Abstract: With the nectarine variety Shuguang as test material and blue light set artificially, the photosynthetic performance and the accumulation of sugar and acid in leaves and fruits were examined, and diurnal changes of the stomata in five key growth periods were observed. Blue light increased the net photosynthetic rate and the contents of chlorophyll a and b. The ratio of chlorophyll a/b was decreased and the leaf area increased. The opening time of the stomata was advanced, and maximum opening was earlier and closing time hysteretic. Fructose, glucose and sorbitol were the main assimilates in leaves, with the highest content being sorbitol. Fructose, glucose, sorbitol and sucrose were the main assimilates in fruit, and sucrose was the main soluble sugar at fruit maturity. As compared with the control, blue light conferred lower content of the three assimilates in leaves, whereas the content of total sugar and sucrose in fruit was higher, which suggests that blue light enhanced the ability to transfer photosynthetic products from leaves to fruit. Oxalic acid was the main organic acid in both leaves and fruits. The content of organic acid under blue light in leaves was decreased, and the sugar to acid ratio was significantly increased, by 30.5%. The key period of peaches improving the sugar to acid ratio under blue light was the late hard nucleus period. Supplementing blue light could be a technical measure to improve photosynthetic performance and fruit quality and could be further applied in the cultivation of greenhouse fruit trees.
1 材料和方法1.1 供试材料实验于2016年2-5月在山东农业大学科技创新园进行。供试材料为6年生曙光油桃(Prunus persica L. var. nectarina Maxim. cv. ‘Shuguang’), 株高2 m, 冠幅1 m×1 m, 株行距为1.5 m×2 m, 常规管理。实验期间设施内昼夜温度范围分别为18-30°C和13- 22°C, 昼夜湿度范围分别为35%-79%和60%-81%。
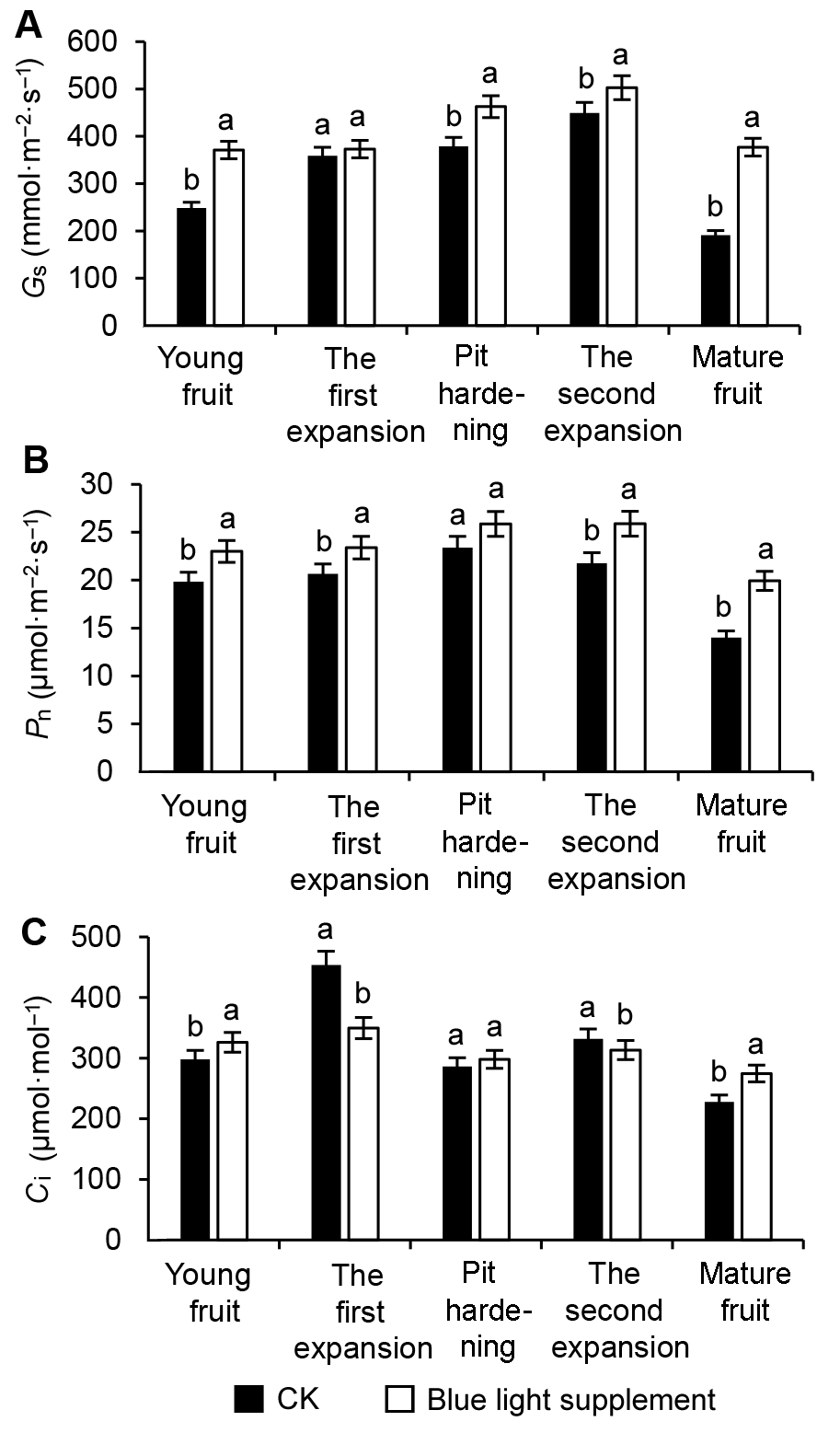
图1 补充蓝光对果实发育时期油桃叶片光合参数的影响 (A) 气孔导度(Gs); (B) 净光合速率(Pn); (C) 胞间CO2浓度(Ci)。图中不同小写字母表示处理间在0.05水平差异显著。 Figure 1 Effects of supplementary blue light on photosynthetic parameters in leaves during peach fruit development (A) Stomatal conductance (Gs); (B) Net photosynthetic rate (Pn); (C) Intercellular CO2 concentration (Ci). Different lowercase letters indicate significant differences at 0.05 level among treatments.
果实生长发育时期, 对照和补充蓝光的油桃叶片的净光合速率(图1B)均呈现前期较为平稳, 成熟期下降的趋势。在果实生长的各个阶段, 补充蓝光处理下油桃叶片的净光合速率均高于对照。对照油桃叶片中Pn在硬核期达到最大值, 为23.4 μmol (CO2)·m-2·s-1; 补充蓝光条件下硬核期和第2次膨大期油桃叶片的净光合速率差异不显著, 约为25.9 μmol (CO2)·m-2·s-1, 较对照提高了11%左右, 补充蓝光处理和对照油桃叶片的净光合速率均在成熟期达到最低值。 胞间CO2浓度的变化方向是确定光合速率变化的主要因素之一, 也是判断是否为气孔限制的必不可少的依据。随着生育期的推进, 对照油桃叶片的胞间CO2浓度(图1C)波动幅度较大, 总体呈下降趋势。补充蓝光的油桃叶片胞间CO2浓度则比较稳定, 基本维持在274.67-349.67 μmol·mol-1之间, 成熟期高于对照, 其余时段则大多低于对照。 2.1.2 对色素含量和叶面积的影响 从表1可以看出, 补充蓝光处理下油桃叶片的叶绿素a、叶绿素b和叶绿素(a+b)含量显著高于对照, 分别提高了14.10%、22.95%和16.17%, 而叶绿素a/b值显著降低, 类胡萝卜素含量与对照相比差异不显著。此外, 补充蓝光能够显著增大油桃的功能叶面积(为对照的1.62倍)。 表1 Table 1 表1 表1 补充蓝光对油桃叶片色素含量和叶面积的影响 Table 1 Effects of supplementary blue light on pigment content and leaf area
Treatment
Chlorophyll a (mg·g-1 FW)
Chlorophyll b (mg·g-1 FW)
Chlorophyll (a+b) (mg·g-1 FW)
Chlorophyll a/b
Carotenoids (mg·g-1 FW)
Leaf area (cm2)
Blue light
2.630*
0.868*
3.498*
3.030
0.491
75.100*
CK
2.305
0.706
3.011
3.265*
0.522
46.458
* 表示相关性达0.05显著水平。* indicate significant correlation at the 0.05 level.
表1 补充蓝光对油桃叶片色素含量和叶面积的影响 Table 1 Effects of supplementary blue light on pigment content and leaf area
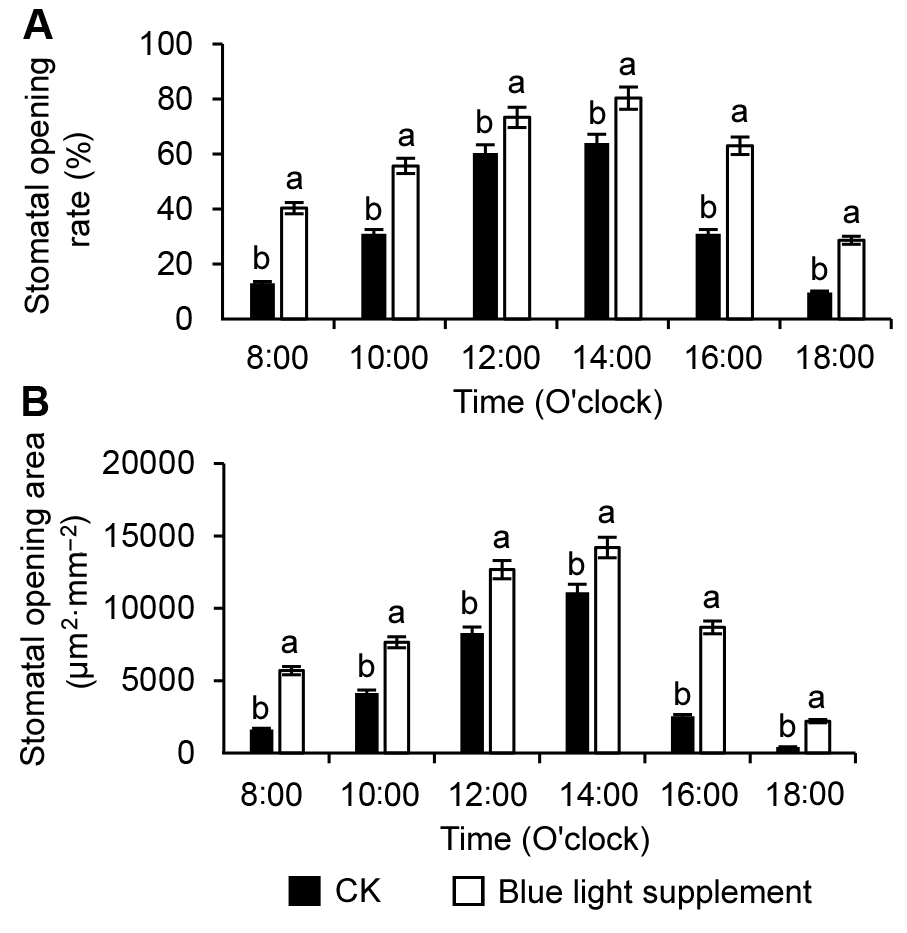
2.2 油桃叶片气孔的日变化由图2和图3可知, 补充蓝光后油桃叶片的气孔开放率和气孔开放面积在一天中的任一时间均显著高于对照。对照和补充蓝光条件下, 油桃叶片的气孔开放率(图2A)和气孔面积(图2B)均呈现先升高后降低的趋势, 并在14:00时达到最大值, 此时, 补充蓝光处理下的气孔开放率为80.33%, 气孔开放面积为1.42× 104 μm2·mm-2, 分别为对照的1.26和1.28倍。18:00时, 补充蓝光处理的油桃叶片仍有28.73%的气孔开放, 对照仅为9.70%, 表明补充蓝光延迟了气孔的关闭。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-227/img_2.png<b>图2</b> 油桃叶片气孔开放率(A)和气孔开放面积(B)的日变化统计分析<br/>图中不同小写字母表示处理间在0.05水平差异显著。<br/><b>Figure 2</b> The statistical analysis of diurnal variation of stomatal opening rate (A) and stomatal opening area (B) of peach leaves<br/>Different lowercase letters indicate significant differences at 0.05 level among treatments. Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-227/img_2.png<b>图2</b> 油桃叶片气孔开放率(A)和气孔开放面积(B)的日变化统计分析<br/>图中不同小写字母表示处理间在0.05水平差异显著。<br/><b>Figure 2</b> The statistical analysis of diurnal variation of stomatal opening rate (A) and stomatal opening area (B) of peach leaves<br/>Different lowercase letters indicate significant differences at 0.05 level among treatments.
图2 油桃叶片气孔开放率(A)和气孔开放面积(B)的日变化统计分析 图中不同小写字母表示处理间在0.05水平差异显著。 Figure 2 The statistical analysis of diurnal variation of stomatal opening rate (A) and stomatal opening area (B) of peach leaves Different lowercase letters indicate significant differences at 0.05 level among treatments.
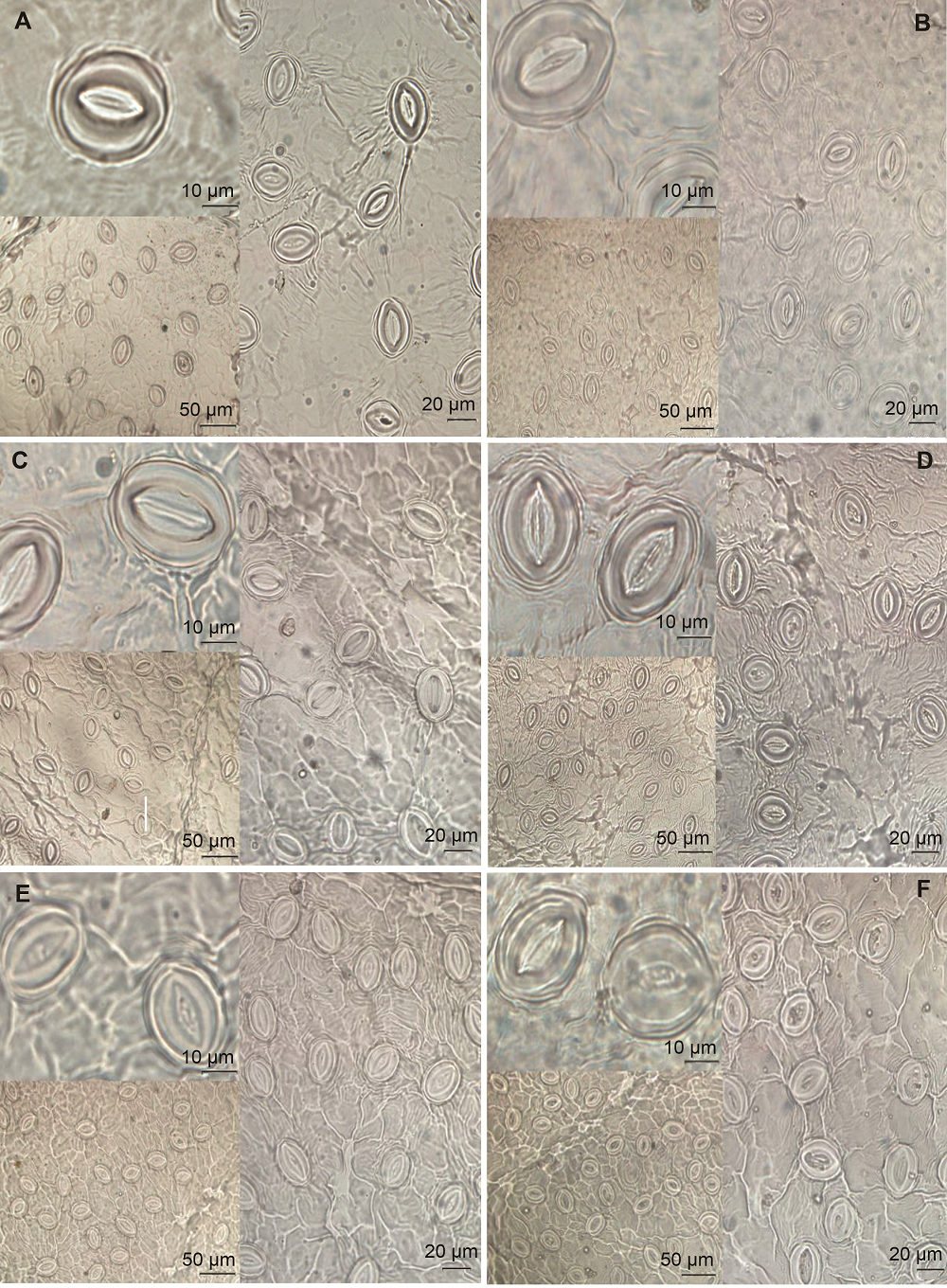
图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-227/img_3.png<b>图3</b> 一天中部分关键时刻油桃叶片的气孔形态观察<br/>(A), (C)和(E) 对照组油桃叶片分别在8:00、14:00和18:00的气孔形态; (B), (D)和(F) 补充蓝光条件下油桃叶片分别在8:00、14:00和18:00的气孔形态。<br/><b>Figure 3</b> Observation of stomatal morphology of peach leaves at some critical moments in a day<br/>(A), (C) and (E) The stomatal morphology in the control of peach leaves at 8:00, 14:00 and 18:00, respectively; (B), (D) and (F) The stomatal morphology in the supplemental blue light of peach leaves at 8:00, 14:00 and 18:00, respectively. Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-227/img_3.png<b>图3</b> 一天中部分关键时刻油桃叶片的气孔形态观察<br/>(A), (C)和(E) 对照组油桃叶片分别在8:00、14:00和18:00的气孔形态; (B), (D)和(F) 补充蓝光条件下油桃叶片分别在8:00、14:00和18:00的气孔形态。<br/><b>Figure 3</b> Observation of stomatal morphology of peach leaves at some critical moments in a day<br/>(A), (C) and (E) The stomatal morphology in the control of peach leaves at 8:00, 14:00 and 18:00, respectively; (B), (D) and (F) The stomatal morphology in the supplemental blue light of peach leaves at 8:00, 14:00 and 18:00, respectively.
图3 一天中部分关键时刻油桃叶片的气孔形态观察 (A), (C)和(E) 对照组油桃叶片分别在8:00、14:00和18:00的气孔形态; (B), (D)和(F) 补充蓝光条件下油桃叶片分别在8:00、14:00和18:00的气孔形态。 Figure 3 Observation of stomatal morphology of peach leaves at some critical moments in a day (A), (C) and (E) The stomatal morphology in the control of peach leaves at 8:00, 14:00 and 18:00, respectively; (B), (D) and (F) The stomatal morphology in the supplemental blue light of peach leaves at 8:00, 14:00 and 18:00, respectively.
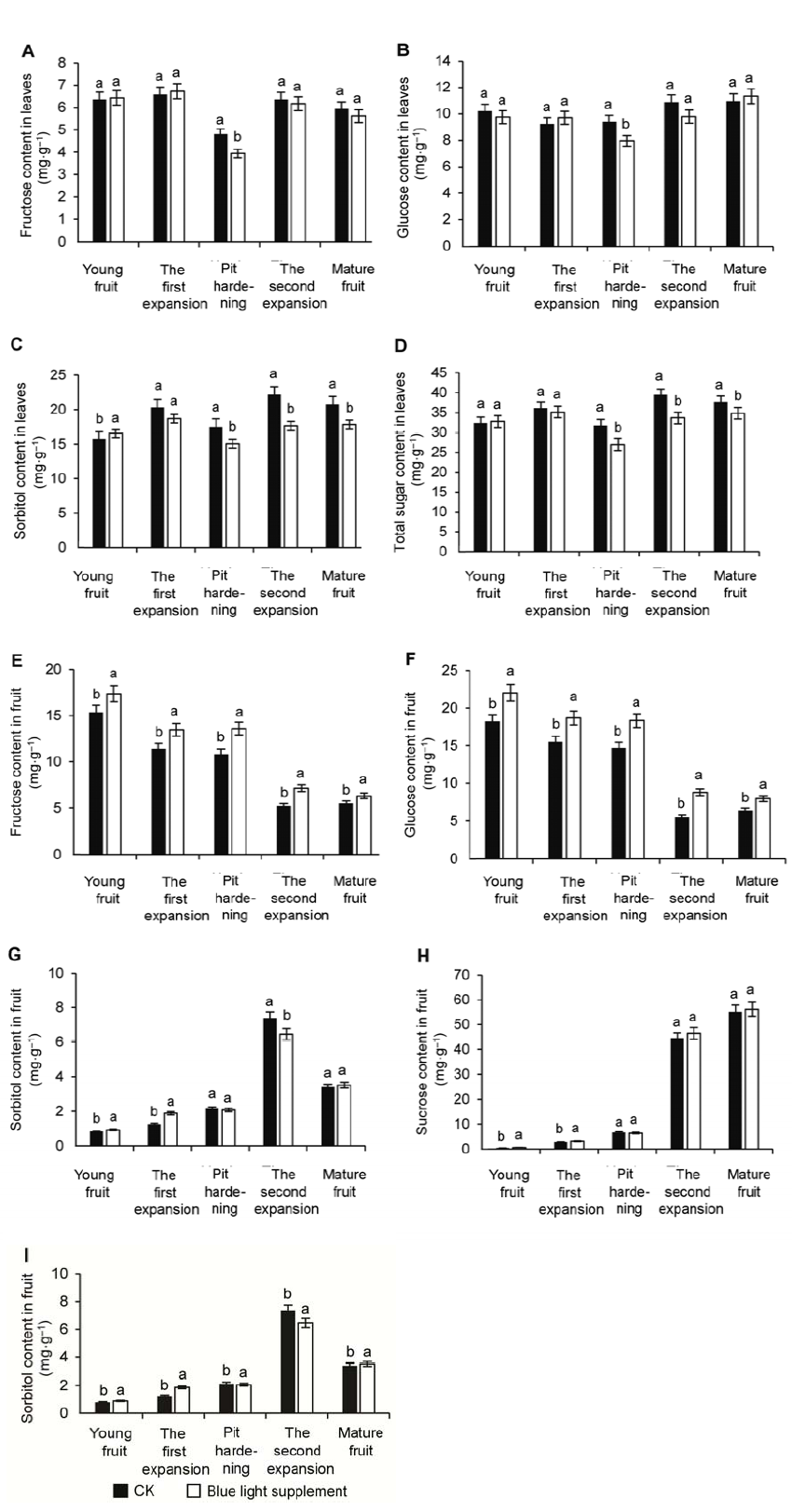
2.3 补充蓝光对叶片和果实光合同化物积累的影响油桃叶片中检测到的光合同化物为3种可溶性糖, 分别是果糖(图4A)、葡萄糖(图4B)和山梨醇(图4C), 山梨醇为叶片中含量最高的糖, 其次为葡萄糖, 再次为果糖。在果实发育期, 这3种糖变化趋势较为一致, 总体表现为先升高后降低再升高, 且均在硬核期含量降至最低。补充蓝光后, 油桃叶片各糖组分含量略低于对照, 总糖(图4D)含量在幼果期与对照相比无显著差异, 其余时期亦均较对照低, 二者总糖含量均在硬核期达到最低值。 图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-227/img_4.png<b>图4</b> 补充蓝光对果实发育时期油桃叶片和果实光合同化物积累的影响<br/>(A) 叶片果糖含量; (B) 叶片葡萄糖含量; (C) 叶片山梨醇含量; (D) 叶片总糖含量; (E) 果实果糖含量; (F) 果实葡萄糖含量; (G) 果实山梨醇含量; (H) 果实蔗糖含量; (I) 果实总糖含量。图中不同小写字母表示处理间在0.05水平差异显著。<br/><b>Figure 4</b> Effects of supplementary blue light on photo-assi- milate in leaves and fruit during peach fruit development<br/>(A) The contents of fructose in leaves; (B) The contents of glucose in leaves; (C) The contents of sorbitol in leaves; (D) The contents of total sugar in leaves; (E) The contents of fructose in fruit; (F) The contents of glucose in fruit; (G) The contents of sorbitol in fruit; (H) The contents of sucrose in fruit; (I) The contents of total sugar in fruit. Different lowercase letters indicate significant differences at 0.05 level among treatments. Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-227/img_4.png<b>图4</b> 补充蓝光对果实发育时期油桃叶片和果实光合同化物积累的影响<br/>(A) 叶片果糖含量; (B) 叶片葡萄糖含量; (C) 叶片山梨醇含量; (D) 叶片总糖含量; (E) 果实果糖含量; (F) 果实葡萄糖含量; (G) 果实山梨醇含量; (H) 果实蔗糖含量; (I) 果实总糖含量。图中不同小写字母表示处理间在0.05水平差异显著。<br/><b>Figure 4</b> Effects of supplementary blue light on photo-assi- milate in leaves and fruit during peach fruit development<br/>(A) The contents of fructose in leaves; (B) The contents of glucose in leaves; (C) The contents of sorbitol in leaves; (D) The contents of total sugar in leaves; (E) The contents of fructose in fruit; (F) The contents of glucose in fruit; (G) The contents of sorbitol in fruit; (H) The contents of sucrose in fruit; (I) The contents of total sugar in fruit. Different lowercase letters indicate significant differences at 0.05 level among treatments.
图4 补充蓝光对果实发育时期油桃叶片和果实光合同化物积累的影响 (A) 叶片果糖含量; (B) 叶片葡萄糖含量; (C) 叶片山梨醇含量; (D) 叶片总糖含量; (E) 果实果糖含量; (F) 果实葡萄糖含量; (G) 果实山梨醇含量; (H) 果实蔗糖含量; (I) 果实总糖含量。图中不同小写字母表示处理间在0.05水平差异显著。 Figure 4 Effects of supplementary blue light on photo-assi- milate in leaves and fruit during peach fruit development (A) The contents of fructose in leaves; (B) The contents of glucose in leaves; (C) The contents of sorbitol in leaves; (D) The contents of total sugar in leaves; (E) The contents of fructose in fruit; (F) The contents of glucose in fruit; (G) The contents of sorbitol in fruit; (H) The contents of sucrose in fruit; (I) The contents of total sugar in fruit. Different lowercase letters indicate significant differences at 0.05 level among treatments.
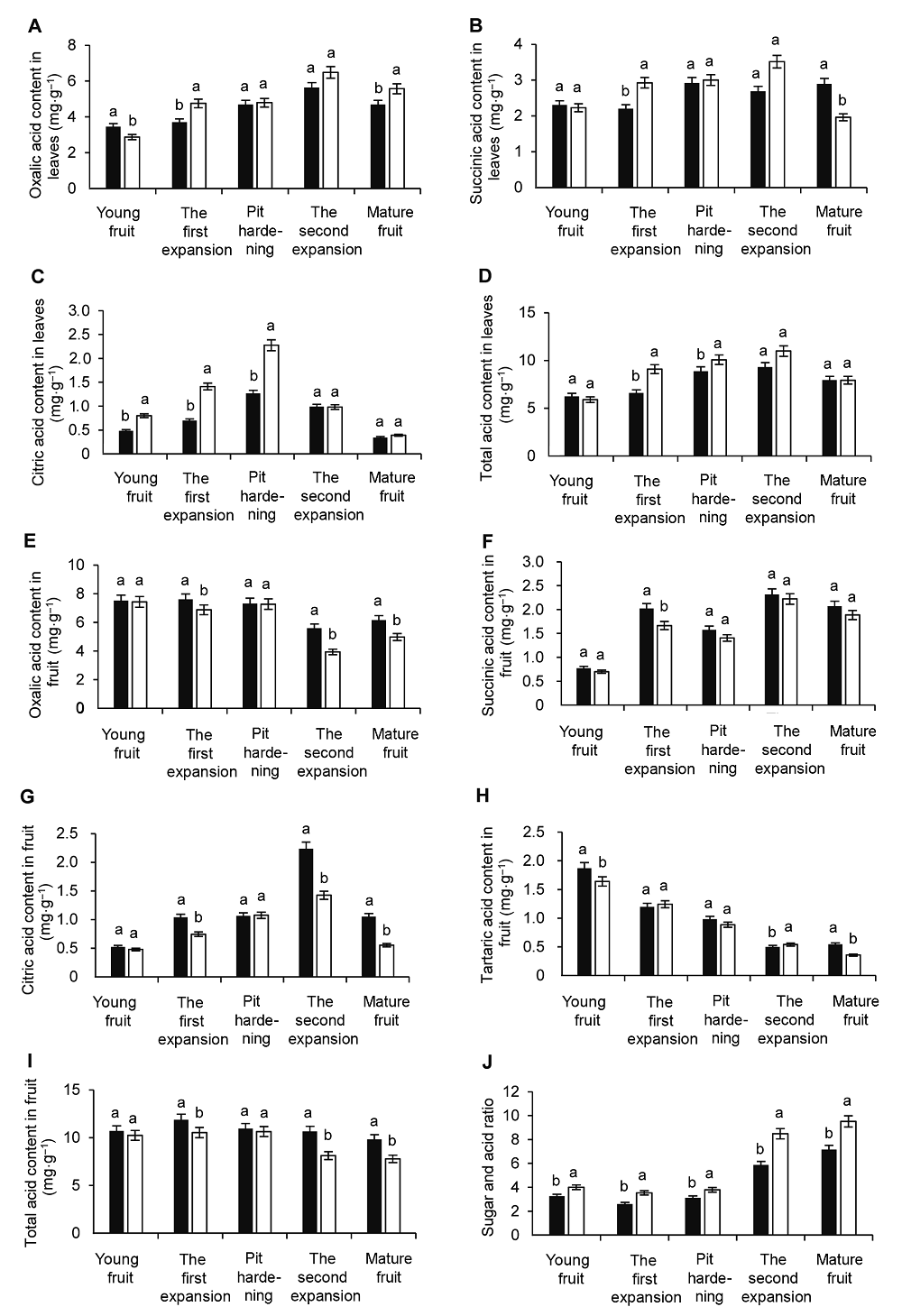
2.4 补充蓝光对叶片和果实有机酸积累的影响叶片中主要的有机酸为3种, 分别是草酸、琥珀酸和柠檬酸, 其中草酸含量最高, 其次为琥珀酸, 柠檬酸含量最低(图5A-C); 草酸和总酸含量随着发育进程呈现出缓慢升高的趋势, 柠檬酸含量先升高后降低且在硬核期时最高, 琥珀酸的含量变化较为平稳(图5D)。由于柠檬酸含量很少, 所以总酸含量主要取决于草酸和琥珀酸。总酸含量呈现出缓慢升高的趋势, 在第2次膨大期最高(对照为9.32 mg·g-1, 补充蓝光处理下为10.99 mg·g-1), 成熟期下降。在果实发育过程中, 补充蓝光时叶片中各酸含量总体上高于对照, 总酸含量除幼果期和成熟期外, 其它时期蓝光处理下均高于对照。 图5https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-227/img_5.png<b>图5</b> 补充蓝光对果实发育时期油桃叶片和果实有机酸积累及果实糖酸比的影响<br/>(A) 叶片草酸含量; (B) 叶片琥珀酸含量; (C) 叶片柠檬酸含量; (D) 叶片总酸含量; (E) 果实草酸含量; (F) 果实琥珀酸含量; (G) 果实柠檬酸含量; (H) 果实酒石酸含量; (I) 果实总酸含量; (J) 果实糖酸比。图中不同小写字母表示处理间在0.05水平差异显著。<br/><b>Figure 5</b> Effects of supplementary blue light on organic acid in leaves and fruit and the sugar and acid ratio of fruit during peach fruit development<br/>(A) The contents of oxalic acid in leaves; (B) The contents of succinic acid in leaves; (C) The contents of citric acid in leaves; (D) The contents of total acid in leaves; (E) The contents of oxalic acid in fruit; (F) The contents of succinic acid in fruit; (G) The contents of citric acid in fruit; (H) The contents of tartaric acid in fruit; (I) The contents of total acid in fruit; (J) Sugar and acid ratio of fruit. Different lowercase letters indicate significant differences at 0.05 level among treatments. Figure 5https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-227/img_5.png<b>图5</b> 补充蓝光对果实发育时期油桃叶片和果实有机酸积累及果实糖酸比的影响<br/>(A) 叶片草酸含量; (B) 叶片琥珀酸含量; (C) 叶片柠檬酸含量; (D) 叶片总酸含量; (E) 果实草酸含量; (F) 果实琥珀酸含量; (G) 果实柠檬酸含量; (H) 果实酒石酸含量; (I) 果实总酸含量; (J) 果实糖酸比。图中不同小写字母表示处理间在0.05水平差异显著。<br/><b>Figure 5</b> Effects of supplementary blue light on organic acid in leaves and fruit and the sugar and acid ratio of fruit during peach fruit development<br/>(A) The contents of oxalic acid in leaves; (B) The contents of succinic acid in leaves; (C) The contents of citric acid in leaves; (D) The contents of total acid in leaves; (E) The contents of oxalic acid in fruit; (F) The contents of succinic acid in fruit; (G) The contents of citric acid in fruit; (H) The contents of tartaric acid in fruit; (I) The contents of total acid in fruit; (J) Sugar and acid ratio of fruit. Different lowercase letters indicate significant differences at 0.05 level among treatments.
图5 补充蓝光对果实发育时期油桃叶片和果实有机酸积累及果实糖酸比的影响 (A) 叶片草酸含量; (B) 叶片琥珀酸含量; (C) 叶片柠檬酸含量; (D) 叶片总酸含量; (E) 果实草酸含量; (F) 果实琥珀酸含量; (G) 果实柠檬酸含量; (H) 果实酒石酸含量; (I) 果实总酸含量; (J) 果实糖酸比。图中不同小写字母表示处理间在0.05水平差异显著。 Figure 5 Effects of supplementary blue light on organic acid in leaves and fruit and the sugar and acid ratio of fruit during peach fruit development (A) The contents of oxalic acid in leaves; (B) The contents of succinic acid in leaves; (C) The contents of citric acid in leaves; (D) The contents of total acid in leaves; (E) The contents of oxalic acid in fruit; (F) The contents of succinic acid in fruit; (G) The contents of citric acid in fruit; (H) The contents of tartaric acid in fruit; (I) The contents of total acid in fruit; (J) Sugar and acid ratio of fruit. Different lowercase letters indicate significant differences at 0.05 level among treatments.
KaraAN, KotovAA, BukhovNG (1997). Specific distribution of gibberellins, cytokinins, indole-3-acetic acid, and abscisic acid in radish plants closely correlates with photomorphogenetic responses to blue or red light.J Plant Physiol 151, 51-59. [本文引用: 1]
[30]
McLachlanDH, LanJ, GeilfusCM, DoddAN, LarsonT, BakerA, HõrakH, KollistH, HeZS, GrahamI, MickelbartMV, HetheringtonAM (2016). The breakdown of stored triacylglycerols is required during light-induced stomatal opening.Curr Biol 26, 707-712. [本文引用: 1]
[31]
MuneerS, KimEJ, ParkJS, LeeJH (2014). Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca sativa L.).Int J Mol Sci 15, 4657-4670. [本文引用: 2]
[32]
ZhangDY, WangXH, ChenY, XuDQ (2005). Determinant of photosynthetic capacity in rice leaves under ambient air conditions.Photosynthetica 43, 273-276. [本文引用: 1]
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}