 ,
, ,1,2,*
,1,2,*Characteristics of shrub leaf carbon, nitrogen and phosphorus stoichiometry and influencing factors in mixed broadleaved-Korean pine forests at different successional stages
Yu-Han SONG1, Peng ZHANG1, Guang-Ze JIN,,1,2,*通讯作者: *金光泽: ORCID:0000-0002-9852-0965(taxus@126.com)
编委: 郝占庆
责任编辑: 赵航
收稿日期:2021-03-19接受日期:2021-06-3
| 基金资助: |
Corresponding authors: *(taxus@126.com)
Received:2021-03-19Accepted:2021-06-3
| Fund supported: |

摘要
灌木是森林生态系统的重要组成部分, 对于演替进程中灌木叶片化学计量特征的研究, 有助于全面理解和预测森林演替过程。该研究以黑龙江凉水国家自然保护区内处于阔叶红松(Pinus koraiensis)林不同演替阶段中的白桦(Betula platyphylla)次生林、落叶阔叶混交林、针阔混交林、阔叶红松林的灌木为研究对象, 分析其叶片的碳(C)、氮(N)、磷(P)化学计量特征差异, 并利用层次分割方法检验其与土壤、物种多样性的关系。主要结果为: 1)随着演替的进行, 阔叶红松林的叶片N含量显著高于其他3种林型, P含量与白桦次生林无显著差异, 但显著高于其他两种林型; 2)土壤N、P含量与个体尺度上的叶片N含量均呈显著正相关关系, 土壤P含量与叶片P含量呈显著正相关关系; 3)群落尺度上, 物种多样性和土壤化学性质共解释叶片N含量变异的82%和叶片P含量变异的62%; 4)群落尺度上Shannon多样性指数与灌木叶片的N、P含量呈显著正相关关系, 与灌木叶片的C:N、C:P呈显著负相关关系。总之, 阔叶红松林4个演替阶段灌木均受到氮限制; 相较于土壤的化学性质, 物种多样性更好地解释了灌木化学计量的变异。
关键词:
Abstract
Aims Shrubs are an important component of forest ecosystems. This study investigated changes in the stoichiometric characteristics of shrub leaves during forest succession in order to understand and predict the processes of forest succession. Methods The study was conducted in the Liangshui National Nature Reserve of Heilongjiang Province, with forest stands at different successional stages of mixed broadleaved-Korean pine (Pinus koraiensis) forest representing secondary birch (Betula platyphylla) forest, mixed deciduous broad-leaved forest, mixed coniferous and broad-leaved forest, and mixed broadleaved-Korean pine forest. Measurements were made on carbon (C), nitrogen (N) and phosphorus (P) contents in leaves of the understory shrubs and soil, and the stoichiometric characteristics of shrub leaves and relationships with soil stoichiometry were examined with hierarchical analysis. Important findings The N content in shrub leaves was significantly higher in the mixed broadleaved-Korean pine forest than in other three forest types; the P content was significantly higher in the mixed broadleaved-Korean pine forest than in two other forest types except the secondary birch forest. Soil N and P contents were significantly and positively correlated with leaf N content at individual scale, and soil P concentration was significantly and positively correlated with leaf P content. At the community level, 82% of leaf N content variation and 62% of leaf P content variation were explained by species diversity and soil chemical properties; the Shannon diversity index was significantly and positively correlated with the N and P contents in shrub leaves, and negatively with the leaf C:N ratio and C:P ratio. In conclusion, shrubs in mixed broadleaved-Korean pine forests at the four successional stages were all N-limited, and species diversity better explains the stoichiometric variations in understory shrubs than soil chemical properties.
Keywords:
PDF (1218KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
宋语涵, 张鹏, 金光泽. 阔叶红松林不同演替阶段灌木叶片碳氮磷化学计量特征及其影响因素. 植物生态学报, 2021, 45(9): 952-960. DOI: 10.17521/cjpe.2021.0101
SONG Yu-Han, ZHANG Peng, JIN Guang-Ze.

作为生物体基本的组成元素, 氮(N)和磷(P)被认为是陆地生态系统中植物体内重要的限制因子(Reich et al., 1997; 贺金生和韩兴国, 2010), 在植物的生长和生理代谢过程中发挥重要作用(Zhang et al., 2020)。揭示植物体内的N、P养分状况对于理解植物的生长状况及对周围环境条件变化的响应至关重要(Su et al., 2021)。
植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al., 2015), 例如, 植物在不同器官合理分配有限的营养物质, 用以适应不同的环境特征(Zhang et al., 2018b)。植物也具有在外界环境变化时保持其体内元素组成相对稳定的能力, 被称为生态化学计量学的内稳态理论(Sterner & Elser, 2002)。在生态化学计量学的研究中, 常利用植物体内N:P来衡量植物的养分利用和受限制状况(Güsewell, 2004; He et al., 2008)。目前, 对于植物化学计量特征的研究主要集中于不同器官的养分分配策略(Yan et al., 2016), 不同环境条件下受N、P限制的模式(Du et al., 2020), 以及植物器官中N、P含量的增长关系(陈婵等, 2016; Zhang et al., 2018b)等。但是, 目前对于演替过程中植物叶片化学计量特征的变异及其驱动因素的研究还较少(Yan et al., 2006)。
植物的化学计量特征可以反映植物的许多特性, 同时也受到周围环境状况的影响(Elser & Urabe, 1999)。以往的研究认为, 土壤化学计量特征是植物体内N、P含量的有效驱动因素(McGroddy et al., 2004), 例如, 在土壤中施加氮肥, 可以导致全球陆生植物内N含量平均增加28.5% (Xia & Wan, 2008)。生物多样性被认为是另一个影响植物化学计量特征的重要因素(Olde Venterink et al., 2003), 但是不同研究中多样性与植物体内N、P的化学计量特征关系并不一致。例如, Braakhekke和Hooftman (1999)研究发现, 一定地区内植物的多样性最高时, 该地区的植物体内N、P含量处于中等比例, 不会过高也不会过低, 而Bobbink等(2010)的研究则发现, 植物体内的单一化学计量与生物多样性呈负相关关系, 因此生物多样性与植物体内N、P含量的关系究竟如何, 还需进一步的研究。
目前对于陆地植物化学计量的研究主要集中于乔木(Cao et al., 2016)和草本(He et al., 2006), 对于灌木的研究较少(Zhang et al., 2018b)。灌木作为森林生态系统的重要组成部分, 在养分利用、土壤肥力改善、调节物种组成(Michalet et al., 2015)、提供栖息地(Boelman et al., 2015)等方面均具有重要作用, 是揭示森林结构的重要指标(Peña-Claros, 2003), 对于群落演替具有至关重要的作用(曹嘉瑜等, 2020)。对于演替进程中灌木化学计量特征的研究, 有助于我们全面地理解森林演替过程中N、P养分的变化。目前对北方温带森林不同演替阶段灌木化学计量特征的研究较少, 尤其是群落尺度上灌木层养分利用特征的研究尚鲜有报道。
阔叶红松(Pinus koraiensis)林是我国东北东部山区的地带性顶极森林植被, 是以红松为建群种的典型温带针阔混交林。本研究以黑龙江凉水国家级自然保护区内的阔叶红松林4个演替阶段群落的灌木叶片为研究对象, 分析其化学计量特征, 旨在探索: (1)不同演替阶段灌木叶片化学计量有何差异; (2)土壤N、P含量对个体尺度上灌木叶片化学计量的影响; (3)物种多样性和土壤化学性质与演替过程中灌木叶片化学计量变异的关系, 以期为理解阔叶红松林演替变化规律提供基础数据和科学依据。
1 材料和方法
1.1 研究区概况
本研究地位于黑龙江凉水国家级自然保护区(47.18° N, 128.89° E)。该区域属于低山丘陵地带, 具有鲜明的温带大陆性季风气候特征, 降水多集中于夏季, 其中6-8月占全年降水量的60%以上, 年降水量为676 mm, 年蒸发量为805 mm, 年平均气温为-0.3 ℃, 地带性土壤为暗棕壤, 占保护区面积的84.91% (徐丽娜和金光泽, 2012)。1.2 样地设置与取样方法
本研究样地为阔叶红松林不同演替阶段的4个森林群落, 根据树种组成和林龄划分为白桦(Betula platyphylla)次生林、落叶阔叶混交林、针阔混交林以及阔叶红松林(后文分别用演替阶段I、II、III、IV来表示)。在4个森林群落中各设置3个20 m × 20 m的样地, 每个样地间隔不小于20 m, 于2019年夏季对样地内所有胸径>1 cm的木本植物(包括乔木和灌木)进行每木调查、挂牌, 并在各样地内采集植物样品和土壤样品。植物样品的采集: 根据样地群落调查的结果(表1), 在每个样地中采集样方中出现的所有灌木种。每个灌木种选择3株大小相似、长势良好的标准木, 采集其未受病虫害的叶片, 并将每株标准木的叶片混合作为一份样品。
Table 1
表1
表1阔叶红松林不同演替阶段乔木和灌木组成
Table 1
| 演替阶段 Successional stage | 乔木组成 Tree composition | 灌木组成 Shrub composition |
|---|---|---|
| 演替阶段I (白桦次生林) Successional stage I (Secondary birch forest) | 5白桦; 2红松; 1兴安落叶松; 1水曲柳+春榆+色木槭+山杨+黄檗-枫桦-红皮云杉-裂叶榆-臭冷杉-胡桃楸-紫椴 5 Betula platyphylla; 2 Pinus koraiensis; 1 Larix gmelinii; 1 Fraxinus mandschurica + Ulmus davidiana var. japonica + Acer pictum subsp. mono + Populus davidiana + Phellodendron amurense - Betula costata - Picea koraiensis - Ulmus laciniata - Abies nephrolepis - Juglans mandshurica - Tilia amurensis | 7暴马丁香; 1东北山梅花; 1刺五加+珍珠梅+早花忍冬+毛榛子-光萼溲疏 7 Syringa reticulata subsp. amurensis; 1 Philadelphus schrenkii; 1 Acanthopanax senticosus + Sorbaria sorbifolia + Lonicera praeflorens + Corylus mandshurica - Deutzia glabrata |
| 演替阶段II (落叶阔叶混交林) Successional stage II (Mixed deciduous broad-leaved forest) | 4水曲柳; 2山杨; 1紫椴; 1色木槭; 1白桦+胡桃楸+鱼鳞云杉-春榆-红松-红皮云杉-青楷槭-稠李-裂叶榆-蒙古栎 4 Fraxinus mandschurica; 2 Populus davidiana; 1 Tilia amurensis; 1 Betula platyphylla + Juglans mandshurica + Picea jezoensis var. microsperma - Ulmus laciniata - Pinus koraiensis - Picea koraiensis - Acer tegmentosum - Padus racemose - Ulmus laciniata - Quercus mongolica | 7暴马丁香; 2毛榛; 1瘤枝卫矛+东北山梅花+刺五加+光萼溲疏-早花忍冬 7 Syringa reticulata subsp. amurensis; 2 Corylus mandshurica; 1 Euonymus verrucosus + Philadelphus schrenkii + Acanthopanax senticosus + Deutzia glabrata - Lonicera praeflorens |
| 演替阶段III (针阔混交林) Succession stage III (Mixed coniferous and broad-leaved forest) | 4臭冷杉; 4红松; 1春榆; 1红皮云杉+鱼鳞云杉+白桦+色木槭-花楷槭-枫桦-紫椴-胡桃楸-稠李 4 Abies nephrolepis; 4 Pinus koraiensis; 1 Ulmus davidiana var. japonica; 1 Picea koraiensis + Picea jezoensis var. microsperma + Betula platyphylla + Acer pictum subsp. mono - Acer ukurunduense - Betula costata - Tilia amurensis - Juglans mandshurica - Padus racemosa | 7暴马丁香; 2毛榛+瘤枝卫矛+早花忍冬-刺五加-光萼溲疏-珍珠梅 7 Syringa reticulata subsp. amurensis; 2 Corylus mandshurica + Euonymus verrucosus + Lonicera praeflorens - Acanthopanax senticosus - Deutzia glabrata - Sorbaria sorbifolia |
| 演替阶段IV (阔叶红松林) Succession stage IV (Mixed broadleaved-Korean pine forest) | 4红松; 2枫桦; 1色木槭; 1鱼鳞云杉; 1红皮云杉+青楷槭+紫椴+白桦+黄檗+裂叶榆-水曲柳-山杨-稠李-臭冷杉-花楷槭-春榆-胡桃楸 4 Pinus koraiensis; 2 Betula costata; 1 Acer pictum subsp. mono; 1 Picea jezoensis var. microsperma; 1 Picea koraiensis + Acer tegmentosum + Tilia amurensis + Betula platyphylla + Phellodendron amurense + Ulmus laciniata - Fraxinus mandschurica - Populus davidiana - Padus racemosa - Abies nephrolepis - Acer ukurunduense - Ulmus davidiana var. japonica - Juglans mandshurica | 2东北山梅花; 2瘤枝卫矛; 1暴马丁香; 1光萼溲疏; 1毛榛; 1早花忍冬; 1刺五加; 1龙牙楤木-绣线菊-东北茶藨子 2 Philadelphus schrenkii; 2 Euonymus verrucosus; 1 Syringa reticulata subsp. amurensis; 1 Deutzia glabrata; 1 Corylus mandshurica; 1 Lonicera praeflorens; 1 Acanthopanax senticosus; 1 Aralia elata - Spiraea salicifolia - Ribes mandshuricum |
新窗口打开|下载CSV
土壤样品的采集: 在每个样地中采用标准五点取样法进行土壤样品的采集, 在每个点取0-20 cm土层的土壤样品, 并去除其中的根系和凋落物。
1.3 样品测定
将采集到的所有样品在65 ℃下烘干至恒质量, 并研磨至通过100目筛。植物和土壤样品的全氮(TN)、全磷(TP)含量在经过浓H2SO4-H2O2消煮后采用AQ400间断分析仪(SEAL Analytical, Mequon, USA)进行测定, 土壤有机碳(SOC)含量采用multiN/ C3000碳氮元素分析仪(Analytik Jena AG, Jena, Germany)进行测定。1.4 数据处理
各样方灌木叶片的化学计量以异速生长方程为基础(李晓娜等, 2010), 利用生物量加权平均法(Zhang et al., 2018a)算出, 公式为:式中, Leaf Ecom表示各样方叶片的N或P含量(g·kg-1), En表示单个灌木叶片N或P含量(g·kg-1), Bn表示单个灌木叶生物量(kg), Bcom表示样方内灌木叶片总生物量(kg)。
不同演替阶段灌木叶片和土壤化学计量特征的差异利用SPSS 22.0中的单因素方差分析进行显著性检验, 若方差齐性, 则使用最小显著差数(LSD)测验法; 若方差非齐性, 则使用Tamhane's T2法进行多重比较, 显著性水平为α = 0.05, 并用Origin 2019制图。叶片与土壤、叶片与生物多样性的相关关系利用SPSS 22.0中的双变量相关分析方法进行分析, 并利用Microsoft Excel 2010制图。采用R 3.6.3的hier.part包(Nally & Walsh, 2004)对群落尺度上灌木叶片化学计量特征及物种多样性和土壤化学性质进行层次分割。
根据群落调查内容, 计算群落内所有胸径>1 cm的木本植物的物种多样性, 计算公式(方精云等, 2004)包括:
式中, Pi为物种i的个体数在全部个体数中的比例。
2 结果
2.1 阔叶红松林不同演替阶段灌木叶片化学计量特征
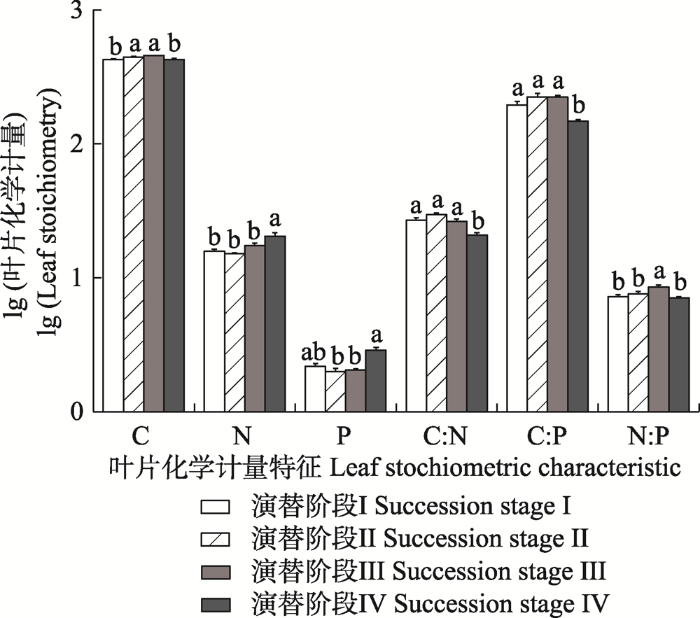
研究区内的阔叶红松林4个演替阶段的灌木叶片N含量的整体平均值为19.1 g·kg-1, 4个林型的平均值的范围为(15.14 ± 0.28)-(20.61 ± 1.37) g·kg-1; 叶片P含量的整体平均值为2.28 g·kg-1, 4个林型的平均值的范围为(2.00 ± 0.11)-(2.92 ± 0.14) g·kg-1。随着演替的进行, 灌木叶片的N、P含量趋势相近, 即演替阶段I、II、III中灌木叶片的N、P含量均无显著差异(p > 0.05), 演替阶段IV的N含量显著高于其他3种林型(p < 0.05), 而P含量与演替阶段I无显著差异(p > 0.05), 但显著高于其他两种林型(p < 0.05) (图1)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1阔叶红松林不同演替阶段群落尺度上叶片的化学计量特征(平均值±标准误)。不同小写字母表示叶片同一化学计量特征间差异显著(p < 0.05)。全碳(C)、全氮(N)、全磷(P)含量的单位为g·kg-1。
Fig. 1Leaf stoichiometric characteristics at community level in mixed broadleaved-Korean pine forests at different successional stages (mean ± SE). Different lowercase letters of the same organ indicate significant differences (p < 0.05). The unit of total carbon (C), total nitrogen (N) and total phosphorus (P) content is g·kg-1.
演替阶段IV的叶片C:N、C:P均显著低于其他3种林型(p < 0.05), 不同演替阶段的N:P的范围为(7.06 ± 0.15)-(8.56 ± 0.34), 其中演替阶段III的N:P显著高于其他3种林型(p < 0.05)。
2.2 土壤氮磷与个体尺度上的叶片化学计量特征的关系
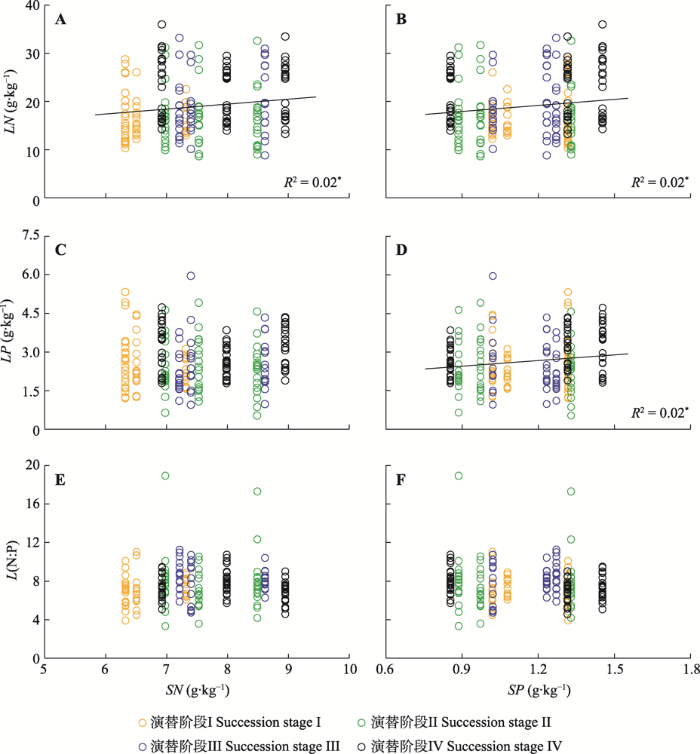
总体上, 相较于土壤N含量, 土壤P含量与个体尺度上的植物叶片的化学含量相关性更强, 土壤P含量与叶片N、P含量均呈显著正相关关系(p < 0.05), 土壤N含量与叶片N含量呈显著正相关关系(p < 0.05), 二者均与叶片N:P无显著相关关系(p > 0.05) (图2)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2阔叶红松林不同演替阶段土壤氮(N)、磷(P)含量与个体尺度上的叶片化学计量特征的关系。*, p < 0.05。LN, 叶片N含量; LP, 叶片P含量; L(N:P), 叶片N:P; SN, 土壤N含量; SP, 土壤P含量。
Fig. 2Relationships of soil nitrogen (N) and phosphorus (P) contents with leaf stoichiometric characteristics at individual scale in mixed broadleaved-Korean pine forests at different successional stages. *, p < 0.05. LN, leaf N content; LP, leaf P content; L(N:P), leaf N:P ratio; SN, soil N content; SP, soil P content.
2.3 物种多样性与土壤化学性质对群落尺度上叶片化学计量特征的影响
层次分割结果显示, 在群落水平上物种多样性和土壤化学性质共解释叶片N含量变异的82%和P含量变异的62%, 其中Shannon多样性指数分别解释叶片N含量变异的26.66%和叶片P含量变异的35.96%; 对于叶片C:N和叶片C:P的变异, 物种多样性和土壤化学性质共解释74%和59% (表2), 说明物种多样性和土壤化学性质对灌木叶片化学计量变异具有较高的解释率。Table 2
表2
表2物种多样性和土壤化学性质对群落尺度上阔叶红松林叶片碳(C)、氮(N)、磷(P)化学计量的层次分割结果
Table 2
| 叶片化学计量 Leaf stoichiometry | R² | 物种多样性解释度 Interpretation of species diversity (%) | 土壤化学性质解释度 Interpretation of soil chemical properties (%) | ||||
|---|---|---|---|---|---|---|---|
| H′ | S | J | SC | SN | SP | ||
| LC | 0.69 | 8.80 | 4.44 | 15.05 | 45.31* | 8.01 | 18.38 |
| LN | 0.82 | 26.66* | 36.11** | 4.94 | 5.73 | 5.14 | 21.42 |
| LP | 0.62 | 35.96* | 40.18* | 6.82 | 9.16 | 3.24 | 4.64 |
| L(C:N) | 0.74 | 28.88* | 33.41** | 4.51 | 13.25 | 6.52 | 13.43 |
| L(C:P) | 0.59 | 34.79* | 33.99* | 8.25 | 16.55 | 4.31 | 2.11 |
| L(N:P) | 0.46 | 22.12 | 16.76 | 23.11 | 9.23 | 3.60 | 25.17 |
新窗口打开|下载CSV
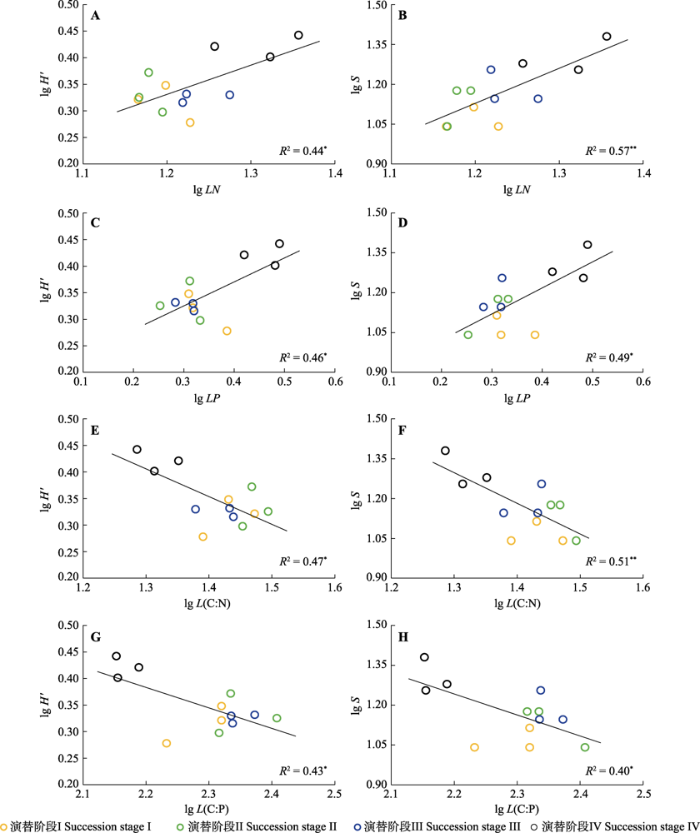
Shannon多样性指数和丰富度均与叶片N、P含量具有显著的正相关关系(p < 0.05)、与叶片C:N和叶片C:P具有显著的负相关关系(p < 0.05)(图3), 与叶片C含量无显著相关关系(p > 0.05)。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3阔叶红松林不同演替阶段叶片氮(N)、磷(P)含量、碳(C):N和N:P与Shannon多样性指数、物种丰富度的线性关系。*, p < 0.05; **, p < 0.01。H', Shannon多样性指数; S, 物种丰富度。LN, 叶片N含量; LP, 叶片P含量; L(C:N), 叶片C:N; L(C:P), 叶片C:P。
Fig. 3Linear relationships of leaf nitrogen (N) and phosphorus (P) contents, carbon (C):N ratio and C:P ratio with Shannon diversity index and Species richness in mixed broadleaved-Korean pine forest at different successional stages. *, p < 0.05; **, p < 0.01. H', Shannon diversity index; S, Species richness. LN, leaf N content; LP, leaf P content; L(C:N), leaf C:N ratio; L(C:P), leaf C:P ratio.
3 讨论
3.1 不同演替阶段灌木叶片化学计量特征的差异
本研究中, 阔叶红松林4个演替阶段的灌木叶片平均N含量为17.22 g·kg-1, 略低于我国灌木叶N平均含量19.1 g·kg-1; 而平均叶P含量为2.28 g·kg-1高于我国的灌木的平均值(1.11 g·kg-1)(Han et al., 2005), 这可能是因为本研究地所处的北方地区气候较为寒冷, 导致落叶灌木叶片寿命缩短、生长速率增高(van Ommen Kloeke et al., 2012), 根据生长速率假说, 生长速率更高的植物需要更多的P来维持植物体内RNA的高速合成, 对P的需求量更大(Güsewell & Koerselman, 2002; Sterner & Elser, 2002)。同时, 植物在这种环境中需要储存更多的脂质P在叶片中, 用以抵抗寒冷, 导致植物体内P含量高(Chapin III et al., 1986; Han et al., 2011)。植物器官的N:P通常可以作为判断植物受何种养分限制的指标(Aerts & Chapin III, 1999)。因为研究区域、研究植物群落有所不同, 不同研究提出作为区分N、P限制位点的N:P指标常常有所差异, 目前常用的指标认为N:P在14和16之间时, 植物不受N、P限制或者受N、P两种营养元素的共同制约; N:P < 14, 植物主要受N限制; N:P > 16, 植物主要受P限制(Koerselman & Meuleman, 1996)。
在本研究的4个演替阶段中灌木叶片的N:P均小于14。这表明本研究中4个演替阶段的灌木均受到N限制。以往的研究认为, 在缺乏灾难性干扰的条件下, 随着森林生态系统的自然演替, 森林生态系统将趋向于受到P限制(Wardle et al., 2004; 刘兴诏等, 2010), 这与本文的研究结果并不相符, 这有两个可能的原因: (1)本研究地区属于温带森林, 年平均气温低, 土壤发育晚(Reich & Oleksyn, 2004), 土壤P的可利用性更高(McGroddy et al., 2004), 所以导致植物P含量较高, 由于植物体内N:P的变化主要是由P浓度的变化所驱动(Tian et al., 2018), 因此灌木叶片表现出低N:P的特征; (2)相对于乔木, 灌木体型小, 生长速率高(Koerselman & Meuleman, 1996), 通常表现出较低的N:P (Makino et al., 2003)。
3.2 土壤氮磷对个体尺度上的灌木叶片化学计量特征的影响
本研究中的4个森林类型土壤的平均N:P为6.73, 低于我国土壤平均N:P (9.3)(Tian et al., 2010), 表明本研究地区土壤N含量相对于P含量而言处于较低的状态。土壤P含量与个体尺度上叶片N、P含量均表现出显著的相关性(p < 0.05), 而土壤N含量则只与叶片N含量具有显著相关性(p < 0.05)。这可能与土壤N的有效性不足时, 植物通过增加吸收土壤中的P来促进体内P的有效性, 进而刺激N吸收有关(Sistla et al., 2015), 因此土壤P含量对个体尺度上的叶片N、P含量均表现出一定的影响。同时, 土壤N含量与个体水平上的叶片N含量显著相关表明, 即使土壤中N含量相对较低, 从土壤中吸收N仍是植物获取N的主要方式。这是因为比起从空气中和衰老叶片中获取N, 植物从土壤中获取N的成本更低、更便捷(Chen et al., 2015; Zheng et al., 2020)。
3.3 物种多样性和土壤碳氮磷对群落尺度上的叶片化学计量特征的影响
土壤化学性质与群落尺度上的叶片化学计量无显著相关关系(表2), 这可能与白桦次生林、落叶阔叶混交林和针阔混交林中具有单一优势灌木——暴马丁香(Syringa reticulata var. amurensis)有关, 即群落中存在单一优势物种时, 此物种的N、P含量对群落尺度的N、P含量具有重要影响, 而在本研究中, 暴马丁香N、P含量虽然随着土壤N、P含量的增加而具有增加的趋势, 但二者之间无显著相关关系(p > 0.05, 未发表数据), 导致土壤化学性质在群落尺度上与叶片化学计量无显著相关关系。群落尺度上的植物和土壤N、P含量的关系也很好地体现了生态化学计量学的内稳态理论, 即生物为保持整个有机体的稳定, 在长期的进化过程中形成了在外界环境变化时保持其化学含量相对稳定的能力(Sterner & Elser, 2002)。物种多样性与群落尺度上灌木叶片的N、P含量呈显著正相关关系(p < 0.05, 图3)。这可以用以下两个原因解释: (1)随着物种多样性的增加, 灌木所面临的光资源的竞争增大, 需要获取更多的营养物质供给叶片用于合成光合作用所需的酶、核糖体等物质, 来维持个体的生存和繁殖(Evans & Poorter, 2001); (2)物种多样性的增加使得灌木可以更彻底地利用有限的资源, 以此增加自身的N、P含量(Roscher et al., 2008)。同时, 随着物种多样性的增加, 相邻树种养分获取策略互补的可能性增加, 这种互补性可以促进植物对养分的吸收(Teste et al., 2014)。由于C在植物内的含量较高且变异较小(Reich & Oleksyn, 2004), 灌木叶片内的C含量与植物多样性无显著相关关系(p > 0.05), 因此随着植物多样性的增加, 植物叶片内C:N和C:P均降低(p < 0.05)。
本研究探讨了阔叶红松林4种演替阶段灌木的化学计量特征。结果显示在群落尺度上4种演替阶段的灌木均受到N限制; 相较于土壤的化学性质, 物种多样性更好地解释了灌木化学计量的变异。为了更好地理解不同演替阶段中物种化学计量的变异规律, 在今后的研究中需要进一步开展对于不同演替阶段中的种间、种内变异的研究。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.1890/08-1140.1URL [本文引用: 1]
DOI:10.1111/gcb.12761URL [本文引用: 1]
DOI:10.2307/3237140URL [本文引用: 1]
DOI:10.17521/cjpe.2020.0024URL [本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
DOI:10.2307/2260357URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/treephys/tpv076URL [本文引用: 1]
DOI:10.1038/s41561-019-0530-4URL [本文引用: 1]
DOI:10.1890/0012-9658(1999)080[0735:TSOCDN]2.0.CO;2URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.17520/biods.2004002 [本文引用: 1]

我国是个多山的国家,拥有丰富的生物多样性资源。为研究我国山地的植物物种多样性垂直格局及其地理分异,北京大学自20世纪90年代中期开始,实施了中国山地植物物种多样性调查计划(Peking University′s Survey Plan for Plant Species Diversity of China′s Mountains, 简称PKU-PSD 计划)。本文简要介绍该计划的主要研究内容、研究的山地以及野外调查和数据分析方法,试图为同类研究提供参考。
DOI:10.1111/j.1469-8137.2004.01192.xPMID:33873556 [本文引用: 1]
Nitrogen (N) and phosphorus (P) availability limit plant growth in most terrestrial ecosystems. This review examines how variation in the relative availability of N and P, as reflected by N : P ratios of plant biomass, influences vegetation composition and functioning. Plastic responses of plants to N and P supply cause up to 50-fold variation in biomass N : P ratios, associated with differences in root allocation, nutrient uptake, biomass turnover and reproductive output. Optimal N : P ratios - those of plants whose growth is equally limited by N and P - depend on species, growth rate, plant age and plant parts. At vegetation level, N : P ratios <10 and >20 often (not always) correspond to N- and P-limited biomass production, as shown by short-term fertilization experiments; however long-term effects of fertilization or effects on individual species can be different. N : P ratios are on average higher in graminoids than in forbs, and in stress-tolerant species compared with ruderals; they correlate negatively with the maximal relative growth rates of species and with their N-indicator values. At vegetation level, N : P ratios often correlate negatively with biomass production; high N : P ratios promote graminoids and stress tolerators relative to other species, whereas relationships with species richness are not consistent. N : P ratios are influenced by global change, increased atmospheric N deposition, and conservation managment. Contents Summary 243 I Introduction 244 II Variability of N : P ratios in response to nutrient supply 244 III Critical N : P ratios as indicators of nutrient limitation 248 IV Interspecific variation in N : P ratios 252 V Vegetation properties in relation to N : P ratios 255 VI Implications of N : P ratios for human impacts on ecosystems 258 VII Conclusions 259 Acknowledgements 259 References 260.
DOI:10.1078/1433-8319-0000022URL [本文引用: 1]
DOI:10.1111/nph.2005.168.issue-2URL [本文引用: 1]
DOI:10.1111/j.1461-0248.2011.01641.xPMID:21692962 [本文引用: 1]
Understanding variation of plant nutrients is largely limited to nitrogen and to a lesser extent phosphorus. Here we analyse patterns of variation in 11 elements (nitrogen/phosphorus/potassium/calcium/magnesium/sulphur/silicon/iron/sodium/manganese/aluminium) in leaves of 1900 plant species across China. The concentrations of these elements show significant latitudinal and longitudinal trends, driven by significant influences of climate, soil and plant functional type. Precipitation explains more variation than temperature for all elements except phosphorus and aluminium, and the 11 elements differentiate in relation to climate, soil and functional type. Variability (assessed as the coefficient of variation) and environmental sensitivity (slope of responses to environmental gradients) are lowest for elements that are required in the highest concentrations, most abundant and most often limiting in nature (the Stability of Limiting Elements Hypothesis). Our findings can help initiate a more holistic approach to ecological plant nutrition and lay the groundwork for the eventual development of multiple element biogeochemical models.© 2011 Blackwell Publishing Ltd/CNRS.
DOI:10.1007/s00442-006-0425-0URL [本文引用: 1]
[本文引用: 1]
DOI:10.3773/j.issn.1005-264x.2010.01.002 [本文引用: 1]
DOI:10.1007/s00442-007-0912-yURL [本文引用: 1]
DOI:10.2307/2404783URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3773/j.issn.1005-264x.2010.01.010 [本文引用: 1]
选择南亚热带森林演替过程3个阶段(初期、中期和后期)的典型森林生态系统为研究对象, 在测定植物与土壤中全N、全P含量的基础上, 阐明了森林演替过程中植物与土壤的N、P化学计量特征。结果显示: 1)土壤中全N含量随演替进行而增加, 马尾松(Pinus massoniana)林(初期)、混交林(中期)和季风林(后期) 0–10 cm土层中全N含量分别为0.440、0.843和1.023 g·kg<sup>–1</sup>; 混交林0–10 cm土层中全P的含量最为丰富, 为0.337 g·kg<sup>–1</sup>, 马尾松林和季风林土壤全P含量分别为0.190和0.283 g·kg<sup>–1</sup>。2)植物叶片中全N、全P的含量随演替呈减少的趋势, 但根系中全N、全P的含量都以马尾松林为最多, 混交林和季风林含量彼此相当。3)各土层中N:P随演替的进行呈现明显增加趋势, 马尾松林、混交林和季风林0–10 cm土层中N:P分别为2.3、2.5和3.6; 植物各器官中N:P随演替的进行也呈增加趋势, 且叶片和根系中的N:P相近, 马尾松林、混交林和季风林叶片中N:P分别为22.7、25.3和29.6。基于上述结果, 探讨了南亚热带森林生态系统植物与土壤中N:P特征、森林演替过程中植物与土壤中N:P变化规律以及P对南亚热带森林生态系统的限制作用。结果表明, P已经成为南亚热带森林生态系统生物生长和重要生态过程的限制因子。
DOI:10.1046/j.1365-2435.2003.00712.xURL [本文引用: 1]
DOI:10.1890/03-0351URL [本文引用: 2]
DOI:10.1111/oik.2015.v124.i9URL [本文引用: 1]
DOI:10.1023/B:BIOC.0000009515.11717.0bURL [本文引用: 1]
DOI:10.1890/01-0639URL [本文引用: 1]
DOI:10.1111/btp.2003.35.issue-4URL [本文引用: 1]
[本文引用: 2]
PMID:9391094 [本文引用: 1]
Despite striking differences in climate, soils, and evolutionary history among diverse biomes ranging from tropical and temperate forests to alpine tundra and desert, we found similar interspecific relationships among leaf structure and function and plant growth in all biomes. Our results thus demonstrate convergent evolution and global generality in plant functioning, despite the enormous diversity of plant species and biomes. For 280 plant species from two global data sets, we found that potential carbon gain (photosynthesis) and carbon loss (respiration) increase in similar proportion with decreasing leaf life-span, increasing leaf nitrogen concentration, and increasing leaf surface area-to-mass ratio. Productivity of individual plants and of leaves in vegetation canopies also changes in constant proportion to leaf life-span and surface area-to-mass ratio. These global plant functional relationships have significant implications for global scale modeling of vegetation-atmosphere CO2 exchange.
DOI:10.1111/j.1365-2745.2008.01353.xURL [本文引用: 1]
DOI:10.1111/oik.2015.v124.i7URL [本文引用: 2]
[本文引用: 3]
URL [本文引用: 1]
DOI:10.1111/fec.2014.28.issue-4URL [本文引用: 1]
DOI:10.1093/nsr/nwx142URL [本文引用: 1]
DOI:10.1007/s10533-009-9382-0URL [本文引用: 1]
DOI:10.1111/j.1466-8238.2011.00667.xURL [本文引用: 1]
PMID:15205475 [本文引用: 1]
During succession, ecosystem development occurs; but in the long-term absence of catastrophic disturbance, a decline phase eventually follows. We studied six long-term chronosequences, in Australia, Sweden, Alaska, Hawaii, and New Zealand; for each, the decline phase was associated with a reduction in tree basal area and an increase in the substrate nitrogen-to-phosphorus ratio, indicating increasing phosphorus limitation over time. These changes were often associated with reductions in litter decomposition rates, phosphorus release from litter, and biomass and activity of decomposer microbes. Our findings suggest that the maximal biomass phase reached during succession cannot be maintained in the long-term absence of major disturbance, and that similar patterns of decline occur in forested ecosystems spanning the tropical, temperate, and boreal zones.
DOI:10.1111/nph.2008.179.issue-2URL [本文引用: 1]
DOI:10.3724/SP.J.1003.2012.12233 [本文引用: 1]
The mixed broadleaved-Korean pine (<em>Pinus koraiensis</em>) forest (MBKF) represents the climax vegetation type of the eastern mountainous area of Northeast China. It is divided into three categories according to community structure and species composition, including southern MBKF, typical MBKF, and northern MBKF. To monitor long-term dynamics of the typical MBKF, we established a 9-ha plot following the field protocol of the 50 ha plot in Panama (Barro Colorado Island, BCI) in 2005 in Liangshui National Nature Reserve of Heilongjiang Province. All free-standing plant species with DBH (diameter at breast height) ≥1 cm were mapped, tagged, and identified to species in the plot. In the 2010 census, we documented 48 woody species with 21,355 individuals, belonging to 34 genera and 20 families. Most of the species belong to the Changbai Mountain plant flora, with a minor subtropical plant species component. The DBH distribution of all individuals showed a reversed “J” type, indicating well regenerated for the community. The DBH distribution for the canopy, sub-canopy, and middle layers and the understory layer showed the reversed "J” and “L” distributions, respectively. The DBH pattern of the nine dominant species can be classified into three categories: approximate normal, reversed “J”, and “L” distribution. Spatial pattern analysis showed that the major tree species were aggregated, and aggregation intensity decreased as DBH increased. Further analysis showed that spatial distribution was closely related to topography and varied with DBH classes. For example, the distribution of <em>Pinus koraiensis</em> and <em>Tilia amurensis</em> was significantly affected by topography (<em>P</em> < 0.05); however, the distribution of <em>Abies nephrolepis</em>, <em>Acer ukurunduense</em>, and <em>Ulmus laciniata</em> correlated significantly with topography at diameter classes of I (DBH < 10 cm) and II (10 cm ≤ DBH < 30 cm). The distribution of <em>Betula costata</em> and <em>Acer tegmentosum</em> was also significantly correlated with topography at a diameter class I (<em>P</em> < 0.05). Overall, the impact of the topographic gradient on species distribution decreased significantly as DBH class increased.
DOI:10.3724/SP.J.1003.2012.12233 [本文引用: 1]
阔叶红松(Pinus koraiensis)林是我国东北东部山区的地带性顶极植被, 按其群落特征和物种组成可分为南部红松林、典型红松林和北部红松林。依照BCI(Barro Colorado Island)50 ha样地的技术规范, 作者于2005年在典型红松林分布的黑龙江凉水国家级自然保护区建立了一块9 ha的固定监测样地, 并于2010年对样地内胸径 ≥ 1 cm的木本植物进行了全面调查。结果表明, 样地内的木本植物共有48种, 独立个体数为21,355株(包括分枝数为34,021棵), 隶属于20科34属。绝大部分种类属于长白山区系小兴安岭亚系, 同时混生有一些亚热带成分。样地内所有个体的径级分布呈倒“J”型, 群落自我更新良好。林冠层、亚冠层和中间层的径级分布均呈倒“J”型, 林下层呈“L”型。主要树种大青杨(Populus ussuriensis)、红松、枫桦(Betula costata)、水曲柳(Fraxinus mandshurica)、红皮云杉(Picea koraiensis)等的径级结构可分为近似于“正态”型、倒“J”型和“L”型3种类型。主要树种在样地的空间分布大多呈聚集分布, 但大部分物种随着径级的增加聚集程度变小。树种分布与地形紧密关联, 不同物种在不同径级表现出对生境有不同的偏好; 红松和紫椴(Tilia amurensis)各径级的分布均与地形显著相关(P Abies nephrolepis)、花楷槭(Acer ukurunduense)、裂叶榆(Ulmus laciniata)、色木槭(Acer mono)的径级I(DBH Acer tegmentosum)的径级I的分布与地形显著相关(P < 0.05), 且随着径级的增加, 地形因子对其分布的影响显著减小。
DOI:10.1007/s11104-006-9128-xURL [本文引用: 1]
URL [本文引用: 1]
DOI:10.1111/gcb.v26.4URL [本文引用: 1]
DOI:10.1111/fec.2018.32.issue-1URL [本文引用: 1]
DOI:10.1007/s11104-018-3655-0URL [本文引用: 3]
DOI:10.1111/ele.v23.2URL [本文引用: 1]
The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns
1
1999
... 植物器官的N:P通常可以作为判断植物受何种养分限制的指标(Aerts & Chapin III,
Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis
1
2010
... 植物的化学计量特征可以反映植物的许多特性, 同时也受到周围环境状况的影响(Elser & Urabe,
Greater shrub dominance alters breeding habitat and food resources for migratory songbirds in Alaskan arctic tundra
1
2015
... 目前对于陆地植物化学计量的研究主要集中于乔木(Cao et al.,
The resource balance hypothesis of plant species diversity in grassland
1
1999
... 植物的化学计量特征可以反映植物的许多特性, 同时也受到周围环境状况的影响(Elser & Urabe,
森林与灌丛的灌木性状揭示不同的生活策略
1
2020
... 目前对于陆地植物化学计量的研究主要集中于乔木(Cao et al.,
森林与灌丛的灌木性状揭示不同的生活策略
1
2020
... 目前对于陆地植物化学计量的研究主要集中于乔木(Cao et al.,
Ecological stoichiometric characteristics and element reserves of three stands in a closed forest on the Chinese loess plateau
1
2016
... 目前对于陆地植物化学计量的研究主要集中于乔木(Cao et al.,
Environmental controls over carbon, nitrogen and phosphorus fractions in Eriophorum vaginatum in Alaskan tussock tundra
1
1986
... 本研究中, 阔叶红松林4个演替阶段的灌木叶片平均N含量为17.22 g·kg-1, 略低于我国灌木叶N平均含量19.1 g·kg-1; 而平均叶P含量为2.28 g·kg-1高于我国的灌木的平均值(1.11 g·kg-1)(Han et al.,
会同杉木器官间C、N、P化学计量比的季节动态与异速生长关系
1
2016
... 植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al.,
会同杉木器官间C、N、P化学计量比的季节动态与异速生长关系
1
2016
... 植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al.,
Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age.
1
2015
... 同时, 土壤N含量与个体水平上的叶片N含量显著相关表明, 即使土壤中N含量相对较低, 从土壤中吸收N仍是植物获取N的主要方式.这是因为比起从空气中和衰老叶片中获取N, 植物从土壤中获取N的成本更低、更便捷(Chen et al.,
Global patterns of terrestrial nitrogen and phosphorus limitation
1
2020
... 植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al.,
The stoichiometry of consumer-driven nutrient recycling: theory, observations, and consequences
1
1999
... 植物的化学计量特征可以反映植物的许多特性, 同时也受到周围环境状况的影响(Elser & Urabe,
Photosynthetic acclimation of plants to growth irradiance: the relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain
1
2001
... 物种多样性与群落尺度上灌木叶片的N、P含量呈显著正相关关系(p < 0.05,
“中国山地植物物种多样性调查计划”及若干技术规范
1
2004
... 根据群落调查内容, 计算群落内所有胸径>1 cm的木本植物的物种多样性, 计算公式(方精云等,
“中国山地植物物种多样性调查计划”及若干技术规范
1
2004
... 根据群落调查内容, 计算群落内所有胸径>1 cm的木本植物的物种多样性, 计算公式(方精云等,
N:P ratios in terrestrial plants: variation and functional significance
1
2004
... 植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al.,
Variation in nitrogen and phosphorus concentrations of wetland plants
1
2002
... 本研究中, 阔叶红松林4个演替阶段的灌木叶片平均N含量为17.22 g·kg-1, 略低于我国灌木叶N平均含量19.1 g·kg-1; 而平均叶P含量为2.28 g·kg-1高于我国的灌木的平均值(1.11 g·kg-1)(Han et al.,
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
1
2005
... 本研究中, 阔叶红松林4个演替阶段的灌木叶片平均N含量为17.22 g·kg-1, 略低于我国灌木叶N平均含量19.1 g·kg-1; 而平均叶P含量为2.28 g·kg-1高于我国的灌木的平均值(1.11 g·kg-1)(Han et al.,
Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China
1
2011
... 本研究中, 阔叶红松林4个演替阶段的灌木叶片平均N含量为17.22 g·kg-1, 略低于我国灌木叶N平均含量19.1 g·kg-1; 而平均叶P含量为2.28 g·kg-1高于我国的灌木的平均值(1.11 g·kg-1)(Han et al.,
Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China
1
2006
... 目前对于陆地植物化学计量的研究主要集中于乔木(Cao et al.,
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... 作为生物体基本的组成元素, 氮(N)和磷(P)被认为是陆地生态系统中植物体内重要的限制因子(Reich et al.,
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... 作为生物体基本的组成元素, 氮(N)和磷(P)被认为是陆地生态系统中植物体内重要的限制因子(Reich et al.,
Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes
1
2008
... 植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al.,
The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation
2
1996
... 植物器官的N:P通常可以作为判断植物受何种养分限制的指标(Aerts & Chapin III,
... 在本研究的4个演替阶段中灌木叶片的N:P均小于14.这表明本研究中4个演替阶段的灌木均受到N限制.以往的研究认为, 在缺乏灾难性干扰的条件下, 随着森林生态系统的自然演替, 森林生态系统将趋向于受到P限制(Wardle et al.,
东北天然次生林下木树种生物量的相对生长
1
2010
... 各样方灌木叶片的化学计量以异速生长方程为基础(李晓娜等,
东北天然次生林下木树种生物量的相对生长
1
2010
... 各样方灌木叶片的化学计量以异速生长方程为基础(李晓娜等,
南亚热带森林不同演替阶段植物与土壤中N、P的化学计量特征
1
2010
... 在本研究的4个演替阶段中灌木叶片的N:P均小于14.这表明本研究中4个演替阶段的灌木均受到N限制.以往的研究认为, 在缺乏灾难性干扰的条件下, 随着森林生态系统的自然演替, 森林生态系统将趋向于受到P限制(Wardle et al.,
南亚热带森林不同演替阶段植物与土壤中N、P的化学计量特征
1
2010
... 在本研究的4个演替阶段中灌木叶片的N:P均小于14.这表明本研究中4个演替阶段的灌木均受到N限制.以往的研究认为, 在缺乏灾难性干扰的条件下, 随着森林生态系统的自然演替, 森林生态系统将趋向于受到P限制(Wardle et al.,
Are bacteria more like plants or animals? Growth rate and resource dependence of bacterial C:N:P stoichiometry
1
2003
... 在本研究的4个演替阶段中灌木叶片的N:P均小于14.这表明本研究中4个演替阶段的灌木均受到N限制.以往的研究认为, 在缺乏灾难性干扰的条件下, 随着森林生态系统的自然演替, 森林生态系统将趋向于受到P限制(Wardle et al.,
Scaling of C:N:P stoichiometry in forests worldwide: implications of terrestrial redfield-type ratios
2
2004
... 植物的化学计量特征可以反映植物的许多特性, 同时也受到周围环境状况的影响(Elser & Urabe,
... 在本研究的4个演替阶段中灌木叶片的N:P均小于14.这表明本研究中4个演替阶段的灌木均受到N限制.以往的研究认为, 在缺乏灾难性干扰的条件下, 随着森林生态系统的自然演替, 森林生态系统将趋向于受到P限制(Wardle et al.,
Disentangling direct and indirect effects of a legume shrub on its understorey community
1
2015
... 目前对于陆地植物化学计量的研究主要集中于乔木(Cao et al.,
Hierarchical partitioning public- domain software
1
2004
... 不同演替阶段灌木叶片和土壤化学计量特征的差异利用SPSS 22.0中的单因素方差分析进行显著性检验, 若方差齐性, 则使用最小显著差数(LSD)测验法; 若方差非齐性, 则使用Tamhane's T2法进行多重比较, 显著性水平为α = 0.05, 并用Origin 2019制图.叶片与土壤、叶片与生物多样性的相关关系利用SPSS 22.0中的双变量相关分析方法进行分析, 并利用Microsoft Excel 2010制图.采用R 3.6.3的hier.part包(Nally & Walsh,
Species richness-productivity patterns differ between N-, P-, and K-limited wetlands
1
2003
... 植物的化学计量特征可以反映植物的许多特性, 同时也受到周围环境状况的影响(Elser & Urabe,
Changes in forest structure and species composition during secondary forest succession in the Bolivian Amazon
1
2003
... 目前对于陆地植物化学计量的研究主要集中于乔木(Cao et al.,
Global patterns of plant leaf N and P in relation to temperature and latitude
2
2004
... 在本研究的4个演替阶段中灌木叶片的N:P均小于14.这表明本研究中4个演替阶段的灌木均受到N限制.以往的研究认为, 在缺乏灾难性干扰的条件下, 随着森林生态系统的自然演替, 森林生态系统将趋向于受到P限制(Wardle et al.,
... 物种多样性与群落尺度上灌木叶片的N、P含量呈显著正相关关系(p < 0.05,
From tropics to tundra: global convergence in plant functioning
1
1997
... 作为生物体基本的组成元素, 氮(N)和磷(P)被认为是陆地生态系统中植物体内重要的限制因子(Reich et al.,
Complementary nitrogen use among potentially dominant species in a biodiversity experiment varies between two years
1
2008
... 物种多样性与群落尺度上灌木叶片的N、P含量呈显著正相关关系(p < 0.05,
Stoichiometric flexibility in response to fertilization along gradients of environmental and organismal nutrient richness
2
2015
... 植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al.,
... 本研究中的4个森林类型土壤的平均N:P为6.73, 低于我国土壤平均N:P (9.3)(Tian et al.,
3
2002
... 植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al.,
... 本研究中, 阔叶红松林4个演替阶段的灌木叶片平均N含量为17.22 g·kg-1, 略低于我国灌木叶N平均含量19.1 g·kg-1; 而平均叶P含量为2.28 g·kg-1高于我国的灌木的平均值(1.11 g·kg-1)(Han et al.,
... 土壤化学性质与群落尺度上的叶片化学计量无显著相关关系(
Responses and drivers of leaf nutrients and resorption to nitrogen enrichment across northern China's grasslands: a meta-analysis
1
2021
... 作为生物体基本的组成元素, 氮(N)和磷(P)被认为是陆地生态系统中植物体内重要的限制因子(Reich et al.,
Complementary plant nutrient-acquisition strategies promote growth of neighbour species
1
2014
... 物种多样性与群落尺度上灌木叶片的N、P含量呈显著正相关关系(p < 0.05,
Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent
1
2018
... 在本研究的4个演替阶段中灌木叶片的N:P均小于14.这表明本研究中4个演替阶段的灌木均受到N限制.以往的研究认为, 在缺乏灾难性干扰的条件下, 随着森林生态系统的自然演替, 森林生态系统将趋向于受到P限制(Wardle et al.,
Pattern and variation of C:N:P ratios in China's soils: a synthesis of observational data
1
2010
... 本研究中的4个森林类型土壤的平均N:P为6.73, 低于我国土壤平均N:P (9.3)(Tian et al.,
Global quantification of contrasting leaf life span strategies for deciduous and evergreen species in response to environmental conditions
1
2012
... 本研究中, 阔叶红松林4个演替阶段的灌木叶片平均N含量为17.22 g·kg-1, 略低于我国灌木叶N平均含量19.1 g·kg-1; 而平均叶P含量为2.28 g·kg-1高于我国的灌木的平均值(1.11 g·kg-1)(Han et al.,
Ecosystem properties and forest decline in contrasting long-term chronosequences
1
2004
... 在本研究的4个演替阶段中灌木叶片的N:P均小于14.这表明本研究中4个演替阶段的灌木均受到N限制.以往的研究认为, 在缺乏灾难性干扰的条件下, 随着森林生态系统的自然演替, 森林生态系统将趋向于受到P限制(Wardle et al.,
Global response patterns of terrestrial plant species to nitrogen addition
1
2008
... 植物的化学计量特征可以反映植物的许多特性, 同时也受到周围环境状况的影响(Elser & Urabe,
小兴安岭凉水典型阔叶红松林动态监测样地: 物种组成与群落结构
1
2012
... 本研究地位于黑龙江凉水国家级自然保护区(47.18° N, 128.89° E).该区域属于低山丘陵地带, 具有鲜明的温带大陆性季风气候特征, 降水多集中于夏季, 其中6-8月占全年降水量的60%以上, 年降水量为676 mm, 年蒸发量为805 mm, 年平均气温为-0.3 ℃, 地带性土壤为暗棕壤, 占保护区面积的84.91% (徐丽娜和金光泽,
小兴安岭凉水典型阔叶红松林动态监测样地: 物种组成与群落结构
1
2012
... 本研究地位于黑龙江凉水国家级自然保护区(47.18° N, 128.89° E).该区域属于低山丘陵地带, 具有鲜明的温带大陆性季风气候特征, 降水多集中于夏季, 其中6-8月占全年降水量的60%以上, 年降水量为676 mm, 年蒸发量为805 mm, 年平均气温为-0.3 ℃, 地带性土壤为暗棕壤, 占保护区面积的84.91% (徐丽娜和金光泽,
Shifts in plant nutrient use strategies under secondary forest succession
1
2006
... 植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al.,
Nutrient allocation strategies of woody plants: an approach from the scaling of nitrogen and phosphorus between twig stems and leaves
1
2016
... 植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al.,
Variation and evolution of C:N ratio among different organs enable plants to adapt to N-limited environments
1
2020
... 作为生物体基本的组成元素, 氮(N)和磷(P)被认为是陆地生态系统中植物体内重要的限制因子(Reich et al.,
C:N:P stoichiometry in China's forests: from organs to ecosystems
1
2018a
... 各样方灌木叶片的化学计量以异速生长方程为基础(李晓娜等,
Nitrogen and phosphorus concentrations and allocation strategies among shrub organs: the effects of plant growth forms and nitrogen-fixation types
3
2018b
... 植物体内的化学计量特征既受到一定的限制, 又会展现出一定的灵活性(Sistla et al.,
... .,
... 目前对于陆地植物化学计量的研究主要集中于乔木(Cao et al.,
Substrate stoichiometry determines nitrogen fixation throughout succession in southern Chinese forests
1
2020
... 同时, 土壤N含量与个体水平上的叶片N含量显著相关表明, 即使土壤中N含量相对较低, 从土壤中吸收N仍是植物获取N的主要方式.这是因为比起从空气中和衰老叶片中获取N, 植物从土壤中获取N的成本更低、更便捷(Chen et al.,

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}