Abstract: In this study, we explored the solo effect of drought stress (control, light, moderate and heavy), heat stress (control 28°C, heat 40°C) and their dual stresses on the antioxidant defense system of Osmanthus fragrans cv. ‘Tian Xiang TaiGe’. The experiment data showed that after drought treatment, reactive oxygen species (ROS) accumulated gradually and membrane lipid peroxidation increased; the activity of antioxidant enzymes increased significantly under light and moderate drought stress. Ascorbic acid (AsA) and its reducing power (AsA/DHA) decreased significantly under heavy drought stress, whereas glutathione (GSH) and its reducing power (GSH/GSSG) as well as the activity of enzymes related to ascorbate glutathione cycle (AsA-GSH cycle) showed a trend of first increasing and then decreasing, with the peak reached after moderate drought stress. Unlike drought stress, ROS accumulation, antioxidant enzyme activity, antioxidant content and AsA-GSH cycle efficiency all improved significantly with heat stress. Furthermore, we found that the dual stresses injured the cultivar more severely than either stress alone. ROS level increased slowly via antioxidant enzyme activity, and the AsA-GSH cycle efficiency was enhanced at first and then decreased significantly with increasing dual stresses, especially after severe stress, and was unable to maintain oxidation reduction equilibrium. Under drought and heat stress, the cultivar quickly activated the antioxidant defense system to eliminate ROS, which improved the reducing power of the organism to reduce the environmental damage.
Key words:Osmanthus fragrans var. semperflorens ; drought ; heat ; reactive oxygen species ; antioxidant enzyme ; ascorbate glutathione cycle
植物细胞器新陈代谢途径对环境变化十分敏感, 环境胁迫会导致细胞代谢失衡, 进而引起氧化胁迫和活性氧(reactive oxygen species, ROS)积累(Suzuki et al., 2012)。ROS过多积累一方面会导致生物膜脂过氧化, 形成有毒产物, 如丙二醛(malondialdehyde, MDA) (Imahori et al., 2008); 另一方面严重破坏叶绿体结构, 削弱植物光合作用的能力, 对植物造成伤害(Lei et al., 2015)。植物体内存在抗氧化防御系统, 可在一定程度上减缓这种伤害(吴永波和叶波, 2016)。植物通过调节超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)和过氧化氢酶(catalase, CAT)等抗氧化酶活性来清除过多的ROS, 维持体内氧化还原平衡(Lei et al., 2015)。诸多研究表明, 植物抗氧化酶对非生物胁迫具有一定的调节作用。低温胁迫下, 油菜(Brassica napus)幼苗SOD和POD活性呈先升后降的趋势, CAT活性逐渐增强(Lei et al., 2015); 盐胁迫下SOD、CAT和POD活性先增后降(Lei et al., 2016)。高温胁迫使小麦(Tri- ticum aestivum) SOD和CAT活性显著增强, POD活性显著降低(Li et al., 2017)。抗坏血酸-谷胱甘肽循环(ascorbate glutathione cycle, AsA-GSH cycle)是维持植物细胞平衡的中心, 可有效抵抗植物氧化胁迫(Sharma et al., 2012)。抗坏血酸(ascorbic acid, AsA)作为抗坏血酸过氧化物酶(ascorbate peroxidase, APX)的电子供体用以清除H2O2。氧化生成的单脱氢抗坏血酸(monodehydroascobate, MDHA)一方面可经单脱氢抗坏血酸还原酶(monodehydroascobate reductase, MDHAR)还原, 另一方面可歧化生成AsA和脱氢抗坏血酸(dehydroascorbate, DHA), 脱氢抗坏血酸还原酶(dehydroascorbate reductase, DHAR)利用还原型谷胱甘肽(glutathione, GSH)作为电子供体可将DHA还原为AsA, 产生的氧化型谷胱甘肽(oxi- dized glutathione, GSSG)又可被谷胱甘肽还原酶(glutathione reductase, GR)再还原为GSH, 从而完成清除H2O2等活性氧并再生AsA和GSH的过程(Sharma et al., 2012)。植物通过增加抗氧化剂含量和循环相关酶活性提高AsA-GSH循环效率以适应环境胁迫。茄子(Solanum melongena)在锌胁迫下及绿豆(Vigna radiata)在高温胁迫下AsA和GSH含量均显著上升, AsA/DHA和GSH/GSSG显著降低, APX、DHAR、MDHAR和GR活性显著增强(Wu et al., 2015; Nahar et al., 2015)。干旱高温协同胁迫下, 植物叶片中的AsA和GSH被大量消耗, 其含量减少, 降幅显著高于单一胁迫(Arab et al., 2016; 谢华英等, 2016)。抗氧化防御系统可有效增强植物的抗逆性, 使其更好地适应逆境。
2 结果与讨论2.1 干旱高温胁迫对四季桂ROS和MDA含量的 影响随着干旱胁迫强度的加剧, 四季桂叶片ROS和MDA含量逐渐增高, 在重度干旱胁迫下, O2-·产生速率、H2O2和MDA含量分别比对照提高了149.2% (P< 0.05)、144.8% (P<0.05)和276.2 % (P<0.05) (表1)。高温胁迫下, O2-·产生速率、H2O2和MDA含量分别比对照增加了44.4% (P<0.05)、31.9% (P<0.05)和123.4% (P<0.05)。干旱高温协同胁迫下, O2-·产生速率与干旱胁迫相比有所下降, 重度协同胁迫较重度干旱胁迫低40.1% (P<0.05); 干旱高温协同胁迫下, H2O2和MDA含量呈先升高后下降的趋势, 在中度协同胁迫时达到峰值, 分别比中度干旱胁迫高12.3% (P<0.05)和14.5% (P<0.05)。由此可见, 干旱和高温胁迫可以诱导四季桂ROS含量和膜脂过氧化程度显著升高。干旱高温协同胁迫使四季桂损伤严重, ROS含量和膜脂过氧化程度的增幅显著低于干旱胁迫。 表1 Table 1 表1 表1 干旱高温胁迫对天香台阁四季桂活性氧和丙二醛含量的影响 Table 1 Effect of drought and heat stress on reactive oxygen species and malondialdehyde (MDA) content in Osmanthus fragrans cv. ‘Tian Xiang TaiGe’
Temperature
Treatment intensity
O2-· (nmol·g-1 FW)
H2O2 (μmol·g-1 FW)
MDA (μmol·g-1 FW)
28°C
CK
7.56±0.77 C
20.11±1.01 D
4.09±0.64 C
Light drought
14.04±0.44 B
33.08±2.33 C
7.42±1.32 B
Moderate drought
15.49±0.45 B
40.60±2.30 B
14.29±2.55 A
Heavy drought
18.84±2.05 A
49.24±1.74 A
15.39±1.69 A
Sum of squares
Between groups (d.f.1=3)
402.69
2743.44
532.18
Within groups (d.f.2=20)
25.94
73.89
57.58
40°C
CK
10.92±0.81 b
26.52±0.51 c
9.14±0.67 b
Light drought
12.79±0.78 a
37.52±3.73 b
14.11±1.51 a
Moderate drought
11.92±0.64 ab
45.60±0.92 a
16.36±2.29 a
Heavy drought
11.30±1.69 ab
34.64±4.18 b
8.48±1.74 b
Sum of squares
Between groups (d.f.1=3)
11.98
1117.58
264.48
Within groups (d.f.2=20)
22.59
162.38
55.13
P: Ft
**
ns
**
P: Fd
**
**
*
P: Ft×Fd
**
**
**
Ft: Effect of different temperature; Fd: Effect of different drought treatment intensity; Ft×Fd: Different responses of plant tissues to drought and heat stress. Each value is the mean±SE (n=6). Different capital letters in the same raw indicate statistically significant differences of drought stress, different lowercase letters in the same column indicate statistically significant differences of heat stress. According to Tukey test, * P<0.05; ** P<0.01; ns: Non-significant Ft: 不同温度的影响; Fd: 不同干旱处理强度的影响; Ft×Fd: 植物组织应对干旱高温胁迫的不同响应。数据为平均值±标准误(n=6)。同列不同大写字母表示不同干旱处理间差异显著, 同列不同小写字母表示不同高温处理间差异显著。根据Tukey多重比较, * P<0.05; ** P<0.01; ns: 不显著
表1 干旱高温胁迫对天香台阁四季桂活性氧和丙二醛含量的影响 Table 1 Effect of drought and heat stress on reactive oxygen species and malondialdehyde (MDA) content in Osmanthus fragrans cv. ‘Tian Xiang TaiGe’
图2 干旱高温胁迫对天香台阁四季桂抗坏血酸含量的影响 (A) 抗坏血酸(AsA)含量; (B) 脱氢抗坏血酸(DHA)含量; (C) AsA/DHA。Ft: 不同温度的影响; Fd: 不同处理强度的影响; Ft×Fd: 植物组织对干旱高温胁迫的不同响应。每个数值为平均值±标准误(n=6)。不同大写字母表示不同干旱处理间差异显著, 不同小写字母表示不同高温处理间差异显著。根据Tukey多重比较, * P<0.05; ** P<0.01; ns: 不显著 Figure 2 The effect of drought and heat stress on the AsA content in Osmanthus fragrans cv. ‘Tian Xiang TaiGe’ (A) Ascorbic acid (AsA) content; (B) Dehydroascorbate (DHA) content; (C) AsA/DHA. Ft: Effect of different temperature; Fd: Effect of different drought treatment intensity; Ft×Fd: Different responses of plant tissues to drought and heat stress. Each value is the mean±SE (n=6). Different capital letters indicate statistically significant differences of drought stress, different lowercase letters indicate statistically significant differences of heat stress. According to Tukey test, * P<0.05; ** P<0.01; ns: Non-significant
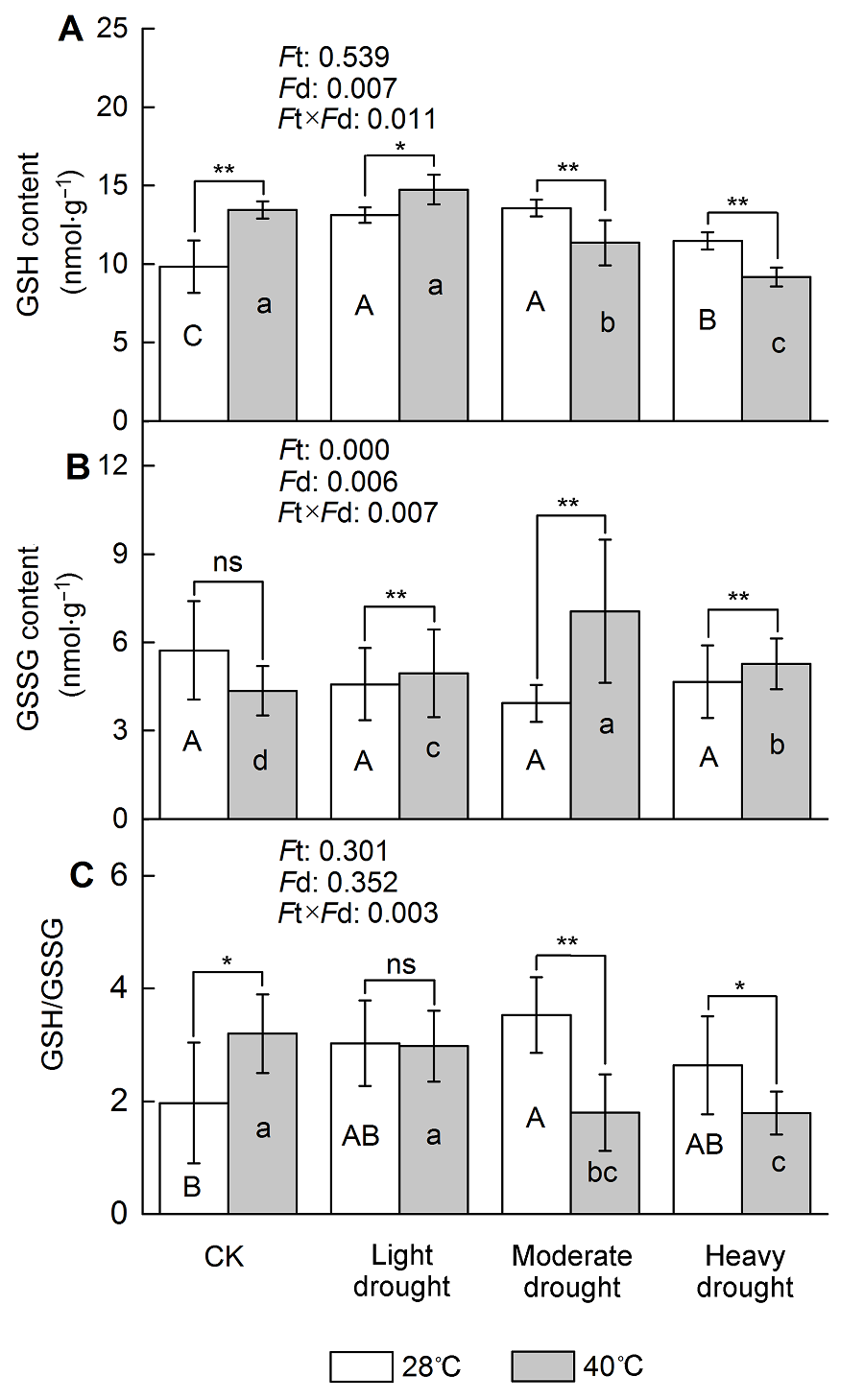
2.4 干旱高温胁迫对四季桂谷胱甘肽水平的影响随着干旱胁迫程度逐渐加强, 四季桂叶片GSH含量也逐渐升高, 中度胁迫时达到峰值(图3A), 与对照相比增加了38.0% (P<0.05); 高温胁迫下, GSH含量比对照提高了36.9% (P<0.05); 高温干旱协同作用时, GSH含量在轻度协同胁迫下达到峰值, 之后逐渐下降, 该值在轻度协同胁迫下较轻度干旱胁迫高出12.3% (P<0.05), 重度协同胁迫下则较重度干旱胁迫低20.0% (P<0.05)。单一干旱胁迫或高温胁迫对GSSG含量无显著影响(图3B); 与单一干旱胁迫相比, 干旱高温协同胁迫下GSSG含量显著增加, 中度协同胁迫比中度干旱胁迫该值提高了79.5% (P<0.05)。干旱胁迫下, GSH/GSSG表现为先升高后下降, 在中度干旱胁迫 时达到峰值, 与对照相比增加了79.6% (P<0.05) (图3C); 单一高温胁迫下, GSH/GSSG较对照高出62.6% (P<0.05); GSH/GSSG在轻度干旱高温协同作用时达到峰值后逐渐降低, 低于干旱胁迫, 且中度协同胁迫较中度干旱胁迫低49.2% (P<0.05)。从上述变化可以看出, 干旱高温胁迫使四季桂受到氧化胁迫, 通过调节GSH和GSSG的含量来维持氧化还原平衡, 以增强抗旱性。 图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-72/img_3.png<b>图3</b> 干旱高温胁迫对天香台阁四季桂谷胱甘肽含量的影响<br/>(A) 谷胱甘肽(GSH)含量; (B) 氧化型谷胱甘肽(GSSG)含量; (C) GSH/GSSG。<i>F</i>t: 不同温度的影响; <i>F</i>d: 不同处理强度的影响; <i>F</i>t×<i>F</i>d: 植物组织对干旱高温胁迫的不同响应。每个数值为平均值±标准误(<i>n</i>=6)。不同大写字母表示不同干旱处理间差异显著, 不同小写字母表示不同高温处理间差异显著。根据Tukey多重比较, * <i>P</i><0.05; ** <i>P</i><0.01; ns: 不显著<br/><b>Figure 3</b> The effect of drought and heat stress on the GSH content in <i>Osmanthus fragrans</i> cv. ‘Tian Xiang TaiGe’ <br/>(A) Glutathione (GSH) content; (B) Oxidized glutathione (GSSG) content; (C) GSH/GSSG.<i> F</i>t: Effect of different temperature; <i>F</i>d: Effect of different drought treatment intensity; <i>F</i>t×<i>F</i>d: Different responses of plant tissues to drought and heat stress. Each value is the mean±SE (<i>n</i>=6). Different capi- tal letters indicate statistically significant differences of drought stress, different lowercase letters indicate statistically significant differences of heat stress. According to Tukey test, * <i>P</i><0.05; ** <i>P</i><0.01; ns: Non-significant Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-72/img_3.png<b>图3</b> 干旱高温胁迫对天香台阁四季桂谷胱甘肽含量的影响<br/>(A) 谷胱甘肽(GSH)含量; (B) 氧化型谷胱甘肽(GSSG)含量; (C) GSH/GSSG。<i>F</i>t: 不同温度的影响; <i>F</i>d: 不同处理强度的影响; <i>F</i>t×<i>F</i>d: 植物组织对干旱高温胁迫的不同响应。每个数值为平均值±标准误(<i>n</i>=6)。不同大写字母表示不同干旱处理间差异显著, 不同小写字母表示不同高温处理间差异显著。根据Tukey多重比较, * <i>P</i><0.05; ** <i>P</i><0.01; ns: 不显著<br/><b>Figure 3</b> The effect of drought and heat stress on the GSH content in <i>Osmanthus fragrans</i> cv. ‘Tian Xiang TaiGe’ <br/>(A) Glutathione (GSH) content; (B) Oxidized glutathione (GSSG) content; (C) GSH/GSSG.<i> F</i>t: Effect of different temperature; <i>F</i>d: Effect of different drought treatment intensity; <i>F</i>t×<i>F</i>d: Different responses of plant tissues to drought and heat stress. Each value is the mean±SE (<i>n</i>=6). Different capi- tal letters indicate statistically significant differences of drought stress, different lowercase letters indicate statistically significant differences of heat stress. According to Tukey test, * <i>P</i><0.05; ** <i>P</i><0.01; ns: Non-significant
图3 干旱高温胁迫对天香台阁四季桂谷胱甘肽含量的影响 (A) 谷胱甘肽(GSH)含量; (B) 氧化型谷胱甘肽(GSSG)含量; (C) GSH/GSSG。Ft: 不同温度的影响; Fd: 不同处理强度的影响; Ft×Fd: 植物组织对干旱高温胁迫的不同响应。每个数值为平均值±标准误(n=6)。不同大写字母表示不同干旱处理间差异显著, 不同小写字母表示不同高温处理间差异显著。根据Tukey多重比较, * P<0.05; ** P<0.01; ns: 不显著 Figure 3 The effect of drought and heat stress on the GSH content in Osmanthus fragrans cv. ‘Tian Xiang TaiGe’ (A) Glutathione (GSH) content; (B) Oxidized glutathione (GSSG) content; (C) GSH/GSSG. Ft: Effect of different temperature; Fd: Effect of different drought treatment intensity; Ft×Fd: Different responses of plant tissues to drought and heat stress. Each value is the mean±SE (n=6). Different capi- tal letters indicate statistically significant differences of drought stress, different lowercase letters indicate statistically significant differences of heat stress. According to Tukey test, * P<0.05; ** P<0.01; ns: Non-significant
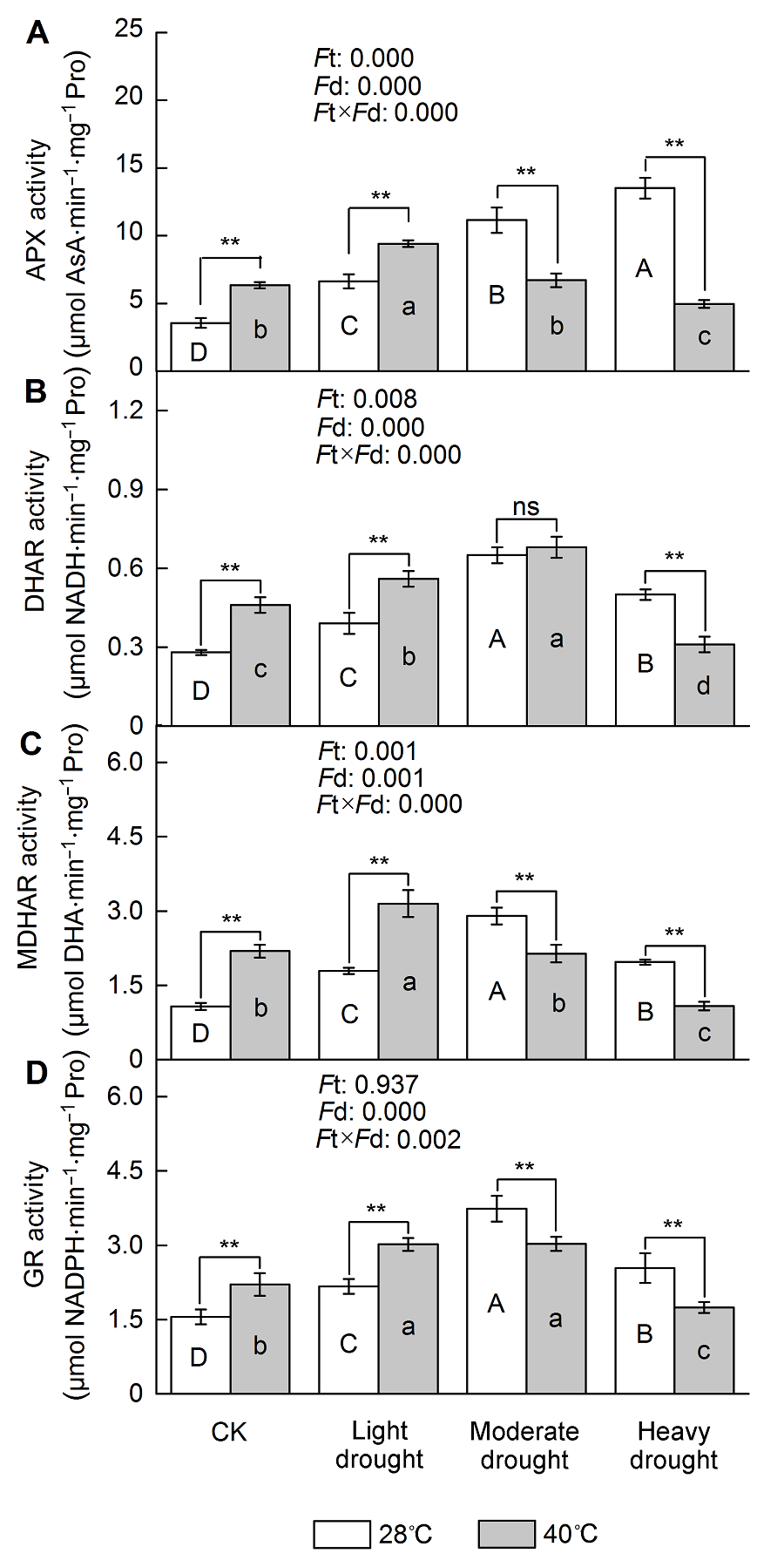
2.5 干旱高温胁迫对四季桂抗坏血酸-谷胱甘肽循环相关酶活性的影响四季桂叶片APX活性随着干旱胁迫程度的加剧逐渐升高, 重度干旱胁迫下, APX活性与对照相比提高了279.7% (P<0.05) (图4A); 单一高温处理下, APX活性较对照增加了78.2% (P<0.05); 轻度干旱高温协同胁迫下APX活性较轻度干旱胁迫增加了42.0% (P< 0.05), 达到峰值, 中度和重度协同胁迫下APX活性逐渐下降, 低于干旱胁迫。干旱胁迫下, DHAR活性逐渐上升并于中度胁迫时达到峰值, 与对照相比增加了42.2% (P<0.05) (图4B); 高温胁迫下, DHAR活性增加了65.9% (P<0.05); 高温干旱协同胁迫, DHAR活性呈先升高后下降的变化趋势, 中度协同胁迫时达到峰值, 重度协同胁迫下降低了37.7% (P<0.05)。MD- HAR活性在干旱胁迫下逐渐上升, 中度干旱胁迫时达到峰值, 与对照相比增加了171.7% (P<0.05) (图4C), 重度胁迫下有所下降; 单一高温胁迫下, MD- HAR活性较对照增加了105.3% (P<0.05); 轻度干旱高温协同胁迫下, MDHAR活性较轻度干旱胁迫提高了75.7% (P<0.05), 达到峰值, 在中度和重度协同胁迫下则逐渐降低。干旱胁迫下, GR活性呈先增加后降低的变化趋势, 中度干旱胁迫时达到峰值, 较对照增加了140.6% (P<0.05), 在重度干旱胁迫时显著下降(图4D)。高温胁迫下, GR活性与对照相比增加了42.1% (P<0.05)。轻度干旱高温协同胁迫下, GR活性比轻度干旱胁迫提高了39.2% (P<0.05), 达到峰值, 在中度和重度协同胁迫时逐渐下降, 显著低于干旱胁迫。由此可见, 单一干旱和高温胁迫下, APX具有较高的清除能力; 随着协同胁迫程度的加深, APX活性显著降低, 清除能力减弱。干旱高温胁迫下四季桂叶片DHAR、MDHAR和GR活性均呈先增加后降低的趋势, 说明其通过增强AsA-GSH循环代谢效率来抵御氧化胁迫, 胁迫程度较轻时防御效果较为明显, 不同酶相互协调, 维持循环的正常运行。 图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-72/img_4.png<b>图4</b> 干旱高温胁迫对天香台阁四季桂AsA-GSH循环相关酶活性的影响<br/>(A) 抗坏血酸过氧化物酶(APX)活性; (B) 脱氢抗坏血酸还原酶(DHAR)活性; (C) 单脱氢抗坏血酸还原酶(MDHAR)活性; (D) 谷胱甘肽还原酶(GR)活性。<i>F</i>t: 不同温度的影响; <i>F</i>d: 不同处理强度的影响; <i>F</i>t×<i>F</i>d: 植物组织对干旱高温胁迫的不同响应。每个数值为平均值±标准误(<i>n</i>=6)。不同大写字母表示不同干旱处理间差异显著, 不同小写字母表示不同高温处理间差异显著。根据Tukey多重比较, * <i>P</i><0.05; ** <i>P</i><0.01; ns: 不显著<br/><b>Figure 4</b> The effect of drought and heat stress on the enzymes activity of AsA-GSH cycle in <i>Osmanthus fragrans</i> cv. ‘Tian Xiang TaiGe’(A) Ascorbate peroxidase (APX) activity; (B) Dehydroascorbate reductase (DHAR) activity; (C) Monodehydroascobate reductase (MDHAR) activity; (D) Glutathione reductase (GR) activity. <i>F</i>t: Effect of different temperature; <i>F</i>d: Effect of different drought treatment intensity; <i>F</i>t×<i>F</i>d: Different responses of plant tissues to drought and heat stress. Each value is the mean±SE (<i>n</i>=6). Different capital letters indicate statistically significant differences of drought stress, different lowercase letters indicate statistically significant differences of heat stress. According to Tukey test, * <i>P</i><0.05; ** <i>P</i><0.01; ns: Non- significant Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-72/img_4.png<b>图4</b> 干旱高温胁迫对天香台阁四季桂AsA-GSH循环相关酶活性的影响<br/>(A) 抗坏血酸过氧化物酶(APX)活性; (B) 脱氢抗坏血酸还原酶(DHAR)活性; (C) 单脱氢抗坏血酸还原酶(MDHAR)活性; (D) 谷胱甘肽还原酶(GR)活性。<i>F</i>t: 不同温度的影响; <i>F</i>d: 不同处理强度的影响; <i>F</i>t×<i>F</i>d: 植物组织对干旱高温胁迫的不同响应。每个数值为平均值±标准误(<i>n</i>=6)。不同大写字母表示不同干旱处理间差异显著, 不同小写字母表示不同高温处理间差异显著。根据Tukey多重比较, * <i>P</i><0.05; ** <i>P</i><0.01; ns: 不显著<br/><b>Figure 4</b> The effect of drought and heat stress on the enzymes activity of AsA-GSH cycle in <i>Osmanthus fragrans</i> cv. ‘Tian Xiang TaiGe’(A) Ascorbate peroxidase (APX) activity; (B) Dehydroascorbate reductase (DHAR) activity; (C) Monodehydroascobate reductase (MDHAR) activity; (D) Glutathione reductase (GR) activity. <i>F</i>t: Effect of different temperature; <i>F</i>d: Effect of different drought treatment intensity; <i>F</i>t×<i>F</i>d: Different responses of plant tissues to drought and heat stress. Each value is the mean±SE (<i>n</i>=6). Different capital letters indicate statistically significant differences of drought stress, different lowercase letters indicate statistically significant differences of heat stress. According to Tukey test, * <i>P</i><0.05; ** <i>P</i><0.01; ns: Non- significant
图4 干旱高温胁迫对天香台阁四季桂AsA-GSH循环相关酶活性的影响 (A) 抗坏血酸过氧化物酶(APX)活性; (B) 脱氢抗坏血酸还原酶(DHAR)活性; (C) 单脱氢抗坏血酸还原酶(MDHAR)活性; (D) 谷胱甘肽还原酶(GR)活性。Ft: 不同温度的影响; Fd: 不同处理强度的影响; Ft×Fd: 植物组织对干旱高温胁迫的不同响应。每个数值为平均值±标准误(n=6)。不同大写字母表示不同干旱处理间差异显著, 不同小写字母表示不同高温处理间差异显著。根据Tukey多重比较, * P<0.05; ** P<0.01; ns: 不显著 Figure 4 The effect of drought and heat stress on the enzymes activity of AsA-GSH cycle in Osmanthus fragrans cv. ‘Tian Xiang TaiGe’(A) Ascorbate peroxidase (APX) activity; (B) Dehydroascorbate reductase (DHAR) activity; (C) Monodehydroascobate reductase (MDHAR) activity; (D) Glutathione reductase (GR) activity. Ft: Effect of different temperature; Fd: Effect of different drought treatment intensity; Ft×Fd: Different responses of plant tissues to drought and heat stress. Each value is the mean±SE (n=6). Different capital letters indicate statistically significant differences of drought stress, different lowercase letters indicate statistically significant differences of heat stress. According to Tukey test, * P<0.05; ** P<0.01; ns: Non- significant
ArabL, KreuzwieserJ, KruseJ, ZimmerI, AcheP, AlfarrajS, Al-rasheidKAS, SchnitzlerJP, HedrichR, RennenbergH (2016). Acclimation to heat and drought- lessons to learn from the date palm ( Phoenix dactylifera). Environ Exp Bot 125, 20-30. [本文引用: 2]
[6]
DoulisAG, DebianN, Kingston-SmithAH, FoyerCH (1997). Differential localization of antioxidants in maize leaves.Plant Physiol 114, 1031-1037.
[7]
FoyerCH, NoctorG (2011). Ascorbate and glutathione: the heart of the redox hub.Plant Physiol 155, 2-18. [本文引用: 1]
[8]
GiannopolitisCN, RiesSK (1977). Superoxide dismutases: I. Occurrence in higher plants.Plant Physiol 59, 309-314. [本文引用: 1]
[9]
HijiokaY, LinE, PereiraJ, CorlettR, CuiX, InsarovG, LascoR, LindgrenE, SurjanA (2014).Climate Change 2014: Impacts, Adaptation, and Vulnerability, Part B: Regional Aspects, Contribution of Working Group II, Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press. pp.1327-1370. [本文引用: 1]
[10]
HodgesDM, DelongJM, ForneyCF, PrangeRK (1999). Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds.Pl- anta 207, 604-611.
[11]
HossainMA, NakanoY, AsadaK (1984). Monodehydroascorbate reductase in spinach chloroplasts and its participation in regeneration of ascorbate for scavenging hydrogen peroxide.Plant Cell Physiol 25, 385-395.
[12]
ImahoriY, TakemuraM, BaiJH (2008). Chilling-induced oxidative stress and antioxidant responses in mume ( Pru- nus mume) fruit during low temperature storage. Postharvest Biol Technol 49, 54-60. [本文引用: 1]
[13]
KumariGJ, ReddyAM, NaikST, KumarSG, PrasanthiJ, SriranganayakuluG, ReddyPC, SudhakarC (2006). Jasmonic acid induced changes in protein pattern, antioxidative enzyme activities and peroxidase isozymes in peanut seedlings.Biol Plant 50, 219-226.
[14]
LeiP, XuZQ, DingY, TangB, ZhangYX, LiHS, FengXH, XuH (2015). Effect of poly (γ-glutamic acid) on the physiological responses and calcium signaling of rape seedlings ( Brassica napus L.) under cold stress. J Agric Food Chem 63, 10399-10406. [本文引用: 3]
[15]
LeiP, XuZQ, LiangJF, LuoXH, ZhangYX, FengXH, XuH (2016). Poly (γ-glutamic acid) enhanced tolerance to salt stress by promoting proline accumulation in Brassica napus L. Plant Growth Regul 78, 233-241. [本文引用: 1]
[16]
LiH, XuHL, ZhangPJ, GaoMQ, WangD, ZhaoHJ (2017). High temperature effects on D1 protein turnover in three wheat varieties with different heat susceptibility.Plant Grow- th Regul 78, 1-9. [本文引用: 1]
[17]
LiuCC, LiuYG, GuoK, FanDY, LiGQ, ZhengYR, YuLF, YangR (2011). Effect of drought on pigments, osmotic adjustment and antioxidant enzymes in six woody plant species in karst habitats of southwestern China.Environ Exp Bot 71, 174-183. [本文引用: 1]
[18]
MaYH, MaFW, WangYH, ZhangJK (2011). The responses of the enzymes related with ascorbate-gluta- thione cycle during drought stress in apple leaves.Acta Physiol Plant 33, 173-180. [本文引用: 1]
[19]
NaharK, HasanuzzamanM, AlamMM, FujitaM (2015). Exogenous glutathione confers high temperature stress tolerance in mung bean ( Vigna radiata L.) by modulating antioxidant defense and methylglyoxal detoxification system. Environ Exp Bot 112, 44-54. [本文引用: 1]
[20]
NakanoY, AsadaK (1981). Hydrogen peroxide is scaven- ged by ascorbate-specific peroxidase in spinach chloroplasts.Plant Cell Physiol 22, 867-880.
[21]
RaiAC, SinghM, ShahK (2012). Effect of water withdrawal on formation of free radical, proline accumulation and activities of antioxidant enzymes in ZAT12-transformed transgenic tomato plants. Plant Physiol Biochem 61, 108-114. [本文引用: 1]
[22]
RootTL, PriceJT, HallKR, SchneiderSH, RosenzweigC, PoundsJA (2003). Fingerprints of global warming on wild animals and plants.Nature 421, 57-60. [本文引用: 1]
SharmaP, JhaAB, DubeyRS, PessarakliM (2012). Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions.J Bot2012, 217037. [本文引用: 2]
[25]
SilvaEN, Ferreira-SilvaSL, FonteneleADV, RibeiroRV, ViégascRA, SilveiraJAG (2010). Photosynthetic chan- ges and protective mechanisms against oxidative damage subjected to isolated and combined drought and heat stresses in Jatropha curcas plants. J Plant Physiol 167, 1157-1164. [本文引用: 3]
[26]
SuzukiN, KoussevitzkyS, MittlerR, MillerG (2012). ROS and redox signaling in the response of plants to abiotic stress.Plant Cell Environ 35, 259-270. [本文引用: 1]
[27]
WangSC, LiangD, LiC, HaoYl, MaFW, ShuHR (2012). Influence of drought stress on the cellular ultrastructure and antioxidant systemin leaves of drought-tolerant and drought-sensitive apple rootstocks.Plant Physiol Biochem 51, 81-89. [本文引用: 2]
[28]
WuXX, HeJ, DingHD, ZhuZW, ChenJL, XuS, ZhaDS (2015). Modulation of zinc-induced oxidative damage in Solanum melongena by 6-benzylaminopurine involves ascorbate-glutathione cyclemetabolism. Environ Exp Bot 116, 1-11. [本文引用: 1]
[29]
ZouMQ, YuanLY, ZhuSD, LiuS, GeJT, WangCG (2016). Response of osmotic adjustment and ascorbate-glutathione cycle to heat stress in a heat-sensitive and a heat-tolerant genotype of wucai ( Brassica campestris L.). Sci Hortic 211, 87-94.
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}