淳雁, 李学勇*, 中国农业科学院作物科学研究所/农作物基因资源与基因改良国家重大科学工程, 北京 100081 ChunYan, LiXueyong*, National Key Facility for Crop Gene Resource and Genetic Improvement/Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China 引用本文 淳雁, 李学勇. 水稻穗型的遗传调控研究进展. , 2017, 52(1): 19-29
Abstract: Panicle architecture, as a key agronomic trait of rice (Oryza sativa), has been a research hot topic in recent years. In this review, we introduce the general development progress of a rice panicle. Genes related to rice panicles are summarized and are classified into 4 categories according to their function in panicle development: regulating initiation of meristem, meristem size, transition from branch meristem to spikelet meristem and elongation of branches. Furthermore, we summarize emerging networks of genes and pathways. Finally, we discuss the problems and prospects of future research into the genetic regulation of rice panicle.
张淑红 (2005). 水稻中控制形态结构建成相关基因的功能研究. 博士论文. 上海: 复旦大学. pp. 72-78.
[本文引用: 1]
[7]
AbeY, MiedaK, AndoT, KonoI, YanoM, KitanoH, IwasakiY (2010). The SMALL AND ROUND SEED1 (SRS1/ DEP2) gene is involved in the regulation of seed size in rice.Genes Genet Syst 85, 327-339. [本文引用: 1]
[8]
AriteT, UmeharaM, IshikawaS, HanadaA, MaekawaM, YamaguchiS, KyozukaJ (2009). d14, a strigolactone- insensitive mutant of rice, shows an accelerated outgrowth of tillers.Plant Cell Physiol 50, 1416-1424. [本文引用: 1]
BenjaminsR, ScheresB (2008). Auxin: the looping star in plant development.Annu Rev Plant Biol 59, 443-465. [本文引用: 1]
[11]
CarmonaMJ, CalonjeM, Martinez-ZapaterJM (2007). The FT/TFL1 gene family in grapevine.Plant Mol Biol 63, 637-650. [本文引用: 1]
[12]
ChengYF, ZhaoYD (2007). A role for auxin in flower development.J Integr Plant Biol 49, 99-104. [本文引用: 1]
[13]
ChuckG, MuszynskiMG, KelloggEA, HakeS, SchmidtRJ (2002). The control of spikelet meristem identity by the branched silkless1 gene in maize.Science 298, 1238-1241. [本文引用: 1]
[14]
DanilevskayaON, MengX, AnanievEV (2010). Concerted modification of flowering time and inflorescence architecture by ectopic expression of TFL1-like genes in maize.Plant Physiol 153, 238-251. [本文引用: 1]
[15]
DelkerC, RaschkeA, QuintM (2008). Auxin dynamics: the dazzling complexity of a small molecule’s message.Planta 227, 929-941. [本文引用: 1]
[16]
FerrándizC, GuQ, MartienssenR, YanofskyMF (2000). Redundant regulation of meristem identity and plant architecture by FRUITFULL, APETALA1, and CAULIFLO- WER.Development 127, 725-734. [本文引用: 1]
[17]
HuangXZ, QianQ, LiuZB, SunHY, HeSY, LuoD, XiaGM, ChuCC, LiJY, FuXD (2009). Natural variation at the DEP1 locus enhances grain yield in rice.Nat Genet 41, 494-497. [本文引用: 2]
IkedaK, SunoharaH, NagatoY (2004). Development course of inflorescence and spikelet in rice.Breeding Sci 54, 147-156. [本文引用: 1]
[20]
IkedakawakatsuK, MaekawaM, IzawaT, ItohJ, NagatoY (2012). ABERRANT PANICLE ORGANIZATION 2/RFL, the rice ortholog of Arabidopsis LEAFY, suppresses the transition from inflorescence meristem to floral meristem through interaction with APO1.Plant J 69, 168-180. [本文引用: 1]
[21]
IshikawaS, MaekawaM, AriteT, OnishiK, TakamureI, KyozukaJ (2005). Suppression of tiller bud activity in tillering dwarf mutants of rice.Plant Cell Physiol 46, 79-86. [本文引用: 1]
KatoT, HoribataA (2012). A novel frameshift mutant allele, fzp-10, affecting the panicle architecture of rice.Euphytica 184, 65-72. [本文引用: 1]
[24]
KempinSA, SavidgeB, YanofskyMF (1995). Molecular basis of the cauliflower phenotype in Arabidopsis.Science 267, 522-525. [本文引用: 1]
[25]
KomatsuK, MaekawaM, UjiieS, SatakeY, FurutaniI, OkamotoH, ShimamotoK, KyozukaJ (2003a). LAX and SPA: major regulators of shoot branching in rice.Proc Natl Acad Sci USA 100, 11765-11770. [本文引用: 4]
[26]
KomatsuM, ChujoA, NagatoY, ShimamotoK, KyozukaJ (2003b). FRIZZY PANICLE is required to prevent the formation of axillary meristems and to establish floral meristem identity in rice spikelets.Development 130, 3841-3850. [本文引用: 2]
[27]
KomatsuM, MaekawaM, ShimamotoK, KyozukaJ (2001). The LAX1 and FRIZZY PANICLE 2 genes determine the inflorescence architecture of rice by controlling rachis-branch and spikelet development.Dev Biol 231, 364-373. [本文引用: 1]
[28]
KurakawaT, UedaN, MaekawaM, KobayashiK, KojimaM, NagatoY, SakakibaraH, KyozukaJ (2007). Direct control of shoot meristem activity by a cytokinin-activating enzyme.Nature 445, 652-655. [本文引用: 1]
[29]
KyozukaJ (2007). Control of shoot and root meristem function by cytokinin.Curr Opin Plant Biol 10, 442-446. [本文引用: 1]
[30]
LiF, LiuWB, TangJY, ChenJF, TongHN, HuB, LiCL, FangJ, ChenMS, ChuCC (2010). Rice DENSE AND ERECT PANICLE 2 is essential for determining panicle outgrowth and elongation.Cell Res 20, 838-849. [本文引用: 2]
[31]
LiM, TangD, WangK, WuXR, LuLL, YuHX, GuMH, YanCJ, ChengZK (2011). Mutations in the F-box gene LARGER PANICLE improve the panicle architecture and enhance the grain yield in rice.Plant Biotechnol J 9, 1002-1013. [本文引用: 2]
[32]
LiSB, QianQ, FuZM, ZengDL, MengXB, KyozukaJ, MaekawaM, ZhuXD, ZhangJ, LiJY, WangYH (2009). Short panicle1 encodes a putative PTR family transporter and determines rice panicle size.Plant J 58, 592-605. [本文引用: 2]
[33]
LiSY, ZhaoBR, YuanDY, DuanMJ, QianQ, TangL, WangB, LiuXQ, ZhangJ, WangJ, SunJQ, LiuZ, FengYQ, YuanLP, LiCY (2013). Rice zinc finger protein DST enhances grain production through controlling Gn1a/OsCKX2 expression.Proc Natl Acad Sci USA 110, 3167-3172. [本文引用: 1]
[34]
LiXY, QianQ, FuZM, WangYH, XiongGS, ZengDL, WangXQ, LiuXF, TengS, HiroshiF, YuanM, LuoD, HanB, LiJY (2003). Control of tillering in rice.Nature 422, 618-621. [本文引用: 1]
[35]
LinH, WangRX, QianQ, YanMX, MengXB, FuZM, YanCY, JiangB, SuZ, LiJY, WangYH (2009). DWARF27, an iron-containing protein required for the biosynthesis of strigolactones, regulates rice tiller bud outgrowth.Plant Cell 21, 1512-1525. [本文引用: 2]
[36]
LindsayDL, SawhneyVK, Bonham-SmithPC (2006). Cytokinin-induced changes in CLAVATA1 and WUSCHEL expression temporally coincide with altered floral deve- lopment in Arabidopsis.Plant Sci 170, 1111-1117. [本文引用: 1]
[37]
LiuC, TeoZW, BiY, SongSY, XiWY, YangXB, YinZC, YuH (2013). A conserved genetic pathway determines inflorescence architecture in Arabidopsis and rice.Dev Cell 24, 612-622. [本文引用: 1]
[38]
MandelMA, Gustafson-BrownC, SavidgeB, YanofskyMF (1992). Molecular characterization of the Arabidopsis floral hopmeotic gene Apetala 1.Nature 360, 273-277. [本文引用: 1]
[39]
MayerKF, SchoofH, HaeckerA, LenhardM, JürgensG, LauxT, EntwicklungsgenetikL (1998). Role of WUS- CHEL in regulating stem cell fate in the Arabidopsis shoot meristem.Cell 95, 805-815. [本文引用: 1]
[40]
MoritaY, KyozukaJ (2007). Characterization of OsPID, the rice ortholog of PINOID, and its possible involvement in the control of polar auxin transport.Plant Cell Physiol 48, 540-549. [本文引用: 1]
[41]
NakagawaM, ShimamotoK, KyozukaJ (2002). Overex- pression of RCN1 and RCN2, rice TERMINAL FLOWER 1/CENTRORADIALIS homologs, confers delay of phase transition and altered panicle morphology in rice.Plant J 29, 743-750. [本文引用: 1]
[42]
OikawaT, KyozukaJ (2009). Two-step regulation of LAX PANICLE 1 protein accumulation in axillary meristem formation in rice.Plant Cell 21, 1095-1108. [本文引用: 2]
[43]
RaoNN, PrasadK, KumarPR, VijayraghavanU (2008). Distinct regulatory role for RFL, the rice LFY homolog, in determining flowering time and plant architecture.Proc Natl Acad Sci USA 105, 3646-3651. [本文引用: 2]
[44]
SazukaT, KamiyaN, NishimuraT, OhmaeK, SatoY, ImamuraK, NagatoY, KoshibaT, NagamuraY, AshikariM, KitanoH, MatsuokaM (2009). A rice tryptophan deficient dwarf mutant, tdd1, contains a reduced level of indole acetic acid and develops abnormal flowers and organless embryos.Plant J 60, 227-241. [本文引用: 1]
[45]
ShenCJ, BaiYH, WangSK, ZhangSN, WuYR, ChenM, JiangDA, QiYH (2010). Expression profile of PIN, AUX/LAX and PGP auxin transpoter gene families in Sorghum bicolor under phytohormone and abiotic stress. FEBS J 277, 2954-2969. [本文引用: 1]
[46]
SuzakiT, SatoM, AshikariM, MiyoshiM, NagatoY, Hi- ranoHY (2004). The gene FLORAL ORGAN NUMBER1 regulates floral meristem size in rice and encodes a leu- cine-rich repeat receptor kinase orthologous to Arab- idopsis CLAVATA1.Development 131, 5649-5657. [本文引用: 1]
[47]
TabuchiH, ZhangY, HattoriS, OmaeM, Shimizu-SatoS, OikawaT, QianQ, NishimuraM, KitanoH, XieH, FangXH, YoshidaH, KyozukaJ, ChenF, SatoY (2011). LAX PANICLE2 of rice encodes a novel nuclear protein and regulates the formation of axillary meristems.Plant Cell 23, 3276-3287. [本文引用: 1]
[48]
WangL, SunSY, JinJY, FuDB, YangXF, WengXS, XuC, LiXH, XiaoJH, ZhangQF (2015). Coordinated regulation of vegetative and reproductive branching in rice. Proc Natl Acad Sci USA 112, 15504-15509. [本文引用: 1]
[49]
WangLM, XieWB, ChenY, TangWJ, YangJY, YeRJ, LiuL, LinYJ, XuC, XiaoJH, ZhangQF (2010). A dynamic gene expression atlas covering the entire life cycle of rice.Plant J 61, 752-766. [本文引用: 1]
WuYZ, FuYC, ZhaoSS, GuP, ZhuZF, SunCQ, TanLB (2015). CLUSTERED PRIMARY BRANCH 1, a new allele of DWARF11, controls panicle architecture and seed size in rice.Plant Biotechnol J 14, 377-386. [本文引用: 1]
[52]
YanWH, WangP, ChenHX, ZhouHJ, LiQP, WangCR, DingZH, ZhangYS, YuSB, XingYZ, ZhangQF (2011). A major QTL, Ghd8, plays pleiotropic roles in regulating grain productivity, plant height, and heading date in rice.Mol Plant 4, 319-330. [本文引用: 1]
[53]
YoshidaA, OhmoriY, KitanoH, Taguchi-ShiobaraF, HiranoHY (2012). ABERRANT SPIKELET AND PANI- CLE1, encoding a TOPLESS-related transcriptional co- repressor, is involved in the regulation of meristem fate in rice.Plant J 70, 327-339. [本文引用: 1]
[54]
YoshidaA, SasaoM, YasunoN, TakagiK, DaimonY, ChenRH, YamazakiR, TokunagaH, KitaguchiY, SatoY, NagamuraY, UshijimaT, KumamaruT, IidaS, MaekawaM, KyozukaJ (2013). TAWAWA1, a regulator of rice inflorescence architecture, functions through the suppression of meristem phase transition.Proc Natl Acad Sci USA 110, 767-772. [本文引用: 1]
[55]
YuHY, MurchieEH, González-CarranzaZH, PykeKA, RobertsJA (2015). Decreased photosynthesis in the erect panicle 3 (ep3) mutant of rice is associated with reduced stomatal conductance and attenuated guard cell development.J Exp Bot 66, 1543-1552. [本文引用: 1]
[56]
ZhangSH, HuWJ, WangLP, LinCF, CongB, SunCR, LuoD (2005). TFL1/CEN-like genes control intercalary meristem activity and phase transition in rice.Plant Sci 168, 1393-1408. [本文引用: 1]
[57]
ZhangZY, LiJJ, YaoGX, ZhangHL, DouHJ, ShiHL, SunXM, LiZC (2011). Fine mapping and cloning of the Grain Number Per-Panicle Gene (Gnp4) on chromosome 4 in rice (Oryza sativa L.).J Integr Agr 10, 1825-1833. [本文引用: 1]
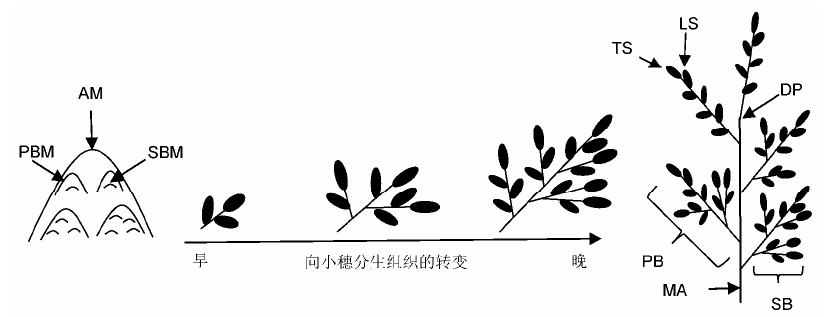
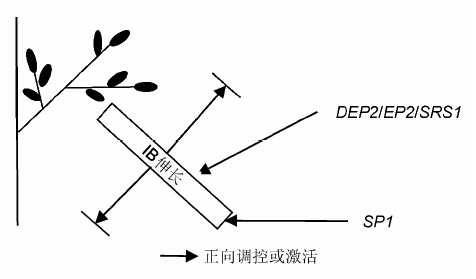
... 直立密穗基因DEP2 (Li et al., 2010)、EP2 (Erect Panicle 2) (Zhu et al., 2010)与小圆粒基因SRS1 (Abe et al., 2010)是同一基因位点.该基因突变后, 株型紧凑, 穗变直立, 但产量无显著变化.研究表明, 该基因突变后不会改变水稻幼穗原基分生组织的形成, 但能够影响穗轴和枝梗的伸长.在突变体中, 茎秆变得粗壮且有更多的维管束, 同时由于外稃纵向细胞长度和数量降低及横向细胞延长, 籽粒变得小而圆.该基因包含10个外显子, 编码1个由1 365 个氨基酸组成的植物特有的蛋白, 该蛋白定位于内质网, 但生物学功能未知. ... ... 水稻分蘖和穗分枝分别是营养生长期和生殖生长期形成的侧生器官.某些情况下, 二者受到共同的调控, 产生同等改变.如MOC1 (MONOCULM 1)和LAX1都参与分蘖和穗分枝, 其中任一基因突变都会导致穗蘖分枝数减少(Komatsu et al., 2003a; Li et al., 2003).但多数情况下, 分蘖数和穗分枝数的改变并不总是保持一致, 如OsCKX2、SP1和DEP1突变能特异地改变水稻穗分枝数, 分蘖数却未表现出相应变化(Ashikari et al., 2005; Huang et al., 2009; Li et al., 2009).甚至在某些情况下还会出现相反的情况, 如NIL-ipa1植株表现为分蘖减少而穗分枝数增加(Li et al., 2010).从一些与激素相关基因, 如Gnla (OsCKX2) (Ashikari et al., 2005)和D27 (Lin et al., 2009)的研究推测, 穗蘖分枝的时空调控可能由不同的基因来完成.以上研究表明, 水稻在营养生长时的分蘖和生殖生长阶段的穗发育可能受不同的机制调控. ...
2 2011
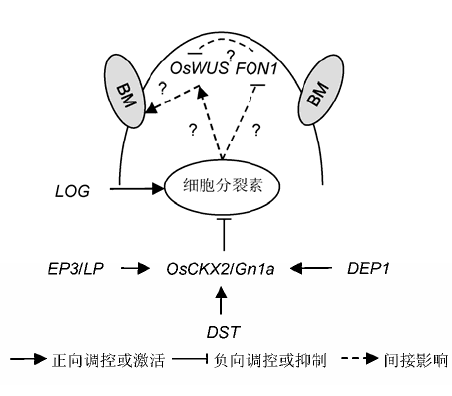
... 目前已报道多个基因通过调控OsCKX2基因的转录水平进而影响生殖分生组织中的细胞分裂素水平.水稻大穗基因LP (Large Panicle)及其等位基因EP3 (Erect Panicle 3), 编码1个定位于内质网上的F-Box蛋白, 可能作为E3泛素连接酶的1个亚基, 调控细胞分裂素氧化酶基因OsCKX2的转录水平, 进而调节植物顶端分生组织内的细胞分裂素水平和分生组织活性, 表明LP介导的蛋白降解可能控制着水稻中细胞分裂素的代谢(Li et al., 2011; Yu et al., 2015).同时, OsCKX2在生殖分生组织中的表达能够被水稻锌指转录因子DST (Drought and Salt Tolerance)直接上调.一个DST的半显性等位基因能够通过降低OsCKX2的表达提高细胞分裂素水平, 从而提高生殖分生组织活性, 产生更多的花序枝梗, 最终提高穗粒数(Li et al., 2013).此外, OsCKX2的表达还能被直立密穗基因DEP1 (Dense and Erect Panicle 1)的等位基因NIL-dep1下调.该位点是1个控制水稻产量性状的主效QTL, 编码产物与角蛋白5-4家族同源, 由3个血管性血友病因子C型结构域、1个跨膜结构域和1个4-二硫化物-核心结构域组成.其等位基因通过下调OsCKX2的表达水平提高水稻分生组织活性, 使得穗长和穗颈节变短, 且穗着粒密度增加, 最终能够促进水稻增产15%-20% (Huang et al., 2009; Zhou et al., 2009). ... ... 当前, 突变体鉴定仍是研究水稻基因功能最为直接有效的方法.多年以来, 水稻相关生物学家和育种专家已发现众多有价值的变异材料, 如叶形突变体(鞠培娜等, 2010)、株高突变体(杨德卫等, 2011)及穗型突变体(刘丹等, 2016)等.而在已获得的众多穗型突变体中, 只有少数几个是功能获得的有利变异, 如大穗突变体lp (Li et al., 2011).如何深入挖掘此类有利等位变异并应用到育种当中必然是将来的重大课题.当前获得变异资源的途径主要有2个, 一是自然变异, 二是人工诱变.针对自然变异, 要扩大相关种质资源的探索范围, 在一些地方品种中或许存在着更优质、更丰富的等位变异.而随着现代生物技术的不断发展, 利用新技术(如近年来兴起的CRISPR/Cas9)创制新的变异类型, 可大大丰富穗型的种质资源. ...
2 2009
... SP1 (Short Panicle 1)参与调控水稻穗长.突变体sp1在早期的幼穗分生组织生长发育无变化, 但在幼穗发育时, 枝梗不能正常伸长, 导致一级枝梗延迟发育甚至退化, 最终穗变短.SP1在幼穗韧皮部中高表达, 编码1个定位于质膜上的多肽转运蛋白(peptide transporter, PTR), 该蛋白含有12个跨膜结构域(Li et al., 2009). ... ... 水稻分蘖和穗分枝分别是营养生长期和生殖生长期形成的侧生器官.某些情况下, 二者受到共同的调控, 产生同等改变.如MOC1 (MONOCULM 1)和LAX1都参与分蘖和穗分枝, 其中任一基因突变都会导致穗蘖分枝数减少(Komatsu et al., 2003a; Li et al., 2003).但多数情况下, 分蘖数和穗分枝数的改变并不总是保持一致, 如OsCKX2、SP1和DEP1突变能特异地改变水稻穗分枝数, 分蘖数却未表现出相应变化(Ashikari et al., 2005; Huang et al., 2009; Li et al., 2009).甚至在某些情况下还会出现相反的情况, 如NIL-ipa1植株表现为分蘖减少而穗分枝数增加(Li et al., 2010).从一些与激素相关基因, 如Gnla (OsCKX2) (Ashikari et al., 2005)和D27 (Lin et al., 2009)的研究推测, 穗蘖分枝的时空调控可能由不同的基因来完成.以上研究表明, 水稻在营养生长时的分蘖和生殖生长阶段的穗发育可能受不同的机制调控. ...
1 2013
... 目前已报道多个基因通过调控OsCKX2基因的转录水平进而影响生殖分生组织中的细胞分裂素水平.水稻大穗基因LP (Large Panicle)及其等位基因EP3 (Erect Panicle 3), 编码1个定位于内质网上的F-Box蛋白, 可能作为E3泛素连接酶的1个亚基, 调控细胞分裂素氧化酶基因OsCKX2的转录水平, 进而调节植物顶端分生组织内的细胞分裂素水平和分生组织活性, 表明LP介导的蛋白降解可能控制着水稻中细胞分裂素的代谢(Li et al., 2011; Yu et al., 2015).同时, OsCKX2在生殖分生组织中的表达能够被水稻锌指转录因子DST (Drought and Salt Tolerance)直接上调.一个DST的半显性等位基因能够通过降低OsCKX2的表达提高细胞分裂素水平, 从而提高生殖分生组织活性, 产生更多的花序枝梗, 最终提高穗粒数(Li et al., 2013).此外, OsCKX2的表达还能被直立密穗基因DEP1 (Dense and Erect Panicle 1)的等位基因NIL-dep1下调.该位点是1个控制水稻产量性状的主效QTL, 编码产物与角蛋白5-4家族同源, 由3个血管性血友病因子C型结构域、1个跨膜结构域和1个4-二硫化物-核心结构域组成.其等位基因通过下调OsCKX2的表达水平提高水稻分生组织活性, 使得穗长和穗颈节变短, 且穗着粒密度增加, 最终能够促进水稻增产15%-20% (Huang et al., 2009; Zhou et al., 2009). ...
1 2003
... 水稻分蘖和穗分枝分别是营养生长期和生殖生长期形成的侧生器官.某些情况下, 二者受到共同的调控, 产生同等改变.如MOC1 (MONOCULM 1)和LAX1都参与分蘖和穗分枝, 其中任一基因突变都会导致穗蘖分枝数减少(Komatsu et al., 2003a; Li et al., 2003).但多数情况下, 分蘖数和穗分枝数的改变并不总是保持一致, 如OsCKX2、SP1和DEP1突变能特异地改变水稻穗分枝数, 分蘖数却未表现出相应变化(Ashikari et al., 2005; Huang et al., 2009; Li et al., 2009).甚至在某些情况下还会出现相反的情况, 如NIL-ipa1植株表现为分蘖减少而穗分枝数增加(Li et al., 2010).从一些与激素相关基因, 如Gnla (OsCKX2) (Ashikari et al., 2005)和D27 (Lin et al., 2009)的研究推测, 穗蘖分枝的时空调控可能由不同的基因来完成.以上研究表明, 水稻在营养生长时的分蘖和生殖生长阶段的穗发育可能受不同的机制调控. ...
2 2009
... 水稻分蘖和穗分枝分别是营养生长期和生殖生长期形成的侧生器官.某些情况下, 二者受到共同的调控, 产生同等改变.如MOC1 (MONOCULM 1)和LAX1都参与分蘖和穗分枝, 其中任一基因突变都会导致穗蘖分枝数减少(Komatsu et al., 2003a; Li et al., 2003).但多数情况下, 分蘖数和穗分枝数的改变并不总是保持一致, 如OsCKX2、SP1和DEP1突变能特异地改变水稻穗分枝数, 分蘖数却未表现出相应变化(Ashikari et al., 2005; Huang et al., 2009; Li et al., 2009).甚至在某些情况下还会出现相反的情况, 如NIL-ipa1植株表现为分蘖减少而穗分枝数增加(Li et al., 2010).从一些与激素相关基因, 如Gnla (OsCKX2) (Ashikari et al., 2005)和D27 (Lin et al., 2009)的研究推测, 穗蘖分枝的时空调控可能由不同的基因来完成.以上研究表明, 水稻在营养生长时的分蘖和生殖生长阶段的穗发育可能受不同的机制调控. ... ... 激素是调控植物生长发育的重要因素, 而植物株型调控机制多是通过内源激素调节分枝原基的产生与伸长.当前, 对植物株型塑造(即分枝)相关激素研究较多的有生长素、细胞分裂素和独脚金内酯(strigolactones).独脚金内酯作为新的植物激素近年来才被发现, 该激素参与抑制植物分枝的伸长生长, 同时与生长素和细胞分裂素一起调控植物的分枝数量.但目前已知的关于独脚金内酯的突变体或基因大多与分蘖相关, 如D3 (Ishikawa et al., 2005)、D14 (Arite et al., 2009)和D27 (Lin et al., 2009)等, 尚未发现其在穗分枝发育中的作用.作为控制分枝发育的重要激素, 独脚金内酯是否参与调控水稻穗型及其相关机理还有待研究.影响穗发育的D11等位基因CPB1参与油菜素内酯(brassinolide)的合成, 然而油菜素内酯对水稻花序发育的调控作用尚未见相关报道.以上研究表明, 植物激素参与的水稻花序发育调控仍有众多未知. ...
1 2006
... 在拟南芥中, 茎端分生组织的维持需要WUS (WUSCHEL)基因表达所激发的信号进行诱导.WUS在茎端分生组织顶层细胞之下表达, 随着细胞的不断分裂和生长, 原分生组织转变为成熟组织, 在其顶端又形成新的分生组织, 因此WUS的表达区域也随之更新.研究表明, CLV1 (CLAVATA 1)能抑制WUS的表达, 而外源细胞分裂素能在激活WUS的同时抑制CLV1 (Mayer et al., 1998; Lindsay et al., 2006).细胞分裂素、WUS和CLV1三者形成了1条反馈通路.越来越多的证据表明, WUS-CLV信号通路在双子叶植物和单子叶植物中是局部保守的.与拟南芥突变体clv相似, CLV1在水稻中的同源基因FON1 (Floral Organ Number 1)突变后也表现花分生组织变大, 花器官数目增多(Suzaki et al., 2004).然而, 水稻顶端分生组织中是否也存在由细胞分裂素、OsWUS和FON1三者组成的反馈路径则尚存疑问. ...
1 2013
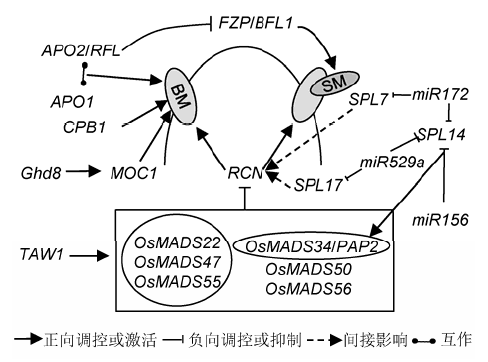
... 拟南芥TFL1 (Terminal Flower 1)/Centroradialis在水稻中的同源基因RCN1 (Rice TFL1/CEN homolog 1)是调控水稻枝梗形成的重要基因, 其表达受到6个MADS-box基因的抑制, 分别为拟南芥SOC1 (Suppressor of Overexpression of Constans 1)在水稻中对应的同源基因OsMADS50和OsMADS56, SVP (Short Vegetative Phase)或AGL24 (AGAMO- US-LIKE 24)对应的OsMADS22、OsMADS47和OsMADS55, 以及SEP4 (SEPALLATA)对应的OsMADS34.在OsMADS34突变背景下敲除其余5个MADS-box基因会导致水稻花序枝梗显著增加(Liu et al., 2013).类TFL1的基因对维持分生组织特性在双子叶和单子叶植物中是高度保守的(Nakagawa et al., 2002; Zhang et al., 2005; Carmona et al., 2007; Danilevskaya et al., 2010).过表达TFL1的同源基因RCN1和RCN2会延长营养生长到生殖生长的转变过程, 具体表现为枝梗生长点分生组织转化为小花结构延迟, 导致枝梗数量显著增加, 穗部变密(张淑红, 2005).MADS-box基因与这些花序身份特性相关基因的关系还有待进一步研究. ...
1 1992
... 水稻花序从枝梗分生组织形成转入小穗形成是水稻穗型塑造中的重要事件, 这取决于分生组织的身份特性.通过对模式植物拟南芥的研究已发现了多个与分生组织身份确定相关的基因, 如对花分生组织身份进行协同调控的基因LFY (LEAFY)、AP1 (APETALA1)和CAL (CAULIFLOWER), 这些基因使得侧生分生组织拥有花的身份特性进而向花分化(Mandel et al., 1992; Weigel et al., 1992; Kempin et al., 1995; Ferrándiz et al., 2000).近年来在水稻中也发现了大量与穗部分生组织身份特性相关的基因, 此类基因变异往往导致小穗分生组织形成提前或延迟发生(图4). ...
1 1998
... 在拟南芥中, 茎端分生组织的维持需要WUS (WUSCHEL)基因表达所激发的信号进行诱导.WUS在茎端分生组织顶层细胞之下表达, 随着细胞的不断分裂和生长, 原分生组织转变为成熟组织, 在其顶端又形成新的分生组织, 因此WUS的表达区域也随之更新.研究表明, CLV1 (CLAVATA 1)能抑制WUS的表达, 而外源细胞分裂素能在激活WUS的同时抑制CLV1 (Mayer et al., 1998; Lindsay et al., 2006).细胞分裂素、WUS和CLV1三者形成了1条反馈通路.越来越多的证据表明, WUS-CLV信号通路在双子叶植物和单子叶植物中是局部保守的.与拟南芥突变体clv相似, CLV1在水稻中的同源基因FON1 (Floral Organ Number 1)突变后也表现花分生组织变大, 花器官数目增多(Suzaki et al., 2004).然而, 水稻顶端分生组织中是否也存在由细胞分裂素、OsWUS和FON1三者组成的反馈路径则尚存疑问. ...
1 2007
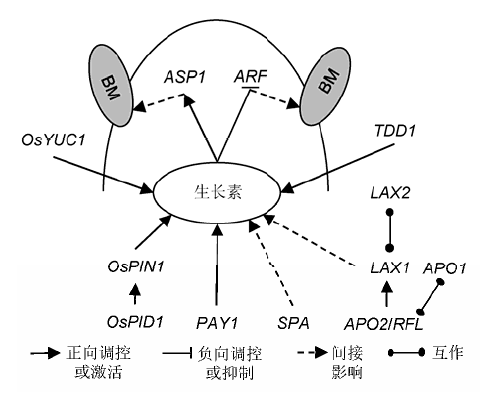
... 水稻中生长素转运机制总体与拟南芥中相似.在生长素极性运输蛋白(polar auxin transporter, PAT)和生长素合成的区域性分布的共同作用下, 生长素在植物体内呈极性分布.与生长素极性分布相适应, PAT的分布也具有极性, 其极性依赖于PIN蛋白家族, 而PIN的分布又受到PID调控(Delker et al., 2008).OsPID能够影响水稻枝梗形成, 其原因是该基因调控生长素的极性运输, 使得水稻花序分生组织腋下分生组织的分化受到影响(Morita and Kyozuka, 2007).最近在水稻中又发现了1个与生长素极性运输相关的基因PAY1 (Plant Architecture and Yield 1), 该基因编码1个类胰蛋白酶的丝氨酸和半胱氨酸蛋白酶, 通过影响生长素极性运输活性以及改变内生吲哚-3-乙酸的分布来改善水稻株型结构.pay1突变体的株高增加, 分蘖减少, 蘖夹角变小, 茎变细而穗变大.过表达PAY1的转基因植株穗粒数增加1倍(Zhao et al., 2015).PAY1的发现进一步丰富了水稻中的生长素转运机制. ...
1 2002
... 拟南芥TFL1 (Terminal Flower 1)/Centroradialis在水稻中的同源基因RCN1 (Rice TFL1/CEN homolog 1)是调控水稻枝梗形成的重要基因, 其表达受到6个MADS-box基因的抑制, 分别为拟南芥SOC1 (Suppressor of Overexpression of Constans 1)在水稻中对应的同源基因OsMADS50和OsMADS56, SVP (Short Vegetative Phase)或AGL24 (AGAMO- US-LIKE 24)对应的OsMADS22、OsMADS47和OsMADS55, 以及SEP4 (SEPALLATA)对应的OsMADS34.在OsMADS34突变背景下敲除其余5个MADS-box基因会导致水稻花序枝梗显著增加(Liu et al., 2013).类TFL1的基因对维持分生组织特性在双子叶和单子叶植物中是高度保守的(Nakagawa et al., 2002; Zhang et al., 2005; Carmona et al., 2007; Danilevskaya et al., 2010).过表达TFL1的同源基因RCN1和RCN2会延长营养生长到生殖生长的转变过程, 具体表现为枝梗生长点分生组织转化为小花结构延迟, 导致枝梗数量显著增加, 穗部变密(张淑红, 2005).MADS-box基因与这些花序身份特性相关基因的关系还有待进一步研究. ...
2 2009
... 与生长素转运机制不同, 参与水稻花序发育的生长素信号通路与双子叶植物相比更为复杂.LAX1 (Lax Panicle 1)编码1个植物特有的bHLH转录因子(Komatsu et al., 2003a).该转录因子为禾本科生殖期生长素信号通路所必需, 参与启动和维持水稻穗颈的腋生分生组织, 促进花序侧生分生组织形成(Shen et al., 2010).lax1突变体的穗部分枝和种子减少, 严重时则只有穗轴, 不产生枝梗, 并且不能结出种子.另外, 有些lax1等位突变体的终端小穗原基无限生长(Komatsu et al., 2001).同时, LAX1是花分生组织特化的基因, 能够启动终端小穗分生组织的形成, 其主要表达区域为地上部顶端分生组织与新形成的分生组织之间的边界, 是参与调控水稻腋芽原基形成的主要调节因子(Oikawa and Kyozuka, 2009). ... ... 水稻穗型是与产量直接相关的性状, 对穗型的研究结果具有重要的应用前景, 已经成为众多分子生物学家和育种学家重点研究的方向之一.过去数十年来, 对控制水稻花序表型的相关因子及调控路径的研究已取得巨大进展, 如能特异地作用于水稻花序发育的生长素信号和细胞分裂素信号.一些开花时间调控因子也能影响水稻穗发育, 如Ghd8 (Grain number, Plant height, and Heading date 8)能促进MOC1的表达(Yan et al., 2011), 而MOC1所编码的GRAS家族核蛋白控制着水稻分蘖和穗分枝, 决定着分蘖数和花序一级枝梗与二级枝梗数(Oikawa and Kyozuka,2009).生殖生长期, 水稻与双子叶植物拟南芥有着共同的调控机制, 包括CLV-WUS信号、生长素和细胞分裂素路径和MADS-box基因.尽管相关基因与路径在水稻中已被发现, 但其作用的具体模式仍有诸多未知之处, 未来应对此进行深入研究, 以期为水稻高产育种提供充分的理论依据. ...
2 2008
... 尽管LAX1在生长素介导的花序形态发生中起着重要作用, 但其作用方式仍不清楚, 仅知其在新的分生组织形成时以非细胞自主性的方式行使功能(Kom- atsu et al., 2003a).LAX1能与LAX2 (Lax Panicle 2)互作.LAX2也被称为Gnp4 (Grain Number Per- Panicle 4), 编码1个核蛋白, 该核蛋白含有植物特异的保守结构域, 与LAX1一起参与调控水稻腋生分生组织形成(Tabuchi et al., 2011; Zhang et al., 2011).在控制水稻腋生分生组织产生路径上, SPA (Small Panicle)是LAX1的功能冗余基因.spa突变体穗分枝异常, 其穗分枝数和每穗小穗数都有所减少, 位于穗基部的一级枝梗缺失显著, 且所有一级枝梗均变短, 二级枝梗减少, 能够形成侧生小穗, 但数量有所降低.当spa和lax1的弱等位突变体lax1-1构成双突变体时, 其穗部变为缺少所有分枝的线状结构.此外, lax或spa任一单突变体对分蘖影响很小, 但双突变体中分蘖几乎全部被抑制(Komatsu et al., 2003a).此外, 拟南 芥LEAFY的直系同源基因APO2 (Aberrant Panicle Organization 2)/RFL (Rice LFY Homolog)作用于花序发育早期, 其表达先于LAX1 (Rao et al., 2008). ... ... FZP的主要负调控因子APO2/RFL是拟南芥中LEAFY的直系同源基因.与LEAFY不同, APO2决定整个花序分生组织的身份而非特定的花分生组织.APO2能与拟南芥中UFO (Unusual Floral Organs)的直系同源基因APO1互作, 二者协同调控水稻花序发育.突变体apo1和apo2均表现出枝梗减少和穗粒数降低的表型, 造成此表型的原因是枝梗原基向小穗原基转变期提前.APO1和APO2还参与叶原基间隔期、花器官特性和花分生组织决定性的调控.APO1编码1个F-box蛋白, 正向调控花器官模型B类基因, APO2编码1个转录因子, 是开花激活因子OsSOC1 (OsMADS50)的调节因子(Ikeda et al., 2007; Rao et al., 2008; Ikedakawakatsu et al., 2012). ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}