王奕丹
1 , 2 , 李亮
2 , 3 , 刘琪璟
, 1 , * , 马泽清
, 2 , 3 , * 1 北京林业大学林学院, 北京 100083 2 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101 3 中国科学院千烟洲亚热带森林生态系统观测研究站, 江西吉安 343700 Lifespan and morphological traits of absorptive fine roots across six typical tree species in subtropical China Yi-Dan WANG
1 , 2 , Liang LI
2 , 3 , Qi-Jing LIU
, 1 , * , Ze-Qing MA
, 2 , 3 , * 1 College of Forestry, Beijing Forestry University, Beijing 100083, China 2 Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Science and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China 3 Qianyanzhou Ecological Research Station, Chinese Academy of Sciences, Ji’an, Jiangxi 343700, China 通讯作者: *Ma ZQ:
mazq@igsnrr.ac.cn ;Liu QJ:
liuqijing@bjfu.edu.cn 编委: 白文明
责任编辑: 李敏
收稿日期: 2021-01-4
接受日期: 2021-03-8
网络出版日期: 2021-04-20
基金资助: 国家自然科学基金 (31822010 )国家自然科学基金 (31971633 )中国科学院基础前沿科学研究计划“从0到1”原始创新项目 (ZDBS-LY-DQC023 )
Corresponding authors: *Ma ZQ:
mazq@igsnrr.ac.cn ;Liu QJ:
liuqijing@bjfu.edu.cn Received: 2021-01-4
Accepted: 2021-03-8
Online: 2021-04-20
Fund supported: National Natural Science Foundation of China (31822010 )National Natural Science Foundation of China (31971633 )“0-1” Original Innovation Project of the Chinese Academy of Sciences (ZDBS-LY-DQC023 )
摘要 根周转是地下生态过程的主要驱动力, 根属性指征了物种生态策略, 根寿命与属性是理解生态系统碳氮循环和群落多样性的关键。目前对亚热带常绿阔叶林根周转等生态过程的直接观测资料缺乏。该研究对中亚热带江西樟树试验林场6个树种吸收细根动态进行了2年观测, 获取了2.8万张微根管照片, 分析了吸收细根寿命年际和季节变化特征及其与根形态属性的关系。结果显示: 1)亚热带6个树种间吸收细根寿命变异为4.6倍, 变异系数可达73%。中值寿命排序为: 红豆杉(
Taxus wallichiana )(426天) >复羽叶栾树(
Koelreuteria bipinnata )(155天) >竹柏(
Nageia nagi )(145天) >樟(
Cinnamomum camphora )(126天) >东京樱花(
Cerasus yedoensis )(93天) >深山含笑(
Michelia maudiae )(92天); 2)树木吸收细根寿命年际、季节变异较大, 可能是适应伏秋旱、雨热不同期、年际变化大的亚热带季风气候的结果; 3)吸收细根寿命与直径呈显著正相关关系, 与比根长呈显著负相关关系, 表明根的构建成本可以在一定程度上预测寿命。这些结果为预测亚热带地下生态过程、揭示亚热带常绿阔叶林碳氮循环、物种共存机制提供依据。
关键词: 细根 ;
吸收根 ;
根周转 ;
根生产 ;
根死亡 ;
地下生态过程 ;
形态属性 ;
净初级生产力 ;
土壤有机碳 Abstract Aims Root turnover is a primary driver of belowground ecological processes, and root functional traits can indicate species ecological strategies, hence root lifespan and morphological traits are essential for understanding ecosystem carbon and nitrogen cycling as well as community diversity. Yet, data on root ecological processes in subtropical evergreen forest is very rare.
Methods We observed root dynamics of six tree species across root orders for two years in an experimental forest farm in Zhangshu, Jiangxi Province. Based on 28 000 minirhizotron photos, we analyzed interannual and seasonal changes of absorptive fine roots in relation to both lifespan and morphology.
Important findings 1) The variation of root lifespan among the six species in subtropical forest was as high as 4.6-fold, the variation of coefficient was 73%, with median lifespan in the sequence of:
Taxus wallichiana (426 d) >
Koelreuteria bipinnata (155 d) >
Nageia nagi (145 d) >
Cinnamomum camphora (126 d) >
Cerasus yedoensis (93 d) >
Michelia maudiae (92 d); 2) Absorptive fine root lifespan appeared remarkable in both seasonal and interannual variations, a pattern seemingly related to the monsoon climate which is characterized by summer-to- autumn drought and the supplies of soil water resources; 3) The lifespan of absorptive roots was positively associated with diameter, but negatively correlated with specific root length, suggesting that the root construction cost is a key predictor of lifespan. These results provide parameters for modeling belowground carbon and nitrogen cycling processes in subtropical evergreen broadleaf forest, and pave the way for exploring species coexistence mechanisms from belowground.
Keywords: fine root ;
absorptive root ;
root turnover ;
root productivity ;
root mortality ;
belowground ecological process ;
morphological trait ;
net primary productivity ;
soil organic carbon PDF (4770KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 王奕丹, 李亮, 刘琪璟, 马泽清. 亚热带6个典型树种吸收细根寿命与形态属性格局. 植物生态学报, 2021, 45(4): 383-393. DOI:
Key words WANG Yi-Dan, LI Liang, LIU Qi-Jing, MA Ze-Qing.
Lifespan and morphological traits of absorptive fine roots across six typical tree species in subtropical China .
Chinese Journal of Plant Ecology , 2021, 45(4): 383-393. DOI:
Key words 植物通过根系的生长、死亡、分解等过程驱动土壤碳氮养分循环, 但准确定量吸收细根周转一直是陆地生态系统研究中的“瓶颈” (
Jacksonet al ., 1997 ;
Norbyet al ., 2004 ;
Guoet al ., 2008a )。根系通常占植物总生物量的63% (
Poorteret al ., 2012 ), 约占全球净初级生产力的22% (
McCormacket al ., 2015 )。由于细根中氮磷含量高(
Gordon & Jackson, 2000 ), 细根死亡后迅速分解, 将养分归还土壤(
Fan & Guo, 2010 ), 将碳以CO
2 的形式释放回大气, 因此吸收细根周转驱动的碳氮循环一直作为陆地生态系统的核心内容来研究(
Warrenet al ., 2015 )。
由于直接观测根系较为困难, 根寿命的研究具有较大的不确定性, 主要表现在3个方面(
Guoet al ., 2008a ;
Huo & Cheng, 2019 )。一是方法的不确定性。目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率。二是根系观测单元的不确定性(
Guoet al ., 2008b )。树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 )。并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b )。三是根系观测周期的影响。目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 )。此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 )。由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 )。
我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 )。亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 )。由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 )。例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说。
为了更好地评估亚热带森林地下生态过程, 发展全球吸收细根数据库, 为生态模型提供参数, 基于2年微根管监测的2.8万张照片数据, 我们对6个代表性树种的根寿命与形态属性进行了研究, 主要目的: 1)揭示6个亚热带典型树种吸收细根的寿命变化格局; 2)探讨这6个树种吸收细根(前两级)形态属性(直径、比根长、组织密度及比根表面积)、根级、出生年份、季节与根寿命之间的关系, 更好地理解根寿命的驱动机制。
1 材料和方法 1.1 研究区域概况 江西省樟树市试验林场是根系生物学与生态过程观测研究网络(RhizoNet)的核心站点之一。研究样地设置在樟树市吴城乡光华山段的试验林场(中心坐标: 28.06° N, 115.23° E), 该林场的林木生活在相同气候、土壤基质条件下, 地形相对平坦、经营管理措施相当。研究区域属于中亚热带季风气候, 具有气候温润, 伏秋旱、雨热不同期、年际变化大, 霜期较短, 日照充足的特点。年平均气温19.2 ℃, 7月平均气温29.7 ℃, 1月平均气温7.5 ℃; 极端最高日平均气温34.9 ℃, 极端最低日平均气温-1.2 ℃; 大于等于5 ℃积温6 992.0 ℃, 大于等于10 ℃积温6 537.5 ℃。年降水量2 019 mm, 多集中在3-6月, 约占全年降水量的50%。年日照时间为1 718 h, 无霜期273天左右。研究设计以同质园观测为理念, 以幼龄林为主, 于1999、2007、2008及2010年分别用苗龄3年的樟(
Cinnamomum camphora ), 苗龄3年的复羽叶栾树(
Koelreuteria bipinnata )、东京樱花(
Cerasus yedoensis )和深山含笑(
Michelia maudiae ), 苗龄5年的竹柏(
Nageia nagi )及苗龄4年的红豆杉(
Taxus wallichiana )的实生苗进行造林实验, 造林面积共32.3 hm
2 ; 造林株行距为2 m × 3 m。
1.2 研究方法 1.2.1 样地设置与微根管安装
综合考虑林场内样地树种分布、亚热带常绿阔叶林建群科属种等信息, 选取了6个树种作为研究对象: 红豆杉、复羽叶栾树、竹柏、樟、东京樱花及深山含笑, 设置观测样地(
图1A )。于2014年6月在每个树种所在样地安装透明丙烯酸塑料微根管, 共8个重复, 以保证图像采集代表性和完整性。所安装的微根管内径为5 cm, 观测的垂直深度为45 cm, 将微根管安装在距离观测样木根茎部30 cm的地方, 与地面夹角为30° (
图1B )。每根微根管的地上裸露部分都加盖密封并缠绕黑色绝缘胶带, 以避免光线穿透。胶带外再覆盖一个铝罐, 喷涂成白色, 最大限度降低太阳辐射对管体的加热及颗粒物的进入。微根管周围地表铺盖与周边类似的覆盖物, 减少填埋等扰动对土壤造成的影响。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1江西省樟树市亚热带典型树种吸收细根观测系统。A, 样地位点分布。B, 微根管观测装置。 Fig. 1Observation system of absorptive root processes for subtropical tree species in Zhangshu, Jiangxi Province. A, Spatial distribution of the sampling sites. B, Illustration of minirhizotron. 1.2.2 图像数据采集
微根管装置安装后自然放置1年, 使土壤微环境及细根恢复至安装前的状态。于2015年5月开始至2016年11月对其进行连续19个月包含两个生长季的根系寿命监测。使用Bartz数码相机和I-CAP 4.01软件对根系图像进行采集, 每次采集的位点相同。每月上旬采集一次图片, 共包含6个样地, 共采集到28 063幅监测图像, 建立了根系寿命图片数据库。对采集的图像利用根系分析软件Rootfly进行处理, 对图像中出现的根的出生、死亡、直径、长度等进行测定。
1.3 数据处理 1.3.1 根系寿命数据提取
利用Rootfly软件对采集到的微根管图像进行分析, 以在图片中首次出现的浅色(白色或棕色)根为活根, 当该根从同位置观察画面消失或者颜色变为深色(黑色或深褐色)或表面皱缩则表示该根死亡; 将观察到的一级及二级根定义为吸收细根(
McCormacket al ., 2015 )。把吸收细根从第一次出现到最后一次出现的时间间隔称为吸收细根寿命。本研究将出生状态被观测到但由于实验结束或其他因素影响导致死亡状态未被观察到的细根定义为删失细根(
McCormacket al ., 2012 )。因此在表达细根寿命时选用平均寿命与中值寿命两个指标, 以提高细根寿命数据的可靠性(
Wells et al ., 2002 )。
1.3.2 根形态属性测定
根据
Pregitzer等(2002) 和
Guo等(2008a) 所描述的分级方法, 将采集到的根系样本分根处理后, 按根级置于培养皿中, 避免交叉重叠, 利用Epson数字化扫描仪进行扫描, 使用WinRHIZO软件对扫描图像进行处理, 获取平均根系直径、总根长、总根体积及总根表面积等数据, 并结合根样本干质量获得比根长(
SRL )、根组织密度(
RTD )及比根表面积(
SRA )等关键的形态属性指标。
SRL 的计算方法为根总长度除以其干质量。
RTD 是根干质量与根体积的
比值。
SRA 定义为总表面积除以其干质量。
1.3.3 数据分析
吸收细根的生存分析主要采用统计软件Origin 2019b中的Kaplan-Meier法, 即直接用概率乘法原理估计生存率, 得到研究树种吸收细根的平均寿命、中值寿命及生存曲线。对于不同树种及不同出生年份、季节与根级的根生存曲线差异采用对数秩检验比较; 种间的吸收细根比较时, 混合前两级根寿命数据。对于不同树种根寿命与形态属性的相关性, 在R 3.5.1软件中采用线性回归方法分析和作图。统计检验显著性水平未特别说明时, 均为
p < 0.05。
2 结果 2.1 吸收细根寿命及形态属性 亚热带6个典型树种的吸收细根的中值寿命变异是4.6倍(92-426天), 物种间差异显著(
p < 0.05), 变异系数达73%。红豆杉的中值寿命最长, 为426天, 深山含笑中值寿命最短, 为92天(
表1 )。平均寿命与中值寿命略有不同, 变异是3.3倍, 变异系数为48% (126-417天), 最长仍为红豆杉, 为417天, 最短是东京樱花, 为126天(
表1 )。本研究追踪了在6个样地内3 556个所观察到吸收细根, 其中复羽叶栾树样地内观察到的细根数量最多, 为1 745个, 占总数的49.1%, 红豆杉仅有31个, 占总数的0.9%。
Table 1 表1 表1 亚热带6个典型树种吸收细根寿命格局(平均值±标准误)
Table 1
Absorptive fine root lifespan across six common subtropical tree species (mean ± SE ) 树种 中值寿命 95%置信区间 平均寿命 观测细根数 删失细根数 LCL UCL 红豆杉 Taxus wallichiana 426 ± 103 d 242 426 417 ± 21 d 31 17 复羽叶栾树 Koelreuteria bipinnata 155 ± 35 c 153 175 216 ± 4 bc 1 745 488 竹柏 Nageia nagi 145 ± 20 bc 128 154 178 ± 7 b 436 102 樟 Cinnamomum camphora 126 ± 11 ab 105 188 179 ± 12 b 128 54 东京樱花 Cerasus yedoensis 93 ± 7 a 93 96 126 ± 3 a 772 155 深山含笑 Michelia maudiae 92 ± 63 ab 92 120 180 ± 9 b 444 192
95% LCL (置信区间下限)和95% UCL (置信区间上限)指中值寿命的95%置信区间下限与上限。“删失细根数”指在观测结束时未观测到根死亡状态或其他影响导致根死亡状态无法观测的根数。同列不同小写字母表示差异显著(p < 0.05, Duncan多重比较)。 95% LCL and 95% UCL refer to the lower and upper confidence interval for root median lifespan, respectively. “Number censored” refers to roots for which death events were not observed by the time the experiment is terminated, or due to other reasons that death events cannot be determined. Different lowercase letters indicate significant differences (p < 0.05, Duncan multiple comparison). 新窗口打开 |
下载CSV 不同树种形态属性差异较大, 根级与直径及根组织密度均呈正比关系, 与比根长及比根表面积呈反比关系(
表2 )。树种间根直径在不同根级间具有显著差异(
p < 0.05), 随着根级增加而增加。一级根直径变异为2倍, 变异系数为25% (0.28-0.57 mm); 二级根直径变异为1.9倍, 变异系数为27% (0.61-1.14 mm)。一级根比根长变异为1.9倍(19.06-35.41 m·g
-1 ), 变异系数是21%; 二级根的比根长变异为2.7倍(3.43- 9.26 m·g
-1 ), 变异系数是30%。一级根的根组织密度变异为2.2倍(0.21-0.47 g·cm
-3 ), 二级根的根组织密度变异为1.8倍(0.29-0.52 g·cm
-3 )。一级根的比根表面积变异为1.4倍(275.3-397.2 cm
2 ·g
-1 ), 变异系数为12%, 二级根的比根表面积变异为1.9倍(110.5- 205.6 cm
2 ·g
-1 ), 变异系数为24%。
Table 2 表2 表2 亚热带6个典型树种吸收细根形态属性特征(平均值±标准误)
Table 2
Morphological traits of absorptive fine root across six common subtropical tree species (mean ± SE ) 树种 Tree species 根级 Root order 直径 Diam (mm) 比根长 SRL (m·g-1 ) 根组织密度 RTD (g·cm-3 ) 比根表面积 SRA (cm2 ·g-1 ) 红豆杉 Taxus wallichiana 1 0.57 ± 0.03 Aa 19.06 ± 0.81 Aa 0.21 ± 0.02 Aa 340.61 ± 27.04 Aa 2 1.14 ± 0.06 Ab 3.43 ± 0.10 Ab 0.29 ± 0.03 Aab 123.26 ± 9.52 Ab 复羽叶栾树 Koelreuteria bipinnata 1 0.33 ± 0.00 Aa 26.62 ± 5.94 Aa 0.47 ± 0.11 Ba 275.29 ± 61.28 Aa 2 0.61 ± 0.03 Ab 7.18 ± 0.11 Ab 0.49 ± 0.05 Bab 137.74 ± 9.50 Ab 竹柏 Nageia nagi 1 0.45 ± 0.00 Aa 23.15 ± 0.27 Aa 0.27 ± 0.00 ABa 328.68 ± 3.77 Aa 2 0.66 ± 0.02 Ab 7.27 ± 0.49 Ab 0.41 ± 0.04 ABab 151.14 ± 11.97 Ab 樟 Cinnamomum camphora 1 0.40 ± 0.01 Aa 25.61 ± 0.25 Aa 0.31 ± 0.01 ABa 323.48 ± 7.77 Aa 2 0.63 ± 0.05 Ab 8.92 ± 1.08 Ab 0.40 ± 0.09 ABab 178.26 ± 35.13 Ab 东京樱花 Cerasus yedoensis 1 0.28 ± 0.01 Aa 35.41 ± 1.39 Aa 0.45 ± 0.04 Ba 317.36 ± 19.73 Aa 2 0.71 ± 0.05 Ab 4.92 ± 0.10 Ab 0.52 ± 0.07 Bab 110.46 ± 9.41 Ab 深山含笑 Michelia maudiae 1 0.45 ± 0.00 Aa 28.26 ± 0.64 Aa 0.23 ± 0.00 Aa 397.19 ± 8.56 Aa 2 0.70 ± 0.05 Ab 9.26 ± 0.16 Ab 0.27 ± 0.05 Aab 205.60 ± 18.67 Ab
不同上标字母表示Duncan多重比较存在显著差异(p < 0.05), 大写字母表示种间差异, 小写字母表示根级间差异。 Diam, diameter; SRL, specific root length; RTD, root tissue density; SRA, specific root area. Different superscript letters denote significant differences based on Duncan multiple comparison (p < 0.05). Uppercase letters indicate differences between species, while lowercase letters between root orders. 新窗口打开 |
下载CSV 2.2 树种间吸收细根寿命的差异 树种的吸收细根中值寿命分布在92-426天, 表现为: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 )。6个树种的分析表明不同生活型的吸收细根寿命没有系统性的差异。例如, 复羽叶栾树与东京樱花同为落叶阔叶树种, 但复羽叶栾树吸收细根中值寿命(155天)高于常绿针叶树种竹柏(145天)而东京樱花(93天)低于竹柏。樟与深山含笑同属常绿阔叶树种, 但深山含笑吸收细根中值寿命(92天)低于落叶阔叶树种东京樱花而樟(126天)高于东京樱花。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2亚热带6个典型树种吸收细根生存曲线。 Fig. 2Survival curves of absorptive fine root across six common subtropical tree species. 2.3 出生年份、季节与根级对吸收细根寿命的影响 出生于2015年的吸收细根中值寿命(152天)显著高于2016年的(124天)(
p < 0.05)(
图3A )。约40%的根出现于夏季, 只有不到5%的根出现于冬季。春季出现的吸收细根中值寿命为93天, 显著低于其他季节。夏、秋季与冬季出现的吸收细根中值寿命分别为155、153、154天, 差异并不显著。但总体来看, 出现于春夏的吸收细根寿命明显长于秋冬(
图3B )。根级对于吸收细根寿命有显著影响(
p < 0.05), 一级根中值寿命(127天)显著低于二级根(278天)(
图3C )。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3亚热带6个典型树种吸收细根不同出生年份(A)、季节(B)及根级(C)的生存曲线。 Fig. 3Annual (A), seasonal (B) absorptive fine root processes and survival probability of different root order (C) across six common subtropical tree species. 2.4 吸收细根寿命与形态属性相关性分析 吸收细根直径与寿命显著正相关(
p < 0.01), 这种关系主要是由不同根级来驱动(
图4A , 黑线); 6个树种的一级根之间有相关但统计不显著(
图4A )。吸收细根比根长与寿命显著负相关(
p < 0.05)(
图4B ); 6个树种的一级根内有相关但二级根无相关关系(
图4B , 蓝点)。吸收细根根组织密度与寿命无相关关系(
p > 0.05)(
图4C ); 6个种的一级根(或二级根)有相关趋势(
R 2 > 0.24), 但统计上不显著(
p > 0.05)(
图4C )。吸收细根比根表面积与细根寿命无相关关系(
R 2 < 0.22,
p > 0.05)(
图4D )。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4亚热带6个典型树种吸收细根中值寿命与根形态属性的关系。ns, 不显著; *, p < 0.05; **, p < 0.01。 Fig. 4Relations between morphological traits and median lifespan of absorptive fine root across six common subtropical tree species. ns, not significant; *, p < 0.05; **, p < 0.01. 2.5 吸收细根寿命受不同因素影响程度分析 出生季节对复羽叶栾树、竹柏、樟及深山含笑的吸收细根寿命具有显著影响(
p < 0.05), 竹柏、樟及深山含笑的吸收细根寿命受根级影响显著(
p < 0.05), 而出生年份仅对红豆杉及竹柏有显著影响(
p < 0.05)(
表3 )。与冬季相比, 复羽叶栾树生于春季与夏季的根寿命相对会延长36%与58%, 樟生于春季的吸收细根死亡危险率降低了95%, 深山含笑生于夏季的吸收细根寿命相对延长了59%。根级对于竹柏的吸收细根寿命影响最大, 二根级寿命相对延长71%。与2016年相比, 出生在2015年的红豆杉吸收细根寿命相对延长97%。
Table 3 表3 表3 3种因素对亚热带不同树种吸收细根寿命的Cox比例风险回归分析
Table 3
Cox analysis for absorptive fine roots lifespan and its drivers across different subtropical tree species 影响因素 红豆杉 Taxus wallichiana 复羽叶栾树 Koelreuteria bipinnata 竹柏 Nageia nagi 樟 Cinnamomum camphora 东京樱花 Cerasus yedoensis 深山含笑 Michelia maudiae p Exp(B) p Exp(B) p Exp(B) p Exp(B) p Exp(B) p Exp(B) 出生 春季 Spring 0.157 0.378 0.001 0.639 0.792 1.055 0.013 0.046 0.152 0.478 0.642 0.823 夏季 Summer 0.000 0.000 0.425 0.426 0.826 0.931 0.916 0.102 0.449 0.037 0.410 秋季 Autumn 0.825 0.644 0.620 0.930 0.021 1.640 0.397 0.414 0.379 0.598 0.384 0.671 冬季 Winter 0.000 0.002 0.000 0.355 0.000 根级 Root order 0.413 0.624 0.073 1.252 0.000 0.291 0.000 9.059 0.135 1.828 0.000 3.355 出生年份 Birth year 0.017 0.029 0.083 0.860 0.001 1.917 0.614 1.188 0.755 0.921 0.922 0.986
风险函数值比值(Exp(B))代表死亡危险率。分析细根寿命的影响因素时, 参考出生季节为“冬季”, 参考根级为“order 2”, 参考出生年份为“2016年”。由于红豆杉冬季未观测到新生吸收细根, 所以红豆杉参考季节为“夏季”。表内加粗值表示p < 0.05, 达到显著水平。 Risk function value ratio (Exp(B)) represents the hazard ratio for mortality. When examining the driving factors on the lifespan of absorptive fine roots across tree species, “winter” was set as the reference season of birth, “order 2” the reference of root order, and “2016” the year of birth. Since no new absorptive fine roots were observed in Taxus chinensis in winter, “summer” was set as the reference season instead. Bold values indicate p < 0.05. 新窗口打开 |
下载CSV 3 讨论 吸收细根寿命是指根系从出生到死亡所经历的时间, 是植物重要的生理生态学特性; 吸收细根个体寿命的变异程度、频度分布的偏态程度是准确评估根系周转在生态系统碳平衡和养分循环中的重要性的先决条件。本研究通过2年的观测, 发现亚热带6种树木吸收细根中值寿命为92-426天, 粗根物种的根寿命是细根物种根寿命的4.6倍(
表1 )。随根直径等构建成本增加, 根个体数量呈线性增加(
图4A )。根寿命随根级增加, 前两级根的中值寿命差异为2倍, 可认为是短寿命吸收细根模块(
图3C ); 不同树种根寿命的年际变异大(
图3A ), 表明物种对根寿命调控起核心的作用。这些结果为理解亚热带季风气候条件的根结构、功能和资源获取策略奠定了基础, 未来需精准评估建群种、伴生种不同根级的根寿命, 提高根系周转对亚热带森林生产力和地下生态过程的贡献。
3.1 亚热带典型树种吸收细根根寿命变异格局 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 )。亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献。根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键。
3.2 树种间的吸收细根寿命具有显著差异 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 )。由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异。从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 )。常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 )。总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 )。总之, 长期的进化过程中, 物种是决定根寿命的关键因素。
亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 )。研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力。深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 )。树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 )。温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合。
此外, 在对粗根树种红豆杉的监测中获取的有效根系数量较少, 一方面是粗根物种根少且根周转较慢, 另一方面是微根管视野小且2年观测时间不够长, 意味着亚热带和热带森林根寿命的原位观测挑战大, 粗根物种要结合菌丝一起观测, 并针对性地延长监测时间、增加微根管数量和观测视野, 从而获得更多可靠的粗根树种根寿命数据。
3.3 亚热带湿润季风气候对吸收细根寿命的影响 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点。在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B)。根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 )。春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 )。在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 )。由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 )。
3.4 根级对吸收细根寿命的影响 亚热带树种的根级对吸收细根寿命有着显著的影响, 随根级增加, 根寿命延长(
图3C )。树木细根是一个高度异质的系统, 根构型是预测寿命的核心因素, 着生在根枝末端的一级根的周转率5倍于着生在根枝基部的五级根(
Xiaet al ., 2010 )。乔木的根级最大可以到10级(
Kajimoto, 2010 ), 前五级根是重点关注的单元(
Guoet al ., 2008b ), 但微根管的观测视野较小, 很难观测到全部5个根级, 针对这个缺点, 根窗法或扫描根系的方法能够全面地理解异质性的根系统, 值得推荐(
Xiaet al ., 2010 ;
Huo & Cheng, 2019 )。通常认为一级根和二级根能被共生真菌侵染且具有皮层, 划分为短寿命的吸收功能模块, 四五级根为长寿命的吸收运输根功能模块, 三级根主要为过渡; 为了方便研究, 把前三级根或前两级根作为短命吸收细根模块(
Xiaet al ., 2010 )。本研究也证实这6个树种前两级根的根寿命基本上是连续的变异, 并未出现突变的结果; 这与北方落叶松的情况有区别, 一级根和二级根的寿命相差4倍(
Huo & Cheng, 2019 ), 表明外生菌根侵染的一级根(根尖)是频繁死亡、半独立、快速周转的单元模块。在草本植物中, 根周转与物种本身的构型具有密切的关系(
Sunet al ., 2016 ;
Baiet al ., 2017 ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 )。解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进。
3.5 根构建成本与寿命正相关 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 )。如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 )。草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 )。
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%。根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 )。从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 )。只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量。本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量。
4 小结 根寿命和根属性是理解物种生态策略的关键, 是发展根经济谱理论的依据, 是理解群落多样性形成不可或缺的环节。本研究揭示了6个亚热带典型树种吸收细根寿命出生年份、季节、根级和4个关键根形态属性对其的影响机制, 为深入理解亚热带地下生态过程提供数据支持。由于研究手段限制, 目前吸收细根寿命数据依旧十分匮乏, 对根构建与寿命的权衡的认识存在较大的不确定性。直接观测的根寿命数据不到叶寿命数据的1/10, 而且数据大多来源于根偏细的温带森林, 对热带和亚热带森林特别是粗根物种(一级根直径>0.6 mm)的根寿命还了解不多。在物种丰富的群落, 很难原位确定目标物种的根寿命; 而在同质园中根寿命可能会被高估, 因为生境避免了种间竞争。总之, 树木根周转存在较大的不确定性, 仍需要更多的研究来跨物种观测根寿命, 研究根寿命的调控机制, 如构建成本、菌根类型、资源类型及供应水平、竞争、边际收益等。
[1] Abramoff RZ Finzi AC 2015 ). Are above- and below-ground phenology in sync?New Phytologist, 205, 1054 -1061 . DOI:
10.1111/nph.2015.205.issue-3 URL [本文引用: 1] [2] Aerts R 1995 ). The advantages of being evergreenTrends in Ecology & Evolution, 10, 402 -407 . DOI:
10.1016/S0169-5347(00)89156-9 URL [本文引用: 1] [3] Bai WM Zhou M Fang Y Zhang WH 2017 ). Differences in spatial and temporal root lifespan of three Stipa grasslands in northern ChinaBiogeochemistry, 293 -306 . DOI:
10.1007/s10533-017-0302-4 URL [本文引用: 1] [4] Bauhus J Messier C 1999 ). Evaluation of fine root length and diameter measurements obtained using RHIZO image analysisAgronomy Journal, 91, 142 -147 . DOI:
10.2134/agronj1999.00021962009100010022x URL [本文引用: 1] [5] Burton AJ Pregitzer KS Hendrick RL 2000 ). Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forestsOecologia, 125, 389 -399 . DOI:
10.1007/s004420000455 PMID:
28547334 [本文引用: 1] Minirhizotrons were used to observe fine root (≤1 mm) production, mortality, and longevity over 2 years in four sugar-maple-dominated northern hardwood forests located along a latitudinal temperature gradient. The sites also differed in N availability, allowing us to assess the relative influences of soil temperature and N availability in controlling fine root lifespans. Root production and mortality occurred throughout the year, with most production occurring in the early portion of the growing season (by mid-July). Mortality was distributed much more evenly throughout the year. For surface fine roots (0-10 cm deep), significant differences in root longevity existed among the sites, with median root lifespans for root cohorts produced in 1994 ranging from 405 to 540 days. Estimates of fine root turnover, based on the average of annual root production and mortality as a proportion of standing crop, ranged from 0.50 to 0.68 year for roots in the upper 30 cm of soil. The patterns across sites in root longevity and turnover did not follow the north to south temperature gradient, but rather corresponded to site differences in N availability, with longer average root lifespans and lower root turnover occurring where N availability was greater. This suggests the possibility that roots are maintained as long as the benefit (nutrients) they provide outweighs the C cost of keeping them alive. Root N concentrations and respiration rates (at a given temperature) were also higher at sites where N availability was greater. It is proposed that greater metabolic activity for roots in nitrogen-rich zones leads to greater carbohydrate allocation to those roots, and that a reduction in root C sink strength when local nutrients are depleted provides a mechanism through which root lifespan is regulated in these forests.
[6] Comas LH Bouma TJ Eissenstat DM 2002 ). Linking root traits to potential growth rate in six temperate tree speciesOecologia, 132, 34 -43 . DOI:
10.1007/s00442-002-0922-8 PMID:
28547288 [本文引用: 1] There is an extremely limited understanding of how plants of different potential growth rate vary in root traits, especially in woody species. We contrasted fine root morphology, physiology, and elemental construction between a fast- and a slow-growing species in each of three families: Aceraceae (maple), Fagaceae (oak), and Pinaceae (pine). Measurements were primarily made on 1-year-old seedlings growing in a growth chamber. Across all three families, first- and second-order roots of fast-growing species had greater specific root length, thinner diameters, and faster respiration rates than those of slow-growing species. These fine roots of fast-growing species in Aceraceae and Fagaceae also had faster phosphorus (P) uptake on a surface area basis than those of slow-growing species, whereas little difference in P uptake was found between Pinaceae species. On a dry weight basis, roots of fast-growing species in Aceraceae and Fagaceae had higher nitrogen concentrations, lower carbon:nitrogen ratios and higher tissue construction costs than roots of slow-growing species (data were unavailable for Pinaceae). Tissue density did not vary in a consistent pattern between fast- and slow-growing species across all three families (P=0.169). To better understand the ecological significance of differences in these root characteristics, a root efficiency model was used to compare P uptake and root carbon (C) cost of each species in simulated field situations in two soils, one with low P buffering capacity (loamy sand) and another with relatively high P buffering capacity (silt loam). For the soil conditions modeled, fast-growing species of Aceraceae and Fagaceae were 17-70% more efficient (defined as cumulative P gain divided by cumulative C cost) at nutrient capture than slow-growing species while the fast-growing Pinaceae species was 20-24% less efficient than the slow-growing species. However, among all three families, roots of fast-growing species reached maximum lifetime efficiency 5-120 days sooner, depending on soil type. Thus, modeling results indicated that root traits of fast- and slow-growing species affected P acquisition in simulated field soil although soil type also had a strong impact.
[7] Comas LH Callahan HS Midford PE 2014 ). Patterns in root traits of woody species hosting arbuscular and ectomycorrhizas: implications for the evolution of belowground strategiesEvolutionary and Ecology, 4, 2979 -2990 . DOI:
10.1002/ece3.2014.4.issue-15 URL [本文引用: 1] [8] Comas LH Eissenstat DM 2004 ). Linking fine root traits to maximum potential growth rate among 11 mature temperate tree speciesFunctional Ecology, 18, 388 -397 . DOI:
10.1111/fec.2004.18.issue-3 URL [本文引用: 2] [9] Courchesne DN Wilson AZ Ryser P 2020 ). Regional distribution patterns of wetland monocots with different root turnover strategies are associated with local variation in soil temperatureNew Phytologist, 226, 86 -97 . DOI:
10.1111/nph.16328 [本文引用: 1] Herbaceous perennial wetland monocots in Northern Ontario, Canada, show dichotomous root overwintering patterns, either with full senescence in autumn or survival over the winter, comparable to deciduous and evergreen leaf habits in trees. To test if these root strategies differ in their associations with growing season length, soil temperatures were recorded in autumn, winter and spring on 38 wetland sites with their dominant species being either of these two root overwintering strategies, altogether 19 monocot species. Traits associated with these strategies were assessed in garden experiments for a subset of these species. Sites with species with autumn-senescing roots were, on average, warmer than sites with species with overwintering roots. The 13 coldest sites were all sites of species with overwintering roots. Spring shoot growth in the field was delayed in species with autumn-senescing roots, despite their higher shoot relative growth rates in the garden, possibly due to the necessity to produce roots first. We conclude that species with autumn-senescing roots are more constrained by temperature than species with overwintering roots, limiting their occurrence in cool climates to locally warm soils and constraining their distribution towards the north. This is comparable to constraints on distribution of deciduous and evergreen trees.
[10] Eissenstat DM 1997 ). Trade-offs in root form and function //Jackson LE . Ecology in Agriculture. Academic Press, San Diego,USA. 173 -199 . [本文引用: 1] [11] Eissenstat DM Wells CE Yanai RD Whitbeck JL 2000 ). Building roots in a changing environment: implications for root longevityNew Phytologist, 147, 33 -42 . DOI:
10.1046/j.1469-8137.2000.00686.x URL [本文引用: 1] [12] Eissenstat DM Yanai RD 1997 ). The ecology of root lifespanAdvances in Ecological Research, 27, 1 -60 . [本文引用: 2] [13] Fan PP Guo DL 2010 ). Slow decomposition of lower order roots: a key mechanism of root carbon and nutrient retention in the soilOecologia, 163, 509 -515 . DOI:
10.1007/s00442-009-1541-4 URL [本文引用: 1] [14] Freschet GT Cornelissen JHC van Logtestijn RSP Aerts R 2010 ). Evidence of the “plant economics spectrum” in a subarctic floraJournal of Ecology, 98, 362 -373 . DOI:
10.1111/jec.2010.98.issue-2 URL [本文引用: 1] [15] Gaudinski J Trumbore S Davidson E Cook A Markewitz D Richter D 2001 ). The age of fine-root carbon in three forests of the eastern United States measured by radiocarbonOecologia, 129, 420 -429 . DOI:
10.1007/s004420100746 PMID:
28547197 [本文引用: 1] Using a new approach involving one-time measurements of radiocarbon (C) in fine (<2 mm diameter) root tissues we have directly measured the mean age of fine-root carbon. We find that the carbon making up the standing stock of fine roots in deciduous and coniferous forests of the eastern United States has a mean age of 3-18 years for live fine roots, 10-18 years for dead fine roots, and 3-18 years for mixed live+dead fine roots. These C-derived mean ages represent the time C was stored in the plant before being allocated for root growth, plus the average lifespan (for live roots), plus the average time for the root to decompose (for dead roots and mixtures). Comparison of the C content of roots known to have grown within 1 year with the C of atmospheric CO for the same period shows that root tissues are derived from recently fixed carbon, and the storage time prior to allocation is <2 years and likely <1 year. Fine-root mean ages tend to increase with depth in the soil. Live roots in the organic horizons are made of C fixed 3-8 years ago compared with 11-18 years in the mineral B horizons. The mean age of C in roots increases with root diameter and also is related to branching order. Our results differ dramatically from previous estimates of fine-root mean ages made using mass balance approaches and root-viewing cameras, which generally report life spans (mean ages for live roots) of a few months to 1-2 years. Each method for estimating fine-root dynamics, including this new radiocarbon method, has biases. Root-viewing approaches tend to emphasize more rapidly cycling roots, while radiocarbon ages tend to reflect those components that persist longest in the soil. Our C-derived estimates of long mean ages can be reconciled with faster estimates only if fine-root populations have varying rates of root mortality and decomposition. Our results indicate that a standard definition of fine roots, as those with diameters of <2 mm, is inadequate to determine the most dynamic portion of the root population. Recognition of the variability in fine-root dynamics is necessary to obtain better estimates of belowground C inputs.
[16] Gordon WS Jackson RB 2000 ). Nutrient concentrations in fine rootsEcology, 81, 275 -280 . DOI:
10.1890/0012-9658(2000)081[0275:NCIFR]2.0.CO;2 URL [本文引用: 1] [17] Gu JC Wang Y Fahey TJ Wang ZQ 2017 ). Effects of root diameter, branch order, soil depth and season of birth on fine root life span in five temperate tree speciesEuropean Journal of Forest Research, 136, 727 -738 . DOI:
10.1007/s10342-017-1068-x URL [本文引用: 1] [18] Guo DL Li H Mitchell RJ Han WX Hendricks JJ Fahey TJ Hendrick RL 2008a ). Fine root heterogeneity by branch order: exploring the discrepancy in root turnover estimates between minirhizotron and carbon isotopic methodsNew Phytologist, 177, 443 -456 . DOI:
10.1111/nph.2008.177.issue-2 URL [本文引用: 4] [19] Guo DL Xia MX Wei X Chang WJ Liu Y Wang ZQ 2008b ). Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree speciesNew Phytologist, 180, 673 -683 . DOI:
10.1111/nph.2008.180.issue-3 URL [本文引用: 3] [20] Hansson K Helmisaari HS Sah SP Lange H 2013 ). Fine root production and turnover of tree and understorey vegetation in Scots pine, silver birch and Norway spruce stands in SW SwedenForest Ecology and Management, 309, 58 -65 . DOI:
10.1016/j.foreco.2013.01.022 URL [本文引用: 1] [21] Huang JX Ling H Yang ZJ Lu ZL Xiong DC Chen GS Yang YS 2012 ). Fine root longevity and controlling factors in subtropical Altingia grlilipes and Castanopsis carlesii forests Acta Ecologica Sinica, 1932 -1942 . DOI:
10.5846/stxb URL [本文引用: 4] [ 黄锦学 , 凌华 , 杨智杰 , 卢正立 , 熊德成 , 陈光水 , 杨玉盛 ( 2012 ). 细柄阿丁枫和米槠细根寿命影响因素生态学报 , 32, 1932 -1942 .] [本文引用: 4] [22] Huo CF Cheng WX 2019 ). Improved root turnover assessment using field scanning rhizotrons with branch order analysisEcosphere, 10, e02793 . DOI: https://www.plant-ecology.com/article/2021/1005-264X/10.1002/ecs2.2793 . URL [本文引用: 6] [23] Iversen CM 2014 ). Using root form to improve our understanding of root functionNew Phytologist, 203, 707 -709 . DOI:
10.1111/nph.2014.203.issue-3 URL [本文引用: 1] [24] Jackson RB Mooney HA Schulze ED 1997 ). A global budget for fine root biomass, surface area, and nutrient contentsProceedings of the National Academy of Sciences of the United States of America, 94, 7362 -7366 . PMID:
11038557 [本文引用: 2] Global biogeochemical models have improved dramatically in the last decade in their representation of the biosphere. Although leaf area data are an important input to such models and are readily available globally, global root distributions for modeling water and nutrient uptake and carbon cycling have not been available. This analysis provides global distributions for fine root biomass, length, and surface area with depth in the soil, and global estimates of nutrient pools in fine roots. Calculated root surface area is almost always greater than leaf area, more than an order of magnitude so in grasslands. The average C:N:P ratio in living fine roots is 450:11:1, and global fine root carbon is more than 5% of all carbon contained in the atmosphere. Assuming conservatively that fine roots turn over once per year, they represent 33% of global annual net primary productivity.
[25] Kajimoto T 2010 ). Root system development of larch trees growing on siberian permafrostPermafrost Ecosystems, 209, 303 -330 . [本文引用: 1] [26] Kong DL Ma CG Zhang Q Li L Chen XY Zeng H Guo DL 2014 ). Leading dimensions in absorptive root trait variation across 96 subtropical forest speciesNew Phytologist, 203, 863 -872 . DOI:
10.1111/nph.2014.203.issue-3 URL [本文引用: 1] [27] Liljeroth E Bryngelsson T 2001 ). DNA fragmentation in cereal roots indicative of programmed root cortical cell deathPhysiologia Plantarum, 111, 365 -372 . DOI:
10.1034/j.1399-3054.2001.1110314.x URL [本文引用: 1] [28] Ling H Yuan YD Yang ZJ Huang JX Chen GS Yang YS 2011 ). Influencing factors of fine root lifespans in two Chinese fir plantations in subtropical ChinaActa Ecologica Sinica, 31, 1130 -1138 . [本文引用: 3] [ 凌华 , 袁一丁 , 杨智杰 , 黄锦学 , 陈光水 , 杨玉盛 ( 2011 ). 杉木人工林细根寿命的影响因素生态学报 , 31, 1130 -1138 .] [本文引用: 3] [29] Liu BT Li HB Zhu B Koide RT Eissenstat DM Guo DL 2015 ). Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree speciesNew Phytologist, 208, 125 -136 . DOI:
10.1111/nph.2015.208.issue-1 URL [本文引用: 1] [30] Liu XJ Trogisch S He JS Niklaus PA Bruelheide H Tang ZY Erfmeier A Scherer-Lorenzen M Pietsch KA Yang B Kühn P Scholten T Huang YY Wang C Staab M Leppert KN Wirth C Schmid B Ma KP 2018 ). Tree species richness increases ecosystem carbon storage in subtropical forestsProceedings of the Royal Society B: Biological Sciences, 285, 1240 . [本文引用: 1] [31] Ma ZQ Guo DL Xu XL Lu MZ Bardgett RD Eissenstat DM McCormack ML Hedin LO 2018 ). Evolutionary history resolves global organization of root functional traitsNature, 555, 94 -97 . DOI:
10.1038/nature25783 URL [本文引用: 3] [32] Majdi H Damm E Nylund JE 2001 ). Longevity of mycorrhizal roots depends on branching order and nutrient availabilityNew Phytologist, 150, 195 -202 . DOI:
10.1046/j.1469-8137.2001.00065.x URL [本文引用: 1] [33] McCormack ML Adams TS Smithwick EAH Eissenstat DM 2012 ). Predicting fine root lifespan from plant functional traits in temperate treesNew Phytologist, 195, 823 -831 . DOI:
10.1111/nph.2012.195.issue-4 URL [本文引用: 5] [34] McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo DL Helmisaari HS Hobbie EA Iversen CM Jackson RB Leppälammi-Kujansuu J Norby RJ Phillips RP Pregitzer KS Pritchard SG Rewald B Zadworny M 2015 ). Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processesNew Phytologist, 207, 505 -518 . DOI:
10.1111/nph.2015.207.issue-3 URL [本文引用: 2] [35] McCormack ML Guo DL 2014 ). Impacts of environmental factors on fine root lifespanFrontiers in Plant Science, 5, 205 . DOI: https://www.plant-ecology.com/article/2021/1005-264X/10.3389/fpls.2014.00205 . PMID:
24904605 [本文引用: 2] The lifespan of fast-cycling roots is a critical parameter determining a large flux of plant carbon into soil through root turnover and is a biological feature regulating the capacity of a plant to capture soil water and nutrients via root-age-related physiological processes. While the importance of root lifespan to whole-plant and ecosystem processes is increasingly recognized, robust descriptions of this dynamic process and its response to changes in climatic and edaphic factors are lacking. Here we synthesize available information and propose testable hypotheses using conceptual models to describe how changes in temperature, water, nitrogen (N), and phosphorus (P) availability impact fine root lifespan within a species. Each model is based on intrinsic responses including root physiological activity and alteration of carbohydrate allocation at the whole-plant level as well as extrinsic factors including mycorrhizal fungi and pressure from pathogens, herbivores, and other microbes. Simplifying interactions among these factors, we propose three general principles describing fine root responses to complex environmental gradients. First, increases in a factor that strongly constrains plant growth (temperature, water, N, or P) should result in increased fine root lifespan. Second, increases in a factor that exceeds plant demand or tolerance should result in decreased lifespan. Third, as multiple factors interact fine root responses should be determined by the most dominant factor controlling plant growth. Moving forward, field experiments should determine which types of species (e.g., coarse vs. fine rooted, obligate vs. facultative mycotrophs) will express greater plasticity in response to environmental gradients while ecosystem models may begin to incorporate more detailed descriptions of root lifespan and turnover. Together these efforts will improve quantitative understanding of root dynamics and help to identify areas where future research should be focused.
[36] Niinemets Ü Ostonen I 2020 ). Plant organ senescence above- and belowground in trees: How to best salvage resources for new growth?Tree Physiology, 40, 981 -986 . DOI:
10.1093/treephys/tpaa060 PMID:
32353147 [本文引用: 1] [37] Norby RJ Ledford J Reilly CD Miller NE O’Neill EG 2004 ). Fine-root production dominates response of a deciduous forest to atmospheric CO2 enrichmentProceedings of the National Academy of Sciences of the United States of America, 101, 9689 -9693 . [本文引用: 1] [38] Poorter H Niklas KJ Reich PB Oleksyn J Poot P Mommer L 2012 ). Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental controlNew Phytologist, 193, 30 -50 . DOI:
10.1111/nph.2011.193.issue-1 URL [本文引用: 1] [39] Pregitzer KS DeForest JL Burton AJ Allen MF Ruess RW Hendrick RL 2002 ). Fine root architecture of nine North American treesEcological Monographs, 72, 293 -309 . DOI:
10.1890/0012-9615(2002)072[0293:FRAONN]2.0.CO;2 URL [本文引用: 2] [40] Radville L McCormack ML Post E Eissenstat DM 2016 ). Root phenology in a changing climateJournal of Experimental Botany, 67, 3617 -3628 . DOI:
10.1093/jxb/erw062 PMID:
26931171 [本文引用: 2] Plant phenology is one of the strongest indicators of ecological responses to climate change, and altered phenology can have pronounced effects on net primary production, species composition in local communities, greenhouse gas fluxes, and ecosystem processes. Although many studies have shown that aboveground plant phenology advances with warmer temperatures, demonstration of a comparable association for belowground phenology has been lacking because the factors that influence root phenology are poorly understood. Because roots can constitute a large fraction of plant biomass, and root phenology may not respond to warming in the same way as shoots, this represents an important knowledge gap in our understanding of how climate change will influence phenology and plant performance. We review studies of root phenology and provide suggestions to direct future research. Only 29% of examined studies approached root phenology quantitatively, strongly limiting interpretation of results across studies. Therefore, we suggest that researchers emphasize quantitative analyses in future phenological studies. We suggest that root initiation, peak growth, and root cessation may be under different controls. Root initiation and cessation may be more constrained by soil temperature and the timing of carbon availability, whereas the timing of peak root growth may represent trade-offs among competing plant sinks. Roots probably do not experience winter dormancy in the same way as shoots: 89% of the studies that examined winter phenology found evidence of growth during winter months. More research is needed to observe root phenology, and future studies should be careful to capture winter and early season phenology. This should be done quantitatively, with direct observations of root growth utilizing rhizotrons or minirhizotrons. © The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
[41] Reich PB 2014 ). The world-wide “fast-slow” plant economics spectrum: a traits manifestoJournal of Ecology, 102, 275 -301 . DOI:
10.1111/1365-2745.12211 URL [本文引用: 1] [42] Roumet C Urcelay C Díaz S 2006 ). Suites of root traits differ between annual and perennial species growing in the fieldNew Phytologist, 170, 357 -368 . DOI:
10.1111/nph.2006.170.issue-2 URL [本文引用: 1] [43] Ryser P 1996 ). The importance of tissue density for growth and life span of leaves and roots: a comparison of five ecologically contrasting grassesFunctional Ecology, 10, 717 . DOI:
10.2307/2390506 URL [本文引用: 3] [44] Sun K McCormack ML Li L Ma ZQ Guo DL 2016 ). Fast-cycling unit of root turnover in perennial herbaceous plants in a cold temperate ecosystemScientific Reports, 6, 19698 . DOI: https://www.plant-ecology.com/article/2021/1005-264X/10.1038/srep19698 . URL [本文引用: 2] [45] Tierney GL Fahey TJ 2001 ). Evaluating minirhizotron estimates of fine root longevity and production in the forest floor of a temperate broadleaf forestPlant and Soil, 229, 167 -176 . DOI:
10.1023/A:1004829423160 URL [本文引用: 2] [46] Warren JM Hanson PJ Iversen CM Kumar J Walker AP Wullschleger SD 2015 ). Root structural and functional dynamics in terrestrial biosphere models: evaluation and recommendationsNew Phytologist, 205, 59 -78 . DOI:
10.1111/nph.2014.205.issue-1 URL [本文引用: 1] [47] Weemstra M Kiorapostolou N van Ruijven J Mommer L de Vries J Sterck F 2020 ). The role of fine-root mass, specific root length and life span in tree performance: a whole-tree explorationFunctional Ecology, 34, 575 -585 . DOI:
10.1111/fec.v34.3 URL [本文引用: 3] [48] Weemstra M Mommer L Visser EJW van Ruijven J Kuyper TW Mohren GMJ Sterck FJ 2016 ). Towards a multidimensional root trait framework: a tree root reviewNew Phytologist, 211, 1159 -1169 . DOI:
10.1111/nph.2016.211.issue-4 URL [本文引用: 1] [49] Wells CE Glenn DM Eissenstat DM 2002 ). Changes in the risk of fine-root mortality with age: a case study in peach, Prunus persica (Rosaceae)American Journal of Botany, 79 -87 . DOI:
10.3732/ajb.89.1.79 URL [本文引用: 2] [50] Withington JM Reich PB Oleksyn J Eissenstat DM 2006 ). Comparisons of structure and life span in roots and leaves among temperate treesEcological Monographs, 76, 381 -397 . DOI:
10.1890/0012-9615(2006)076[0381:COSALS]2.0.CO;2 URL [本文引用: 3] [51] Wu C Wang ZQ Fan ZQ 2004 ). Significance of senescence study on tree roots and its advancesChinese Journal of Applied Ecology, 15, 1276 -1280 . [本文引用: 1] [ 吴楚 , 王政权 , 范志强 ( 2004 ). 树木根系衰老研究的意义与现状应用生态学报 , 15, 1276 -1280 .] [本文引用: 1] [52] Xia MX Guo DL Pregitzer KS 2010 ). Ephemeral root modules in Fraxinus mandshurica New Phytologist, 1065 -1074 . DOI:
10.1111/nph.2010.188.issue-4 URL [本文引用: 5] [53] Yu GR Chen Z Piao SL Peng CH Ciais P Wang QF Li XR Zhu XJ 2014 ). High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon regionProceedings of the National Academy of Sciences of the United States of America, 111, 4910 -4915 . [本文引用: 1] [54] Yu SQ Wang JB Hao QW Wang WF Wang Q Zhan LF 2020 ). Fine root lifespan and influencing factors of four tree species with different life formsActa Ecologica Sinica, 40, 3040 -3047 . [本文引用: 5] [ 于水强 , 王静波 , 郝倩葳 , 王维枫 , 王琪 , 詹龙飞 ( 2020 ). 四种不同生活型树种细根寿命及影响因素生态学报 , 40, 3040 -3047 .] [本文引用: 5] [55] Zadworny M McCormack ML Żytkowiak R Karolewski P Mucha J Oleksyn J 2017 ). Patterns of structural and defense investments in fine roots of Scots pine (Pinus sylvestris L.) across a strong temperature and latitudinal gradient in EuropeGlobal Change Biology, 1218 -1231 . DOI:
10.1111/gcb.13514 PMID:
27670838 [本文引用: 1] Plant functional traits may be altered as plants adapt to various environmental constraints. Cold, low fertility growing conditions are often associated with root adjustments to increase acquisition of limiting nutrient resources, but they may also result in construction of roots with reduced uptake potential but higher tissue persistence. It is ultimately unclear whether plants produce fine roots of different structure in response to decreasing temperatures and whether these changes represent a trade-off between root function or potential root persistence. We assessed patterns of root construction based on various root morphological, biochemical and defense traits including root diameter, specific root length (SRL), root tissue density (RTD), C:N ratio, phenolic compounds, and number of phellem layers across up to 10 root orders in diverse populations of Scots pine along a 2000-km climatic gradient in Europe. Our results showed that different root traits are related to mean annual temperature (MAT) and expressed a pattern of higher root diameter and lower SRL and RTD in northern sites with lower MAT. Among absorptive roots, we observed a gradual decline in chemical defenses (phenolic compounds) with decreasing MAT. In contrast, decreasing MAT resulted in an increase of structural protection (number of phellem layers) in transport fine roots. This indicated that absorptive roots with high capacity for nutrient uptake, and transport roots with low uptake capacity, were characterized by distinct and contrasting trade-offs. Our observations suggest that diminishing structural and chemical investments into the more distal, absorptive roots in colder climates is consistent with building roots of higher absorptive capacity. At the same time, roots that play a more prominent role in transport of nutrients and water within the root system saw an increase in structural investment, which can increase persistence and reduce long-term costs associated with their frequent replacement. © 2016 John Wiley & Sons Ltd.
[56] Zheng JX Huang JX Wang ZZ Xiong DC Yang ZJ Chen GS 2012 ). Fine root longevity and controlling factors in aPhoebe bournei plantation Acta Ecologica Sinica, 7532 -7539 . DOI:
10.5846/stxb URL [本文引用: 3] [ 郑金兴 , 黄锦学 , 王珍珍 , 熊德成 , 杨智杰 , 陈光水 ( 2012 ). 闽楠人工林细根寿命及其影响因素生态学报 , 32, 7532 -7539 .] [本文引用: 3] Are above- and below-ground phenology in sync?
1
2015
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
The advantages of being evergreen
1
1995
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
Differences in spatial and temporal root lifespan of three
Stipa grasslands in northern China
1
2017
... 亚热带树种的根级对吸收细根寿命有着显著的影响, 随根级增加, 根寿命延长(
图3C ).树木细根是一个高度异质的系统, 根构型是预测寿命的核心因素, 着生在根枝末端的一级根的周转率5倍于着生在根枝基部的五级根(
Xiaet al ., 2010 ).乔木的根级最大可以到10级(
Kajimoto, 2010 ), 前五级根是重点关注的单元(
Guoet al ., 2008b ), 但微根管的观测视野较小, 很难观测到全部5个根级, 针对这个缺点, 根窗法或扫描根系的方法能够全面地理解异质性的根系统, 值得推荐(
Xiaet al ., 2010 ;
Huo & Cheng, 2019 ).通常认为一级根和二级根能被共生真菌侵染且具有皮层, 划分为短寿命的吸收功能模块, 四五级根为长寿命的吸收运输根功能模块, 三级根主要为过渡; 为了方便研究, 把前三级根或前两级根作为短命吸收细根模块(
Xiaet al ., 2010 ).本研究也证实这6个树种前两级根的根寿命基本上是连续的变异, 并未出现突变的结果; 这与北方落叶松的情况有区别, 一级根和二级根的寿命相差4倍(
Huo & Cheng, 2019 ), 表明外生菌根侵染的一级根(根尖)是频繁死亡、半独立、快速周转的单元模块.在草本植物中, 根周转与物种本身的构型具有密切的关系(
Sunet al ., 2016 ;
Baiet al ., 2017 ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
Evaluation of fine root length and diameter measurements obtained using RHIZO image analysis
1
1999
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests
1
2000
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
Linking root traits to potential growth rate in six temperate tree species
1
2002
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
Patterns in root traits of woody species hosting arbuscular and ectomycorrhizas: implications for the evolution of belowground strategies
1
2014
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
Linking fine root traits to maximum potential growth rate among 11 mature temperate tree species
2
2004
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
Regional distribution patterns of wetland monocots with different root turnover strategies are associated with local variation in soil temperature
1
2020
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
1
1997
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
Building roots in a changing environment: implications for root longevity
1
2000
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
The ecology of root lifespan
2
1997
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
Slow decomposition of lower order roots: a key mechanism of root carbon and nutrient retention in the soil
1
2010
... 植物通过根系的生长、死亡、分解等过程驱动土壤碳氮养分循环, 但准确定量吸收细根周转一直是陆地生态系统研究中的“瓶颈” (
Jacksonet al ., 1997 ;
Norbyet al ., 2004 ;
Guoet al ., 2008a ).根系通常占植物总生物量的63% (
Poorteret al ., 2012 ), 约占全球净初级生产力的22% (
McCormacket al ., 2015 ).由于细根中氮磷含量高(
Gordon & Jackson, 2000 ), 细根死亡后迅速分解, 将养分归还土壤(
Fan & Guo, 2010 ), 将碳以CO
2 的形式释放回大气, 因此吸收细根周转驱动的碳氮循环一直作为陆地生态系统的核心内容来研究(
Warrenet al ., 2015 ). ...
Evidence of the “plant economics spectrum” in a subarctic flora
1
2010
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
The age of fine-root carbon in three forests of the eastern United States measured by radiocarbon
1
2001
... 由于直接观测根系较为困难, 根寿命的研究具有较大的不确定性, 主要表现在3个方面(
Guoet al ., 2008a ;
Huo & Cheng, 2019 ).一是方法的不确定性.目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率.二是根系观测单元的不确定性(
Guoet al ., 2008b ).树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
Nutrient concentrations in fine roots
1
2000
... 植物通过根系的生长、死亡、分解等过程驱动土壤碳氮养分循环, 但准确定量吸收细根周转一直是陆地生态系统研究中的“瓶颈” (
Jacksonet al ., 1997 ;
Norbyet al ., 2004 ;
Guoet al ., 2008a ).根系通常占植物总生物量的63% (
Poorteret al ., 2012 ), 约占全球净初级生产力的22% (
McCormacket al ., 2015 ).由于细根中氮磷含量高(
Gordon & Jackson, 2000 ), 细根死亡后迅速分解, 将养分归还土壤(
Fan & Guo, 2010 ), 将碳以CO
2 的形式释放回大气, 因此吸收细根周转驱动的碳氮循环一直作为陆地生态系统的核心内容来研究(
Warrenet al ., 2015 ). ...
Effects of root diameter, branch order, soil depth and season of birth on fine root life span in five temperate tree species
1
2017
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
Fine root heterogeneity by branch order: exploring the discrepancy in root turnover estimates between minirhizotron and carbon isotopic methods
4
2008a
... 植物通过根系的生长、死亡、分解等过程驱动土壤碳氮养分循环, 但准确定量吸收细根周转一直是陆地生态系统研究中的“瓶颈” (
Jacksonet al ., 1997 ;
Norbyet al ., 2004 ;
Guoet al ., 2008a ).根系通常占植物总生物量的63% (
Poorteret al ., 2012 ), 约占全球净初级生产力的22% (
McCormacket al ., 2015 ).由于细根中氮磷含量高(
Gordon & Jackson, 2000 ), 细根死亡后迅速分解, 将养分归还土壤(
Fan & Guo, 2010 ), 将碳以CO
2 的形式释放回大气, 因此吸收细根周转驱动的碳氮循环一直作为陆地生态系统的核心内容来研究(
Warrenet al ., 2015 ). ...
... 由于直接观测根系较为困难, 根寿命的研究具有较大的不确定性, 主要表现在3个方面(
Guoet al ., 2008a ;
Huo & Cheng, 2019 ).一是方法的不确定性.目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率.二是根系观测单元的不确定性(
Guoet al ., 2008b ).树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... ).一是方法的不确定性.目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率.二是根系观测单元的不确定性(
Guoet al ., 2008b ).树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... 根据
Pregitzer等(2002) 和
Guo等(2008a) 所描述的分级方法, 将采集到的根系样本分根处理后, 按根级置于培养皿中, 避免交叉重叠, 利用Epson数字化扫描仪进行扫描, 使用WinRHIZO软件对扫描图像进行处理, 获取平均根系直径、总根长、总根体积及总根表面积等数据, 并结合根样本干质量获得比根长(
SRL )、根组织密度(
RTD )及比根表面积(
SRA )等关键的形态属性指标.
SRL 的计算方法为根总长度除以其干质量.
RTD 是根干质量与根体积的 ...
Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species
3
2008b
... 由于直接观测根系较为困难, 根寿命的研究具有较大的不确定性, 主要表现在3个方面(
Guoet al ., 2008a ;
Huo & Cheng, 2019 ).一是方法的不确定性.目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率.二是根系观测单元的不确定性(
Guoet al ., 2008b ).树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... 亚热带树种的根级对吸收细根寿命有着显著的影响, 随根级增加, 根寿命延长(
图3C ).树木细根是一个高度异质的系统, 根构型是预测寿命的核心因素, 着生在根枝末端的一级根的周转率5倍于着生在根枝基部的五级根(
Xiaet al ., 2010 ).乔木的根级最大可以到10级(
Kajimoto, 2010 ), 前五级根是重点关注的单元(
Guoet al ., 2008b ), 但微根管的观测视野较小, 很难观测到全部5个根级, 针对这个缺点, 根窗法或扫描根系的方法能够全面地理解异质性的根系统, 值得推荐(
Xiaet al ., 2010 ;
Huo & Cheng, 2019 ).通常认为一级根和二级根能被共生真菌侵染且具有皮层, 划分为短寿命的吸收功能模块, 四五级根为长寿命的吸收运输根功能模块, 三级根主要为过渡; 为了方便研究, 把前三级根或前两级根作为短命吸收细根模块(
Xiaet al ., 2010 ).本研究也证实这6个树种前两级根的根寿命基本上是连续的变异, 并未出现突变的结果; 这与北方落叶松的情况有区别, 一级根和二级根的寿命相差4倍(
Huo & Cheng, 2019 ), 表明外生菌根侵染的一级根(根尖)是频繁死亡、半独立、快速周转的单元模块.在草本植物中, 根周转与物种本身的构型具有密切的关系(
Sunet al ., 2016 ;
Baiet al ., 2017 ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
Fine root production and turnover of tree and understorey vegetation in Scots pine, silver birch and Norway spruce stands in SW Sweden
1
2013
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
细柄阿丁枫和米槠细根寿命影响因素
4
2012
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
细柄阿丁枫和米槠细根寿命影响因素
4
2012
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
Improved root turnover assessment using field scanning rhizotrons with branch order analysis
6
2019
... 由于直接观测根系较为困难, 根寿命的研究具有较大的不确定性, 主要表现在3个方面(
Guoet al ., 2008a ;
Huo & Cheng, 2019 ).一是方法的不确定性.目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率.二是根系观测单元的不确定性(
Guoet al ., 2008b ).树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
... 亚热带树种的根级对吸收细根寿命有着显著的影响, 随根级增加, 根寿命延长(
图3C ).树木细根是一个高度异质的系统, 根构型是预测寿命的核心因素, 着生在根枝末端的一级根的周转率5倍于着生在根枝基部的五级根(
Xiaet al ., 2010 ).乔木的根级最大可以到10级(
Kajimoto, 2010 ), 前五级根是重点关注的单元(
Guoet al ., 2008b ), 但微根管的观测视野较小, 很难观测到全部5个根级, 针对这个缺点, 根窗法或扫描根系的方法能够全面地理解异质性的根系统, 值得推荐(
Xiaet al ., 2010 ;
Huo & Cheng, 2019 ).通常认为一级根和二级根能被共生真菌侵染且具有皮层, 划分为短寿命的吸收功能模块, 四五级根为长寿命的吸收运输根功能模块, 三级根主要为过渡; 为了方便研究, 把前三级根或前两级根作为短命吸收细根模块(
Xiaet al ., 2010 ).本研究也证实这6个树种前两级根的根寿命基本上是连续的变异, 并未出现突变的结果; 这与北方落叶松的情况有区别, 一级根和二级根的寿命相差4倍(
Huo & Cheng, 2019 ), 表明外生菌根侵染的一级根(根尖)是频繁死亡、半独立、快速周转的单元模块.在草本植物中, 根周转与物种本身的构型具有密切的关系(
Sunet al ., 2016 ;
Baiet al ., 2017 ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
... ).本研究也证实这6个树种前两级根的根寿命基本上是连续的变异, 并未出现突变的结果; 这与北方落叶松的情况有区别, 一级根和二级根的寿命相差4倍(
Huo & Cheng, 2019 ), 表明外生菌根侵染的一级根(根尖)是频繁死亡、半独立、快速周转的单元模块.在草本植物中, 根周转与物种本身的构型具有密切的关系(
Sunet al ., 2016 ;
Baiet al ., 2017 ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
... ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
Using root form to improve our understanding of root function
1
2014
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
A global budget for fine root biomass, surface area, and nutrient contents
2
1997
... 植物通过根系的生长、死亡、分解等过程驱动土壤碳氮养分循环, 但准确定量吸收细根周转一直是陆地生态系统研究中的“瓶颈” (
Jacksonet al ., 1997 ;
Norbyet al ., 2004 ;
Guoet al ., 2008a ).根系通常占植物总生物量的63% (
Poorteret al ., 2012 ), 约占全球净初级生产力的22% (
McCormacket al ., 2015 ).由于细根中氮磷含量高(
Gordon & Jackson, 2000 ), 细根死亡后迅速分解, 将养分归还土壤(
Fan & Guo, 2010 ), 将碳以CO
2 的形式释放回大气, 因此吸收细根周转驱动的碳氮循环一直作为陆地生态系统的核心内容来研究(
Warrenet al ., 2015 ). ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
Root system development of larch trees growing on siberian permafrost
1
2010
... 亚热带树种的根级对吸收细根寿命有着显著的影响, 随根级增加, 根寿命延长(
图3C ).树木细根是一个高度异质的系统, 根构型是预测寿命的核心因素, 着生在根枝末端的一级根的周转率5倍于着生在根枝基部的五级根(
Xiaet al ., 2010 ).乔木的根级最大可以到10级(
Kajimoto, 2010 ), 前五级根是重点关注的单元(
Guoet al ., 2008b ), 但微根管的观测视野较小, 很难观测到全部5个根级, 针对这个缺点, 根窗法或扫描根系的方法能够全面地理解异质性的根系统, 值得推荐(
Xiaet al ., 2010 ;
Huo & Cheng, 2019 ).通常认为一级根和二级根能被共生真菌侵染且具有皮层, 划分为短寿命的吸收功能模块, 四五级根为长寿命的吸收运输根功能模块, 三级根主要为过渡; 为了方便研究, 把前三级根或前两级根作为短命吸收细根模块(
Xiaet al ., 2010 ).本研究也证实这6个树种前两级根的根寿命基本上是连续的变异, 并未出现突变的结果; 这与北方落叶松的情况有区别, 一级根和二级根的寿命相差4倍(
Huo & Cheng, 2019 ), 表明外生菌根侵染的一级根(根尖)是频繁死亡、半独立、快速周转的单元模块.在草本植物中, 根周转与物种本身的构型具有密切的关系(
Sunet al ., 2016 ;
Baiet al ., 2017 ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
Leading dimensions in absorptive root trait variation across 96 subtropical forest species
1
2014
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
DNA fragmentation in cereal roots indicative of programmed root cortical cell death
1
2001
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
杉木人工林细根寿命的影响因素
3
2011
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
杉木人工林细根寿命的影响因素
3
2011
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree species
1
2015
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
Tree species richness increases ecosystem carbon storage in subtropical forests
1
2018
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
Evolutionary history resolves global organization of root functional traits
3
2018
... 由于直接观测根系较为困难, 根寿命的研究具有较大的不确定性, 主要表现在3个方面(
Guoet al ., 2008a ;
Huo & Cheng, 2019 ).一是方法的不确定性.目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率.二是根系观测单元的不确定性(
Guoet al ., 2008b ).树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
Longevity of mycorrhizal roots depends on branching order and nutrient availability
1
2001
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
Predicting fine root lifespan from plant functional traits in temperate trees
5
2012
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 利用Rootfly软件对采集到的微根管图像进行分析, 以在图片中首次出现的浅色(白色或棕色)根为活根, 当该根从同位置观察画面消失或者颜色变为深色(黑色或深褐色)或表面皱缩则表示该根死亡; 将观察到的一级及二级根定义为吸收细根(
McCormacket al ., 2015 ).把吸收细根从第一次出现到最后一次出现的时间间隔称为吸收细根寿命.本研究将出生状态被观测到但由于实验结束或其他因素影响导致死亡状态未被观察到的细根定义为删失细根(
McCormacket al ., 2012 ).因此在表达细根寿命时选用平均寿命与中值寿命两个指标, 以提高细根寿命数据的可靠性(
Wells et al ., 2002 ). ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes
2
2015
... 植物通过根系的生长、死亡、分解等过程驱动土壤碳氮养分循环, 但准确定量吸收细根周转一直是陆地生态系统研究中的“瓶颈” (
Jacksonet al ., 1997 ;
Norbyet al ., 2004 ;
Guoet al ., 2008a ).根系通常占植物总生物量的63% (
Poorteret al ., 2012 ), 约占全球净初级生产力的22% (
McCormacket al ., 2015 ).由于细根中氮磷含量高(
Gordon & Jackson, 2000 ), 细根死亡后迅速分解, 将养分归还土壤(
Fan & Guo, 2010 ), 将碳以CO
2 的形式释放回大气, 因此吸收细根周转驱动的碳氮循环一直作为陆地生态系统的核心内容来研究(
Warrenet al ., 2015 ). ...
... 利用Rootfly软件对采集到的微根管图像进行分析, 以在图片中首次出现的浅色(白色或棕色)根为活根, 当该根从同位置观察画面消失或者颜色变为深色(黑色或深褐色)或表面皱缩则表示该根死亡; 将观察到的一级及二级根定义为吸收细根(
McCormacket al ., 2015 ).把吸收细根从第一次出现到最后一次出现的时间间隔称为吸收细根寿命.本研究将出生状态被观测到但由于实验结束或其他因素影响导致死亡状态未被观察到的细根定义为删失细根(
McCormacket al ., 2012 ).因此在表达细根寿命时选用平均寿命与中值寿命两个指标, 以提高细根寿命数据的可靠性(
Wells et al ., 2002 ). ...
Impacts of environmental factors on fine root lifespan
2
2014
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
Plant organ senescence above- and belowground in trees: How to best salvage resources for new growth?
1
2020
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
Fine-root production dominates response of a deciduous forest to atmospheric CO
2 enrichment
1
2004
... 植物通过根系的生长、死亡、分解等过程驱动土壤碳氮养分循环, 但准确定量吸收细根周转一直是陆地生态系统研究中的“瓶颈” (
Jacksonet al ., 1997 ;
Norbyet al ., 2004 ;
Guoet al ., 2008a ).根系通常占植物总生物量的63% (
Poorteret al ., 2012 ), 约占全球净初级生产力的22% (
McCormacket al ., 2015 ).由于细根中氮磷含量高(
Gordon & Jackson, 2000 ), 细根死亡后迅速分解, 将养分归还土壤(
Fan & Guo, 2010 ), 将碳以CO
2 的形式释放回大气, 因此吸收细根周转驱动的碳氮循环一直作为陆地生态系统的核心内容来研究(
Warrenet al ., 2015 ). ...
Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control
1
2012
... 植物通过根系的生长、死亡、分解等过程驱动土壤碳氮养分循环, 但准确定量吸收细根周转一直是陆地生态系统研究中的“瓶颈” (
Jacksonet al ., 1997 ;
Norbyet al ., 2004 ;
Guoet al ., 2008a ).根系通常占植物总生物量的63% (
Poorteret al ., 2012 ), 约占全球净初级生产力的22% (
McCormacket al ., 2015 ).由于细根中氮磷含量高(
Gordon & Jackson, 2000 ), 细根死亡后迅速分解, 将养分归还土壤(
Fan & Guo, 2010 ), 将碳以CO
2 的形式释放回大气, 因此吸收细根周转驱动的碳氮循环一直作为陆地生态系统的核心内容来研究(
Warrenet al ., 2015 ). ...
Fine root architecture of nine North American trees
2
2002
... 由于直接观测根系较为困难, 根寿命的研究具有较大的不确定性, 主要表现在3个方面(
Guoet al ., 2008a ;
Huo & Cheng, 2019 ).一是方法的不确定性.目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率.二是根系观测单元的不确定性(
Guoet al ., 2008b ).树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... 根据
Pregitzer等(2002) 和
Guo等(2008a) 所描述的分级方法, 将采集到的根系样本分根处理后, 按根级置于培养皿中, 避免交叉重叠, 利用Epson数字化扫描仪进行扫描, 使用WinRHIZO软件对扫描图像进行处理, 获取平均根系直径、总根长、总根体积及总根表面积等数据, 并结合根样本干质量获得比根长(
SRL )、根组织密度(
RTD )及比根表面积(
SRA )等关键的形态属性指标.
SRL 的计算方法为根总长度除以其干质量.
RTD 是根干质量与根体积的 ...
Root phenology in a changing climate
2
2016
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
The world-wide “fast-slow” plant economics spectrum: a traits manifesto
1
2014
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
Suites of root traits differ between annual and perennial species growing in the field
1
2006
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
The importance of tissue density for growth and life span of leaves and roots: a comparison of five ecologically contrasting grasses
3
1996
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
Fast-cycling unit of root turnover in perennial herbaceous plants in a cold temperate ecosystem
2
2016
... 亚热带树种的根级对吸收细根寿命有着显著的影响, 随根级增加, 根寿命延长(
图3C ).树木细根是一个高度异质的系统, 根构型是预测寿命的核心因素, 着生在根枝末端的一级根的周转率5倍于着生在根枝基部的五级根(
Xiaet al ., 2010 ).乔木的根级最大可以到10级(
Kajimoto, 2010 ), 前五级根是重点关注的单元(
Guoet al ., 2008b ), 但微根管的观测视野较小, 很难观测到全部5个根级, 针对这个缺点, 根窗法或扫描根系的方法能够全面地理解异质性的根系统, 值得推荐(
Xiaet al ., 2010 ;
Huo & Cheng, 2019 ).通常认为一级根和二级根能被共生真菌侵染且具有皮层, 划分为短寿命的吸收功能模块, 四五级根为长寿命的吸收运输根功能模块, 三级根主要为过渡; 为了方便研究, 把前三级根或前两级根作为短命吸收细根模块(
Xiaet al ., 2010 ).本研究也证实这6个树种前两级根的根寿命基本上是连续的变异, 并未出现突变的结果; 这与北方落叶松的情况有区别, 一级根和二级根的寿命相差4倍(
Huo & Cheng, 2019 ), 表明外生菌根侵染的一级根(根尖)是频繁死亡、半独立、快速周转的单元模块.在草本植物中, 根周转与物种本身的构型具有密切的关系(
Sunet al ., 2016 ;
Baiet al ., 2017 ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
... ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
Evaluating minirhizotron estimates of fine root longevity and production in the forest floor of a temperate broadleaf forest
2
2001
... 由于直接观测根系较为困难, 根寿命的研究具有较大的不确定性, 主要表现在3个方面(
Guoet al ., 2008a ;
Huo & Cheng, 2019 ).一是方法的不确定性.目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率.二是根系观测单元的不确定性(
Guoet al ., 2008b ).树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
Root structural and functional dynamics in terrestrial biosphere models: evaluation and recommendations
1
2015
... 植物通过根系的生长、死亡、分解等过程驱动土壤碳氮养分循环, 但准确定量吸收细根周转一直是陆地生态系统研究中的“瓶颈” (
Jacksonet al ., 1997 ;
Norbyet al ., 2004 ;
Guoet al ., 2008a ).根系通常占植物总生物量的63% (
Poorteret al ., 2012 ), 约占全球净初级生产力的22% (
McCormacket al ., 2015 ).由于细根中氮磷含量高(
Gordon & Jackson, 2000 ), 细根死亡后迅速分解, 将养分归还土壤(
Fan & Guo, 2010 ), 将碳以CO
2 的形式释放回大气, 因此吸收细根周转驱动的碳氮循环一直作为陆地生态系统的核心内容来研究(
Warrenet al ., 2015 ). ...
The role of fine-root mass, specific root length and life span in tree performance: a whole-tree exploration
3
2020
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
Towards a multidimensional root trait framework: a tree root review
1
2016
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
Changes in the risk of fine-root mortality with age: a case study in peach,
Prunus persica (Rosaceae)
2
2002
... 由于直接观测根系较为困难, 根寿命的研究具有较大的不确定性, 主要表现在3个方面(
Guoet al ., 2008a ;
Huo & Cheng, 2019 ).一是方法的不确定性.目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率.二是根系观测单元的不确定性(
Guoet al ., 2008b ).树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... 利用Rootfly软件对采集到的微根管图像进行分析, 以在图片中首次出现的浅色(白色或棕色)根为活根, 当该根从同位置观察画面消失或者颜色变为深色(黑色或深褐色)或表面皱缩则表示该根死亡; 将观察到的一级及二级根定义为吸收细根(
McCormacket al ., 2015 ).把吸收细根从第一次出现到最后一次出现的时间间隔称为吸收细根寿命.本研究将出生状态被观测到但由于实验结束或其他因素影响导致死亡状态未被观察到的细根定义为删失细根(
McCormacket al ., 2012 ).因此在表达细根寿命时选用平均寿命与中值寿命两个指标, 以提高细根寿命数据的可靠性(
Wells et al ., 2002 ). ...
Comparisons of structure and life span in roots and leaves among temperate trees
3
2006
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
树木根系衰老研究的意义与现状
1
2004
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
树木根系衰老研究的意义与现状
1
2004
... 吸收细根寿命与直径显著正相关, 与比根长显著负相关, 与根组织密度无相关关系, 表明根的构建成本可以在一定程度上预测寿命, 支持弱的根经济谱的理论(
Weemstraet al ., 2016 ,
2020 ;
Maet al ., 2018 ).如果根直径之间变异范围足够大, 根直径足够粗, 碳构建成本越高, 它就需要用较长的寿命来确保高碳投资回报率(
McCormacket al ., 2012 ).草本植物中根直径普遍细, 根构建成本与根寿命之间权衡不太明显(
Roumetet al ., 2006 ), 但根组织密度能够在一定程度上预测根的寿命(
Ryser, 1996 ).
Tierney和Fahey (2001) 发现美国东北部硬木林的腐殖质层和矿质表层中细根的存活率随根直径的增加而显著提高: 直径每增加0.1 mm, 存活率提高43%; 本研究发现亚热带6个树种根直径每增加0.1 mm, 存活率提高近30%.根寿命符合成本效益比, 服从植物整体生态策略, 受控于器官内碳水化合物数量及分配(
Eissenstat & Yanai, 1997 ).从细胞水平上讲, 吸收细根衰老是衰老皮层细胞的细胞核逐渐解体, 成熟区细胞壁栓质加厚, 吸收细根成熟区细胞吸收能力下降; 物质吸收和运输通道衰解的过程, 说明根衰老是一种程序化死亡的结果(
Liljeroth & Bryngelsson, 2001 ), 目前对单根尺度根皮层细胞死亡的机制还缺乏研究(
吴楚等, 2004 ).只有准确测定不同直径和根级的根寿命, 我们才能定量出根系通过周转和分解向土壤系统输送的碳和养分的数量.本研究由于涉及的物种较少, 同一根级的根寿命与属性关系不显著, 未来需增加样本量. ...
Ephemeral root modules in
Fraxinus mandshurica 5
2010
... 由于直接观测根系较为困难, 根寿命的研究具有较大的不确定性, 主要表现在3个方面(
Guoet al ., 2008a ;
Huo & Cheng, 2019 ).一是方法的不确定性.目前通用的微根管中值寿命(一般为1年左右)大大低估寿命, 高估了细根周转率(
Guoet al ., 2008a ), 而碳同位素(
14 C)法测得的根寿命(一般为5年左右)则明显高估寿命(
Gaudinskiet al ., 2001 ), 低估了细根周转率.二是根系观测单元的不确定性(
Guoet al ., 2008b ).树木细根是一个高度异质的系统(
Pregitzeret al ., 2002 ), 着生在根枝末端的一级根的周转率5倍于着生在根基部的五级根(
Xiaet al ., 2010 ).并且, 随根系分枝等级增加, 细根个体数量呈指数下降(
Guoet al ., 2008b ).三是根系观测周期的影响.目前观测细根寿命的研究很少有长于3年的, 而部分细根的寿命可能大于3年(
Huo & Cheng, 2019 ), 导致难以获得所观测的所有吸收细根的完整周转过程; 因此吸收细根寿命的频率分布往往呈非正态分布(
Tierney & Fahey, 2001 ).此外, 根观测频率对寿命估计具有不可忽视的影响(
Wellset al ., 2002 ).由于观测困难, 研究费时耗力, 目前全球直接观测的根寿命数据缺乏, 特别是热带、亚热带木本植物的根寿命数据就更少(
Maet al ., 2018 ). ...
... 亚热带树种的根级对吸收细根寿命有着显著的影响, 随根级增加, 根寿命延长(
图3C ).树木细根是一个高度异质的系统, 根构型是预测寿命的核心因素, 着生在根枝末端的一级根的周转率5倍于着生在根枝基部的五级根(
Xiaet al ., 2010 ).乔木的根级最大可以到10级(
Kajimoto, 2010 ), 前五级根是重点关注的单元(
Guoet al ., 2008b ), 但微根管的观测视野较小, 很难观测到全部5个根级, 针对这个缺点, 根窗法或扫描根系的方法能够全面地理解异质性的根系统, 值得推荐(
Xiaet al ., 2010 ;
Huo & Cheng, 2019 ).通常认为一级根和二级根能被共生真菌侵染且具有皮层, 划分为短寿命的吸收功能模块, 四五级根为长寿命的吸收运输根功能模块, 三级根主要为过渡; 为了方便研究, 把前三级根或前两级根作为短命吸收细根模块(
Xiaet al ., 2010 ).本研究也证实这6个树种前两级根的根寿命基本上是连续的变异, 并未出现突变的结果; 这与北方落叶松的情况有区别, 一级根和二级根的寿命相差4倍(
Huo & Cheng, 2019 ), 表明外生菌根侵染的一级根(根尖)是频繁死亡、半独立、快速周转的单元模块.在草本植物中, 根周转与物种本身的构型具有密切的关系(
Sunet al ., 2016 ;
Baiet al ., 2017 ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
... ), 但微根管的观测视野较小, 很难观测到全部5个根级, 针对这个缺点, 根窗法或扫描根系的方法能够全面地理解异质性的根系统, 值得推荐(
Xiaet al ., 2010 ;
Huo & Cheng, 2019 ).通常认为一级根和二级根能被共生真菌侵染且具有皮层, 划分为短寿命的吸收功能模块, 四五级根为长寿命的吸收运输根功能模块, 三级根主要为过渡; 为了方便研究, 把前三级根或前两级根作为短命吸收细根模块(
Xiaet al ., 2010 ).本研究也证实这6个树种前两级根的根寿命基本上是连续的变异, 并未出现突变的结果; 这与北方落叶松的情况有区别, 一级根和二级根的寿命相差4倍(
Huo & Cheng, 2019 ), 表明外生菌根侵染的一级根(根尖)是频繁死亡、半独立、快速周转的单元模块.在草本植物中, 根周转与物种本身的构型具有密切的关系(
Sunet al ., 2016 ;
Baiet al ., 2017 ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
... ).通常认为一级根和二级根能被共生真菌侵染且具有皮层, 划分为短寿命的吸收功能模块, 四五级根为长寿命的吸收运输根功能模块, 三级根主要为过渡; 为了方便研究, 把前三级根或前两级根作为短命吸收细根模块(
Xiaet al ., 2010 ).本研究也证实这6个树种前两级根的根寿命基本上是连续的变异, 并未出现突变的结果; 这与北方落叶松的情况有区别, 一级根和二级根的寿命相差4倍(
Huo & Cheng, 2019 ), 表明外生菌根侵染的一级根(根尖)是频繁死亡、半独立、快速周转的单元模块.在草本植物中, 根周转与物种本身的构型具有密切的关系(
Sunet al ., 2016 ;
Baiet al ., 2017 ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
... ), 不同植物根系的构型和生长是否具有功能模块化的构造, 是否能够预测寿命等功能, 也是一个颇有研究价值的问题(
Xiaet al ., 2010 ;
Sunet al ., 2016 ;
Huo & Cheng, 2019 ).解决这一问题需要采集完整根系的形态结构指标, 这是一项困难的工作, 其成功很大程度上依赖于根系观测技术和采样方法的改进. ...
High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region
1
2014
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
四种不同生活型树种细根寿命及影响因素
5
2020
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
四种不同生活型树种细根寿命及影响因素
5
2020
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 亚热带常绿针叶树种寿命是常绿阔叶树种根寿命的1.5倍; 主要树种吸收细根中值寿命格局: 红豆杉(常绿针叶) >复羽叶栾树(落叶阔叶) >竹柏(常绿针叶) >樟(常绿阔叶) >东京樱花(落叶阔叶) >深山含笑(常绿阔叶)(
图2 ).由于观测物种较少, 本研究中并没有发现生活型具有系统性的差异.从全球来看, 北方寒温带树种松树根寿命较长, 如欧洲赤松(
Pinus sylvestris )的细根寿命为924天(
Hanssonet al ., 2013 ), 红松(
Pinus koraiensis )的细根寿命最长为434天(
Guet al ., 2017 ), 帽儿山的落叶松(
Larix gmelinii )的一级根寿命为264天, 二级根可以达到990天(
Huo & Cheng, 2019 ).常绿针叶树种红豆杉(460天)(
图1 )和柳杉(280天)的根寿命也较长(
于水强等, 2020 ), 而杉科树木与松科不同, 其获取土壤资源更依赖内生菌根(
Comaset al ., 2014 ).总体来看, 针叶裸子植物的根寿命通常较长(
Majdiet al ., 2001 ;
Withingtonet al ., 2006 ), 这类常绿针叶生活型是应对资源稀缺的一种生态对策(
Aerts, 1995 ), 具有更粗的根和更低的比根长(
Bauhus & Messier, 1999 ;
Comas & Eissenstat, 2004 ), 有可能会表现为慢策略(
Reich, 2014 ).总之, 长期的进化过程中, 物种是决定根寿命的关键因素. ...
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
Patterns of structural and defense investments in fine roots of Scots pine (
Pinus sylvestris L.) across a strong temperature and latitudinal gradient in Europe
1
2017
... 亚热带常绿阔叶林代表性树种以壳斗科、木兰科和樟科植物为主(
Liu et al ., 2018 ).研究表明壳斗科麻栎(
Quercus acutissima )(约69天)和石栎(
Lithocarpus glaber )(约124天)的根周转较快(
于水强等, 2020 ), 米槠根寿命约为212天(
黄锦学等, 2012 ); 本研究表明木兰科的深山含笑和樟科的樟的根寿命较短, 分别为92和126天, 即地带性植被具有较高的根周转速率和获取土壤资源的能力.深山含笑的比根长与其他5个物种相似, 但组织密度较小, 说明其物理防御功能较弱(
Zadwornyet al ., 2017 ); 物种间获取土壤资源、防御功能的差异与寿命相关(
Comas & Eissenstat, 2004 ;
Courchesneet al ., 2020 ;
Niinemets & Ostonen, 2020 ), 一定程度上可以预测根的寿命(
Ryser, 1996 ).树木整体而言, 理论上认为速生树种具有较快的根周转率(
McCormacket al ., 2012 ;
Weemstraet al ., 2020 ).温带槭树科和壳斗科生长快的植物细根对磷的吸收也比生长慢的植物快, 而松科植物对磷的吸收差异不大(
Comaset al ., 2002 ), 据此推断地带性的亚热带优势植物理论上应该有较高的磷获取效率, 未来根寿命研究工作需与养分获取速率研究相结合. ...
闽楠人工林细根寿命及其影响因素
3
2012
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
闽楠人工林细根寿命及其影响因素
3
2012
... 我国亚热带森林生产力较高, 特别是大面积人工林的碳汇功能显著, 对于深入理解北半球生态系统能量流动和物质循环具有重要意义(
Yuet al ., 2014 ).亚热带不同树种根寿命与周转是理解森林生态系统物质循环的关键, 然而从全球来看这个区域的数据较为匮乏(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
Maet al ., 2018 ;
于水强等, 2020 ).由于亚热带森林演化时间长, 树种丰富, 不同树种根功能属性往往能够表征地下资源获取策略(
Liuet al ., 2015 ), 反映出根系构建成本的选择(
Eissenstat, 1997 ;
Eissenstatet al ., 2000 ;
Konget al ., 2014 ).例如, 同一气候区不同树种快速周转的根模块的寿命是否大致相同(
Withingtonet al ., 2006 )? 树种的根寿命主要受限于根系自身形态属性(
Ryser, 1996 ;
McCormacket al ., 2012 ;
Iversen, 2014 ), 还是受植物整体的影响(
Freschetet al ., 2010 ;
Weemstraet al ., 2020 ); 或是更容易受到气候的影响(
McCormack & Guo, 2014 ), 如独特的亚热带季风气候、季节性干旱等, 目前还缺乏有效数据检验这些科学假说. ...
... 亚热带6个树种的吸收细根中值寿命为92-426天(
表1 ), 变异范围略高于北美温带森林12种树木吸收细根寿命(95-336天)(
McCormacket al ., 2012 ), 低于北温带波兰同质园内11个乔木树种细根寿命(231-900天)(
Withingtonet al ., 2006 ).亚热带常绿阔叶林代表性树种如细柄蕈树(
Altingia gracilipes )(约184天)、米槠(
Castanopsis carlesii )(约212天)、杉木(
Cunninghamia lanceolata )(约220天)、闽楠(
Phoebe bournei )(约89天)、柳杉(
Cryptomeria japonica )(280天)的吸收细根寿命基本也位于该区间(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ); 表明绝大多数亚热带树种根系周转至少1年1次; 如果沿用1年的一次周转率(
Jacksonet al ., 1997 ), 可能会低估亚热带森林生态系统的根碳周转的贡献.根系的萌生和停止可能更多地受到土壤温度和资源供应有效性的控制(
Radvilleet al ., 2016 ), 尽管亚热带冬季不受冻融的影响, 但这6个树种细根冬季出生率较低, 死亡率较高, 因此亚热带植物冬季的根叶物候的权衡是理解该区域树木碳分配的关键. ...
... 东亚亚热带季风气候具有伏秋旱、雨热不同期、年际变化大等特点.在这种气候条件下, 亚热带常绿阔叶林典型树种的根寿命年际间变异和季节变化较大(
图3A 、3B).根寿命与水分等资源的供应关系密切(
Eissenstat & Yanai, 1997 ), 实验期内2015和 2016年的年平均气温分别为19.04和19.35 ℃, 差异不显著; 但2015年的降水量2 213 mm高于2016年的1 825 mm, 表明干旱会一定程度加速根的死亡(
Mccormack & Guo, 2014 ).春季出生的吸收细根具有中值寿命较小的特点, 树种水平上具有出生于秋季>冬季而夏季>春季的季节性寿命规律(
图3B ); 夏季为生长高峰期, 观察到新生根数量最大为38.8%, 冬季为相对休眠期, 观察到的根数量只有4.8%, 基本上符合春夏两季出生, 冬季死亡(
Burtonet al ., 2000 ).在北方、温带和亚热带生物群落中, 根和地上部生长与月平均气温和月降水量呈正相关关系(
Abramoff & Finzi, 2015 ); 根叶生长具有很强的协同性, 根系萌发和停止可能更多地受到土壤温度和碳有效性限制(
Radvilleet al ., 2016 ).由于资源供应时间上的异质性, 物种间根生长的时间生态位分化, 目前数据还不足以描述亚热带树木根物候及其季节动态变化规律(
凌华等, 2011 ;
黄锦学等, 2012 ;
郑金兴等, 2012 ;
于水强等, 2020 ). ...
 ,1,*, 马泽清
,1,*, 马泽清

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT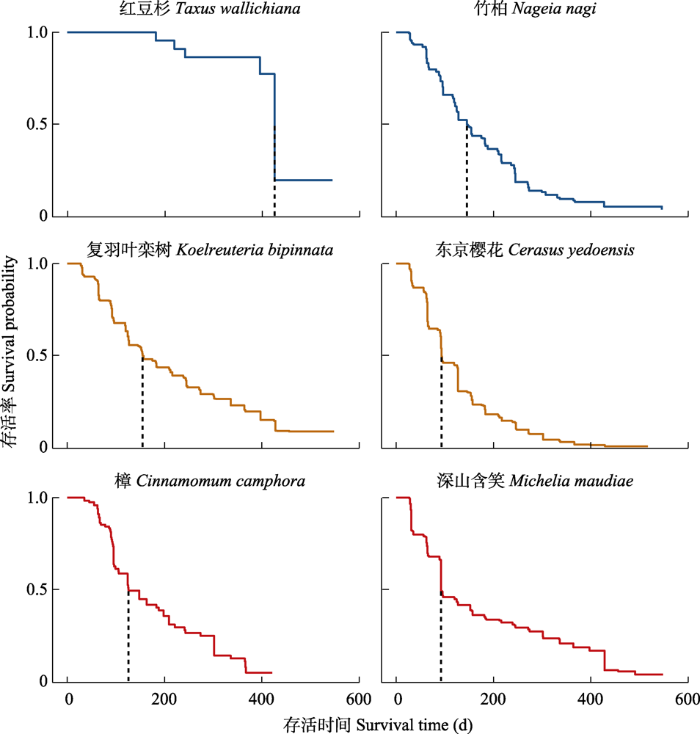 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT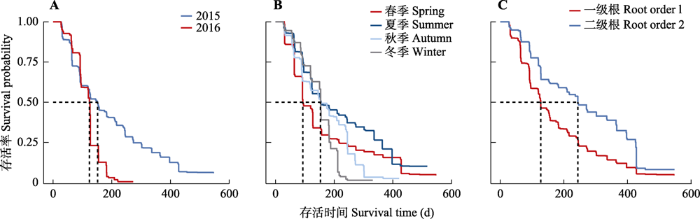 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT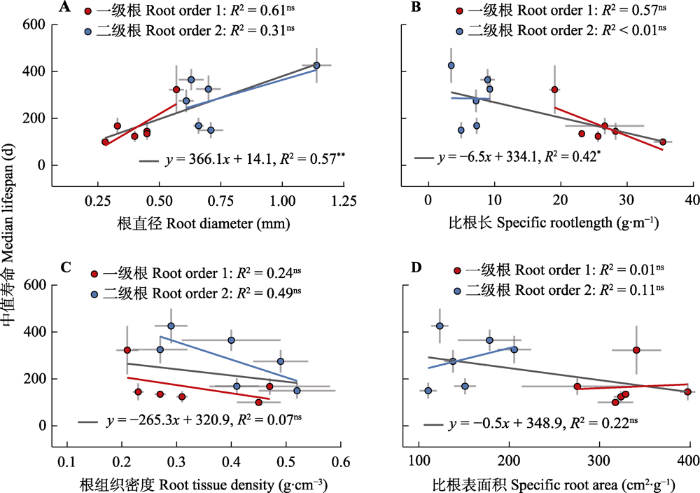 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}