Pressure-volume curve analysis of epiphytic lichens and its applicability in subtropical forests
Lin-Lin DONG1, Xiao-Yan PU2, Lu-Lu ZHANG,1,*, Liang SONG2, Zhi-Yun LU2,3, Su LI,,2,*1Institute of Environment and Ecology, Shandong Normal University, Jinan 250358, China 2CAS Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Kunming 650223, China 3Ailaoshan Station for Subtropical Forest Ecosystem Studies, Chinese Academy of Sciences, Jingdong, Yunnan 676209, China
Corresponding authors: * (Li S:lis@xtbg.ac.cn;Zhang LL:675359138@qq.com) Received:2020-10-20Accepted:2021-01-2Online:2021-03-20
Fund supported:
National Natural Science Foundation of China(31770494) National Natural Science Foundation of China(3175001) Youth Innovation Promotion Association of the Chinese Academy of Sciences (CAS)(2017441) CAS “Light of West China” Program, and the CAS 135 Program(2017XTBG-T01)
Abstract Aims Epiphytic lichens are a widespread and significant component of forest ecosystems, and play an important role in biodiversity maintaining and water and nutrient cycling. This study aims to illustrate the water potential characteristics of the epiphytic lichen group and to assess whether the pressure-volume (PV) curve and relative parameters are suitable for exploring the response of lichens to drought stress. Methods The water potential parameters and PV curves of 15 epiphytic lichen species from five functional groups were measured in the subtropical forests in the Ailao Mountains, Yunnan Province. Important findings The internal water content (WCinternal) and symplast water content (Rs) of cyanolichens were significantly higher than those of chlorolichens, while other parameters showed no significant differences. We only observed significant differences in WCinternal, Rs and relative water content at turgor loss point (RWCTLP) among different functional groups of lichens. All parameters differed significantly among species. Results of principal component analysis (PCA) further indicated that there are extensive limitations for PV curves and water potential parameters in evaluating the overall response of lichen species to water stress and in revealing the selection strategy of water-related habitats, largely due to the influence of the photobiont type and growth form. However, the adaptation strategies of cyanolichens to habitat could be explained by the maximum internal water holding capacity, while the adaptation to water conditions could be explained by the saturated water osmotic potential (Ψsat) in narrowly lobed foliose lichens, and by the RWCTLP in broadly lobed foliose and fruticose lichens. Our study suggests that PV curves and water potential parameters are not suitable for the general evaluation of the drought resistance of lichen communities, and should be used carefully in comparing the drought resistances between lichens and other plant groups. Keywords:epiphytic lichen;water potential;internal water content;pressure-volume curve;principal component analysis;poikilohydric plant
PDF (1856KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 董琳琳, 普晓妍, 张璐璐, 宋亮, 鲁志云, 李苏. 亚热带森林附生地衣压力-体积曲线分析及其适用性. 植物生态学报, 2021, 45(3): 274-285. DOI: 10.17521/cjpe.2020.0344 DONG Lin-Lin, PU Xiao-Yan, ZHANG Lu-Lu, SONG Liang, LU Zhi-Yun, LI Su. Pressure-volume curve analysis of epiphytic lichens and its applicability in subtropical forests. Chinese Journal of Plant Ecology, 2021, 45(3): 274-285. DOI: 10.17521/cjpe.2020.0344
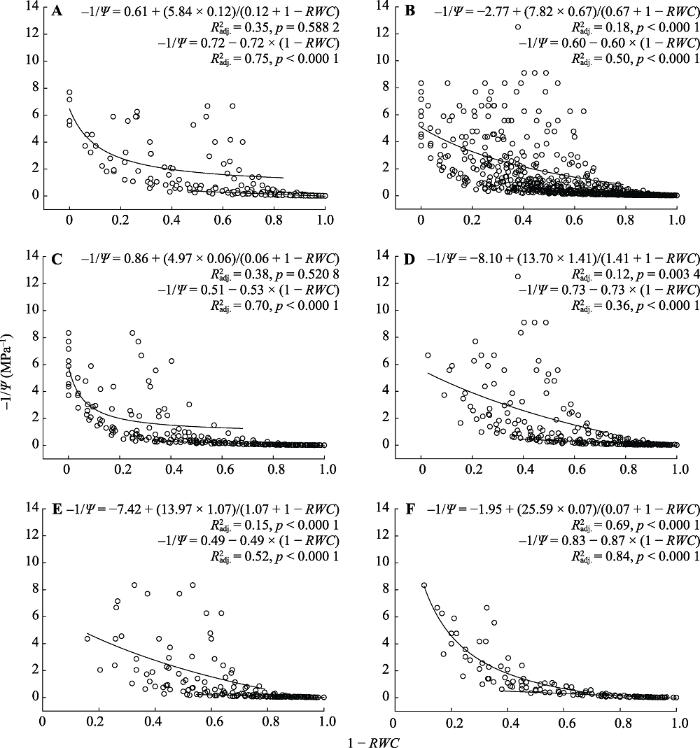
Fig. 1Pressure-volume curves of epiphytic lichen functional groups in subtropical forests in the Ailao Mountains, Yunnan. A, Cyanolichens. B, Chlorolichens. C, Broadly lobed foliose chlorolichens. D, Narrowly lobed foliose chlorolichens. E, Shrubby fruticose chlorolichens. F, Pendent fruticose chlorolichens. Ψ, water potential; RWC, relative water content.
Table 2 表2 表2云南哀牢山亚热带森林附生地衣的水势参数(平均值±标准误) Table 2Water potential parameters (mean ± SE) of epiphytic lichens in subtropical forests in the Ailao Mountains, Yunnan
共生藻型 Photobi-ont type
生长型 Growth form
物种 Species
内部含水量 WCinternal (%, Dry mass)
共质体水 Rs (%, Dry mass)
质外体水 Ra (%, Dry mass)
膨压损失点的 相对含水量 RWCTLP(%)
膨压损失点的 渗透势ΨTLP (MPa)
饱和渗透势 Ψsat (MPa)
蓝藻 Cyanob-acteria
328.66 ± 36.84 Ac
322.08 ± 36.79 Ac
6.58 ± 1.19 Aab
40.96 ± 3.97 Abc
-3.44 ± 0.33 Aa
-1.29 ± 0.13 Aab
叶状 Foliose
网肺衣 Lobaria retigera
214.86 ± 11.54
208.02 ± 11.14
6.85 ± 2.64
55.34 ± 2.95
-2.84 ± 0.35
-1.55 ± 0.24
猫耳衣 Leptogium menziesii
497.80 ± 43.93
492.40 ± 43.56
5.40 ± 1.44
22.68 ± 1.49
-3.86 ± 0.53
-0.85 ± 0.14
黑芽牛皮叶 Sticta fuliginosa
273.31 ± 32.08
265.81 ± 29.86
7.49 ± 2.30
44.86 ± 3.91
-3.63 ± 0.77
-1.46 ± 0.20
绿藻 Green alga
155.28 ± 8.95 B
149.71 ± 8.73 B
5.57 ± 0.45 A
46.93 ± 1.54 A
-3.80 ± 0.22 A
-1.71 ± 0.11 A
阔叶型 Broadly lobed foliose
137.05 ± 7.67 ab
130.05 ± 7.18 ab
7.00 ± 0.80 a
56.89 ± 2.73 a
-3.45 ± 0.30 a
-1.89 ± 0.18 c
针芽肺衣 Lobaria isidiophora
133.11 ± 10.40
128.03 ± 9.95
5.08 ± 1.20
65.68 ± 3.14
-3.85 ± 0.37
-2.47 ± 0.24
网大叶梅 Parmotrema reticulatum
119.18 ± 12.49
112.58 ± 11.77
6.60 ± 0.79
45.66 ± 3.44
-3.41 ± 0.78
-1.44 ± 0.32
平滑牛皮叶 Sticta nylanderiana
158.87 ± 12.26
149.55 ± 11.44
9.32 ± 1.57
59.34 ± 2.00
-3.08 ± 0.34
-1.75 ± 0.18
狭叶型 Narrowly lobed foliose
185.42 ± 23.36 ab
179.71 ± 22.70 ab
5.71 ± 0.93 ab
45.75 ± 2.23 bc
-3.74 ± 0.43 ab
-1.72 ± 0.23 abc
黑腹绵腹衣 Anzia hypomelaena
349.87 ± 25.89
339.44 ± 25.63
10.43 ± 1.08
33.88 ± 1.99
-1.58 ± 0.19
-0.50 ± 0.07
短根条衣 Everniastrum nepalense
115.29 ± 6.35
111.44 ± 6.40
3.86 ± 0.40
48.52 ± 2.02
-4.74 ± 0.75
-2.28 ± 0.44
卷梢哑铃孢 Heterodermia boryi
109.69 ± 9.46
107.74 ± 9.72
1.95 ± 0.46
51.66 ± 1.29
-4.98 ± 0.81
-2.52 ± 0.42
云南袋衣 Hypogymnia yunnanensis
166.81 ± 7.57
160.20 ± 7.77
6.61 ± 2.20
48.94 ± 6.04
-3.64 ± 0.72
-1.56 ± 0.25
直立枝状 Shrubby fruticose
153.06 ± 10.08 a
149.03 ± 9.88 a
4.03 ± 0.77 b
39.15 ± 2.46 c
-4.82 ± 0.45 b
-1.85 ± 0.23 bc
裂髓树花 Ramalina conduplicans
148.43 ± 14.35
142.97 ± 13.80
5.46 ± 1.10
48.02 ± 2.01
-5.82 ± 0.89
-2.70 ± 0.42
槽枝衣 Sulcaria sulcata
128.74 ± 8.78
125.69 ± 8.03
3.05 ± 1.80
40.92 ± 2.38
-3.97 ± 0.54
-1.56 ± 0.16
多花松萝 Usnea florida
182.00 ± 20.55
178.44 ± 20.20
3.57 ± 0.96
28.52 ± 2.35
-4.68 ± 0.79
-1.29 ± 0.26
悬垂枝状 Pendent fruticose
125.67 ± 10.85 b
120.23 ± 10.56 b
5.44 ± 0.78 ab
46.03 ± 3.63 b
-2.92 ± 0.28 a
-1.19 ± 0.06 a
巢松萝 Usnea dasopoga
153.47 ± 10.12
148.05 ± 9.08
5.43 ± 1.20
40.86 ± 3.71
-3.05 ± 0.33
-1.14 ± 0.10
长松萝 Usnea longissima
97.86 ± 6.33
92.41 ± 5.72
5.45 ± 1.15
51.20 ± 5.65
-2.79 ± 0.48
-1.24 ± 0.08
不同字母表示差异显著(p< 0.05)。大写字母表示共生藻类型之间的比较, 小写字母表示不同功能群的比较。 Ψsat, saturation water osmotic potential; ΨTLP, water potential at turgor loss point;Ra, apoplastic water;Rs, symplastic water;RWCTLP, relative water content at turgor loss point; WCinternal, internal water content. Different letters indicate significant difference (p < 0.05), the uppercase letters indicate the comparison between different photobiont types, and the lowercase letters indicate the comparison between different functional groups.
Table 3 表3 表3云南哀牢山亚热带森林15种附生地衣水势参数在主成分分析(PCA)中的载荷和解释方差 Table 3Principal component analysis (PCA) loading and interpretation variances of water potential parameters of 15 epiphytic lichens in subtropical forests in the Ailao Mountains, Yunnan
水势参数 Water potential parameter
PC1
PC2
共质体水 Rs
0.584
-
质外体水 Ra
0.307
-0.818
质壁分离时相对含水量 RWCTLP
-0.494
-0.565
饱和渗透势 Ψsat
0.566
-
标准差 Standard deviation
1.451
1.038
方差贡献率 Proportion of variance
52.66%
26.94%
方差累计贡献率 Cumulative proportion
52.66%
79.60%
Ψsat, saturation water osmotic potential; Ra, apoplastic water;Rs, symplastic water;RWCTLP, relative water content at turgor loss point.
Supplement III PV curve parameters of epiphytic lichens in subtropical forests in the Ailao Mountains in Yunnan and results of one sample t-test between liches and other plant growth forms (mean ± SE)
BarkmanJJ (1958). Phytosociology and Ecology of Cryptogamic Epiphytes. van Gorcum, Assen, Netherlands. [本文引用: 1]
BeckettRP (1997). Pressure-volume analysis of a range of poikilohydric plants implies the existence of negative turgor in vegetative cells Annals of Botany, 79, 145-152. DOI:10.1006/anbo.1996.0318URL [本文引用: 1]
CaiJR, QianTH, LeiJL (2015). Evaluation of drought resistance of five wild shrub species in South China Ecological Science, 34, 94-103. [本文引用: 1]
Castro-DíezP, NavarroJ (2007). Water relations of seedlings of three Quercus species: variations across and within species grown in contrasting light and water regimes Tree Physiology, 27, 1011-1018. DOI:10.1093/treephys/27.7.1011URL [本文引用: 1]
ChenK, LiuWY, LiS, SongL (2014). Photosynthetic characteristics of three epiphytic lichens under different water conditions Plant Diversity and Resources, 36, 603-610. [本文引用: 2]
ErikssonA, GauslaaY, PalmqvistK, Ekstr?mM, EsseenPA (2018). Morphology drives water storage traits in the globally widespread lichen genus Usnea Fungal Ecology, 35, 51-61. DOI:10.1016/j.funeco.2018.06.007URL [本文引用: 4]
EsseenPA, R?nnqvistM, GauslaaY, CoxsonDS (2017). Externally held water—A key factor for hair lichens in boreal forest canopies Fungal Ecology, 30, 29-38. DOI:10.1016/j.funeco.2017.08.003URL [本文引用: 3]
GauslaaY, CoxsonDS (2011). Interspecific and intraspecific variations in water storage in epiphytic old forest foliose lichens Botany, 89, 787-798. DOI:10.1139/b11-070URL [本文引用: 1]
HartardB, CuntzM, MáguasC, LakatosM (2009). Water isotopes in desiccating lichens Planta, 231, 179-193. DOI:10.1007/s00425-009-1038-8PMID:19888598 [本文引用: 1] The stable isotopic composition of water is routinely used as a tracer to study water exchange processes in vascular plants and ecosystems. To date, no study has focussed on isotope processes in non-vascular, poikilohydric organisms such as lichens and bryophytes. To understand basic isotope exchange processes of non-vascular plants, thallus water isotopic composition was studied in various green-algal lichens exposed to desiccation. The study indicates that lichens equilibrate with the isotopic composition of surrounding water vapour. A model was developed as a proof of concept that accounts for the specific water relations of these poikilohydric organisms. The approach incorporates first their variable thallus water potential and second a compartmentation of the thallus water into two isotopically distinct but connected water pools. Moreover, the results represent first steps towards the development of poikilohydric organisms as a recorder of ambient vapour isotopic composition.
HeXD, CongPF, GaoYB, LuJG, WangHT, XuePP, ZhangX (2007). Drought resistance of four grasses using pressure- volume curve Frontiers of Biology in China, 2, 425-430. DOI:10.1007/s11515-007-0065-8URL [本文引用: 1]
HoneggerR, PeterM, ScherrerS (1996). Drought-induced structural alterations at the mycobiont-photobiont interface in a range of foliose macrolichens Protoplasma, 190, 221-232. DOI:10.1007/BF01281320URL [本文引用: 1]
HuT, LiS, LiuS, LiuWY, ChenX, SongL, ChenQ (2016). Water relations and photosynthetic characteristics in different functional groups of epiphytic lichens in montane forest of Ailaoshan Chinese Journal of Plant Ecology, 40, 810-826. DOI:10.17521/cjpe.2015.0445URL [本文引用: 3]
LangeOL (2003). Photosynthetic productivity if the epilithic lichen Lecanora muralis: long-term field monitoring of CO2 exchanges physiological interpretation. Diel, seasonal, and annual carbon budgets Flora, 198, 277-292. [本文引用: 1]
LangeOL, ZieglerH (1986). Different Limiting Processes of Photosynthesis in Lichens. Biological Control of Photosynthesis. Martinus Nijhoff Publishers, Dordrecht.147-161. [本文引用: 3]
LiHJ, DiXY, ChenJW, WangMB (2004). A method of using SigmaPlot to calculate the moisture parameter ΨTLP of the PV curve Plant Research, 24, 71-75. [本文引用: 1]
LiS, LiuWY, LiDW (2013a). Bole epiphytic lichens as potential indicators of environmental change in subtropical forest ecosystems in southwest China Ecological Indicators, 29, 93-104. DOI:10.1016/j.ecolind.2012.12.012URL [本文引用: 7]
LiS, LiuWY, LiDW (2013b). Epiphytic lichens in subtropical forest ecosystems in southwest China: species diversity and implications for conservation Biological Conservation, 159, 88-95. DOI:10.1016/j.biocon.2012.12.027URL [本文引用: 2]
LiS, LiuWY, WangLS, MaWZ, SongL (2011). Biomass, diversity and composition of epiphytic macrolichens in primary and secondary forests in the subtropical Ailao Mountains, SW China Forest Ecology and Management, 261, 1760-1770. DOI:10.1016/j.foreco.2011.01.037URL [本文引用: 1]
LiS, LiuWY, WangLS, YangGP, LiDW (2007). Species diversity and distribution of epiphytic lichens in the primary and secondary forests in Ailao Mountain, Yunnan Biodiversity Science, 15, 445-455. DOI:10.1360/biodiv.070078URL [本文引用: 1]
LonginottiS, SolhaugK, GauslaaY (2017). Hydration traits in cephalolichen members of the epiphytic old forest genus Lobaria (s. lat.) The Lichenologist, 49, 493-596. DOI:10.1017/S0024282917000445URL [本文引用: 1]
McCuneB (1993). Gradients in epiphyte biomass in three Pseudotsuga-Tsuga forests of different ages in western Oregon and Washington The Bryologist, 96, 405-411. DOI:10.2307/3243870URL [本文引用: 1]
MotalebifardR, NajafiN, OustanS, NyshabouriMR, ValizadehM (2013). The combined effects of phosphorus and zinc on evapotranspiration, leaf water potential, water use efficiency and tuber attributes of potato under water deficit conditions Scientia Horticulturae, 162, 31-38. DOI:10.1016/j.scienta.2013.07.043URL [本文引用: 1]
NardiniA, MarchettoA, TretiachM (2013). Water relation parameters of six Peltigera species correlate with their habitat preferences Fungal Ecology, 6, 397-407. DOI:10.1016/j.funeco.2013.05.004URL [本文引用: 3]
NimisPL, MartellosS (2008). ITALIC-The Information System on Italian Lichens. Version 4.0. [2015-12-07]. http://dbiodbs.univ.trieste.it/. URL [本文引用: 1]
OlsonDM, DinersteinE (1998). The global 200: a representation approach to conserving the earth’s most biologically valuable ecoregions Conservation Biology, 12, 502-515. DOI:10.1046/j.1523-1739.1998.012003502.xURL [本文引用: 1]
SaruwatariMW, DavisSD (1989). Tissue water relations of three chaparral shrub species after wildfire Oecologia, 80, 303-308. DOI:10.1007/BF00379031PMID:28312057 [本文引用: 1] We compared the tissue water relations among resprouts and seedlings of three chaparral species during the first summer drought after wildfire. Two of the species, Rhus laurina and Ceanothus spinosus recover after fire by a combination of resprouting and seedling establishment (facultative resprouters), whereas a third species, Ceanothus megacarpus recovers by seedling establishment alone (obligate seeder). Our objectives were to document any differences in tissue water characteristics that might arise between resprouts and seedlings and to test the hypothesis that seedlings of obligate seeders develop more drought tolerant characteristics of their tissues than seedlings of facultative resprouters. We found that resprouts had much higher predawn values of water potential, osmotic potential, and turgor potentials than seedlings. Predawn turgor potentials of resprouts were 1.5 MPa through July and August when turgor potentials for seedlings remained near 0 MPa. During summer months, midday water potentials were 2 to 3 MPa higher for resprouts than seedlings and midday conductances of resprouts were two to five fold greater than those of seedlings. Even though resprouts did not experience severe water stress like seedlings, their tissue water characteristics, as determined by pressure-volume curve analyses, were similar by the peak of the drought in August. Further-more, the tissue water characteristics of seedlings from the obligate seeder, C. megacarpus, were similar to those of facultative resprouters - R. laurina, and C. spinosus. We attribute the observed differences in plant water status between resprouts and seedlings to differences in rooting depths and access to soil moisture reserves during summer drought. We conclude that the higher growth rates, photosynthetic performance, and survivorship of postfire resprouts are primarily a result of higher water availability to resprouting tissues during summer months. It appears that the greater seedling survivorship during summer drought observed for the obligate seeder, C. megacarpus, is not associated with more favorable tissue water characteristics.
SatoT, AbdallaOS, OweisTY, SakurataniT (2006). The validity of predawn leaf water potential as an irrigation- timing indicator for field-grown wheat in northern Syria Agricultural Water Management, 82, 223-236. DOI:10.1016/j.agwat.2005.07.023URL [本文引用: 1]
SmithDC, MolesworthS (1973). Lichen physiology xiii. Effects of rewetting dry lichens New Phytologist, 72, 525-533. DOI:10.1111/nph.1973.72.issue-3URL [本文引用: 1]
SongL, ZhangYJ, ChenX, LiS, LuHZ, WuCS, TanZH, LiuWY, ShiXM (2015). Water relations and gas exchange of fan bryophytes and their adaptations to microhabitats in an Asian subtropical montane cloud forest Journal of Plant Research, 128, 573-584. DOI:10.1007/s10265-015-0721-zURL [本文引用: 4]
TyreeMT, HammelHT (1972). The measurement of the turgor pressure and the water relations of plants by the pressure- bomb technique Journal of Experimental Botany, 23, 267-282. DOI:10.1093/jxb/23.1.267URL [本文引用: 3]
VriesMC, WatlingJR (2008). Differences in the utilization of water vapour and free water in two co-occurring foliose lichens from semi-arid southern Australia Austral Ecology, 33, 975-985. DOI:10.1111/j.1442-9993.2008.01868.xURL [本文引用: 1]
WangLS (1995). Anzia physowea, a lichen new to China Mycosystema, 14, 313-314. [本文引用: 1]
WenkertW, LemonER, SinclairTR (1978). Water content potential relationship in soya bean: changes in component potentials for mature and immature leaves under ?eld conditions Annals of Botany, 42, 295-307. DOI:10.1093/oxfordjournals.aob.a085460URL [本文引用: 1]
YanMJ, YamamotoM, YamanakaN, YamamotoF, LiuGB, DuS (2013). A comparison of pressure-volume curves with and without rehydration pretreatment in eight woody species of the semiarid Loess Plateau Acta Physiologiae Plantarum, 35, 1051-1060. [本文引用: 3]
ZhangLS, ZhangHT, HuJJ, QuanJ, XuSR, HanMY, MaFW (2013). The response of pressure volume curve water parameters and root system hydraulic architecture of two apple rootstocks to drought stress Acta Ecologica Sinica, 33, 3324-3331. DOI:10.5846/stxbURL [本文引用: 1]
ZhengYX, WuJC, CaoFL, ZhangYP (2010). Effects of water stress on photosynthetic activity, dry mass partitioning and some associated metabolic changes in four provenances of neem (Azadirachta indica A. Juss) Photosynthetica, 48, 361-369. DOI:10.1007/s11099-010-0047-yURL [本文引用: 1]
ZhuH, YanLC (2009). List of Seed Plants in the Ailao Mts. of Yunnan Province, China. Yunnan Science and Technology Press, Kunming. [本文引用: 1]
Photosynthetic productivity if the epilithic lichen Lecanora muralis: long-term field monitoring of CO2 exchanges physiological interpretation. Diel, seasonal, and annual carbon budgets 1 2003
The combined effects of phosphorus and zinc on evapotranspiration, leaf water potential, water use efficiency and tuber attributes of potato under water deficit conditions 1 2013
... 压力-体积曲线(PV曲线)是使用最广泛的表征植物水分状况最广泛使用的工具(Wenkert et al., 1978).通过测量植物组织从完全饱和状态直至脱水干燥以后失水全过程的水势(Ψ)与相对含水量(RWC)的变化, 绘制饱和叶片失水过程中叶水势的负倒数(-1/Ψ)和相对含水量(1 - RWC)之间关系的PV曲线, 获取和评估共质体含水量(Rs)、质外体含水量(Ra)、膨压损失点的相对含水量(RWCTLP)和渗透势(ΨTLP)、以及饱和渗透势(Ψsat)等参数(附录I).这些参数综合体现了植物组织内部的水分状况及潜在忍耐力, 是衡量植物应对环境干旱胁迫能力的重要指标, 广泛应用于草本(Sato et al., 2006; Motalebifard et al., 2013), 灌木(Saruwatari & Davis, 1989; 蔡静如等, 2015), 乔木(Zheng et al., 2010; Yan et al., 2013; 张林森等, 2013)类群耐旱性的评价.然而, 对于变水生物如苔藓和地衣来说, PV曲线及相关参数是否适用于其干旱胁迫响应的分析, 尚未得到广泛研究和一致结论.少量研究证明, 部分地卷属(Peltigera)地衣具有较低的Ψsat和ΨTLP, 在干燥生境中具有明显的生存优势(Nardini et al., 2013). ...
Water relation parameters of six Peltigera species correlate with their habitat preferences 3 2013
... 压力-体积曲线(PV曲线)是使用最广泛的表征植物水分状况最广泛使用的工具(Wenkert et al., 1978).通过测量植物组织从完全饱和状态直至脱水干燥以后失水全过程的水势(Ψ)与相对含水量(RWC)的变化, 绘制饱和叶片失水过程中叶水势的负倒数(-1/Ψ)和相对含水量(1 - RWC)之间关系的PV曲线, 获取和评估共质体含水量(Rs)、质外体含水量(Ra)、膨压损失点的相对含水量(RWCTLP)和渗透势(ΨTLP)、以及饱和渗透势(Ψsat)等参数(附录I).这些参数综合体现了植物组织内部的水分状况及潜在忍耐力, 是衡量植物应对环境干旱胁迫能力的重要指标, 广泛应用于草本(Sato et al., 2006; Motalebifard et al., 2013), 灌木(Saruwatari & Davis, 1989; 蔡静如等, 2015), 乔木(Zheng et al., 2010; Yan et al., 2013; 张林森等, 2013)类群耐旱性的评价.然而, 对于变水生物如苔藓和地衣来说, PV曲线及相关参数是否适用于其干旱胁迫响应的分析, 尚未得到广泛研究和一致结论.少量研究证明, 部分地卷属(Peltigera)地衣具有较低的Ψsat和ΨTLP, 在干燥生境中具有明显的生存优势(Nardini et al., 2013). ...
两种苹果砧木根系水力结构及其PV曲线水分参数对干旱胁迫的响应 1 2013
... 压力-体积曲线(PV曲线)是使用最广泛的表征植物水分状况最广泛使用的工具(Wenkert et al., 1978).通过测量植物组织从完全饱和状态直至脱水干燥以后失水全过程的水势(Ψ)与相对含水量(RWC)的变化, 绘制饱和叶片失水过程中叶水势的负倒数(-1/Ψ)和相对含水量(1 - RWC)之间关系的PV曲线, 获取和评估共质体含水量(Rs)、质外体含水量(Ra)、膨压损失点的相对含水量(RWCTLP)和渗透势(ΨTLP)、以及饱和渗透势(Ψsat)等参数(附录I).这些参数综合体现了植物组织内部的水分状况及潜在忍耐力, 是衡量植物应对环境干旱胁迫能力的重要指标, 广泛应用于草本(Sato et al., 2006; Motalebifard et al., 2013), 灌木(Saruwatari & Davis, 1989; 蔡静如等, 2015), 乔木(Zheng et al., 2010; Yan et al., 2013; 张林森等, 2013)类群耐旱性的评价.然而, 对于变水生物如苔藓和地衣来说, PV曲线及相关参数是否适用于其干旱胁迫响应的分析, 尚未得到广泛研究和一致结论.少量研究证明, 部分地卷属(Peltigera)地衣具有较低的Ψsat和ΨTLP, 在干燥生境中具有明显的生存优势(Nardini et al., 2013). ...
Effects of water stress on photosynthetic activity, dry mass partitioning and some associated metabolic changes in four provenances of neem (Azadirachta indica A. Juss) 1 2010
... 压力-体积曲线(PV曲线)是使用最广泛的表征植物水分状况最广泛使用的工具(Wenkert et al., 1978).通过测量植物组织从完全饱和状态直至脱水干燥以后失水全过程的水势(Ψ)与相对含水量(RWC)的变化, 绘制饱和叶片失水过程中叶水势的负倒数(-1/Ψ)和相对含水量(1 - RWC)之间关系的PV曲线, 获取和评估共质体含水量(Rs)、质外体含水量(Ra)、膨压损失点的相对含水量(RWCTLP)和渗透势(ΨTLP)、以及饱和渗透势(Ψsat)等参数(附录I).这些参数综合体现了植物组织内部的水分状况及潜在忍耐力, 是衡量植物应对环境干旱胁迫能力的重要指标, 广泛应用于草本(Sato et al., 2006; Motalebifard et al., 2013), 灌木(Saruwatari & Davis, 1989; 蔡静如等, 2015), 乔木(Zheng et al., 2010; Yan et al., 2013; 张林森等, 2013)类群耐旱性的评价.然而, 对于变水生物如苔藓和地衣来说, PV曲线及相关参数是否适用于其干旱胁迫响应的分析, 尚未得到广泛研究和一致结论.少量研究证明, 部分地卷属(Peltigera)地衣具有较低的Ψsat和ΨTLP, 在干燥生境中具有明显的生存优势(Nardini et al., 2013). ...
1 2009
... 哀牢山位于云贵高原西南部、横断山区南段以东, 属云岭山脉向南分支的余脉, 是全球生物多样性保护热点Indo-Burma区域的一部分(Olson & Dinerstein, 1998).本研究位于哀牢山国家级自然保护区核心区域景东徐家坝片区(23.35°-24.44° N, 100.53°-101.30° E), 海拔约为2 400-2 750 m.该地区降雨丰沛, 终年温凉潮湿, 年降水量约1 947 mm, 年蒸发量约1 192 mm, 相对湿度约为85%; 月平均最高气温为15.8 ℃, 月平均最低气温为4.5 ℃, 年平均气温11.3 ℃ (李苏等,2007; 朱华和闫丽春, 2009).中山湿性常绿阔叶林是哀牢山徐家坝地区的主要原生森林植被, 林内附生生物种类繁多, 地衣是附生类群的重要组成部分, 目前已记录217种(Li et al., 2013a, 2013b). ...
1 2009
... 哀牢山位于云贵高原西南部、横断山区南段以东, 属云岭山脉向南分支的余脉, 是全球生物多样性保护热点Indo-Burma区域的一部分(Olson & Dinerstein, 1998).本研究位于哀牢山国家级自然保护区核心区域景东徐家坝片区(23.35°-24.44° N, 100.53°-101.30° E), 海拔约为2 400-2 750 m.该地区降雨丰沛, 终年温凉潮湿, 年降水量约1 947 mm, 年蒸发量约1 192 mm, 相对湿度约为85%; 月平均最高气温为15.8 ℃, 月平均最低气温为4.5 ℃, 年平均气温11.3 ℃ (李苏等,2007; 朱华和闫丽春, 2009).中山湿性常绿阔叶林是哀牢山徐家坝地区的主要原生森林植被, 林内附生生物种类繁多, 地衣是附生类群的重要组成部分, 目前已记录217种(Li et al., 2013a, 2013b). ...
 ,1,*, 宋亮2, 鲁志云2,3, 李苏
,1,*, 宋亮2, 鲁志云2,3, 李苏 ,
,

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT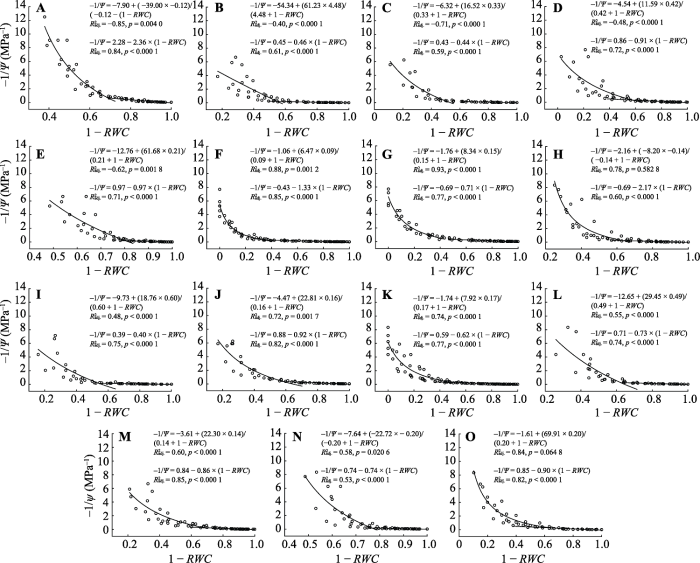 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT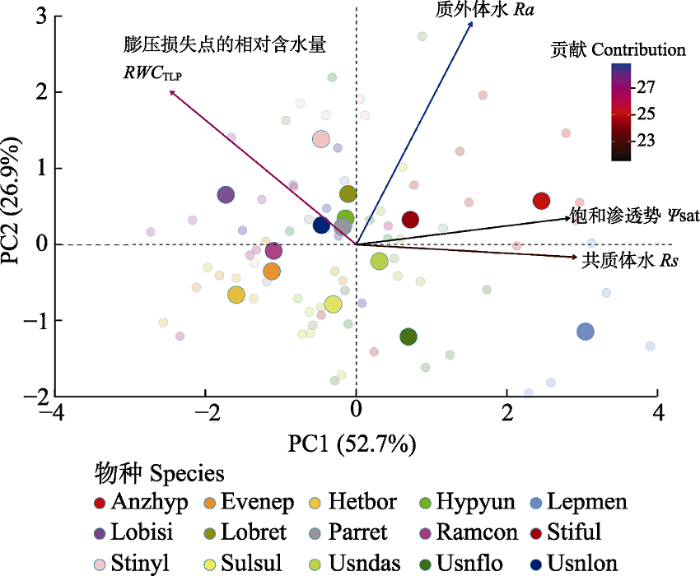 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}