靳川
1 , 2 , 李鑫豪
1 , 2 , 蒋燕
1 , 2 , 徐铭泽
1 , 2 , 田赟
1 , 2 , 刘鹏
1 , 2 , 贾昕
1 , 2 , 3 , 查天山
, 1 , 2 , 3 , * 1 北京林业大学水土保持学院, 北京 1000832 宁夏盐池毛乌素沙地生态系统国家定位观测研究站, 北京 1000833 北京林业大学水土保持国家林业局重点实验室, 北京 100083Relative changes and regulation of photosynthetic energy partitioning components in Artemisia ordosica during growing season Chuan JIN
1 , 2 , Xin-Hao LI
1 , 2 , Yan JIANG
1 , 2 , Ming-Ze XU
1 , 2 , Yun TIAN
1 , 2 , Peng LIU
1 , 2 , Xin JIA
1 , 2 , 3 , Tian- Shan ZHA
, 1 , 2 , 3 , * 1 School of Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China2 Yanchi Ecology Research Station of Mau US Desert, Beijing 100083, China3 Key Laboratory of State Forestry Administration on Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China通讯作者: *
tianshanzha@bjfu.edu.cn 编委: 彭长连
责任编辑: 李敏
收稿日期: 2021-04-19
修回日期: 2021-06-26
基金资助: 国家自然科学基金 (32071842 )国家自然科学基金 (31901366 )国家自然科学基金 (32071843 )中央高校基本科研业务费专项资金 (2015ZCQ-SB-02 )
Corresponding authors: *
tianshanzha@bjfu.edu.cn Received: 2021-04-19
Revised: 2021-06-26
Fund supported: National Natural Science Foundation of China (32071842 )National Natural Science Foundation of China (31901366 )National Natural Science Foundation of China (32071843 )Fundamental Research Funds for the Central Universities (2015ZCQ-SB-02 )
摘要 为了探明典型荒漠灌木优势物种黑沙蒿(俗名油蒿,
Artemisia ordosica )光合过程能量中分配对环境波动的相对变化及其长期调节机制, 该研究于2018年4-10月在宁夏盐池毛乌素沙地, 同时使用MONITORING-PAM多通道荧光监测仪和LI-6400XT便携式光合测量仪对黑沙蒿叶片的最小荧光产量(
F o )、最大荧光产量(
F m )、稳态荧光产量(
F s )、光下最大荧光产量(
F m ′)、净光合速率(
P n )、暗呼吸速率(
R d )、蒸腾速率(
E )和叶片气孔导度(
g s )进行现场测定, 在实验室内计算比叶面积(
SLA )、单位面积氮含量(
N area )、叶绿素含量(
C Chl )和叶绿素a/b (Chl a/b), 分析黑沙蒿光合过程能量分配中固碳耗能占比(
Φ A )、光呼吸耗能占比(
Φ PR )、调节性热耗散耗能占比(
Φ NPQ )和非调节性热耗散耗能占比(
Φ NO )与环境参数和叶性状参数之间的关系以及能量分配各组分之间的相对变化。结果表明, 光化学反应组分(
Φ A 、
Φ PR )和热耗散组分(
Φ NPQ 、
Φ NO )之间呈负相关竞争关系, 两组分内部呈正相关协同关系,
E 和
Φ A 、
Φ PR 正相关, 和
Φ NPQ 、
Φ NO 负相关。在低土壤含水量(
SWC )和高饱和水汽压差(
VPD )环境条件下, 黑沙蒿
Φ A 、
Φ PR 和
SLA 显著降低,
Φ NPQ 和
Φ NO 显著增加。研究认为, 在长期干旱或高蒸散条件下, 黑沙蒿通过降低
SLA 等途径避免水分的过度流失, 同时将部分过剩光能由光呼吸代谢途径转移到热耗散组分进行耗散。波动环境下黑沙蒿形态性状的变异和光合过程能量分配的长期调节机制, 反映了其利用形态与生理的协同可塑性对逆境的适应。
关键词: 荒漠植物 ;
光合作用 ;
能量分配 ;
光呼吸 ;
热耗散 Abstract Aims This study is to examine the relative changes and regulation in photosynthetic energy partitioning components of a typical desert shrub
Artemisia ordosica in Northwest China.
Methods The minimum fluorescence yield (
F o ), maximal fluorescence yield (
F m ), actual ?uorescence yield (
F s ), maximal fluorescence yield in the light-adapted state (
F m ′), net photosynthetic rate (
P n ), dark respiration (
R d ), transpiration rate (
E ) and stomatal conductance (
g s ) of
A. ordosica were investigated from April to October, 2018 using a multi-channel fluorometer and a portable photosynthesis analyzer. The specific leaf area (
SLA ), nitrogen per leaf area (
N area ), chlorophyll content (
C Chl ) and the ratio of chlorophyll a to chlorophyll b content (Chl a/b) were also measured. We analyzed the fraction of energy used for CO
2 assimilation (
Φ A ), the fraction of energy used for photorespiration (
Φ PR ), the fraction of regulate heat dissipation (
Φ NPQ ) and the fraction of non-regulate heat dissipation in energy allocation (
Φ NO ) during the photosynthetic process in
A. ordosica and their responses to fluctuating environment.
Important findings The photochemical reaction component (
Φ A and
Φ PR ) and the heat dissipation component (
Φ NPQ and
Φ NO ) showed a negative competition relationship, and exhibited positive synergistic relationship within components.
E was positively correlated with
Φ A and
Φ PR , and negatively correlated with
Φ NPQ and
Φ NO . Under low soil water content (
SWC ) and high saturated water vapor pressure (
VPD ) conditions,
Φ A ,
Φ PR and
SLA were significantly decreased, while
Φ NPQ and
Φ NO were significantly increased. The results indicate that
A. ordosica avoids excessive loss of water under the condition of the long drought or high evapotranspiration via reducing
SLA. Synergistically, excessive light energy is transferred from photorespiration to thermal dissipation pathway. The relative changes and long-term regulation of photosynthetic energy partitioning in
A. ordosica under fluctuating environment, fully reflect the acclimation of plants to adversity via its synergistic morphologic and physiological plasticity.
Keywords: desert plant ;
photosynthesis ;
energy partitioning ;
photorespiration ;
heat dissipation PDF (1018KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 靳川, 李鑫豪, 蒋燕, 徐铭泽, 田赟, 刘鹏, 贾昕, 查天山. 黑沙蒿光合能量分配组分在生长季的相对变化与调控机制. 植物生态学报, 2021, 45(8): 870-879. DOI:
10.17521/cjpe.2021.0146 JIN Chuan, LI Xin-Hao, JIANG Yan, XU Ming-Ze, TIAN Yun, LIU Peng, JIA Xin, ZHA Tian- Shan.
Relative changes and regulation of photosynthetic energy partitioning components in Artemisia ordosica during growing season .
Chinese Journal of Plant Ecology , 2021, 45(8): 870-879. DOI:
10.17521/cjpe.2021.0146 干旱半干旱区是维持生态稳定的关键屏障, 也是我国植被恢复的重点区域(
陈亚宁等, 2019 )。近年来气候恶化以及极端天气事件的频发, 该区植被经常遭受多种环境胁迫, 处于急剧退化的风险当中(
Huang et al ., 2017 )。植物通过光合作用将光能转化为化学能, 驱动生态系统碳循环(
赵风华和于贵瑞, 2018 )。光合过程各个步骤对外界环境的波动非常敏感, 根据植物光合特性能够快速了解其生理状态(
孙安安等, 2019 )。因此, 探究荒漠植物光合作用对环境因子的响应, 可以进一步明确其在逆境条件下的耐受力、恢复力和适应力(
Chaves et al ., 2004 )。
光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 )。调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 )。基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 )。植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 )。然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 )。随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 )。目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 )。受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究。通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 )。
荒漠生态系统具有干旱、高太阳辐射、高蒸腾和极端温度等环境特征(
Jia et al ., 2016 )。夏季荒漠的强烈太阳辐射远超植物光合所需, 已有研究证明荒漠植物通过调整热耗散等途径进行调节, 但实际上仍然会产生大量活性氧, 对植物光合机构造成极大威胁, 此时光呼吸过程的存在价值就显得尤为重要(
何炎红等, 2015 )。其次, 荒漠气候极端温度也会抑制卡尔文循环相关蛋白酶, 从而导致光合效率降低(
孙勇等, 2015 )。另外, 荒漠干燥空气导致的高蒸腾会促使植物气孔关闭, 影响了水的传输, 限制了光合过程中相关电子传输速率(
阮成江和李代琼, 2001 )。因此, 波动环境下荒漠植物光合过程能量分配的相对变化和内在调节机制可能具有明显特征。
黑沙蒿(俗名油蒿,
Artemisia ordosica )属半灌木菊科蒿属, 广泛分布于我国北部及西北沙地, 具有生长快、耐干旱和抗风沙侵蚀等优点(
杨洪晓等, 2004 )。作为毛乌素沙地最为主要的固沙物种, 黑沙蒿在维护荒漠生态稳定方面发挥着重要作用。本研究运用叶绿素荧光和气体交换技术, 对宁夏盐池毛乌素沙地荒漠灌木优势种黑沙蒿进行长期原位监测, 从光合过程能量分配角度出发, 旨在揭示: (1)波动环境下黑沙蒿光合过程能量分配各组分对环境因子的响应; (2)波动环境下黑沙蒿光合过程能量分配各组分之间相对变化及其内在调节机制。以期为构建荒漠生态系统能量交换模型提供数据支撑, 丰富荒漠植物的生理可塑性理论。
1 材料和方法 1.1 样地概况 研究区选取位于毛乌素沙地南部的宁夏盐池荒漠生态系统国家定位观测研究站, 37.07° N, 106.50° E, 海拔1 550 m。该区属温带大陆性季风气候, 年平均气温为7.7 ℃, 昼夜温差可达25 ℃以上。年降水量为292 mm, 年潜在蒸散量为2 024 mm。研究区植被以旱生灌木为主, 主要包括黑沙蒿、北沙柳(
Salix psammophila )、蒙古山竹子(
Corethrodendron fruticosum var.
mongolicum )和细枝山竹子(
Corethrodendron scoparium )。本研究选取黑沙蒿为研究对象, 黑沙蒿通常4月开始萌发, 7-8月积累生物量达到最大, 10月开始枯萎凋落。
于2018年4月在研究区选取地势平坦的天然黑沙蒿群落, 布设一块10 m × 10 m的样地。样地内黑沙蒿共27株, 平均高度为67.3 cm, 平均冠幅为0.83 m
2 , 植被盖度为45%。在样地内选择5株长势良好、有代表性(冠幅直径约为135 cm, 高度约为65 cm)的黑沙蒿作为样本植株, 在每个月的上、中和下旬分别进行叶绿素荧光和气体交换的原位监测, 以及叶性状测定取样。
1.2 叶绿素荧光和光合气体交换的测量 在晴朗无风的9:00-11:00, 使用MONITORING- PAM多通道荧光监测仪(Walz, Effeltrich, Germany)对5株黑沙蒿样株进行叶绿素荧光的测量。在向阳朝南方向中部枝条前端选取健康、无病虫害的2-3簇叶片, 使用特制遮光套袋同时盖住5个荧光探头和叶片, 进行20 min的暗适应后选取手动模式测定最小荧光产量(
F o )和最大荧光产量(
F m )。随后设置光强为1 500 μmol·m
-2 ·s
-1 , 进行10 min的光适应后得到稳态荧光产量(
F s )和光下最大荧光产量(
F m ′), 每个样株重复测量3次, 测定完成的叶片进行标记。
叶绿素荧光测量完成后, 使用配有LI-6400-02红蓝光源叶室的LI-6400便携式光合测量仪(LI-COR, Lincoln, USA)进行黑沙蒿光合气体交换测定。夹取测定叶绿素荧光后带有标记的样株叶片, 将叶室温度设置为25 ℃, 湿度设置为55%, CO
2 浓度设置为400 μmol·mol
-1 。首先将光强设置为0, 进行20 min的暗适应后, 待仪器显示实时数值小数点后一位稳定不变时, 记录相应数据。随后将光强设置为1 500 μmol·m
-2 ·s
-1 , 进行10 min后的光适应, 同样待数值稳定后记录数据。测量指标有净光合速率(
P n )、蒸腾速率(
E )和叶片气孔导度(
g s )等。每个样株重复测量3次, 采用轮回方法测定。
1.3 叶性状的测量与计算 由于黑沙蒿叶片小且形状不规则, 将测定光合气体交换的叶片收集, 平铺于带有黑色方格的背景纸上进行拍照, 利用图片像素提取方法识别出叶片和方格的区域, 根据比例关系计算出实际叶片面积。使用烘箱在70 ℃下烘干48 h, 称量叶片干质量计算比叶面积(
SLA )。使用研磨机分别将烘干的每份叶片制成粉末装袋, 委托中国科学研究院植物研究所测试中心采用凯氏定氮法测定叶片的氮含量, 后期计算单位面积氮含量(
N area )。每周期测量结束时, 选择生长良好的叶片取样, 用95%乙醇进行研磨浸提, 充分静置后使用紫外分光光度计(UV-2100, Unico, Calabasas, USA)在665和649 nm分别测定其吸光值, 计算叶绿素含量(
C Chl )和叶绿素a/b (Chl a/b)。
1.4 环境因子的观测与计算 光合有效辐射(
PAR )由距离样地150 m处通量塔(高6.2 m)的光合有效辐射传感器(PAR-LITE, Kipp and Zonen, Delft, the Netherlands)测得, 相对湿度(
RH )和空气温度(
T a )由通量塔上的温湿度传感器(HMP155A, Vaisala, Finland)测得,
PAR 和
T a 的缺失数据由荧光仪连续同步监测数据进行补充。采用以下公式求得饱和水汽压差(
VPD ):
(1) VPD = 0.611exp[17.27T a /(T a + 237.3)] × (1 - RH /100)土壤含水量(
SWC )由布设在样地内的土壤温湿度传感器(ECH2O-5TE, Decagon Devices, Pullman, USA)测定(深度为30 cm)。降水量(
P )由距离样地200 m空旷处的翻斗式雨量筒(TE525W, Campbell Scientific, Logan, USA)测定, 上述环境因子的观测间隔均为30 min。
1.5 光合过程能量分配参数的计算 采用以下公式计算最大光化学效率(
F v /
F m )、实际光化学效率(
Φ PSII )、调节性热耗散(
Φ NPQ )和非调节性热耗散(
Φ NO ):
(2) F v /F m = (F m - F o )/F m (3) Φ PSII = (F m ′ - F s )/F m ′(4) Φ NPQ = F s /F m ′ - F s /F m (5) Φ NO = 1 - Φ PSII - Φ NPQ 同时, 计算光化学反应的电子传输速率(
ETR ):
(6) ETR = APAR × f × Φ PSII (7) APAR = PAR × α l 式中,
APAR 为吸收光强,
f 为植物光系统II和光系统I的比例, 假设两个光系统均分光能,
f 取值0.5,
Φ PSII 为实际光化学效率。
PAR 为光合有效辐射,
α l 为经验性吸光系数, 取值0.84。同时根据能量守恒定律, 又可以得出:
(8) ETR = ETR A + ETR P 式中,
ETR A 为用于进行固碳部分的电子流,
ETR P 为用于进行光呼吸代谢部分的电子流。由于光呼吸过程中的甘氨酸脱羧作用, 一个羧化和氧化循环需要4个电子, 同时光呼吸过程中每2个O
2 释放1个CO
2 , 那么就有:
(9) ETR A = 4P n + 4R d + 0.5ETR P 式中,
P n 为净光合速率,
R d 为光下暗呼吸速率, 即为暗适应下的
P n 。将公式(8)和(9)重新整理就有:
(10) ETR A = 1/3(ETR + 8P n + 8R d )(11) ETR P = 2/3(ETR - 4P n - 4R d )(12) Φ A = Φ PSII × ETR A /ETR (13) Φ PR = Φ PSII × ETR P /ETR 式中,
Φ A 为固碳耗能占比, 表示光化学反应中用于实际固碳的比例。
Φ PR 为光呼吸耗能占比, 表示光化学反应中光呼吸代谢途径的比例, 根据能量守恒,
Φ A 、
Φ PR 、
Φ NO 和
Φ NPQ 相加等于1。
1.6 数据分析 基于整个生长季所有测量数据, 对黑沙蒿光合能量分配各组分、环境参数和叶性状参数进行相关性分析, 显著性水平设为0.05, 极显著水平设为0.01。建立基于叶性状参数的黑沙蒿光合能量分配组分的多元线性回归模型, 使用拟合优度
R 2 和赤池信息量准则(AIC)对模型进行评价, 通常情况下AIC值为负, 且模型的拟合效果越好AIC值越低。根据以往相关研究结果, 以0.1 m
3 ·m
-3 作为
SWC 的阈值, 1.0 kPa作为
VPD 的阈值(
Wu et al ., 2018 ), 将数据分为4组, 进行黑沙蒿光合能量分配和
SLA 的重复测量方差分析的显著性分析。因可用数据点数量较少, 分析前进行方差齐性检验和正态分布检验, 对质量不符合要求的数据进行对数转换, 以达到方差齐性。
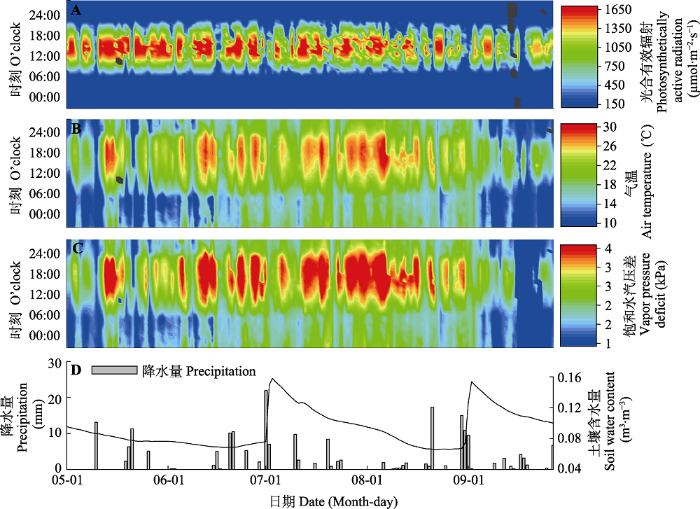
2 结果和分析 2.1 环境因子的季节动态 PAR 、
T a 和
VPD 都呈现先升高后降低的日变化趋势。如
图1A 所示,
PAR 在14:00左右达到峰值, 在季节尺度, 5-7月较高, 最高为1 890 μmol·m
-2 ·s
-1 (6月12日)。在
图1B 中,
T a 约在16:00达到峰值, 在季节尺度, 呈现出先升高后逐渐降低的趋势, 最高为31.6 ℃ (8月5日), 最低为1.4 ℃ (5月2日)。如
图1C 所示,
VPD 约在16:00达到峰值, 在季节尺度, 6-7月较高, 最高为4.4 kPa (8月5日)。由
图1D 可知,
SWC 在观测期持续下降, 随
P 变化, 在2次
P 较高的降雨事件后有较大抬升, 日平均值最高为0.16 m
3 ·m
-3 (7月2日), 最低为0.06 m
3 ·m
-3 (8月21日), 观测期内
P 为212.7 mm。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1毛乌素沙漠黑沙蒿样地环境因子的动态变化。 Fig. 1Dynamics in environmental factors in the sample plots of Artemisia ordosica in Mau Us desert. 2.2 黑沙蒿光合能量分配和叶性状参数的季节动态 由
图2A 可知, 观测期内的黑沙蒿光合能量分配各组分有明显的季节变化,
Φ A 均值为0.11, 在0.07-0.13内波动;
Φ PR 均值为0.18, 在0.11-0.23内波动;
Φ NPQ 均值为0.32, 在0.29-0.35内波动;
Φ NO 平均为0.39, 在0.34-0.46内波动。观测期内,
P n (
图2B )均值为23.3 μmol·m
-2 ·s
-1 , 在15.9-29.7 μmol·m
-2 ·s
-1 内波动。观测期内,
F v /
F m (
图2C )均值为0.79, 在0.75-0.81内波动。
E (
图2D )均值为11.45 mmol·m
2 ·s
-1 , 在5.30-17.93 mmol·m
2 ·s
-1 内波动。
g s (
图2E )均值为0.51 mol·m
2 ·s
-1 , 在0.36-0.72 mol·m
2 ·s
-1 内波动。
N area (
图2F )均值为3.98 g·m
-2 , 在3.61-4.36 g·m
-2 内波动。
SLA (
图2G )均值为63.28 cm
2 ·g
-1 , 在43.22-83.51 cm
2 ·g
-1 内波动。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2毛乌素沙漠黑沙蒿光合能量分配和叶性状参数动态变化(平均值±标准差)。 Fig. 2Dynamics in photosynthetic energy partitioning and leaf trait parameters of Artemisia ordosica in Mau Us desert (mean ± SD ). 2.3 黑沙蒿光合能量分配组分、环境参数和叶性状参数之间的相关性 由
表1 可知, 光合能量分配的光化学反应组分(
Φ A 和
Φ PR )呈极显著正相关关系, 热耗散组分(
Φ NO 和
Φ NPQ )呈极显著正相关关系。而
Φ A 和
Φ NO 、
Φ A 和
Φ NPQ 、
Φ PR 和
Φ NO 、
Φ PR 和
Φ NPQ 之间呈极显著负相关关系。
E 和
Φ A 、
E 和
Φ PR 呈极显著正相关关系,
E 和
Φ NO 极显著负相关。环境因子之间,
PAR 和
T a 极显著正相关,
PAR 和
VPD 显著正相关。对于叶性状参数,
g s 和
F v /
F m 、
N area 和
VPD 、
SLA 和
SWC 、
C Chl 和
PAR 显著正相关,
N area 和
SWC 、
SLA 和
VPD 显著负相关。
Table 1 表1 表1 黑沙蒿光合能量分配组分、环境参数和叶性状参数之间的相关性
Table 1
Relationship between photosynthetic energy partitioning, environmental and leaf trait parameters of Artemisia ordosica Φ A Φ PR Φ NPQ Φ NO PAR T a VPD SWC F v /F m E g s N area SLA C Chl Chl a/b Φ A 1.000 0 Φ PR 0.995 0** 1.000 0 Φ NPQ -0.921 6** -0.916 6** 1.000 0 Φ NO -0.960 8** -0.966 7** 0.785 9** 1.000 0 PAR -0.194 7 -0.214 2 0.170 1 0.214 8 1.000 0 T a -0.231 2 -0.224 3 0.199 6 0.225 0 0.540 2** 1.000 0 VPD -0.114 8 -0.053 7 0.110 6 0.043 6 0.706 3* 0.341 5 1.000 0 SWC 0.361 7 0.426 4 -0.347 9 -0.407 8 -0.506 8 -0.097 9 -0.526 0 1.000 0 F v /F m 0.058 9 0.099 8 -0.006 6 -0.131 3 -0.038 3 0.030 1 -0.252 6 0.251 5 1.000 0 E 0.619 5** 0.634 5** -0.521 1* -0.647 0** 0.185 0 0.339 0 0.184 4 0.176 1 -0.339 2 1.000 0 g s 0.225 7 0.271 4 -0.225 4 -0.254 6 -0.205 3 -0.419 4 -0.206 4 0.221 9 0.414 5* 0.130 5 1.000 0 N area 0.105 1 0.014 8 -0.109 4 0.001 5 0.132 2 -0.271 7 0.583 3* -0.646 3* -0.294 9 -0.144 6 -0.035 5 1.000 0 SLA -0.192 1 -0.127 2 0.057 8 0.195 9 -0.306 8 -0.055 1 -0.586 3* 0.569 2* 0.438 9 -0.101 8 0.389 7 -0.530 6 1.000 0 C Chl -0.266 9 -0.242 1 0.254 8 0.226 0 0.612 1* 0.423 2 0.345 0 -0.133 7 0.445 8 0.164 9 0.290 9 -0.120 4 -0.041 4 1.000 0 Chl a/b -0.030 8 -0.066 9 0.168 9 -0.024 8 0.171 5 -0.106 6 0.197 2 -0.144 8 -0.112 6 0.034 8 0.065 3 0.444 5 -0.004 7 0.251 4 1.000 0
Φ A , 固碳耗能占比; Φ NO , 非调节性热耗散耗能占比; Φ NPQ , 调节性热耗散耗能占比; Φ PR , 光呼吸耗能占比; Chl a/b, 叶绿素a/b比值; C Chl , 叶绿素含量; E, 蒸腾速率; F v /F m , 最大光化学效率; g s , 气孔导度; N area , 单位面积氮含量; PAR, 光合有效辐射; SLA, 比叶面积; SWC, 土壤含水量; T a , 空气温度; VPD, 饱和水汽压差。*, 在p < 0.05水平双侧显著相关; **, 在p < 0.01水平双侧极显著相关。Φ A , the fraction of energy used for CO2 assimilation; Φ NO , the fraction of non-regulated heat dissipation; Φ NPQ , the fraction of regulated heat dissipation; Φ PR , the fraction of energy used for photorespiration; Chl a/b, ratio of chlorophyll a to chlorophyll b; C Chl , chlorophyll content; E, transpiration rate; F v /F m , maximum photochemical efficiency; g s , stomatal conductance; N area , nitrogen per leaf area; PAR, photosynthetically active radiation; SLA, specific leaf area; SWC, soil water content; T a , air temperature; VPD, vapor pressure deficit. *, p < 0.05; **, p < 0.01.新窗口打开 |
下载CSV 2.4 黑沙蒿光合能量分配的多重叶片资源限制 由
表2 可知,
Φ A 和
Φ PR 模型内所有叶性状参数都为正相关系数,
Φ NO 和
Φ NPQ 模型内所有叶性状参数都为负相关系数。叶性状参数对黑沙蒿光合能量分配模型的解释度在57%-69%之间,
p 在0.11-0.29之间,
Φ A 的模型最优, AIC值为-50.72, AIC值最高为
Φ PR 的-42.29。在土壤水分限制(
SWC < 0.1 m
3 ·m
-3 )和空气水分限制(
VPD > 1.0 kPa)条件下,
Φ A 和
Φ PR 显著降低,
Φ NO 和
Φ NPQ 显著增加(
图3A ),
SLA 显著降低(
图3B )。
Table 2 表2 表2 黑沙蒿光合能量分配组分与叶性状参数的模型拟合参数
Table 2
Fitting parameters of the relationship between photosynthetic energy partitioning components of Artemisia ordosica and leaf trait parameters E g s N area SLA C Chl Chl a/b Y SSE R 2 p AIC Φ A 0.000 8 (0.001 2) 0.018 9 (0.051 5) 0.069 1 (0.033 8) 0.001 9 (0.000 7) 0.0347 0.004 2 (0.010 1) 0.075 3 (0.168 3) 0.000 3 0.57 0.29 -50.72 Φ PR 0.001 5 (0.336 7) 0.050 5 (0.103 0) 0.001 1 (0.067 6) 0.000 3 (0.001 3) 0.069 3 (0.112 2) 0.008 5 (0.020 8) 0.150 7 (0.336 7) 0.001 3 0.58 0.26 -42.29 Φ NPQ -0.000 9 (0.001 8) -0.014 9 (0.078 8) -0.049 3 (0.051 7) -0.001 1 (0.001 0) -0.007 4 (0.085 9) -0.007 3 (0.015 9) 0.572 9 (0.257 6) 0.000 8 0.59 0.25 -45.54 Φ NO -0.001 4 (0.002 1) -0.084 4 (0.093 7) -0.041 2 (0.061 5) -0.000 5 (0.001 2) -0.111 5 (0.102 1) -0.020 1 (0.018 9) 0.200 9 (0.306 3) 0.001 1 0.69 0.11 -43.44
Φ A , 固碳耗能占比; Φ NO , 非调节性热耗散耗能占比; Φ NPQ , 调节性热耗散耗能占比; Φ PR , 光呼吸耗能占比; AIC, 模型优良程度; Chl a/b, 叶绿素a/b比值; C Chl , 叶绿素含量; E, 蒸腾速率; g s , 气孔导度; N area , 单位面积氮含量; p, 拟合的显著程度; R 2 , 拟合程度; SLA, 比叶面积; SSE, 标准误差平方和; Y, 方程截距。括号内为对应参数95%置信区间。Φ A , the fraction of energy used for CO2 assimilation; Φ NO , the fraction of non-regulated heat dissipation; Φ NPQ , the fraction of regulated heat dissipation; Φ PR , the fraction of energy used for photorespiration; AIC, the goodness of the model; Chl a/b, ratio of chlorophyll a to chlorophyll b; C Chl , chlorophyll content; E, transpiration rate; g s , stomatal conductance; N area , nitrogen per leaf area; p, the degree of significance of the fit; R 2 , the degree of fit; SLA, specific leaf area; SSE, the sum of squared errors; Y, the intercept of the equation. Values in the parentheses are 95% confidence interval.新窗口打开 |
下载CSV 图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3不同土壤和空气水分条件下黑沙蒿的光合过程能量分配和比叶面积(平均值±标准差)。SWC, 土壤含水量; VPD, 饱和水汽压差。不同小写字母表示差异显著(p < 0.05)。 Fig. 3Photosynthetic energy partitioning and specific leaf area of Artemisia ordosica under different soil and air moisture conditions (mean ± SD ). SWC, soil water content; VPD, vapor pressure deficit. Different lowercase letters indicate significant differences (p < 0.05). 3 讨论 3.1 环境因子对黑沙蒿光合过程能量分配的影响 环境对植物光合过程能量分配的影响存在时间尺度的差异, 短期响应发生在秒至分钟内, 如光系统II的叶黄素循环(
靳川等, 2021 )。长期影响则可以通过调整光合色素浓度、Chl a/b和
SLA 等叶性状参数或光系统II的特定蛋白等途径实现, 通常存在几天到几周内(
李鑫豪等, 2019 )。在生长季内, 黑沙蒿光合过程能量分配与各环境因子均有很强的相关性(
表1 ), 结果表明光化学反应组分(
Φ A 、
Φ PR )和
PAR 呈负相关关系, 这是由于
PAR 的升高产生了更多的过剩光能, 黑沙蒿会瞬时激发叶黄素循环等持续性热耗散, 将过剩光能转移到热耗散组分(
Φ NPQ 和
Φ NO )。光化学反应组分和
T a 负相关, 说明高温可能抑制了黑沙蒿光系统II光反应酶或捕光蛋白的活性, 导致光合过程电子传递受限。植物细胞的原生质层抗凝结特性对于增加细胞内渗透压意义重大(
Savitch et al ., 2009 )。同地区相关研究结果也证明黑沙蒿光系统II对温度极敏感, 在清晨的低温条件下也发生了相似的抑制现象(
吴雅娟等, 2015 )。研究认为黑沙蒿能够通过原生质层的抗凝结特性增加细胞内渗透压, 从而以减小代谢速率的方式抵御极端温度胁迫。
水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 )。黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失。气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 )。然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累。另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少。在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 )。与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 )。蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤。黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 )。
3.2 黑沙蒿光合过程能量分配的调节机制 在荒漠地区高辐射、极端温度和干旱等多种恶劣气候条件下, 植物通过对光合过程能量分配的调节, 避免过剩光能对光合单元机构造成破坏。结果表明, 黑沙蒿光合过程能量分配的光化学组分和热耗散组分之间在生长季内表现出竞争关系(
表1 )。高辐射条件下, 叶片叶绿素分子产生的大量过剩激发能对光系统II反应中心造成极大的威胁, 植物将启动一系列能量耗散的权衡调节以维持光能吸收利用的平衡关系(
王燕凌等, 2015 ;
Ruban, 2016 )。
E 与
Φ A 和
Φ PR 极显著正相关, 说明黑沙蒿以提高蒸腾为代价增加其光合固碳能力, 并且过剩光能可以通过转移到光呼吸代谢途径进行消耗。研究发现光呼吸不仅与光合固碳、氮代谢、氨基酸形成等基础循环有关, 同时还参与了对环境逆境的抵抗(
姜闯道等, 2005 )。光呼吸虽然被看作光合作用的竞争过程, 但通过光呼吸可以清除大量有害代谢产物。另外光呼吸产生的CO
2 可以限制甘氨酸脱羧酶的活动范围, 从而提高核酮糖二磷酸羧化酶的羧化效率, 因此本研究中
Φ A 和
Φ PR 呈现出正相关的协同关系。
E 与
Φ NPQ 和
Φ NO 表现为负相关关系, 说明在蒸腾受限情况下, 黑沙蒿能够通过增加热耗散组分来耗散过剩光能。依赖于叶黄素循环的热耗散机制是保护植物光合单元机构免受灼伤的主要途径, 当过剩光能累积导致跨膜质子梯度增加时, 在脱环氧化酶的催化下, 紫黄质经由花药黄质后最终转化为玉米黄质, 同时产生大量的热量耗散过剩光能(
孙艳等, 2006 )。
研究发现植物可通过叶片厚度、叶绿素含量、叶面积等性状可塑性反应和相应的光合生理状态调控来适应环境变化(
李鑫豪等, 2019 )。单一叶性状参数与黑沙蒿光合能量分配各组分的关系并不显著, 但基于多叶性状参数的多元线性回归模型却有较低的AIC值(
表2 ), 说明黑沙蒿发挥生理可塑性对其叶性状整体进行调节, 而不是由单一性状决定的(
Burnham et al ., 2011 )。作为指示植物光合单元机构健康程度的重要参数,
F v /
F m 正常情况处于0.75以上, 若小于该范围则表示植物光合单元机构受损失活(
Murchie et al ., 2013 )。观测期内
F v /
F m 在0.75-0.85之间波动, 认为黑沙蒿光合单元机构通过不断自我调整和修复, 可以长期处于相对健康稳定的状态, 具有一定的适应和恢复力。在高
VPD 和低
SWC 的限制条件下, 黑沙蒿的
SLA 显著降低(
图3B )。
SLA 反映了植物在环境胁迫条件下的光合策略, 与叶片含水量、单位面积气孔数量和光能拦截均存在密切关联(
史作民等, 2015 )。本研究认为黑沙蒿在长期干旱或高蒸散条件下, 通过降低比叶面积等途径减少叶片水分流失, 以优化其蒸腾和固碳能力的匹配, 同时将大量过剩光能由光呼吸代谢途径转移到热耗散途径, 这一系列调节机制及过程充分反映出黑沙蒿对环境异质性的适应性和其生理可塑性。
致谢 感谢北京林业大学卫腾宙、周文君在野外测量工作中给予的帮助。
[1] Burnham KP Anderson DR Huyvaert KP 2011 ). AIC model selection and multimodel inference in behavioral ecology: some background, observations, and comparisonsBehavioral Ecology and Sociobiology 23 -35 . [本文引用: 1] [2] Chaves MM Osório J Pereira JS 2004 ). Water use efficiency and photosynthesis //Bacon MA. Water Use Efficiency in Plant Biology. Blackwell Publishing, Oxford, UK. [本文引用: 1] [3] Chen YN Chen YP Zhu CG Li WH 2019 ). The concept and mode of ecosystem sustainable management in arid desert areas in northwest ChinaActa Ecologica Sinica 7410 -7417 . [本文引用: 1] [ 陈亚宁 , 陈亚鹏 , 朱成刚 , 李卫红 (2019 ). 西北干旱荒漠区生态系统可持续管理理念与模式生态学报 , 39, 7410 -7417 .] [本文引用: 1] [4] Chong PF Li Y Su SP 2010 ). Diurnal change in chlorophyll fluorescence parameters of desert plant Reaumuria soongorica and its relationship with environmental factorsJournal of Desert Research 539 -545 . [本文引用: 1] [ 种培芳 , 李毅 , 苏世平 (2010 ). 荒漠植物红砂叶绿素荧光参数日变化及其与环境因子的关系中国沙漠 , 30, 539 -545 .] [本文引用: 1] [5] Galmés J Medrano H Flexas J 2007 ). Photosynthetic limitations in response to water stress and recovery in Mediterranean plants with different growth formsNew Phytologist 81 -93 . [本文引用: 1] [6] He YH Bai YE Wang HY Lin T Tian YL 2015 ). Effect of light stress on chlorophyll fluorescence and photorespiration of Ammopiptanthus mongolicusActa Agriculturae Boreali-Occidentalis Sinica 124 -130 . [本文引用: 1] [ 何炎红 , 白玉娥 , 王海燕 , 林涛 , 田有亮 (2015 ). 光胁迫对沙冬青叶绿素荧光特征和光呼吸的影响西北农业学报 , 24, 124 -130 .] [本文引用: 1] [7] Hou XW Li YJ Zhong Q Peng XX 2019 ). Recent progress of photorespiration pathway and its regulationPlant Physiology Journal 255 -264 . [本文引用: 1] [ 侯学文 , 李英杰 , 钟琪 , 彭新湘 (2019 ). 光呼吸代谢途径及其调控的研究进展植物生理学报 , 55, 255 -264 .] [本文引用: 1] [8] Huang JP Yu HP Dai AG Wei Y Kang LT 2017 ). Drylands face potential threat under 2 °C global warming targetNature Climate Change 417 -422 . [本文引用: 1] [9] Jia X Zha TS Gong JN Wang B Zhang YQ Wu B Qin SG Peltola H 2016 ). Carbon and water exchange over a temperate semi-arid shrubland during three years of contrasting precipitation and soil moisture patternsAgricultural and Forest Meteorology 120 -129 . [本文引用: 1] [10] Jiang CD Gao HY Zou Q Jiang GM Li LH 2005 ). The co-operation of leaf orientation, photorespiration and thermal dissipation alleviate photoinhibition in young leaves of soybean plantsActa Ecologica Sinica 319 -325 . [本文引用: 2] [ 姜闯道 , 高辉远 , 邹琦 , 蒋高明 , 李凌浩 (2005 ). 叶角、光呼吸和热耗散协同作用减轻大豆幼叶光抑制生态学报 , 25, 319 -325 .] [本文引用: 2] [11] Jin C Jiang Y Li XH Xu MZ Gao SJ Wei NN Jia X Tian Y Zha TS 2021 ). Multi-time scale property of environmental responses to photosystem II of Artemisia ordosica in Mu Us desertTransactions of the Chinese Society of Agricultural Engineering 152 -160 . [本文引用: 1] [ 靳川 , 蒋燕 , 李鑫豪 , 徐铭泽 , 高圣杰 , 魏宁宁 , 贾昕 , 田赟 , 查天山 (2021 ). 毛乌素沙地油蒿光系统II多时间尺度的环境响应特征农业工程学报 , 37, 152 -160 .] [本文引用: 1] [12] Jin C Zha TS Jia X Tian Y Zhou WJ Yang SB Guo ZF 2020 ). Dynamics of chlorophyll fluorescence parameters under drought condition for three desert shrub speciesJournal of Beijing Forestry University 8 ), 72 -80 . [本文引用: 1] [ 靳川 , 查天山 , 贾昕 , 田赟 , 周文君 , 杨双宝 , 郭子繁 (2020 ). 干旱环境3种荒漠灌木叶绿素荧光参数动态北京林业大学学报 , 42(8 ), 72 -80 .] [本文引用: 1] [13] Kang BW Liu JJ Sun JH Li YF 2010 ). Study on root distribution of Artemisa ordosica in Mu Us Sandy landResearch of Soil and Water Conservation 119 -123 . [本文引用: 1] [ 康博文 , 刘建军 , 孙建华 , 李岩峰 (2010 ). 陕北毛乌素沙漠黑沙蒿根系分布特征研究水土保持研究 , 17, 119 -123 .] [本文引用: 1] [14] Kramer DM Johnson G Kiirats O Edwards GE 2004 ). New fluorescence parameters for the determination of QA redox state and excitation energy fluxesPhotosynthesis Research 209 -218 . [本文引用: 1] [15] Li XH Yan HJ Wei TZ Zhou WJ Jia X Zha TS 2019 ). Relative changes of resource use efficiencies and their responses to environmental factors in Artemisia ordosica during growing seasonChinese Journal of Plant Ecology 889 -898 . [本文引用: 2] [ 李鑫豪 , 闫慧娟 , 卫腾宙 , 周文君 , 贾昕 , 查天山 (2019 ). 油蒿资源利用效率在生长季的相对变化及对环境因子的响应植物生态学报 , 43, 889 -898 .] [本文引用: 2] [16] Lin NF Tang J 2001 ). Study on the environmental evolution and the causes of desertification in arid and semiarid regions in ChinaScientia Geographica Sinica 24 -29 . [本文引用: 1] [ 林年丰 , 汤洁 (2001 ). 中国干旱半干旱区的环境演变与荒漠化的成因地理科学 , 21, 24 -29 .] [本文引用: 1] [17] Luo DD Wang CK Jin Y 2019 ). Stomatal regulation of plants in response to drought stressChinese Journal of Applied Ecology 4333 -4343 . [本文引用: 1] [ 罗丹丹 , 王传宽 , 金鹰 (2019 ). 植物应对干旱胁迫的气孔调节应用生态学报 , 30, 4333 -4343 .] [本文引用: 1] [18] Murchie EH Lawson T 2013 ). Chlorophyll fluorescence analysis: a guide to good practice and understanding some new applicationsJournal of Experimental Botany 3983 -3998 . [本文引用: 2] [19] Müller P Li XP Niyogi KK 2001 ). Non-photochemical quenching. A response to excess light energyPlant physiology 1558 -1566 . [本文引用: 1] [20] Ruan CJ Li DQ 2001 ). Stomatal conductance and influence factors of seabuckthorn in Loess Hilly RegionActa Botanica Boreali-occidentalia Sinica 30 -36 . [本文引用: 1] [ 阮成江 , 李代琼 (2001 ). 黄土丘陵区沙棘气孔导度及其影响因子西北植物学报 , 21, 30 -36 .] [本文引用: 1] [21] Ruban AV 2016 ). Nonphotochemical chlorophyll fluorescence quenching: mechanism and effectiveness in protecting plants from photodamagePlant Physiology 1903 -1916 . [本文引用: 2] [22] Savitch LV Ivanov AG Gudynaite-Savitch L Huner NPA Simmonds J 2009 ). Effects of low temperature stress on excitation energy partitioning and photoprotection in Zea mays Functional Plant Biology 37 -49 . [本文引用: 1] [23] Shi ZM Tang JC Cheng RM Luo D Liu SR 2015 ). A review of nitrogen allocation in leaves and factors in its effectsActa Ecologica Sinica 5909 -5919 . [本文引用: 1] [ 史作民 , 唐敬超 , 程瑞梅 , 罗达 , 刘世荣 (2015 ). 植物叶片氮分配及其影响因子研究进展生态学报 , 35, 5909 -5919 .] [本文引用: 1] [24] Sun AA Zhi YB Jiang PP Lü K Zhang DJ Li HL Zhang HL Wang YF Hua YP Hong G Gao JB 2019 ). Characteristics of and differences in photosynthesis in four desert plants in western OrdosActa Ecologica Sinica 4944 -4952 . [本文引用: 1] [ 孙安安 , 智颖飙 , 姜平平 , 吕凯 , 张德健 , 李红丽 , 张荷亮 , 王云飞 , 华宇鹏 , 红鸽 , 高健斌 (2019 ). 西鄂尔多斯4种荒漠植物光合作用特征与差异性生态学报 , 39, 4944 -4952 .] [本文引用: 1] [25] Sun Y Wang D Tong Z Yang Q Chang LL Wang LM He LP Wang XC 2015 ). Proteomic analysis of banana seedling leaf response to low temperatureChinese Agricultural Science Bulletin 216 -228 . [本文引用: 1] [ 孙勇 , 王丹 , 仝征 , 杨倩 , 常丽丽 , 王力敏 , 何丽平 , 王旭初 (2015 ). 香蕉幼苗叶片响应低温胁迫的比较蛋白质组学研究中国农学通报 , 31, 216 -228 .] [本文引用: 1] [26] Sun Y Xu WJ Fan AL 2006 ). Effects of salicylic acid on chlorophyll fluorescence and xanthophyll cycle in cucumber leaves under high temperature and strong lightChinese Journal of Applied Ecology 3399 -3402 . [本文引用: 1] [ 孙艳 , 徐伟君 , 范爱丽 (2006 ). 高温强光下水杨酸对黄瓜叶片叶绿素荧光和叶黄素循环的影响应用生态学报 , 17, 3399 -3402 .] [本文引用: 1] [27] Wang Y Lü GH Gao LJ Ren ML Su Q Sun LJ 2013 ). Stomatal conductance characteristics of desert species Poacynum pictum (Schrenk.) Baill of and the impact factorsJournal of Arid Land Resources and Environment 158 -163 . [本文引用: 1] [ 王芸 , 吕光辉 , 高丽娟 , 任曼丽 , 苏前 , 孙丽君 (2013 ). 荒漠植物白麻气孔导度特征及其影响因子研究干旱区资源与环境 , 27, 158 -163 .] [本文引用: 1] [28] Wang YL Liu J Li WB Li F 2015 ). Study on characteristics in photosynthesis, transpiration and water use efficiency of Tamarix hispida willdin the lower reaches of the Tarim river. Xinjiang Agricultural Sciences , 52, 292 -299 . [本文引用: 1] [ 王燕凌 , 刘君 , 李文兵 , 李芳 (2015 ). 塔里木河下游刚毛柽柳光合作用、蒸腾作用及水分利用效率特性研究新疆农业科学 , 52, 292 -299 .] [本文引用: 1] [29] Wu YJ Ren C Tian Y Zha TS Liu P Bai YJ Ma JY Lai ZR Bourque CPA 2018 ). Photosynthetic gas-exchange and PSII photochemical acclimation to drought in a native and non-native xerophytic species (Artemisia ordosica and Salix psammophila )Ecological Indicators 130 -138 . [本文引用: 1] [30] Wu YJ Zha TS Jia X Qin SG Li Y Wang B 2015 ). Temporal variation and controlling factors of photochemical efficiency and non-photo-chemical quenching in Artemisia ordosica Chinese Journal of Ecology 319 -325 . [本文引用: 1] [ 吴雅娟 , 查天山 , 贾昕 , 秦树高 , 李媛 , 王奔 (2015 ). 油蒿(Artemisia ordosica )光化学量子效率和非光化学淬灭的动态及其影响因子生态学杂志 , 34, 319 -325 .] [本文引用: 1] [31] Yang HX Zhang J Wu B Wang Y Li XS Xu B 2004 ). Adaptation of Artemisia ordosica to temperate arid sandy land and its roles in habitat shiftJournal of Beijing Normal University (Natural Science )684 -690 . [本文引用: 2] [ 杨洪晓 , 张金屯 , 吴波 , 王妍 , 李晓松 , 许彬 (2004 ). 油蒿(Artemisia ordosica )对半干旱区沙地生境的适应及其生态作用北京师范大学学报(自然科学版) , 40, 684 -690 .] [本文引用: 2] [32] Zha TS Wu YJ Jia X Zhang MY Bai YJ Liu P Ma JY Bourque CPA Peltola H 2017 ). Diurnal response of effective quantum yield of PSII photochemistry to irradiance as an indicator of photosynthetic acclimation to stressed environments revealed in a xerophytic speciesEcological Indicators , 74, 191 -197 . [本文引用: 1] [33] Zhang C Zhan DX Zhang PP Zhang YL Luo HH Zhang WF 2014 ). Responses of photorespiration and thermal dissipation in PSII to soil water in cotton bractsChinese Journal of Plant Ecology 387 -395 . [本文引用: 1] [ 张超 , 占东霞 , 张鹏鹏 , 张亚黎 , 罗宏海 , 张旺锋 (2014 ). 棉花苞叶光呼吸和PSII热耗散对土壤水分的响应植物生态学报 , 38, 387 -395 .] [本文引用: 1] [34] Zhao FH Yu GR 2008 ). A review on the coupled carbon and water cycles in the terrestrial ecosystemsProgress in Geography 32 -38 . [本文引用: 1] [ 赵风华 , 于贵瑞 (2008 ). 陆地生态系统碳-水耦合机制初探地理科学进展 , 27, 32 -38 .] [本文引用: 1] AIC model selection and multimodel inference in behavioral ecology: some background, observations, and comparisons
1
2011
... 研究发现植物可通过叶片厚度、叶绿素含量、叶面积等性状可塑性反应和相应的光合生理状态调控来适应环境变化(
李鑫豪等, 2019 ).单一叶性状参数与黑沙蒿光合能量分配各组分的关系并不显著, 但基于多叶性状参数的多元线性回归模型却有较低的AIC值(
表2 ), 说明黑沙蒿发挥生理可塑性对其叶性状整体进行调节, 而不是由单一性状决定的(
Burnham et al ., 2011 ).作为指示植物光合单元机构健康程度的重要参数,
F v /
F m 正常情况处于0.75以上, 若小于该范围则表示植物光合单元机构受损失活(
Murchie et al ., 2013 ).观测期内
F v /
F m 在0.75-0.85之间波动, 认为黑沙蒿光合单元机构通过不断自我调整和修复, 可以长期处于相对健康稳定的状态, 具有一定的适应和恢复力.在高
VPD 和低
SWC 的限制条件下, 黑沙蒿的
SLA 显著降低(
图3B ).
SLA 反映了植物在环境胁迫条件下的光合策略, 与叶片含水量、单位面积气孔数量和光能拦截均存在密切关联(
史作民等, 2015 ).本研究认为黑沙蒿在长期干旱或高蒸散条件下, 通过降低比叶面积等途径减少叶片水分流失, 以优化其蒸腾和固碳能力的匹配, 同时将大量过剩光能由光呼吸代谢途径转移到热耗散途径, 这一系列调节机制及过程充分反映出黑沙蒿对环境异质性的适应性和其生理可塑性. ...
1
2004
... 干旱半干旱区是维持生态稳定的关键屏障, 也是我国植被恢复的重点区域(
陈亚宁等, 2019 ).近年来气候恶化以及极端天气事件的频发, 该区植被经常遭受多种环境胁迫, 处于急剧退化的风险当中(
Huang et al ., 2017 ).植物通过光合作用将光能转化为化学能, 驱动生态系统碳循环(
赵风华和于贵瑞, 2018 ).光合过程各个步骤对外界环境的波动非常敏感, 根据植物光合特性能够快速了解其生理状态(
孙安安等, 2019 ).因此, 探究荒漠植物光合作用对环境因子的响应, 可以进一步明确其在逆境条件下的耐受力、恢复力和适应力(
Chaves et al ., 2004 ). ...
西北干旱荒漠区生态系统可持续管理理念与模式
1
2019
... 干旱半干旱区是维持生态稳定的关键屏障, 也是我国植被恢复的重点区域(
陈亚宁等, 2019 ).近年来气候恶化以及极端天气事件的频发, 该区植被经常遭受多种环境胁迫, 处于急剧退化的风险当中(
Huang et al ., 2017 ).植物通过光合作用将光能转化为化学能, 驱动生态系统碳循环(
赵风华和于贵瑞, 2018 ).光合过程各个步骤对外界环境的波动非常敏感, 根据植物光合特性能够快速了解其生理状态(
孙安安等, 2019 ).因此, 探究荒漠植物光合作用对环境因子的响应, 可以进一步明确其在逆境条件下的耐受力、恢复力和适应力(
Chaves et al ., 2004 ). ...
西北干旱荒漠区生态系统可持续管理理念与模式
1
2019
... 干旱半干旱区是维持生态稳定的关键屏障, 也是我国植被恢复的重点区域(
陈亚宁等, 2019 ).近年来气候恶化以及极端天气事件的频发, 该区植被经常遭受多种环境胁迫, 处于急剧退化的风险当中(
Huang et al ., 2017 ).植物通过光合作用将光能转化为化学能, 驱动生态系统碳循环(
赵风华和于贵瑞, 2018 ).光合过程各个步骤对外界环境的波动非常敏感, 根据植物光合特性能够快速了解其生理状态(
孙安安等, 2019 ).因此, 探究荒漠植物光合作用对环境因子的响应, 可以进一步明确其在逆境条件下的耐受力、恢复力和适应力(
Chaves et al ., 2004 ). ...
荒漠植物红砂叶绿素荧光参数日变化及其与环境因子的关系
1
2010
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
荒漠植物红砂叶绿素荧光参数日变化及其与环境因子的关系
1
2010
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
Photosynthetic limitations in response to water stress and recovery in Mediterranean plants with different growth forms
1
2007
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
光胁迫对沙冬青叶绿素荧光特征和光呼吸的影响
1
2015
... 荒漠生态系统具有干旱、高太阳辐射、高蒸腾和极端温度等环境特征(
Jia et al ., 2016 ).夏季荒漠的强烈太阳辐射远超植物光合所需, 已有研究证明荒漠植物通过调整热耗散等途径进行调节, 但实际上仍然会产生大量活性氧, 对植物光合机构造成极大威胁, 此时光呼吸过程的存在价值就显得尤为重要(
何炎红等, 2015 ).其次, 荒漠气候极端温度也会抑制卡尔文循环相关蛋白酶, 从而导致光合效率降低(
孙勇等, 2015 ).另外, 荒漠干燥空气导致的高蒸腾会促使植物气孔关闭, 影响了水的传输, 限制了光合过程中相关电子传输速率(
阮成江和李代琼, 2001 ).因此, 波动环境下荒漠植物光合过程能量分配的相对变化和内在调节机制可能具有明显特征. ...
光胁迫对沙冬青叶绿素荧光特征和光呼吸的影响
1
2015
... 荒漠生态系统具有干旱、高太阳辐射、高蒸腾和极端温度等环境特征(
Jia et al ., 2016 ).夏季荒漠的强烈太阳辐射远超植物光合所需, 已有研究证明荒漠植物通过调整热耗散等途径进行调节, 但实际上仍然会产生大量活性氧, 对植物光合机构造成极大威胁, 此时光呼吸过程的存在价值就显得尤为重要(
何炎红等, 2015 ).其次, 荒漠气候极端温度也会抑制卡尔文循环相关蛋白酶, 从而导致光合效率降低(
孙勇等, 2015 ).另外, 荒漠干燥空气导致的高蒸腾会促使植物气孔关闭, 影响了水的传输, 限制了光合过程中相关电子传输速率(
阮成江和李代琼, 2001 ).因此, 波动环境下荒漠植物光合过程能量分配的相对变化和内在调节机制可能具有明显特征. ...
光呼吸代谢途径及其调控的研究进展
1
2019
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
光呼吸代谢途径及其调控的研究进展
1
2019
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
Drylands face potential threat under 2 °C global warming target
1
2017
... 干旱半干旱区是维持生态稳定的关键屏障, 也是我国植被恢复的重点区域(
陈亚宁等, 2019 ).近年来气候恶化以及极端天气事件的频发, 该区植被经常遭受多种环境胁迫, 处于急剧退化的风险当中(
Huang et al ., 2017 ).植物通过光合作用将光能转化为化学能, 驱动生态系统碳循环(
赵风华和于贵瑞, 2018 ).光合过程各个步骤对外界环境的波动非常敏感, 根据植物光合特性能够快速了解其生理状态(
孙安安等, 2019 ).因此, 探究荒漠植物光合作用对环境因子的响应, 可以进一步明确其在逆境条件下的耐受力、恢复力和适应力(
Chaves et al ., 2004 ). ...
Carbon and water exchange over a temperate semi-arid shrubland during three years of contrasting precipitation and soil moisture patterns
1
2016
... 荒漠生态系统具有干旱、高太阳辐射、高蒸腾和极端温度等环境特征(
Jia et al ., 2016 ).夏季荒漠的强烈太阳辐射远超植物光合所需, 已有研究证明荒漠植物通过调整热耗散等途径进行调节, 但实际上仍然会产生大量活性氧, 对植物光合机构造成极大威胁, 此时光呼吸过程的存在价值就显得尤为重要(
何炎红等, 2015 ).其次, 荒漠气候极端温度也会抑制卡尔文循环相关蛋白酶, 从而导致光合效率降低(
孙勇等, 2015 ).另外, 荒漠干燥空气导致的高蒸腾会促使植物气孔关闭, 影响了水的传输, 限制了光合过程中相关电子传输速率(
阮成江和李代琼, 2001 ).因此, 波动环境下荒漠植物光合过程能量分配的相对变化和内在调节机制可能具有明显特征. ...
叶角、光呼吸和热耗散协同作用减轻大豆幼叶光抑制
2
2005
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
... 在荒漠地区高辐射、极端温度和干旱等多种恶劣气候条件下, 植物通过对光合过程能量分配的调节, 避免过剩光能对光合单元机构造成破坏.结果表明, 黑沙蒿光合过程能量分配的光化学组分和热耗散组分之间在生长季内表现出竞争关系(
表1 ).高辐射条件下, 叶片叶绿素分子产生的大量过剩激发能对光系统II反应中心造成极大的威胁, 植物将启动一系列能量耗散的权衡调节以维持光能吸收利用的平衡关系(
王燕凌等, 2015 ;
Ruban, 2016 ).
E 与
Φ A 和
Φ PR 极显著正相关, 说明黑沙蒿以提高蒸腾为代价增加其光合固碳能力, 并且过剩光能可以通过转移到光呼吸代谢途径进行消耗.研究发现光呼吸不仅与光合固碳、氮代谢、氨基酸形成等基础循环有关, 同时还参与了对环境逆境的抵抗(
姜闯道等, 2005 ).光呼吸虽然被看作光合作用的竞争过程, 但通过光呼吸可以清除大量有害代谢产物.另外光呼吸产生的CO
2 可以限制甘氨酸脱羧酶的活动范围, 从而提高核酮糖二磷酸羧化酶的羧化效率, 因此本研究中
Φ A 和
Φ PR 呈现出正相关的协同关系.
E 与
Φ NPQ 和
Φ NO 表现为负相关关系, 说明在蒸腾受限情况下, 黑沙蒿能够通过增加热耗散组分来耗散过剩光能.依赖于叶黄素循环的热耗散机制是保护植物光合单元机构免受灼伤的主要途径, 当过剩光能累积导致跨膜质子梯度增加时, 在脱环氧化酶的催化下, 紫黄质经由花药黄质后最终转化为玉米黄质, 同时产生大量的热量耗散过剩光能(
孙艳等, 2006 ). ...
叶角、光呼吸和热耗散协同作用减轻大豆幼叶光抑制
2
2005
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
... 在荒漠地区高辐射、极端温度和干旱等多种恶劣气候条件下, 植物通过对光合过程能量分配的调节, 避免过剩光能对光合单元机构造成破坏.结果表明, 黑沙蒿光合过程能量分配的光化学组分和热耗散组分之间在生长季内表现出竞争关系(
表1 ).高辐射条件下, 叶片叶绿素分子产生的大量过剩激发能对光系统II反应中心造成极大的威胁, 植物将启动一系列能量耗散的权衡调节以维持光能吸收利用的平衡关系(
王燕凌等, 2015 ;
Ruban, 2016 ).
E 与
Φ A 和
Φ PR 极显著正相关, 说明黑沙蒿以提高蒸腾为代价增加其光合固碳能力, 并且过剩光能可以通过转移到光呼吸代谢途径进行消耗.研究发现光呼吸不仅与光合固碳、氮代谢、氨基酸形成等基础循环有关, 同时还参与了对环境逆境的抵抗(
姜闯道等, 2005 ).光呼吸虽然被看作光合作用的竞争过程, 但通过光呼吸可以清除大量有害代谢产物.另外光呼吸产生的CO
2 可以限制甘氨酸脱羧酶的活动范围, 从而提高核酮糖二磷酸羧化酶的羧化效率, 因此本研究中
Φ A 和
Φ PR 呈现出正相关的协同关系.
E 与
Φ NPQ 和
Φ NO 表现为负相关关系, 说明在蒸腾受限情况下, 黑沙蒿能够通过增加热耗散组分来耗散过剩光能.依赖于叶黄素循环的热耗散机制是保护植物光合单元机构免受灼伤的主要途径, 当过剩光能累积导致跨膜质子梯度增加时, 在脱环氧化酶的催化下, 紫黄质经由花药黄质后最终转化为玉米黄质, 同时产生大量的热量耗散过剩光能(
孙艳等, 2006 ). ...
毛乌素沙地油蒿光系统II多时间尺度的环境响应特征
1
2021
... 环境对植物光合过程能量分配的影响存在时间尺度的差异, 短期响应发生在秒至分钟内, 如光系统II的叶黄素循环(
靳川等, 2021 ).长期影响则可以通过调整光合色素浓度、Chl a/b和
SLA 等叶性状参数或光系统II的特定蛋白等途径实现, 通常存在几天到几周内(
李鑫豪等, 2019 ).在生长季内, 黑沙蒿光合过程能量分配与各环境因子均有很强的相关性(
表1 ), 结果表明光化学反应组分(
Φ A 、
Φ PR )和
PAR 呈负相关关系, 这是由于
PAR 的升高产生了更多的过剩光能, 黑沙蒿会瞬时激发叶黄素循环等持续性热耗散, 将过剩光能转移到热耗散组分(
Φ NPQ 和
Φ NO ).光化学反应组分和
T a 负相关, 说明高温可能抑制了黑沙蒿光系统II光反应酶或捕光蛋白的活性, 导致光合过程电子传递受限.植物细胞的原生质层抗凝结特性对于增加细胞内渗透压意义重大(
Savitch et al ., 2009 ).同地区相关研究结果也证明黑沙蒿光系统II对温度极敏感, 在清晨的低温条件下也发生了相似的抑制现象(
吴雅娟等, 2015 ).研究认为黑沙蒿能够通过原生质层的抗凝结特性增加细胞内渗透压, 从而以减小代谢速率的方式抵御极端温度胁迫. ...
毛乌素沙地油蒿光系统II多时间尺度的环境响应特征
1
2021
... 环境对植物光合过程能量分配的影响存在时间尺度的差异, 短期响应发生在秒至分钟内, 如光系统II的叶黄素循环(
靳川等, 2021 ).长期影响则可以通过调整光合色素浓度、Chl a/b和
SLA 等叶性状参数或光系统II的特定蛋白等途径实现, 通常存在几天到几周内(
李鑫豪等, 2019 ).在生长季内, 黑沙蒿光合过程能量分配与各环境因子均有很强的相关性(
表1 ), 结果表明光化学反应组分(
Φ A 、
Φ PR )和
PAR 呈负相关关系, 这是由于
PAR 的升高产生了更多的过剩光能, 黑沙蒿会瞬时激发叶黄素循环等持续性热耗散, 将过剩光能转移到热耗散组分(
Φ NPQ 和
Φ NO ).光化学反应组分和
T a 负相关, 说明高温可能抑制了黑沙蒿光系统II光反应酶或捕光蛋白的活性, 导致光合过程电子传递受限.植物细胞的原生质层抗凝结特性对于增加细胞内渗透压意义重大(
Savitch et al ., 2009 ).同地区相关研究结果也证明黑沙蒿光系统II对温度极敏感, 在清晨的低温条件下也发生了相似的抑制现象(
吴雅娟等, 2015 ).研究认为黑沙蒿能够通过原生质层的抗凝结特性增加细胞内渗透压, 从而以减小代谢速率的方式抵御极端温度胁迫. ...
干旱环境3种荒漠灌木叶绿素荧光参数动态
1
2020
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
干旱环境3种荒漠灌木叶绿素荧光参数动态
1
2020
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
陕北毛乌素沙漠黑沙蒿根系分布特征研究
1
2010
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
陕北毛乌素沙漠黑沙蒿根系分布特征研究
1
2010
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
New fluorescence parameters for the determination of QA redox state and excitation energy fluxes
1
2004
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
油蒿资源利用效率在生长季的相对变化及对环境因子的响应
2
2019
... 环境对植物光合过程能量分配的影响存在时间尺度的差异, 短期响应发生在秒至分钟内, 如光系统II的叶黄素循环(
靳川等, 2021 ).长期影响则可以通过调整光合色素浓度、Chl a/b和
SLA 等叶性状参数或光系统II的特定蛋白等途径实现, 通常存在几天到几周内(
李鑫豪等, 2019 ).在生长季内, 黑沙蒿光合过程能量分配与各环境因子均有很强的相关性(
表1 ), 结果表明光化学反应组分(
Φ A 、
Φ PR )和
PAR 呈负相关关系, 这是由于
PAR 的升高产生了更多的过剩光能, 黑沙蒿会瞬时激发叶黄素循环等持续性热耗散, 将过剩光能转移到热耗散组分(
Φ NPQ 和
Φ NO ).光化学反应组分和
T a 负相关, 说明高温可能抑制了黑沙蒿光系统II光反应酶或捕光蛋白的活性, 导致光合过程电子传递受限.植物细胞的原生质层抗凝结特性对于增加细胞内渗透压意义重大(
Savitch et al ., 2009 ).同地区相关研究结果也证明黑沙蒿光系统II对温度极敏感, 在清晨的低温条件下也发生了相似的抑制现象(
吴雅娟等, 2015 ).研究认为黑沙蒿能够通过原生质层的抗凝结特性增加细胞内渗透压, 从而以减小代谢速率的方式抵御极端温度胁迫. ...
... 研究发现植物可通过叶片厚度、叶绿素含量、叶面积等性状可塑性反应和相应的光合生理状态调控来适应环境变化(
李鑫豪等, 2019 ).单一叶性状参数与黑沙蒿光合能量分配各组分的关系并不显著, 但基于多叶性状参数的多元线性回归模型却有较低的AIC值(
表2 ), 说明黑沙蒿发挥生理可塑性对其叶性状整体进行调节, 而不是由单一性状决定的(
Burnham et al ., 2011 ).作为指示植物光合单元机构健康程度的重要参数,
F v /
F m 正常情况处于0.75以上, 若小于该范围则表示植物光合单元机构受损失活(
Murchie et al ., 2013 ).观测期内
F v /
F m 在0.75-0.85之间波动, 认为黑沙蒿光合单元机构通过不断自我调整和修复, 可以长期处于相对健康稳定的状态, 具有一定的适应和恢复力.在高
VPD 和低
SWC 的限制条件下, 黑沙蒿的
SLA 显著降低(
图3B ).
SLA 反映了植物在环境胁迫条件下的光合策略, 与叶片含水量、单位面积气孔数量和光能拦截均存在密切关联(
史作民等, 2015 ).本研究认为黑沙蒿在长期干旱或高蒸散条件下, 通过降低比叶面积等途径减少叶片水分流失, 以优化其蒸腾和固碳能力的匹配, 同时将大量过剩光能由光呼吸代谢途径转移到热耗散途径, 这一系列调节机制及过程充分反映出黑沙蒿对环境异质性的适应性和其生理可塑性. ...
油蒿资源利用效率在生长季的相对变化及对环境因子的响应
2
2019
... 环境对植物光合过程能量分配的影响存在时间尺度的差异, 短期响应发生在秒至分钟内, 如光系统II的叶黄素循环(
靳川等, 2021 ).长期影响则可以通过调整光合色素浓度、Chl a/b和
SLA 等叶性状参数或光系统II的特定蛋白等途径实现, 通常存在几天到几周内(
李鑫豪等, 2019 ).在生长季内, 黑沙蒿光合过程能量分配与各环境因子均有很强的相关性(
表1 ), 结果表明光化学反应组分(
Φ A 、
Φ PR )和
PAR 呈负相关关系, 这是由于
PAR 的升高产生了更多的过剩光能, 黑沙蒿会瞬时激发叶黄素循环等持续性热耗散, 将过剩光能转移到热耗散组分(
Φ NPQ 和
Φ NO ).光化学反应组分和
T a 负相关, 说明高温可能抑制了黑沙蒿光系统II光反应酶或捕光蛋白的活性, 导致光合过程电子传递受限.植物细胞的原生质层抗凝结特性对于增加细胞内渗透压意义重大(
Savitch et al ., 2009 ).同地区相关研究结果也证明黑沙蒿光系统II对温度极敏感, 在清晨的低温条件下也发生了相似的抑制现象(
吴雅娟等, 2015 ).研究认为黑沙蒿能够通过原生质层的抗凝结特性增加细胞内渗透压, 从而以减小代谢速率的方式抵御极端温度胁迫. ...
... 研究发现植物可通过叶片厚度、叶绿素含量、叶面积等性状可塑性反应和相应的光合生理状态调控来适应环境变化(
李鑫豪等, 2019 ).单一叶性状参数与黑沙蒿光合能量分配各组分的关系并不显著, 但基于多叶性状参数的多元线性回归模型却有较低的AIC值(
表2 ), 说明黑沙蒿发挥生理可塑性对其叶性状整体进行调节, 而不是由单一性状决定的(
Burnham et al ., 2011 ).作为指示植物光合单元机构健康程度的重要参数,
F v /
F m 正常情况处于0.75以上, 若小于该范围则表示植物光合单元机构受损失活(
Murchie et al ., 2013 ).观测期内
F v /
F m 在0.75-0.85之间波动, 认为黑沙蒿光合单元机构通过不断自我调整和修复, 可以长期处于相对健康稳定的状态, 具有一定的适应和恢复力.在高
VPD 和低
SWC 的限制条件下, 黑沙蒿的
SLA 显著降低(
图3B ).
SLA 反映了植物在环境胁迫条件下的光合策略, 与叶片含水量、单位面积气孔数量和光能拦截均存在密切关联(
史作民等, 2015 ).本研究认为黑沙蒿在长期干旱或高蒸散条件下, 通过降低比叶面积等途径减少叶片水分流失, 以优化其蒸腾和固碳能力的匹配, 同时将大量过剩光能由光呼吸代谢途径转移到热耗散途径, 这一系列调节机制及过程充分反映出黑沙蒿对环境异质性的适应性和其生理可塑性. ...
中国干旱半干旱区的环境演变与荒漠化的成因
1
2001
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
中国干旱半干旱区的环境演变与荒漠化的成因
1
2001
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
植物应对干旱胁迫的气孔调节
1
2019
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
植物应对干旱胁迫的气孔调节
1
2019
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
Chlorophyll fluorescence analysis: a guide to good practice and understanding some new applications
2
2013
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
... 研究发现植物可通过叶片厚度、叶绿素含量、叶面积等性状可塑性反应和相应的光合生理状态调控来适应环境变化(
李鑫豪等, 2019 ).单一叶性状参数与黑沙蒿光合能量分配各组分的关系并不显著, 但基于多叶性状参数的多元线性回归模型却有较低的AIC值(
表2 ), 说明黑沙蒿发挥生理可塑性对其叶性状整体进行调节, 而不是由单一性状决定的(
Burnham et al ., 2011 ).作为指示植物光合单元机构健康程度的重要参数,
F v /
F m 正常情况处于0.75以上, 若小于该范围则表示植物光合单元机构受损失活(
Murchie et al ., 2013 ).观测期内
F v /
F m 在0.75-0.85之间波动, 认为黑沙蒿光合单元机构通过不断自我调整和修复, 可以长期处于相对健康稳定的状态, 具有一定的适应和恢复力.在高
VPD 和低
SWC 的限制条件下, 黑沙蒿的
SLA 显著降低(
图3B ).
SLA 反映了植物在环境胁迫条件下的光合策略, 与叶片含水量、单位面积气孔数量和光能拦截均存在密切关联(
史作民等, 2015 ).本研究认为黑沙蒿在长期干旱或高蒸散条件下, 通过降低比叶面积等途径减少叶片水分流失, 以优化其蒸腾和固碳能力的匹配, 同时将大量过剩光能由光呼吸代谢途径转移到热耗散途径, 这一系列调节机制及过程充分反映出黑沙蒿对环境异质性的适应性和其生理可塑性. ...
Non-photochemical quenching. A response to excess light energy
1
2001
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
黄土丘陵区沙棘气孔导度及其影响因子
1
2001
... 荒漠生态系统具有干旱、高太阳辐射、高蒸腾和极端温度等环境特征(
Jia et al ., 2016 ).夏季荒漠的强烈太阳辐射远超植物光合所需, 已有研究证明荒漠植物通过调整热耗散等途径进行调节, 但实际上仍然会产生大量活性氧, 对植物光合机构造成极大威胁, 此时光呼吸过程的存在价值就显得尤为重要(
何炎红等, 2015 ).其次, 荒漠气候极端温度也会抑制卡尔文循环相关蛋白酶, 从而导致光合效率降低(
孙勇等, 2015 ).另外, 荒漠干燥空气导致的高蒸腾会促使植物气孔关闭, 影响了水的传输, 限制了光合过程中相关电子传输速率(
阮成江和李代琼, 2001 ).因此, 波动环境下荒漠植物光合过程能量分配的相对变化和内在调节机制可能具有明显特征. ...
黄土丘陵区沙棘气孔导度及其影响因子
1
2001
... 荒漠生态系统具有干旱、高太阳辐射、高蒸腾和极端温度等环境特征(
Jia et al ., 2016 ).夏季荒漠的强烈太阳辐射远超植物光合所需, 已有研究证明荒漠植物通过调整热耗散等途径进行调节, 但实际上仍然会产生大量活性氧, 对植物光合机构造成极大威胁, 此时光呼吸过程的存在价值就显得尤为重要(
何炎红等, 2015 ).其次, 荒漠气候极端温度也会抑制卡尔文循环相关蛋白酶, 从而导致光合效率降低(
孙勇等, 2015 ).另外, 荒漠干燥空气导致的高蒸腾会促使植物气孔关闭, 影响了水的传输, 限制了光合过程中相关电子传输速率(
阮成江和李代琼, 2001 ).因此, 波动环境下荒漠植物光合过程能量分配的相对变化和内在调节机制可能具有明显特征. ...
Nonphotochemical chlorophyll fluorescence quenching: mechanism and effectiveness in protecting plants from photodamage
2
2016
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
... 在荒漠地区高辐射、极端温度和干旱等多种恶劣气候条件下, 植物通过对光合过程能量分配的调节, 避免过剩光能对光合单元机构造成破坏.结果表明, 黑沙蒿光合过程能量分配的光化学组分和热耗散组分之间在生长季内表现出竞争关系(
表1 ).高辐射条件下, 叶片叶绿素分子产生的大量过剩激发能对光系统II反应中心造成极大的威胁, 植物将启动一系列能量耗散的权衡调节以维持光能吸收利用的平衡关系(
王燕凌等, 2015 ;
Ruban, 2016 ).
E 与
Φ A 和
Φ PR 极显著正相关, 说明黑沙蒿以提高蒸腾为代价增加其光合固碳能力, 并且过剩光能可以通过转移到光呼吸代谢途径进行消耗.研究发现光呼吸不仅与光合固碳、氮代谢、氨基酸形成等基础循环有关, 同时还参与了对环境逆境的抵抗(
姜闯道等, 2005 ).光呼吸虽然被看作光合作用的竞争过程, 但通过光呼吸可以清除大量有害代谢产物.另外光呼吸产生的CO
2 可以限制甘氨酸脱羧酶的活动范围, 从而提高核酮糖二磷酸羧化酶的羧化效率, 因此本研究中
Φ A 和
Φ PR 呈现出正相关的协同关系.
E 与
Φ NPQ 和
Φ NO 表现为负相关关系, 说明在蒸腾受限情况下, 黑沙蒿能够通过增加热耗散组分来耗散过剩光能.依赖于叶黄素循环的热耗散机制是保护植物光合单元机构免受灼伤的主要途径, 当过剩光能累积导致跨膜质子梯度增加时, 在脱环氧化酶的催化下, 紫黄质经由花药黄质后最终转化为玉米黄质, 同时产生大量的热量耗散过剩光能(
孙艳等, 2006 ). ...
Effects of low temperature stress on excitation energy partitioning and photoprotection in
Zea mays 1
2009
... 环境对植物光合过程能量分配的影响存在时间尺度的差异, 短期响应发生在秒至分钟内, 如光系统II的叶黄素循环(
靳川等, 2021 ).长期影响则可以通过调整光合色素浓度、Chl a/b和
SLA 等叶性状参数或光系统II的特定蛋白等途径实现, 通常存在几天到几周内(
李鑫豪等, 2019 ).在生长季内, 黑沙蒿光合过程能量分配与各环境因子均有很强的相关性(
表1 ), 结果表明光化学反应组分(
Φ A 、
Φ PR )和
PAR 呈负相关关系, 这是由于
PAR 的升高产生了更多的过剩光能, 黑沙蒿会瞬时激发叶黄素循环等持续性热耗散, 将过剩光能转移到热耗散组分(
Φ NPQ 和
Φ NO ).光化学反应组分和
T a 负相关, 说明高温可能抑制了黑沙蒿光系统II光反应酶或捕光蛋白的活性, 导致光合过程电子传递受限.植物细胞的原生质层抗凝结特性对于增加细胞内渗透压意义重大(
Savitch et al ., 2009 ).同地区相关研究结果也证明黑沙蒿光系统II对温度极敏感, 在清晨的低温条件下也发生了相似的抑制现象(
吴雅娟等, 2015 ).研究认为黑沙蒿能够通过原生质层的抗凝结特性增加细胞内渗透压, 从而以减小代谢速率的方式抵御极端温度胁迫. ...
植物叶片氮分配及其影响因子研究进展
1
2015
... 研究发现植物可通过叶片厚度、叶绿素含量、叶面积等性状可塑性反应和相应的光合生理状态调控来适应环境变化(
李鑫豪等, 2019 ).单一叶性状参数与黑沙蒿光合能量分配各组分的关系并不显著, 但基于多叶性状参数的多元线性回归模型却有较低的AIC值(
表2 ), 说明黑沙蒿发挥生理可塑性对其叶性状整体进行调节, 而不是由单一性状决定的(
Burnham et al ., 2011 ).作为指示植物光合单元机构健康程度的重要参数,
F v /
F m 正常情况处于0.75以上, 若小于该范围则表示植物光合单元机构受损失活(
Murchie et al ., 2013 ).观测期内
F v /
F m 在0.75-0.85之间波动, 认为黑沙蒿光合单元机构通过不断自我调整和修复, 可以长期处于相对健康稳定的状态, 具有一定的适应和恢复力.在高
VPD 和低
SWC 的限制条件下, 黑沙蒿的
SLA 显著降低(
图3B ).
SLA 反映了植物在环境胁迫条件下的光合策略, 与叶片含水量、单位面积气孔数量和光能拦截均存在密切关联(
史作民等, 2015 ).本研究认为黑沙蒿在长期干旱或高蒸散条件下, 通过降低比叶面积等途径减少叶片水分流失, 以优化其蒸腾和固碳能力的匹配, 同时将大量过剩光能由光呼吸代谢途径转移到热耗散途径, 这一系列调节机制及过程充分反映出黑沙蒿对环境异质性的适应性和其生理可塑性. ...
植物叶片氮分配及其影响因子研究进展
1
2015
... 研究发现植物可通过叶片厚度、叶绿素含量、叶面积等性状可塑性反应和相应的光合生理状态调控来适应环境变化(
李鑫豪等, 2019 ).单一叶性状参数与黑沙蒿光合能量分配各组分的关系并不显著, 但基于多叶性状参数的多元线性回归模型却有较低的AIC值(
表2 ), 说明黑沙蒿发挥生理可塑性对其叶性状整体进行调节, 而不是由单一性状决定的(
Burnham et al ., 2011 ).作为指示植物光合单元机构健康程度的重要参数,
F v /
F m 正常情况处于0.75以上, 若小于该范围则表示植物光合单元机构受损失活(
Murchie et al ., 2013 ).观测期内
F v /
F m 在0.75-0.85之间波动, 认为黑沙蒿光合单元机构通过不断自我调整和修复, 可以长期处于相对健康稳定的状态, 具有一定的适应和恢复力.在高
VPD 和低
SWC 的限制条件下, 黑沙蒿的
SLA 显著降低(
图3B ).
SLA 反映了植物在环境胁迫条件下的光合策略, 与叶片含水量、单位面积气孔数量和光能拦截均存在密切关联(
史作民等, 2015 ).本研究认为黑沙蒿在长期干旱或高蒸散条件下, 通过降低比叶面积等途径减少叶片水分流失, 以优化其蒸腾和固碳能力的匹配, 同时将大量过剩光能由光呼吸代谢途径转移到热耗散途径, 这一系列调节机制及过程充分反映出黑沙蒿对环境异质性的适应性和其生理可塑性. ...
西鄂尔多斯4种荒漠植物光合作用特征与差异性
1
2019
... 干旱半干旱区是维持生态稳定的关键屏障, 也是我国植被恢复的重点区域(
陈亚宁等, 2019 ).近年来气候恶化以及极端天气事件的频发, 该区植被经常遭受多种环境胁迫, 处于急剧退化的风险当中(
Huang et al ., 2017 ).植物通过光合作用将光能转化为化学能, 驱动生态系统碳循环(
赵风华和于贵瑞, 2018 ).光合过程各个步骤对外界环境的波动非常敏感, 根据植物光合特性能够快速了解其生理状态(
孙安安等, 2019 ).因此, 探究荒漠植物光合作用对环境因子的响应, 可以进一步明确其在逆境条件下的耐受力、恢复力和适应力(
Chaves et al ., 2004 ). ...
西鄂尔多斯4种荒漠植物光合作用特征与差异性
1
2019
... 干旱半干旱区是维持生态稳定的关键屏障, 也是我国植被恢复的重点区域(
陈亚宁等, 2019 ).近年来气候恶化以及极端天气事件的频发, 该区植被经常遭受多种环境胁迫, 处于急剧退化的风险当中(
Huang et al ., 2017 ).植物通过光合作用将光能转化为化学能, 驱动生态系统碳循环(
赵风华和于贵瑞, 2018 ).光合过程各个步骤对外界环境的波动非常敏感, 根据植物光合特性能够快速了解其生理状态(
孙安安等, 2019 ).因此, 探究荒漠植物光合作用对环境因子的响应, 可以进一步明确其在逆境条件下的耐受力、恢复力和适应力(
Chaves et al ., 2004 ). ...
香蕉幼苗叶片响应低温胁迫的比较蛋白质组学研究
1
2015
... 荒漠生态系统具有干旱、高太阳辐射、高蒸腾和极端温度等环境特征(
Jia et al ., 2016 ).夏季荒漠的强烈太阳辐射远超植物光合所需, 已有研究证明荒漠植物通过调整热耗散等途径进行调节, 但实际上仍然会产生大量活性氧, 对植物光合机构造成极大威胁, 此时光呼吸过程的存在价值就显得尤为重要(
何炎红等, 2015 ).其次, 荒漠气候极端温度也会抑制卡尔文循环相关蛋白酶, 从而导致光合效率降低(
孙勇等, 2015 ).另外, 荒漠干燥空气导致的高蒸腾会促使植物气孔关闭, 影响了水的传输, 限制了光合过程中相关电子传输速率(
阮成江和李代琼, 2001 ).因此, 波动环境下荒漠植物光合过程能量分配的相对变化和内在调节机制可能具有明显特征. ...
香蕉幼苗叶片响应低温胁迫的比较蛋白质组学研究
1
2015
... 荒漠生态系统具有干旱、高太阳辐射、高蒸腾和极端温度等环境特征(
Jia et al ., 2016 ).夏季荒漠的强烈太阳辐射远超植物光合所需, 已有研究证明荒漠植物通过调整热耗散等途径进行调节, 但实际上仍然会产生大量活性氧, 对植物光合机构造成极大威胁, 此时光呼吸过程的存在价值就显得尤为重要(
何炎红等, 2015 ).其次, 荒漠气候极端温度也会抑制卡尔文循环相关蛋白酶, 从而导致光合效率降低(
孙勇等, 2015 ).另外, 荒漠干燥空气导致的高蒸腾会促使植物气孔关闭, 影响了水的传输, 限制了光合过程中相关电子传输速率(
阮成江和李代琼, 2001 ).因此, 波动环境下荒漠植物光合过程能量分配的相对变化和内在调节机制可能具有明显特征. ...
高温强光下水杨酸对黄瓜叶片叶绿素荧光和叶黄素循环的影响
1
2006
... 在荒漠地区高辐射、极端温度和干旱等多种恶劣气候条件下, 植物通过对光合过程能量分配的调节, 避免过剩光能对光合单元机构造成破坏.结果表明, 黑沙蒿光合过程能量分配的光化学组分和热耗散组分之间在生长季内表现出竞争关系(
表1 ).高辐射条件下, 叶片叶绿素分子产生的大量过剩激发能对光系统II反应中心造成极大的威胁, 植物将启动一系列能量耗散的权衡调节以维持光能吸收利用的平衡关系(
王燕凌等, 2015 ;
Ruban, 2016 ).
E 与
Φ A 和
Φ PR 极显著正相关, 说明黑沙蒿以提高蒸腾为代价增加其光合固碳能力, 并且过剩光能可以通过转移到光呼吸代谢途径进行消耗.研究发现光呼吸不仅与光合固碳、氮代谢、氨基酸形成等基础循环有关, 同时还参与了对环境逆境的抵抗(
姜闯道等, 2005 ).光呼吸虽然被看作光合作用的竞争过程, 但通过光呼吸可以清除大量有害代谢产物.另外光呼吸产生的CO
2 可以限制甘氨酸脱羧酶的活动范围, 从而提高核酮糖二磷酸羧化酶的羧化效率, 因此本研究中
Φ A 和
Φ PR 呈现出正相关的协同关系.
E 与
Φ NPQ 和
Φ NO 表现为负相关关系, 说明在蒸腾受限情况下, 黑沙蒿能够通过增加热耗散组分来耗散过剩光能.依赖于叶黄素循环的热耗散机制是保护植物光合单元机构免受灼伤的主要途径, 当过剩光能累积导致跨膜质子梯度增加时, 在脱环氧化酶的催化下, 紫黄质经由花药黄质后最终转化为玉米黄质, 同时产生大量的热量耗散过剩光能(
孙艳等, 2006 ). ...
高温强光下水杨酸对黄瓜叶片叶绿素荧光和叶黄素循环的影响
1
2006
... 在荒漠地区高辐射、极端温度和干旱等多种恶劣气候条件下, 植物通过对光合过程能量分配的调节, 避免过剩光能对光合单元机构造成破坏.结果表明, 黑沙蒿光合过程能量分配的光化学组分和热耗散组分之间在生长季内表现出竞争关系(
表1 ).高辐射条件下, 叶片叶绿素分子产生的大量过剩激发能对光系统II反应中心造成极大的威胁, 植物将启动一系列能量耗散的权衡调节以维持光能吸收利用的平衡关系(
王燕凌等, 2015 ;
Ruban, 2016 ).
E 与
Φ A 和
Φ PR 极显著正相关, 说明黑沙蒿以提高蒸腾为代价增加其光合固碳能力, 并且过剩光能可以通过转移到光呼吸代谢途径进行消耗.研究发现光呼吸不仅与光合固碳、氮代谢、氨基酸形成等基础循环有关, 同时还参与了对环境逆境的抵抗(
姜闯道等, 2005 ).光呼吸虽然被看作光合作用的竞争过程, 但通过光呼吸可以清除大量有害代谢产物.另外光呼吸产生的CO
2 可以限制甘氨酸脱羧酶的活动范围, 从而提高核酮糖二磷酸羧化酶的羧化效率, 因此本研究中
Φ A 和
Φ PR 呈现出正相关的协同关系.
E 与
Φ NPQ 和
Φ NO 表现为负相关关系, 说明在蒸腾受限情况下, 黑沙蒿能够通过增加热耗散组分来耗散过剩光能.依赖于叶黄素循环的热耗散机制是保护植物光合单元机构免受灼伤的主要途径, 当过剩光能累积导致跨膜质子梯度增加时, 在脱环氧化酶的催化下, 紫黄质经由花药黄质后最终转化为玉米黄质, 同时产生大量的热量耗散过剩光能(
孙艳等, 2006 ). ...
荒漠植物白麻气孔导度特征及其影响因子研究
1
2013
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
荒漠植物白麻气孔导度特征及其影响因子研究
1
2013
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
塔里木河下游刚毛柽柳光合作用、蒸腾作用及水分利用效率特性研究
1
2015
... 在荒漠地区高辐射、极端温度和干旱等多种恶劣气候条件下, 植物通过对光合过程能量分配的调节, 避免过剩光能对光合单元机构造成破坏.结果表明, 黑沙蒿光合过程能量分配的光化学组分和热耗散组分之间在生长季内表现出竞争关系(
表1 ).高辐射条件下, 叶片叶绿素分子产生的大量过剩激发能对光系统II反应中心造成极大的威胁, 植物将启动一系列能量耗散的权衡调节以维持光能吸收利用的平衡关系(
王燕凌等, 2015 ;
Ruban, 2016 ).
E 与
Φ A 和
Φ PR 极显著正相关, 说明黑沙蒿以提高蒸腾为代价增加其光合固碳能力, 并且过剩光能可以通过转移到光呼吸代谢途径进行消耗.研究发现光呼吸不仅与光合固碳、氮代谢、氨基酸形成等基础循环有关, 同时还参与了对环境逆境的抵抗(
姜闯道等, 2005 ).光呼吸虽然被看作光合作用的竞争过程, 但通过光呼吸可以清除大量有害代谢产物.另外光呼吸产生的CO
2 可以限制甘氨酸脱羧酶的活动范围, 从而提高核酮糖二磷酸羧化酶的羧化效率, 因此本研究中
Φ A 和
Φ PR 呈现出正相关的协同关系.
E 与
Φ NPQ 和
Φ NO 表现为负相关关系, 说明在蒸腾受限情况下, 黑沙蒿能够通过增加热耗散组分来耗散过剩光能.依赖于叶黄素循环的热耗散机制是保护植物光合单元机构免受灼伤的主要途径, 当过剩光能累积导致跨膜质子梯度增加时, 在脱环氧化酶的催化下, 紫黄质经由花药黄质后最终转化为玉米黄质, 同时产生大量的热量耗散过剩光能(
孙艳等, 2006 ). ...
塔里木河下游刚毛柽柳光合作用、蒸腾作用及水分利用效率特性研究
1
2015
... 在荒漠地区高辐射、极端温度和干旱等多种恶劣气候条件下, 植物通过对光合过程能量分配的调节, 避免过剩光能对光合单元机构造成破坏.结果表明, 黑沙蒿光合过程能量分配的光化学组分和热耗散组分之间在生长季内表现出竞争关系(
表1 ).高辐射条件下, 叶片叶绿素分子产生的大量过剩激发能对光系统II反应中心造成极大的威胁, 植物将启动一系列能量耗散的权衡调节以维持光能吸收利用的平衡关系(
王燕凌等, 2015 ;
Ruban, 2016 ).
E 与
Φ A 和
Φ PR 极显著正相关, 说明黑沙蒿以提高蒸腾为代价增加其光合固碳能力, 并且过剩光能可以通过转移到光呼吸代谢途径进行消耗.研究发现光呼吸不仅与光合固碳、氮代谢、氨基酸形成等基础循环有关, 同时还参与了对环境逆境的抵抗(
姜闯道等, 2005 ).光呼吸虽然被看作光合作用的竞争过程, 但通过光呼吸可以清除大量有害代谢产物.另外光呼吸产生的CO
2 可以限制甘氨酸脱羧酶的活动范围, 从而提高核酮糖二磷酸羧化酶的羧化效率, 因此本研究中
Φ A 和
Φ PR 呈现出正相关的协同关系.
E 与
Φ NPQ 和
Φ NO 表现为负相关关系, 说明在蒸腾受限情况下, 黑沙蒿能够通过增加热耗散组分来耗散过剩光能.依赖于叶黄素循环的热耗散机制是保护植物光合单元机构免受灼伤的主要途径, 当过剩光能累积导致跨膜质子梯度增加时, 在脱环氧化酶的催化下, 紫黄质经由花药黄质后最终转化为玉米黄质, 同时产生大量的热量耗散过剩光能(
孙艳等, 2006 ). ...
Photosynthetic gas-exchange and PSII photochemical acclimation to drought in a native and non-native xerophytic species (
Artemisia ordosica and
Salix psammophila )
1
2018
... 基于整个生长季所有测量数据, 对黑沙蒿光合能量分配各组分、环境参数和叶性状参数进行相关性分析, 显著性水平设为0.05, 极显著水平设为0.01.建立基于叶性状参数的黑沙蒿光合能量分配组分的多元线性回归模型, 使用拟合优度
R 2 和赤池信息量准则(AIC)对模型进行评价, 通常情况下AIC值为负, 且模型的拟合效果越好AIC值越低.根据以往相关研究结果, 以0.1 m
3 ·m
-3 作为
SWC 的阈值, 1.0 kPa作为
VPD 的阈值(
Wu et al ., 2018 ), 将数据分为4组, 进行黑沙蒿光合能量分配和
SLA 的重复测量方差分析的显著性分析.因可用数据点数量较少, 分析前进行方差齐性检验和正态分布检验, 对质量不符合要求的数据进行对数转换, 以达到方差齐性. ...
油蒿(
Artemisia ordosica )光化学量子效率和非光化学淬灭的动态及其影响因子
1
2015
... 环境对植物光合过程能量分配的影响存在时间尺度的差异, 短期响应发生在秒至分钟内, 如光系统II的叶黄素循环(
靳川等, 2021 ).长期影响则可以通过调整光合色素浓度、Chl a/b和
SLA 等叶性状参数或光系统II的特定蛋白等途径实现, 通常存在几天到几周内(
李鑫豪等, 2019 ).在生长季内, 黑沙蒿光合过程能量分配与各环境因子均有很强的相关性(
表1 ), 结果表明光化学反应组分(
Φ A 、
Φ PR )和
PAR 呈负相关关系, 这是由于
PAR 的升高产生了更多的过剩光能, 黑沙蒿会瞬时激发叶黄素循环等持续性热耗散, 将过剩光能转移到热耗散组分(
Φ NPQ 和
Φ NO ).光化学反应组分和
T a 负相关, 说明高温可能抑制了黑沙蒿光系统II光反应酶或捕光蛋白的活性, 导致光合过程电子传递受限.植物细胞的原生质层抗凝结特性对于增加细胞内渗透压意义重大(
Savitch et al ., 2009 ).同地区相关研究结果也证明黑沙蒿光系统II对温度极敏感, 在清晨的低温条件下也发生了相似的抑制现象(
吴雅娟等, 2015 ).研究认为黑沙蒿能够通过原生质层的抗凝结特性增加细胞内渗透压, 从而以减小代谢速率的方式抵御极端温度胁迫. ...
油蒿(
Artemisia ordosica )光化学量子效率和非光化学淬灭的动态及其影响因子
1
2015
... 环境对植物光合过程能量分配的影响存在时间尺度的差异, 短期响应发生在秒至分钟内, 如光系统II的叶黄素循环(
靳川等, 2021 ).长期影响则可以通过调整光合色素浓度、Chl a/b和
SLA 等叶性状参数或光系统II的特定蛋白等途径实现, 通常存在几天到几周内(
李鑫豪等, 2019 ).在生长季内, 黑沙蒿光合过程能量分配与各环境因子均有很强的相关性(
表1 ), 结果表明光化学反应组分(
Φ A 、
Φ PR )和
PAR 呈负相关关系, 这是由于
PAR 的升高产生了更多的过剩光能, 黑沙蒿会瞬时激发叶黄素循环等持续性热耗散, 将过剩光能转移到热耗散组分(
Φ NPQ 和
Φ NO ).光化学反应组分和
T a 负相关, 说明高温可能抑制了黑沙蒿光系统II光反应酶或捕光蛋白的活性, 导致光合过程电子传递受限.植物细胞的原生质层抗凝结特性对于增加细胞内渗透压意义重大(
Savitch et al ., 2009 ).同地区相关研究结果也证明黑沙蒿光系统II对温度极敏感, 在清晨的低温条件下也发生了相似的抑制现象(
吴雅娟等, 2015 ).研究认为黑沙蒿能够通过原生质层的抗凝结特性增加细胞内渗透压, 从而以减小代谢速率的方式抵御极端温度胁迫. ...
油蒿(
Artemisia ordosica )对半干旱区沙地生境的适应及其生态作用
2
2004
... 黑沙蒿(俗名油蒿,
Artemisia ordosica )属半灌木菊科蒿属, 广泛分布于我国北部及西北沙地, 具有生长快、耐干旱和抗风沙侵蚀等优点(
杨洪晓等, 2004 ).作为毛乌素沙地最为主要的固沙物种, 黑沙蒿在维护荒漠生态稳定方面发挥着重要作用.本研究运用叶绿素荧光和气体交换技术, 对宁夏盐池毛乌素沙地荒漠灌木优势种黑沙蒿进行长期原位监测, 从光合过程能量分配角度出发, 旨在揭示: (1)波动环境下黑沙蒿光合过程能量分配各组分对环境因子的响应; (2)波动环境下黑沙蒿光合过程能量分配各组分之间相对变化及其内在调节机制.以期为构建荒漠生态系统能量交换模型提供数据支撑, 丰富荒漠植物的生理可塑性理论. ...
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
油蒿(
Artemisia ordosica )对半干旱区沙地生境的适应及其生态作用
2
2004
... 黑沙蒿(俗名油蒿,
Artemisia ordosica )属半灌木菊科蒿属, 广泛分布于我国北部及西北沙地, 具有生长快、耐干旱和抗风沙侵蚀等优点(
杨洪晓等, 2004 ).作为毛乌素沙地最为主要的固沙物种, 黑沙蒿在维护荒漠生态稳定方面发挥着重要作用.本研究运用叶绿素荧光和气体交换技术, 对宁夏盐池毛乌素沙地荒漠灌木优势种黑沙蒿进行长期原位监测, 从光合过程能量分配角度出发, 旨在揭示: (1)波动环境下黑沙蒿光合过程能量分配各组分对环境因子的响应; (2)波动环境下黑沙蒿光合过程能量分配各组分之间相对变化及其内在调节机制.以期为构建荒漠生态系统能量交换模型提供数据支撑, 丰富荒漠植物的生理可塑性理论. ...
... 水分是限制荒漠植物生长发育的关键因素, 土壤干旱和空气干燥等情况在荒漠地区经常出现(
林年丰和汤洁, 2001 ;
王芸等, 2013 ;
罗丹丹等, 2019 ).黑沙蒿蒸腾作用在高
VPD 条件下被加强, 通过气孔关闭避免水分的过度流失.气孔导度限制卡尔文循环造成ATP消耗速率降低, 进而抑制ATP合酶活性(
Müller et al ., 2001 ).然而跨类囊体质子梯度受电子传递速率和ATP合酶活性的双向调节, ATP合酶活性的抑制会降低类囊体膜的质子导度, 进而不可避免地提高了跨类囊体质子梯度, 不宜于维持黑沙蒿光化学过程和暗反应中碳同化物的积累.另外, 过低的
SWC 难以维持植物的正常水分需求, 抑制了光反应过程中由水裂解释放的电子传输速率, 进而导致光合效率的减少.在水分亏缺条件下, 黑沙蒿通过增加
Φ PR 、
Φ NPQ 和
Φ NO 耗散不利条件下的过剩光能, 同时利用其发达的根系增加吸水能力等形态学优势抵御干旱(
杨洪晓等, 2004 ;
康博文等, 2010 ).与其他荒漠物种的研究结果比较, 高辐射对黑沙蒿光合过程能量分配的影响要小于对蒙古山竹子和北沙柳的影响(
靳川等, 2020 ).蒙古山竹子为互生阔线状叶片, 沙柳为互生披针形叶片, 它们对于光能的截获能力都高于黑沙蒿的针型叶片, 同时光合机构也更容易受到高辐射灼伤.黑沙蒿对极端温度的适应能力要优于红砂(
Reaumuria soongarica ), 这可能是因为黑沙蒿原生质层的高抗凝结能力(
种培芳等, 2010 ). ...
Diurnal response of effective quantum yield of PSII photochemistry to irradiance as an indicator of photosynthetic acclimation to stressed environments revealed in a xerophytic species
1
2017
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
棉花苞叶光呼吸和PSII热耗散对土壤水分的响应
1
2014
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
棉花苞叶光呼吸和PSII热耗散对土壤水分的响应
1
2014
... 光能被植物叶片吸收后, 通过以下3种途径进行耗散: 激发能在叶绿素分子间传递, 用于光化学反应; 激发态叶绿素分子回迁到基态, 激发能直接变成热能; 重新释放光子, 产生叶绿素荧光(
Murchie & Lawson, 2013 ).调制荧光技术已然成为探究植物光合的有效工具, 其应用逐渐延伸到植物生态学、农学、林学等多个领域(
Zha et al ., 2017 ).基于高等植物的“湖泊”模型, 可将吸收光能划分为调节性热耗散(
Φ NPQ )、非调节性热耗散(
Φ NO )和实际光化学效率(
Φ PSII )三部分(
Kramer et al ., 2004 ).植物通过热耗散可以有效地消散过剩光能, 减少活性氧生成, 避免光合机构灼伤(
Ruban, 2016 ).然而, 热耗散机制并不是消除过剩能量的唯一途径, 其中部分能量会用于光呼吸和氮代谢等过程(
侯学文等, 2019 ).随着研究深入, 越来越多报道指出光呼吸过程能够消耗活性氧, 构成植物主动抵抗各种逆境的光破坏防御机制(
姜闯道等, 2005 ).目前, 国内外研究对于光呼吸代谢途径等生化过程已相对明确, 但是对于植物光呼吸过程在自然状态下是如何受环境调控等问题的认识仍然有限(
张超等, 2014 ).受测量技术限制, 常规测量仪器难以在叶片上直接测量光呼吸, 导致目前几乎没有将热耗散和光呼吸相关联的研究.通过气体交换技术可以准确测量出植物的净光合速率(
P n )和暗呼吸速率(
R d ), 因此利用叶绿素荧光和气体交换的同步测量, 进而计算差值得出植物光呼吸速率(
R p ), 或将成为植物生态学的相关热点(
Galmés et al ., 2007 ). ...
陆地生态系统碳-水耦合机制初探
1
2008
... 干旱半干旱区是维持生态稳定的关键屏障, 也是我国植被恢复的重点区域(
陈亚宁等, 2019 ).近年来气候恶化以及极端天气事件的频发, 该区植被经常遭受多种环境胁迫, 处于急剧退化的风险当中(
Huang et al ., 2017 ).植物通过光合作用将光能转化为化学能, 驱动生态系统碳循环(
赵风华和于贵瑞, 2018 ).光合过程各个步骤对外界环境的波动非常敏感, 根据植物光合特性能够快速了解其生理状态(
孙安安等, 2019 ).因此, 探究荒漠植物光合作用对环境因子的响应, 可以进一步明确其在逆境条件下的耐受力、恢复力和适应力(
Chaves et al ., 2004 ). ...
陆地生态系统碳-水耦合机制初探
1
2008
... 干旱半干旱区是维持生态稳定的关键屏障, 也是我国植被恢复的重点区域(
陈亚宁等, 2019 ).近年来气候恶化以及极端天气事件的频发, 该区植被经常遭受多种环境胁迫, 处于急剧退化的风险当中(
Huang et al ., 2017 ).植物通过光合作用将光能转化为化学能, 驱动生态系统碳循环(
赵风华和于贵瑞, 2018 ).光合过程各个步骤对外界环境的波动非常敏感, 根据植物光合特性能够快速了解其生理状态(
孙安安等, 2019 ).因此, 探究荒漠植物光合作用对环境因子的响应, 可以进一步明确其在逆境条件下的耐受力、恢复力和适应力(
Chaves et al ., 2004 ). ...
 ,1,2,3,*
,1,2,3,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT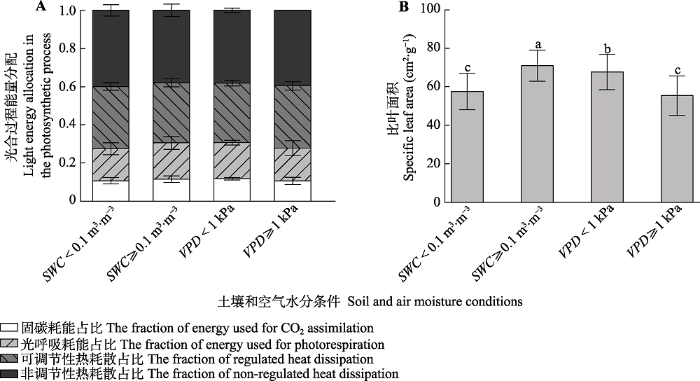 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}