Abstract: Vitis vinifera cv. ‘Merlot’ roots undergo freezing stress (0°C) or chilling stress (2°C) during cold treatment of grapevine leaves. Fast chlorophyll fluorescence was measured after cold treatment and used to analyze the effect of low root temperature on frost damage in leaves. Low root temperature to different extents led to different frost damage on leaves. Freezing stress (0°C) on roots led to more severe damage on leaves with cold treatment; this kind of damage to PSII was unable to recover. Chilling stress (2°C) applied to roots prevented the severe damage to leaves and was beneficial for recovery in PSII.
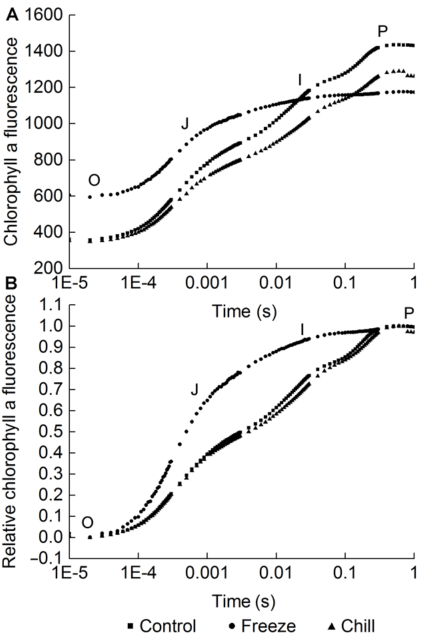
2 结果与讨论2.1 根系低温处理对葡萄叶片低温胁迫后快速叶绿素荧光的影响2.1.1 对快速叶绿素荧光诱导动力学曲线的影响 与对照相比, 经过低温胁迫后美乐葡萄叶片叶绿素荧光诱导动力学曲线(O-J-I-P曲线)发生了显著的变化(图1A)。低温胁迫造成叶片最大荧光(maximal fluorescence, Fm)强度下降, O-J-I-P曲线形状发生变化。但是不同根系低温胁迫对叶片O-J-I-P曲线的影响不同。对根系进行低温冷胁迫, 叶片经过低温胁迫后O-J-I-P曲线形状基本保持不变, 但曲线整体下移, P点、J点及I点等典型位点荧光值均有所下降, 而初始荧光(initial fluorescence, F0)强度无显著变化, 说明低温黑暗处理造成PSII反应中心可逆性失活。在叶片受低温胁迫时, 若对根系进行低温冻胁迫, 叶片O-J-I-P曲线失去典型形状, 最大荧光强度较对照显著降低, 且低于根系低温冷胁迫的最大荧光, 初始荧光强度显著升高, 说明叶片PSII反应中心在低温处理过程中发生了不可逆性失活或破坏。 图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-159/img_1.png<b>图1</b> 低温胁迫刚结束时不同根系低温胁迫的葡萄叶片快速叶绿素荧光诱导动力学曲线(O-J-I-P曲线)<br/>(A) 叶绿素荧光原始数据; (B) 叶绿素荧光数据标准化后的相对值<br/><b>Figure 1</b> Fast chlorophyll fluorescence induction dynamics curves of grape leaves with different low root temperature stress as soon as the frost treatment finished <br/>(A) Original data of chlorophyll fluorescence; (B) Standardized data of chlorophyll fluorescence Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-159/img_1.png<b>图1</b> 低温胁迫刚结束时不同根系低温胁迫的葡萄叶片快速叶绿素荧光诱导动力学曲线(O-J-I-P曲线)<br/>(A) 叶绿素荧光原始数据; (B) 叶绿素荧光数据标准化后的相对值<br/><b>Figure 1</b> Fast chlorophyll fluorescence induction dynamics curves of grape leaves with different low root temperature stress as soon as the frost treatment finished <br/>(A) Original data of chlorophyll fluorescence; (B) Standardized data of chlorophyll fluorescence
图1 低温胁迫刚结束时不同根系低温胁迫的葡萄叶片快速叶绿素荧光诱导动力学曲线(O-J-I-P曲线) (A) 叶绿素荧光原始数据; (B) 叶绿素荧光数据标准化后的相对值 Figure 1 Fast chlorophyll fluorescence induction dynamics curves of grape leaves with different low root temperature stress as soon as the frost treatment finished (A) Original data of chlorophyll fluorescence; (B) Standardized data of chlorophyll fluorescence
对原始荧光数据进行标准化处理, 得到相对荧光变化曲线, 可以看出不同位点的相对变化情况(图1 B)。与对照相比, 对根系进行低温冷胁迫和冻胁迫, 叶片相对荧光变化曲线呈现不同的特征。对根系进行低温冷胁迫后相对荧光曲线形状不变, 但整体下移, I点下降最突出, 其它位点变化不大。对根系进行低温冻胁迫, 叶片相对荧光变化曲线失去典型形状, J-I-P段上升明显。 表1 Table 1 表1 表1 低温胁迫刚结束时不同根系低温胁迫的葡萄叶片的快速叶绿素荧光参数(平均值±标准误, n=5) Table 1 Fast chlorophyll fluorescence parameters of grape leaves with different low root temperature stress as soon as the frost treatment finished (means±SE, n=5)
Chlorophyll fluorescence parameters
Control
Freeze
Cool
Fv/Fm
0.75±0.01 a
0.51±0.01 b
0.74±0.01 a
F0
335.65±7.26 b
551.74±161.76 a
329.55±7.93 b
PIabs
1.3±0.06 a
0.39±0.12 c
1.13±0.05 b
Wk
0.47±0.01 a
0.51±0.01 a
0.48±0.01 a
Vj
0.5±0.02 b
0.76±0.05 a
0.47±0.02 b
ψ0
0.5±0.02 a
0.24±0.05 b
0.53±0.02 a
RC/CSm
557.05±54.61 a
298.99±11.76 c
491.31±39.15 b
ABS/CSm
1401.52±54.61 a
1176.33±1.9 b
1279.59±39.15 a
TR0/CSm
1065.87±48.49 a
604.33±14.68 c
950.04±33.89 b
ET0/CSm
544.17±37 a
220±63.65 b
510.73±27.14 a
DI0/CSm
335.65±7.26 b
551.74±161.76 a
329.55±7.93 b
Fv/Fm: Maximum quantum yield of PSII; F0: Initial fluorescence; PIabs: Performance index on absorption basis; Wk: Ratio of the variable fluorescence Fk to the amplitude Fj-F0; Vj: Relative variable fluorescence at J step; ψ0: The excitation efficiency of electron transport beyond QA; RC/CSm: Number of active reaction centers per unit area; ABS/CSm: Absorption flux per unit area; TR0/CSm: Trapped energy flux per unit area; ET0/CSm: Electron transport flux per unit area; DI0/CSm: Dissipated energy flux per unit area. Different letters mean significant difference at α=0.05 according to LSD test. Fv/Fm: 最大光化学效率; F0: 初始荧光; PIabs: 叶片性能指数; Wk: 可变荧光Fk占Fj-F0振幅的比例; Vj: J点可变荧光强度; ψ0: 经过QA进入电子传递链的能量指数; RC/CSm: 单位面积有活性的反应中心数目; ABS/CSm: 单位面积吸收的能量; TR0/CSm: 单位面积捕获光能; ET0/CSm: 单位面积电子传递能量; DI0/CSm: 单位面积热耗散能量。采用LSD法进行多重比较, 同一行中不同字母表示差异显著(α=0.05)。
表1 低温胁迫刚结束时不同根系低温胁迫的葡萄叶片的快速叶绿素荧光参数(平均值±标准误, n=5) Table 1 Fast chlorophyll fluorescence parameters of grape leaves with different low root temperature stress as soon as the frost treatment finished (means±SE, n=5)
2.1.2 对快速叶绿素荧光参数的影响 快速叶绿素荧光动力学曲线参数包含大量关于PSII原初光化学反应的信息。表1显示, 与对照相比, 黑暗低温胁迫后, 葡萄叶绿素各荧光参数均发生显著变化。根系受冻导致叶片经过黑暗低温胁迫后, 最大光化学效率(maximum quantum yield of PSII, Fv/Fm)降低了32%, 初始荧光(F0)升高了64.4%, 叶片性能指数(performance index on absorption basis, PIabs)降低了70%, J点可变荧光强度(relative variable fluorescence at J step, Vj)上升了52%, 经过QA进入电子传递链的能量指数(the excitation efficiency of electron transport begond QA, ψ0)降低了52%, 单位面积有活性的反应中心数目(number of active reaction centers per unit area, RC/CSm)减少了46.3%, 单位面积吸收的能量(absorption flux per unit area, ABS/CSm)下降了16.1%, 单位面积捕获光能(trapped energy flux per unit area, TR0/CSm)下降了43.3%, 单位面积电子传递能量(electron transport flux per unit area, ET0/CSm)下降了59.6%, 单位面积热耗散能量(dissipated energy flux per unit area, DI0/CSm)增加了64.4%; 而反映PSII供体侧伤害程度的Wk (ratio of the variable fluorescence Fk to the amplitude Fj-F0)无显著差异。 与根系冻胁迫不同, 根系冷胁迫导致显著变化的指标有PIabs、RC/CSm和TR0/CSm。PIabs降低了13.1%, RC/CSm减少了11.8%, TR0/CSm下降了10.9%; 其它参数与对照相比无显著差异。
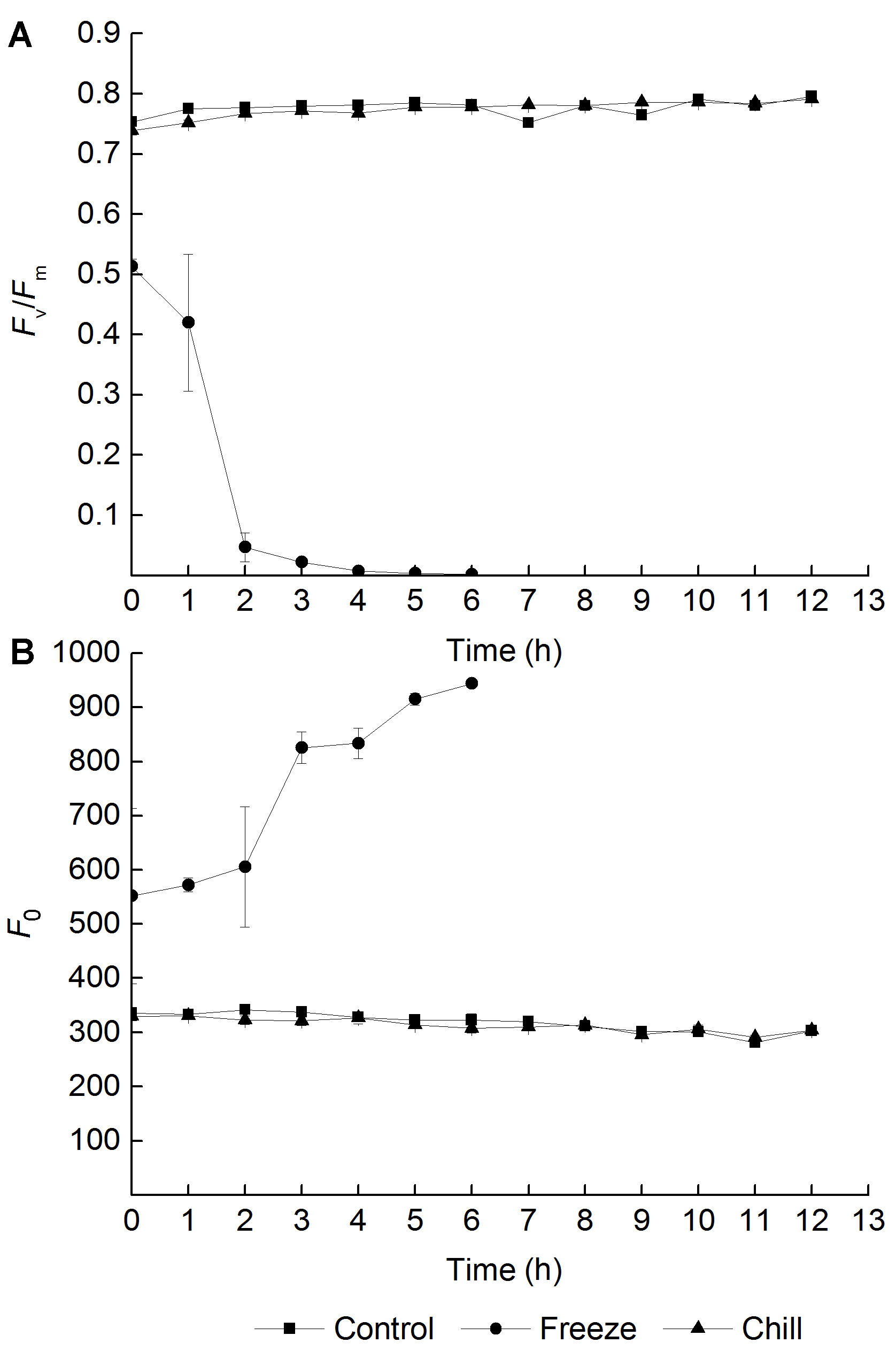
2.2 叶片低温胁迫期间根系冷、冻处理对葡萄叶片恢复期PSII光抑制的影响Fv/Fm是检测光抑制的典型指标(Takahashi and Murata, 2008)。图2显示了根系冷、冻胁迫后葡萄叶片恢复期PSII光抑制情况。结果表明, 对根系进行低温冻胁迫后, 叶片在恢复期间发生显著的光抑制。Fv/Fm在恢复期间持续下降, 并降到难以测定的程度(图2A), 而初始荧光(F0)却不断增大(图2B), 说明PSII反应中心发生了不可逆转的伤害。另外, 在实验中观察到恢复后7小时, 叶片因干裂而无法测定叶绿素荧光, 其伤害程度远大于对照和根系冷处理组。对根系进行低温冷处理后, 恢复期Fv/Fm与对照相比没有显著差异, 其变化趋势也与对照相同, 说明根系进行低温冷处理后叶片在遭受低温胁迫后的恢复过程中并未发生光抑制。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-159/img_2.png<b>图2</b> 不同根系低温处理后葡萄叶片恢复期的光抑制情况(<i>n</i>=5)<br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: 最大光化学效率; <i>F</i><sub>0</sub>: 初始荧光<br/><b>Figure 2</b> Photoinhibition of grape leaves during recovery period after different low root temperature treatment (<i>n</i>=5)<br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: Maximum quantum yield of PSII; <i>F</i><sub>0</sub>: Initial fluorescence Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-159/img_2.png<b>图2</b> 不同根系低温处理后葡萄叶片恢复期的光抑制情况(<i>n</i>=5)<br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: 最大光化学效率; <i>F</i><sub>0</sub>: 初始荧光<br/><b>Figure 2</b> Photoinhibition of grape leaves during recovery period after different low root temperature treatment (<i>n</i>=5)<br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: Maximum quantum yield of PSII; <i>F</i><sub>0</sub>: Initial fluorescence
图2 不同根系低温处理后葡萄叶片恢复期的光抑制情况(n=5) Fv/Fm: 最大光化学效率; F0: 初始荧光 Figure 2 Photoinhibition of grape leaves during recovery period after different low root temperature treatment (n=5) Fv/Fm: Maximum quantum yield of PSII; F0: Initial fluorescence
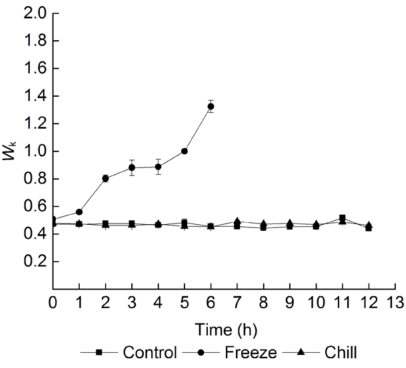
2.3 叶片低温胁迫期间根系低温处理对葡萄叶片恢复期PSII供体侧伤害的影响PSII反应中心供体侧放氧复合体(oxygen-evolving complex, OEC)造成伤害会导致Wk迅速上升, 因此Wk往往被用于检测PSII反应中心供体侧的伤害程度(Strasser, 1997; Srivastava et al., 1997; Chen et al., 2004)。图3显示, 叶片经过低温胁迫后, 根系冷胁迫和冻胁迫均未造成Wk的显著升高。但是在恢复期, 经过不同根系低温处理的叶片Wk变化不同。在叶片低温胁迫期间对根系进行冻胁迫后, 叶片恢复期间Wk显著升高, 随着恢复时间的延长, Wk呈逐渐升高的趋势, 说明在恢复期间, 供体侧放氧复合体受到显著伤害。在叶片低温胁迫同时对根系进行冷胁迫, 叶片在恢复期间Wk没有显著升高, 且随着恢复时间的延长, 其变化趋势与对照相同, 说明根系冷胁迫并未造成叶片低温胁迫后恢复期供体侧的伤害。 图3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-159/img_3.png<b>图3</b> 不同根系低温处理后葡萄叶片恢复期PSII供体侧伤害情况(<i>n</i>=5)<br/><b>Figure 3</b> Damage of donor side of PSII in grape leaves during recovery period after different low root temperature treatments (<i>n</i>=5) Figure 3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-159/img_3.png<b>图3</b> 不同根系低温处理后葡萄叶片恢复期PSII供体侧伤害情况(<i>n</i>=5)<br/><b>Figure 3</b> Damage of donor side of PSII in grape leaves during recovery period after different low root temperature treatments (<i>n</i>=5)
图3 不同根系低温处理后葡萄叶片恢复期PSII供体侧伤害情况(n=5) Figure 3 Damage of donor side of PSII in grape leaves during recovery period after different low root temperature treatments (n=5)
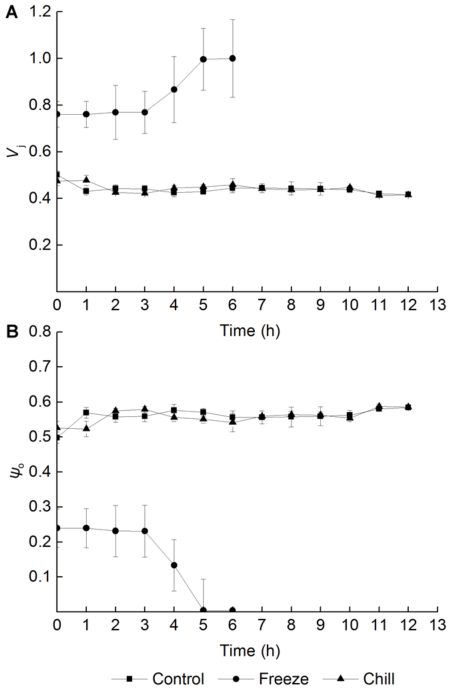
2.4 叶片低温胁迫期间根系低温处理对葡萄叶片恢复期PSII受体侧伤害的影响J点可变荧光强度(Vj)反映了QA--QB之间的电子传递情况和QA-的积累情况(李鹏民等, 2005)。图4A显示, 在叶片受低温胁迫刚结束时, 对根系进行低温冻胁迫处理造成Vj显著上升, 随着进入恢复期, Vj进一步上升。而对根系进行低温冷胁迫, 叶片低温胁迫后与对照相比Vj没有显著差别, 恢复期第1小时略高于对照, 从恢复期第2小时开始到恢复期结束始终保持在与对照相同的水平。 图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-159/img_4.png<b>图4</b> 不同根系低温处理后葡萄叶片恢复期PSII受体侧的伤害情况(<i>n</i>=5)<br/><i>V</i><sub>j</sub>: J点可变荧光强度; <i>ψ</i><sub>0</sub>: 经过Q<sub>A</sub>进入电子传递链的能量指数<br/><b>Figure 4</b> Damage of acceptor side of PSII in grape leaves during recovery period after different low root temperature treatments (<i>n</i>=5)<br/><i>V</i><sub>j</sub>: Relative variable fluorescence at J step; <i>ψ</i><sub>0</sub>: The excitation efficiency of electron transport beyond Q<sub>A</sub> Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-159/img_4.png<b>图4</b> 不同根系低温处理后葡萄叶片恢复期PSII受体侧的伤害情况(<i>n</i>=5)<br/><i>V</i><sub>j</sub>: J点可变荧光强度; <i>ψ</i><sub>0</sub>: 经过Q<sub>A</sub>进入电子传递链的能量指数<br/><b>Figure 4</b> Damage of acceptor side of PSII in grape leaves during recovery period after different low root temperature treatments (<i>n</i>=5)<br/><i>V</i><sub>j</sub>: Relative variable fluorescence at J step; <i>ψ</i><sub>0</sub>: The excitation efficiency of electron transport beyond Q<sub>A</sub>
图4 不同根系低温处理后葡萄叶片恢复期PSII受体侧的伤害情况(n=5) Vj: J点可变荧光强度; ψ0: 经过QA进入电子传递链的能量指数 Figure 4 Damage of acceptor side of PSII in grape leaves during recovery period after different low root temperature treatments (n=5) Vj: Relative variable fluorescence at J step; ψ0: The excitation efficiency of electron transport beyond QA
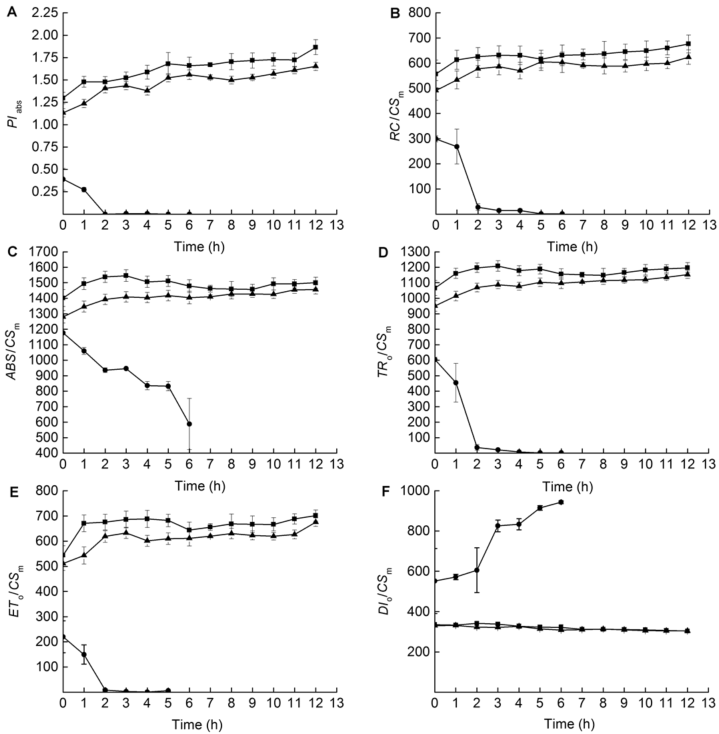
2.5 叶片低温胁迫期间根系低温处理对葡萄叶片恢复期PSII反应中心的影响图5显示了叶片低温胁迫期间, 对根系进行冷、冻胁迫对叶片低温胁迫后及恢复过程中PSII反应中心的影响。从图5可以看出, 与对照相比, 低温胁迫造成PIabs、RC/CSm、ABS/CSm、TR0/CSm及ET0/CSm值的下降, 但不同的根系低温处理所造成的影响不同。根系冻胁迫造成相应指标显著下降, 并且在恢复期间进一步降低, 甚至达到不可测定的程度。根系冷胁迫造成相应指标在低温胁迫刚结束时下降, 但与对照相比差异不显著, 且在恢复期间有所恢复, 虽然没有恢复到对照水平, 但与对照相比无显著差异。根系低温冻胁迫造成DI0/CSm值显著升高, 并在恢复期进一步升高; 而根系低温冷胁迫并未导致DI0/CSm值发生显著变化。 图5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-159/img_5.png<b>图5</b> 不同根系低温处理后葡萄叶片恢复期PSII反应中心的伤害情况(<i>n</i>=5)<br/><i>PI</i><sub>abs</sub>、<i>RC</i>/<i>CS</i><sub>m</sub>、<i>ABS</i>/<i>CS</i><sub>m</sub>、<i>TR</i><sub>0</sub>/<i>CS</i><sub>m</sub>、<i>ET</i><sub>0</sub>/<i>CS</i><sub>m</sub>和<i>DI</i><sub>0</sub>/<i>CS</i><sub>m</sub>同<xref ref-type="table" rid="T1-1674-3466-52-2-159">表1</xref>。<br/><b>Figure 5</b> Damage of PSII reaction center in grape leaves during recovery period after different low root temperature treatments (<i>n</i>=5)<br/><i>PI</i><sub>abs</sub>, <i>RC</i>/<i>CS</i><sub>m</sub>, <i>ABS</i>/<i>CS</i><sub>m</sub>, <i>TR</i><sub>0</sub>/<i>CS</i><sub>m</sub>, <i>ET</i><sub>0</sub>/<i>CS</i><sub>m</sub>, and <i>DI</i><sub>0</sub>/<i>CS</i><sub>m</sub> see <xref ref-type="table" rid="T1-1674-3466-52-2-159">Table 1</xref>. Figure 5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-159/img_5.png<b>图5</b> 不同根系低温处理后葡萄叶片恢复期PSII反应中心的伤害情况(<i>n</i>=5)<br/><i>PI</i><sub>abs</sub>、<i>RC</i>/<i>CS</i><sub>m</sub>、<i>ABS</i>/<i>CS</i><sub>m</sub>、<i>TR</i><sub>0</sub>/<i>CS</i><sub>m</sub>、<i>ET</i><sub>0</sub>/<i>CS</i><sub>m</sub>和<i>DI</i><sub>0</sub>/<i>CS</i><sub>m</sub>同<xref ref-type="table" rid="T1-1674-3466-52-2-159">表1</xref>。<br/><b>Figure 5</b> Damage of PSII reaction center in grape leaves during recovery period after different low root temperature treatments (<i>n</i>=5)<br/><i>PI</i><sub>abs</sub>, <i>RC</i>/<i>CS</i><sub>m</sub>, <i>ABS</i>/<i>CS</i><sub>m</sub>, <i>TR</i><sub>0</sub>/<i>CS</i><sub>m</sub>, <i>ET</i><sub>0</sub>/<i>CS</i><sub>m</sub>, and <i>DI</i><sub>0</sub>/<i>CS</i><sub>m</sub> see <xref ref-type="table" rid="T1-1674-3466-52-2-159">Table 1</xref>.
图5 不同根系低温处理后葡萄叶片恢复期PSII反应中心的伤害情况(n=5) PIabs、RC/CSm、ABS/CSm、TR0/CSm、ET0/CSm和DI0/CSm同表1。 Figure 5 Damage of PSII reaction center in grape leaves during recovery period after different low root temperature treatments (n=5) PIabs, RC/CSm, ABS/CSm, TR0/CSm, ET0/CSm, and DI0/CSm see Table 1.
AroEM, VirginI, AnderssonB (1993). Photoinhibition of photosystem II inactivation, protein damage and turnover.BBA-Bioenergetics 1143, 113-134. [本文引用: 1]
[10]
ChenHX, LiWJ, AnSZ, GaoHY (2004). Characterization of PSII photochemistry and thermostability in salt-treatedRumex leaves. J Plant Physiol 161, 257-264. [本文引用: 1]
[11]
HavauxN, DavaudA (1994). Photoinhibition of photosynthesis in chilled potato leaves with a loss of photosystem- activity II.Photosynthesis Res 40, 75-92. [本文引用: 1]
[12]
HuangLK, WongSC, TerashimaI, ZhangX, LinDX, OsmondCB (1989). Chilling injury in mature leaves of rice. I. Varietal differences in the effects of chilling on canopy photosynthesis under simulated ‘dry cold dew wind’ conditions experienced in southeast China.Aust J Plant Physiol 16, 321-337. [本文引用: 1]
[13]
NeunerG, ErlerA, LadinigU, HackerJ, WagnerJ (2013). Frost resistance of reproductive tissues during various stages of development in high mountain plants.Physiol Plantarum 147, 88-100. [本文引用: 1]
[14]
SrivastavaA, GuisséB, GreppinH, StrasserRJ (1997). Regulation of antenna structure and electron transport in PSII ofPisum sativum under elevated temperature probed by the fast polyphasic chlorophyll a fluorescence transient: OKJIP. BBA-Bioenergetics 1320, 95-106. [本文引用: 1]
[15]
StrasserBJ (1997). Donor side capacity of photosystem II probed by chlorophyll a fluorescence transients. Photosynth Res 52, 147-155. [本文引用: 1]
[16]
StrasserRJ, Tsimilli-MichaelM, QiangS, GoltsevV (2010). Simultaneousin vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. BBA-Bioenergetics 1797, 1313-1326.
[17]
SuzukiK, NagasugaK, OkadaM (2008). The chilling injury induced by high root temperature in the leaves of rice seedlings.Plant Cell Physiol 49, 433-442. [本文引用: 1]
[18]
SuzukiK, OhmoriY, NagaoM (2013). Accumulation of nitrate and nitrite in chilled leaves of rice seedlings is induced by high root temperature.Plant Cell Physiol 54, 1769-1779. [本文引用: 1]
[19]
TakahashiS, MurataN (2008). How do environmental stresses accelerate photoinhibition?Trends Plant Sci 13, 178-182. [本文引用: 1]
[20]
TeicherHB, MøllerBL, SchellerHV (2000). Photoinhibition of photosystem I in field-grown barley (Hordeum vulgare L.): induction, recovery and acclimation. Photosynthesis Res 64, 53-61. [本文引用: 1]
[21]
ZhouY, HuangL, ZhangY, ShiK, YuJ, NoguešS (2007). Chill-induced decrease in capacity of RuBP carboxylation and associated H2O2 accumulation in cucumber leaves are alleviated by grafting onto figleaf gourd.Ann Bot 100, 839-848. [本文引用: 1]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}