National Natural Science Foundation of China(41771051) National Natural Science Foundation of China(41630750) National Key R&D Program of China(2018YFA0606101)
Abstract Aims The high-precision data measured by the Dendrometer includes not only the stem radial growth process caused by the enlargement of xylem cells but also the reversible changes caused by the consumption and replenishment of the stem water. Our objectives were to assess the difference in stem water relations of Picea meyeri obtained by different de-trending methods and their responses to water conditions in soil and air. Methods Hourly stem radius variations in P. meyeri and corresponding environmental factors were monitored in the lower limit of the coniferous forest on the northern slope of Luya Mountain, northern Shanxi Province, China. Gompertz growth model (GPZ), linear growth model (LG), and zero growth model (ZG), daily approach (D), and stem cycle approach (SC) were used to fit the stem growth trend in the growing season of 2015. Then, the growth trend and extract five different types of tree water deficit-induced stem shrinkage (TWDGPZ, TWDLG, TWDZG, TWDD, and TWDSC) were removed to characterize the dynamic of stem water relations. Moving window correlation (31 days) and ordinary least square regression were further employed to analyze the responses of different types of stem water relations to soil and air moisture conditions. Important findings The results showed that: 1) stem water relations derived from different de-trending methods had contrasting trends and amplitude, which could be clustered into three groups: TWDLG and TWDZG, TWDGPZ, and TWDD and TWDSC. Each month, significant correlations between stem water relations in the same group or a lower clustering distance showed, however, TWDLG, TWDZG, and TWDGPZ had weaker correlations with TWD and TWDSC in August. 2) TWDD and TWDSC had a closer and more stable positive relationship with vapor pressure deficit (VPD) than TWDGPZ, TWDLG, and TWDZG did. The responses of all types of stem water relations to soil water content (SWC) varied greatly during the growing season. 3) All stem water relations significantly increased as the water stress (VPD/SWC) intensified (p < 0.05). When the stress was low, TWDSC was most sensitive to the changes of VPD/SWC (R2 = 0.39, p < 0.001), whereas it was not much different from TWDZG (R2 = 0.37, p < 0.001); when the stress was high, TWDZG showed the greatest sensitivity to VPD/SWC (R2 = 0.59, p < 0.001). Our results suggested that the zero-growth model was more suitable to detrend the stem radius variations during the growing season, and provided a crucial reference for predicting the stem water status and dynamics, especially under drought stress. Keywords:Dendrometer;growth detrend;stem shrinkage;moving window correlation;drought
PDF (865KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 薛峰, 江源, 董满宇, 王明昌, 丁新原, 杨显基, 崔明皓, 康慕谊. 不同去趋势方法对基于Dendrometer数据的茎干水分动态分析的影响——以白扦为例. 植物生态学报, 2021, 45(8): 880-890. DOI: 10.17521/cjpe.2021.0025 XUE Feng, JIANG Yuan, DONG Man-Yu, WANG Ming-Chang, DING Xin-Yuan, YANG Xian-Ji, CUI Ming-Hao, KANG Mu-Yi. Influence of different de-trending methods on stem water relations of Picea meyeri derived from Dendrometer measurements. Chinese Journal of Plant Ecology, 2021, 45(8): 880-890. DOI: 10.17521/cjpe.2021.0025
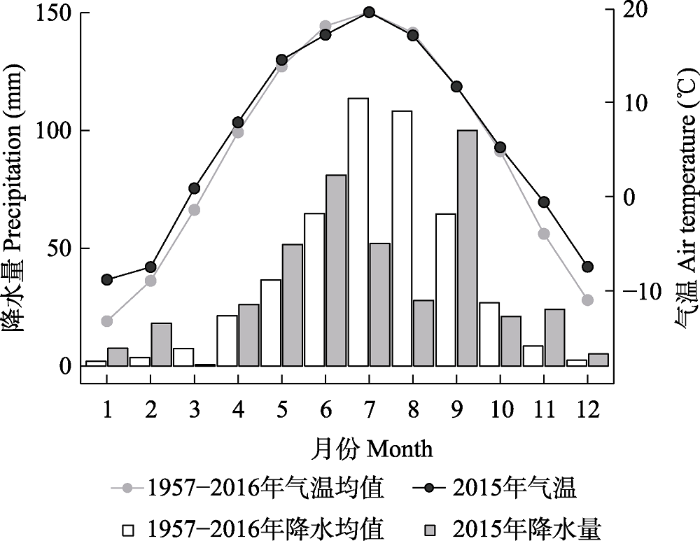
研究在山西省北部的芦芽山开展。气候记录(1957-2016年)显示该地区年平均气温5.3 ℃, 年降水量459.8 mm, 主要集中于夏季6-9月份之间, 占全年降水量的75%。该地区2015年的年平均气温为6.5 ℃, 高于多年气温平均值, 年降水量为464.3 mm, 稍高于多年降水量的平均值。然而, 当年降水的季节分布与多年的平均状况差异很大。其中, 2015年最大月降水量出现在9月份, 7和8月份的降水量(52.0和27.8 mm)远小于多年月降水量的平均值(113.7和108.2 mm)(图1)。研究点位于芦芽山北坡寒温性针叶林分布的下限林缘(38.75° N, 111.82° E), 海拔2 040 m, 主要建群种为白扦和华北落叶松。样地(10 m × 10 m)平均树高9 m, 林内郁闭度为0.9, 林下草本和灌木稀少。坡面缓平, 坡度为10°。土壤主要发育在残积和/或坡积母质上, 厚度约50 cm, 土壤类型为山地棕壤。
Fig. 1Comparison of the mean monthly air temperatures and the monthly precipitation for the year 2015 and the 60 a average (1957-2016) of Luya Mountain, Shanxi Province, China.
Fig. 2Hourly time series of stem radial variations (SRV) and growth process calculated by different detrending approaches for Picea meyeri of Luya Mountain, Shanxi Province, China in the growing season of 2015. DOY, day of the year.
Table 2 表2 表2山西芦芽山白扦不同去趋势的茎干水分亏缺(TWD)的特征参数 Table 2Main parameters of tree water deficit-induced stem shrinkage (TWD) calculated by different detrending methods of Picea meyeri on Luya Mountain, Shanxi Province, China
Fig. 3Tree water deficit-induced stem shrinkage (TWD) of Picea meyeri on Luya Mountain, Shanxi Province, China, calculated by different detrending approaches. D, daily approach; GPZ, Gompertz model; LG, linear growth model; SC, stem cycle approach; ZG, zero growth model. Dashed lines indicate the average. DOY, day of the year.
Fig. 4Cluster analysis of tree water deficit-induced stem shrinkage (TWD) of Picea meyeri on Luya Mountain, Shanxi Province, China, with different detrending methods. D, daily approach; GPZ, Gompertz model; LG, linear growth model; SC, stem cycle approach; ZG, zero growth model.
图5
新窗口打开|下载原图ZIP|生成PPT 图5山西芦芽山白扦不同去趋势茎干水分亏缺(TWD)在不同月份的相关系数矩阵。D, 日值法; GPZ, Gompertz生长模型; LG, 线性生长模型; SC, 茎干循环法; ZG, 零生长模型。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 5Correlation matrix of tree water deficit-induced stem shrinkage (TWD) relations of Picea meyeri on Luya Mountain, Shanxi Province, China, developed from different detrending approaches in different months. D, daily approach; GPZ, Gompertz model; LG, linear growth model; SC, stem cycle approach; ZG, zero growth model. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Fig. 6Moving window correlation (31 days) between tree water deficit-induced stem shrinkage (TWD) of Picea meyeri on Luya Mountain, Shanxi Province, China, and vapor pressure (VPD) and soil water content (SWC). D, daily approach; GPZ, Gompertz model; LG, linear growth model; SC, stem cycle approach; ZG, zero growth model. DOY, day of the year. Dotted lines indicate p < 0.05.
2.4 不同去趋势方法的茎干水分亏缺对干旱的敏感性
树木茎干的水分亏缺是蒸腾失水减少和土壤吸水补充动态平衡结果。我们计算空气饱和水汽压差和土壤含水量的比值(VPD/SWC)来表征样点环境中的水分胁迫程度, 其值反映了随着空气和土壤水分地减少, 茎干维持原来饱和状态的难度逐渐增大。整个生长季内, VPD/SWC在0-7.8 kPa·m3·m-3之间波动, 均值为2.7 kPa·m3·m-3。其中, 从8月4日(DOY 216)到9月3日(DOY 246)的时间段内其值都在生长季平均值之上, 持续长达30天(图7)。随着水分胁迫程度的升高, 不同去趋势方法得到的茎干水分波动都展示出显著的上升趋势(图8; p < 0.01)。当胁迫程度较低时(VPD/SWC ≤ 2.7 kPa·m3·m-3), TWDSC对VPD/SWC的变化最为敏感(R2 = 0.39, p < 0.001), TWDZG、TWDLG和TWDD次之(R2 = 0.37, 0.35和0.31; p < 0.001), TWDGPZ最弱(R2 = 0.11, p < 0.01)。而当胁迫程度较高时(VPD/SWC > 2.7 kPa·m3·m-3), TWDZG对VPD/SWC的变化敏感性最高, 解释率可达0.59。TWDLG和TWDGPZ对VPD/SWC的敏感性差别不大(R2 = 0.50; p < 0.001), 但是明显高于TWDD和TWDSC的(R2 = 0.10; p < 0.001)。
Fig. 7Seasonal variations of water stress factor at monitoring site on Luya Mountain, Shanxi Province, China. SWC, soil water content; VPD, vapor pressure deficit. DOY, day of the year. The dashed line represents the average.
Fig. 8Sensitivity of tree water dificit-induced stem shrinkage (TWD) relations of Picea meyeri on Luya Mountain, Shanxi Province, China, with different de-trending methods to water stress factors. D, daily approach; GPZ, Gompertz model; LG, linear growth model; SC, stem cycle approach; ZG, zero growth model. SWC, soil water content; VPD, vapor pressure deficit. The dashed line represents the average of VPD/SWC = 2.7 kPa·m3·m-3.
3 讨论
3.1 不同去趋势方法对茎干水分亏缺特征的影响
在从高精度茎干变化数据中提取茎干水分信息的过程中, 不同去趋势方法对计算得到的茎干水分亏缺特征有一定的影响。这主要是因为不同方法模拟的年内茎干生长过程不同, 并且时间尺度也有所差别。聚类分析也将不同的茎干水分亏缺序列划分为3组: TWDGPZ, TWDLG和TWDZG, TWDD和TWDSC。TWDGPZ是Gompertz函数曲线拟合去趋势的结果。高拟合度的Gompertz生长模型被认为更符合木质部细胞的实际生长过程(董满宇等, 2012; Zhang et al., 2019), 而去除拟合生长趋势接近于茎干半径变化数据标准化的过程(Oberhuber, 2017; van der Maaten et al., 2018), 因此, TWDGPZ的平均值最小。线性生长模型和零生长模型以及日值法和茎干循环法根据茎干收缩和膨胀变化来模拟生长过程, 其构建机理是形成层细胞活动和茎干水势以及细胞膨压的紧密关联(Zweifel et al., 2005; Zweifel, 2016)。虽然很难量化不同时间尺度内木质部细胞在茎干收缩期的真实生理活动, 但是长时间尺度去趋势的TWDLG和TWDZG与短时间尺度的TWDD和TWDSC之间的波动特征展现出很大差别。TWDLG和TWDZG能够很好地捕捉到一段时间内树木茎干水分亏缺, 而TWDD和TWDSC主要反映短时间内的茎干水分流失(Sallo et al., 2017)。本研究的结果也显示, TWDLG和TWDZG的平均值、最大值以及波动幅度都远大于TWDD和TWDSC。
CampbellGS, NormanJM (1998). An Introduction to Environmental Biophysics Springer,New York. [本文引用: 1]
De SwaefT, De SchepperV, VandegehuchteMW, SteppeK (2015). Stem diameter variations as a versatile research tool in ecophysiology Tree Physiology, 35, 1047-1061. [本文引用: 1]
DeslauriersA, MorinH, UrbinatiC, CarrerM (2003). Daily weather response of balsam fir (Abies balsamea(L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada) Trees, 17, 477-484. [本文引用: 1]
DeslauriersA, RossiS, AnfodilloT (2007). Dendrometer and intra-annual tree growth: What kind of information can be inferred? Dendrochronologia, 25, 113-124. [本文引用: 3]
DeslauriersA, RossiS, TurcotteA, MorinH, KrauseC (2011). A three-step procedure in SAS to analyze the time series from automatic dendrometers Dendrochronologia, 29, 151-161. [本文引用: 1]
DevineWD, HarringtonCA (2011). Factors affecting diurnal stem contraction in young Douglas-fir Agricultural and Forest Meteorology, 151, 414-419. [本文引用: 1]
DongMY, JiangY, YangHC, WangMC, ZhangWT, GuoYY (2012). Dynamics of stem radial growth of Picea meyeri during the growing season at the treeline of Luya Mountain, China Chinese Journal of Plant Ecology, 36, 956-964. [本文引用: 3]
DongMY, WangBQ, JiangY, DingXY (2019). Environmental controls of diurnal and seasonal variations in the stem radius of Platycladus orientalis in Northern China Forests, 10, 784. DOI: 10.3390/f10090784. [本文引用: 1]
FernándezJE, CuevasMV (2010). Irrigation scheduling from stem diameter variations: a review Agricultural and Forest Meteorology, 150, 135-151. [本文引用: 1]
GaoJN, YangB, HeMH, ShishovV (2019). Intra-annual stem radial increment patterns of Chinese pine, Helan Mountains, Northern Central China Trees, 33, 751-763. [本文引用: 2]
JiQW, ZhengCY, ZhangL, ZengFX (2020). Stem radial growth dynamics of Pinus sylvestris var. mongolica and their relationship with meteorological factor in Saihanba, Hebei, China Chinese Journal of Plant Ecology, 44, 257-265. [本文引用: 1]
KingG, FontiP, NievergeltD, BüntgenU, FrankD (2013). Climatic drivers of hourly to yearly tree radius variations along a 6 °C natural warming gradient Agricultural and Forest Meteorology, 168, 36-46. [本文引用: 3]
LiXH, LiuRP, MaoZJ, SongY, LiuLX, SunT (2014). Dailystem radial variation of Pinus koraiensis and its responseto meteorological parameters in Xiaoxing’an Mountain Acta Ecologica Sinica, 34, 1635-1644. [本文引用: 2]
LiuXS, NieYQ, QiuXW, WenF, ZhangL, LuoTX (2015). A review on tree growth and its water use in cold environments based on Dendrometer World Forestry Research, 28, 19-25. [本文引用: 1]
MaJ, GuoJB, WangYH, LiuZB, GaoD, HongL, ZhangZY (2020). Variations in stem radii of Larix principis- rupprechtii to environmental factors at two slope locations in the Liupan Mountains, northwest China Journal of Forestry Research, 32, 513-527. [本文引用: 1]
MelvinTM, BriffaKR (2008). A “signal-free” approach to dendroclimatic standardisation Dendrochronologia, 26, 71-86. [本文引用: 1]
NiuHG, ZhangF, YuAL, WangF, ZhangJZ, GouXH (2018). Intra-annual stem radial growth dynamics of Picea wilsorii in response to climate in the eastern Qilian Mountains Acta Ecologica Sinica, 38, 7412-7420. [本文引用: 2]
OberhuberW (2017). Soil water availability and evaporative demand affect seasonal growth dynamics and use of stored water in co-occurring saplings and mature conifers under drought Trees, 31, 467-478. [本文引用: 1]
OberhuberW, HammerleA, KoflerW (2015). Tree water status and growth of saplings and mature Norway spruce (Picea abies) at a dry distribution limit Frontiers in Plant Science, 6, 703. DOI: 10.3389/fpls.2015.00703. [本文引用: 1]
OkiT, KanaeS (2006). Global hydrological cycles and world water resources Science, 313, 1068-1072. [本文引用: 1]
SalloFdS, SanchesL, de Morais DiasVR, PalaciosRdS, NogueiraJdS (2017). Stem water storage dynamics of Vochysia divergens in a seasonally flooded environment Agricultural and Forest Meteorology, 232, 566-575. [本文引用: 4]
SchlesingerWH, JasechkoS (2014). Transpiration in the global water cycle Agricultural and Forest Meteorology, 189- 190, 115-117. [本文引用: 1]
SunSJ, MengP, ZhangJS, JiaCR, RenYF (2012). Diurnal variation of Quercus variabilis trunk diameter in response to environmental factors at south aspect of Taihang Mountains Chinese Journal of Applied Ecology, 23, 2141-2148. [本文引用: 3]
TianQY, HeZB, XiaoSC, PengXM, DingAJ, LinPF (2017). Response of stem radial growth of Qinghai spruce (Picea crassifolia) to environmental factors in the Qilian Mountains of China Dendrochronologia, 44, 76-83. [本文引用: 3]
TurcotteA, RossiS, DeslauriersA, KrauseC, MorinH (2011). Dynamics of depletion and replenishment of water storage in stem and roots of black spruce measured by Dendrometers Frontiers in Plant Science, 2, 21. DOI: 10.3389/fpls.2011.00021. [本文引用: 1]
van der MaatenE, PapeJ, van der Maaten-TheunissenM, ScharnweberT, SmiljanićM, Cruz-GarcíaR, WilmkingM (2018). Distinct growth phenology but similar daily stem dynamics in three co-occurring broadleaved tree species Tree Physiology, 38, 1820-1828. [本文引用: 2]
WangZY, YangB, DeslauriersA, BräuningA (2015). Intra- annual stem radial increment response of Qilian juniper to temperature and precipitation along an altitudinal gradient in northwestern China Trees, 29, 25-34. [本文引用: 1]
ZhangYP, JiangY, WenY, DingXY, WangB, XuJL (2019). Comparing primary and secondary growth of co-occurring deciduous and evergreen conifers in an alpine habitat Forests, 10, 574. DOI: 10.3390/f10070574. [本文引用: 1]
ZhaoYD, GaoC, ZhangX, CaiX (2016). Review of real-time detecting methods of water stress for plants Transactions of the Chinese Society for Agricultural Machiner, 47, 290-300. [本文引用: 1]
ZweifelR (2016). Radial stem variations—A source of tree physiological information not fully exploited yet Plant, Cell, & Environment, 39, 231-232. [本文引用: 5]
ZweifelR, HaeniM, BuchmannN, EugsterW (2016). Are trees able to grow in periods of stem shrinkage? New Phytologist, 211, 839-849.
ZweifelR, ZimmermannL, NewberyDM (2005). Modeling tree water deficit from microclimate: an approach to quantifying drought stress Tree Physiology, 25, 147-156. [本文引用: 3]
An Introduction to Environmental Biophysics 1 1998
Stem diameter variations as a versatile research tool in ecophysiology 1 2015
... 树木茎干半径变化记录仪(Dendrometer)因其可以无伤害、自动和连续监测的特点而被广泛应用于树木的径向变化研究.研究人员基于茎干变化监测数据用Gompertz方程模拟了我国北方主要的针叶树——油松(Pinus tabuliformis)(Gao et al., 2019), 华北落叶松(Larix gmelinii var. principis-rupprechtii)(Ma et al., 2020), 青海云杉(Picea crassifolia)(Tian et al., 2017), 白扦(Picea meyeri)(董满宇等, 2012), 青扦(Picea wilsonii)(牛豪阁等, 2018)和红松(Pinus koraiensis)(李兴欢等, 2014)的季节生长过程并以此确定了各树种径向生长的物候期.依据茎干半径昼夜波动规律, 以往的研究也会使用日值法(Deslauriers et al., 2003; Gao et al., 2019)或茎干循环法(Deslauriers et al., 2007)提取茎干生长信息, 并以此分析短时间尺度下径向生长特征.进一步的研究也表明, 日值法和茎干循环法获得的日茎干生长的时间序列高度相关(Deslauriers et al., 2007).高精度的茎干变化数据不仅包含形成层细胞分裂和扩大导致的不可逆变化, 还包含由茎干水分的消耗和补充引起的可逆变化(Zweifel, 2016).有研究甚至认为茎干变化可以比液流、茎干水势和蒸腾速率等更为敏感有效地指示植物水分动态(赵燕东等, 2016).为了将茎干水分信息从茎干变化中剥离出来, 研究人员采用多种去生长趋势方法.van der Maaten等(2018)通过函数拟合去趋势的方法发现, 德国3种阔叶树的茎干水分波动非常相似并且都主要受到空气湿度、饱和水汽压差和降水的影响.Zweifel等(2016)根据茎干在收缩期线性生长或不生长的假设, 计算并提出以茎干的收缩量代表树木水分缺失, 而近年来的观测表明, 其能够较好地追踪叶水势和茎干水势等的变化(De Swaef et al., 2015).另外, 茎干日最大收缩量, 茎干收缩和恢复速率等也常被用来表征茎干水分的动态(Sallo et al., 2017; Tian et al., 2017).孙守家等(2012)对太行山南麓栓皮栎(Quercus variabilis)的研究发现, 茎干日最大收缩值与液流通量和叶片水势的变化显著相关, 并且主要受到季节性干旱和雨季土壤含水量的影响.然而, 迄今为止, 我们对不同去趋势方法获得的茎干水分波动序列之间的关系以及它们对环境因子敏感度的差异性仍然知之甚少. ...
Daily weather response of balsam fir (Abies balsamea(L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada) 1 2003
... 树木茎干半径变化记录仪(Dendrometer)因其可以无伤害、自动和连续监测的特点而被广泛应用于树木的径向变化研究.研究人员基于茎干变化监测数据用Gompertz方程模拟了我国北方主要的针叶树——油松(Pinus tabuliformis)(Gao et al., 2019), 华北落叶松(Larix gmelinii var. principis-rupprechtii)(Ma et al., 2020), 青海云杉(Picea crassifolia)(Tian et al., 2017), 白扦(Picea meyeri)(董满宇等, 2012), 青扦(Picea wilsonii)(牛豪阁等, 2018)和红松(Pinus koraiensis)(李兴欢等, 2014)的季节生长过程并以此确定了各树种径向生长的物候期.依据茎干半径昼夜波动规律, 以往的研究也会使用日值法(Deslauriers et al., 2003; Gao et al., 2019)或茎干循环法(Deslauriers et al., 2007)提取茎干生长信息, 并以此分析短时间尺度下径向生长特征.进一步的研究也表明, 日值法和茎干循环法获得的日茎干生长的时间序列高度相关(Deslauriers et al., 2007).高精度的茎干变化数据不仅包含形成层细胞分裂和扩大导致的不可逆变化, 还包含由茎干水分的消耗和补充引起的可逆变化(Zweifel, 2016).有研究甚至认为茎干变化可以比液流、茎干水势和蒸腾速率等更为敏感有效地指示植物水分动态(赵燕东等, 2016).为了将茎干水分信息从茎干变化中剥离出来, 研究人员采用多种去生长趋势方法.van der Maaten等(2018)通过函数拟合去趋势的方法发现, 德国3种阔叶树的茎干水分波动非常相似并且都主要受到空气湿度、饱和水汽压差和降水的影响.Zweifel等(2016)根据茎干在收缩期线性生长或不生长的假设, 计算并提出以茎干的收缩量代表树木水分缺失, 而近年来的观测表明, 其能够较好地追踪叶水势和茎干水势等的变化(De Swaef et al., 2015).另外, 茎干日最大收缩量, 茎干收缩和恢复速率等也常被用来表征茎干水分的动态(Sallo et al., 2017; Tian et al., 2017).孙守家等(2012)对太行山南麓栓皮栎(Quercus variabilis)的研究发现, 茎干日最大收缩值与液流通量和叶片水势的变化显著相关, 并且主要受到季节性干旱和雨季土壤含水量的影响.然而, 迄今为止, 我们对不同去趋势方法获得的茎干水分波动序列之间的关系以及它们对环境因子敏感度的差异性仍然知之甚少. ...
Dendrometer and intra-annual tree growth: What kind of information can be inferred? 3 2007
... 树木茎干半径变化记录仪(Dendrometer)因其可以无伤害、自动和连续监测的特点而被广泛应用于树木的径向变化研究.研究人员基于茎干变化监测数据用Gompertz方程模拟了我国北方主要的针叶树——油松(Pinus tabuliformis)(Gao et al., 2019), 华北落叶松(Larix gmelinii var. principis-rupprechtii)(Ma et al., 2020), 青海云杉(Picea crassifolia)(Tian et al., 2017), 白扦(Picea meyeri)(董满宇等, 2012), 青扦(Picea wilsonii)(牛豪阁等, 2018)和红松(Pinus koraiensis)(李兴欢等, 2014)的季节生长过程并以此确定了各树种径向生长的物候期.依据茎干半径昼夜波动规律, 以往的研究也会使用日值法(Deslauriers et al., 2003; Gao et al., 2019)或茎干循环法(Deslauriers et al., 2007)提取茎干生长信息, 并以此分析短时间尺度下径向生长特征.进一步的研究也表明, 日值法和茎干循环法获得的日茎干生长的时间序列高度相关(Deslauriers et al., 2007).高精度的茎干变化数据不仅包含形成层细胞分裂和扩大导致的不可逆变化, 还包含由茎干水分的消耗和补充引起的可逆变化(Zweifel, 2016).有研究甚至认为茎干变化可以比液流、茎干水势和蒸腾速率等更为敏感有效地指示植物水分动态(赵燕东等, 2016).为了将茎干水分信息从茎干变化中剥离出来, 研究人员采用多种去生长趋势方法.van der Maaten等(2018)通过函数拟合去趋势的方法发现, 德国3种阔叶树的茎干水分波动非常相似并且都主要受到空气湿度、饱和水汽压差和降水的影响.Zweifel等(2016)根据茎干在收缩期线性生长或不生长的假设, 计算并提出以茎干的收缩量代表树木水分缺失, 而近年来的观测表明, 其能够较好地追踪叶水势和茎干水势等的变化(De Swaef et al., 2015).另外, 茎干日最大收缩量, 茎干收缩和恢复速率等也常被用来表征茎干水分的动态(Sallo et al., 2017; Tian et al., 2017).孙守家等(2012)对太行山南麓栓皮栎(Quercus variabilis)的研究发现, 茎干日最大收缩值与液流通量和叶片水势的变化显著相关, 并且主要受到季节性干旱和雨季土壤含水量的影响.然而, 迄今为止, 我们对不同去趋势方法获得的茎干水分波动序列之间的关系以及它们对环境因子敏感度的差异性仍然知之甚少. ...
... )提取茎干生长信息, 并以此分析短时间尺度下径向生长特征.进一步的研究也表明, 日值法和茎干循环法获得的日茎干生长的时间序列高度相关(Deslauriers et al., 2007).高精度的茎干变化数据不仅包含形成层细胞分裂和扩大导致的不可逆变化, 还包含由茎干水分的消耗和补充引起的可逆变化(Zweifel, 2016).有研究甚至认为茎干变化可以比液流、茎干水势和蒸腾速率等更为敏感有效地指示植物水分动态(赵燕东等, 2016).为了将茎干水分信息从茎干变化中剥离出来, 研究人员采用多种去生长趋势方法.van der Maaten等(2018)通过函数拟合去趋势的方法发现, 德国3种阔叶树的茎干水分波动非常相似并且都主要受到空气湿度、饱和水汽压差和降水的影响.Zweifel等(2016)根据茎干在收缩期线性生长或不生长的假设, 计算并提出以茎干的收缩量代表树木水分缺失, 而近年来的观测表明, 其能够较好地追踪叶水势和茎干水势等的变化(De Swaef et al., 2015).另外, 茎干日最大收缩量, 茎干收缩和恢复速率等也常被用来表征茎干水分的动态(Sallo et al., 2017; Tian et al., 2017).孙守家等(2012)对太行山南麓栓皮栎(Quercus variabilis)的研究发现, 茎干日最大收缩值与液流通量和叶片水势的变化显著相关, 并且主要受到季节性干旱和雨季土壤含水量的影响.然而, 迄今为止, 我们对不同去趋势方法获得的茎干水分波动序列之间的关系以及它们对环境因子敏感度的差异性仍然知之甚少. ...
... (4)日值法(D).日值法通过计算每天的最大值、最小值或平均值来分析径向变化特征.研究表明3种方法提取的茎干生长信息高度相关(Deslauriers et al., 2007), 因此本研究只采用了日最大值法(图2C), 即从茎干变化记录仪所获取的每日24个原始数据中提取最大值, 计算连续两日的最大值之差.当差值大于0时得到树木茎干的日生长值, 当差值小于0时日生长计为0. ...
A three-step procedure in SAS to analyze the time series from automatic dendrometers 1 2011
Variations in stem radii of Larix principis- rupprechtii to environmental factors at two slope locations in the Liupan Mountains, northwest China 1 2020
... 树木茎干半径变化记录仪(Dendrometer)因其可以无伤害、自动和连续监测的特点而被广泛应用于树木的径向变化研究.研究人员基于茎干变化监测数据用Gompertz方程模拟了我国北方主要的针叶树——油松(Pinus tabuliformis)(Gao et al., 2019), 华北落叶松(Larix gmelinii var. principis-rupprechtii)(Ma et al., 2020), 青海云杉(Picea crassifolia)(Tian et al., 2017), 白扦(Picea meyeri)(董满宇等, 2012), 青扦(Picea wilsonii)(牛豪阁等, 2018)和红松(Pinus koraiensis)(李兴欢等, 2014)的季节生长过程并以此确定了各树种径向生长的物候期.依据茎干半径昼夜波动规律, 以往的研究也会使用日值法(Deslauriers et al., 2003; Gao et al., 2019)或茎干循环法(Deslauriers et al., 2007)提取茎干生长信息, 并以此分析短时间尺度下径向生长特征.进一步的研究也表明, 日值法和茎干循环法获得的日茎干生长的时间序列高度相关(Deslauriers et al., 2007).高精度的茎干变化数据不仅包含形成层细胞分裂和扩大导致的不可逆变化, 还包含由茎干水分的消耗和补充引起的可逆变化(Zweifel, 2016).有研究甚至认为茎干变化可以比液流、茎干水势和蒸腾速率等更为敏感有效地指示植物水分动态(赵燕东等, 2016).为了将茎干水分信息从茎干变化中剥离出来, 研究人员采用多种去生长趋势方法.van der Maaten等(2018)通过函数拟合去趋势的方法发现, 德国3种阔叶树的茎干水分波动非常相似并且都主要受到空气湿度、饱和水汽压差和降水的影响.Zweifel等(2016)根据茎干在收缩期线性生长或不生长的假设, 计算并提出以茎干的收缩量代表树木水分缺失, 而近年来的观测表明, 其能够较好地追踪叶水势和茎干水势等的变化(De Swaef et al., 2015).另外, 茎干日最大收缩量, 茎干收缩和恢复速率等也常被用来表征茎干水分的动态(Sallo et al., 2017; Tian et al., 2017).孙守家等(2012)对太行山南麓栓皮栎(Quercus variabilis)的研究发现, 茎干日最大收缩值与液流通量和叶片水势的变化显著相关, 并且主要受到季节性干旱和雨季土壤含水量的影响.然而, 迄今为止, 我们对不同去趋势方法获得的茎干水分波动序列之间的关系以及它们对环境因子敏感度的差异性仍然知之甚少. ...
A “signal-free” approach to dendroclimatic standardisation 1 2008
Soil water availability and evaporative demand affect seasonal growth dynamics and use of stored water in co-occurring saplings and mature conifers under drought 1 2017
... 在从高精度茎干变化数据中提取茎干水分信息的过程中, 不同去趋势方法对计算得到的茎干水分亏缺特征有一定的影响.这主要是因为不同方法模拟的年内茎干生长过程不同, 并且时间尺度也有所差别.聚类分析也将不同的茎干水分亏缺序列划分为3组: TWDGPZ, TWDLG和TWDZG, TWDD和TWDSC.TWDGPZ是Gompertz函数曲线拟合去趋势的结果.高拟合度的Gompertz生长模型被认为更符合木质部细胞的实际生长过程(董满宇等, 2012; Zhang et al., 2019), 而去除拟合生长趋势接近于茎干半径变化数据标准化的过程(Oberhuber, 2017; van der Maaten et al., 2018), 因此, TWDGPZ的平均值最小.线性生长模型和零生长模型以及日值法和茎干循环法根据茎干收缩和膨胀变化来模拟生长过程, 其构建机理是形成层细胞活动和茎干水势以及细胞膨压的紧密关联(Zweifel et al., 2005; Zweifel, 2016).虽然很难量化不同时间尺度内木质部细胞在茎干收缩期的真实生理活动, 但是长时间尺度去趋势的TWDLG和TWDZG与短时间尺度的TWDD和TWDSC之间的波动特征展现出很大差别.TWDLG和TWDZG能够很好地捕捉到一段时间内树木茎干水分亏缺, 而TWDD和TWDSC主要反映短时间内的茎干水分流失(Sallo et al., 2017).本研究的结果也显示, TWDLG和TWDZG的平均值、最大值以及波动幅度都远大于TWDD和TWDSC. ...
Tree water status and growth of saplings and mature Norway spruce (Picea abies) at a dry distribution limit 1 2015
... 不同去趋势方法的茎干水分亏缺与饱和水汽压差有着相对稳定的正相关关系, 而与土壤含水量的关系则变化很大.并且, 短时间尺度的茎干水分亏缺(TWDD和TWDSC)与饱和水汽压差有更好更稳定的相关性, 而与土壤含水量关系不大.这一结果与其他树种不同时间尺度上茎干水分波动的研究 (King et al., 2013; Oberhuber et al., 2015; Tian et al., 2017)一致.短时间内茎干水分动态主要受到蒸腾作用的驱动(Fernández & Cuevas, 2010; 季倩雯等, 2020).随着饱和水汽压差的增大, 叶片和空气之间水势差升高, 茎干储水的加速流失导致茎干收缩量变大(孙守家等, 2012; Dong et al., 2019).这也从侧面说明了茎干储水对异常蒸腾需求的缓冲作用.尽管土壤水分是茎干水分的重要来源, 但是大多数研究也都发现土壤含水量对短时间内的茎干变化影响很弱(Devine & Harrington, 2011; King et al., 2013).与我们的观察相似, 这主要是因为土壤水分昼夜变化小, 而且其在生长季内不断下降的趋势不符合茎干水分高频率波动.相比之下, TWDGPZ、TWDLG和TWDZG与饱和水汽压的关系较不稳定, 其相关性在8月份的干旱期未达到显著水平, 而与土壤含水量显著负相关.在土壤-植被-空气的连续体中, 随着土壤含水量的降低, 植株水分无法得到充足补充, 茎干储水量不断减少.在长时间持续的干旱期, 当土壤和茎干水分损耗到一定程度时, 储水组织间水分的流动阻力大于蒸腾拉力, 茎干水分波动将和空气水分状况解耦(Sallo et al., 2017). ...
Global hydrological cycles and world water resources 1 2006
Distinct growth phenology but similar daily stem dynamics in three co-occurring broadleaved tree species 2 2018
... 树木茎干半径变化记录仪(Dendrometer)因其可以无伤害、自动和连续监测的特点而被广泛应用于树木的径向变化研究.研究人员基于茎干变化监测数据用Gompertz方程模拟了我国北方主要的针叶树——油松(Pinus tabuliformis)(Gao et al., 2019), 华北落叶松(Larix gmelinii var. principis-rupprechtii)(Ma et al., 2020), 青海云杉(Picea crassifolia)(Tian et al., 2017), 白扦(Picea meyeri)(董满宇等, 2012), 青扦(Picea wilsonii)(牛豪阁等, 2018)和红松(Pinus koraiensis)(李兴欢等, 2014)的季节生长过程并以此确定了各树种径向生长的物候期.依据茎干半径昼夜波动规律, 以往的研究也会使用日值法(Deslauriers et al., 2003; Gao et al., 2019)或茎干循环法(Deslauriers et al., 2007)提取茎干生长信息, 并以此分析短时间尺度下径向生长特征.进一步的研究也表明, 日值法和茎干循环法获得的日茎干生长的时间序列高度相关(Deslauriers et al., 2007).高精度的茎干变化数据不仅包含形成层细胞分裂和扩大导致的不可逆变化, 还包含由茎干水分的消耗和补充引起的可逆变化(Zweifel, 2016).有研究甚至认为茎干变化可以比液流、茎干水势和蒸腾速率等更为敏感有效地指示植物水分动态(赵燕东等, 2016).为了将茎干水分信息从茎干变化中剥离出来, 研究人员采用多种去生长趋势方法.van der Maaten等(2018)通过函数拟合去趋势的方法发现, 德国3种阔叶树的茎干水分波动非常相似并且都主要受到空气湿度、饱和水汽压差和降水的影响.Zweifel等(2016)根据茎干在收缩期线性生长或不生长的假设, 计算并提出以茎干的收缩量代表树木水分缺失, 而近年来的观测表明, 其能够较好地追踪叶水势和茎干水势等的变化(De Swaef et al., 2015).另外, 茎干日最大收缩量, 茎干收缩和恢复速率等也常被用来表征茎干水分的动态(Sallo et al., 2017; Tian et al., 2017).孙守家等(2012)对太行山南麓栓皮栎(Quercus variabilis)的研究发现, 茎干日最大收缩值与液流通量和叶片水势的变化显著相关, 并且主要受到季节性干旱和雨季土壤含水量的影响.然而, 迄今为止, 我们对不同去趋势方法获得的茎干水分波动序列之间的关系以及它们对环境因子敏感度的差异性仍然知之甚少. ...
... 在从高精度茎干变化数据中提取茎干水分信息的过程中, 不同去趋势方法对计算得到的茎干水分亏缺特征有一定的影响.这主要是因为不同方法模拟的年内茎干生长过程不同, 并且时间尺度也有所差别.聚类分析也将不同的茎干水分亏缺序列划分为3组: TWDGPZ, TWDLG和TWDZG, TWDD和TWDSC.TWDGPZ是Gompertz函数曲线拟合去趋势的结果.高拟合度的Gompertz生长模型被认为更符合木质部细胞的实际生长过程(董满宇等, 2012; Zhang et al., 2019), 而去除拟合生长趋势接近于茎干半径变化数据标准化的过程(Oberhuber, 2017; van der Maaten et al., 2018), 因此, TWDGPZ的平均值最小.线性生长模型和零生长模型以及日值法和茎干循环法根据茎干收缩和膨胀变化来模拟生长过程, 其构建机理是形成层细胞活动和茎干水势以及细胞膨压的紧密关联(Zweifel et al., 2005; Zweifel, 2016).虽然很难量化不同时间尺度内木质部细胞在茎干收缩期的真实生理活动, 但是长时间尺度去趋势的TWDLG和TWDZG与短时间尺度的TWDD和TWDSC之间的波动特征展现出很大差别.TWDLG和TWDZG能够很好地捕捉到一段时间内树木茎干水分亏缺, 而TWDD和TWDSC主要反映短时间内的茎干水分流失(Sallo et al., 2017).本研究的结果也显示, TWDLG和TWDZG的平均值、最大值以及波动幅度都远大于TWDD和TWDSC. ...
Intra- annual stem radial increment response of Qilian juniper to temperature and precipitation along an altitudinal gradient in northwestern China 1 2015
 ,, 江源
,, 江源 ,*, 董满宇, 王明昌, 丁新原, 杨显基, 崔明皓, 康慕谊北京师范大学地理科学学部, 北京 100875
,*, 董满宇, 王明昌, 丁新原, 杨显基, 崔明皓, 康慕谊北京师范大学地理科学学部, 北京 100875

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT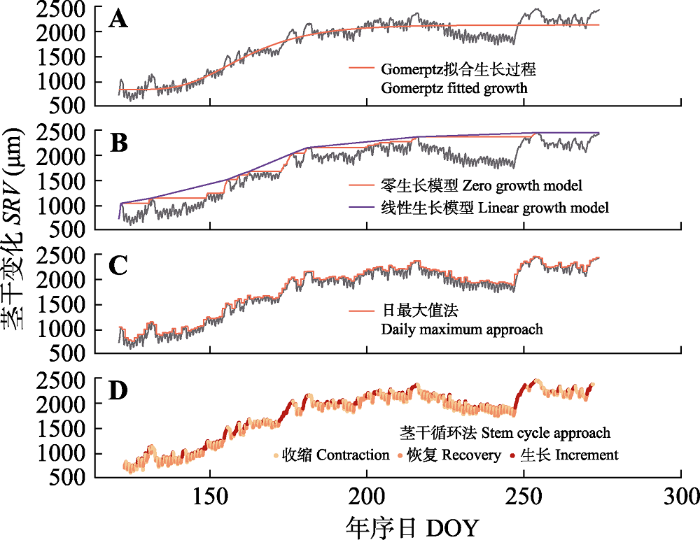 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT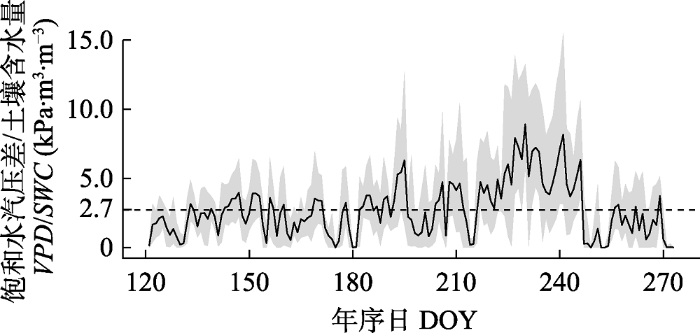 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT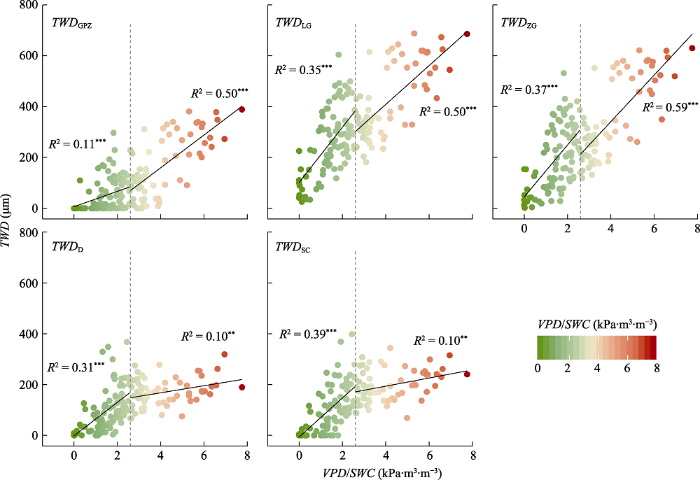 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}