Foliar condensate absorption and its pathways of two typical shrub species in the Mu Us Desert
Zi-Yang GUI1, Shu-Gao QIN1,2, Zhao HU1, Feng BAI1, Hui-Shu SHI3, Yu-Qing ZHANG,1,2,*1Yanchi Research Station, School of Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China 2Key Laboratory of National Forestry and Grassland Administration on Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China 3Ningxia Habahu National Nature Reserve Administration, Yanchi, Ningxia 751500, China
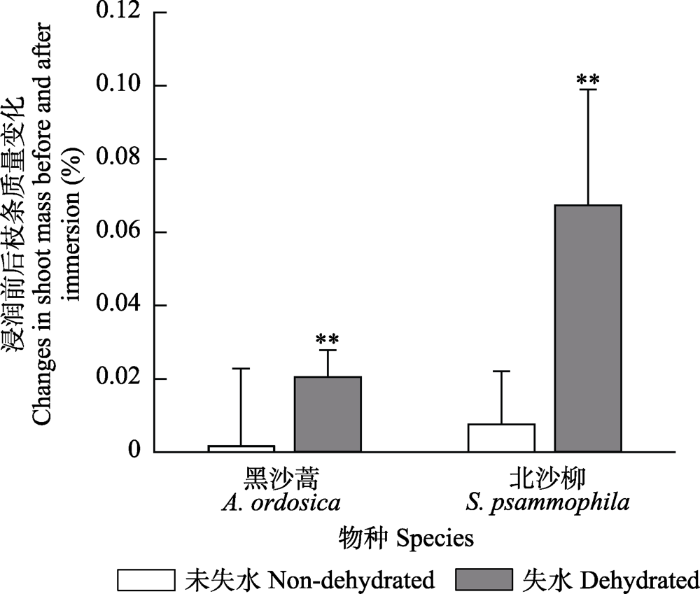
Abstract Aims Condensate is an important water source for plants in the ecosystems of drylands. Previous studies have found that some desert plants can absorb condensate via leaves. This study aimed to determine the capacity of the foliar condensate absorption of typical shrub species (Artemisia ordosica and Salix psammophila) in the Mu Us Desert, and to explore the pathways of foliar condensate absorption and transport. Methods The dehydrated and non-dehydrated detached shoots of A. ordosica and S. psammophilawere placed in an artificial chamber and exposed to deuterium labelled condensate, and the foliar condensate absorption was determined by comparing the differences of shoot masses and isotopic signals between pre- and post-immersion. The potted whole plants of A. ordosica and S. psammophilawere placed in an artificial chamber and exposed to fluorescent tracer solution, and the pathways of foliar water uptake and transport were determined by comparing the differences of fluorescent tracing in leaves and twigs between pre- and post-immersion. Important findings (1) After the deuterium labelled dew exposure, no significant differences were found in shoot masses between pre- and post-immersion of non-dehydrated detached shoots of A. ordosica and S. psammophila. However, the dehydrated shoot masses significantly increased by 2.04% and 6.74% in A. ordosica and S. psammophila, respectively; the δD (stable isotope ratio of hydrogen) of leaf water increased by 170.10‰ and 104.09‰ in A. ordosica and S. psammophila, respectively; and the δD of xylem water increased by 10.52‰ and 12.72‰ in A. ordosica and S. psammophila, respectively. (2) After the fluorescent tracer solution exposure, fluorescence was observed in the cuticles, stomata, spongy mesophyll, palisade cells and vascular bundle of the leaves of A. ordosica and S. psammophila. The fluorescence was also found in collenchyma of the leaves of A. ordosica. In addition, the fluorescence was observed in phloem, xylem, and pith of twigs of two shrub species. This study found that two typical shrub species in the Mu Us Desert had the capacity to absorb condensate via their leaves, and the plants undergoing water stress had the higher capacity of foliar condensate absorption. The leaves of A. ordosica and S. psammophilaabsorbed condensate through cuticles or stomata, and the absorbed water was transported to vascular bundle and even twigs. Foliar condensate absorption may be an important water use strategy to survive for A. ordosica and S. psammophila during dry periods. Keywords:Artemisia ordosica; Salix psammophila;foliar condensate absorption;condensate;water use strategy
PDF (4905KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 桂子洋, 秦树高, 胡朝, 白凤, 石慧书, 张宇清. 毛乌素沙地两种典型灌木叶片凝结水吸收能力及吸水途径. 植物生态学报, 2021, 45(6): 583-593. DOI: 10.17521/cjpe.2021.0066 GUI Zi-Yang, QIN Shu-Gao, HU Zhao, BAI Feng, SHI Hui-Shu, ZHANG Yu-Qing. Foliar condensate absorption and its pathways of two typical shrub species in the Mu Us Desert. Chinese Journal of Plant Ecology, 2021, 45(6): 583-593. DOI: 10.17521/cjpe.2021.0066
$\delta D(‰)=(R_{sample}-R_{standard}/R_{sample}×1000)$ 式中, Rsample和Rstandard分别表示植物组织水和标准水(标准平均海水, SMOW)的同位素比值。δD的测量精度为±0.3‰。鉴于样品中残留的有机污染物会干扰同位素分析(Schultz et al., 2011), 使用基于同位素比值红外光谱法的LWIA光谱污染识别软件来校正δD (West et al., 2011)。
$f_{dew}=\frac{\delta D _{post-immersion}-\delta D _{pre-immersion}}{\delta D _{dew}-\delta D _{pre-immersion}}×100$ 式中, δDpost-immersion为浸润后每个植物样品的氘同位素丰度, δDpre-immersion为浸润前植物样品的平均氘同位素丰度, δDdew为凝结水的氘同位素丰度。
新窗口打开|下载原图ZIP|生成PPT 图2黑沙蒿和北沙柳在浸润前后的叶片水与茎水氢稳定同位素比值(δD)变化(平均值±标准差)。**, p < 0.01。
Fig. 2Changes in hydrogen stable isotope ratio (δD) for leaf water and xylem water of Artemisia ordosica and Salix psammophila before and after immersion (mean ± SD). **, p < 0.01.
新窗口打开|下载原图ZIP|生成PPT 图3凝结水对黑沙蒿和北沙柳叶片水与茎水的相对贡献率(平均值±标准差)。*, p < 0.05; **, p < 0.01。
Fig. 3Relative contributions of condensate to total water in leaf and xylem of Artemisia ordosica and Salix psammophila (mean ± SD). *, p < 0.05; **, p < 0.01.
Fig. 4Leaf surface and stomata of Artemisia ordosica and Salix psammophila. A, Leaf surface of A. ordosica. B, C, Stomata of A. ordosica. D, Abaxial side of leaf of S. psammophila. E, Adaxial side of leaf of S. psammophila. F, Stomata of S. psammophila.
Fig. 5Cross-sections of leaf and twig of Artemisia ordosica and Salix psammophila in fluorescent tracing before and after immersion. A, E, Cross-sections of the leaf of A. ordosica and S. psammophila before immersion. B, F, Cross-sections of the leaf of A. ordosica and S. psammophila after immersion. C, G, Cross-sections of the twig of A. ordosica and S. psammophila before immersion. D, H, Cross-sections of the twig of A. ordosica and S. psammophila after immersion. Ca, collenchyma; Ct, cuticles; Pc, palisade cells; Ph, phloem; Pi, pith; Sc, spongy mesophyll; Vb, vascular bundle; Xl, xylem.
Fig. 6Cross-sections of leaf of Artemisia ordosica and Salix psammophila in fluorescent tracing (confocal laser scanning). A, D, Cross-sections of the leaf of A. ordosica and S. psammophil under fluorescence and bright light. B, E, Cross-sections of the leaf of A. ordosica and S. psammophil under fluorescence. C, F, Fluorescent bright spot in cross-sections of the leaf of A. ordosica and S. psammophil under fluorescence. Ca, collenchyma; Ct, cuticles; Pc, palisade cells; Sc, spongy mesophyll; St, stomata; Vb, vascular bundle.
BaiYX, SheWW, MichaletR, ZhengJ, QinSG, ZhangYQ (2018). Benefactor facilitation and beneficiary feedback effects drive shrub-dominated community succession in a semi-arid dune ecosystem Applied Vegetation Science, 21, 595-606. DOI:10.1111/avsc.12388URL [本文引用: 1]
BerryZC, EmeryNC, GotschSG, GoldsmithGR (2019). Foliar water uptake: processes, pathways, and integration into plant water budgets Plant, Cell & Environment, 42, 410-423. [本文引用: 4]
BerryZC, HughesNM, SmithWK (2014). Cloud immersion: an important water source for spruce and fir saplings in the southern Appalachian Mountains Oecologia, 174, 319-326. DOI:10.1007/s00442-013-2770-0URL [本文引用: 1]
BrewerCA, SmithWK, VogelmannTC (1991). Functional interaction between leaf trichomes, leaf wettability and the optical properties of water droplets Plant, Cell & Environment, 14, 955-962. [本文引用: 1]
BurkhardtJ, KaiserH, KappenL, GoldbachHE (2001). The possible role of aerosols on stomatal conductivity for water vapour Basic and Applied Ecology, 2, 351-364. DOI:10.1078/1439-1791-00062URL [本文引用: 1]
CairdMA, RichardsJH, DonovanLA (2007). Nighttime stomatal conductance and transpiration in C3 and C4 plants Plant Physiology, 143, 4-10. DOI:10.1104/pp.106.092940URL [本文引用: 2]
CassanaFF, EllerCB, OliveiraRS, DillenburgLR (2016). Effects of soil water availability on foliar water uptake of Araucaria angustifolia. Plant and Soil, 399, 147-157. [本文引用: 2]
CavallaroA, Carbonell SilletaL, PereyraDA, GoldsteinG, ScholzFG, BucciSJ (2020). Foliar water uptake in arid ecosystems: seasonal variability and ecophysiological consequences Oecologia, 193, 337-348. DOI:10.1007/s00442-020-04673-1PMID:32474806 [本文引用: 1] Foliar water uptake (FWU) has been reported for different species across several ecosystems types. However, little attention has been given to arid ecosystems, where FWU during dew formation or small rain events could ameliorate water deficits. FWU and their effects on leaf water potential (Ψ) were evaluated in grasses and shrubs exploring different soil water sources in a Patagonian steppe. Also, seasonal variability in FWU and the role of cell wall elasticity in determining the effects on Ψ were assessed. Eleven small rain events (<?8 mm) and 45 days with dew formation were recorded during the study period. All species exhibited FWU after experimental wetting. There was a large variability in FWU across species, from 0.04 mmol m s in species with deep roots to 0.75 mmol m s in species with shallow roots. Species-specific mean FWU rates were positively correlated with mean transpiration rates. The increase in Ψ after leaf wetting varied between 0.65 MPa and 1.67 MPa across species and seasons. The effects of FWU on Ψ were inversely correlated with cell wall elasticity. FWU integrated over both seasons varied between 28 mol m in species with deep roots to 361 mol m in species with shallow roots. Taking into account the percentage of coverage of each species, accumulated FWU represented 1.6% of the total annual transpiration of grasses and shrubs in this ecosystem. Despite this low FWU integrated over time compared to transpiration, wetting leaves surfaces can help to avoid larger water deficit during the dry season.
ChenD, ZhouHY, LiPG, ChenYL, WangYL, ZhaoX(2015). Circadian variations and regulation mechanism of eco-physiological characteristics of Artemisia ordosica and Caragana korshinskii. Journal of Desert Research, 35, 1549-1556. [本文引用: 1]
ChengXL, AnSQ, LiB, ChenJQ, LinGH, LiuYH, LuoYQ, LiuSR (2006). Summer rain pulse size and rainwater uptake by three dominant desert plants in a desertified grassland ecosystem in northwestern China Plant Ecology, 184, 1-12. DOI:10.1007/s11258-005-9047-6URL [本文引用: 1]
DawsonTE, EhleringerJR (1993). Isotopic enrichment of water in the “woody” tissues of plants: implications for plant water source, water uptake, and other studies which use the stable isotopic composition of cellulose Geochimica et Cosmochimica Acta, 57, 3487-3492. DOI:10.1016/0016-7037(93)90554-AURL [本文引用: 1]
DongmannG, NürnbergHW, FörstelH, WagenerK (1974). On the enrichment of H218O in the leaves of transpiring plants Radiation and Environmental Biophysics, 11, 41-52. PMID:4832051 [本文引用: 1]
EllerCB, LimaAL, OliveiraRS (2016). Cloud forest trees with higher foliar water uptake capacity and anisohydric behavior are more vulnerable to drought and climate change New Phytologist, 211, 489-501. DOI:10.1111/nph.2016.211.issue-2URL [本文引用: 5]
GoldsmithGR, LehmannMM, CernusakLA, ArendM, SiegwolfRTW (2017). Inferring foliar water uptake using stable isotopes of water Oecologia, 184, 763-766. DOI:10.1007/s00442-017-3917-1PMID:28735456 [本文引用: 1] A growing number of studies have described the direct absorption of water into leaves, a phenomenon known as foliar water uptake. The resultant increase in the amount of water in the leaf can be important for plant function. Exposing leaves to isotopically enriched or depleted water sources has become a common method for establishing whether or not a plant is capable of carrying out foliar water uptake. However, a careful inspection of our understanding of the fluxes of water isotopes between leaves and the atmosphere under high humidity conditions shows that there can clearly be isotopic exchange between the two pools even in the absence of a change in the mass of water in the leaf. We provide experimental evidence that while leaf water isotope ratios may change following exposure to a fog event using water with a depleted oxygen isotope ratio, leaf mass only changes when leaves are experiencing a water deficit that creates a driving gradient for the uptake of water by the leaf. Studies that rely on stable isotopes of water as a means of studying plant water use, particularly with respect to foliar water uptake, must consider the effects of these isotopic exchange processes.
GongXW, LüGH, HeXM, SarkarB, YangXD (2019). High air humidity causes atmospheric water absorption via assimilating branches in the deep-rooted tree Haloxylon ammodendron in an arid desert region of northwest China Frontiers in Plant Science, 10, 573. DOI: 10.3389/fpls2019.00573. URL [本文引用: 1]
GotschSG, AsbjornsenH, HolwerdaF, GoldsmithGR, WeintraubAE, DawsonTE (2014). Foggy days and dry nights determine crown-level water balance in a seasonal tropical montane cloud forest Plant, Cell & Environment, 37, 261-272. [本文引用: 1]
GotschSG, NadkarniN, DarbyA, GlunkA, DixM, DavidsonK, DawsonTE (2015). Life in the treetops: ecophysiological strategies of canopy epiphytes in a tropical montane cloud forest Ecological Monographs, 85, 393-412. DOI:10.1890/14-1076.1URL [本文引用: 1]
GuoXN, ZhaTS, JiaX, WuB, FengW, XieJ, GongJN, ZhangYQ, PeltolaH (2016). Dynamics of dew in a cold desert-shrub ecosystem and its abiotic controls Atmosphere, 7, 32. DOI: 10.3390/atmos7030032. URL [本文引用: 1]
GuoXN(2017). Dynamics and Environmental Control of Nocturnal Vapor Exchange in a Desert Shrub Ecosystem in Mu Us Desert PhD dissertation, Beijing Forestry University,Beijing. [本文引用: 3]
HalesS (1727). Vegetable Staticks Isaac Newton, London. [本文引用: 1]
HaoXM, LiC, GuoB, MaJX, AyupM, ChenZS (2012). Dew formation and its long-term trend in a desert riparian forest ecosystem on the eastern edge of the Taklimakan Desert in China Journal of Hydrology, 472-473, 90-98. [本文引用: 1]
HeJS, ChenWL, WangXL(1994). Morphological and anatomical features of quercus section suber and its adaptation to the ecological environment Acta Phytoecologica Sinica, 18, 219-227. [本文引用: 1]
JiaX, ZhaTS, GongJN, ZhangYQ, WuB, QinSG, PeltolaH (2018). Multi-scale dynamics and environmental controls on net ecosystem CO2 exchange over a temperate semiarid shrubland Agricultural and Forest Meteorology, 259, 250-259. DOI:10.1016/j.agrformet.2018.05.009URL [本文引用: 2]
KimK, LeeX (2011). Transition of stable isotope ratios of leaf water under simulated dew formation Plant, Cell & Environment, 34, 1790-1801. [本文引用: 2]
LaurJ, HackeUG (2014). Exploring Picea glauca aquaporins in the context of needle water uptake and xylem refilling New Phytologist, 203, 388-400. DOI:10.1111/nph.2014.203.issue-2URL [本文引用: 1]
LiLC, GuiZY, QinSG, ZhangYQ, LiuL, YangKJ(2021). Foliar condensate absorption capacity of four typical plant species and their physiological responses to water in the Mu Us Sandy Land of northwestern China Journal of Beijing Forestry University, 43(2), 72-80. [本文引用: 1]
LiuMZ, CenY, WangCD, GuX, BowlerP, WuDX, ZhangL, JiangGM, BeysensD (2020). Foliar uptake of dew in the sandy ecosystem of the Mongolia Plateau: a life-sustaining and carbon accumulation strategy shared differently by C3 and C4 grasses Agricultural and Forest Meteorology, 287, 107941. DOI: 10.1016/j.agrformet.2020.107941. URL [本文引用: 1]
LiuWJ, ZengJM, WangCM, LiHM, DuanWP(2001). On the relationship between forests and occult precipitation (dew and fog precipitation) Journal of Natural Resources, 16, 571-575. [本文引用: 1]
MalekE, McCurdyG, GilesB (1999). Dew contribution to the annual water balances in semi-arid desert valleys Journal of Arid Environments, 42, 71-80. DOI:10.1006/jare.1999.0506URL [本文引用: 1]
MartinCE, von WillertAD (2000). Leaf epidermal hydathodes and the ecophysiological consequences of foliar water uptake in species of Crassula from the Namib Desert in southern Africa. Plant Biology, 2, 229-242. [本文引用: 1]
MayrS, SchmidP, LaurJ, RosnerS, Charra-VaskouK, DämonB, HackeUG (2014). Uptake of water via branches helps timberline conifers refill embolized xylem in late winter Plant Physiology, 164, 1731-1740. DOI:10.1104/pp.114.236646URL [本文引用: 3]
McCullohKA, JohnsonDM, MeinzerFC, LachenbruchB (2011). An annual pattern of native embolism in upper branches of four tall conifer species American Journal of Botany, 98, 1007-1015. DOI:10.3732/ajb.1000503URL [本文引用: 1]
MitchellD, HenschelJR, HetemRS, WassenaarTD, StraussWM, HanrahanSA, SeelyMK (2020). Fog and fauna of the Namib Desert: past and future Ecosphere, 11, e02996. DOI: 10.1002/ecs2.2996. [本文引用: 1]
Munné-BoschS, AlegreL (1999). Role of dew on the recovery of water-stressed Melissa officinalis L. plants Journal of Plant Physiology, 154, 759-766. DOI:10.1016/S0176-1617(99)80255-7URL [本文引用: 1]
PanLZ, GuoW, WangT, LiYP, YangSJ(2021). Research progress on foliar water uptake Plant Physiology Journal, 57, 19-32. [本文引用: 1]
RiedererM, SchreiberL (2001). Protecting against water loss: analysis of the barrier properties of plant cuticles Journal of Experimental Botany, 52, 2023-2032. PMID:11559738 [本文引用: 1] The cuticle is the major barrier against uncontrolled water loss from leaves, fruits and other primary parts of higher plants. More than 100 mean values for water permeabilities determined with isolated leaf and fruit cuticles from 61 plant species are compiled and discussed in relation to plant organ, natural habitat and morphology. The maximum barrier properties of plant cuticles exceed that of synthetic polymeric films of equal thickness. Cuticular water permeability is not correlated to the thickness of the cuticle or to wax coverage. Relationships between cuticular permeability, wax composition and physical properties of the cuticle are evaluated. Cuticular permeability to water increases on the average by a factor of 2 when leaf surface temperature is raised from 15 degrees C to 35 degrees C. Organic compounds of anthropogenic and biogenic origin may enhance cuticular permeability. The pathway taken by water across the cuticular transport barrier is reviewed. The conclusion from this discussion is that the bulk of water diffuses as single molecules across a lipophilic barrier while a minor fraction travels along polar pores. Open questions concerning the mechanistic understanding of the plant cuticular transport barrier and the role the plant cuticle plays in ensuring the survival and reproductive success of an individual plant are indicated.
SchönherrJ (2006). Characterization of aqueous pores in plant cuticles and permeation of ionic solutes Journal of Experimental Botany, 57, 2471-2491. PMID:16825315 [本文引用: 1] Plant cuticles are lipid membranes with separate diffusion paths for lipophilic non-electrolytes and hydrated ionic compounds. Ions are lipid insoluble and require an aqueous pathway across cuticles. Based on experimental data, the aqueous pathway in cuticles has been characterized. Aqueous pores arise by hydration of permanent dipoles and ionic functional groups. They can be localized using ionic fluorescent dyes, silver nitrate, and mercuric chloride. Aqueous pores preferentially occur in cuticular ledges, at the base of trichomes, and in cuticles over anticlinal walls. Average pore radii ranged from 0.45 to 1.18 nm. Penetration of ions was a first order process as the fraction of the salt remaining on the cuticle surface decreased exponentially with time. Permeability of cuticles to ions depended on humidity and was highest at 100% humidity. Wetting agents increased rate constants by factors of up to 12, which indicates that the pore openings are surrounded by waxes. The pores in cuticular ledges of Helxine soleirolii allowed passage of berberine sulphate, which has a molecular weight of 769 g mol(-1). Increasing the molecular weight of solutes from 100 to 500 g mol(-1) decreased the rate constants of penetration by factors of 7 (Vicia faba) and 13 (Populus canescens), respectively. Half-times of penetration of inorganic salts and organic ions across Populus cuticles and Vicia leaf surfaces varied between 1 and 12 h. This shows that penetration of ionic compounds can be fairly rapid, and ions with molecular weights of up to 800 g mol(-1) can penetrate cuticles that possess aqueous pores.
SchreelJDM, van de WalBAE, Hervé-FernandezP, BoeckxP, SteppeK (2019a). Hydraulic redistribution of foliar absorbed water causes turgor-driven growth in mangrove seedlings Plant, Cell & Environment, 42, 2437-2447. [本文引用: 1]
SchreelJDM, von der CroneJS, KangurO, SteppeK (2019b). Influence of drought on foliar water uptake capacity of temperate tree species Forests, 10, 562. DOI: 10.3390/f10070562. URL [本文引用: 1]
SchultzNM, GriffisTJ, LeeX, BakerJM (2011). Identification and correction of spectral contamination in 2H/1H and 18O/16O measured in leaf, stem, and soil water Rapid Communications in Mass Spectrometry, 25, 3360-3368. DOI:10.1002/rcm.5236URL [本文引用: 1]
SnyderKA, RichardsJH, DonovanLA (2003). Night-time conductance in C3 and C4 species: Do plants lose water at night? Journal of Experimental Botany, 54, 861-865. PMID:12554729 [本文引用: 1] Significant night-time stomatal conductance and transpiration were found for 11 out of 17 species with a range of life histories (herbaceous annual, perennial grass, shrub, tree), photosynthetic pathways (C(3), C(4)), and habitats in the western United States. Across species and habitats, higher night-time conductance and transpiration were associated with higher daytime values. The prevalence, mechanisms and ecological implications of substantial night-time water loss deserve further investigation.
SparksJP, CampbellGS, BlackAR (2001). Water content, hydraulic conductivity, and ice formation in winter stems of Pinus contorta: a TDR case study Oecologia, 127, 468-475. DOI:10.1007/s004420000587PMID:28547483 [本文引用: 1] Stem water content, ice fraction, and losses in xylem conductivity were monitored from November 1996 to October 1997 in an even-aged stand of Pinus contorta (lodgepole pine) near Potlatch, Idaho, USA. A time domain reflectometry (TDR) probe was used to continuously monitor stem water contents and ice fractions. Stem sapwood water contents measured with TDR were not different from water contents measured gravimetrically. The liquid water content of stems ranged from 0.70 m m to 0.20 m m associated with freezing and thawing of the wood tissue. Ice fraction of the stem varied from 0-75% during the winter suggesting liquid water was always present even at ambient temperatures below -20°C. Shoot xylem tensions decreased through the winter to a minimum of ca. -1.4 MPa in February then increased to -0.4 MPa in May. Shoot xylem tensions decreased during the growing season reaching -1.7 MPa by September. Annually, low shoot water potentials were not correlated to decreases in stem hydraulic conductivity. Xylem conductivity decreased due to cavitation through the winter and was 70% of summer values by March. Decreases in xylem conductivity were correlated to low shoot water potentials and cumulative freezing and thawing events within the xylem. Xylem conductivity increased to pre-winter values by May and no reductions in xylem conductivity were observed during the growing season.
WangF, GuoSJ, HanFG, WangFL, ZhangWX, ZhangYN(2020). Study on leaf water uptake traits of desert plants in Minqin Arid Zone Research, 37, 1256-1263. [本文引用: 1]
WangHX, DingJ, DaiLM, WangXG, LinT (2010). Directional water-transfer through fabrics induced by asymmetric wettability Journal of Materials Chemistry, 20, 7938-7940. DOI:10.1039/c0jm02364gURL [本文引用: 1]
WangXH, XiaoHL, ChengYB, RenJ (2016a). Leaf epidermal water-absorbing scales and their absorption of unsaturated atmospheric water in Reaumuria soongorica, a desert plant from the northwest arid region of China Journal of Arid Environments, 128, 17-29. DOI:10.1016/j.jaridenv.2016.01.005URL [本文引用: 1]
WangXH, XiaoHL, RenJ, ChengYB, YangQ (2016b). An ultrasonic humidification fluorescent tracing method for detecting unsaturated atmospheric water absorption by the aerial parts of desert plants Journal of Arid Land, 8, 272-283. DOI:10.1007/s40333-015-0018-zURL [本文引用: 1]
WestAG, GoldsmithGR, MatimatiI, DawsonTE (2011). Spectral analysis software improves confidence in plant and soil water stable isotope analyses performed by isotope ratio infrared spectroscopy (IRIS) Rapid Communications in Mass Spectrometry, 25, 2268-2274. [本文引用: 1]
YanX, ZhouMX, DongXC, ZouSB, XiaoHL, MaXF (2015). Molecular mechanisms of foliar water uptake in a desert tree AoB Plants, 7, plv129. DOI: 10.1093/aobpla/plv129. [本文引用: 1]
YangLZ, FengL, YangGS, HuangL(2020). Water absorption potential and influencing factors of leaf in Caragana korshinskii, Artemisia ordosica, Hedysarum scoparium in a revegetated area of the Tengger Desert, China Journal of Desert Research, 40, 214-221. [本文引用: 2]
YangQ(2016). Effect of Rainfall on Transpiration of Artemisia ordosica and Salix psammophila in the Mu Us Desert. Master degree dissertation, Beijing Forestry University Beijing. [本文引用: 1]
YatesDJ, HutleyLB (1995). Foliar uptake of water by wet leaves of Sloanea woollsii, an Australian subtropical rainforest tree Australian Journal of Botany, 43, 157. DOI: 10.1071/bt9950157. URL [本文引用: 1]
YinLH, HuangJT, WangXY, MaHY, ZhangJ, DongJQ(2016). Analyses on change in leaf water potential of four species in Maowusu sandland and its influence factors Journal of Plant Resources and Environment, 25, 17-23. [本文引用: 1]
ZangvilA (1996). Six years of dew observations in the Negev Desert, Israel Journal of Arid Environments, 32, 361-371. DOI:10.1006/jare.1996.0030URL [本文引用: 1]
ZhangJ, ZhangYM, DowningA, ChengJH, ZhouXB, ZhangBC (2009). The influence of biological soil crusts on dew deposition in Gurbantunggut Desert, Northwestern China Journal of Hydrology, 379, 220-228. DOI:10.1016/j.jhydrol.2009.09.053URL [本文引用: 1]
ZhengXJ, LiS, LiY(2011). Leaf water uptake strategy of desert plants in the Junggar Basin, China Chinese Journal of Plant Ecology, 35, 893-905. DOI:10.3724/SP.J.1258.2011.00893URL [本文引用: 3]
ZhuYJ, JiaZQ, LuQ, HaoYG, ZhangJB, LiL, QiYL(2010). Water use strategy of five shrubs in Ulanbuh Desert Scientia Silvae Sinicae, 46(4), 15-21. [本文引用: 1]
ZhuangYL, RatcliffeS (2012). Relationship between dew presence and Bassia dasyphylla plant growth Journal of Arid Land, 4, 11-18. DOI:10.3724/SP.J.1227.2012.00011URL [本文引用: 2]
ZhuangYL, ZhaoWZ(2010). Experimental study of effects of artificial dew on Bassia dasyphylla and Agriophyllum squarrosum Journal of Desert Research, 30, 1068-1074. [本文引用: 2]
Isotopic enrichment of water in the “woody” tissues of plants: implications for plant water source, water uptake, and other studies which use the stable isotopic composition of cellulose 1 1993
On the enrichment of H218O in the leaves of transpiring plants 1 1974
... 模拟凝结水浸润后, 黑沙蒿和北沙柳离体枝条叶片水同位素组成均发生了显著改变.根据叶片水同位素组成改变计算出的凝结水对叶片水和茎水的贡献率远大于枝条质量增加率(图1), 表明叶片在凝结水浸润中, 发生了与凝结水之间水汽净通量为0的双向交换.双向交换过程中, 凝结水置换叶水, 并改变了叶片水同位素组成(Goldsmith et al., 2017).在离体枝条浸润实验中, 凝结水对失水枝条的叶水和茎水相对贡献率小于未失水枝条, 这是由于双向交换强度主要受大气湿度和气孔导度影响(Dongmann et al., 1974), 缺水叶片的气孔部分关闭降低了水汽交换速率(Schreel et al., 2019b), 但并未改变失水叶片通过水势梯度吸收凝结水. ...
Cloud forest trees with higher foliar water uptake capacity and anisohydric behavior are more vulnerable to drought and climate change 5 2016
... 植物叶片具有多种吸水结构, 露水可通过叶毛(Eller et al., 2016)、角质层(Goldsmith et al., 2013; Eller et al., 2016)、气孔(Kim & Lee, 2011)、水孔(叶片尖端或边缘的排水结构)(Martin & von Willert, 2000)等结构进入叶片, 并经过叶肉组织向维管束运移(Kim & Lee, 2011; Eller et al., 2016).由于气孔的水力导度远大于其他结构, 一般认为气孔是主要的吸水通道; 而当夜间气孔关闭时, 叶片吸水可能主要通过角质层(Eller et al., 2016).沙生植物的叶片通常具有特殊的耐旱特征, 如下陷的气孔和致密的角质层, 但目前并不清楚这些叶片特性如何影响吸水.此外, 一些沙生灌木如红砂(Reaumuria soongarica), 具有特定的吸水结构(Wang et al., 2016a), 但其他沙生植物能否通过特殊结构吸收水分并不清楚. ...
... ; Eller et al., 2016)、气孔(Kim & Lee, 2011)、水孔(叶片尖端或边缘的排水结构)(Martin & von Willert, 2000)等结构进入叶片, 并经过叶肉组织向维管束运移(Kim & Lee, 2011; Eller et al., 2016).由于气孔的水力导度远大于其他结构, 一般认为气孔是主要的吸水通道; 而当夜间气孔关闭时, 叶片吸水可能主要通过角质层(Eller et al., 2016).沙生植物的叶片通常具有特殊的耐旱特征, 如下陷的气孔和致密的角质层, 但目前并不清楚这些叶片特性如何影响吸水.此外, 一些沙生灌木如红砂(Reaumuria soongarica), 具有特定的吸水结构(Wang et al., 2016a), 但其他沙生植物能否通过特殊结构吸收水分并不清楚. ...
... ; Eller et al., 2016).由于气孔的水力导度远大于其他结构, 一般认为气孔是主要的吸水通道; 而当夜间气孔关闭时, 叶片吸水可能主要通过角质层(Eller et al., 2016).沙生植物的叶片通常具有特殊的耐旱特征, 如下陷的气孔和致密的角质层, 但目前并不清楚这些叶片特性如何影响吸水.此外, 一些沙生灌木如红砂(Reaumuria soongarica), 具有特定的吸水结构(Wang et al., 2016a), 但其他沙生植物能否通过特殊结构吸收水分并不清楚. ...
... ).由于气孔的水力导度远大于其他结构, 一般认为气孔是主要的吸水通道; 而当夜间气孔关闭时, 叶片吸水可能主要通过角质层(Eller et al., 2016).沙生植物的叶片通常具有特殊的耐旱特征, 如下陷的气孔和致密的角质层, 但目前并不清楚这些叶片特性如何影响吸水.此外, 一些沙生灌木如红砂(Reaumuria soongarica), 具有特定的吸水结构(Wang et al., 2016a), 但其他沙生植物能否通过特殊结构吸收水分并不清楚. ...
... 模拟凝结水浸润后, 未失水的黑沙蒿和北沙柳枝条茎水氘同位素显著富集(图2), 这可能是叶片吸收的水分运移至茎干, 改变了茎水同位素组成.叶片吸水运移在很多植物种均有发现, 当叶片水势高于茎干和根系, 叶片吸收的水分可以向下运移至茎、根乃至根际土壤(Laur & Hacke, 2014; Eller et al., 2016).沙生植物频繁经受水分胁迫, 叶片吸水向下运移可以缓解植物栓塞, 减少植物在干旱期的死亡率(McCulloh et al., 2011; Mayr et al., 2014).此外, 一些研究发现, 同化枝及未栓化的枝条也具有吸水能力(Sparks et al., 2001; Mayr et al., 2014), 而且枝条吸收的水分可以通过渗透势向内运输, 直接补充木质部以缓解栓塞(Mayr et al., 2014).在本研究中, 凝结水对北沙柳茎水的相对贡献率较高, 很可能是北沙柳枝条直接吸水的结果. ...
Inferring foliar water uptake using stable isotopes of water 1 2017
... 模拟凝结水浸润后, 黑沙蒿和北沙柳离体枝条叶片水同位素组成均发生了显著改变.根据叶片水同位素组成改变计算出的凝结水对叶片水和茎水的贡献率远大于枝条质量增加率(图1), 表明叶片在凝结水浸润中, 发生了与凝结水之间水汽净通量为0的双向交换.双向交换过程中, 凝结水置换叶水, 并改变了叶片水同位素组成(Goldsmith et al., 2017).在离体枝条浸润实验中, 凝结水对失水枝条的叶水和茎水相对贡献率小于未失水枝条, 这是由于双向交换强度主要受大气湿度和气孔导度影响(Dongmann et al., 1974), 缺水叶片的气孔部分关闭降低了水汽交换速率(Schreel et al., 2019b), 但并未改变失水叶片通过水势梯度吸收凝结水. ...
The incidence and implications of clouds for cloud forest plant water relations 2 2013
... 植物叶片具有多种吸水结构, 露水可通过叶毛(Eller et al., 2016)、角质层(Goldsmith et al., 2013; Eller et al., 2016)、气孔(Kim & Lee, 2011)、水孔(叶片尖端或边缘的排水结构)(Martin & von Willert, 2000)等结构进入叶片, 并经过叶肉组织向维管束运移(Kim & Lee, 2011; Eller et al., 2016).由于气孔的水力导度远大于其他结构, 一般认为气孔是主要的吸水通道; 而当夜间气孔关闭时, 叶片吸水可能主要通过角质层(Eller et al., 2016).沙生植物的叶片通常具有特殊的耐旱特征, 如下陷的气孔和致密的角质层, 但目前并不清楚这些叶片特性如何影响吸水.此外, 一些沙生灌木如红砂(Reaumuria soongarica), 具有特定的吸水结构(Wang et al., 2016a), 但其他沙生植物能否通过特殊结构吸收水分并不清楚. ...
... 角质层也被认为是叶片吸水的主要途径(Yates & Hutley, 1995; Goldsmith et al., 2013).角质层由带有羟基和羧基的长碳链脂肪酸聚合物组成, 其中的分子间隙及分子上的羟基、羧基亲水基团可使水渗透进入叶内.在本研究中, 荧光剂分布于黑沙蒿表面和北沙柳叶片两面.此外, 荧光示踪显示, 北沙柳叶片背面的角质层比腹面富集了更多的荧光剂(图6F), 这可能是腹面的叶毛簇具有一定的斥水性.尽管一些植物的叶毛具有亲水性, 能够促进叶面浸润乃至直接吸收水分(Schönherr, 2006; Wang et al., 2010), 但叶毛的性质和密度会影响叶面的润湿性, 当叶毛密度大于25 Ind.·mm-2时, 叶面具有很强的斥水性(Brewer et al., 1991).北沙柳腹面密布的叶毛具有遮蔽气孔、阻止蒸发的功能, 但也降低了叶面润湿性和气孔导度, 从而降低了腹面的吸水能力. ...
High air humidity causes atmospheric water absorption via assimilating branches in the deep-rooted tree Haloxylon ammodendron in an arid desert region of northwest China 1 2019
... 自Hales (1727)首次提出叶片吸水以来, 已经在200多种植物、6个主要生态系统中发现此种现象(Berry et al., 2019).作为一种广泛存在的功能, 叶片吸水可显著改善植物水分状况.对两种常绿灌木的研究结果表明, 具有叶片吸水功能的植物种经历长期干旱后, 凝结水的输入使叶片组织相对含水量和小枝水势均有大幅提高, 水分亏缺状况得到缓解, 而不具吸水能力的植物种持续受到干旱胁迫影响(Munné-Bosch & Alegre, 1999).在云雾林中的观测发现, 利用氢氧同位素示踪计算得到的凝结水对欧洲云杉(Picea abies)水分来源的贡献率可达31% (Berry et al., 2014), 吸收凝结水产生的茎流逆流占旱季蒸腾量的9% (Gotsch et al., 2014).长期凝结水添加可显著提高供试植物的株高、生物量(庄艳丽和赵文智, 2010; Zhuang & Ratchiffe, 2012; Liu et al., 2020).已有研究发现, 荒漠区植物普遍具有叶片吸水潜力(郑新军等, 2011; 王飞等, 2020; 杨利贞等, 2020), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
Foggy days and dry nights determine crown-level water balance in a seasonal tropical montane cloud forest 1 2014
... 自Hales (1727)首次提出叶片吸水以来, 已经在200多种植物、6个主要生态系统中发现此种现象(Berry et al., 2019).作为一种广泛存在的功能, 叶片吸水可显著改善植物水分状况.对两种常绿灌木的研究结果表明, 具有叶片吸水功能的植物种经历长期干旱后, 凝结水的输入使叶片组织相对含水量和小枝水势均有大幅提高, 水分亏缺状况得到缓解, 而不具吸水能力的植物种持续受到干旱胁迫影响(Munné-Bosch & Alegre, 1999).在云雾林中的观测发现, 利用氢氧同位素示踪计算得到的凝结水对欧洲云杉(Picea abies)水分来源的贡献率可达31% (Berry et al., 2014), 吸收凝结水产生的茎流逆流占旱季蒸腾量的9% (Gotsch et al., 2014).长期凝结水添加可显著提高供试植物的株高、生物量(庄艳丽和赵文智, 2010; Zhuang & Ratchiffe, 2012; Liu et al., 2020).已有研究发现, 荒漠区植物普遍具有叶片吸水潜力(郑新军等, 2011; 王飞等, 2020; 杨利贞等, 2020), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
Life in the treetops: ecophysiological strategies of canopy epiphytes in a tropical montane cloud forest 1 2015
Multi-scale dynamics and environmental controls on net ecosystem CO2 exchange over a temperate semiarid shrubland 2 2018
... 本研究在毛乌素沙漠西南边缘的宁夏盐池毛乌素沙地生态系统国家定位观测研究站(37.07°- 38.17° N, 106.50°-107.68° E, 海拔1 530 m)开展.研究区属半干旱大陆性季风气候, 年平均气温为8.1 ℃, 冬夏温差为28 ℃, 昼夜温差为20 ℃.年降水量为292 mm, 雨季为6-9月, 降水量约占全年的80% (Jia et al., 2018).凝结水年发生天数占全年的78%, 日凝结水量为(0.15 ± 0.08) mm (郭晓楠, 2017).研究区植被主要由黑沙蒿、北沙柳、蒙古山竹子(Corethrodendron fruticosum var. mongolicum)、柠条锦鸡儿(Caragana korshinskii)等灌木, 以及赖草(Leymus secalinus)、沙蓬(Agriophyllum squarrosum)等草本植物组成(Bai et al., 2018; Jia et al., 2018). ...
... ; Jia et al., 2018). ...
Transition of stable isotope ratios of leaf water under simulated dew formation 2 2011
... 植物叶片具有多种吸水结构, 露水可通过叶毛(Eller et al., 2016)、角质层(Goldsmith et al., 2013; Eller et al., 2016)、气孔(Kim & Lee, 2011)、水孔(叶片尖端或边缘的排水结构)(Martin & von Willert, 2000)等结构进入叶片, 并经过叶肉组织向维管束运移(Kim & Lee, 2011; Eller et al., 2016).由于气孔的水力导度远大于其他结构, 一般认为气孔是主要的吸水通道; 而当夜间气孔关闭时, 叶片吸水可能主要通过角质层(Eller et al., 2016).沙生植物的叶片通常具有特殊的耐旱特征, 如下陷的气孔和致密的角质层, 但目前并不清楚这些叶片特性如何影响吸水.此外, 一些沙生灌木如红砂(Reaumuria soongarica), 具有特定的吸水结构(Wang et al., 2016a), 但其他沙生植物能否通过特殊结构吸收水分并不清楚. ...
... )等结构进入叶片, 并经过叶肉组织向维管束运移(Kim & Lee, 2011; Eller et al., 2016).由于气孔的水力导度远大于其他结构, 一般认为气孔是主要的吸水通道; 而当夜间气孔关闭时, 叶片吸水可能主要通过角质层(Eller et al., 2016).沙生植物的叶片通常具有特殊的耐旱特征, 如下陷的气孔和致密的角质层, 但目前并不清楚这些叶片特性如何影响吸水.此外, 一些沙生灌木如红砂(Reaumuria soongarica), 具有特定的吸水结构(Wang et al., 2016a), 但其他沙生植物能否通过特殊结构吸收水分并不清楚. ...
Exploring Picea glauca aquaporins in the context of needle water uptake and xylem refilling 1 2014
... 模拟凝结水浸润后, 未失水的黑沙蒿和北沙柳枝条茎水氘同位素显著富集(图2), 这可能是叶片吸收的水分运移至茎干, 改变了茎水同位素组成.叶片吸水运移在很多植物种均有发现, 当叶片水势高于茎干和根系, 叶片吸收的水分可以向下运移至茎、根乃至根际土壤(Laur & Hacke, 2014; Eller et al., 2016).沙生植物频繁经受水分胁迫, 叶片吸水向下运移可以缓解植物栓塞, 减少植物在干旱期的死亡率(McCulloh et al., 2011; Mayr et al., 2014).此外, 一些研究发现, 同化枝及未栓化的枝条也具有吸水能力(Sparks et al., 2001; Mayr et al., 2014), 而且枝条吸收的水分可以通过渗透势向内运输, 直接补充木质部以缓解栓塞(Mayr et al., 2014).在本研究中, 凝结水对北沙柳茎水的相对贡献率较高, 很可能是北沙柳枝条直接吸水的结果. ...
毛乌素沙地4种典型植物叶片凝结水吸收能力及其水分生理响应 1 2021
... 水分是荒漠生态系统最主要的环境限制要素.凝结水是干旱半干旱区重要的补充性水源, 发挥着不可忽视的作用(Zhang et al., 2009; Hao et al., 2012).在有些地区, 如纳米布沙漠, 凝结水量可超过年降水量(Mitchell et al., 2020).而在以色列内盖夫沙漠、美国内华达沙漠、中国毛乌素沙地, 凝结水量可达年降水量的7.2%-16.2% (Zangvil, 1996; Malek et al., 1999; 郭晓楠, 2017).在凝结水事件中, 土壤表层水分可以得到一定改善(刘文杰等, 2001), 叶片温度及气孔导度降低, 致使植物蒸腾耗水减少(Zhuang & Ratchiffe, 2012; 李鹭辰等, 2021), 叶片还可以直接吸收利用叶片表面的凝结水. ...
毛乌素沙地4种典型植物叶片凝结水吸收能力及其水分生理响应 1 2021
... 水分是荒漠生态系统最主要的环境限制要素.凝结水是干旱半干旱区重要的补充性水源, 发挥着不可忽视的作用(Zhang et al., 2009; Hao et al., 2012).在有些地区, 如纳米布沙漠, 凝结水量可超过年降水量(Mitchell et al., 2020).而在以色列内盖夫沙漠、美国内华达沙漠、中国毛乌素沙地, 凝结水量可达年降水量的7.2%-16.2% (Zangvil, 1996; Malek et al., 1999; 郭晓楠, 2017).在凝结水事件中, 土壤表层水分可以得到一定改善(刘文杰等, 2001), 叶片温度及气孔导度降低, 致使植物蒸腾耗水减少(Zhuang & Ratchiffe, 2012; 李鹭辰等, 2021), 叶片还可以直接吸收利用叶片表面的凝结水. ...
Foliar uptake of dew in the sandy ecosystem of the Mongolia Plateau: a life-sustaining and carbon accumulation strategy shared differently by C3 and C4 grasses 1 2020
... 自Hales (1727)首次提出叶片吸水以来, 已经在200多种植物、6个主要生态系统中发现此种现象(Berry et al., 2019).作为一种广泛存在的功能, 叶片吸水可显著改善植物水分状况.对两种常绿灌木的研究结果表明, 具有叶片吸水功能的植物种经历长期干旱后, 凝结水的输入使叶片组织相对含水量和小枝水势均有大幅提高, 水分亏缺状况得到缓解, 而不具吸水能力的植物种持续受到干旱胁迫影响(Munné-Bosch & Alegre, 1999).在云雾林中的观测发现, 利用氢氧同位素示踪计算得到的凝结水对欧洲云杉(Picea abies)水分来源的贡献率可达31% (Berry et al., 2014), 吸收凝结水产生的茎流逆流占旱季蒸腾量的9% (Gotsch et al., 2014).长期凝结水添加可显著提高供试植物的株高、生物量(庄艳丽和赵文智, 2010; Zhuang & Ratchiffe, 2012; Liu et al., 2020).已有研究发现, 荒漠区植物普遍具有叶片吸水潜力(郑新军等, 2011; 王飞等, 2020; 杨利贞等, 2020), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
森林与雾露水关系研究进展 1 2001
... 水分是荒漠生态系统最主要的环境限制要素.凝结水是干旱半干旱区重要的补充性水源, 发挥着不可忽视的作用(Zhang et al., 2009; Hao et al., 2012).在有些地区, 如纳米布沙漠, 凝结水量可超过年降水量(Mitchell et al., 2020).而在以色列内盖夫沙漠、美国内华达沙漠、中国毛乌素沙地, 凝结水量可达年降水量的7.2%-16.2% (Zangvil, 1996; Malek et al., 1999; 郭晓楠, 2017).在凝结水事件中, 土壤表层水分可以得到一定改善(刘文杰等, 2001), 叶片温度及气孔导度降低, 致使植物蒸腾耗水减少(Zhuang & Ratchiffe, 2012; 李鹭辰等, 2021), 叶片还可以直接吸收利用叶片表面的凝结水. ...
森林与雾露水关系研究进展 1 2001
... 水分是荒漠生态系统最主要的环境限制要素.凝结水是干旱半干旱区重要的补充性水源, 发挥着不可忽视的作用(Zhang et al., 2009; Hao et al., 2012).在有些地区, 如纳米布沙漠, 凝结水量可超过年降水量(Mitchell et al., 2020).而在以色列内盖夫沙漠、美国内华达沙漠、中国毛乌素沙地, 凝结水量可达年降水量的7.2%-16.2% (Zangvil, 1996; Malek et al., 1999; 郭晓楠, 2017).在凝结水事件中, 土壤表层水分可以得到一定改善(刘文杰等, 2001), 叶片温度及气孔导度降低, 致使植物蒸腾耗水减少(Zhuang & Ratchiffe, 2012; 李鹭辰等, 2021), 叶片还可以直接吸收利用叶片表面的凝结水. ...
Dew contribution to the annual water balances in semi-arid desert valleys 1 1999
... 水分是荒漠生态系统最主要的环境限制要素.凝结水是干旱半干旱区重要的补充性水源, 发挥着不可忽视的作用(Zhang et al., 2009; Hao et al., 2012).在有些地区, 如纳米布沙漠, 凝结水量可超过年降水量(Mitchell et al., 2020).而在以色列内盖夫沙漠、美国内华达沙漠、中国毛乌素沙地, 凝结水量可达年降水量的7.2%-16.2% (Zangvil, 1996; Malek et al., 1999; 郭晓楠, 2017).在凝结水事件中, 土壤表层水分可以得到一定改善(刘文杰等, 2001), 叶片温度及气孔导度降低, 致使植物蒸腾耗水减少(Zhuang & Ratchiffe, 2012; 李鹭辰等, 2021), 叶片还可以直接吸收利用叶片表面的凝结水. ...
Leaf epidermal hydathodes and the ecophysiological consequences of foliar water uptake in species of 1 2000
... 植物叶片具有多种吸水结构, 露水可通过叶毛(Eller et al., 2016)、角质层(Goldsmith et al., 2013; Eller et al., 2016)、气孔(Kim & Lee, 2011)、水孔(叶片尖端或边缘的排水结构)(Martin & von Willert, 2000)等结构进入叶片, 并经过叶肉组织向维管束运移(Kim & Lee, 2011; Eller et al., 2016).由于气孔的水力导度远大于其他结构, 一般认为气孔是主要的吸水通道; 而当夜间气孔关闭时, 叶片吸水可能主要通过角质层(Eller et al., 2016).沙生植物的叶片通常具有特殊的耐旱特征, 如下陷的气孔和致密的角质层, 但目前并不清楚这些叶片特性如何影响吸水.此外, 一些沙生灌木如红砂(Reaumuria soongarica), 具有特定的吸水结构(Wang et al., 2016a), 但其他沙生植物能否通过特殊结构吸收水分并不清楚. ...
Uptake of water via branches helps timberline conifers refill embolized xylem in late winter 3 2014
... 模拟凝结水浸润后, 未失水的黑沙蒿和北沙柳枝条茎水氘同位素显著富集(图2), 这可能是叶片吸收的水分运移至茎干, 改变了茎水同位素组成.叶片吸水运移在很多植物种均有发现, 当叶片水势高于茎干和根系, 叶片吸收的水分可以向下运移至茎、根乃至根际土壤(Laur & Hacke, 2014; Eller et al., 2016).沙生植物频繁经受水分胁迫, 叶片吸水向下运移可以缓解植物栓塞, 减少植物在干旱期的死亡率(McCulloh et al., 2011; Mayr et al., 2014).此外, 一些研究发现, 同化枝及未栓化的枝条也具有吸水能力(Sparks et al., 2001; Mayr et al., 2014), 而且枝条吸收的水分可以通过渗透势向内运输, 直接补充木质部以缓解栓塞(Mayr et al., 2014).在本研究中, 凝结水对北沙柳茎水的相对贡献率较高, 很可能是北沙柳枝条直接吸水的结果. ...
... ; Mayr et al., 2014), 而且枝条吸收的水分可以通过渗透势向内运输, 直接补充木质部以缓解栓塞(Mayr et al., 2014).在本研究中, 凝结水对北沙柳茎水的相对贡献率较高, 很可能是北沙柳枝条直接吸水的结果. ...
... ), 而且枝条吸收的水分可以通过渗透势向内运输, 直接补充木质部以缓解栓塞(Mayr et al., 2014).在本研究中, 凝结水对北沙柳茎水的相对贡献率较高, 很可能是北沙柳枝条直接吸水的结果. ...
An annual pattern of native embolism in upper branches of four tall conifer species 1 2011
... 模拟凝结水浸润后, 未失水的黑沙蒿和北沙柳枝条茎水氘同位素显著富集(图2), 这可能是叶片吸收的水分运移至茎干, 改变了茎水同位素组成.叶片吸水运移在很多植物种均有发现, 当叶片水势高于茎干和根系, 叶片吸收的水分可以向下运移至茎、根乃至根际土壤(Laur & Hacke, 2014; Eller et al., 2016).沙生植物频繁经受水分胁迫, 叶片吸水向下运移可以缓解植物栓塞, 减少植物在干旱期的死亡率(McCulloh et al., 2011; Mayr et al., 2014).此外, 一些研究发现, 同化枝及未栓化的枝条也具有吸水能力(Sparks et al., 2001; Mayr et al., 2014), 而且枝条吸收的水分可以通过渗透势向内运输, 直接补充木质部以缓解栓塞(Mayr et al., 2014).在本研究中, 凝结水对北沙柳茎水的相对贡献率较高, 很可能是北沙柳枝条直接吸水的结果. ...
Fog and fauna of the Namib Desert: past and future 1 2020
... 水分是荒漠生态系统最主要的环境限制要素.凝结水是干旱半干旱区重要的补充性水源, 发挥着不可忽视的作用(Zhang et al., 2009; Hao et al., 2012).在有些地区, 如纳米布沙漠, 凝结水量可超过年降水量(Mitchell et al., 2020).而在以色列内盖夫沙漠、美国内华达沙漠、中国毛乌素沙地, 凝结水量可达年降水量的7.2%-16.2% (Zangvil, 1996; Malek et al., 1999; 郭晓楠, 2017).在凝结水事件中, 土壤表层水分可以得到一定改善(刘文杰等, 2001), 叶片温度及气孔导度降低, 致使植物蒸腾耗水减少(Zhuang & Ratchiffe, 2012; 李鹭辰等, 2021), 叶片还可以直接吸收利用叶片表面的凝结水. ...
Role of dew on the recovery of water-stressed Melissa officinalis L. plants 1 1999
... 自Hales (1727)首次提出叶片吸水以来, 已经在200多种植物、6个主要生态系统中发现此种现象(Berry et al., 2019).作为一种广泛存在的功能, 叶片吸水可显著改善植物水分状况.对两种常绿灌木的研究结果表明, 具有叶片吸水功能的植物种经历长期干旱后, 凝结水的输入使叶片组织相对含水量和小枝水势均有大幅提高, 水分亏缺状况得到缓解, 而不具吸水能力的植物种持续受到干旱胁迫影响(Munné-Bosch & Alegre, 1999).在云雾林中的观测发现, 利用氢氧同位素示踪计算得到的凝结水对欧洲云杉(Picea abies)水分来源的贡献率可达31% (Berry et al., 2014), 吸收凝结水产生的茎流逆流占旱季蒸腾量的9% (Gotsch et al., 2014).长期凝结水添加可显著提高供试植物的株高、生物量(庄艳丽和赵文智, 2010; Zhuang & Ratchiffe, 2012; Liu et al., 2020).已有研究发现, 荒漠区植物普遍具有叶片吸水潜力(郑新军等, 2011; 王飞等, 2020; 杨利贞等, 2020), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
... 在模拟凝结水浸润实验中, 水分亏缺的黑沙蒿和北沙柳具有更强的叶片吸水能力.荒漠地区土壤干旱事件频发, 但土壤干旱期不乏小降水事件, 以及小降水事件后的凝结水事件(Guo et al., 2016).缺水植物的叶片吸水能够显著改善植物的水分生理状况(Cassana et al., 2016; Schreel & Steppe, 2020), 叶片吸收凝结水可能是沙生植物适应沙地频繁土壤干旱的重要抗旱策略. ...
Hydraulic redistribution of foliar absorbed water causes turgor-driven growth in mangrove seedlings 1 2019a
... 使用两箱线性模型(Schreel et al., 2019a)计算凝结水对叶片水和茎水的相对贡献: ...
Influence of drought on foliar water uptake capacity of temperate tree species 1 2019b
... 模拟凝结水浸润后, 黑沙蒿和北沙柳离体枝条叶片水同位素组成均发生了显著改变.根据叶片水同位素组成改变计算出的凝结水对叶片水和茎水的贡献率远大于枝条质量增加率(图1), 表明叶片在凝结水浸润中, 发生了与凝结水之间水汽净通量为0的双向交换.双向交换过程中, 凝结水置换叶水, 并改变了叶片水同位素组成(Goldsmith et al., 2017).在离体枝条浸润实验中, 凝结水对失水枝条的叶水和茎水相对贡献率小于未失水枝条, 这是由于双向交换强度主要受大气湿度和气孔导度影响(Dongmann et al., 1974), 缺水叶片的气孔部分关闭降低了水汽交换速率(Schreel et al., 2019b), 但并未改变失水叶片通过水势梯度吸收凝结水. ...
Identification and correction of spectral contamination in 2H/1H and 18O/16O measured in leaf, stem, and soil water 1 2011
... 式中, Rsample和Rstandard分别表示植物组织水和标准水(标准平均海水, SMOW)的同位素比值.δD的测量精度为±0.3‰.鉴于样品中残留的有机污染物会干扰同位素分析(Schultz et al., 2011), 使用基于同位素比值红外光谱法的LWIA光谱污染识别软件来校正δD (West et al., 2011). ...
Night-time conductance in C3 and C4 species: Do plants lose water at night? 1 2003
... 荧光标记凝结水浸润后, 黑沙蒿和北沙柳叶片的气孔和角质层均有示踪剂聚集(图5, 图6).叶片的气孔吸水现象已在番茄(Solanum lycopersicum)、梭梭(Haloxylon ammodendron)等植物中发现(Burkhardt et al., 2012; Wang et al., 2016b).当气孔被水分浸润时, 凝结水中的盐离子、气溶胶会降低水面张力, 促进液态水通过气孔(Burkhardt et al., 2001, 2012).由于气孔导度占叶面导度的95%以上, 一般认为白天气孔未关闭时, 气孔是叶片吸水的主要通道(Riederer & Schreiber, 2001; Berry et al., 2019).本项研究模拟了夜间露水浸润, 发现叶片吸水仍存在明显的气孔途径.近些年来的研究发现, 尽管气孔在夜晚部分关闭, 但仍能保持15%的气孔导度(Caird et al., 2007), 一些荒漠植物甚至可达30% (Snyder et al., 2003), 黑沙蒿也存在夜间气孔开放的现象(陈栋等, 2015).夜间气孔开放可以帮助植物补充养分和缓解栓塞(Caird et al., 2007), 也间接促进了叶片吸水. ...
Water content, hydraulic conductivity, and ice formation in winter stems of Pinus contorta: a TDR case study 1 2001
... 模拟凝结水浸润后, 未失水的黑沙蒿和北沙柳枝条茎水氘同位素显著富集(图2), 这可能是叶片吸收的水分运移至茎干, 改变了茎水同位素组成.叶片吸水运移在很多植物种均有发现, 当叶片水势高于茎干和根系, 叶片吸收的水分可以向下运移至茎、根乃至根际土壤(Laur & Hacke, 2014; Eller et al., 2016).沙生植物频繁经受水分胁迫, 叶片吸水向下运移可以缓解植物栓塞, 减少植物在干旱期的死亡率(McCulloh et al., 2011; Mayr et al., 2014).此外, 一些研究发现, 同化枝及未栓化的枝条也具有吸水能力(Sparks et al., 2001; Mayr et al., 2014), 而且枝条吸收的水分可以通过渗透势向内运输, 直接补充木质部以缓解栓塞(Mayr et al., 2014).在本研究中, 凝结水对北沙柳茎水的相对贡献率较高, 很可能是北沙柳枝条直接吸水的结果. ...
民勤荒漠植物叶片水分吸收性状研究 1 2020
... 自Hales (1727)首次提出叶片吸水以来, 已经在200多种植物、6个主要生态系统中发现此种现象(Berry et al., 2019).作为一种广泛存在的功能, 叶片吸水可显著改善植物水分状况.对两种常绿灌木的研究结果表明, 具有叶片吸水功能的植物种经历长期干旱后, 凝结水的输入使叶片组织相对含水量和小枝水势均有大幅提高, 水分亏缺状况得到缓解, 而不具吸水能力的植物种持续受到干旱胁迫影响(Munné-Bosch & Alegre, 1999).在云雾林中的观测发现, 利用氢氧同位素示踪计算得到的凝结水对欧洲云杉(Picea abies)水分来源的贡献率可达31% (Berry et al., 2014), 吸收凝结水产生的茎流逆流占旱季蒸腾量的9% (Gotsch et al., 2014).长期凝结水添加可显著提高供试植物的株高、生物量(庄艳丽和赵文智, 2010; Zhuang & Ratchiffe, 2012; Liu et al., 2020).已有研究发现, 荒漠区植物普遍具有叶片吸水潜力(郑新军等, 2011; 王飞等, 2020; 杨利贞等, 2020), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
民勤荒漠植物叶片水分吸收性状研究 1 2020
... 自Hales (1727)首次提出叶片吸水以来, 已经在200多种植物、6个主要生态系统中发现此种现象(Berry et al., 2019).作为一种广泛存在的功能, 叶片吸水可显著改善植物水分状况.对两种常绿灌木的研究结果表明, 具有叶片吸水功能的植物种经历长期干旱后, 凝结水的输入使叶片组织相对含水量和小枝水势均有大幅提高, 水分亏缺状况得到缓解, 而不具吸水能力的植物种持续受到干旱胁迫影响(Munné-Bosch & Alegre, 1999).在云雾林中的观测发现, 利用氢氧同位素示踪计算得到的凝结水对欧洲云杉(Picea abies)水分来源的贡献率可达31% (Berry et al., 2014), 吸收凝结水产生的茎流逆流占旱季蒸腾量的9% (Gotsch et al., 2014).长期凝结水添加可显著提高供试植物的株高、生物量(庄艳丽和赵文智, 2010; Zhuang & Ratchiffe, 2012; Liu et al., 2020).已有研究发现, 荒漠区植物普遍具有叶片吸水潜力(郑新军等, 2011; 王飞等, 2020; 杨利贞等, 2020), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
Directional water-transfer through fabrics induced by asymmetric wettability 1 2010
... 角质层也被认为是叶片吸水的主要途径(Yates & Hutley, 1995; Goldsmith et al., 2013).角质层由带有羟基和羧基的长碳链脂肪酸聚合物组成, 其中的分子间隙及分子上的羟基、羧基亲水基团可使水渗透进入叶内.在本研究中, 荧光剂分布于黑沙蒿表面和北沙柳叶片两面.此外, 荧光示踪显示, 北沙柳叶片背面的角质层比腹面富集了更多的荧光剂(图6F), 这可能是腹面的叶毛簇具有一定的斥水性.尽管一些植物的叶毛具有亲水性, 能够促进叶面浸润乃至直接吸收水分(Schönherr, 2006; Wang et al., 2010), 但叶毛的性质和密度会影响叶面的润湿性, 当叶毛密度大于25 Ind.·mm-2时, 叶面具有很强的斥水性(Brewer et al., 1991).北沙柳腹面密布的叶毛具有遮蔽气孔、阻止蒸发的功能, 但也降低了叶面润湿性和气孔导度, 从而降低了腹面的吸水能力. ...
Leaf epidermal water-absorbing scales and their absorption of unsaturated atmospheric water in Reaumuria soongorica, a desert plant from the northwest arid region of China 1 2016a
... 植物叶片具有多种吸水结构, 露水可通过叶毛(Eller et al., 2016)、角质层(Goldsmith et al., 2013; Eller et al., 2016)、气孔(Kim & Lee, 2011)、水孔(叶片尖端或边缘的排水结构)(Martin & von Willert, 2000)等结构进入叶片, 并经过叶肉组织向维管束运移(Kim & Lee, 2011; Eller et al., 2016).由于气孔的水力导度远大于其他结构, 一般认为气孔是主要的吸水通道; 而当夜间气孔关闭时, 叶片吸水可能主要通过角质层(Eller et al., 2016).沙生植物的叶片通常具有特殊的耐旱特征, 如下陷的气孔和致密的角质层, 但目前并不清楚这些叶片特性如何影响吸水.此外, 一些沙生灌木如红砂(Reaumuria soongarica), 具有特定的吸水结构(Wang et al., 2016a), 但其他沙生植物能否通过特殊结构吸收水分并不清楚. ...
An ultrasonic humidification fluorescent tracing method for detecting unsaturated atmospheric water absorption by the aerial parts of desert plants 1 2016b
... 荧光标记凝结水浸润后, 黑沙蒿和北沙柳叶片的气孔和角质层均有示踪剂聚集(图5, 图6).叶片的气孔吸水现象已在番茄(Solanum lycopersicum)、梭梭(Haloxylon ammodendron)等植物中发现(Burkhardt et al., 2012; Wang et al., 2016b).当气孔被水分浸润时, 凝结水中的盐离子、气溶胶会降低水面张力, 促进液态水通过气孔(Burkhardt et al., 2001, 2012).由于气孔导度占叶面导度的95%以上, 一般认为白天气孔未关闭时, 气孔是叶片吸水的主要通道(Riederer & Schreiber, 2001; Berry et al., 2019).本项研究模拟了夜间露水浸润, 发现叶片吸水仍存在明显的气孔途径.近些年来的研究发现, 尽管气孔在夜晚部分关闭, 但仍能保持15%的气孔导度(Caird et al., 2007), 一些荒漠植物甚至可达30% (Snyder et al., 2003), 黑沙蒿也存在夜间气孔开放的现象(陈栋等, 2015).夜间气孔开放可以帮助植物补充养分和缓解栓塞(Caird et al., 2007), 也间接促进了叶片吸水. ...
Spectral analysis software improves confidence in plant and soil water stable isotope analyses performed by isotope ratio infrared spectroscopy (IRIS) 1 2011
... 式中, Rsample和Rstandard分别表示植物组织水和标准水(标准平均海水, SMOW)的同位素比值.δD的测量精度为±0.3‰.鉴于样品中残留的有机污染物会干扰同位素分析(Schultz et al., 2011), 使用基于同位素比值红外光谱法的LWIA光谱污染识别软件来校正δD (West et al., 2011). ...
Molecular mechanisms of foliar water uptake in a desert tree 1 2015
... 自Hales (1727)首次提出叶片吸水以来, 已经在200多种植物、6个主要生态系统中发现此种现象(Berry et al., 2019).作为一种广泛存在的功能, 叶片吸水可显著改善植物水分状况.对两种常绿灌木的研究结果表明, 具有叶片吸水功能的植物种经历长期干旱后, 凝结水的输入使叶片组织相对含水量和小枝水势均有大幅提高, 水分亏缺状况得到缓解, 而不具吸水能力的植物种持续受到干旱胁迫影响(Munné-Bosch & Alegre, 1999).在云雾林中的观测发现, 利用氢氧同位素示踪计算得到的凝结水对欧洲云杉(Picea abies)水分来源的贡献率可达31% (Berry et al., 2014), 吸收凝结水产生的茎流逆流占旱季蒸腾量的9% (Gotsch et al., 2014).长期凝结水添加可显著提高供试植物的株高、生物量(庄艳丽和赵文智, 2010; Zhuang & Ratchiffe, 2012; Liu et al., 2020).已有研究发现, 荒漠区植物普遍具有叶片吸水潜力(郑新军等, 2011; 王飞等, 2020; 杨利贞等, 2020), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
Relationship between dew presence and Bassia dasyphylla plant growth 2 2012
... 水分是荒漠生态系统最主要的环境限制要素.凝结水是干旱半干旱区重要的补充性水源, 发挥着不可忽视的作用(Zhang et al., 2009; Hao et al., 2012).在有些地区, 如纳米布沙漠, 凝结水量可超过年降水量(Mitchell et al., 2020).而在以色列内盖夫沙漠、美国内华达沙漠、中国毛乌素沙地, 凝结水量可达年降水量的7.2%-16.2% (Zangvil, 1996; Malek et al., 1999; 郭晓楠, 2017).在凝结水事件中, 土壤表层水分可以得到一定改善(刘文杰等, 2001), 叶片温度及气孔导度降低, 致使植物蒸腾耗水减少(Zhuang & Ratchiffe, 2012; 李鹭辰等, 2021), 叶片还可以直接吸收利用叶片表面的凝结水. ...
... 自Hales (1727)首次提出叶片吸水以来, 已经在200多种植物、6个主要生态系统中发现此种现象(Berry et al., 2019).作为一种广泛存在的功能, 叶片吸水可显著改善植物水分状况.对两种常绿灌木的研究结果表明, 具有叶片吸水功能的植物种经历长期干旱后, 凝结水的输入使叶片组织相对含水量和小枝水势均有大幅提高, 水分亏缺状况得到缓解, 而不具吸水能力的植物种持续受到干旱胁迫影响(Munné-Bosch & Alegre, 1999).在云雾林中的观测发现, 利用氢氧同位素示踪计算得到的凝结水对欧洲云杉(Picea abies)水分来源的贡献率可达31% (Berry et al., 2014), 吸收凝结水产生的茎流逆流占旱季蒸腾量的9% (Gotsch et al., 2014).长期凝结水添加可显著提高供试植物的株高、生物量(庄艳丽和赵文智, 2010; Zhuang & Ratchiffe, 2012; Liu et al., 2020).已有研究发现, 荒漠区植物普遍具有叶片吸水潜力(郑新军等, 2011; 王飞等, 2020; 杨利贞等, 2020), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
荒漠植物雾冰藜和沙米叶片对凝结水响应的模拟实验 2 2010
... 自Hales (1727)首次提出叶片吸水以来, 已经在200多种植物、6个主要生态系统中发现此种现象(Berry et al., 2019).作为一种广泛存在的功能, 叶片吸水可显著改善植物水分状况.对两种常绿灌木的研究结果表明, 具有叶片吸水功能的植物种经历长期干旱后, 凝结水的输入使叶片组织相对含水量和小枝水势均有大幅提高, 水分亏缺状况得到缓解, 而不具吸水能力的植物种持续受到干旱胁迫影响(Munné-Bosch & Alegre, 1999).在云雾林中的观测发现, 利用氢氧同位素示踪计算得到的凝结水对欧洲云杉(Picea abies)水分来源的贡献率可达31% (Berry et al., 2014), 吸收凝结水产生的茎流逆流占旱季蒸腾量的9% (Gotsch et al., 2014).长期凝结水添加可显著提高供试植物的株高、生物量(庄艳丽和赵文智, 2010; Zhuang & Ratchiffe, 2012; Liu et al., 2020).已有研究发现, 荒漠区植物普遍具有叶片吸水潜力(郑新军等, 2011; 王飞等, 2020; 杨利贞等, 2020), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
... ), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
荒漠植物雾冰藜和沙米叶片对凝结水响应的模拟实验 2 2010
... 自Hales (1727)首次提出叶片吸水以来, 已经在200多种植物、6个主要生态系统中发现此种现象(Berry et al., 2019).作为一种广泛存在的功能, 叶片吸水可显著改善植物水分状况.对两种常绿灌木的研究结果表明, 具有叶片吸水功能的植物种经历长期干旱后, 凝结水的输入使叶片组织相对含水量和小枝水势均有大幅提高, 水分亏缺状况得到缓解, 而不具吸水能力的植物种持续受到干旱胁迫影响(Munné-Bosch & Alegre, 1999).在云雾林中的观测发现, 利用氢氧同位素示踪计算得到的凝结水对欧洲云杉(Picea abies)水分来源的贡献率可达31% (Berry et al., 2014), 吸收凝结水产生的茎流逆流占旱季蒸腾量的9% (Gotsch et al., 2014).长期凝结水添加可显著提高供试植物的株高、生物量(庄艳丽和赵文智, 2010; Zhuang & Ratchiffe, 2012; Liu et al., 2020).已有研究发现, 荒漠区植物普遍具有叶片吸水潜力(郑新军等, 2011; 王飞等, 2020; 杨利贞等, 2020), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
... ), 巴丹吉林沙漠、腾格里沙漠、库布齐沙漠的部分沙生植物叶片具有吸水功能, 并通过吸收大气凝结水, 改善自身水分状况, 维持生理生态功能(庄艳丽和赵文智, 2010; Yan et al., 2015; Gong et al., 2019).然而, 目前并不清楚毛乌素沙地两种典型的沙生灌木黑沙蒿(又名油蒿)(Artemisia ordosica)和北沙柳(Salix psammophila)是否具有此种功能. ...
 ,1,2,*
,1,2,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT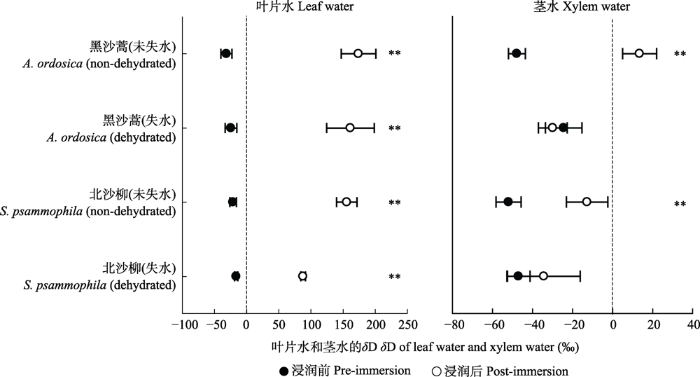 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT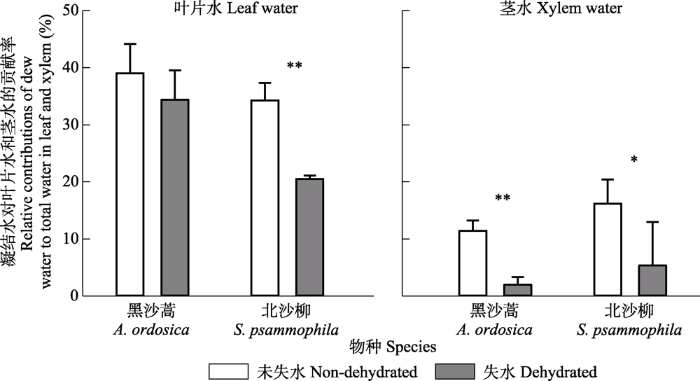 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT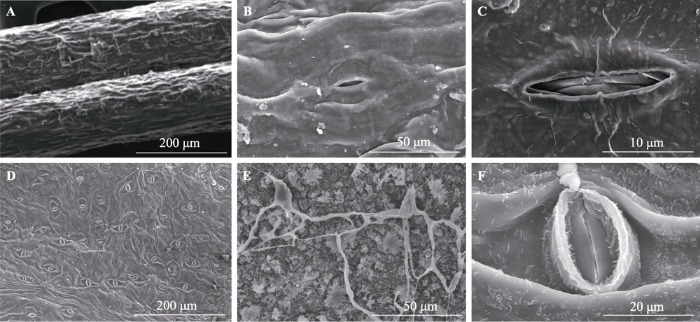 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT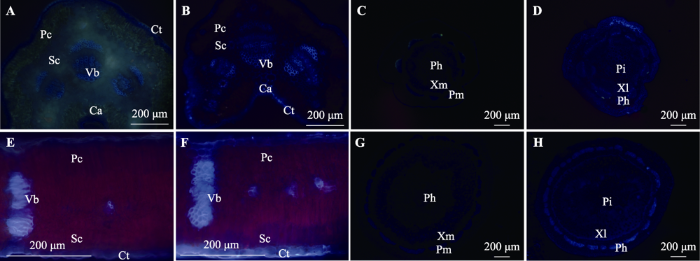 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT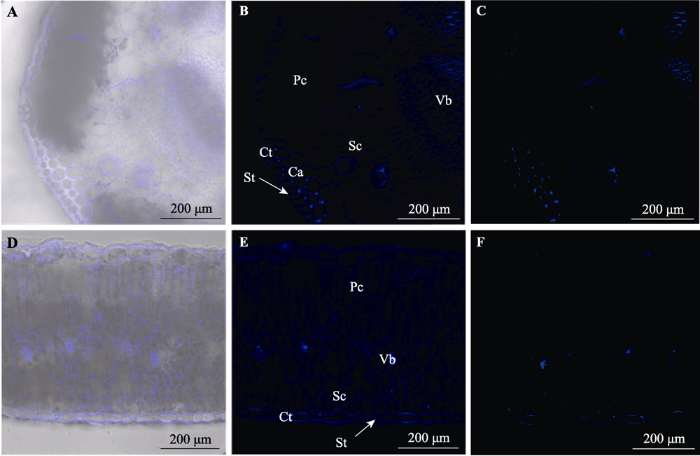 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}