National Natural Science Foundation of China(31971636) Special Foundation for National Science and Technology Basic Resources Investigation of China(2019FY202300) Young Elite Scientists Sponsorship Program by CAST(2018QNRC001)
Abstract Aims Leaf age is an important factor affecting trait variation during ontogenetic development of plants. The variation of traits across a gradient of leaf ages can reflect the response of plants to environmental changes. Our aim was to investigate the variation of needle morphological traits of Pinus koraiensis at different needle ages and the influence of latitude and needle age on the correlation of morphological traits. Methods In this study, adult Korean pine (Pinus koraiensis) of four mixed broadleaved-Korean pine forests in northeast China was selected as the research object. We measured the leaf lifespan, as well as specific leaf area (SLA), leaf dry matter content (LDMC), needle density (ND) and needle volume (NV) of needles at different ages. The correlation between morphological traits of pine leaves was discussed and the effects of latitude and needle age on these correlations were examined. Important findings (1) Leaf lifespan of P. koraiensis significantly varied with latitudes, and showed a unimodal pattern with increasing latitude. (2) Needle age accounted for the largest proportion (34.2%-80.1%) in trait variations, and had a significant influence on all the four leaf morphological traits. SLA, LDMC and ND of the current year leaves showed higher plasticity than other age classes, while NVshowed high plasticity at all age classes. (3) Most of these trait-trait correlations were significant at different latitudes, while the patterns of slopes of the regression lines among traits did vary across the latitudinal gradient. (4) The correlations among traits within different needle ages classes were all significant, but showing different patterns across needle ages. For example, the regression slope of SLA and NDdecreased with the increase of needle age, while the regression slope of ND and NV was opposite. The results showed that the variation and correlation of morphological traits were significantly influenced by latitude and needle age, the collaboration among these traits responded to the environmental changes at different latitudes, and the resource utilization strategies of needles were different in different ages. Keywords:Pinus koraiensis;needle age;latitude;specific leaf area;plasticity
PDF (988KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张自琰, 金光泽, 刘志理. 不同区域针叶年龄对红松叶性状及相关关系的影响. 植物生态学报, 2021, 45(3): 253-264. DOI: 10.17521/cjpe.2020.0352 ZHANG Zi-Yan, JIN Guang-Ze, LIU Zhi-Li. Effects of needle age on leaf traits and their correlations of Pinus koraiensis across different regions . Chinese Journal of Plant Ecology, 2021, 45(3): 253-264. DOI: 10.17521/cjpe.2020.0352
近年来, 探究区域尺度上植物功能性状的种内变异模式备受关注(Fajardo & Piper, 2011; Martin et al., 2017; Thomas et al., 2020)。植株个体在发育过程中, 其性状变化的不同可能源于遗传差异(固定个体发生)、引起表型变化的环境差异(可塑性)或两者组合(Carlson & Holsinger, 2012)。为了响应产生表型变异的进化机制, 性状在不同生态尺度上通常存在差异, 因此, 性状对不同变异驱动因素(如遗传变异、环境差异或个体发育)的敏感性差异较大(Messier et al., 2017)。表型可塑性作为一种由局部环境条件所触发的特性, 是各个组织和器官感知环境信号并对其做出反应的综合体现(De Kroon et al., 2005)。在小的分类尺度上叶片性状之间广泛的进化联系可能被可塑性模糊或逆转(Anderegg et al., 2018), 所以探究区域尺度植物叶片性状的变异规律及可塑性有助于深入了解区域范围内植物对环境的响应模式。
Dong和Liu (2017)的研究结果表明红松(Pinus koraiensis)的多个功能性状受纬度不同所导致的环境差异的影响。多个叶片性状与纬度变化密切相关, 如叶片氮磷比随纬度升高而减小, 比叶面积(SLA)和叶寿命随纬度升高而增大(张林和罗天祥, 2004; De et al., 2013)。叶寿命与叶片性状的显著相关性也多有论述, 如单位质量叶氮含量与叶寿命通常呈负相关关系(张林和罗天祥, 2004; Mediavilla et al., 2014; Osada et al., 2015), SLA随叶寿命的增大而减小(Kikuzawa & Lechowicz, 2011; de la Riva et al., 2016)。He和Yan (2018)认为常绿树种性状的二元关系强度并不高, 但已有多个研究证实在广泛的植物类型和生物群落之间叶性状存在普遍相关性(Reich et al., 1999; Wright et al., 2004; Niinemets, 2015)。红松针叶的SLA与叶片厚度呈显著的正线性相关关系, 但与叶干物质含量(LDMC)呈显著的负线性相关关系(Dong & Liu, 2017)。叶片的SLA与体积也表现为显著负相关, 且SLA较小而叶片体积大的植物资源利用性更高(王晓洁等, 2015)。叶片性状之间的关系往往反映了植物对不同环境下养分资源的适应(Chen et al., 2012), 因此, 揭示种内叶片性状的共变模式, 对于确定叶片是如何调整资源利用策略来适应环境变化是至关重要的, 但这种共变策略是否受纬度梯度的影响仍不清晰。
针叶年龄是影响性状变异及其相关关系的主要因素(Reich et al., 1999; Gratani & Bombelli, 2000)。许多物种会表现出一种被称为异胚性的发育变化(Zotz et al., 2011; Kuusk et al., 2018b), 即使是同一株植物但不同年龄的叶片形态也会存在明显差异(Grotkopp et al., 2002)。然而, 以往关于常绿树种叶性状的研究通常以完全展开的当年生叶片为研究对象, 而忽略了叶片衰老对性状的影响(Han et al., 2008)。黄海侠等(2013)通过对常绿树种当年生和多年生叶片的研究认为其SLA和LDMC因老龄叶片密度和厚度变大会发生较大变异。Kuusk等(2018a)对地中海3种松树的研究发现不同针叶年龄的针叶体积(NV)存在显著差异, 而海岸松(Pinus pinaster)的针叶密度(ND)也受到针叶年龄的显著影响(Eimil- Fraga et al., 2015)。这种不同针叶年龄之间叶片形态的差异受年际间温度和降水等环境因素的影响(曾瑞琪等, 2018), 也有可能是由发育阶段不同导致叶片生活史策略发生转变造成的(黄海侠等, 2013; Robakowski & Bielinis, 2017)。但目前在区域尺度上探究针叶年龄是如何影响红松叶性状及其相关性的研究较少。
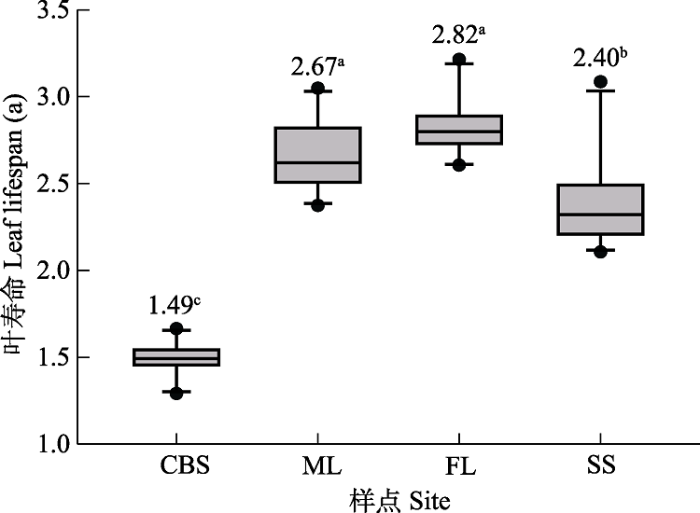
Fig. 1Comparison of leaf lifespan of Pinus koraiensis among sites across different regions. CBS, Changbai Mountain; ML, Muling; FL, Fenglin; SS, Shengshan. Different lowercase letters indicate that there were significant differences in leaf lifespan among sites (p < 0.05).
Fig. 3Variation and plasticity of leaf morphological traits of Pinus koraiensis along the gradient of needle ages (mean ± SD). LDMC, leaf dry matter content; ND, needle density; NV, needle volume; SLA, specific leaf area. Different lowercase letters indicate that the traits were significantly different at different needle ages (p < 0.05).
Table 2 表2 表2不同区域红松叶形态性状的当年生值和所有龄级平均值(标准差)的差异 Table 2Differences in leaf morphological traits of Pinus koraiensis across different regions between the mean values of the current year and those of overall age classes (SD)
样点 Site
龄级 Age classes
比叶面积 SLA (cm2·g-1)
叶干物质含量 LDMC (g·g-1)
针叶密度 ND (g·cm-3)
针叶体积 NV (cm3)
长白山 Changbai Mountain
当年生 Current year
105.6 (8.06)Ab
0.29 (0.01)Bc
0.32 (0.03)Ba
0.05 (0.01)Ac
所有龄级 Overall age
89.7 (4.29)Ba
0.36 (0.00)Ac
0.37 (0.02)Aa
0.04 (0.00)Bd
穆棱 Muling
当年生 Current year
88.0 (7.37)Ac
0.34 (0.02)Ba
0.29 (0.01)Bb
0.10 (0.02)Aa
所有龄级 Overall age
72.3 (5.24)Bc
0.40 (0.00)Aa
0.36 (0.03)Aab
0.11 (0.02)Aa
丰林 Fenglin
当年生 Current year
113.7 (9.06)Aa
0.34 (0.02)Bc
0.26 (0.01)Bc
0.07 (0.01)Bb
所有龄级 Overall age
85.3 (8.17)Ba
0.38 (0.01)Ab
0.34 (0.02)Ab
0.08 (0.01)Ab
胜山 Shengshan
当年生 Current year
104.2 (7.26)Ab
0.31 (0.01)Bb
0.32 (0.03)Ba
0.07 (0.01)Ab
所有龄级 Overall age
78.1 (5.27)Bb
0.37 (0.02)Abc
0.36 (0.03)Aab
0.06 (0.01)Bc
可塑性 Plasticity (%)
当年生 Current year
22.6
16.4
19.4
51.5
所有龄级 Overall age
19.4
9.5
8.9
64.5
LDMC, 叶干物质含量; ND, 针叶密度; NV, 针叶体积; SLA, 比叶面积。所有龄级包括当年生。不同大写字母表示同一地点性状的当年生值和所有龄级均值差异显著, 不同小写字母表示性状的当年生值或所有龄级均值在4个样点间差异显著(p< 0.05)。 LDMC, leaf dry matter content; ND, needle density; NV, needle volume;SLA, specific leaf area. Overall years includes current year. Different uppercase letters indicate that there are significant differences between the mean values of the current year and those of overall age classes in the same site. Different lowercase letters indicate that the mean values of the current year or those of overall age classes differ significantly among the four sites (p< 0.05).
Fig. 4Effects of latitude on correlations between leaf morphological traits of Pinus koraiensis. Different lowercase letters indicate that there were significant differences between the slopes of different sites, and the solid lines indicate that the regressions were significant (p < 0.05). CBS, Changbai Mountain; FL, Fenglin; ML, Muling; SS, Shengshan. LDMC, leaf dry matter content; ND, needle density; NV, needle volume; SLA, specific leaf area.
Fig. 5Effects of needle age on correlations between leaf morphological traits of Pinus koraiensis. Different lowercase letters indicate that there were significant differences between the slopes of different needle ages. The solid lines indicate that the regressions were significant (p < 0.05). LDMC, leaf dry matter content; ND, needle density; NV, needle volume; SLA, specific leaf area.
王瑞丽等(2015)对中国东部南北样带的多种植物调查分析后发现SLA和LDMC存在明显的纬度格局, 而在本研究中红松4个叶形态性状在不同纬度梯度虽大多有显著差异, 但并未表现出规律性(表2), 可能是前者研究区域范围的纬度跨度更大、气候条件差异显著导致的结果。本研究中穆棱样点的叶寿命在4个样点中相对较长, 但SLA最小且与其他样点存在显著差异(表2), 这与SLA低则叶寿命长的普遍结论(Gratani & Bombelli, 2000; de la Riva et al., 2016; Onoda et al., 2017)相符合。Withington等(2006)基于全球数据也得到相似的结论, 即具有厚叶、低SLA、低单位质量氮浓度和低光合速率的物种通常表现出较长的叶寿命。叶片具有较高的SLA通常单位叶面积所需的碳投资很少, 生理上更活跃, 所以低SLA的叶片具有较长的叶寿命可以在防御结构中每单位叶面积投入更多的碳, 使它们更能抵御物理胁迫和食草动物啃食(Sakschewski et al., 2015)。纬度最高的胜山样点红松的NV最小ND最大(表2), 这可能是资源的生产能力和存储量受低温环境限制而不足, 因此NV变小, 红松叶片投资策略转向防御并选择了更保守的叶片性状来缓冲低温对生长的影响(Buzzard et al., 2019), 且NV的变异速率大于叶干质量增加的速率导致ND变大。
3.2 针叶年龄对叶形态性状的影响
针叶年龄对针长、针厚以及针叶总投影面积等性状有显著影响(Eimil-Fraga et al., 2015), 本研究结果强调了针叶年龄在解释红松叶形态性状变异中的重要地位(图2)。红松针叶的SLA随针叶年龄增加而减小(图3), 这与以往研究结果(Anderegg et al., 2018; Liu et al., 2020)相符。植物通过合成不同化合物从而以不同化学成分和结构来分配能量(Villar & Merino, 2001), 韧皮部的二次生长、薄壁组织细胞酚类衍生物的增加和碳汇减少(Gilmore et al., 1995; Kuusk et al., 2018b)都是导致SLA随针叶年龄增加而减小的重要原因。另外, 叶片年龄尚小时维管束的体积较大进而叶片密度增大, 也会导致SLA的变化(Grotkopp et al., 2002)。LDMC和ND均随针叶年龄增加而增大(图3), 与Eimil-Fraga等(2015)和Kuusk等(2018a)对海岸松和意大利松(Pinus pinea)等树种的研究结果相符。LDMC可以指示结构细胞含量与液体细胞含量(Kikuzawa & Lechowicz, 2011), 对细胞壁投入更多以及老叶的代谢物沉积是LDMC随针叶年龄增加而增大的主要原因(黄海侠等, 2013; Kuusk et al., 2018b)。而叶片在年龄增加的过程中为了增加叶片韧性, 空气占据的空间逐渐减小而木质化组织大量增加(Evans & Poorter, 2001; Kuusk et al., 2018a; Poorter et al., 2009), 这可能是导致ND随针叶年龄增加而增大的重要原因。Kuusk等(2018b)的研究结果表明幼年针叶的NV更小, 本研究结果与之相符, 但增加到一定程度后NV又有所下降, 这可能是因为老叶不仅是常绿乔木的同化器官还是同化产物的贮藏器官(Suzuki & Takahashi, 2020), 叶片年龄越大可利用资源存储越多, 而叶片体积作为反映植物本身对资源利用能力的综合指标(王晓洁等, 2015), 表现出随年龄增加而减小的趋势, 说明了红松老叶对资源有效利用的能力下降。
叶经济谱性状很大程度上保留了各自独立响应表型变异驱动因素的能力, 驱动因素的影响因尺度而异(Martin et al., 2017)。作为叶经济谱的一个重要性状, 很多研究已证明SLA对种内变异和环境普遍敏感(Adler et al., 2014; Messier et al., 2017)。SLA可塑性相对较大, LDMC的可塑性低于SLA (表2), 这与de la Riva等(2016)的研究结论相符。当年生叶片与所有龄级表现的可塑性大小不同, 这可能是不同年龄针叶之间的光适应能力差异导致的, 尤其是当年生针叶容易受日照强度影响做出适应性调整(Wyka et al., 2007)。不同针叶年龄下响应策略不同也可能是叶片可塑性大小存在较大差异的原因, De Kroon等(2005)曾提出, 在生长发育阶段植物做出的可塑性响应分别包括决定分生组织的营养或生殖去向以及对植物各部分尺寸和形状的调整, 而在成熟阶段表达为诱导防御等。
当年生叶片与所有龄级叶片间的性状存在显著差异(表2), 这与以往研究结论相符。Mediavilla和Escudero (2003)曾在叶片寿命与光合特性的线性关系中发现, 用所有龄级平均值计算比当年生叶片数据得到的斜率更陡。黄海侠等(2013)也确定了当年生叶片的性状值较为发散, 往年生叶片的性状值更为聚拢和稳定。导致这种差异的主要原因可能是叶片发育阶段的不同致使投资策略不同(Climent et al., 2009), 幼年叶片倾向于通过增大光合效率积累生物量, 而成年叶片以支撑结构的投资为主, 投资策略的调整可能是受叶片发育期间生长条件的影响(Gilmore et al., 1995)而形成的响应机制。
3.3 纬度和针叶年龄对叶性状之间相关关系的影响
样点对叶性状变异及相关关系有一定的影响, 尤其对由叶片构建期间的环境波动所决定的形态性状影响很大(Reich et al., 1999; Martin et al., 2017), 而王瑞丽等(2015)的研究显示来源于样地间环境差异的叶功能属性的变异不到1/3。Anderegg等(2018)对美国西北地区进行调查后发现, 地块级别的环境特征并不是种间、种内或性状间变异的强预测因子。本研究结果证明样点(纬度)是叶片种内性状变异的原因之一, 但对除NV以外其他3个性状变异的解释占比很小(图2)。
AdlerPB, Salguero-GómezR, CompagnoniA, HsuJS, Ray-MukherjeeJ, Mbeau-AcheC, FrancoM (2014). Functional traits explain variation in plant life history strategies Proceedings of the National Academy of Sciences of the United States of America, 111, 740-745. [本文引用: 1]
AndereggLDL, BernerLT, BadgleyG, SethiML, LawBE, HilleRisLambersJ (2018). Within-species patterns challenge our understanding of the leaf economics spectrum Ecology Letters, 21, 734-744. DOI:10.1111/ele.12945PMID:29569818 [本文引用: 3] The utility of plant functional traits for predictive ecology relies on our ability to interpret trait variation across multiple taxonomic and ecological scales. Using extensive data sets of trait variation within species, across species and across communities, we analysed whether and at what scales leaf economics spectrum (LES) traits show predicted trait-trait covariation. We found that most variation in LES traits is often, but not universally, at high taxonomic levels (between families or genera in a family). However, we found that trait covariation shows distinct taxonomic scale dependence, with some trait correlations showing opposite signs within vs. across species. LES traits responded independently to environmental gradients within species, with few shared environmental responses across traits or across scales. We conclude that, at small taxonomic scales, plasticity may obscure or reverse the broad evolutionary linkages between leaf traits, meaning that variation in LES traits cannot always be interpreted as differences in resource use strategy.? 2018 John Wiley & Sons Ltd/CNRS.
CarlsonJE, HolsingerKE (2012). Developmental plasticity in Protea as an evolutionary response to environmental clines in the Cape Floristic Region PLOS ONE, 7, e52035. DOI: 10.1371/journal.pone.0052035. DOI:10.1371/journal.pone.0052035URL [本文引用: 1]
ChenFS, NiklasKJ, ChenGS, GuoDL (2012). Leaf traits and relationships differ with season as well as among species groupings in a managed Southeastern China forest landscape Plant Ecology, 213, 1489-1502. DOI:10.1007/s11258-012-0106-5URL [本文引用: 1]
ClimentJ, SilvaFCE, ChambelMR, PardosM, AlmeidaMH (2009). Freezing injury in primary and secondary needles of mediterranean pine species of contrasting ecological niches Annals of Forest Science, 66, 407-407. DOI:10.1051/forest/2009016URL [本文引用: 1]
DeFrenne P, GraaeBJ, Rodríguez-SánchezF, KolbA, ChabrerieO, DecocqG, DeKort H, DeSchrijver A, DiekmannM, ErikssonO, GruwezR, HermyM, LenoirJ, PlueJ, CoomesDA, VerheyenK (2013). Latitudinal gradients as natural laboratories to infer species’ responses to temperature Journal of Ecology, 101, 784-795. DOI:10.1111/jec.2013.101.issue-3URL [本文引用: 1]
DeKroon H, HuberH, StueferJF, van GroenendaelJM (2005). A modular concept of phenotypic plasticity in plants New Phytologist, 166, 73-82. PMID:15760352 Based on empirical evidence from the literature we propose that, in nature, phenotypic plasticity in plants is usually expressed at a subindividual level. While reaction norms (i.e. the type and the degree of plant responses to environmental variation) are a property of genotypes, they are expressed at the level of modular subunits in most plants. We thus contend that phenotypic plasticity is not a whole-plant response, but a property of individual meristems, leaves, branches and roots, triggered by local environmental conditions. Communication and behavioural integration of interconnected modules can change the local responses in different ways: it may enhance or diminish local plastic effects, thereby increasing or decreasing the differences between integrated modules exposed to different conditions. Modular integration can also induce qualitatively different responses, which are not expressed if all modules experience the same conditions. We propose that the response of a plant to its environment is the sum of all modular responses to their local conditions plus all interaction effects that are due to integration. The local response rules to environmental variation, and the modular interaction rules may be seen as evolving traits targeted by natural selection. Following this notion, whole-plant reaction norms are an integrative by-product of modular plasticity, which has far-reaching methodological, ecological and evolutionary implications.Copyright New Phytologist (2005).
dela Riva EG, OlmoM, PoorterH, UberaJL, VillarR (2016). Leaf mass per area (LMA) and its relationship with leaf structure and anatomy in 34 Mediterranean woody species along a water availability gradient PLOS ONE, 11, e0148788. DOI:10.1371/journal.pone.0148788URL
DongYC, LiuYH (2017). Response of Korean pine’s functional traits to geography and climate PLOS ONE, 12, e0184051. DOI: 10.1371/journal.pone.0184051. DOI:10.1371/journal.pone.0184051 [本文引用: 3]
Eimil-FragaC, Sánchez-RodríguezF, álvarez-RodríguezE, Rodríguez-SoalleiroR (2015). Relationships between needle traits, needle age and site and stand parameters in Pinus pinaster Trees, 29, 1103-1113. DOI:10.1007/s00468-015-1190-7URL [本文引用: 2]
EvansJR, PoorterH (2001). Photosynthetic acclimation of plants to growth irradiance: the relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain Plant, Cell & Environment, 24, 755-767. [本文引用: 1]
FajardoA, PiperFI (2011). Intraspecific trait variation and covariation in a widespread tree species (Nothofagus pumilio) in southern Chile New Phytologist, 189, 259-271. DOI:10.1111/nph.2010.189.issue-1URL [本文引用: 2]
GenetH, BredaN, DufreneE (2010). Age-related variation in carbon allocation at tree and stand scales in beech (Fagus sylvatica L.) and sessile oak (Quercus petraea (Matt.) Liebl.) using a chronosequence approach Tree Physiology, 30, 177-192. DOI:10.1093/treephys/tpp105URL [本文引用: 1]
GilmoreDW, SeymourRS, HaltemanWA, GreenwoodMS (1995). Canopy dynamics and the morphological development of Abies balsamea: effects of foliage age on specific leaf area and secondary vascular development Tree Physiology, 15, 47-55. PMID:14966011 [本文引用: 2] Data were collected from two branches from each whorl of nine open-grown Abies balsamea (L.) Miller trees to test the hypothesis that specific leaf area (SLA, m(2) projected fresh leaf area kg(-1) oven-dry foliage) is constant among five foliage age classes (current-year, 1-year-old, 2-year-old, 3-year-old and 4-year-old-plus). Between-tree variation in SLA was greater than within-tree variation. Differences in SLA among the foliage age classes were small, but statistically significant, showing a trend of decreasing SLA with increasing foliage age. Using data from two previous biomass studies, we found that three different methods of calculating SLA of individual trees produced the same projected leaf area estimates. To test the hypothesis that foliage mass increases with foliage age as a result of secondary xylem or phloem development, we examined the secondary vascular development of foliage collected from five age classes and three crown sections in an open-grown A. balsamea. The number of rows of xylem cells was not constant among foliage age classes, but the differences were small and showed no consistent pattern of change with foliage age. Total number of rows of phloem cells increased, number of living rows of phloem cells decreased, and the number of rows of nonliving crushed phloem cells increased with foliage age.
GrataniL, BombelliA (2000). Correlation between leaf age and other leaf traits in three Mediterranean maquis shrub species: Quercus ilex, Phillyrea latifolia and Cistus incanus Environmental and Experimental Botany, 43, 141-153. DOI:10.1016/S0098-8472(99)00052-0URL [本文引用: 2]
GrotkoppE, RejmánekM, RostTL (2002). Toward a causal explanation of plant invasiveness: seedling growth and life-history strategies of 29 pine (Pinus) species The American Naturalist, 159, 396-419. DOI:10.1086/338995PMID:18707424 [本文引用: 2] We studied 29 pine (Pinus) species to test the hypothesis that invasive species in disturbed habitats have distinct attributes. Seedling relative growth rate (RGR) and measures of invasiveness were positively associated across species as well as within phylogenetically independent contrasts. High RGR, small seed masses, and short generation times characterize pine species that are successful invaders in disturbed habitats. Discriminant analysis and logistic regression revealed that RGR was the most significant factor among these life-history traits separating invasive and noninvasive species. We also explored the causes of differences in RGR among invasive and noninvasive species. While net assimilation rate, leaf mass ratio, and specific leaf area (SLA) were all found to be contributing positively to RGR, SLA was found to be the main component responsible for differences in RGR between invasive and noninvasive pines. We investigated differences in SLA further by studying leaf anatomy, leaf density, and leaf thickness. We also evaluated relative leaf production rate as an important aspect of SLA. We proposed a hypothetical causal network of all relevant variables.
HanQM, KawasakiT, NakanoT, ChibaY (2008). Leaf-age effects on seasonal variability in photosynthetic parameters and its relationships with leaf mass per area and leaf nitrogen concentration within a Pinus densiflora crown Tree Physiology, 28, 551-558. DOI:10.1093/treephys/28.4.551URL [本文引用: 1]
HeD, YanER (2018). Size-dependent variations in individual traits and trait scaling relationships within a shade-tolerant evergreen tree species American Journal of Botany, 105, 1165-1174. DOI:10.1002/ajb2.2018.105.issue-7URL [本文引用: 1]
HuangHX, YangXD, SunBW, ZhangZH, YanER (2013). Variability and association of leaf traits between current- year and former-year leaves in evergreen trees in Tiantong, Zhejiang, China Chinese Journal of Plant Ecology, 37, 912-921. DOI:10.3724/SP.J.1258.2013.00094URL [本文引用: 5]
HughesSW (2005). Archimedes revisited: a faster, better, cheaper method of accurately measuring the volume of small objects Physics Education, 40, 468-474. DOI:10.1088/0031-9120/40/5/008URL [本文引用: 1]
HulshofCM, SwensonNG (2010). Variation in leaf functional trait values within and across individuals and species: an example from a Costa Rican dry forest Functional Ecology, 24, 217-223. DOI:10.1111/fec.2010.24.issue-1URL [本文引用: 1]
KayamaM, SasaK, KoikeT (2002). Needle life span, photosynthetic rate and nutrient concentration of Picea glehnii, P. jezoensis and P. abies planted on serpentine soil in northern Japan Tree Physiology, 22, 707-716. DOI:10.1093/treephys/22.10.707URL [本文引用: 1]
KikuzawaK, LechowiczMJ (2011). Theories of Leaf Longevity. Ecological Research Monographs. Springer, Tokyo.41-56. [本文引用: 5]
KuuskV, Niinemetsü, ValladaresF (2018a). A major trade-off between structural and photosynthetic investments operative across plant and needle ages in three Mediterranean pines Tree Physiology, 38, 543-557. DOI:10.1093/treephys/tpx139URL [本文引用: 3]
KuuskV, Niinemetsü, ValladaresF (2018b). Structural controls on photosynthetic capacity through juvenile-to-adult transition and needle ageing in Mediterranean pines Functional Ecology, 32, 1479-1491. DOI:10.1111/fec.2018.32.issue-6URL [本文引用: 5]
LiMH, Kr?uchiN, DobbertinM (2006). Biomass distribution of different-aged needles in young and old Pinus cembra trees at highland and lowland sites Trees, 20, 611-618. DOI:10.1007/s00468-006-0076-0URL [本文引用: 1]
LiuZL, HikosakaK, LiFR, JinGZ (2020). Variations in leaf economics spectrum traits for an evergreen coniferous species: tree size dominates over environment factors Functional Ecology, 34, 458-467. DOI:10.1111/fec.v34.2URL [本文引用: 2]
LiuZL, JinGZ, ZhouM (2014). Measuring seasonal dynamics of leaf area index in a mixed conifer-broadleaved forest with direct and indirect methods Chinese Journal of Plant Ecology, 38, 843-856. DOI:10.3724/SP.J.1258.2014.00079URL [本文引用: 1]
MarshallJD, MonserudRA (2003). Foliage height influences specific leaf area of three conifer species Canadian Journal of Forest Research, 33, 164-170. DOI:10.1139/x02-158URL [本文引用: 2]
MartinAR, RapidelB, RoupsardO, vanden Meersche K, deMelo Virginio Filho E, BarriosM, IsaacME (2017). Intraspecific trait variation across multiple scales: the leaf economics spectrum in coffee Functional Ecology, 31, 604-612. DOI:10.1111/fec.2017.31.issue-3URL [本文引用: 3]
MediavillaS, EscuderoA (2003). Photosynthetic capacity, integrated over the lifetime of a leaf, is predicted to be independent of leaf longevity in some tree species New Phytologist, 159, 203-211. DOI:10.1046/j.1469-8137.2003.00798.xURL [本文引用: 1]
MediavillaS, HerranzM, González-ZurdoP, EscuderoA (2014). Ontogenetic transition in leaf traits: a new cost associated with the increase in leaf longevity Journal of Plant Ecology, 7, 567-575. DOI:10.1093/jpe/rtt059URL [本文引用: 1]
MessierJ, McGillBJ, EnquistBJ, LechowiczMJ (2017). Trait variation and integration across scales: Is the leaf economic spectrum present at local scales Ecography, 40, 685-697. DOI:10.1111/ecog.02006URL [本文引用: 3]
Niinemetsü (2015). Is there a species spectrum within the world-wide leaf economics spectrum? Major variations in leaf functional traits in the Mediterranean sclerophyll Quercus ilex New Phytologist, 205, 79-96. DOI:10.1111/nph.2014.205.issue-1URL [本文引用: 1]
Niinemetsü, CescattiA, RodeghieroM, TosensT (2005). Leaf internal diffusion conductance limits photosynthesis more strongly in older leaves of Mediterranean evergreen broad-leaved species Plant, Cell & Environment, 28, 1552-1566. [本文引用: 1]
OnodaY, WrightIJ, EvansJR, HikosakaK, KitajimaK, Niinemetsü, PoorterH, TosensT, WestobyM (2017). Physiological and structural tradeoffs underlying the leaf economics spectrum New Phytologist, 214, 1447-1463. DOI:10.1111/nph.2017.214.issue-4URL [本文引用: 1]
OsadaN, NabeshimaE, HiuraT (2015). Geographic variation in shoot traits and branching intensity in relation to leaf size in Fagus crenata: a common garden experiment American Journal of Botany, 102, 878-887. DOI:10.3732/ajb.1400559URL [本文引用: 1]
ParadisE, ClaudeJ, StrimmerK (2004). APE: analyses of phylogenetics and evolution in R language Bioinformatics, 20, 289-290. DOI:10.1093/bioinformatics/btg412URL [本文引用: 1]
PoorterH, Niinemetsü, PoorterL, WrightIJ, VillarR (2009). Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis New Phytologist, 182, 565-588. DOI:10.1111/nph.2009.182.issue-3URL [本文引用: 1]
RCore Team (2017). R: a language and environment for statistical computing R Foundation for Statistical Computing, Vienna, Austria. [本文引用: 1]
ReichPB, OleksynJ, ModrzynskiJ, TjoelkerMG (1996). Evidence that longer needle retention of spruce and pine populations at high elevations and high latitudes is largely a phenotypic response Tree Physiology, 16, 643-647. PMID:14871702 [本文引用: 1] There is abundant evidence that evergreen conifers living at high elevations or at high latitudes have longer-lived needles than trees of the same species living elsewhere. This pattern is likely caused by the influence of low temperature in combination with related factors such as a short growing season and low nutrient availability. Because it is not known to what degree such patterns result from phenotypic versus genotypic variation, we evaluated needle longevity for common-garden-grown lowland populations of European Scots pine (Pinus sylvestris L.) of wide latitudinal origin and Norway spruce (Picea abies L.) of wide elevational origin. Nine-year-old trees of 16 Scots pine populations ranging in origin from 47 degrees to 60 degrees N were studied in Kórnik, Poland (52 degrees N) and 18-year-old trees of 18 Norway spruce populations ranging in origin from 670 to 1235 m elevation in southwestern Poland were studied near Morawina, Poland (51 degrees N, 180 m elevation). There was no tendency in either species for populations from northern or high elevation origins to retain needles longer than other populations. All of the Scots pine populations had between 2.5 to 3.0 needle age cohorts and all of the Norway spruce populations had between 6.4 and 7.2 needle age cohorts. Thus, extended needle retention in Scots pine and Norway spruce populations in low-temperature habitats at high elevations and high latitudes appears to be largely an environmentally regulated phenotypic acclimation.
RobakowskiP, BielinisE (2017). Needle age dependence of photosynthesis along a light gradient within an Abies alba crown Acta Physiologiae Plantarum, 39, 1-12. DOI:10.1007/s11738-016-2300-xURL [本文引用: 1]
SakschewskiB, vonBloh W, BoitA, RammigA, KattgeJ, PoorterL, Pe?uelasJ, ThonickeK (2015). Leaf and stem economics spectra drive diversity of functional plant traits in a dynamic global vegetation model Global Change Biology, 21, 2711-2725. DOI:10.1111/gcb.12870PMID:25611734 [本文引用: 1] Functional diversity is critical for ecosystem dynamics, stability and productivity. However, dynamic global vegetation models (DGVMs) which are increasingly used to simulate ecosystem functions under global change, condense functional diversity to plant functional types (PFTs) with constant parameters. Here, we develop an individual- and trait-based version of the DGVM LPJmL (Lund-Potsdam-Jena managed Land) called LPJmL- flexible individual traits (LPJmL-FIT) with flexible individual traits) which we apply to generate plant trait maps for the Amazon basin. LPJmL-FIT incorporates empirical ranges of five traits of tropical trees extracted from the TRY global plant trait database, namely specific leaf area (SLA), leaf longevity (LL), leaf nitrogen content (N ), the maximum carboxylation rate of Rubisco per leaf area (vcmaxarea), and wood density (WD). To scale the individual growth performance of trees, the leaf traits are linked by trade-offs based on the leaf economics spectrum, whereas wood density is linked to tree mortality. No preselection of growth strategies is taking place, because individuals with unique trait combinations are uniformly distributed at tree establishment. We validate the modeled trait distributions by empirical trait data and the modeled biomass by a remote sensing product along a climatic gradient. Including trait variability and trade-offs successfully predicts natural trait distributions and achieves a more realistic representation of functional diversity at the local to regional scale. As sites of high climatic variability, the fringes of the Amazon promote trait divergence and the coexistence of multiple tree growth strategies, while lower plant trait diversity is found in the species-rich center of the region with relatively low climatic variability. LPJmL-FIT enables to test hypotheses on the effects of functional biodiversity on ecosystem functioning and to apply the DGVM to current challenges in ecosystem management from local to global scales, that is, deforestation and climate change effects.? 2015 John Wiley & Sons Ltd.
SuzukiR, TakahashiK (2020). Effects of leaf age, elevation and light conditions on photosynthesis and leaf traits in saplings of two evergreen conifers, Abies veitchii and A. mariesii Journal of Plant Ecology, 13, 460-469. DOI:10.1093/jpe/rtaa034URL [本文引用: 3]
ThomasHJD, BjorkmanAD, Myers-SmithIH, ElmendorfSC, KattgeJ, DiazS, VellendM, BlokD, CornelissenJHC, ForbesBC, HenryGHR, HollisterRD, NormandS, PrevéyJS, RixenC, et al. (2020). Global plant trait relationships extend to the climatic extremes of the tundra biome Nature Communications, 11, 1351. DOI: 10.1038/s41467-020-15014-4. DOI:10.1038/s41467-020-15014-4 [本文引用: 1]
VillarR, MerinoJ (2001). Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems New Phytologist, 151, 213-226. DOI:10.1046/j.1469-8137.2001.00147.xURL [本文引用: 1]
WangHL, ZhangY, XiaXL, YinWL, GuoHW, LiZH (2020). Research advances in leaf senescence of woody plants Scientia Sinica (Vitae), 50, 196-206. [本文引用: 1]
WangN, PalmrothS, MaierCA, DomecJC, OrenR (2019). Anatomical changes with needle length are correlated with leaf structural and physiological traits across five Pinus species Plant, Cell & Environment, 42, 1690-1704. [本文引用: 1]
WangRL, YuGR, HeNP, WangQF, ZhaoN, XuZW (2015). Latitudinal patterns and influencing factors of leaf functional traits in Chinese forest ecosystems Acta Geographica Sinica, 70, 1735-1746. [本文引用: 2]
WangXJ, ZhangK, XiaoD, HouJH (2015). Leaf traits and their interrelationships of main plant species in Liangshui natural broadleaved Korean pine mixed forest Journal of Central South University of Forestry & Technology, 35(9), 52-58. [本文引用: 3]
WrightIJ, CannonK (2001). Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora Functional Ecology, 15, 351-359. DOI:10.1046/j.1365-2435.2001.00522.xURL
WrightIJ, ReichPB, WestobyM, AckerlyDD, BaruchZ, BongersF, Cavender-BaresJ, ChapinT, CornelissenJHC, DiemerM, FlexasJ, GarnierE, GroomPK, GuliasJ, HikosakaK, et al. (2004). The worldwide leaf economics spectrum Nature, 428, 821-827. PMID:15103368 [本文引用: 1] Bringing together leaf trait data spanning 2,548 species and 175 sites we describe, for the first time at global scale, a universal spectrum of leaf economics consisting of key chemical, structural and physiological properties. The spectrum runs from quick to slow return on investments of nutrients and dry mass in leaves, and operates largely independently of growth form, plant functional type or biome. Categories along the spectrum would, in general, describe leaf economic variation at the global scale better than plant functional types, because functional types overlap substantially in their leaf traits. Overall, modulation of leaf traits and trait relationships by climate is surprisingly modest, although some striking and significant patterns can be seen. Reliable quantification of the leaf economics spectrum and its interaction with climate will prove valuable for modelling nutrient fluxes and vegetation boundaries under changing land-use and climate.
WykaT, RobakowskiP, ?ytkowiakR (2007). Leaf age as a factor in anatomical and physiological acclimative responses of Taxus baccata L. needles to contrasting irradiance environments Photosynthesis Research, 95, 87-99. DOI:10.1007/s11120-007-9238-1URL [本文引用: 1]
WykaTP, OleksynJ (2014). Photosynthetic ecophysiology of evergreen leaves in the woody angiosperms: a review Dendrobiology, 72, 3-27. DOI:10.12657/denbio.072.001URL [本文引用: 1]
YangDM, ZhangJJ, ZhouD, QianMJ, ZhengY, JinLM (2012). Leaf and twig functional traits of woody plants and their relationships with environmental change: a review Chinese Journal of Ecology, 31, 702-713. [本文引用: 1]
ZengRQ, ZhaoJG, LiuYZ, ChenGP, XianJR (2018). Long-term warming effects on leaf traits of different-aged needles of Abies faxoniana seedlings in a treeline ecotone in western Sichuan Acta Ecologica Sinica, 38, 4008-4017. [本文引用: 2]
Functional traits explain variation in plant life history strategies 1 2014
... 叶经济谱性状很大程度上保留了各自独立响应表型变异驱动因素的能力, 驱动因素的影响因尺度而异(Martin et al., 2017).作为叶经济谱的一个重要性状, 很多研究已证明SLA对种内变异和环境普遍敏感(Adler et al., 2014; Messier et al., 2017).SLA可塑性相对较大, LDMC的可塑性低于SLA (表2), 这与de la Riva等(2016)的研究结论相符.当年生叶片与所有龄级表现的可塑性大小不同, 这可能是不同年龄针叶之间的光适应能力差异导致的, 尤其是当年生针叶容易受日照强度影响做出适应性调整(Wyka et al., 2007).不同针叶年龄下响应策略不同也可能是叶片可塑性大小存在较大差异的原因, De Kroon等(2005)曾提出, 在生长发育阶段植物做出的可塑性响应分别包括决定分生组织的营养或生殖去向以及对植物各部分尺寸和形状的调整, 而在成熟阶段表达为诱导防御等. ...
Within-species patterns challenge our understanding of the leaf economics spectrum 3 2018
... 近年来, 探究区域尺度上植物功能性状的种内变异模式备受关注(Fajardo & Piper, 2011; Martin et al., 2017; Thomas et al., 2020).植株个体在发育过程中, 其性状变化的不同可能源于遗传差异(固定个体发生)、引起表型变化的环境差异(可塑性)或两者组合(Carlson & Holsinger, 2012).为了响应产生表型变异的进化机制, 性状在不同生态尺度上通常存在差异, 因此, 性状对不同变异驱动因素(如遗传变异、环境差异或个体发育)的敏感性差异较大(Messier et al., 2017).表型可塑性作为一种由局部环境条件所触发的特性, 是各个组织和器官感知环境信号并对其做出反应的综合体现(De Kroon et al., 2005).在小的分类尺度上叶片性状之间广泛的进化联系可能被可塑性模糊或逆转(Anderegg et al., 2018), 所以探究区域尺度植物叶片性状的变异规律及可塑性有助于深入了解区域范围内植物对环境的响应模式. ...
... 针叶年龄对针长、针厚以及针叶总投影面积等性状有显著影响(Eimil-Fraga et al., 2015), 本研究结果强调了针叶年龄在解释红松叶形态性状变异中的重要地位(图2).红松针叶的SLA随针叶年龄增加而减小(图3), 这与以往研究结果(Anderegg et al., 2018; Liu et al., 2020)相符.植物通过合成不同化合物从而以不同化学成分和结构来分配能量(Villar & Merino, 2001), 韧皮部的二次生长、薄壁组织细胞酚类衍生物的增加和碳汇减少(Gilmore et al., 1995; Kuusk et al., 2018b)都是导致SLA随针叶年龄增加而减小的重要原因.另外, 叶片年龄尚小时维管束的体积较大进而叶片密度增大, 也会导致SLA的变化(Grotkopp et al., 2002).LDMC和ND均随针叶年龄增加而增大(图3), 与Eimil-Fraga等(2015)和Kuusk等(2018a)对海岸松和意大利松(Pinus pinea)等树种的研究结果相符.LDMC可以指示结构细胞含量与液体细胞含量(Kikuzawa & Lechowicz, 2011), 对细胞壁投入更多以及老叶的代谢物沉积是LDMC随针叶年龄增加而增大的主要原因(黄海侠等, 2013; Kuusk et al., 2018b).而叶片在年龄增加的过程中为了增加叶片韧性, 空气占据的空间逐渐减小而木质化组织大量增加(Evans & Poorter, 2001; Kuusk et al., 2018a; Poorter et al., 2009), 这可能是导致ND随针叶年龄增加而增大的重要原因.Kuusk等(2018b)的研究结果表明幼年针叶的NV更小, 本研究结果与之相符, 但增加到一定程度后NV又有所下降, 这可能是因为老叶不仅是常绿乔木的同化器官还是同化产物的贮藏器官(Suzuki & Takahashi, 2020), 叶片年龄越大可利用资源存储越多, 而叶片体积作为反映植物本身对资源利用能力的综合指标(王晓洁等, 2015), 表现出随年龄增加而减小的趋势, 说明了红松老叶对资源有效利用的能力下降. ...
... 样点对叶性状变异及相关关系有一定的影响, 尤其对由叶片构建期间的环境波动所决定的形态性状影响很大(Reich et al., 1999; Martin et al., 2017), 而王瑞丽等(2015)的研究显示来源于样地间环境差异的叶功能属性的变异不到1/3.Anderegg等(2018)对美国西北地区进行调查后发现, 地块级别的环境特征并不是种间、种内或性状间变异的强预测因子.本研究结果证明样点(纬度)是叶片种内性状变异的原因之一, 但对除NV以外其他3个性状变异的解释占比很小(图2). ...
Continental scale structuring of forest and soil diversity via functional traits 1 2019
... 王瑞丽等(2015)对中国东部南北样带的多种植物调查分析后发现SLA和LDMC存在明显的纬度格局, 而在本研究中红松4个叶形态性状在不同纬度梯度虽大多有显著差异, 但并未表现出规律性(表2), 可能是前者研究区域范围的纬度跨度更大、气候条件差异显著导致的结果.本研究中穆棱样点的叶寿命在4个样点中相对较长, 但SLA最小且与其他样点存在显著差异(表2), 这与SLA低则叶寿命长的普遍结论(Gratani & Bombelli, 2000; de la Riva et al., 2016; Onoda et al., 2017)相符合.Withington等(2006)基于全球数据也得到相似的结论, 即具有厚叶、低SLA、低单位质量氮浓度和低光合速率的物种通常表现出较长的叶寿命.叶片具有较高的SLA通常单位叶面积所需的碳投资很少, 生理上更活跃, 所以低SLA的叶片具有较长的叶寿命可以在防御结构中每单位叶面积投入更多的碳, 使它们更能抵御物理胁迫和食草动物啃食(Sakschewski et al., 2015).纬度最高的胜山样点红松的NV最小ND最大(表2), 这可能是资源的生产能力和存储量受低温环境限制而不足, 因此NV变小, 红松叶片投资策略转向防御并选择了更保守的叶片性状来缓冲低温对生长的影响(Buzzard et al., 2019), 且NV的变异速率大于叶干质量增加的速率导致ND变大. ...
Developmental plasticity in Protea as an evolutionary response to environmental clines in the Cape Floristic Region 1 2012
... 近年来, 探究区域尺度上植物功能性状的种内变异模式备受关注(Fajardo & Piper, 2011; Martin et al., 2017; Thomas et al., 2020).植株个体在发育过程中, 其性状变化的不同可能源于遗传差异(固定个体发生)、引起表型变化的环境差异(可塑性)或两者组合(Carlson & Holsinger, 2012).为了响应产生表型变异的进化机制, 性状在不同生态尺度上通常存在差异, 因此, 性状对不同变异驱动因素(如遗传变异、环境差异或个体发育)的敏感性差异较大(Messier et al., 2017).表型可塑性作为一种由局部环境条件所触发的特性, 是各个组织和器官感知环境信号并对其做出反应的综合体现(De Kroon et al., 2005).在小的分类尺度上叶片性状之间广泛的进化联系可能被可塑性模糊或逆转(Anderegg et al., 2018), 所以探究区域尺度植物叶片性状的变异规律及可塑性有助于深入了解区域范围内植物对环境的响应模式. ...
Leaf traits and relationships differ with season as well as among species groupings in a managed Southeastern China forest landscape 1 2012
... Dong和Liu (2017)的研究结果表明红松(Pinus koraiensis)的多个功能性状受纬度不同所导致的环境差异的影响.多个叶片性状与纬度变化密切相关, 如叶片氮磷比随纬度升高而减小, 比叶面积(SLA)和叶寿命随纬度升高而增大(张林和罗天祥, 2004; De et al., 2013).叶寿命与叶片性状的显著相关性也多有论述, 如单位质量叶氮含量与叶寿命通常呈负相关关系(张林和罗天祥, 2004; Mediavilla et al., 2014; Osada et al., 2015), SLA随叶寿命的增大而减小(Kikuzawa & Lechowicz, 2011; de la Riva et al., 2016).He和Yan (2018)认为常绿树种性状的二元关系强度并不高, 但已有多个研究证实在广泛的植物类型和生物群落之间叶性状存在普遍相关性(Reich et al., 1999; Wright et al., 2004; Niinemets, 2015).红松针叶的SLA与叶片厚度呈显著的正线性相关关系, 但与叶干物质含量(LDMC)呈显著的负线性相关关系(Dong & Liu, 2017).叶片的SLA与体积也表现为显著负相关, 且SLA较小而叶片体积大的植物资源利用性更高(王晓洁等, 2015).叶片性状之间的关系往往反映了植物对不同环境下养分资源的适应(Chen et al., 2012), 因此, 揭示种内叶片性状的共变模式, 对于确定叶片是如何调整资源利用策略来适应环境变化是至关重要的, 但这种共变策略是否受纬度梯度的影响仍不清晰. ...
Freezing injury in primary and secondary needles of mediterranean pine species of contrasting ecological niches 1 2009
... 当年生叶片与所有龄级叶片间的性状存在显著差异(表2), 这与以往研究结论相符.Mediavilla和Escudero (2003)曾在叶片寿命与光合特性的线性关系中发现, 用所有龄级平均值计算比当年生叶片数据得到的斜率更陡.黄海侠等(2013)也确定了当年生叶片的性状值较为发散, 往年生叶片的性状值更为聚拢和稳定.导致这种差异的主要原因可能是叶片发育阶段的不同致使投资策略不同(Climent et al., 2009), 幼年叶片倾向于通过增大光合效率积累生物量, 而成年叶片以支撑结构的投资为主, 投资策略的调整可能是受叶片发育期间生长条件的影响(Gilmore et al., 1995)而形成的响应机制. ...
Latitudinal gradients as natural laboratories to infer species’ responses to temperature 1 2013
... Dong和Liu (2017)的研究结果表明红松(Pinus koraiensis)的多个功能性状受纬度不同所导致的环境差异的影响.多个叶片性状与纬度变化密切相关, 如叶片氮磷比随纬度升高而减小, 比叶面积(SLA)和叶寿命随纬度升高而增大(张林和罗天祥, 2004; De et al., 2013).叶寿命与叶片性状的显著相关性也多有论述, 如单位质量叶氮含量与叶寿命通常呈负相关关系(张林和罗天祥, 2004; Mediavilla et al., 2014; Osada et al., 2015), SLA随叶寿命的增大而减小(Kikuzawa & Lechowicz, 2011; de la Riva et al., 2016).He和Yan (2018)认为常绿树种性状的二元关系强度并不高, 但已有多个研究证实在广泛的植物类型和生物群落之间叶性状存在普遍相关性(Reich et al., 1999; Wright et al., 2004; Niinemets, 2015).红松针叶的SLA与叶片厚度呈显著的正线性相关关系, 但与叶干物质含量(LDMC)呈显著的负线性相关关系(Dong & Liu, 2017).叶片的SLA与体积也表现为显著负相关, 且SLA较小而叶片体积大的植物资源利用性更高(王晓洁等, 2015).叶片性状之间的关系往往反映了植物对不同环境下养分资源的适应(Chen et al., 2012), 因此, 揭示种内叶片性状的共变模式, 对于确定叶片是如何调整资源利用策略来适应环境变化是至关重要的, 但这种共变策略是否受纬度梯度的影响仍不清晰. ...
A modular concept of phenotypic plasticity in plants 2005
Leaf mass per area (LMA) and its relationship with leaf structure and anatomy in 34 Mediterranean woody species along a water availability gradient 2016
Response of Korean pine’s functional traits to geography and climate 3 2017
... Dong和Liu (2017)的研究结果表明红松(Pinus koraiensis)的多个功能性状受纬度不同所导致的环境差异的影响.多个叶片性状与纬度变化密切相关, 如叶片氮磷比随纬度升高而减小, 比叶面积(SLA)和叶寿命随纬度升高而增大(张林和罗天祥, 2004; De et al., 2013).叶寿命与叶片性状的显著相关性也多有论述, 如单位质量叶氮含量与叶寿命通常呈负相关关系(张林和罗天祥, 2004; Mediavilla et al., 2014; Osada et al., 2015), SLA随叶寿命的增大而减小(Kikuzawa & Lechowicz, 2011; de la Riva et al., 2016).He和Yan (2018)认为常绿树种性状的二元关系强度并不高, 但已有多个研究证实在广泛的植物类型和生物群落之间叶性状存在普遍相关性(Reich et al., 1999; Wright et al., 2004; Niinemets, 2015).红松针叶的SLA与叶片厚度呈显著的正线性相关关系, 但与叶干物质含量(LDMC)呈显著的负线性相关关系(Dong & Liu, 2017).叶片的SLA与体积也表现为显著负相关, 且SLA较小而叶片体积大的植物资源利用性更高(王晓洁等, 2015).叶片性状之间的关系往往反映了植物对不同环境下养分资源的适应(Chen et al., 2012), 因此, 揭示种内叶片性状的共变模式, 对于确定叶片是如何调整资源利用策略来适应环境变化是至关重要的, 但这种共变策略是否受纬度梯度的影响仍不清晰. ...
Relationships between needle traits, needle age and site and stand parameters in Pinus pinaster 2 2015
... 针叶年龄对针长、针厚以及针叶总投影面积等性状有显著影响(Eimil-Fraga et al., 2015), 本研究结果强调了针叶年龄在解释红松叶形态性状变异中的重要地位(图2).红松针叶的SLA随针叶年龄增加而减小(图3), 这与以往研究结果(Anderegg et al., 2018; Liu et al., 2020)相符.植物通过合成不同化合物从而以不同化学成分和结构来分配能量(Villar & Merino, 2001), 韧皮部的二次生长、薄壁组织细胞酚类衍生物的增加和碳汇减少(Gilmore et al., 1995; Kuusk et al., 2018b)都是导致SLA随针叶年龄增加而减小的重要原因.另外, 叶片年龄尚小时维管束的体积较大进而叶片密度增大, 也会导致SLA的变化(Grotkopp et al., 2002).LDMC和ND均随针叶年龄增加而增大(图3), 与Eimil-Fraga等(2015)和Kuusk等(2018a)对海岸松和意大利松(Pinus pinea)等树种的研究结果相符.LDMC可以指示结构细胞含量与液体细胞含量(Kikuzawa & Lechowicz, 2011), 对细胞壁投入更多以及老叶的代谢物沉积是LDMC随针叶年龄增加而增大的主要原因(黄海侠等, 2013; Kuusk et al., 2018b).而叶片在年龄增加的过程中为了增加叶片韧性, 空气占据的空间逐渐减小而木质化组织大量增加(Evans & Poorter, 2001; Kuusk et al., 2018a; Poorter et al., 2009), 这可能是导致ND随针叶年龄增加而增大的重要原因.Kuusk等(2018b)的研究结果表明幼年针叶的NV更小, 本研究结果与之相符, 但增加到一定程度后NV又有所下降, 这可能是因为老叶不仅是常绿乔木的同化器官还是同化产物的贮藏器官(Suzuki & Takahashi, 2020), 叶片年龄越大可利用资源存储越多, 而叶片体积作为反映植物本身对资源利用能力的综合指标(王晓洁等, 2015), 表现出随年龄增加而减小的趋势, 说明了红松老叶对资源有效利用的能力下降. ...
... ), 与Eimil-Fraga等(2015)和Kuusk等(2018a)对海岸松和意大利松(Pinus pinea)等树种的研究结果相符.LDMC可以指示结构细胞含量与液体细胞含量(Kikuzawa & Lechowicz, 2011), 对细胞壁投入更多以及老叶的代谢物沉积是LDMC随针叶年龄增加而增大的主要原因(黄海侠等, 2013; Kuusk et al., 2018b).而叶片在年龄增加的过程中为了增加叶片韧性, 空气占据的空间逐渐减小而木质化组织大量增加(Evans & Poorter, 2001; Kuusk et al., 2018a; Poorter et al., 2009), 这可能是导致ND随针叶年龄增加而增大的重要原因.Kuusk等(2018b)的研究结果表明幼年针叶的NV更小, 本研究结果与之相符, 但增加到一定程度后NV又有所下降, 这可能是因为老叶不仅是常绿乔木的同化器官还是同化产物的贮藏器官(Suzuki & Takahashi, 2020), 叶片年龄越大可利用资源存储越多, 而叶片体积作为反映植物本身对资源利用能力的综合指标(王晓洁等, 2015), 表现出随年龄增加而减小的趋势, 说明了红松老叶对资源有效利用的能力下降. ...
Photosynthetic acclimation of plants to growth irradiance: the relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain 1 2001
... 针叶年龄对针长、针厚以及针叶总投影面积等性状有显著影响(Eimil-Fraga et al., 2015), 本研究结果强调了针叶年龄在解释红松叶形态性状变异中的重要地位(图2).红松针叶的SLA随针叶年龄增加而减小(图3), 这与以往研究结果(Anderegg et al., 2018; Liu et al., 2020)相符.植物通过合成不同化合物从而以不同化学成分和结构来分配能量(Villar & Merino, 2001), 韧皮部的二次生长、薄壁组织细胞酚类衍生物的增加和碳汇减少(Gilmore et al., 1995; Kuusk et al., 2018b)都是导致SLA随针叶年龄增加而减小的重要原因.另外, 叶片年龄尚小时维管束的体积较大进而叶片密度增大, 也会导致SLA的变化(Grotkopp et al., 2002).LDMC和ND均随针叶年龄增加而增大(图3), 与Eimil-Fraga等(2015)和Kuusk等(2018a)对海岸松和意大利松(Pinus pinea)等树种的研究结果相符.LDMC可以指示结构细胞含量与液体细胞含量(Kikuzawa & Lechowicz, 2011), 对细胞壁投入更多以及老叶的代谢物沉积是LDMC随针叶年龄增加而增大的主要原因(黄海侠等, 2013; Kuusk et al., 2018b).而叶片在年龄增加的过程中为了增加叶片韧性, 空气占据的空间逐渐减小而木质化组织大量增加(Evans & Poorter, 2001; Kuusk et al., 2018a; Poorter et al., 2009), 这可能是导致ND随针叶年龄增加而增大的重要原因.Kuusk等(2018b)的研究结果表明幼年针叶的NV更小, 本研究结果与之相符, 但增加到一定程度后NV又有所下降, 这可能是因为老叶不仅是常绿乔木的同化器官还是同化产物的贮藏器官(Suzuki & Takahashi, 2020), 叶片年龄越大可利用资源存储越多, 而叶片体积作为反映植物本身对资源利用能力的综合指标(王晓洁等, 2015), 表现出随年龄增加而减小的趋势, 说明了红松老叶对资源有效利用的能力下降. ...
Intraspecific trait variation and covariation in a widespread tree species (Nothofagus pumilio) in southern Chile 2 2011
... 近年来, 探究区域尺度上植物功能性状的种内变异模式备受关注(Fajardo & Piper, 2011; Martin et al., 2017; Thomas et al., 2020).植株个体在发育过程中, 其性状变化的不同可能源于遗传差异(固定个体发生)、引起表型变化的环境差异(可塑性)或两者组合(Carlson & Holsinger, 2012).为了响应产生表型变异的进化机制, 性状在不同生态尺度上通常存在差异, 因此, 性状对不同变异驱动因素(如遗传变异、环境差异或个体发育)的敏感性差异较大(Messier et al., 2017).表型可塑性作为一种由局部环境条件所触发的特性, 是各个组织和器官感知环境信号并对其做出反应的综合体现(De Kroon et al., 2005).在小的分类尺度上叶片性状之间广泛的进化联系可能被可塑性模糊或逆转(Anderegg et al., 2018), 所以探究区域尺度植物叶片性状的变异规律及可塑性有助于深入了解区域范围内植物对环境的响应模式. ...
Age-related variation in carbon allocation at tree and stand scales in beech (Fagus sylvatica L.) and sessile oak (Quercus petraea (Matt.) Liebl.) using a chronosequence approach 1 2010
Canopy dynamics and the morphological development of Abies balsamea: effects of foliage age on specific leaf area and secondary vascular development 2 1995
... 针叶年龄对针长、针厚以及针叶总投影面积等性状有显著影响(Eimil-Fraga et al., 2015), 本研究结果强调了针叶年龄在解释红松叶形态性状变异中的重要地位(图2).红松针叶的SLA随针叶年龄增加而减小(图3), 这与以往研究结果(Anderegg et al., 2018; Liu et al., 2020)相符.植物通过合成不同化合物从而以不同化学成分和结构来分配能量(Villar & Merino, 2001), 韧皮部的二次生长、薄壁组织细胞酚类衍生物的增加和碳汇减少(Gilmore et al., 1995; Kuusk et al., 2018b)都是导致SLA随针叶年龄增加而减小的重要原因.另外, 叶片年龄尚小时维管束的体积较大进而叶片密度增大, 也会导致SLA的变化(Grotkopp et al., 2002).LDMC和ND均随针叶年龄增加而增大(图3), 与Eimil-Fraga等(2015)和Kuusk等(2018a)对海岸松和意大利松(Pinus pinea)等树种的研究结果相符.LDMC可以指示结构细胞含量与液体细胞含量(Kikuzawa & Lechowicz, 2011), 对细胞壁投入更多以及老叶的代谢物沉积是LDMC随针叶年龄增加而增大的主要原因(黄海侠等, 2013; Kuusk et al., 2018b).而叶片在年龄增加的过程中为了增加叶片韧性, 空气占据的空间逐渐减小而木质化组织大量增加(Evans & Poorter, 2001; Kuusk et al., 2018a; Poorter et al., 2009), 这可能是导致ND随针叶年龄增加而增大的重要原因.Kuusk等(2018b)的研究结果表明幼年针叶的NV更小, 本研究结果与之相符, 但增加到一定程度后NV又有所下降, 这可能是因为老叶不仅是常绿乔木的同化器官还是同化产物的贮藏器官(Suzuki & Takahashi, 2020), 叶片年龄越大可利用资源存储越多, 而叶片体积作为反映植物本身对资源利用能力的综合指标(王晓洁等, 2015), 表现出随年龄增加而减小的趋势, 说明了红松老叶对资源有效利用的能力下降. ...
... 当年生叶片与所有龄级叶片间的性状存在显著差异(表2), 这与以往研究结论相符.Mediavilla和Escudero (2003)曾在叶片寿命与光合特性的线性关系中发现, 用所有龄级平均值计算比当年生叶片数据得到的斜率更陡.黄海侠等(2013)也确定了当年生叶片的性状值较为发散, 往年生叶片的性状值更为聚拢和稳定.导致这种差异的主要原因可能是叶片发育阶段的不同致使投资策略不同(Climent et al., 2009), 幼年叶片倾向于通过增大光合效率积累生物量, 而成年叶片以支撑结构的投资为主, 投资策略的调整可能是受叶片发育期间生长条件的影响(Gilmore et al., 1995)而形成的响应机制. ...
Correlation between leaf age and other leaf traits in three Mediterranean maquis shrub species: Quercus ilex, Phillyrea latifolia and Cistus incanus 2 2000
... 针叶年龄是影响性状变异及其相关关系的主要因素(Reich et al., 1999; Gratani & Bombelli, 2000).许多物种会表现出一种被称为异胚性的发育变化(Zotz et al., 2011; Kuusk et al., 2018b), 即使是同一株植物但不同年龄的叶片形态也会存在明显差异(Grotkopp et al., 2002).然而, 以往关于常绿树种叶性状的研究通常以完全展开的当年生叶片为研究对象, 而忽略了叶片衰老对性状的影响(Han et al., 2008).黄海侠等(2013)通过对常绿树种当年生和多年生叶片的研究认为其SLA和LDMC因老龄叶片密度和厚度变大会发生较大变异.Kuusk等(2018a)对地中海3种松树的研究发现不同针叶年龄的针叶体积(NV)存在显著差异, 而海岸松(Pinus pinaster)的针叶密度(ND)也受到针叶年龄的显著影响(Eimil- Fraga et al., 2015).这种不同针叶年龄之间叶片形态的差异受年际间温度和降水等环境因素的影响(曾瑞琪等, 2018), 也有可能是由发育阶段不同导致叶片生活史策略发生转变造成的(黄海侠等, 2013; Robakowski & Bielinis, 2017).但目前在区域尺度上探究针叶年龄是如何影响红松叶性状及其相关性的研究较少. ...
... 王瑞丽等(2015)对中国东部南北样带的多种植物调查分析后发现SLA和LDMC存在明显的纬度格局, 而在本研究中红松4个叶形态性状在不同纬度梯度虽大多有显著差异, 但并未表现出规律性(表2), 可能是前者研究区域范围的纬度跨度更大、气候条件差异显著导致的结果.本研究中穆棱样点的叶寿命在4个样点中相对较长, 但SLA最小且与其他样点存在显著差异(表2), 这与SLA低则叶寿命长的普遍结论(Gratani & Bombelli, 2000; de la Riva et al., 2016; Onoda et al., 2017)相符合.Withington等(2006)基于全球数据也得到相似的结论, 即具有厚叶、低SLA、低单位质量氮浓度和低光合速率的物种通常表现出较长的叶寿命.叶片具有较高的SLA通常单位叶面积所需的碳投资很少, 生理上更活跃, 所以低SLA的叶片具有较长的叶寿命可以在防御结构中每单位叶面积投入更多的碳, 使它们更能抵御物理胁迫和食草动物啃食(Sakschewski et al., 2015).纬度最高的胜山样点红松的NV最小ND最大(表2), 这可能是资源的生产能力和存储量受低温环境限制而不足, 因此NV变小, 红松叶片投资策略转向防御并选择了更保守的叶片性状来缓冲低温对生长的影响(Buzzard et al., 2019), 且NV的变异速率大于叶干质量增加的速率导致ND变大. ...
Toward a causal explanation of plant invasiveness: seedling growth and life-history strategies of 29 pine (Pinus) species 2 2002
... 针叶年龄是影响性状变异及其相关关系的主要因素(Reich et al., 1999; Gratani & Bombelli, 2000).许多物种会表现出一种被称为异胚性的发育变化(Zotz et al., 2011; Kuusk et al., 2018b), 即使是同一株植物但不同年龄的叶片形态也会存在明显差异(Grotkopp et al., 2002).然而, 以往关于常绿树种叶性状的研究通常以完全展开的当年生叶片为研究对象, 而忽略了叶片衰老对性状的影响(Han et al., 2008).黄海侠等(2013)通过对常绿树种当年生和多年生叶片的研究认为其SLA和LDMC因老龄叶片密度和厚度变大会发生较大变异.Kuusk等(2018a)对地中海3种松树的研究发现不同针叶年龄的针叶体积(NV)存在显著差异, 而海岸松(Pinus pinaster)的针叶密度(ND)也受到针叶年龄的显著影响(Eimil- Fraga et al., 2015).这种不同针叶年龄之间叶片形态的差异受年际间温度和降水等环境因素的影响(曾瑞琪等, 2018), 也有可能是由发育阶段不同导致叶片生活史策略发生转变造成的(黄海侠等, 2013; Robakowski & Bielinis, 2017).但目前在区域尺度上探究针叶年龄是如何影响红松叶性状及其相关性的研究较少. ...
... 针叶年龄对针长、针厚以及针叶总投影面积等性状有显著影响(Eimil-Fraga et al., 2015), 本研究结果强调了针叶年龄在解释红松叶形态性状变异中的重要地位(图2).红松针叶的SLA随针叶年龄增加而减小(图3), 这与以往研究结果(Anderegg et al., 2018; Liu et al., 2020)相符.植物通过合成不同化合物从而以不同化学成分和结构来分配能量(Villar & Merino, 2001), 韧皮部的二次生长、薄壁组织细胞酚类衍生物的增加和碳汇减少(Gilmore et al., 1995; Kuusk et al., 2018b)都是导致SLA随针叶年龄增加而减小的重要原因.另外, 叶片年龄尚小时维管束的体积较大进而叶片密度增大, 也会导致SLA的变化(Grotkopp et al., 2002).LDMC和ND均随针叶年龄增加而增大(图3), 与Eimil-Fraga等(2015)和Kuusk等(2018a)对海岸松和意大利松(Pinus pinea)等树种的研究结果相符.LDMC可以指示结构细胞含量与液体细胞含量(Kikuzawa & Lechowicz, 2011), 对细胞壁投入更多以及老叶的代谢物沉积是LDMC随针叶年龄增加而增大的主要原因(黄海侠等, 2013; Kuusk et al., 2018b).而叶片在年龄增加的过程中为了增加叶片韧性, 空气占据的空间逐渐减小而木质化组织大量增加(Evans & Poorter, 2001; Kuusk et al., 2018a; Poorter et al., 2009), 这可能是导致ND随针叶年龄增加而增大的重要原因.Kuusk等(2018b)的研究结果表明幼年针叶的NV更小, 本研究结果与之相符, 但增加到一定程度后NV又有所下降, 这可能是因为老叶不仅是常绿乔木的同化器官还是同化产物的贮藏器官(Suzuki & Takahashi, 2020), 叶片年龄越大可利用资源存储越多, 而叶片体积作为反映植物本身对资源利用能力的综合指标(王晓洁等, 2015), 表现出随年龄增加而减小的趋势, 说明了红松老叶对资源有效利用的能力下降. ...
Leaf-age effects on seasonal variability in photosynthetic parameters and its relationships with leaf mass per area and leaf nitrogen concentration within a Pinus densiflora crown 1 2008
... 针叶年龄是影响性状变异及其相关关系的主要因素(Reich et al., 1999; Gratani & Bombelli, 2000).许多物种会表现出一种被称为异胚性的发育变化(Zotz et al., 2011; Kuusk et al., 2018b), 即使是同一株植物但不同年龄的叶片形态也会存在明显差异(Grotkopp et al., 2002).然而, 以往关于常绿树种叶性状的研究通常以完全展开的当年生叶片为研究对象, 而忽略了叶片衰老对性状的影响(Han et al., 2008).黄海侠等(2013)通过对常绿树种当年生和多年生叶片的研究认为其SLA和LDMC因老龄叶片密度和厚度变大会发生较大变异.Kuusk等(2018a)对地中海3种松树的研究发现不同针叶年龄的针叶体积(NV)存在显著差异, 而海岸松(Pinus pinaster)的针叶密度(ND)也受到针叶年龄的显著影响(Eimil- Fraga et al., 2015).这种不同针叶年龄之间叶片形态的差异受年际间温度和降水等环境因素的影响(曾瑞琪等, 2018), 也有可能是由发育阶段不同导致叶片生活史策略发生转变造成的(黄海侠等, 2013; Robakowski & Bielinis, 2017).但目前在区域尺度上探究针叶年龄是如何影响红松叶性状及其相关性的研究较少. ...
Size-dependent variations in individual traits and trait scaling relationships within a shade-tolerant evergreen tree species 1 2018
... Dong和Liu (2017)的研究结果表明红松(Pinus koraiensis)的多个功能性状受纬度不同所导致的环境差异的影响.多个叶片性状与纬度变化密切相关, 如叶片氮磷比随纬度升高而减小, 比叶面积(SLA)和叶寿命随纬度升高而增大(张林和罗天祥, 2004; De et al., 2013).叶寿命与叶片性状的显著相关性也多有论述, 如单位质量叶氮含量与叶寿命通常呈负相关关系(张林和罗天祥, 2004; Mediavilla et al., 2014; Osada et al., 2015), SLA随叶寿命的增大而减小(Kikuzawa & Lechowicz, 2011; de la Riva et al., 2016).He和Yan (2018)认为常绿树种性状的二元关系强度并不高, 但已有多个研究证实在广泛的植物类型和生物群落之间叶性状存在普遍相关性(Reich et al., 1999; Wright et al., 2004; Niinemets, 2015).红松针叶的SLA与叶片厚度呈显著的正线性相关关系, 但与叶干物质含量(LDMC)呈显著的负线性相关关系(Dong & Liu, 2017).叶片的SLA与体积也表现为显著负相关, 且SLA较小而叶片体积大的植物资源利用性更高(王晓洁等, 2015).叶片性状之间的关系往往反映了植物对不同环境下养分资源的适应(Chen et al., 2012), 因此, 揭示种内叶片性状的共变模式, 对于确定叶片是如何调整资源利用策略来适应环境变化是至关重要的, 但这种共变策略是否受纬度梯度的影响仍不清晰. ...
浙江天童常绿植物当年生与往年生叶片性状的变异与关联 5 2013
... 针叶年龄是影响性状变异及其相关关系的主要因素(Reich et al., 1999; Gratani & Bombelli, 2000).许多物种会表现出一种被称为异胚性的发育变化(Zotz et al., 2011; Kuusk et al., 2018b), 即使是同一株植物但不同年龄的叶片形态也会存在明显差异(Grotkopp et al., 2002).然而, 以往关于常绿树种叶性状的研究通常以完全展开的当年生叶片为研究对象, 而忽略了叶片衰老对性状的影响(Han et al., 2008).黄海侠等(2013)通过对常绿树种当年生和多年生叶片的研究认为其SLA和LDMC因老龄叶片密度和厚度变大会发生较大变异.Kuusk等(2018a)对地中海3种松树的研究发现不同针叶年龄的针叶体积(NV)存在显著差异, 而海岸松(Pinus pinaster)的针叶密度(ND)也受到针叶年龄的显著影响(Eimil- Fraga et al., 2015).这种不同针叶年龄之间叶片形态的差异受年际间温度和降水等环境因素的影响(曾瑞琪等, 2018), 也有可能是由发育阶段不同导致叶片生活史策略发生转变造成的(黄海侠等, 2013; Robakowski & Bielinis, 2017).但目前在区域尺度上探究针叶年龄是如何影响红松叶性状及其相关性的研究较少. ...
Needle life span, photosynthetic rate and nutrient concentration of Picea glehnii, P. jezoensis and P. abies planted on serpentine soil in northern Japan 1 2002
Intraspecific trait variation across multiple scales: the leaf economics spectrum in coffee 3 2017
... 近年来, 探究区域尺度上植物功能性状的种内变异模式备受关注(Fajardo & Piper, 2011; Martin et al., 2017; Thomas et al., 2020).植株个体在发育过程中, 其性状变化的不同可能源于遗传差异(固定个体发生)、引起表型变化的环境差异(可塑性)或两者组合(Carlson & Holsinger, 2012).为了响应产生表型变异的进化机制, 性状在不同生态尺度上通常存在差异, 因此, 性状对不同变异驱动因素(如遗传变异、环境差异或个体发育)的敏感性差异较大(Messier et al., 2017).表型可塑性作为一种由局部环境条件所触发的特性, 是各个组织和器官感知环境信号并对其做出反应的综合体现(De Kroon et al., 2005).在小的分类尺度上叶片性状之间广泛的进化联系可能被可塑性模糊或逆转(Anderegg et al., 2018), 所以探究区域尺度植物叶片性状的变异规律及可塑性有助于深入了解区域范围内植物对环境的响应模式. ...
... 叶经济谱性状很大程度上保留了各自独立响应表型变异驱动因素的能力, 驱动因素的影响因尺度而异(Martin et al., 2017).作为叶经济谱的一个重要性状, 很多研究已证明SLA对种内变异和环境普遍敏感(Adler et al., 2014; Messier et al., 2017).SLA可塑性相对较大, LDMC的可塑性低于SLA (表2), 这与de la Riva等(2016)的研究结论相符.当年生叶片与所有龄级表现的可塑性大小不同, 这可能是不同年龄针叶之间的光适应能力差异导致的, 尤其是当年生针叶容易受日照强度影响做出适应性调整(Wyka et al., 2007).不同针叶年龄下响应策略不同也可能是叶片可塑性大小存在较大差异的原因, De Kroon等(2005)曾提出, 在生长发育阶段植物做出的可塑性响应分别包括决定分生组织的营养或生殖去向以及对植物各部分尺寸和形状的调整, 而在成熟阶段表达为诱导防御等. ...
... 样点对叶性状变异及相关关系有一定的影响, 尤其对由叶片构建期间的环境波动所决定的形态性状影响很大(Reich et al., 1999; Martin et al., 2017), 而王瑞丽等(2015)的研究显示来源于样地间环境差异的叶功能属性的变异不到1/3.Anderegg等(2018)对美国西北地区进行调查后发现, 地块级别的环境特征并不是种间、种内或性状间变异的强预测因子.本研究结果证明样点(纬度)是叶片种内性状变异的原因之一, 但对除NV以外其他3个性状变异的解释占比很小(图2). ...
Photosynthetic capacity, integrated over the lifetime of a leaf, is predicted to be independent of leaf longevity in some tree species 1 2003
... 当年生叶片与所有龄级叶片间的性状存在显著差异(表2), 这与以往研究结论相符.Mediavilla和Escudero (2003)曾在叶片寿命与光合特性的线性关系中发现, 用所有龄级平均值计算比当年生叶片数据得到的斜率更陡.黄海侠等(2013)也确定了当年生叶片的性状值较为发散, 往年生叶片的性状值更为聚拢和稳定.导致这种差异的主要原因可能是叶片发育阶段的不同致使投资策略不同(Climent et al., 2009), 幼年叶片倾向于通过增大光合效率积累生物量, 而成年叶片以支撑结构的投资为主, 投资策略的调整可能是受叶片发育期间生长条件的影响(Gilmore et al., 1995)而形成的响应机制. ...
Ontogenetic transition in leaf traits: a new cost associated with the increase in leaf longevity 1 2014
... Dong和Liu (2017)的研究结果表明红松(Pinus koraiensis)的多个功能性状受纬度不同所导致的环境差异的影响.多个叶片性状与纬度变化密切相关, 如叶片氮磷比随纬度升高而减小, 比叶面积(SLA)和叶寿命随纬度升高而增大(张林和罗天祥, 2004; De et al., 2013).叶寿命与叶片性状的显著相关性也多有论述, 如单位质量叶氮含量与叶寿命通常呈负相关关系(张林和罗天祥, 2004; Mediavilla et al., 2014; Osada et al., 2015), SLA随叶寿命的增大而减小(Kikuzawa & Lechowicz, 2011; de la Riva et al., 2016).He和Yan (2018)认为常绿树种性状的二元关系强度并不高, 但已有多个研究证实在广泛的植物类型和生物群落之间叶性状存在普遍相关性(Reich et al., 1999; Wright et al., 2004; Niinemets, 2015).红松针叶的SLA与叶片厚度呈显著的正线性相关关系, 但与叶干物质含量(LDMC)呈显著的负线性相关关系(Dong & Liu, 2017).叶片的SLA与体积也表现为显著负相关, 且SLA较小而叶片体积大的植物资源利用性更高(王晓洁等, 2015).叶片性状之间的关系往往反映了植物对不同环境下养分资源的适应(Chen et al., 2012), 因此, 揭示种内叶片性状的共变模式, 对于确定叶片是如何调整资源利用策略来适应环境变化是至关重要的, 但这种共变策略是否受纬度梯度的影响仍不清晰. ...
Trait variation and integration across scales: Is the leaf economic spectrum present at local scales 3 2017
... 近年来, 探究区域尺度上植物功能性状的种内变异模式备受关注(Fajardo & Piper, 2011; Martin et al., 2017; Thomas et al., 2020).植株个体在发育过程中, 其性状变化的不同可能源于遗传差异(固定个体发生)、引起表型变化的环境差异(可塑性)或两者组合(Carlson & Holsinger, 2012).为了响应产生表型变异的进化机制, 性状在不同生态尺度上通常存在差异, 因此, 性状对不同变异驱动因素(如遗传变异、环境差异或个体发育)的敏感性差异较大(Messier et al., 2017).表型可塑性作为一种由局部环境条件所触发的特性, 是各个组织和器官感知环境信号并对其做出反应的综合体现(De Kroon et al., 2005).在小的分类尺度上叶片性状之间广泛的进化联系可能被可塑性模糊或逆转(Anderegg et al., 2018), 所以探究区域尺度植物叶片性状的变异规律及可塑性有助于深入了解区域范围内植物对环境的响应模式. ...
... 叶经济谱性状很大程度上保留了各自独立响应表型变异驱动因素的能力, 驱动因素的影响因尺度而异(Martin et al., 2017).作为叶经济谱的一个重要性状, 很多研究已证明SLA对种内变异和环境普遍敏感(Adler et al., 2014; Messier et al., 2017).SLA可塑性相对较大, LDMC的可塑性低于SLA (表2), 这与de la Riva等(2016)的研究结论相符.当年生叶片与所有龄级表现的可塑性大小不同, 这可能是不同年龄针叶之间的光适应能力差异导致的, 尤其是当年生针叶容易受日照强度影响做出适应性调整(Wyka et al., 2007).不同针叶年龄下响应策略不同也可能是叶片可塑性大小存在较大差异的原因, De Kroon等(2005)曾提出, 在生长发育阶段植物做出的可塑性响应分别包括决定分生组织的营养或生殖去向以及对植物各部分尺寸和形状的调整, 而在成熟阶段表达为诱导防御等. ...
Is there a species spectrum within the world-wide leaf economics spectrum? Major variations in leaf functional traits in the Mediterranean sclerophyll Quercus ilex 1 2015
... Dong和Liu (2017)的研究结果表明红松(Pinus koraiensis)的多个功能性状受纬度不同所导致的环境差异的影响.多个叶片性状与纬度变化密切相关, 如叶片氮磷比随纬度升高而减小, 比叶面积(SLA)和叶寿命随纬度升高而增大(张林和罗天祥, 2004; De et al., 2013).叶寿命与叶片性状的显著相关性也多有论述, 如单位质量叶氮含量与叶寿命通常呈负相关关系(张林和罗天祥, 2004; Mediavilla et al., 2014; Osada et al., 2015), SLA随叶寿命的增大而减小(Kikuzawa & Lechowicz, 2011; de la Riva et al., 2016).He和Yan (2018)认为常绿树种性状的二元关系强度并不高, 但已有多个研究证实在广泛的植物类型和生物群落之间叶性状存在普遍相关性(Reich et al., 1999; Wright et al., 2004; Niinemets, 2015).红松针叶的SLA与叶片厚度呈显著的正线性相关关系, 但与叶干物质含量(LDMC)呈显著的负线性相关关系(Dong & Liu, 2017).叶片的SLA与体积也表现为显著负相关, 且SLA较小而叶片体积大的植物资源利用性更高(王晓洁等, 2015).叶片性状之间的关系往往反映了植物对不同环境下养分资源的适应(Chen et al., 2012), 因此, 揭示种内叶片性状的共变模式, 对于确定叶片是如何调整资源利用策略来适应环境变化是至关重要的, 但这种共变策略是否受纬度梯度的影响仍不清晰. ...
Leaf internal diffusion conductance limits photosynthesis more strongly in older leaves of Mediterranean evergreen broad-leaved species 1 2005
Generality of leaf trait relationships: a test across six biomes 3 1999
... Dong和Liu (2017)的研究结果表明红松(Pinus koraiensis)的多个功能性状受纬度不同所导致的环境差异的影响.多个叶片性状与纬度变化密切相关, 如叶片氮磷比随纬度升高而减小, 比叶面积(SLA)和叶寿命随纬度升高而增大(张林和罗天祥, 2004; De et al., 2013).叶寿命与叶片性状的显著相关性也多有论述, 如单位质量叶氮含量与叶寿命通常呈负相关关系(张林和罗天祥, 2004; Mediavilla et al., 2014; Osada et al., 2015), SLA随叶寿命的增大而减小(Kikuzawa & Lechowicz, 2011; de la Riva et al., 2016).He和Yan (2018)认为常绿树种性状的二元关系强度并不高, 但已有多个研究证实在广泛的植物类型和生物群落之间叶性状存在普遍相关性(Reich et al., 1999; Wright et al., 2004; Niinemets, 2015).红松针叶的SLA与叶片厚度呈显著的正线性相关关系, 但与叶干物质含量(LDMC)呈显著的负线性相关关系(Dong & Liu, 2017).叶片的SLA与体积也表现为显著负相关, 且SLA较小而叶片体积大的植物资源利用性更高(王晓洁等, 2015).叶片性状之间的关系往往反映了植物对不同环境下养分资源的适应(Chen et al., 2012), 因此, 揭示种内叶片性状的共变模式, 对于确定叶片是如何调整资源利用策略来适应环境变化是至关重要的, 但这种共变策略是否受纬度梯度的影响仍不清晰. ...
... 针叶年龄是影响性状变异及其相关关系的主要因素(Reich et al., 1999; Gratani & Bombelli, 2000).许多物种会表现出一种被称为异胚性的发育变化(Zotz et al., 2011; Kuusk et al., 2018b), 即使是同一株植物但不同年龄的叶片形态也会存在明显差异(Grotkopp et al., 2002).然而, 以往关于常绿树种叶性状的研究通常以完全展开的当年生叶片为研究对象, 而忽略了叶片衰老对性状的影响(Han et al., 2008).黄海侠等(2013)通过对常绿树种当年生和多年生叶片的研究认为其SLA和LDMC因老龄叶片密度和厚度变大会发生较大变异.Kuusk等(2018a)对地中海3种松树的研究发现不同针叶年龄的针叶体积(NV)存在显著差异, 而海岸松(Pinus pinaster)的针叶密度(ND)也受到针叶年龄的显著影响(Eimil- Fraga et al., 2015).这种不同针叶年龄之间叶片形态的差异受年际间温度和降水等环境因素的影响(曾瑞琪等, 2018), 也有可能是由发育阶段不同导致叶片生活史策略发生转变造成的(黄海侠等, 2013; Robakowski & Bielinis, 2017).但目前在区域尺度上探究针叶年龄是如何影响红松叶性状及其相关性的研究较少. ...
... 样点对叶性状变异及相关关系有一定的影响, 尤其对由叶片构建期间的环境波动所决定的形态性状影响很大(Reich et al., 1999; Martin et al., 2017), 而王瑞丽等(2015)的研究显示来源于样地间环境差异的叶功能属性的变异不到1/3.Anderegg等(2018)对美国西北地区进行调查后发现, 地块级别的环境特征并不是种间、种内或性状间变异的强预测因子.本研究结果证明样点(纬度)是叶片种内性状变异的原因之一, 但对除NV以外其他3个性状变异的解释占比很小(图2). ...
Evidence that longer needle retention of spruce and pine populations at high elevations and high latitudes is largely a phenotypic response 1 1996
Needle age dependence of photosynthesis along a light gradient within an Abies alba crown 1 2017
... 针叶年龄是影响性状变异及其相关关系的主要因素(Reich et al., 1999; Gratani & Bombelli, 2000).许多物种会表现出一种被称为异胚性的发育变化(Zotz et al., 2011; Kuusk et al., 2018b), 即使是同一株植物但不同年龄的叶片形态也会存在明显差异(Grotkopp et al., 2002).然而, 以往关于常绿树种叶性状的研究通常以完全展开的当年生叶片为研究对象, 而忽略了叶片衰老对性状的影响(Han et al., 2008).黄海侠等(2013)通过对常绿树种当年生和多年生叶片的研究认为其SLA和LDMC因老龄叶片密度和厚度变大会发生较大变异.Kuusk等(2018a)对地中海3种松树的研究发现不同针叶年龄的针叶体积(NV)存在显著差异, 而海岸松(Pinus pinaster)的针叶密度(ND)也受到针叶年龄的显著影响(Eimil- Fraga et al., 2015).这种不同针叶年龄之间叶片形态的差异受年际间温度和降水等环境因素的影响(曾瑞琪等, 2018), 也有可能是由发育阶段不同导致叶片生活史策略发生转变造成的(黄海侠等, 2013; Robakowski & Bielinis, 2017).但目前在区域尺度上探究针叶年龄是如何影响红松叶性状及其相关性的研究较少. ...
Leaf and stem economics spectra drive diversity of functional plant traits in a dynamic global vegetation model 1 2015
... 王瑞丽等(2015)对中国东部南北样带的多种植物调查分析后发现SLA和LDMC存在明显的纬度格局, 而在本研究中红松4个叶形态性状在不同纬度梯度虽大多有显著差异, 但并未表现出规律性(表2), 可能是前者研究区域范围的纬度跨度更大、气候条件差异显著导致的结果.本研究中穆棱样点的叶寿命在4个样点中相对较长, 但SLA最小且与其他样点存在显著差异(表2), 这与SLA低则叶寿命长的普遍结论(Gratani & Bombelli, 2000; de la Riva et al., 2016; Onoda et al., 2017)相符合.Withington等(2006)基于全球数据也得到相似的结论, 即具有厚叶、低SLA、低单位质量氮浓度和低光合速率的物种通常表现出较长的叶寿命.叶片具有较高的SLA通常单位叶面积所需的碳投资很少, 生理上更活跃, 所以低SLA的叶片具有较长的叶寿命可以在防御结构中每单位叶面积投入更多的碳, 使它们更能抵御物理胁迫和食草动物啃食(Sakschewski et al., 2015).纬度最高的胜山样点红松的NV最小ND最大(表2), 这可能是资源的生产能力和存储量受低温环境限制而不足, 因此NV变小, 红松叶片投资策略转向防御并选择了更保守的叶片性状来缓冲低温对生长的影响(Buzzard et al., 2019), 且NV的变异速率大于叶干质量增加的速率导致ND变大. ...
Effects of leaf age, elevation and light conditions on photosynthesis and leaf traits in saplings of two evergreen conifers, Abies veitchii and A. mariesii 3 2020
... 针叶年龄对针长、针厚以及针叶总投影面积等性状有显著影响(Eimil-Fraga et al., 2015), 本研究结果强调了针叶年龄在解释红松叶形态性状变异中的重要地位(图2).红松针叶的SLA随针叶年龄增加而减小(图3), 这与以往研究结果(Anderegg et al., 2018; Liu et al., 2020)相符.植物通过合成不同化合物从而以不同化学成分和结构来分配能量(Villar & Merino, 2001), 韧皮部的二次生长、薄壁组织细胞酚类衍生物的增加和碳汇减少(Gilmore et al., 1995; Kuusk et al., 2018b)都是导致SLA随针叶年龄增加而减小的重要原因.另外, 叶片年龄尚小时维管束的体积较大进而叶片密度增大, 也会导致SLA的变化(Grotkopp et al., 2002).LDMC和ND均随针叶年龄增加而增大(图3), 与Eimil-Fraga等(2015)和Kuusk等(2018a)对海岸松和意大利松(Pinus pinea)等树种的研究结果相符.LDMC可以指示结构细胞含量与液体细胞含量(Kikuzawa & Lechowicz, 2011), 对细胞壁投入更多以及老叶的代谢物沉积是LDMC随针叶年龄增加而增大的主要原因(黄海侠等, 2013; Kuusk et al., 2018b).而叶片在年龄增加的过程中为了增加叶片韧性, 空气占据的空间逐渐减小而木质化组织大量增加(Evans & Poorter, 2001; Kuusk et al., 2018a; Poorter et al., 2009), 这可能是导致ND随针叶年龄增加而增大的重要原因.Kuusk等(2018b)的研究结果表明幼年针叶的NV更小, 本研究结果与之相符, 但增加到一定程度后NV又有所下降, 这可能是因为老叶不仅是常绿乔木的同化器官还是同化产物的贮藏器官(Suzuki & Takahashi, 2020), 叶片年龄越大可利用资源存储越多, 而叶片体积作为反映植物本身对资源利用能力的综合指标(王晓洁等, 2015), 表现出随年龄增加而减小的趋势, 说明了红松老叶对资源有效利用的能力下降. ...
Global plant trait relationships extend to the climatic extremes of the tundra biome 1 2020
... 近年来, 探究区域尺度上植物功能性状的种内变异模式备受关注(Fajardo & Piper, 2011; Martin et al., 2017; Thomas et al., 2020).植株个体在发育过程中, 其性状变化的不同可能源于遗传差异(固定个体发生)、引起表型变化的环境差异(可塑性)或两者组合(Carlson & Holsinger, 2012).为了响应产生表型变异的进化机制, 性状在不同生态尺度上通常存在差异, 因此, 性状对不同变异驱动因素(如遗传变异、环境差异或个体发育)的敏感性差异较大(Messier et al., 2017).表型可塑性作为一种由局部环境条件所触发的特性, 是各个组织和器官感知环境信号并对其做出反应的综合体现(De Kroon et al., 2005).在小的分类尺度上叶片性状之间广泛的进化联系可能被可塑性模糊或逆转(Anderegg et al., 2018), 所以探究区域尺度植物叶片性状的变异规律及可塑性有助于深入了解区域范围内植物对环境的响应模式. ...
Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems 1 2001
... 针叶年龄对针长、针厚以及针叶总投影面积等性状有显著影响(Eimil-Fraga et al., 2015), 本研究结果强调了针叶年龄在解释红松叶形态性状变异中的重要地位(图2).红松针叶的SLA随针叶年龄增加而减小(图3), 这与以往研究结果(Anderegg et al., 2018; Liu et al., 2020)相符.植物通过合成不同化合物从而以不同化学成分和结构来分配能量(Villar & Merino, 2001), 韧皮部的二次生长、薄壁组织细胞酚类衍生物的增加和碳汇减少(Gilmore et al., 1995; Kuusk et al., 2018b)都是导致SLA随针叶年龄增加而减小的重要原因.另外, 叶片年龄尚小时维管束的体积较大进而叶片密度增大, 也会导致SLA的变化(Grotkopp et al., 2002).LDMC和ND均随针叶年龄增加而增大(图3), 与Eimil-Fraga等(2015)和Kuusk等(2018a)对海岸松和意大利松(Pinus pinea)等树种的研究结果相符.LDMC可以指示结构细胞含量与液体细胞含量(Kikuzawa & Lechowicz, 2011), 对细胞壁投入更多以及老叶的代谢物沉积是LDMC随针叶年龄增加而增大的主要原因(黄海侠等, 2013; Kuusk et al., 2018b).而叶片在年龄增加的过程中为了增加叶片韧性, 空气占据的空间逐渐减小而木质化组织大量增加(Evans & Poorter, 2001; Kuusk et al., 2018a; Poorter et al., 2009), 这可能是导致ND随针叶年龄增加而增大的重要原因.Kuusk等(2018b)的研究结果表明幼年针叶的NV更小, 本研究结果与之相符, 但增加到一定程度后NV又有所下降, 这可能是因为老叶不仅是常绿乔木的同化器官还是同化产物的贮藏器官(Suzuki & Takahashi, 2020), 叶片年龄越大可利用资源存储越多, 而叶片体积作为反映植物本身对资源利用能力的综合指标(王晓洁等, 2015), 表现出随年龄增加而减小的趋势, 说明了红松老叶对资源有效利用的能力下降. ...
... 王瑞丽等(2015)对中国东部南北样带的多种植物调查分析后发现SLA和LDMC存在明显的纬度格局, 而在本研究中红松4个叶形态性状在不同纬度梯度虽大多有显著差异, 但并未表现出规律性(表2), 可能是前者研究区域范围的纬度跨度更大、气候条件差异显著导致的结果.本研究中穆棱样点的叶寿命在4个样点中相对较长, 但SLA最小且与其他样点存在显著差异(表2), 这与SLA低则叶寿命长的普遍结论(Gratani & Bombelli, 2000; de la Riva et al., 2016; Onoda et al., 2017)相符合.Withington等(2006)基于全球数据也得到相似的结论, 即具有厚叶、低SLA、低单位质量氮浓度和低光合速率的物种通常表现出较长的叶寿命.叶片具有较高的SLA通常单位叶面积所需的碳投资很少, 生理上更活跃, 所以低SLA的叶片具有较长的叶寿命可以在防御结构中每单位叶面积投入更多的碳, 使它们更能抵御物理胁迫和食草动物啃食(Sakschewski et al., 2015).纬度最高的胜山样点红松的NV最小ND最大(表2), 这可能是资源的生产能力和存储量受低温环境限制而不足, 因此NV变小, 红松叶片投资策略转向防御并选择了更保守的叶片性状来缓冲低温对生长的影响(Buzzard et al., 2019), 且NV的变异速率大于叶干质量增加的速率导致ND变大. ...
... 样点对叶性状变异及相关关系有一定的影响, 尤其对由叶片构建期间的环境波动所决定的形态性状影响很大(Reich et al., 1999; Martin et al., 2017), 而王瑞丽等(2015)的研究显示来源于样地间环境差异的叶功能属性的变异不到1/3.Anderegg等(2018)对美国西北地区进行调查后发现, 地块级别的环境特征并不是种间、种内或性状间变异的强预测因子.本研究结果证明样点(纬度)是叶片种内性状变异的原因之一, 但对除NV以外其他3个性状变异的解释占比很小(图2). ...
中国森林叶片功能属性的纬度格局及其影响因素 2 2015
... 王瑞丽等(2015)对中国东部南北样带的多种植物调查分析后发现SLA和LDMC存在明显的纬度格局, 而在本研究中红松4个叶形态性状在不同纬度梯度虽大多有显著差异, 但并未表现出规律性(表2), 可能是前者研究区域范围的纬度跨度更大、气候条件差异显著导致的结果.本研究中穆棱样点的叶寿命在4个样点中相对较长, 但SLA最小且与其他样点存在显著差异(表2), 这与SLA低则叶寿命长的普遍结论(Gratani & Bombelli, 2000; de la Riva et al., 2016; Onoda et al., 2017)相符合.Withington等(2006)基于全球数据也得到相似的结论, 即具有厚叶、低SLA、低单位质量氮浓度和低光合速率的物种通常表现出较长的叶寿命.叶片具有较高的SLA通常单位叶面积所需的碳投资很少, 生理上更活跃, 所以低SLA的叶片具有较长的叶寿命可以在防御结构中每单位叶面积投入更多的碳, 使它们更能抵御物理胁迫和食草动物啃食(Sakschewski et al., 2015).纬度最高的胜山样点红松的NV最小ND最大(表2), 这可能是资源的生产能力和存储量受低温环境限制而不足, 因此NV变小, 红松叶片投资策略转向防御并选择了更保守的叶片性状来缓冲低温对生长的影响(Buzzard et al., 2019), 且NV的变异速率大于叶干质量增加的速率导致ND变大. ...
... 样点对叶性状变异及相关关系有一定的影响, 尤其对由叶片构建期间的环境波动所决定的形态性状影响很大(Reich et al., 1999; Martin et al., 2017), 而王瑞丽等(2015)的研究显示来源于样地间环境差异的叶功能属性的变异不到1/3.Anderegg等(2018)对美国西北地区进行调查后发现, 地块级别的环境特征并不是种间、种内或性状间变异的强预测因子.本研究结果证明样点(纬度)是叶片种内性状变异的原因之一, 但对除NV以外其他3个性状变异的解释占比很小(图2). ...
凉水天然红松阔叶混交林主要植物叶片性状相互关系研究 3 2015
... Dong和Liu (2017)的研究结果表明红松(Pinus koraiensis)的多个功能性状受纬度不同所导致的环境差异的影响.多个叶片性状与纬度变化密切相关, 如叶片氮磷比随纬度升高而减小, 比叶面积(SLA)和叶寿命随纬度升高而增大(张林和罗天祥, 2004; De et al., 2013).叶寿命与叶片性状的显著相关性也多有论述, 如单位质量叶氮含量与叶寿命通常呈负相关关系(张林和罗天祥, 2004; Mediavilla et al., 2014; Osada et al., 2015), SLA随叶寿命的增大而减小(Kikuzawa & Lechowicz, 2011; de la Riva et al., 2016).He和Yan (2018)认为常绿树种性状的二元关系强度并不高, 但已有多个研究证实在广泛的植物类型和生物群落之间叶性状存在普遍相关性(Reich et al., 1999; Wright et al., 2004; Niinemets, 2015).红松针叶的SLA与叶片厚度呈显著的正线性相关关系, 但与叶干物质含量(LDMC)呈显著的负线性相关关系(Dong & Liu, 2017).叶片的SLA与体积也表现为显著负相关, 且SLA较小而叶片体积大的植物资源利用性更高(王晓洁等, 2015).叶片性状之间的关系往往反映了植物对不同环境下养分资源的适应(Chen et al., 2012), 因此, 揭示种内叶片性状的共变模式, 对于确定叶片是如何调整资源利用策略来适应环境变化是至关重要的, 但这种共变策略是否受纬度梯度的影响仍不清晰. ...
... 针叶年龄对针长、针厚以及针叶总投影面积等性状有显著影响(Eimil-Fraga et al., 2015), 本研究结果强调了针叶年龄在解释红松叶形态性状变异中的重要地位(图2).红松针叶的SLA随针叶年龄增加而减小(图3), 这与以往研究结果(Anderegg et al., 2018; Liu et al., 2020)相符.植物通过合成不同化合物从而以不同化学成分和结构来分配能量(Villar & Merino, 2001), 韧皮部的二次生长、薄壁组织细胞酚类衍生物的增加和碳汇减少(Gilmore et al., 1995; Kuusk et al., 2018b)都是导致SLA随针叶年龄增加而减小的重要原因.另外, 叶片年龄尚小时维管束的体积较大进而叶片密度增大, 也会导致SLA的变化(Grotkopp et al., 2002).LDMC和ND均随针叶年龄增加而增大(图3), 与Eimil-Fraga等(2015)和Kuusk等(2018a)对海岸松和意大利松(Pinus pinea)等树种的研究结果相符.LDMC可以指示结构细胞含量与液体细胞含量(Kikuzawa & Lechowicz, 2011), 对细胞壁投入更多以及老叶的代谢物沉积是LDMC随针叶年龄增加而增大的主要原因(黄海侠等, 2013; Kuusk et al., 2018b).而叶片在年龄增加的过程中为了增加叶片韧性, 空气占据的空间逐渐减小而木质化组织大量增加(Evans & Poorter, 2001; Kuusk et al., 2018a; Poorter et al., 2009), 这可能是导致ND随针叶年龄增加而增大的重要原因.Kuusk等(2018b)的研究结果表明幼年针叶的NV更小, 本研究结果与之相符, 但增加到一定程度后NV又有所下降, 这可能是因为老叶不仅是常绿乔木的同化器官还是同化产物的贮藏器官(Suzuki & Takahashi, 2020), 叶片年龄越大可利用资源存储越多, 而叶片体积作为反映植物本身对资源利用能力的综合指标(王晓洁等, 2015), 表现出随年龄增加而减小的趋势, 说明了红松老叶对资源有效利用的能力下降. ...
Comparisons of structure and life span in roots and leaves among temperate trees 1 2006
... 王瑞丽等(2015)对中国东部南北样带的多种植物调查分析后发现SLA和LDMC存在明显的纬度格局, 而在本研究中红松4个叶形态性状在不同纬度梯度虽大多有显著差异, 但并未表现出规律性(表2), 可能是前者研究区域范围的纬度跨度更大、气候条件差异显著导致的结果.本研究中穆棱样点的叶寿命在4个样点中相对较长, 但SLA最小且与其他样点存在显著差异(表2), 这与SLA低则叶寿命长的普遍结论(Gratani & Bombelli, 2000; de la Riva et al., 2016; Onoda et al., 2017)相符合.Withington等(2006)基于全球数据也得到相似的结论, 即具有厚叶、低SLA、低单位质量氮浓度和低光合速率的物种通常表现出较长的叶寿命.叶片具有较高的SLA通常单位叶面积所需的碳投资很少, 生理上更活跃, 所以低SLA的叶片具有较长的叶寿命可以在防御结构中每单位叶面积投入更多的碳, 使它们更能抵御物理胁迫和食草动物啃食(Sakschewski et al., 2015).纬度最高的胜山样点红松的NV最小ND最大(表2), 这可能是资源的生产能力和存储量受低温环境限制而不足, 因此NV变小, 红松叶片投资策略转向防御并选择了更保守的叶片性状来缓冲低温对生长的影响(Buzzard et al., 2019), 且NV的变异速率大于叶干质量增加的速率导致ND变大. ...
Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora 2001
The worldwide leaf economics spectrum 1 2004
... Dong和Liu (2017)的研究结果表明红松(Pinus koraiensis)的多个功能性状受纬度不同所导致的环境差异的影响.多个叶片性状与纬度变化密切相关, 如叶片氮磷比随纬度升高而减小, 比叶面积(SLA)和叶寿命随纬度升高而增大(张林和罗天祥, 2004; De et al., 2013).叶寿命与叶片性状的显著相关性也多有论述, 如单位质量叶氮含量与叶寿命通常呈负相关关系(张林和罗天祥, 2004; Mediavilla et al., 2014; Osada et al., 2015), SLA随叶寿命的增大而减小(Kikuzawa & Lechowicz, 2011; de la Riva et al., 2016).He和Yan (2018)认为常绿树种性状的二元关系强度并不高, 但已有多个研究证实在广泛的植物类型和生物群落之间叶性状存在普遍相关性(Reich et al., 1999; Wright et al., 2004; Niinemets, 2015).红松针叶的SLA与叶片厚度呈显著的正线性相关关系, 但与叶干物质含量(LDMC)呈显著的负线性相关关系(Dong & Liu, 2017).叶片的SLA与体积也表现为显著负相关, 且SLA较小而叶片体积大的植物资源利用性更高(王晓洁等, 2015).叶片性状之间的关系往往反映了植物对不同环境下养分资源的适应(Chen et al., 2012), 因此, 揭示种内叶片性状的共变模式, 对于确定叶片是如何调整资源利用策略来适应环境变化是至关重要的, 但这种共变策略是否受纬度梯度的影响仍不清晰. ...
Leaf age as a factor in anatomical and physiological acclimative responses of Taxus baccata L. needles to contrasting irradiance environments 1 2007
... 叶经济谱性状很大程度上保留了各自独立响应表型变异驱动因素的能力, 驱动因素的影响因尺度而异(Martin et al., 2017).作为叶经济谱的一个重要性状, 很多研究已证明SLA对种内变异和环境普遍敏感(Adler et al., 2014; Messier et al., 2017).SLA可塑性相对较大, LDMC的可塑性低于SLA (表2), 这与de la Riva等(2016)的研究结论相符.当年生叶片与所有龄级表现的可塑性大小不同, 这可能是不同年龄针叶之间的光适应能力差异导致的, 尤其是当年生针叶容易受日照强度影响做出适应性调整(Wyka et al., 2007).不同针叶年龄下响应策略不同也可能是叶片可塑性大小存在较大差异的原因, De Kroon等(2005)曾提出, 在生长发育阶段植物做出的可塑性响应分别包括决定分生组织的营养或生殖去向以及对植物各部分尺寸和形状的调整, 而在成熟阶段表达为诱导防御等. ...
Photosynthetic ecophysiology of evergreen leaves in the woody angiosperms: a review 1 2014
 ,1,2,*
,1,2,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT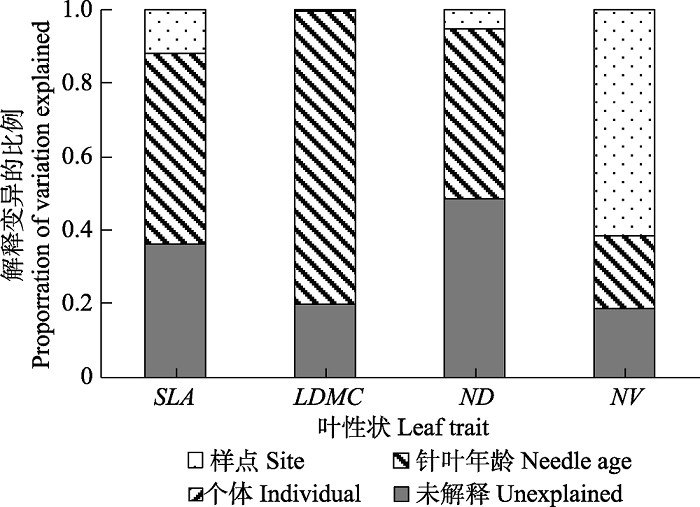 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT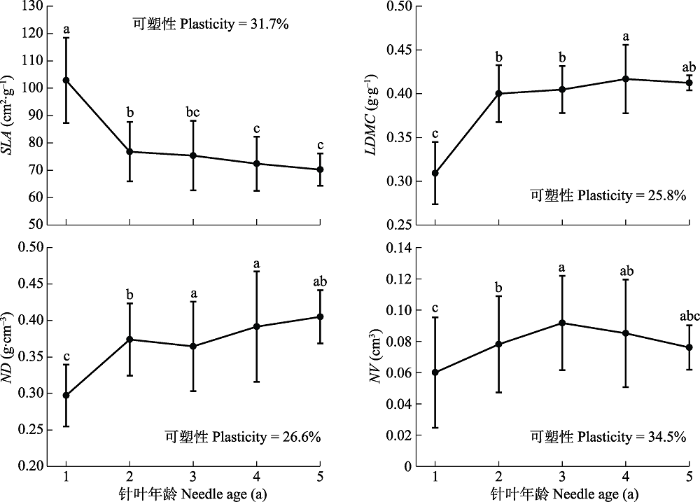 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT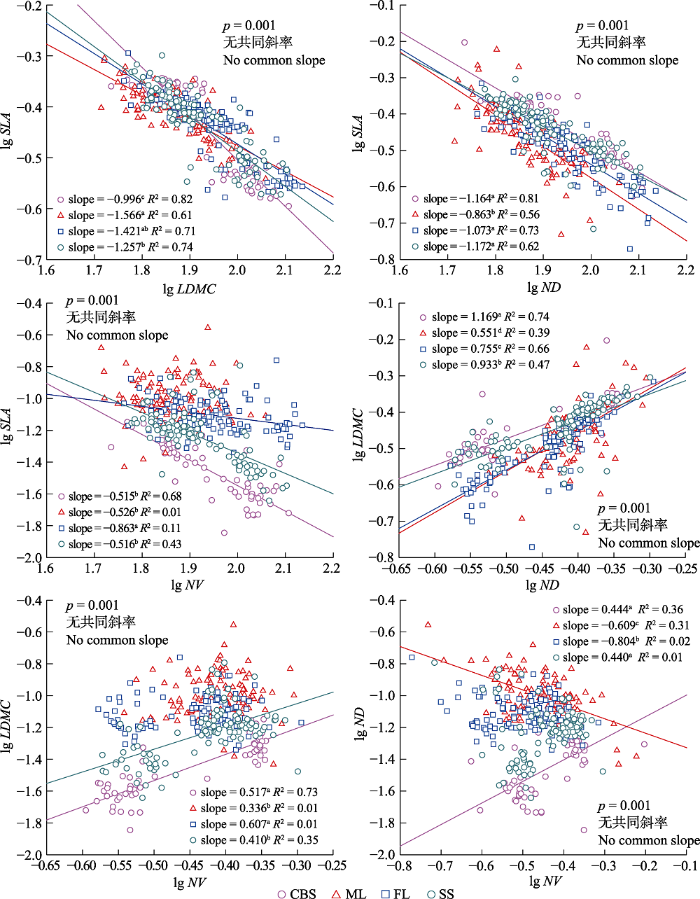 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT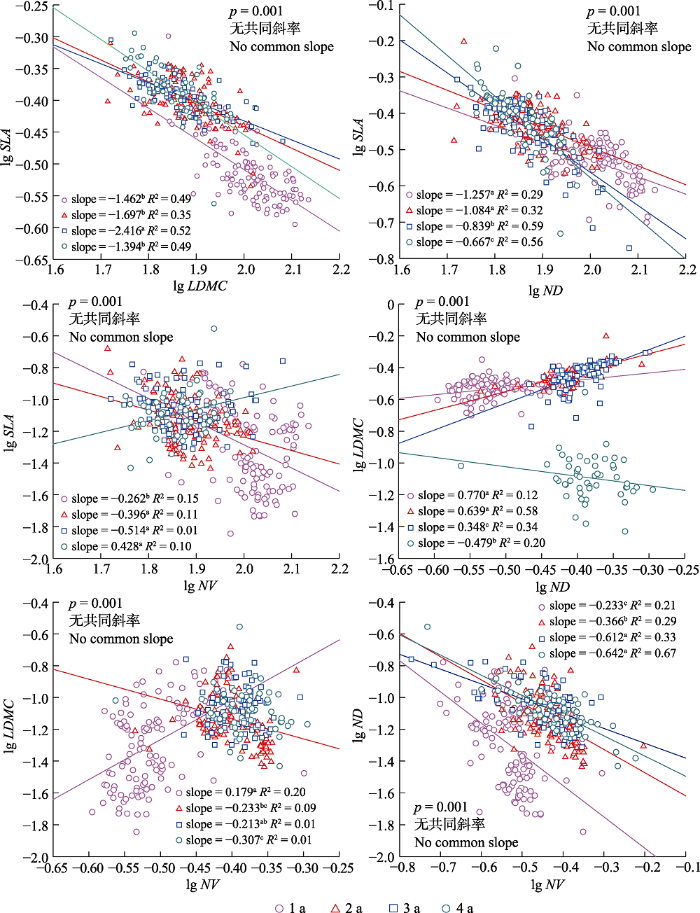 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}