相连的阶段, 受到种子发育的内源因素和外界环境因素共同调控.Finch-Savage和Leubner-Metzger (2006)对种子休眠和萌发进行了定义: 休眠是指具有活力的种子在适宜的环境条件下不能萌发的状态; 而萌发则是种子内胚根突破外周的组织结构(胚乳和种皮)而展露在种皮之外的过程.脱落酸(abscisic acid, ABA)是调控种子休眠和萌发过程最重要的植物激素, 具有诱导和维持种子休眠、抑制种子萌发及幼苗生长发育的作用.十几年来, ABA调控种子休眠和萌发的研究取得了重大进展(Holdsworth et al., 2008; Weitbrecht et al., 2011; Shu et al., 2016).刘一灵等(2014)从ABA代谢与种子发育、穗萌和种子萌发等方面进行了综述.操春燕等(2017)则从ABA与细胞分裂素(cytokinin, CK)和赤霉素(gibberellin, GA)等相互作用调控种子萌发的角度概述了相关的最新研究进展.近年来, 随着参与种子ABA代谢和ABA信号基因相关突变体研究的深入, ABA调控种子休眠和萌发的分子机制已逐渐明晰, 本文参考ABA代谢基因、种子成熟基因以及ABA信号基因的遗传功能和表达调控的最新研究成果(表1), 综述ABA调控种子休眠和萌发的分子机制, 以期为植物激素的作用机理研究和农作物分子设计育种提供重要的理论依据. ... ... ABA在促进种子成熟及诱导和维持种子休眠中发挥重要作用.在种子发育过程中, ABA对种子休眠的诱导和维持与种子内源ABA水平及种胚发育信号密切相关(Holdsworth et al., 2008; Shu et al., 2016).在种子发育过程中, 内源ABA水平的降低导致成熟种子的休眠能力下降甚至丧失, 而ABA含量的增加则会使种子休眠能力增强及萌发推迟(Okamoto et al., 2006; Nonogaki et al., 2014). ...
1 2015
... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
... ABA通过特定的信号转导途径抑制种子萌发.ABA信号转导起始于ABA受体对ABA信号的感知.2009年, 2个实验室各自独立发现1个含有START (STeroidogenic Acute Regulatory Related Lipid Transfer)特征区的蛋白质PYR (PYRABACTIN RESISTANCE)/ PYL (PYR1-LIKE)/RCAR (REGULATORY COMPO- NENTS OF ABA RECEPTORS)家族成员能够结合ABA, 是一种细胞内ABA受体.其作用机理是PYR/ PYL/RCAR通过与下游信号调节子PP2C直接相互作用, 抑制PP2C的活性, 解除PP2C对其下游信号调节子SNF1相关蛋白激酶2 (SNF1-related protein kinase 2, SnRK2)的抑制, 从而使SnRK2能够通过磷酸化激活下游转录因子ABI5/ABF4 (ABRE binding factors), 实现ABA生理效应(图1) (Ma et al., 2009; Park et al., 2009; Hubbard et al., 2010; Hauser et al., 2011). ...
1 2014
... APETALA 2/乙烯应答元件结合因子(AP2/ERF)是植物特有的转录因子家族, 调控植物生长发育及胁迫响应.目前已证实在拟南芥和水稻(Oryza sativa)中, AP2型转录因子通过调节ABA/GA的平衡来调控种子的休眠和萌发.拟南芥ABI4通过结合CYP707A1和CYP707A2基因启动子的CCAC顺式元件抑制其表达, 同时直接促进NCED6和GA2ox7 (GA2 oxidase 7)基因的表达, 且ABI4缺失突变体能恢复ga1-t、cyp707a1和cyp707a2突变体不萌发的表型, 证明ABI4通过正调控ABA合成和负调控GA合成来调控种子的休眠与萌发(Shu et al., 2013, 2018).ABI4基因的转录受到控制种子休眠的关键转录因子SPT (SPATULA)的调控, 提示SPT-ABI4在种子休眠的诱导和维持方面具有重要作用(Vaistij et al., 2013).拟南芥CHO1 (CHOTTO1)具有双重的AP2结构, CHO1在ABI4下游, 正调控种子休眠, 通过促进ABA1/ZEP和NCED9的表达, 抑制CYP707A2和CYP707A3的表达, 调控种子萌发过程中ABA介导的GA合成抑制过程(Yano et al., 2009).拟南芥ORA47 (octadeca- noid-responsive AP2/ERF-domain transcription fac- tor 47)和DREB2C通过正调控NCED3和NCED9的表达, 促进ABA合成, 从而对非生物胁迫作出响应(Je et al., 2014; Chen et al., 2016).此外, 转录因子MYB96通过调控ABI4和ABA合成基因NCED2与NCED6的表达, 促进种子休眠而抑制萌发(Lee et al., 2015a, 2015b). ...
1 2014
... 拟南芥中, 调控种胚发育和种子成熟的转录因子主要由B3型转录因子的AFL支线ABI3 (ABA INSENSITIVE 3)、 FUS3 (FUSCA 3)和LEC2 (LEAFY COTY- LEDON 2), 以及2个LEC1型NF-YB (NUCLEAR FA- CTOR Y B subunit)家族成员LEC1和L1L (LEC1- LIKE)组成, 统称为LAFL.B3型转录因子最先是从玉米(Zea mays) vp1 (viviparous1)和拟南芥abi3 (abscisic acid insensitive 3)突变体中被发现, 属于植物特有的B3型DNA结合结构域蛋白家族.该家族蛋白能识别种子特异基因启动子的RY重复或Sph元件的保守序列, 具有促进种胚发育和成熟的功能(Jia et al., 2014).其中, ABI3是种子休眠与萌发的主要调控因子, 具有调控种皮色素改变、诱导种子休眠和促进贮藏蛋白积累等功能(Dekkers et al., 2016).在ABI3上游, Barrero等(2010)发现1个RING-finger蛋白(一类E3泛素连接酶)的编码基因DEP (DESPIERTO), 在种子发育期间调控ABI3表达并影响休眠的诱导, 突变体dep-1种子休眠完全丧失.FUS3在种子发育中后期增强NCED6、NCED9和ABA2的表达, 促进种子干旱耐受性的获得, 并决定子叶上皮细胞的一致性(Tiedemann et al., 2008; Yamamoto et al., 2010).LEC2在胚胎发育早中期表达, 促进贮藏蛋白和油体蛋白的积累, 并参与种子成熟阶段的基因表达调控, 如DOG1 (DELAY OF GERMINATION 1) (Braybr- ook et al., 2006).拟南芥DOG1蛋白是种子休眠的关键调控因子, 通过抑制特定蛋白磷酸酶2C (protein phosphatase 2Cs, PP2C)活性来调控休眠, 且其蛋白水平决定了休眠释放的时机.dog1突变体种子休眠完全缺失(Née et al., 2017).LEC1型NF-YB家族转录因子具有结合CCAAT-box的结构域, 通过调控AFL基因促进种子贮藏蛋白(seed storage protein, SSP)基因的表达(Kagaya et al., 2005).LEC1在胚胎发育早期表达并促进种子胚胎形成(Lotan et al., 1998).L1L在发育中的胚胎(稍晚于LEC1)以及营养器官中表达, 表明除正向调控种胚发育外, L1L还具有与LEC1不同的调控功能(Kwong et al., 2003). ...
2 2009
... 研究表明, 拟南芥ABAR/CHLH过表达株系表现出对ABA的高度敏感, 而突变体abar-2和ABAR RN- Ai株系的种子萌发对ABA不敏感.上述结果与ABAR作为ABA受体的结论相符.WRKY40是细胞内关键的ABA信号负调控因子, 在细胞核中通过抑制ABI5的表达而抑制下游的ABA应答反应.WRKY40缺失突变体种子萌发表型对ABA高度敏感(Jiang and Yu, 2009; Shang et al., 2010).拟南芥种子萌发及之后早期幼苗生长发育阶段, ABA诱导WRKY2的积累需要ABI3、ABI5、ABA2和ABA3的存在.wrky2缺失突变体种子萌发及幼苗生长对ABA超敏感, 说明WRKY2也负调控ABA抑制的种子萌发和幼苗早期生长发育(Jiang and Yu, 2009).另外, 早期研究表明, 野燕麦(Avena fatua) WRKY蛋白ABF1和ABF2能结合在α-淀粉酶编码基因启动子的box2/W-box元件上, 促进胚乳中淀粉的水解, 从而间接促进种子萌发(Rush- ton et al., 1995). ... ... 也负调控ABA抑制的种子萌发和幼苗早期生长发育(Jiang and Yu, 2009).另外, 早期研究表明, 野燕麦(Avena fatua) WRKY蛋白ABF1和ABF2能结合在α-淀粉酶编码基因启动子的box2/W-box元件上, 促进胚乳中淀粉的水解, 从而间接促进种子萌发(Rush- ton et al., 1995). ...
1 2012
... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
2 2005
... 拟南芥中, 调控种胚发育和种子成熟的转录因子主要由B3型转录因子的AFL支线ABI3 (ABA INSENSITIVE 3)、 FUS3 (FUSCA 3)和LEC2 (LEAFY COTY- LEDON 2), 以及2个LEC1型NF-YB (NUCLEAR FA- CTOR Y B subunit)家族成员LEC1和L1L (LEC1- LIKE)组成, 统称为LAFL.B3型转录因子最先是从玉米(Zea mays) vp1 (viviparous1)和拟南芥abi3 (abscisic acid insensitive 3)突变体中被发现, 属于植物特有的B3型DNA结合结构域蛋白家族.该家族蛋白能识别种子特异基因启动子的RY重复或Sph元件的保守序列, 具有促进种胚发育和成熟的功能(Jia et al., 2014).其中, ABI3是种子休眠与萌发的主要调控因子, 具有调控种皮色素改变、诱导种子休眠和促进贮藏蛋白积累等功能(Dekkers et al., 2016).在ABI3上游, Barrero等(2010)发现1个RING-finger蛋白(一类E3泛素连接酶)的编码基因DEP (DESPIERTO), 在种子发育期间调控ABI3表达并影响休眠的诱导, 突变体dep-1种子休眠完全丧失.FUS3在种子发育中后期增强NCED6、NCED9和ABA2的表达, 促进种子干旱耐受性的获得, 并决定子叶上皮细胞的一致性(Tiedemann et al., 2008; Yamamoto et al., 2010).LEC2在胚胎发育早中期表达, 促进贮藏蛋白和油体蛋白的积累, 并参与种子成熟阶段的基因表达调控, 如DOG1 (DELAY OF GERMINATION 1) (Braybr- ook et al., 2006).拟南芥DOG1蛋白是种子休眠的关键调控因子, 通过抑制特定蛋白磷酸酶2C (protein phosphatase 2Cs, PP2C)活性来调控休眠, 且其蛋白水平决定了休眠释放的时机.dog1突变体种子休眠完全缺失(Née et al., 2017).LEC1型NF-YB家族转录因子具有结合CCAAT-box的结构域, 通过调控AFL基因促进种子贮藏蛋白(seed storage protein, SSP)基因的表达(Kagaya et al., 2005).LEC1在胚胎发育早期表达并促进种子胚胎形成(Lotan et al., 1998).L1L在发育中的胚胎(稍晚于LEC1)以及营养器官中表达, 表明除正向调控种胚发育外, L1L还具有与LEC1不同的调控功能(Kwong et al., 2003). ... ... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
1 2010
... 转录组分析显示, 拟南芥干种子中含有种类丰富的mRNA, 包括种子发育晚期丰富蛋白(late embryogenesis abundant, LEA)和其它贮藏蛋白基因的mRNA.绝大多数高丰度mRNA编码基因的启动子中含有ABA应答元件(ABA response element, ABRE).拟南芥bZIP型转录因子具有亮氨酸拉链结构(leucine zipper), 能特异识别基因启动子ABRE应答元件中的ACGT核心序列, 调控相应基因的表达.ABI5作为bZIP型转录因子家族中的重要成员, 能够激活种胚内ABA调控基因的表达, 如LEA蛋白的编码基因EM1 (EARLY METHIONINE-LABELED 1)、EM6和LEAD- 34 (Finkelstein and Lynch, 2000).酵母单杂交实验结果显示, ABI5直接与EM6启动子的ABRE结合从而促进其表达(Nakamura et al., 2001; Carles et al., 2002).种子休眠的关键转录因子DOG1是通过促进ABI5转录, 从而激活下游LEA和HSP (HEAT SHO- CK PROTEIN)基因的表达(Dekkers et al., 2016).ABI5不直接参与种子休眠调控, 但负调控种子的萌发, 说明ABA调控种子休眠和萌发是通过2个不同的信号转导途径(Piskurewicz et al., 2008; Kanai et al., 2010).ABI5能与ABI3进行体外蛋白互作, 且位于ABI3的下游, 激活下游基因表达(Nakamura et al., 2001; Lopez-Molina et al., 2002).此外, 拟南芥转录因子bZIP53通过结合种子贮藏蛋白基因(如SSP和LEA)启动子G-box元件, 促进相关基因表达, 进而调控种子成熟.体外结合实验显示, 当bZIP53与bZIP10或bZIP25异源二聚化后, 能增强bZIP53与G-box的结合活性(Alonso et al., 2009).同时, bZIP10和bZIP25也能促进SSP基因的表达(Lara et al., 2003), 表明这3个转录因子在促进种子成熟过程中具有协同作用. ...
1 2010
... 高等植物种子发育包括胚和胚乳发育(形态发生)及种子成熟, 在该过程中内源ABA的含量会出现2个高峰.拟南芥种子进入成熟阶段(形成心形胚)前, 首个ABA积累高峰主要来自母体韧皮部中ABA的合成, ABA由母体运输至胚内, 抑制种子成熟前的胎萌; 进入成熟阶段后, 内源ABA积累以胚内合成为主, 种子成熟时达到第2个高峰; 且ABA缺陷型突变体在成熟阶段, 母体合成的ABA仍会向胚内运输(Kanno et al., 2010).对拟南芥ABA合成途径相关基因突变体的研究表明, 拟南芥ABA1 (与烟草(Nicotiana tabacum) ABA2同源)缺失突变体aba1呈现植株萎蔫、内源ABA缺乏及ABA诱导的基因表达下调等表型.aba2和aba3表型与aba1相同, 且种子休眠能力下降(Finke- lstein et al., 2002).AtABA2过量表达株系种子中内源ABA含量高于野生型, 萌发延迟(Lin et al., 2007).AtNCED6和AtNCED9均在种子发育期特异性表达, 前者仅在胚乳中表达, 而后者在胚和胚乳中都表达, 是诱导和维持胚休眠的主要基因(Lefebvre et al., 2006).AtNCED6和AtNCED9存在功能冗余, 拟南芥nced6/nced9双突变体种子休眠程度减弱而单突变体无相关表型(Lefebvre et al., 2006).种子成熟后期, 胚内含水量下降, AtNCED5表达上调, 与AtNCED6和AtNCED9协同调控种子休眠, 三突变体nced5/ nced6/nced9休眠水平显著低于三者间的双突变体(Frey et al., 2012).值得关注的是, 只有胚内合成的ABA能诱导和维持种子的初级休眠, 母体合成或胚发育期间外源施加ABA均不能诱导种子的初级休眠(Karssen et al., 1983; Koornneef and Karssen, 1994). ...
1 1983
... 高等植物种子发育包括胚和胚乳发育(形态发生)及种子成熟, 在该过程中内源ABA的含量会出现2个高峰.拟南芥种子进入成熟阶段(形成心形胚)前, 首个ABA积累高峰主要来自母体韧皮部中ABA的合成, ABA由母体运输至胚内, 抑制种子成熟前的胎萌; 进入成熟阶段后, 内源ABA积累以胚内合成为主, 种子成熟时达到第2个高峰; 且ABA缺陷型突变体在成熟阶段, 母体合成的ABA仍会向胚内运输(Kanno et al., 2010).对拟南芥ABA合成途径相关基因突变体的研究表明, 拟南芥ABA1 (与烟草(Nicotiana tabacum) ABA2同源)缺失突变体aba1呈现植株萎蔫、内源ABA缺乏及ABA诱导的基因表达下调等表型.aba2和aba3表型与aba1相同, 且种子休眠能力下降(Finke- lstein et al., 2002).AtABA2过量表达株系种子中内源ABA含量高于野生型, 萌发延迟(Lin et al., 2007).AtNCED6和AtNCED9均在种子发育期特异性表达, 前者仅在胚乳中表达, 而后者在胚和胚乳中都表达, 是诱导和维持胚休眠的主要基因(Lefebvre et al., 2006).AtNCED6和AtNCED9存在功能冗余, 拟南芥nced6/nced9双突变体种子休眠程度减弱而单突变体无相关表型(Lefebvre et al., 2006).种子成熟后期, 胚内含水量下降, AtNCED5表达上调, 与AtNCED6和AtNCED9协同调控种子休眠, 三突变体nced5/ nced6/nced9休眠水平显著低于三者间的双突变体(Frey et al., 2012).值得关注的是, 只有胚内合成的ABA能诱导和维持种子的初级休眠, 母体合成或胚发育期间外源施加ABA均不能诱导种子的初级休眠(Karssen et al., 1983; Koornneef and Karssen, 1994). ...
1 2014
... 种子休眠的维持取决于种子内源ABA与GA含量的比值(ABA/GA), 而休眠解除是由种子内源ABA含量下降和GA含量升高所致(Cadman et al., 2006).拟南芥休眠型Cvi (cape verde islands)和非休眠型Col (Columbia)种子吸胀2-3小时后, CYP707A2转录均被激活, 导致ABA含量迅速下降, 说明CYP707A2是种子吸胀过程中ABA降解的主要代谢酶(Preston et al., 2009).研究表明, 休眠型Cvi种子后熟作用的休眠释放(dormancy release)与CYP707A2和GA3ox1基因表达相关.cyp707a2缺失突变体种子中GA4含量和GA3ox1的转录水平均有所下降, 表明种子萌发时ABA通过负调控GA合成来抑制种子萌发(Yano et al., 2009).同样, GA也能够抑制ABA的合成途径, ABA与GA的拮抗作用主要受AP2型转录因子的调控(Oh et al., 2007; Shu et al., 2013).种子吸胀过程中, GA合成基因GA3ox和GA20ox高表达, 促使GA在种胚内大量积累(Ogawa et al., 2003).种子萌发早期, ABA/GA比值下降至原有水平的1/3; 而在萌发后期, ABA/GA比值下降至1/10 (Weitbrecht et al., 2011).淀粉酶作为种子萌发过程中重要的水解酶, 通过分解胚乳中的营养物质来为萌发中的胚提供能量和营养.Kondhare等(2014)使用外源激素处理小麦(Triticum aestivum)和水稻种子时, 发现ABA抑制而GA促进α-淀粉酶的活性, 可见ABA/GA平衡还能调控种子内部营养物质的改变. ...
1 1994
... 高等植物种子发育包括胚和胚乳发育(形态发生)及种子成熟, 在该过程中内源ABA的含量会出现2个高峰.拟南芥种子进入成熟阶段(形成心形胚)前, 首个ABA积累高峰主要来自母体韧皮部中ABA的合成, ABA由母体运输至胚内, 抑制种子成熟前的胎萌; 进入成熟阶段后, 内源ABA积累以胚内合成为主, 种子成熟时达到第2个高峰; 且ABA缺陷型突变体在成熟阶段, 母体合成的ABA仍会向胚内运输(Kanno et al., 2010).对拟南芥ABA合成途径相关基因突变体的研究表明, 拟南芥ABA1 (与烟草(Nicotiana tabacum) ABA2同源)缺失突变体aba1呈现植株萎蔫、内源ABA缺乏及ABA诱导的基因表达下调等表型.aba2和aba3表型与aba1相同, 且种子休眠能力下降(Finke- lstein et al., 2002).AtABA2过量表达株系种子中内源ABA含量高于野生型, 萌发延迟(Lin et al., 2007).AtNCED6和AtNCED9均在种子发育期特异性表达, 前者仅在胚乳中表达, 而后者在胚和胚乳中都表达, 是诱导和维持胚休眠的主要基因(Lefebvre et al., 2006).AtNCED6和AtNCED9存在功能冗余, 拟南芥nced6/nced9双突变体种子休眠程度减弱而单突变体无相关表型(Lefebvre et al., 2006).种子成熟后期, 胚内含水量下降, AtNCED5表达上调, 与AtNCED6和AtNCED9协同调控种子休眠, 三突变体nced5/ nced6/nced9休眠水平显著低于三者间的双突变体(Frey et al., 2012).值得关注的是, 只有胚内合成的ABA能诱导和维持种子的初级休眠, 母体合成或胚发育期间外源施加ABA均不能诱导种子的初级休眠(Karssen et al., 1983; Koornneef and Karssen, 1994). ...
1 2013
... 近年来, 一些NF-Y转录因子也被发现通过影响ABA信号调节种子萌发.酵母双杂交分析表明, NF- YB2能与ABF3 (ABA RESPONSE ELEMENT BIND- ING PROTEINS 3/ABA BINDING FACTORS 3)结合, NF-YC9能与ABF2结合; NF-YB2和NF-YB3过量表达时表现出对ABA敏感性的显著改变, 但缺失突变体无相关表型, 提示NF-YB2和NF-YB3并不是影响种子萌发的关键转录因子.三突变体nf-yc3/nf-yc4/ nf-yc9种子萌发对ABA敏感性增强, 而nf-yc4单突变体种子萌发对ABA敏感性下降, 提示NF-YC3、NF-YC4和NF-YC9基因之间存在功能冗余(Kumimo- to et al., 2013).进一步研究表明, NF-YC (NF-YC3、NF-YC4和NF-YC9)能与DELLA蛋白RGL2 (GA信号的关键负调控蛋白)构成复合物NF-YC-RGL2, 通过识别ABI5启动子的CCAAT元件, 促进ABI5表达从而激活下游的ABA和GA介导的种子萌发过程(Liu et al., 2016).此外, 研究人员还发现位于叶绿体内的ABA- R/CHLH互作蛋白——伴侣蛋白20 (co-chaperonin 20, CPN20)负调控ABA信号转导通路.cpn20-1突变体中WRKY40表达下调, 且CPN20通过其N-Signa- ture结构域与WRKY40竞争性结合ABAR, 从而减少ABAR-WRKY40互作, 抑制ABA下游信号.相反, ABA又通过直接抑制ABAR-CPN20的互作降低CPN- 20的表达, 同时促进ABAR-WRKY40互作来促进信号传递(图1) (Zhang et al., 2013, 2014). ...
... 转录组分析显示, 拟南芥干种子中含有种类丰富的mRNA, 包括种子发育晚期丰富蛋白(late embryogenesis abundant, LEA)和其它贮藏蛋白基因的mRNA.绝大多数高丰度mRNA编码基因的启动子中含有ABA应答元件(ABA response element, ABRE).拟南芥bZIP型转录因子具有亮氨酸拉链结构(leucine zipper), 能特异识别基因启动子ABRE应答元件中的ACGT核心序列, 调控相应基因的表达.ABI5作为bZIP型转录因子家族中的重要成员, 能够激活种胚内ABA调控基因的表达, 如LEA蛋白的编码基因EM1 (EARLY METHIONINE-LABELED 1)、EM6和LEAD- 34 (Finkelstein and Lynch, 2000).酵母单杂交实验结果显示, ABI5直接与EM6启动子的ABRE结合从而促进其表达(Nakamura et al., 2001; Carles et al., 2002).种子休眠的关键转录因子DOG1是通过促进ABI5转录, 从而激活下游LEA和HSP (HEAT SHO- CK PROTEIN)基因的表达(Dekkers et al., 2016).ABI5不直接参与种子休眠调控, 但负调控种子的萌发, 说明ABA调控种子休眠和萌发是通过2个不同的信号转导途径(Piskurewicz et al., 2008; Kanai et al., 2010).ABI5能与ABI3进行体外蛋白互作, 且位于ABI3的下游, 激活下游基因表达(Nakamura et al., 2001; Lopez-Molina et al., 2002).此外, 拟南芥转录因子bZIP53通过结合种子贮藏蛋白基因(如SSP和LEA)启动子G-box元件, 促进相关基因表达, 进而调控种子成熟.体外结合实验显示, 当bZIP53与bZIP10或bZIP25异源二聚化后, 能增强bZIP53与G-box的结合活性(Alonso et al., 2009).同时, bZIP10和bZIP25也能促进SSP基因的表达(Lara et al., 2003), 表明这3个转录因子在促进种子成熟过程中具有协同作用. ...
1 1998
... 拟南芥中, 调控种胚发育和种子成熟的转录因子主要由B3型转录因子的AFL支线ABI3 (ABA INSENSITIVE 3)、 FUS3 (FUSCA 3)和LEC2 (LEAFY COTY- LEDON 2), 以及2个LEC1型NF-YB (NUCLEAR FA- CTOR Y B subunit)家族成员LEC1和L1L (LEC1- LIKE)组成, 统称为LAFL.B3型转录因子最先是从玉米(Zea mays) vp1 (viviparous1)和拟南芥abi3 (abscisic acid insensitive 3)突变体中被发现, 属于植物特有的B3型DNA结合结构域蛋白家族.该家族蛋白能识别种子特异基因启动子的RY重复或Sph元件的保守序列, 具有促进种胚发育和成熟的功能(Jia et al., 2014).其中, ABI3是种子休眠与萌发的主要调控因子, 具有调控种皮色素改变、诱导种子休眠和促进贮藏蛋白积累等功能(Dekkers et al., 2016).在ABI3上游, Barrero等(2010)发现1个RING-finger蛋白(一类E3泛素连接酶)的编码基因DEP (DESPIERTO), 在种子发育期间调控ABI3表达并影响休眠的诱导, 突变体dep-1种子休眠完全丧失.FUS3在种子发育中后期增强NCED6、NCED9和ABA2的表达, 促进种子干旱耐受性的获得, 并决定子叶上皮细胞的一致性(Tiedemann et al., 2008; Yamamoto et al., 2010).LEC2在胚胎发育早中期表达, 促进贮藏蛋白和油体蛋白的积累, 并参与种子成熟阶段的基因表达调控, 如DOG1 (DELAY OF GERMINATION 1) (Braybr- ook et al., 2006).拟南芥DOG1蛋白是种子休眠的关键调控因子, 通过抑制特定蛋白磷酸酶2C (protein phosphatase 2Cs, PP2C)活性来调控休眠, 且其蛋白水平决定了休眠释放的时机.dog1突变体种子休眠完全缺失(Née et al., 2017).LEC1型NF-YB家族转录因子具有结合CCAAT-box的结构域, 通过调控AFL基因促进种子贮藏蛋白(seed storage protein, SSP)基因的表达(Kagaya et al., 2005).LEC1在胚胎发育早期表达并促进种子胚胎形成(Lotan et al., 1998).L1L在发育中的胚胎(稍晚于LEC1)以及营养器官中表达, 表明除正向调控种胚发育外, L1L还具有与LEC1不同的调控功能(Kwong et al., 2003). ...
2 2009
... ABA通过特定的信号转导途径抑制种子萌发.ABA信号转导起始于ABA受体对ABA信号的感知.2009年, 2个实验室各自独立发现1个含有START (STeroidogenic Acute Regulatory Related Lipid Transfer)特征区的蛋白质PYR (PYRABACTIN RESISTANCE)/ PYL (PYR1-LIKE)/RCAR (REGULATORY COMPO- NENTS OF ABA RECEPTORS)家族成员能够结合ABA, 是一种细胞内ABA受体.其作用机理是PYR/ PYL/RCAR通过与下游信号调节子PP2C直接相互作用, 抑制PP2C的活性, 解除PP2C对其下游信号调节子SNF1相关蛋白激酶2 (SNF1-related protein kinase 2, SnRK2)的抑制, 从而使SnRK2能够通过磷酸化激活下游转录因子ABI5/ABF4 (ABRE binding factors), 实现ABA生理效应(图1) (Ma et al., 2009; Park et al., 2009; Hubbard et al., 2010; Hauser et al., 2011). ... ... 拟南芥pyr1/pyl1/pyl2/prl4四突变体种子萌发表现出对ABA高度不敏感(Park et al., 2009).PP2C是一类丝氨酸/苏氨酸残基蛋白磷酸酶, 包括ABI1 (ABA INSENSITIVE 1)、ABI2、HAB1 (HOMOLOGY TO ABA INSENSITIVE 1)、HAB2和AHG3/PP2CA (ABA-HYPERSENSITIVE GERMINATION 3), 在ABA信号转导中起着关键的负调控作用(Hauser et al., 2011).ABI1和ABI2的显性负突变体abi1-1和abi2-1由于蛋白质结构的改变而不能传递ABA信号, 呈现出种子休眠水平下降和对外源ABA不敏感表型(Ma et al., 2009; Park et al., 2009).hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004).AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006).SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014).其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程.双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007).三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009).RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ...
... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
... 转录组分析显示, 拟南芥干种子中含有种类丰富的mRNA, 包括种子发育晚期丰富蛋白(late embryogenesis abundant, LEA)和其它贮藏蛋白基因的mRNA.绝大多数高丰度mRNA编码基因的启动子中含有ABA应答元件(ABA response element, ABRE).拟南芥bZIP型转录因子具有亮氨酸拉链结构(leucine zipper), 能特异识别基因启动子ABRE应答元件中的ACGT核心序列, 调控相应基因的表达.ABI5作为bZIP型转录因子家族中的重要成员, 能够激活种胚内ABA调控基因的表达, 如LEA蛋白的编码基因EM1 (EARLY METHIONINE-LABELED 1)、EM6和LEAD- 34 (Finkelstein and Lynch, 2000).酵母单杂交实验结果显示, ABI5直接与EM6启动子的ABRE结合从而促进其表达(Nakamura et al., 2001; Carles et al., 2002).种子休眠的关键转录因子DOG1是通过促进ABI5转录, 从而激活下游LEA和HSP (HEAT SHO- CK PROTEIN)基因的表达(Dekkers et al., 2016).ABI5不直接参与种子休眠调控, 但负调控种子的萌发, 说明ABA调控种子休眠和萌发是通过2个不同的信号转导途径(Piskurewicz et al., 2008; Kanai et al., 2010).ABI5能与ABI3进行体外蛋白互作, 且位于ABI3的下游, 激活下游基因表达(Nakamura et al., 2001; Lopez-Molina et al., 2002).此外, 拟南芥转录因子bZIP53通过结合种子贮藏蛋白基因(如SSP和LEA)启动子G-box元件, 促进相关基因表达, 进而调控种子成熟.体外结合实验显示, 当bZIP53与bZIP10或bZIP25异源二聚化后, 能增强bZIP53与G-box的结合活性(Alonso et al., 2009).同时, bZIP10和bZIP25也能促进SSP基因的表达(Lara et al., 2003), 表明这3个转录因子在促进种子成熟过程中具有协同作用. ... ... ).ABI5能与ABI3进行体外蛋白互作, 且位于ABI3的下游, 激活下游基因表达(Nakamura et al., 2001; Lopez-Molina et al., 2002).此外, 拟南芥转录因子bZIP53通过结合种子贮藏蛋白基因(如SSP和LEA)启动子G-box元件, 促进相关基因表达, 进而调控种子成熟.体外结合实验显示, 当bZIP53与bZIP10或bZIP25异源二聚化后, 能增强bZIP53与G-box的结合活性(Alonso et al., 2009).同时, bZIP10和bZIP25也能促进SSP基因的表达(Lara et al., 2003), 表明这3个转录因子在促进种子成熟过程中具有协同作用. ...
1 2009
... 拟南芥pyr1/pyl1/pyl2/prl4四突变体种子萌发表现出对ABA高度不敏感(Park et al., 2009).PP2C是一类丝氨酸/苏氨酸残基蛋白磷酸酶, 包括ABI1 (ABA INSENSITIVE 1)、ABI2、HAB1 (HOMOLOGY TO ABA INSENSITIVE 1)、HAB2和AHG3/PP2CA (ABA-HYPERSENSITIVE GERMINATION 3), 在ABA信号转导中起着关键的负调控作用(Hauser et al., 2011).ABI1和ABI2的显性负突变体abi1-1和abi2-1由于蛋白质结构的改变而不能传递ABA信号, 呈现出种子休眠水平下降和对外源ABA不敏感表型(Ma et al., 2009; Park et al., 2009).hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004).AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006).SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014).其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程.双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007).三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009).RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ...
1 2017
... 拟南芥中, 调控种胚发育和种子成熟的转录因子主要由B3型转录因子的AFL支线ABI3 (ABA INSENSITIVE 3)、 FUS3 (FUSCA 3)和LEC2 (LEAFY COTY- LEDON 2), 以及2个LEC1型NF-YB (NUCLEAR FA- CTOR Y B subunit)家族成员LEC1和L1L (LEC1- LIKE)组成, 统称为LAFL.B3型转录因子最先是从玉米(Zea mays) vp1 (viviparous1)和拟南芥abi3 (abscisic acid insensitive 3)突变体中被发现, 属于植物特有的B3型DNA结合结构域蛋白家族.该家族蛋白能识别种子特异基因启动子的RY重复或Sph元件的保守序列, 具有促进种胚发育和成熟的功能(Jia et al., 2014).其中, ABI3是种子休眠与萌发的主要调控因子, 具有调控种皮色素改变、诱导种子休眠和促进贮藏蛋白积累等功能(Dekkers et al., 2016).在ABI3上游, Barrero等(2010)发现1个RING-finger蛋白(一类E3泛素连接酶)的编码基因DEP (DESPIERTO), 在种子发育期间调控ABI3表达并影响休眠的诱导, 突变体dep-1种子休眠完全丧失.FUS3在种子发育中后期增强NCED6、NCED9和ABA2的表达, 促进种子干旱耐受性的获得, 并决定子叶上皮细胞的一致性(Tiedemann et al., 2008; Yamamoto et al., 2010).LEC2在胚胎发育早中期表达, 促进贮藏蛋白和油体蛋白的积累, 并参与种子成熟阶段的基因表达调控, 如DOG1 (DELAY OF GERMINATION 1) (Braybr- ook et al., 2006).拟南芥DOG1蛋白是种子休眠的关键调控因子, 通过抑制特定蛋白磷酸酶2C (protein phosphatase 2Cs, PP2C)活性来调控休眠, 且其蛋白水平决定了休眠释放的时机.dog1突变体种子休眠完全缺失(Née et al., 2017).LEC1型NF-YB家族转录因子具有结合CCAAT-box的结构域, 通过调控AFL基因促进种子贮藏蛋白(seed storage protein, SSP)基因的表达(Kagaya et al., 2005).LEC1在胚胎发育早期表达并促进种子胚胎形成(Lotan et al., 1998).L1L在发育中的胚胎(稍晚于LEC1)以及营养器官中表达, 表明除正向调控种胚发育外, L1L还具有与LEC1不同的调控功能(Kwong et al., 2003). ...
1 2014
... ABA在促进种子成熟及诱导和维持种子休眠中发挥重要作用.在种子发育过程中, ABA对种子休眠的诱导和维持与种子内源ABA水平及种胚发育信号密切相关(Holdsworth et al., 2008; Shu et al., 2016).在种子发育过程中, 内源ABA水平的降低导致成熟种子的休眠能力下降甚至丧失, 而ABA含量的增加则会使种子休眠能力增强及萌发推迟(Okamoto et al., 2006; Nonogaki et al., 2014). ...
1 2003
... 种子休眠的维持取决于种子内源ABA与GA含量的比值(ABA/GA), 而休眠解除是由种子内源ABA含量下降和GA含量升高所致(Cadman et al., 2006).拟南芥休眠型Cvi (cape verde islands)和非休眠型Col (Columbia)种子吸胀2-3小时后, CYP707A2转录均被激活, 导致ABA含量迅速下降, 说明CYP707A2是种子吸胀过程中ABA降解的主要代谢酶(Preston et al., 2009).研究表明, 休眠型Cvi种子后熟作用的休眠释放(dormancy release)与CYP707A2和GA3ox1基因表达相关.cyp707a2缺失突变体种子中GA4含量和GA3ox1的转录水平均有所下降, 表明种子萌发时ABA通过负调控GA合成来抑制种子萌发(Yano et al., 2009).同样, GA也能够抑制ABA的合成途径, ABA与GA的拮抗作用主要受AP2型转录因子的调控(Oh et al., 2007; Shu et al., 2013).种子吸胀过程中, GA合成基因GA3ox和GA20ox高表达, 促使GA在种胚内大量积累(Ogawa et al., 2003).种子萌发早期, ABA/GA比值下降至原有水平的1/3; 而在萌发后期, ABA/GA比值下降至1/10 (Weitbrecht et al., 2011).淀粉酶作为种子萌发过程中重要的水解酶, 通过分解胚乳中的营养物质来为萌发中的胚提供能量和营养.Kondhare等(2014)使用外源激素处理小麦(Triticum aestivum)和水稻种子时, 发现ABA抑制而GA促进α-淀粉酶的活性, 可见ABA/GA平衡还能调控种子内部营养物质的改变. ...
1 2007
... 种子休眠的维持取决于种子内源ABA与GA含量的比值(ABA/GA), 而休眠解除是由种子内源ABA含量下降和GA含量升高所致(Cadman et al., 2006).拟南芥休眠型Cvi (cape verde islands)和非休眠型Col (Columbia)种子吸胀2-3小时后, CYP707A2转录均被激活, 导致ABA含量迅速下降, 说明CYP707A2是种子吸胀过程中ABA降解的主要代谢酶(Preston et al., 2009).研究表明, 休眠型Cvi种子后熟作用的休眠释放(dormancy release)与CYP707A2和GA3ox1基因表达相关.cyp707a2缺失突变体种子中GA4含量和GA3ox1的转录水平均有所下降, 表明种子萌发时ABA通过负调控GA合成来抑制种子萌发(Yano et al., 2009).同样, GA也能够抑制ABA的合成途径, ABA与GA的拮抗作用主要受AP2型转录因子的调控(Oh et al., 2007; Shu et al., 2013).种子吸胀过程中, GA合成基因GA3ox和GA20ox高表达, 促使GA在种胚内大量积累(Ogawa et al., 2003).种子萌发早期, ABA/GA比值下降至原有水平的1/3; 而在萌发后期, ABA/GA比值下降至1/10 (Weitbrecht et al., 2011).淀粉酶作为种子萌发过程中重要的水解酶, 通过分解胚乳中的营养物质来为萌发中的胚提供能量和营养.Kondhare等(2014)使用外源激素处理小麦(Triticum aestivum)和水稻种子时, 发现ABA抑制而GA促进α-淀粉酶的活性, 可见ABA/GA平衡还能调控种子内部营养物质的改变. ...
3 2006
... ABA在促进种子成熟及诱导和维持种子休眠中发挥重要作用.在种子发育过程中, ABA对种子休眠的诱导和维持与种子内源ABA水平及种胚发育信号密切相关(Holdsworth et al., 2008; Shu et al., 2016).在种子发育过程中, 内源ABA水平的降低导致成熟种子的休眠能力下降甚至丧失, 而ABA含量的增加则会使种子休眠能力增强及萌发推迟(Okamoto et al., 2006; Nonogaki et al., 2014). ... ... 在ABA代谢方面, AtCYP707A1主要在种子成熟中期的种皮和胚乳中表达, 导致ABA水平下降, 突变体cyp707a1种子表现出较野生型更强的休眠能力.AtCYP707A2主要在种子成熟晚期的种皮、胚乳和胚中以及吸胀的种子中表达; cyp707a2干种子ABA含量较野生型高5倍, 呈深度休眠表型; 吸胀后AtCYP- 707A2表达量显著增强, 表明CYP707A2是种子萌发过程中的主效基因.AtCYP707A3主要在胚以及成苗莲座叶中表达, 受种子萌发和干旱胁迫的诱导, 但cyp707a3突变体种子休眠程度不如cyp707a1和cyp707a2变化显著, 干种子ABA含量与野生型相近, 说明CYP707A3可能并不调控种子的发育, 而是调控萌发后幼苗生长及胁迫应答过程中ABA的积累.At- CYP707A4主要在种子发育中后期的角果皮中表达, 而在种子中无表达活性, 暗示AtCYP707A4并不调控种子发育过程中的ABA积累(Kushiro et al., 2004; Okamoto et al., 2006).双突变体表型分析显示, cyp707a1/cyp707a2种子休眠程度高于cyp707a2/ cyp707a3, 而后者又高于cyp707a1/cyp707a3 (Ok- amoto et al., 2006). ... ... (Ok- amoto et al., 2006). ...
1 2010
... 种子休眠的维持可能与染色质结构相关, 促进种子萌发的基因由于空间位阻不能被转录因子结合而处于静止状态(Shu et al., 2016).拟南芥KYP/SUVH4编码一种H3 Lys 9甲基转移酶, 为H3K9甲基化所需; 同时KYP/SUVH4负调控种子休眠, 其缺失突变体休眠程度增强.研究表明, ABA可能通过抑制KYP/SUVH4的表达从而促进种子休眠(Zheng et al., 2012).组蛋白乙酰转移酶(histone acetyltransferases)通过调节染色质活力来调控基因表达.大麦(Hordeum vulgare)种子经外源ABA处理后, 组蛋白乙酰转移酶基因HvGNAT/MYST表达上调(Papaefthimiou et al., 2010).表观遗传调控因子HUB1 (HISTONE MON- OU-BIQUITINATION 1 )和RDO2 (REDUCED DOR- MANCY 2)在休眠诱导期受到ABA的强烈诱导(Liu et al., 2011).此外, 组蛋白脱甲基酶LDL1 (LYSINES- PECIFIC DEMETHYLASE LIKE 1)和LDL2通过抑制DOG1、ABA2和ABI3的表达, 负调控种子休眠(Zhao et al., 2015).KYP/SUVH4和SUVH5在种子成熟期抑制DOG1和ABI3的表达(Zheng et al., 2012), 且DO- G1的染色质结构重塑参与种子的休眠循环过程(Foo- titt et al., 2015), 表明DOG1可能通过2条不同的途 径诱导种子休眠.关于ABA通过影响染色质结构重塑调节种子休眠与萌发的分子机制, 目前尚不完全明确. ...
3 2009
... ABA通过特定的信号转导途径抑制种子萌发.ABA信号转导起始于ABA受体对ABA信号的感知.2009年, 2个实验室各自独立发现1个含有START (STeroidogenic Acute Regulatory Related Lipid Transfer)特征区的蛋白质PYR (PYRABACTIN RESISTANCE)/ PYL (PYR1-LIKE)/RCAR (REGULATORY COMPO- NENTS OF ABA RECEPTORS)家族成员能够结合ABA, 是一种细胞内ABA受体.其作用机理是PYR/ PYL/RCAR通过与下游信号调节子PP2C直接相互作用, 抑制PP2C的活性, 解除PP2C对其下游信号调节子SNF1相关蛋白激酶2 (SNF1-related protein kinase 2, SnRK2)的抑制, 从而使SnRK2能够通过磷酸化激活下游转录因子ABI5/ABF4 (ABRE binding factors), 实现ABA生理效应(图1) (Ma et al., 2009; Park et al., 2009; Hubbard et al., 2010; Hauser et al., 2011). ... ... 拟南芥pyr1/pyl1/pyl2/prl4四突变体种子萌发表现出对ABA高度不敏感(Park et al., 2009).PP2C是一类丝氨酸/苏氨酸残基蛋白磷酸酶, 包括ABI1 (ABA INSENSITIVE 1)、ABI2、HAB1 (HOMOLOGY TO ABA INSENSITIVE 1)、HAB2和AHG3/PP2CA (ABA-HYPERSENSITIVE GERMINATION 3), 在ABA信号转导中起着关键的负调控作用(Hauser et al., 2011).ABI1和ABI2的显性负突变体abi1-1和abi2-1由于蛋白质结构的改变而不能传递ABA信号, 呈现出种子休眠水平下降和对外源ABA不敏感表型(Ma et al., 2009; Park et al., 2009).hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004).AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006).SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014).其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程.双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007).三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009).RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ... ... ; Park et al., 2009).hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004).AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006).SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014).其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程.双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007).三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009).RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ...
1 2003
... B3型转录因子家族的VAL (VIVIPAROUS1/ ABI3-LIKE)支线VAL1/HSI2 (HIGH-LEVEL EXPRESSION OF SUGAR-INDUCIBLE GENE 2)、VAL2/HSL1 (HSI2-LIKE1)和VAL3/HSL2 (HSI2- LIKE2)参与对LAFL的转录抑制(Suzuki et al., 2007).VAL蛋白除含有B3结构域外, 还具有与染色质因子相关的PHD-L (plant homeodomain-like)和CW (cys- and trp-containing domain)结构域, 以及EAR (ethy- lene response factor-associated repression)转录抑制结构域(Perry and Zhao, 2003; Bienz, 2006).在种子萌发过程中, VAL蛋白通过CW结构域, 将组蛋白脱乙酰酶复合物(histone deacetylase complex, HDAC)招募至含有Sph/RY元件的LAFL基因上, 并抑制它们的表达, 从而正调控种子萌发及幼苗生长.有趣的是, 已有研究显示VAL1是FUS3的直接靶标(Wang and Perry, 2013). ...
1 2008
... 转录组分析显示, 拟南芥干种子中含有种类丰富的mRNA, 包括种子发育晚期丰富蛋白(late embryogenesis abundant, LEA)和其它贮藏蛋白基因的mRNA.绝大多数高丰度mRNA编码基因的启动子中含有ABA应答元件(ABA response element, ABRE).拟南芥bZIP型转录因子具有亮氨酸拉链结构(leucine zipper), 能特异识别基因启动子ABRE应答元件中的ACGT核心序列, 调控相应基因的表达.ABI5作为bZIP型转录因子家族中的重要成员, 能够激活种胚内ABA调控基因的表达, 如LEA蛋白的编码基因EM1 (EARLY METHIONINE-LABELED 1)、EM6和LEAD- 34 (Finkelstein and Lynch, 2000).酵母单杂交实验结果显示, ABI5直接与EM6启动子的ABRE结合从而促进其表达(Nakamura et al., 2001; Carles et al., 2002).种子休眠的关键转录因子DOG1是通过促进ABI5转录, 从而激活下游LEA和HSP (HEAT SHO- CK PROTEIN)基因的表达(Dekkers et al., 2016).ABI5不直接参与种子休眠调控, 但负调控种子的萌发, 说明ABA调控种子休眠和萌发是通过2个不同的信号转导途径(Piskurewicz et al., 2008; Kanai et al., 2010).ABI5能与ABI3进行体外蛋白互作, 且位于ABI3的下游, 激活下游基因表达(Nakamura et al., 2001; Lopez-Molina et al., 2002).此外, 拟南芥转录因子bZIP53通过结合种子贮藏蛋白基因(如SSP和LEA)启动子G-box元件, 促进相关基因表达, 进而调控种子成熟.体外结合实验显示, 当bZIP53与bZIP10或bZIP25异源二聚化后, 能增强bZIP53与G-box的结合活性(Alonso et al., 2009).同时, bZIP10和bZIP25也能促进SSP基因的表达(Lara et al., 2003), 表明这3个转录因子在促进种子成熟过程中具有协同作用. ...
1 2009
... 种子休眠的维持取决于种子内源ABA与GA含量的比值(ABA/GA), 而休眠解除是由种子内源ABA含量下降和GA含量升高所致(Cadman et al., 2006).拟南芥休眠型Cvi (cape verde islands)和非休眠型Col (Columbia)种子吸胀2-3小时后, CYP707A2转录均被激活, 导致ABA含量迅速下降, 说明CYP707A2是种子吸胀过程中ABA降解的主要代谢酶(Preston et al., 2009).研究表明, 休眠型Cvi种子后熟作用的休眠释放(dormancy release)与CYP707A2和GA3ox1基因表达相关.cyp707a2缺失突变体种子中GA4含量和GA3ox1的转录水平均有所下降, 表明种子萌发时ABA通过负调控GA合成来抑制种子萌发(Yano et al., 2009).同样, GA也能够抑制ABA的合成途径, ABA与GA的拮抗作用主要受AP2型转录因子的调控(Oh et al., 2007; Shu et al., 2013).种子吸胀过程中, GA合成基因GA3ox和GA20ox高表达, 促使GA在种胚内大量积累(Ogawa et al., 2003).种子萌发早期, ABA/GA比值下降至原有水平的1/3; 而在萌发后期, ABA/GA比值下降至1/10 (Weitbrecht et al., 2011).淀粉酶作为种子萌发过程中重要的水解酶, 通过分解胚乳中的营养物质来为萌发中的胚提供能量和营养.Kondhare等(2014)使用外源激素处理小麦(Triticum aestivum)和水稻种子时, 发现ABA抑制而GA促进α-淀粉酶的活性, 可见ABA/GA平衡还能调控种子内部营养物质的改变. ...
1 2017
... 植物特有的转录因子DOF蛋白(DNA BINDING WITH ONE FINGER)具有1个锌指型DNA结合域, 参与调控植物不同的生理过程.转录因子DAG1 (DOF AFFECTING GERMINATION 1)在种子成熟过程中表达, 影响种子胚胎发育.dag1缺失突变体种子休眠能力减弱, 且胚胎畸形(Boccaccini et al., 2014).DAG1受表观遗传调控, 直接与GA3ox1 (GA3 oxidase 1)和CYP707A2的启动子结合, 抑制其转录, 从而抑制GA并促进ABA积累, 调控种子休眠.突变体dag1种子内源ABA含量降低, 而GA含量升高(Bocc- accini et al., 2016).在种子成熟和休眠过程中, DAG1通过结合DAG2的启动子抑制DAG2的表达, 但dag2突变体种胚表型与野生型无差异, 表明DAG2并不影响胚胎发育和种子休眠.进一步研究表明, DAG2在PIL5 (PHYTOCHROME INTERACTING FACTOR3- LIKE 5)的下游, 正调控光促进的种子萌发过程(Santopolo et al., 2015).最新研究表明, DOF6蛋白能与RGL2 (RGA-LIKE2) (GA信号途径的负调控蛋白)构成复合体, 共同结合在RGL2下游靶标GATA12的DOF相关基序上, 激活其表达, 从而促进种子初级休眠的维持; DOF6表达上调株系种子的休眠能力显著增强(Ravindran et al., 2017). ...
1 2001
... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ... ... ; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ... ... 种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
1 1995
... 研究表明, 拟南芥ABAR/CHLH过表达株系表现出对ABA的高度敏感, 而突变体abar-2和ABAR RN- Ai株系的种子萌发对ABA不敏感.上述结果与ABAR作为ABA受体的结论相符.WRKY40是细胞内关键的ABA信号负调控因子, 在细胞核中通过抑制ABI5的表达而抑制下游的ABA应答反应.WRKY40缺失突变体种子萌发表型对ABA高度敏感(Jiang and Yu, 2009; Shang et al., 2010).拟南芥种子萌发及之后早期幼苗生长发育阶段, ABA诱导WRKY2的积累需要ABI3、ABI5、ABA2和ABA3的存在.wrky2缺失突变体种子萌发及幼苗生长对ABA超敏感, 说明WRKY2也负调控ABA抑制的种子萌发和幼苗早期生长发育(Jiang and Yu, 2009).另外, 早期研究表明, 野燕麦(Avena fatua) WRKY蛋白ABF1和ABF2能结合在α-淀粉酶编码基因启动子的box2/W-box元件上, 促进胚乳中淀粉的水解, 从而间接促进种子萌发(Rush- ton et al., 1995). ...
1 2004
... 拟南芥pyr1/pyl1/pyl2/prl4四突变体种子萌发表现出对ABA高度不敏感(Park et al., 2009).PP2C是一类丝氨酸/苏氨酸残基蛋白磷酸酶, 包括ABI1 (ABA INSENSITIVE 1)、ABI2、HAB1 (HOMOLOGY TO ABA INSENSITIVE 1)、HAB2和AHG3/PP2CA (ABA-HYPERSENSITIVE GERMINATION 3), 在ABA信号转导中起着关键的负调控作用(Hauser et al., 2011).ABI1和ABI2的显性负突变体abi1-1和abi2-1由于蛋白质结构的改变而不能传递ABA信号, 呈现出种子休眠水平下降和对外源ABA不敏感表型(Ma et al., 2009; Park et al., 2009).hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004).AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006).SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014).其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程.双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007).三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009).RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ...
1 2015
... 植物特有的转录因子DOF蛋白(DNA BINDING WITH ONE FINGER)具有1个锌指型DNA结合域, 参与调控植物不同的生理过程.转录因子DAG1 (DOF AFFECTING GERMINATION 1)在种子成熟过程中表达, 影响种子胚胎发育.dag1缺失突变体种子休眠能力减弱, 且胚胎畸形(Boccaccini et al., 2014).DAG1受表观遗传调控, 直接与GA3ox1 (GA3 oxidase 1)和CYP707A2的启动子结合, 抑制其转录, 从而抑制GA并促进ABA积累, 调控种子休眠.突变体dag1种子内源ABA含量降低, 而GA含量升高(Bocc- accini et al., 2016).在种子成熟和休眠过程中, DAG1通过结合DAG2的启动子抑制DAG2的表达, 但dag2突变体种胚表型与野生型无差异, 表明DAG2并不影响胚胎发育和种子休眠.进一步研究表明, DAG2在PIL5 (PHYTOCHROME INTERACTING FACTOR3- LIKE 5)的下游, 正调控光促进的种子萌发过程(Santopolo et al., 2015).最新研究表明, DOF6蛋白能与RGL2 (RGA-LIKE2) (GA信号途径的负调控蛋白)构成复合体, 共同结合在RGL2下游靶标GATA12的DOF相关基序上, 激活其表达, 从而促进种子初级休眠的维持; DOF6表达上调株系种子的休眠能力显著增强(Ravindran et al., 2017). ...
2 2010
... 清华大学张大鹏研究组发现, 定位于叶绿体被膜上的蛋白质镁螯合酶H亚基(Mg2+-chelatase H subunit, CHLH)能够结合ABA, 他们提出1条CHLH/ ABAR (ABA receptor)介导的始于叶绿体的解除转录抑制的ABA信号通路.该模型认为在生理状态或低浓度ABA时, WRKY40抑制一组ABA信号正调节基因(ABF4、ABI4和ABI5)的表达; 高浓度ABA时, ABAR结合ABA后, 刺激WRKY40从细胞核转移到细胞质, 促进ABAR与WRKY40的相互作用, 进而激发一种未知因子(或信号系统), 阻遏WRKY40的表达, 从而解除WRKY40对ABA响应基因转录的抑制, 最终实现ABA的生理效应(Shen et al., 2006; Shang et al., 2010; 张大鹏, 2011). ... ... 研究表明, 拟南芥ABAR/CHLH过表达株系表现出对ABA的高度敏感, 而突变体abar-2和ABAR RN- Ai株系的种子萌发对ABA不敏感.上述结果与ABAR作为ABA受体的结论相符.WRKY40是细胞内关键的ABA信号负调控因子, 在细胞核中通过抑制ABI5的表达而抑制下游的ABA应答反应.WRKY40缺失突变体种子萌发表型对ABA高度敏感(Jiang and Yu, 2009; Shang et al., 2010).拟南芥种子萌发及之后早期幼苗生长发育阶段, ABA诱导WRKY2的积累需要ABI3、ABI5、ABA2和ABA3的存在.wrky2缺失突变体种子萌发及幼苗生长对ABA超敏感, 说明WRKY2也负调控ABA抑制的种子萌发和幼苗早期生长发育(Jiang and Yu, 2009).另外, 早期研究表明, 野燕麦(Avena fatua) WRKY蛋白ABF1和ABF2能结合在α-淀粉酶编码基因启动子的box2/W-box元件上, 促进胚乳中淀粉的水解, 从而间接促进种子萌发(Rush- ton et al., 1995). ...
1 2006
... 清华大学张大鹏研究组发现, 定位于叶绿体被膜上的蛋白质镁螯合酶H亚基(Mg2+-chelatase H subunit, CHLH)能够结合ABA, 他们提出1条CHLH/ ABAR (ABA receptor)介导的始于叶绿体的解除转录抑制的ABA信号通路.该模型认为在生理状态或低浓度ABA时, WRKY40抑制一组ABA信号正调节基因(ABF4、ABI4和ABI5)的表达; 高浓度ABA时, ABAR结合ABA后, 刺激WRKY40从细胞核转移到细胞质, 促进ABAR与WRKY40的相互作用, 进而激发一种未知因子(或信号系统), 阻遏WRKY40的表达, 从而解除WRKY40对ABA响应基因转录的抑制, 最终实现ABA的生理效应(Shen et al., 2006; Shang et al., 2010; 张大鹏, 2011). ...
3 2016
... 种子休眠和萌发是植物生长发育过程中2个紧密相连的阶段, 受到种子发育的内源因素和外界环境因素共同调控.Finch-Savage和Leubner-Metzger (2006)对种子休眠和萌发进行了定义: 休眠是指具有活力的种子在适宜的环境条件下不能萌发的状态; 而萌发则是种子内胚根突破外周的组织结构(胚乳和种皮)而展露在种皮之外的过程.脱落酸(abscisic acid, ABA)是调控种子休眠和萌发过程最重要的植物激素, 具有诱导和维持种子休眠、抑制种子萌发及幼苗生长发育的作用.十几年来, ABA调控种子休眠和萌发的研究取得了重大进展(Holdsworth et al., 2008; Weitbrecht et al., 2011; Shu et al., 2016).刘一灵等(2014)从ABA代谢与种子发育、穗萌和种子萌发等方面进行了综述.操春燕等(2017)则从ABA与细胞分裂素(cytokinin, CK)和赤霉素(gibberellin, GA)等相互作用调控种子萌发的角度概述了相关的最新研究进展.近年来, 随着参与种子ABA代谢和ABA信号基因相关突变体研究的深入, ABA调控种子休眠和萌发的分子机制已逐渐明晰, 本文参考ABA代谢基因、种子成熟基因以及ABA信号基因的遗传功能和表达调控的最新研究成果(表1), 综述ABA调控种子休眠和萌发的分子机制, 以期为植物激素的作用机理研究和农作物分子设计育种提供重要的理论依据. ... ... ABA在促进种子成熟及诱导和维持种子休眠中发挥重要作用.在种子发育过程中, ABA对种子休眠的诱导和维持与种子内源ABA水平及种胚发育信号密切相关(Holdsworth et al., 2008; Shu et al., 2016).在种子发育过程中, 内源ABA水平的降低导致成熟种子的休眠能力下降甚至丧失, 而ABA含量的增加则会使种子休眠能力增强及萌发推迟(Okamoto et al., 2006; Nonogaki et al., 2014). ... ... 种子休眠的维持可能与染色质结构相关, 促进种子萌发的基因由于空间位阻不能被转录因子结合而处于静止状态(Shu et al., 2016).拟南芥KYP/SUVH4编码一种H3 Lys 9甲基转移酶, 为H3K9甲基化所需; 同时KYP/SUVH4负调控种子休眠, 其缺失突变体休眠程度增强.研究表明, ABA可能通过抑制KYP/SUVH4的表达从而促进种子休眠(Zheng et al., 2012).组蛋白乙酰转移酶(histone acetyltransferases)通过调节染色质活力来调控基因表达.大麦(Hordeum vulgare)种子经外源ABA处理后, 组蛋白乙酰转移酶基因HvGNAT/MYST表达上调(Papaefthimiou et al., 2010).表观遗传调控因子HUB1 (HISTONE MON- OU-BIQUITINATION 1 )和RDO2 (REDUCED DOR- MANCY 2)在休眠诱导期受到ABA的强烈诱导(Liu et al., 2011).此外, 组蛋白脱甲基酶LDL1 (LYSINES- PECIFIC DEMETHYLASE LIKE 1)和LDL2通过抑制DOG1、ABA2和ABI3的表达, 负调控种子休眠(Zhao et al., 2015).KYP/SUVH4和SUVH5在种子成熟期抑制DOG1和ABI3的表达(Zheng et al., 2012), 且DO- G1的染色质结构重塑参与种子的休眠循环过程(Foo- titt et al., 2015), 表明DOG1可能通过2条不同的途 径诱导种子休眠.关于ABA通过影响染色质结构重塑调节种子休眠与萌发的分子机制, 目前尚不完全明确. ...
2 2013
... APETALA 2/乙烯应答元件结合因子(AP2/ERF)是植物特有的转录因子家族, 调控植物生长发育及胁迫响应.目前已证实在拟南芥和水稻(Oryza sativa)中, AP2型转录因子通过调节ABA/GA的平衡来调控种子的休眠和萌发.拟南芥ABI4通过结合CYP707A1和CYP707A2基因启动子的CCAC顺式元件抑制其表达, 同时直接促进NCED6和GA2ox7 (GA2 oxidase 7)基因的表达, 且ABI4缺失突变体能恢复ga1-t、cyp707a1和cyp707a2突变体不萌发的表型, 证明ABI4通过正调控ABA合成和负调控GA合成来调控种子的休眠与萌发(Shu et al., 2013, 2018).ABI4基因的转录受到控制种子休眠的关键转录因子SPT (SPATULA)的调控, 提示SPT-ABI4在种子休眠的诱导和维持方面具有重要作用(Vaistij et al., 2013).拟南芥CHO1 (CHOTTO1)具有双重的AP2结构, CHO1在ABI4下游, 正调控种子休眠, 通过促进ABA1/ZEP和NCED9的表达, 抑制CYP707A2和CYP707A3的表达, 调控种子萌发过程中ABA介导的GA合成抑制过程(Yano et al., 2009).拟南芥ORA47 (octadeca- noid-responsive AP2/ERF-domain transcription fac- tor 47)和DREB2C通过正调控NCED3和NCED9的表达, 促进ABA合成, 从而对非生物胁迫作出响应(Je et al., 2014; Chen et al., 2016).此外, 转录因子MYB96通过调控ABI4和ABA合成基因NCED2与NCED6的表达, 促进种子休眠而抑制萌发(Lee et al., 2015a, 2015b). ... ... 种子休眠的维持取决于种子内源ABA与GA含量的比值(ABA/GA), 而休眠解除是由种子内源ABA含量下降和GA含量升高所致(Cadman et al., 2006).拟南芥休眠型Cvi (cape verde islands)和非休眠型Col (Columbia)种子吸胀2-3小时后, CYP707A2转录均被激活, 导致ABA含量迅速下降, 说明CYP707A2是种子吸胀过程中ABA降解的主要代谢酶(Preston et al., 2009).研究表明, 休眠型Cvi种子后熟作用的休眠释放(dormancy release)与CYP707A2和GA3ox1基因表达相关.cyp707a2缺失突变体种子中GA4含量和GA3ox1的转录水平均有所下降, 表明种子萌发时ABA通过负调控GA合成来抑制种子萌发(Yano et al., 2009).同样, GA也能够抑制ABA的合成途径, ABA与GA的拮抗作用主要受AP2型转录因子的调控(Oh et al., 2007; Shu et al., 2013).种子吸胀过程中, GA合成基因GA3ox和GA20ox高表达, 促使GA在种胚内大量积累(Ogawa et al., 2003).种子萌发早期, ABA/GA比值下降至原有水平的1/3; 而在萌发后期, ABA/GA比值下降至1/10 (Weitbrecht et al., 2011).淀粉酶作为种子萌发过程中重要的水解酶, 通过分解胚乳中的营养物质来为萌发中的胚提供能量和营养.Kondhare等(2014)使用外源激素处理小麦(Triticum aestivum)和水稻种子时, 发现ABA抑制而GA促进α-淀粉酶的活性, 可见ABA/GA平衡还能调控种子内部营养物质的改变. ...
1 2018
... APETALA 2/乙烯应答元件结合因子(AP2/ERF)是植物特有的转录因子家族, 调控植物生长发育及胁迫响应.目前已证实在拟南芥和水稻(Oryza sativa)中, AP2型转录因子通过调节ABA/GA的平衡来调控种子的休眠和萌发.拟南芥ABI4通过结合CYP707A1和CYP707A2基因启动子的CCAC顺式元件抑制其表达, 同时直接促进NCED6和GA2ox7 (GA2 oxidase 7)基因的表达, 且ABI4缺失突变体能恢复ga1-t、cyp707a1和cyp707a2突变体不萌发的表型, 证明ABI4通过正调控ABA合成和负调控GA合成来调控种子的休眠与萌发(Shu et al., 2013, 2018).ABI4基因的转录受到控制种子休眠的关键转录因子SPT (SPATULA)的调控, 提示SPT-ABI4在种子休眠的诱导和维持方面具有重要作用(Vaistij et al., 2013).拟南芥CHO1 (CHOTTO1)具有双重的AP2结构, CHO1在ABI4下游, 正调控种子休眠, 通过促进ABA1/ZEP和NCED9的表达, 抑制CYP707A2和CYP707A3的表达, 调控种子萌发过程中ABA介导的GA合成抑制过程(Yano et al., 2009).拟南芥ORA47 (octadeca- noid-responsive AP2/ERF-domain transcription fac- tor 47)和DREB2C通过正调控NCED3和NCED9的表达, 促进ABA合成, 从而对非生物胁迫作出响应(Je et al., 2014; Chen et al., 2016).此外, 转录因子MYB96通过调控ABI4和ABA合成基因NCED2与NCED6的表达, 促进种子休眠而抑制萌发(Lee et al., 2015a, 2015b). ...
1 2008
... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
1 2007
... B3型转录因子家族的VAL (VIVIPAROUS1/ ABI3-LIKE)支线VAL1/HSI2 (HIGH-LEVEL EXPRESSION OF SUGAR-INDUCIBLE GENE 2)、VAL2/HSL1 (HSI2-LIKE1)和VAL3/HSL2 (HSI2- LIKE2)参与对LAFL的转录抑制(Suzuki et al., 2007).VAL蛋白除含有B3结构域外, 还具有与染色质因子相关的PHD-L (plant homeodomain-like)和CW (cys- and trp-containing domain)结构域, 以及EAR (ethy- lene response factor-associated repression)转录抑制结构域(Perry and Zhao, 2003; Bienz, 2006).在种子萌发过程中, VAL蛋白通过CW结构域, 将组蛋白脱乙酰酶复合物(histone deacetylase complex, HDAC)招募至含有Sph/RY元件的LAFL基因上, 并抑制它们的表达, 从而正调控种子萌发及幼苗生长.有趣的是, 已有研究显示VAL1是FUS3的直接靶标(Wang and Perry, 2013). ...
1 2008
... 拟南芥中, 调控种胚发育和种子成熟的转录因子主要由B3型转录因子的AFL支线ABI3 (ABA INSENSITIVE 3)、 FUS3 (FUSCA 3)和LEC2 (LEAFY COTY- LEDON 2), 以及2个LEC1型NF-YB (NUCLEAR FA- CTOR Y B subunit)家族成员LEC1和L1L (LEC1- LIKE)组成, 统称为LAFL.B3型转录因子最先是从玉米(Zea mays) vp1 (viviparous1)和拟南芥abi3 (abscisic acid insensitive 3)突变体中被发现, 属于植物特有的B3型DNA结合结构域蛋白家族.该家族蛋白能识别种子特异基因启动子的RY重复或Sph元件的保守序列, 具有促进种胚发育和成熟的功能(Jia et al., 2014).其中, ABI3是种子休眠与萌发的主要调控因子, 具有调控种皮色素改变、诱导种子休眠和促进贮藏蛋白积累等功能(Dekkers et al., 2016).在ABI3上游, Barrero等(2010)发现1个RING-finger蛋白(一类E3泛素连接酶)的编码基因DEP (DESPIERTO), 在种子发育期间调控ABI3表达并影响休眠的诱导, 突变体dep-1种子休眠完全丧失.FUS3在种子发育中后期增强NCED6、NCED9和ABA2的表达, 促进种子干旱耐受性的获得, 并决定子叶上皮细胞的一致性(Tiedemann et al., 2008; Yamamoto et al., 2010).LEC2在胚胎发育早中期表达, 促进贮藏蛋白和油体蛋白的积累, 并参与种子成熟阶段的基因表达调控, 如DOG1 (DELAY OF GERMINATION 1) (Braybr- ook et al., 2006).拟南芥DOG1蛋白是种子休眠的关键调控因子, 通过抑制特定蛋白磷酸酶2C (protein phosphatase 2Cs, PP2C)活性来调控休眠, 且其蛋白水平决定了休眠释放的时机.dog1突变体种子休眠完全缺失(Née et al., 2017).LEC1型NF-YB家族转录因子具有结合CCAAT-box的结构域, 通过调控AFL基因促进种子贮藏蛋白(seed storage protein, SSP)基因的表达(Kagaya et al., 2005).LEC1在胚胎发育早期表达并促进种子胚胎形成(Lotan et al., 1998).L1L在发育中的胚胎(稍晚于LEC1)以及营养器官中表达, 表明除正向调控种胚发育外, L1L还具有与LEC1不同的调控功能(Kwong et al., 2003). ...
2 2006
... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ... ... 的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
1 2013
... APETALA 2/乙烯应答元件结合因子(AP2/ERF)是植物特有的转录因子家族, 调控植物生长发育及胁迫响应.目前已证实在拟南芥和水稻(Oryza sativa)中, AP2型转录因子通过调节ABA/GA的平衡来调控种子的休眠和萌发.拟南芥ABI4通过结合CYP707A1和CYP707A2基因启动子的CCAC顺式元件抑制其表达, 同时直接促进NCED6和GA2ox7 (GA2 oxidase 7)基因的表达, 且ABI4缺失突变体能恢复ga1-t、cyp707a1和cyp707a2突变体不萌发的表型, 证明ABI4通过正调控ABA合成和负调控GA合成来调控种子的休眠与萌发(Shu et al., 2013, 2018).ABI4基因的转录受到控制种子休眠的关键转录因子SPT (SPATULA)的调控, 提示SPT-ABI4在种子休眠的诱导和维持方面具有重要作用(Vaistij et al., 2013).拟南芥CHO1 (CHOTTO1)具有双重的AP2结构, CHO1在ABI4下游, 正调控种子休眠, 通过促进ABA1/ZEP和NCED9的表达, 抑制CYP707A2和CYP707A3的表达, 调控种子萌发过程中ABA介导的GA合成抑制过程(Yano et al., 2009).拟南芥ORA47 (octadeca- noid-responsive AP2/ERF-domain transcription fac- tor 47)和DREB2C通过正调控NCED3和NCED9的表达, 促进ABA合成, 从而对非生物胁迫作出响应(Je et al., 2014; Chen et al., 2016).此外, 转录因子MYB96通过调控ABI4和ABA合成基因NCED2与NCED6的表达, 促进种子休眠而抑制萌发(Lee et al., 2015a, 2015b). ...
... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ... ... B3型转录因子家族的VAL (VIVIPAROUS1/ ABI3-LIKE)支线VAL1/HSI2 (HIGH-LEVEL EXPRESSION OF SUGAR-INDUCIBLE GENE 2)、VAL2/HSL1 (HSI2-LIKE1)和VAL3/HSL2 (HSI2- LIKE2)参与对LAFL的转录抑制(Suzuki et al., 2007).VAL蛋白除含有B3结构域外, 还具有与染色质因子相关的PHD-L (plant homeodomain-like)和CW (cys- and trp-containing domain)结构域, 以及EAR (ethy- lene response factor-associated repression)转录抑制结构域(Perry and Zhao, 2003; Bienz, 2006).在种子萌发过程中, VAL蛋白通过CW结构域, 将组蛋白脱乙酰酶复合物(histone deacetylase complex, HDAC)招募至含有Sph/RY元件的LAFL基因上, 并抑制它们的表达, 从而正调控种子萌发及幼苗生长.有趣的是, 已有研究显示VAL1是FUS3的直接靶标(Wang and Perry, 2013). ...
2 2011
... 种子休眠和萌发是植物生长发育过程中2个紧密相连的阶段, 受到种子发育的内源因素和外界环境因素共同调控.Finch-Savage和Leubner-Metzger (2006)对种子休眠和萌发进行了定义: 休眠是指具有活力的种子在适宜的环境条件下不能萌发的状态; 而萌发则是种子内胚根突破外周的组织结构(胚乳和种皮)而展露在种皮之外的过程.脱落酸(abscisic acid, ABA)是调控种子休眠和萌发过程最重要的植物激素, 具有诱导和维持种子休眠、抑制种子萌发及幼苗生长发育的作用.十几年来, ABA调控种子休眠和萌发的研究取得了重大进展(Holdsworth et al., 2008; Weitbrecht et al., 2011; Shu et al., 2016).刘一灵等(2014)从ABA代谢与种子发育、穗萌和种子萌发等方面进行了综述.操春燕等(2017)则从ABA与细胞分裂素(cytokinin, CK)和赤霉素(gibberellin, GA)等相互作用调控种子萌发的角度概述了相关的最新研究进展.近年来, 随着参与种子ABA代谢和ABA信号基因相关突变体研究的深入, ABA调控种子休眠和萌发的分子机制已逐渐明晰, 本文参考ABA代谢基因、种子成熟基因以及ABA信号基因的遗传功能和表达调控的最新研究成果(表1), 综述ABA调控种子休眠和萌发的分子机制, 以期为植物激素的作用机理研究和农作物分子设计育种提供重要的理论依据. ... ... 种子休眠的维持取决于种子内源ABA与GA含量的比值(ABA/GA), 而休眠解除是由种子内源ABA含量下降和GA含量升高所致(Cadman et al., 2006).拟南芥休眠型Cvi (cape verde islands)和非休眠型Col (Columbia)种子吸胀2-3小时后, CYP707A2转录均被激活, 导致ABA含量迅速下降, 说明CYP707A2是种子吸胀过程中ABA降解的主要代谢酶(Preston et al., 2009).研究表明, 休眠型Cvi种子后熟作用的休眠释放(dormancy release)与CYP707A2和GA3ox1基因表达相关.cyp707a2缺失突变体种子中GA4含量和GA3ox1的转录水平均有所下降, 表明种子萌发时ABA通过负调控GA合成来抑制种子萌发(Yano et al., 2009).同样, GA也能够抑制ABA的合成途径, ABA与GA的拮抗作用主要受AP2型转录因子的调控(Oh et al., 2007; Shu et al., 2013).种子吸胀过程中, GA合成基因GA3ox和GA20ox高表达, 促使GA在种胚内大量积累(Ogawa et al., 2003).种子萌发早期, ABA/GA比值下降至原有水平的1/3; 而在萌发后期, ABA/GA比值下降至1/10 (Weitbrecht et al., 2011).淀粉酶作为种子萌发过程中重要的水解酶, 通过分解胚乳中的营养物质来为萌发中的胚提供能量和营养.Kondhare等(2014)使用外源激素处理小麦(Triticum aestivum)和水稻种子时, 发现ABA抑制而GA促进α-淀粉酶的活性, 可见ABA/GA平衡还能调控种子内部营养物质的改变. ...
2 2010
... 拟南芥中, 调控种胚发育和种子成熟的转录因子主要由B3型转录因子的AFL支线ABI3 (ABA INSENSITIVE 3)、 FUS3 (FUSCA 3)和LEC2 (LEAFY COTY- LEDON 2), 以及2个LEC1型NF-YB (NUCLEAR FA- CTOR Y B subunit)家族成员LEC1和L1L (LEC1- LIKE)组成, 统称为LAFL.B3型转录因子最先是从玉米(Zea mays) vp1 (viviparous1)和拟南芥abi3 (abscisic acid insensitive 3)突变体中被发现, 属于植物特有的B3型DNA结合结构域蛋白家族.该家族蛋白能识别种子特异基因启动子的RY重复或Sph元件的保守序列, 具有促进种胚发育和成熟的功能(Jia et al., 2014).其中, ABI3是种子休眠与萌发的主要调控因子, 具有调控种皮色素改变、诱导种子休眠和促进贮藏蛋白积累等功能(Dekkers et al., 2016).在ABI3上游, Barrero等(2010)发现1个RING-finger蛋白(一类E3泛素连接酶)的编码基因DEP (DESPIERTO), 在种子发育期间调控ABI3表达并影响休眠的诱导, 突变体dep-1种子休眠完全丧失.FUS3在种子发育中后期增强NCED6、NCED9和ABA2的表达, 促进种子干旱耐受性的获得, 并决定子叶上皮细胞的一致性(Tiedemann et al., 2008; Yamamoto et al., 2010).LEC2在胚胎发育早中期表达, 促进贮藏蛋白和油体蛋白的积累, 并参与种子成熟阶段的基因表达调控, 如DOG1 (DELAY OF GERMINATION 1) (Braybr- ook et al., 2006).拟南芥DOG1蛋白是种子休眠的关键调控因子, 通过抑制特定蛋白磷酸酶2C (protein phosphatase 2Cs, PP2C)活性来调控休眠, 且其蛋白水平决定了休眠释放的时机.dog1突变体种子休眠完全缺失(Née et al., 2017).LEC1型NF-YB家族转录因子具有结合CCAAT-box的结构域, 通过调控AFL基因促进种子贮藏蛋白(seed storage protein, SSP)基因的表达(Kagaya et al., 2005).LEC1在胚胎发育早期表达并促进种子胚胎形成(Lotan et al., 1998).L1L在发育中的胚胎(稍晚于LEC1)以及营养器官中表达, 表明除正向调控种胚发育外, L1L还具有与LEC1不同的调控功能(Kwong et al., 2003). ... ... LAFL转录因子间通过复杂的相互调节及自我调节作用, 共同促进种胚发育和成熟.研究表明, LEC1能激活ABI3、FUS3和LEC2的表达(Kagaya et al., 2005; To et al., 2006); 营养组织中LEC2的异位表达使LEC1和FUS3表达上调, 提示LEC2和LEC1之间形成了双向正调控机制(Stone et al., 2008).种子发育中后期LEC1和LEC2停止转录, ABI3和FUS3的自身反馈及相互正调控机制使它们能够维持平衡(To et al., 2006).转录组分析显示, L1L受FUS3调控(Yamamoto et al., 2010), 同时L1L也是LEC1潜在的直接靶标(Junker et al., 2012); FUS3是ABI3的靶标(Mönke et al., 2012), 且FUS3能与LEC1、FUS3和ABI3的调控区域进行物理互作(Wang and Perry, 2013).突变体分析显示, 拟南芥fus3、abi3、lec1和lec2的种子均表现休眠能力下降(Raz et al., 2001)及种子贮藏蛋白表达降低(Gutierrez et al., 2007).但它们各自也有一些特殊表型, 如fus3突变体种子内贮藏蛋白和脂肪酸含量显著减少(Roscoe et al., 2015); abi3表现出明显的种子成熟缺陷, 主要表现为绿色种子(叶绿素II降解缺失)、蛋白质无法贮藏(大量蛋白质变性)以及不能获得干旱耐受性(Finkelstein et al., 2002; Roscoe et al., 2015).突变体lec1呈现胚胎缺陷、种子发育异常及子叶长毛状体的表型(Huang et al., 2015).突变体lec2种子主要表现为贮藏蛋白和脂肪酸的积累量减少(Roscoe et al., 2015). ...
2 2009
... APETALA 2/乙烯应答元件结合因子(AP2/ERF)是植物特有的转录因子家族, 调控植物生长发育及胁迫响应.目前已证实在拟南芥和水稻(Oryza sativa)中, AP2型转录因子通过调节ABA/GA的平衡来调控种子的休眠和萌发.拟南芥ABI4通过结合CYP707A1和CYP707A2基因启动子的CCAC顺式元件抑制其表达, 同时直接促进NCED6和GA2ox7 (GA2 oxidase 7)基因的表达, 且ABI4缺失突变体能恢复ga1-t、cyp707a1和cyp707a2突变体不萌发的表型, 证明ABI4通过正调控ABA合成和负调控GA合成来调控种子的休眠与萌发(Shu et al., 2013, 2018).ABI4基因的转录受到控制种子休眠的关键转录因子SPT (SPATULA)的调控, 提示SPT-ABI4在种子休眠的诱导和维持方面具有重要作用(Vaistij et al., 2013).拟南芥CHO1 (CHOTTO1)具有双重的AP2结构, CHO1在ABI4下游, 正调控种子休眠, 通过促进ABA1/ZEP和NCED9的表达, 抑制CYP707A2和CYP707A3的表达, 调控种子萌发过程中ABA介导的GA合成抑制过程(Yano et al., 2009).拟南芥ORA47 (octadeca- noid-responsive AP2/ERF-domain transcription fac- tor 47)和DREB2C通过正调控NCED3和NCED9的表达, 促进ABA合成, 从而对非生物胁迫作出响应(Je et al., 2014; Chen et al., 2016).此外, 转录因子MYB96通过调控ABI4和ABA合成基因NCED2与NCED6的表达, 促进种子休眠而抑制萌发(Lee et al., 2015a, 2015b). ... ... 种子休眠的维持取决于种子内源ABA与GA含量的比值(ABA/GA), 而休眠解除是由种子内源ABA含量下降和GA含量升高所致(Cadman et al., 2006).拟南芥休眠型Cvi (cape verde islands)和非休眠型Col (Columbia)种子吸胀2-3小时后, CYP707A2转录均被激活, 导致ABA含量迅速下降, 说明CYP707A2是种子吸胀过程中ABA降解的主要代谢酶(Preston et al., 2009).研究表明, 休眠型Cvi种子后熟作用的休眠释放(dormancy release)与CYP707A2和GA3ox1基因表达相关.cyp707a2缺失突变体种子中GA4含量和GA3ox1的转录水平均有所下降, 表明种子萌发时ABA通过负调控GA合成来抑制种子萌发(Yano et al., 2009).同样, GA也能够抑制ABA的合成途径, ABA与GA的拮抗作用主要受AP2型转录因子的调控(Oh et al., 2007; Shu et al., 2013).种子吸胀过程中, GA合成基因GA3ox和GA20ox高表达, 促使GA在种胚内大量积累(Ogawa et al., 2003).种子萌发早期, ABA/GA比值下降至原有水平的1/3; 而在萌发后期, ABA/GA比值下降至1/10 (Weitbrecht et al., 2011).淀粉酶作为种子萌发过程中重要的水解酶, 通过分解胚乳中的营养物质来为萌发中的胚提供能量和营养.Kondhare等(2014)使用外源激素处理小麦(Triticum aestivum)和水稻种子时, 发现ABA抑制而GA促进α-淀粉酶的活性, 可见ABA/GA平衡还能调控种子内部营养物质的改变. ...
1 2006
... 拟南芥pyr1/pyl1/pyl2/prl4四突变体种子萌发表现出对ABA高度不敏感(Park et al., 2009).PP2C是一类丝氨酸/苏氨酸残基蛋白磷酸酶, 包括ABI1 (ABA INSENSITIVE 1)、ABI2、HAB1 (HOMOLOGY TO ABA INSENSITIVE 1)、HAB2和AHG3/PP2CA (ABA-HYPERSENSITIVE GERMINATION 3), 在ABA信号转导中起着关键的负调控作用(Hauser et al., 2011).ABI1和ABI2的显性负突变体abi1-1和abi2-1由于蛋白质结构的改变而不能传递ABA信号, 呈现出种子休眠水平下降和对外源ABA不敏感表型(Ma et al., 2009; Park et al., 2009).hab1-1种子萌发对ABA高度敏感, 其萌发受到严重抑制(Saez et al., 2004).AHG3/AtPP2CA缺失突变体ahg3-1种子对ABA的敏感性高于其它成员(Yoshida et al., 2006).SnRK2通过激活ABA信号下游转录因子ABI5、ABF4和RAV1 (Related to ABI3/VP1)的表达正向调控ABA信号(Nakashima et al., 2009; Feng et al., 2014).其中SnRK2.2 (SnRK2D)、SnRK2.3 (SnRK2I)和SnRK2.6 (OPEN STOMATA1/OST1/SnRK2E)参与种子萌发过程中ABA信号转导过程.双突变体snrk2.2/snrk2.3与野生型和单突变体相比, 种子萌发表现出显著的对ABA不敏感(Fujii et al., 2007).三突变体snrk2.2/snrk2.3/snrk2.6种子萌发对ABA高度不敏感, 且三突变体种子中ABI5及其下游的AtEM6和AtEM1表达下调(Fujii and Zhu, 2009).RAV1转录因子属于AP2型转录因子的RAV亚家族, 其表达下调株系种子萌发对ABA敏感性增强, 而过表达株系对ABA高度不敏感; 被SnRK2激活后, RAV1直接结合在ABI3、ABI4和ABI5启动子上并抑制它们的表达, 负调控ABA信号转导(Feng et al., 2014). ...

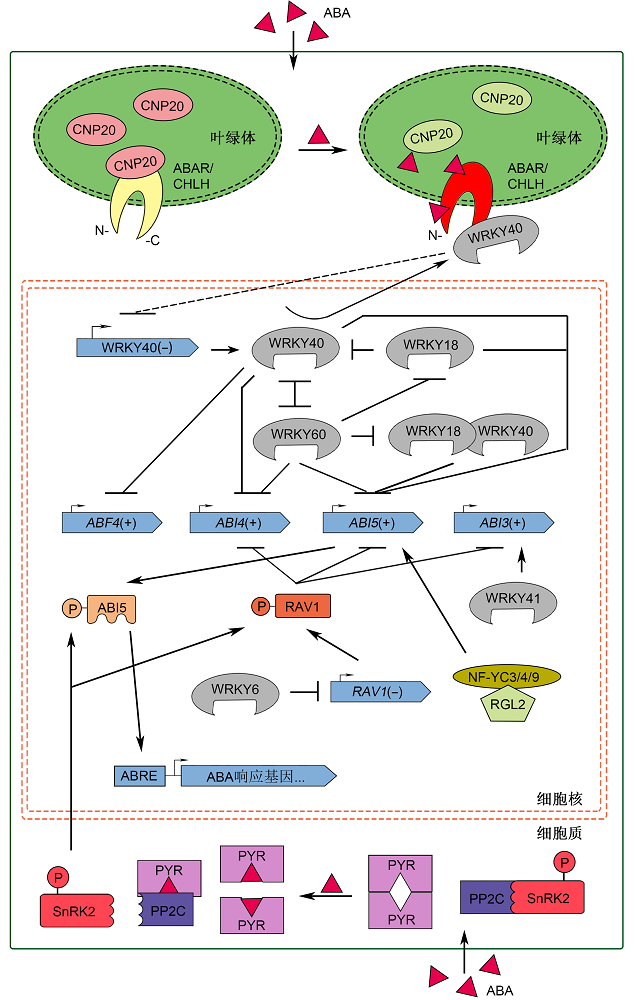

{kind=link}
{kind=link}