Abstract: FLORICAULA/LEAFY encodes an evolutionarily conserved land plant-specific transcription factor. It functions by regulating the expression of downstream genes. RFL, the FLORICAULA/LEAFY homolog in rice, controls several important developments in rice: flowering time, plastochron and panicle branching. In this study, we detected the binding specificity of RFL by using the method systematic evolution of ligands by exponential enrichment (SELEX). After seven SELEX processes and sequencing, the binding site of RFL was identified as (C/A)(C/T)NN(T/C/A)G(G/T). The result contributes to clarifying the molecular function of RFL.
Key words:rice ; transcription factor ; SELEX ; DNA binding site
LFY (LEAFY)是拟南芥(Arabidopsis thaliana)花原基形成的必需基因, 该基因能够整合外部环境信号与内部开花信号来精确调控拟南芥的开花时间(Blaz- quez and Weigel, 2000)。Kyozuka等(1998)克隆了拟南芥LFY在水稻(Oryza sativa)中的同源基因RFL, 并利用原位杂交技术检测了RFL的表达情况。在一次枝梗和二次枝梗分化期, 该基因不在原基位置表达, 而是在原基周围区域表达。当小穗原基分化起始后, RFL基因就不在花序中的任何位置表达了, 这表明RFL与穗分化过程中分生特性的维持与转换密切相关(Kyozuka et al., 1998; Prasad et al., 2003)。
通过对RFL:RNAi植株和突变体的研究表明, 基因功能丧失后水稻穗部枝梗数减少甚至没有分枝, 说明RFL在幼穗分化中发挥非常重要的作用(Rao et al., 2008; Ikeda-Kawakatsu et al., 2012)。在RFL敲减实验中发现RFL能够通过调控CUC (CUP SHAPED COTELYDON)和LAX1 (LAX PANICLE 1)的表达量影响分蘖的发生(Deshpande et al., 2015)。在RFL基因控制下, 花序分生组织中的小穗分生组织的数量减少, 并表现出异常的花器官。在野生型的花序中, 穗轴分生组织通常形成10-12个一次枝梗后停止发育。然而, 在rfl突变体中, 枝梗分生组织在产生少量枝梗后就提前转化为小穗分生组织原基, 说明RFL基因的突变加
BlazquezMA, WeigelD (2000). Integration of floral inductive signals in Arabidopsis.Nature 404, 889-892. [本文引用: 1]
[3]
BuschMA, BombliesK, WeigelD (1999). Activation of a floral homeotic gene in Arabidopsis.Science 285, 585-587. [本文引用: 2]
[4]
DeshpandeGM, RamakrishnaK, ChongloiGL, VijayraghavanU (2015). Functions for rice RFL in vegetative axillary meristem specification and outgrowth.J Exp Bot 66, 2773-2784. [本文引用: 1]
[5]
HamesC, PtchelkineD, GrimmC, ThevenonE, MoyroudE, GerardF, MartielJL, BenllochR, ParcyF, MullerCW (2008). Structural basis for LEAFY floral switch function and similarity with helix-turn-helix proteins.EMBO J 27, 2628-2637. [本文引用: 1]
[6]
Ikeda-KawakatsuK, MaekawaM, IzawaT, ItohJ, NagatoY (2012). ABERRANT PANICLE ORGANIZATION 2/RFL, the rice ortholog of Arabidopsis LEAFY, suppresses the transition from inflorescence meristem to floral meristem through interaction with APO1.Plant J 69, 168-180. [本文引用: 4]
[7]
KlugSJ, FamulokM (1994). All you wanted to know about selex. Mol Biol Rep 20, 97-107. [本文引用: 2]
[8]
KyozukaJ, KonishiS, NemotoK, IzawaT, ShimamotoK (1998). Down-regulation of RFL, the FLO/LFY homolog of rice, accompanied with panicle branch initiation.Proc Natl Acad Sci USA 95, 1979-1982. [本文引用: 1]
[9]
PastoreJJ, LimpuangthipA, YamaguchiN, WuMF, SangY, HanSK, MalaspinaL, ChavdaroffN, YamaguchiA, WagnerD (2011). LATE MERISTEM IDENTITY2 acts together with LEAFY to activate APETALA1.Development 138, 3189-3198. [本文引用: 2]
[10]
PrasadK, KushalappaK, VijayraghavanU (2003). Mechanism underlying regulated expression of RFL, a conserved transcription factor, in the developing rice inflorescence.Mechan Dev 120, 491-502. [本文引用: 1]
[11]
RaoNN, PrasadK, KumarPR, VijayraghavanU (2008). Distinct regulatory role for RFL, the rice LFY homolog, in determining flowering time and plant architecture. Proc Natl Acad Sci USA 105, 3646-3651. [本文引用: 2]
[12]
SaddicLA, HuvermannB, BezhaniS, SuY, WinterCM, KwonCS, CollumRP, WagnerD (2006). The LEAFY target LMI1 is a meristem identity regulator and acts together with LEAFY to regulate expression of CAULIFLOWER.Development 133, 1673-1682. [本文引用: 1]
[13]
SayouC, MonniauxM, NanaoMH, MoyroudE, BrockingtonSF, ThevenonE, ChahtaneH, WarthmannN, MelkonianM, ZhangY, WongGK, WeigelD, ParcyF, DumasR (2014). A promiscuous intermediate underlies the evolution of LEAFY DNA binding specificity.Science 343, 645-648. [本文引用: 2]
[14]
SiriwardanaNS, LambRS (2012). A conserved domain in the N-terminus is important for LEAFY dimerization and function in Arabidopsis thaliana.Plant J 71, 736-749. [本文引用: 1]
[15]
WagnerD, SablowskiRW, MeyerowitzEM (1999). Trans- criptional activation of APETALA1 by LEAFY.Science 285, 582-584. [本文引用: 1]
[16]
WilliamDA, SuY, SmithMR, LuM, BaldwinDA, WagnerD (2004). Genomic identification of direct target genes of LEAFY.Proc Natl Acad Sci USA 101, 1775-1780. [本文引用: 1]
[17]
WinterCM, AustinRS, Blanvillain-BaufumeS, RebackMA, MonniauxM, WuMF, SangY, YamaguchiA, YamaguchiN, ParkerJE, ParcyF, JensenST, LiH, WagnerD (2011). LEAFY target genes reveal floral regulatory logic, cis motifs, and a link to biotic stimulus response.Dev Cell 20, 430-443. [本文引用: 3]
植物LEAFY同源基因的研究进展 1 2005
... RFL的突变使一次枝梗和二次枝梗数量急剧减少, 大大降低了水稻单产.研究RFL基因的功能有助于揭示水稻幼穗的分化过程.RFL作为一个转录因子, 其功能的行使主要依赖于对下游基因的调控.在拟南芥中已经鉴定得到了几个直接的下游基因, 如LMI1 (LATE MERISTEM IDENTITY1) (Saddic et al., 2006)、CAL (CAULIFLOWER) (William et al., 2004)、LMI2 (AtMYB17/LATE MERISTEM IDENTITY2) (Pastore et al., 2011)、APETALA1、AG (AGAMOUS) (Busch et al., 1999)和AP1 (APETALA1) (Wagner et al., 1999; Pastore et al., 2011).除此之外, 利用ChIP-seq技术还鉴定到多个潜在的直接下游基因(Hames et al., 2008; Winter et al., 2011), 但结果仍需通过遗传学及分子生物学技术进行验证.值得注意的是, 拟南芥LFY突变后植株不能正常形成花原基, 而水稻中幼穗各个发育进程似乎加快了, 促进了花原基的形成.研究表明尽管该基因在物种之间蛋白质序列相对保守, 但在进化过程中, 其功能发生了分化(马月萍等, 2005).目前水稻RFL的直接下游基因尚未被鉴定得到, 其转录调控功能需进一步研究.本研究鉴定了该基因的DNA结合位点, 为下一步确定直接下游调控基因奠定了重要基础. ...
1 2000
... LFY (LEAFY)是拟南芥(Arabidopsis thaliana)花原基形成的必需基因, 该基因能够整合外部环境信号与内部开花信号来精确调控拟南芥的开花时间(Blaz- quez and Weigel, 2000).Kyozuka等(1998)克隆了拟南芥LFY在水稻(Oryza sativa)中的同源基因RFL, 并利用原位杂交技术检测了RFL的表达情况.在一次枝梗和二次枝梗分化期, 该基因不在原基位置表达, 而是在原基周围区域表达.当小穗原基分化起始后, RFL基因就不在花序中的任何位置表达了, 这表明RFL与穗分化过程中分生特性的维持与转换密切相关(Kyozuka et al., 1998; Prasad et al., 2003). ...
2 1999
... LFY基因是调控陆生植物开花和细胞分裂的关键基因, 在植物中通常只有1个拷贝(Sayou et al., 2014).Sayou等(2014)系统研究了LFY作为转录因子在几种陆生植物中的DNA结合位点, 发现LFY通过改变结合位点的特异性使其在进化过程中保持单拷贝.目前关于拟南芥LFY的DNA结合位点研究得最为清楚(Bus- ch et al., 1999; Winter et al., 2011; Sayou et al., 2014).Busch等(1999)在拟南芥AP1和AG1基因的启动子上发现了LFY的DNA结合位点, 序列分别为CC- AGTGG和CCAATG(G/T).进一步研究确认了该特异性的结合位点可以表示为CC(A/G)N(T/C)GG (Win- ter et al., 2011).而关于水稻中RFL的DNA结合位点的研究尚未见报道.本研究利用原核表达系统表达RFL蛋白质, 利用重组蛋白进行蛋白质的功能研究.原核表达系统具有表达周期短、易于培养且成本低廉等特点, 已被广泛应用于外源蛋白质的重组表达.本研究使用原核表达蛋白RFL, 利用SELEX技术, 体外筛选鉴定到81条与之特异结合的DNA序列, 通过生物信息学分析鉴定到其特异结合位点.该结合位点与拟南芥中LFY略微不同, 首先在第3位上, RFL没有特异性要求, 而LFY更加偏向于A或者G; 在第6位上, RFL对碱基的要求更加特异, 只能是G.由于进行SELEX分析时使用的蛋白长度不同可能引起DNA结合结构域的构象发生变化, 从而影响序列结合的特异性.此外, 水稻RFL与拟南芥LFY蛋白中DNA结合结构域中氨基酸的不同可能也直接影响了特异性.Sayou等(2014)在研究中使用了2种不同长度的RFL蛋白进行SELEX分析, 发现水稻RFL结合位点, 其研究结果与 Winter等(2011)对于拟南芥LFY结合位点的研究类似.这似乎意味着表达蛋白的长度对于特异性分析更为重要.我们的研究表明, RFL蛋白对于结合序列中的第3位没有特异性要求, 这与Sayou等(2014)的研究结果不同.但由于所有的研究都使用不完全的RFL蛋白进行分析, 所以下一步的研究需要改进原核表达的方法, 使用全长蛋白进行SELEX分析, 进一步验证该蛋白DNA结合序列的特异性. ... ... RFL的突变使一次枝梗和二次枝梗数量急剧减少, 大大降低了水稻单产.研究RFL基因的功能有助于揭示水稻幼穗的分化过程.RFL作为一个转录因子, 其功能的行使主要依赖于对下游基因的调控.在拟南芥中已经鉴定得到了几个直接的下游基因, 如LMI1 (LATE MERISTEM IDENTITY1) (Saddic et al., 2006)、CAL (CAULIFLOWER) (William et al., 2004)、LMI2 (AtMYB17/LATE MERISTEM IDENTITY2) (Pastore et al., 2011)、APETALA1、AG (AGAMOUS) (Busch et al., 1999)和AP1 (APETALA1) (Wagner et al., 1999; Pastore et al., 2011).除此之外, 利用ChIP-seq技术还鉴定到多个潜在的直接下游基因(Hames et al., 2008; Winter et al., 2011), 但结果仍需通过遗传学及分子生物学技术进行验证.值得注意的是, 拟南芥LFY突变后植株不能正常形成花原基, 而水稻中幼穗各个发育进程似乎加快了, 促进了花原基的形成.研究表明尽管该基因在物种之间蛋白质序列相对保守, 但在进化过程中, 其功能发生了分化(马月萍等, 2005).目前水稻RFL的直接下游基因尚未被鉴定得到, 其转录调控功能需进一步研究.本研究鉴定了该基因的DNA结合位点, 为下一步确定直接下游调控基因奠定了重要基础. ...
1 2015
... 通过对RFL:RNAi植株和突变体的研究表明, 基因功能丧失后水稻穗部枝梗数减少甚至没有分枝, 说明RFL在幼穗分化中发挥非常重要的作用(Rao et al., 2008; Ikeda-Kawakatsu et al., 2012).在RFL敲减实验中发现RFL能够通过调控CUC (CUP SHAPED COTELYDON)和LAX1 (LAX PANICLE 1)的表达量影响分蘖的发生(Deshpande et al., 2015).在RFL基因控制下, 花序分生组织中的小穗分生组织的数量减少, 并表现出异常的花器官.在野生型的花序中, 穗轴分生组织通常形成10-12个一次枝梗后停止发育.然而, 在rfl突变体中, 枝梗分生组织在产生少量枝梗后就提前转化为小穗分生组织原基, 说明RFL基因的突变加 ...
1 2008
... RFL的突变使一次枝梗和二次枝梗数量急剧减少, 大大降低了水稻单产.研究RFL基因的功能有助于揭示水稻幼穗的分化过程.RFL作为一个转录因子, 其功能的行使主要依赖于对下游基因的调控.在拟南芥中已经鉴定得到了几个直接的下游基因, 如LMI1 (LATE MERISTEM IDENTITY1) (Saddic et al., 2006)、CAL (CAULIFLOWER) (William et al., 2004)、LMI2 (AtMYB17/LATE MERISTEM IDENTITY2) (Pastore et al., 2011)、APETALA1、AG (AGAMOUS) (Busch et al., 1999)和AP1 (APETALA1) (Wagner et al., 1999; Pastore et al., 2011).除此之外, 利用ChIP-seq技术还鉴定到多个潜在的直接下游基因(Hames et al., 2008; Winter et al., 2011), 但结果仍需通过遗传学及分子生物学技术进行验证.值得注意的是, 拟南芥LFY突变后植株不能正常形成花原基, 而水稻中幼穗各个发育进程似乎加快了, 促进了花原基的形成.研究表明尽管该基因在物种之间蛋白质序列相对保守, 但在进化过程中, 其功能发生了分化(马月萍等, 2005).目前水稻RFL的直接下游基因尚未被鉴定得到, 其转录调控功能需进一步研究.本研究鉴定了该基因的DNA结合位点, 为下一步确定直接下游调控基因奠定了重要基础. ...
4 2012
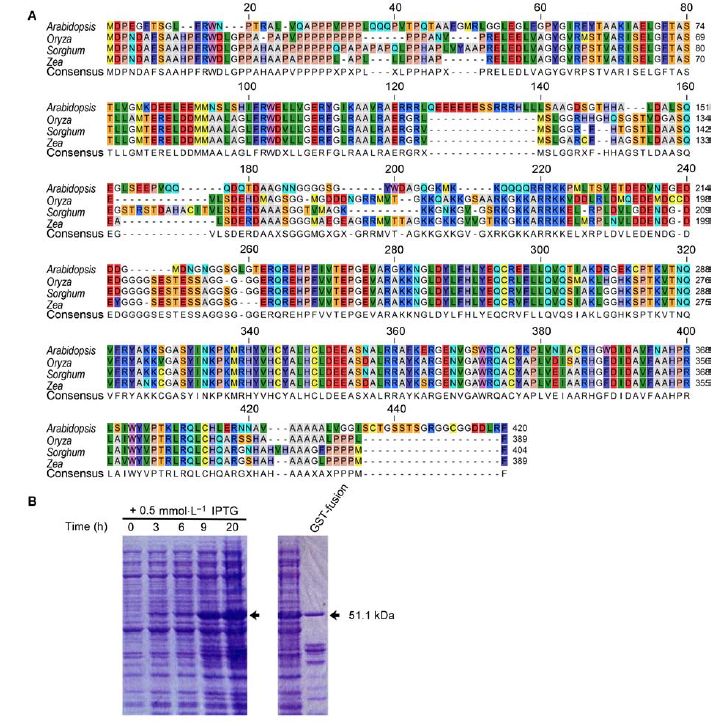
... 通过对RFL:RNAi植株和突变体的研究表明, 基因功能丧失后水稻穗部枝梗数减少甚至没有分枝, 说明RFL在幼穗分化中发挥非常重要的作用(Rao et al., 2008; Ikeda-Kawakatsu et al., 2012).在RFL敲减实验中发现RFL能够通过调控CUC (CUP SHAPED COTELYDON)和LAX1 (LAX PANICLE 1)的表达量影响分蘖的发生(Deshpande et al., 2015).在RFL基因控制下, 花序分生组织中的小穗分生组织的数量减少, 并表现出异常的花器官.在野生型的花序中, 穗轴分生组织通常形成10-12个一次枝梗后停止发育.然而, 在rfl突变体中, 枝梗分生组织在产生少量枝梗后就提前转化为小穗分生组织原基, 说明RFL基因的突变加 ... ... 快了花序分生组织分生特性的转化(Rao et al., 2008; Ikeda-Kawakatsu et al., 2012).此外, RFL基因的突变使水稻的出叶速率加快, 并延长了营养生长时间(Ikeda-Kawakatsu et al., 2012). ... ... 基因的突变使水稻的出叶速率加快, 并延长了营养生长时间(Ikeda-Kawakatsu et al., 2012). ... ... RFL蛋白质有2个主要结构域(图1A), 分别位于C端和N端.其中N端保守结构域是RFL蛋白形成同源复合体所必需的; C端为DNA结合结构域(Ikeda-Kawakatsu et al., 2012; Siriwardana and Lamb, 2012).首先, 我们克隆了RFL全长基因进行原核表达.但经过反复优化, 难以得到可溶表达的蛋白全长.其原因可能是由于编码序列较长, 蛋白难以在原核表达系统中进行完整翻译.根据RFL蛋白结构域, 只克隆DNA结合结构域进行原核表达, 可提高蛋白质的原核表达效率.本研究克隆了RFL蛋白的第189-389位氨基酸, 通过改变诱导温度、时间以及IPTG浓度, 探索各种条件对蛋白表达的影响.结果表明, IPTG浓度为0.5 mmol·L-1时诱导表达效果最好, 在诱导后20小时蛋白质就已经大量表达, 且均为可溶表达(图1B). ...
2 1994
... RFL作为特异性转录因子, 其功能的行使主要依赖于调控下游基因的表达.鉴定转录因子的DNA结合序列是研究转录因子基因调控功能的重要一步.指数富集配体的系统进化(systematic evolution of ligands by exponential enrichment, SELEX)是20世纪90年代发展起来的一项组合化学技术, 可以用于鉴定转录因子的DNA结合序列(Klug and Famulok, 1994).SELEX技术是一个从人工合成的大容量寡核苷酸库中, 结合PCR扩增、指数级富集与靶分子特异结合的寡核苷酸, 经过几轮或数十轮筛选, 获得高亲和力、高特异性的核苷酸配基, 即适体(Klug and Famulok, 1994).该技术已经成为一种重要的研究手段和工具, 可以从随机核酸序列库中筛选到与转录因子特异性结合的DNA序列, 即确认转录因子的DNA结合位点.如Sayou等(2014)利用该技术鉴定了多种陆生植物中LFY同源基因的DNA结合序列, 发现在陆生植物中, LFY同源基因的结合序列可以分为3类, 其中被子植物、裸子植物、蕨类植物和苔藓植物都划归为第1类.本研究利用SELEX技术鉴定了水稻RFL的DNA结合 ... ... ).SELEX技术是一个从人工合成的大容量寡核苷酸库中, 结合PCR扩增、指数级富集与靶分子特异结合的寡核苷酸, 经过几轮或数十轮筛选, 获得高亲和力、高特异性的核苷酸配基, 即适体(Klug and Famulok, 1994).该技术已经成为一种重要的研究手段和工具, 可以从随机核酸序列库中筛选到与转录因子特异性结合的DNA序列, 即确认转录因子的DNA结合位点.如Sayou等(2014)利用该技术鉴定了多种陆生植物中LFY同源基因的DNA结合序列, 发现在陆生植物中, LFY同源基因的结合序列可以分为3类, 其中被子植物、裸子植物、蕨类植物和苔藓植物都划归为第1类.本研究利用SELEX技术鉴定了水稻RFL的DNA结合 ...
1 1998
... LFY (LEAFY)是拟南芥(Arabidopsis thaliana)花原基形成的必需基因, 该基因能够整合外部环境信号与内部开花信号来精确调控拟南芥的开花时间(Blaz- quez and Weigel, 2000).Kyozuka等(1998)克隆了拟南芥LFY在水稻(Oryza sativa)中的同源基因RFL, 并利用原位杂交技术检测了RFL的表达情况.在一次枝梗和二次枝梗分化期, 该基因不在原基位置表达, 而是在原基周围区域表达.当小穗原基分化起始后, RFL基因就不在花序中的任何位置表达了, 这表明RFL与穗分化过程中分生特性的维持与转换密切相关(Kyozuka et al., 1998; Prasad et al., 2003). ...
2 2011
... RFL的突变使一次枝梗和二次枝梗数量急剧减少, 大大降低了水稻单产.研究RFL基因的功能有助于揭示水稻幼穗的分化过程.RFL作为一个转录因子, 其功能的行使主要依赖于对下游基因的调控.在拟南芥中已经鉴定得到了几个直接的下游基因, 如LMI1 (LATE MERISTEM IDENTITY1) (Saddic et al., 2006)、CAL (CAULIFLOWER) (William et al., 2004)、LMI2 (AtMYB17/LATE MERISTEM IDENTITY2) (Pastore et al., 2011)、APETALA1、AG (AGAMOUS) (Busch et al., 1999)和AP1 (APETALA1) (Wagner et al., 1999; Pastore et al., 2011).除此之外, 利用ChIP-seq技术还鉴定到多个潜在的直接下游基因(Hames et al., 2008; Winter et al., 2011), 但结果仍需通过遗传学及分子生物学技术进行验证.值得注意的是, 拟南芥LFY突变后植株不能正常形成花原基, 而水稻中幼穗各个发育进程似乎加快了, 促进了花原基的形成.研究表明尽管该基因在物种之间蛋白质序列相对保守, 但在进化过程中, 其功能发生了分化(马月萍等, 2005).目前水稻RFL的直接下游基因尚未被鉴定得到, 其转录调控功能需进一步研究.本研究鉴定了该基因的DNA结合位点, 为下一步确定直接下游调控基因奠定了重要基础. ... ... ; Pastore et al., 2011).除此之外, 利用ChIP-seq技术还鉴定到多个潜在的直接下游基因(Hames et al., 2008; Winter et al., 2011), 但结果仍需通过遗传学及分子生物学技术进行验证.值得注意的是, 拟南芥LFY突变后植株不能正常形成花原基, 而水稻中幼穗各个发育进程似乎加快了, 促进了花原基的形成.研究表明尽管该基因在物种之间蛋白质序列相对保守, 但在进化过程中, 其功能发生了分化(马月萍等, 2005).目前水稻RFL的直接下游基因尚未被鉴定得到, 其转录调控功能需进一步研究.本研究鉴定了该基因的DNA结合位点, 为下一步确定直接下游调控基因奠定了重要基础. ...
1 2003
... LFY (LEAFY)是拟南芥(Arabidopsis thaliana)花原基形成的必需基因, 该基因能够整合外部环境信号与内部开花信号来精确调控拟南芥的开花时间(Blaz- quez and Weigel, 2000).Kyozuka等(1998)克隆了拟南芥LFY在水稻(Oryza sativa)中的同源基因RFL, 并利用原位杂交技术检测了RFL的表达情况.在一次枝梗和二次枝梗分化期, 该基因不在原基位置表达, 而是在原基周围区域表达.当小穗原基分化起始后, RFL基因就不在花序中的任何位置表达了, 这表明RFL与穗分化过程中分生特性的维持与转换密切相关(Kyozuka et al., 1998; Prasad et al., 2003). ...
2 2008
... 通过对RFL:RNAi植株和突变体的研究表明, 基因功能丧失后水稻穗部枝梗数减少甚至没有分枝, 说明RFL在幼穗分化中发挥非常重要的作用(Rao et al., 2008; Ikeda-Kawakatsu et al., 2012).在RFL敲减实验中发现RFL能够通过调控CUC (CUP SHAPED COTELYDON)和LAX1 (LAX PANICLE 1)的表达量影响分蘖的发生(Deshpande et al., 2015).在RFL基因控制下, 花序分生组织中的小穗分生组织的数量减少, 并表现出异常的花器官.在野生型的花序中, 穗轴分生组织通常形成10-12个一次枝梗后停止发育.然而, 在rfl突变体中, 枝梗分生组织在产生少量枝梗后就提前转化为小穗分生组织原基, 说明RFL基因的突变加 ... ... 快了花序分生组织分生特性的转化(Rao et al., 2008; Ikeda-Kawakatsu et al., 2012).此外, RFL基因的突变使水稻的出叶速率加快, 并延长了营养生长时间(Ikeda-Kawakatsu et al., 2012). ...
1 2006
... RFL的突变使一次枝梗和二次枝梗数量急剧减少, 大大降低了水稻单产.研究RFL基因的功能有助于揭示水稻幼穗的分化过程.RFL作为一个转录因子, 其功能的行使主要依赖于对下游基因的调控.在拟南芥中已经鉴定得到了几个直接的下游基因, 如LMI1 (LATE MERISTEM IDENTITY1) (Saddic et al., 2006)、CAL (CAULIFLOWER) (William et al., 2004)、LMI2 (AtMYB17/LATE MERISTEM IDENTITY2) (Pastore et al., 2011)、APETALA1、AG (AGAMOUS) (Busch et al., 1999)和AP1 (APETALA1) (Wagner et al., 1999; Pastore et al., 2011).除此之外, 利用ChIP-seq技术还鉴定到多个潜在的直接下游基因(Hames et al., 2008; Winter et al., 2011), 但结果仍需通过遗传学及分子生物学技术进行验证.值得注意的是, 拟南芥LFY突变后植株不能正常形成花原基, 而水稻中幼穗各个发育进程似乎加快了, 促进了花原基的形成.研究表明尽管该基因在物种之间蛋白质序列相对保守, 但在进化过程中, 其功能发生了分化(马月萍等, 2005).目前水稻RFL的直接下游基因尚未被鉴定得到, 其转录调控功能需进一步研究.本研究鉴定了该基因的DNA结合位点, 为下一步确定直接下游调控基因奠定了重要基础. ...
2 2014
... LFY基因是调控陆生植物开花和细胞分裂的关键基因, 在植物中通常只有1个拷贝(Sayou et al., 2014).Sayou等(2014)系统研究了LFY作为转录因子在几种陆生植物中的DNA结合位点, 发现LFY通过改变结合位点的特异性使其在进化过程中保持单拷贝.目前关于拟南芥LFY的DNA结合位点研究得最为清楚(Bus- ch et al., 1999; Winter et al., 2011; Sayou et al., 2014).Busch等(1999)在拟南芥AP1和AG1基因的启动子上发现了LFY的DNA结合位点, 序列分别为CC- AGTGG和CCAATG(G/T).进一步研究确认了该特异性的结合位点可以表示为CC(A/G)N(T/C)GG (Win- ter et al., 2011).而关于水稻中RFL的DNA结合位点的研究尚未见报道.本研究利用原核表达系统表达RFL蛋白质, 利用重组蛋白进行蛋白质的功能研究.原核表达系统具有表达周期短、易于培养且成本低廉等特点, 已被广泛应用于外源蛋白质的重组表达.本研究使用原核表达蛋白RFL, 利用SELEX技术, 体外筛选鉴定到81条与之特异结合的DNA序列, 通过生物信息学分析鉴定到其特异结合位点.该结合位点与拟南芥中LFY略微不同, 首先在第3位上, RFL没有特异性要求, 而LFY更加偏向于A或者G; 在第6位上, RFL对碱基的要求更加特异, 只能是G.由于进行SELEX分析时使用的蛋白长度不同可能引起DNA结合结构域的构象发生变化, 从而影响序列结合的特异性.此外, 水稻RFL与拟南芥LFY蛋白中DNA结合结构域中氨基酸的不同可能也直接影响了特异性.Sayou等(2014)在研究中使用了2种不同长度的RFL蛋白进行SELEX分析, 发现水稻RFL结合位点, 其研究结果与 Winter等(2011)对于拟南芥LFY结合位点的研究类似.这似乎意味着表达蛋白的长度对于特异性分析更为重要.我们的研究表明, RFL蛋白对于结合序列中的第3位没有特异性要求, 这与Sayou等(2014)的研究结果不同.但由于所有的研究都使用不完全的RFL蛋白进行分析, 所以下一步的研究需要改进原核表达的方法, 使用全长蛋白进行SELEX分析, 进一步验证该蛋白DNA结合序列的特异性. ... ... ; Sayou et al., 2014).Busch等(1999)在拟南芥AP1和AG1基因的启动子上发现了LFY的DNA结合位点, 序列分别为CC- AGTGG和CCAATG(G/T).进一步研究确认了该特异性的结合位点可以表示为CC(A/G)N(T/C)GG (Win- ter et al., 2011).而关于水稻中RFL的DNA结合位点的研究尚未见报道.本研究利用原核表达系统表达RFL蛋白质, 利用重组蛋白进行蛋白质的功能研究.原核表达系统具有表达周期短、易于培养且成本低廉等特点, 已被广泛应用于外源蛋白质的重组表达.本研究使用原核表达蛋白RFL, 利用SELEX技术, 体外筛选鉴定到81条与之特异结合的DNA序列, 通过生物信息学分析鉴定到其特异结合位点.该结合位点与拟南芥中LFY略微不同, 首先在第3位上, RFL没有特异性要求, 而LFY更加偏向于A或者G; 在第6位上, RFL对碱基的要求更加特异, 只能是G.由于进行SELEX分析时使用的蛋白长度不同可能引起DNA结合结构域的构象发生变化, 从而影响序列结合的特异性.此外, 水稻RFL与拟南芥LFY蛋白中DNA结合结构域中氨基酸的不同可能也直接影响了特异性.Sayou等(2014)在研究中使用了2种不同长度的RFL蛋白进行SELEX分析, 发现水稻RFL结合位点, 其研究结果与 Winter等(2011)对于拟南芥LFY结合位点的研究类似.这似乎意味着表达蛋白的长度对于特异性分析更为重要.我们的研究表明, RFL蛋白对于结合序列中的第3位没有特异性要求, 这与Sayou等(2014)的研究结果不同.但由于所有的研究都使用不完全的RFL蛋白进行分析, 所以下一步的研究需要改进原核表达的方法, 使用全长蛋白进行SELEX分析, 进一步验证该蛋白DNA结合序列的特异性. ...




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}