Effects of sand burial on litter decomposition rate and salt content dynamics in an extremely arid region
Lin-Jie Fan1,2,4, Cheng-Dao LI1,2,4, Xiang-Yi LI,,1,3,*, J. SUN Henry5, Li-Sha LIN1,2,3, Bo LIU61Xinjiang Key Laboratory of Desert Plant Roots Ecology and Vegetation Restoration, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, ürümqi 830011, China 2State Key Laboratory of Desert and Oasis Ecology, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, ürümqi 830011, China 3Cele National Station of Observation and Research for Desert-Grassland Ecosystems, Cele, Xinjiang 848300, China 4University of Chinese Academy of Sciences, Beijing 100049, China 5Desert Research Institute, Las Vegas 89119, USA 6Linyi University, Linyi, Shandong 276000, China
Abstract Aims Due to the extremely low precipitation, low vegetation coverage, strong solar radiation, and poor soil stability, litter turnover in extremely arid areas differs from that in non-arid areas. This study aimed to determine the patterns of leaf litter decomposition of contrasting initial qualities in an extremely arid region. Methods We used the litter bag method to investigate changes of the mass and water-soluble salt content in the leaf litter of three dominant species, Karelinia caspia, Alhagi sparsifoliaand Populus euphratica,in the desert- oasis transitional zone of the southern edge of the Taklimakan Desert, in responses to three levels of sand burial treatments, including placement of letter samples at the surface, and 2 cm and 15 cm soil depths, respectively, that represented different incubation environments under natural conditions. Important findings The relationships of litter decomposition rate with the initial litter quality indicators, including carbon (C) content, nitrogen (N) content, C:N and lignin content, differed between the extremely arid sites and the non-arid sites. The litter placed on the surface had higher lignin content and faster mass loss than those subjected to other treatments. The losses of litter mass and changes in water-soluble salt content significantly varied with the level of burial treatments. Litter samples placed on the surface and at 2 cm depth had a significantly greater rate of losses in mass and water-soluble salt content than those at 15 cm depth. The surface litter had a greater amount of dissolved water-soluble salt in the early stage of decomposition. This study shows that the driving mechanism of litter decomposition in the extremely arid areas is unique. Under conditions of extremely low precipitation and the low activity of soil microorganisms, the buried depth is not the main factor driving the litter decomposition, whilst other abiotic processes such as solar radiation controlled the rate of decomposition. Keywords:litter decomposition;extremely arid area;burial depth;salt
PDF (1338KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 范琳杰, 李成道, 李向义, Henry J. SUN, 林丽莎, 刘波. 极端干旱区沙土掩埋对凋落物分解速率及盐分含量动态的影响. 植物生态学报, 2021, 45(2): 144-153. DOI: 10.17521/cjpe.2020.0273 Fan Lin-Jie, LI Cheng-Dao, LI Xiang-Yi, Henry J. SUN, LIN Li-Sha, LIU Bo. Effects of sand burial on litter decomposition rate and salt content dynamics in an extremely arid region. Chinese Journal of Plant Ecology, 2021, 45(2): 144-153. DOI: 10.17521/cjpe.2020.0273
凋落物在维持生态系统物质循环和能量流动方面发挥着重要作用(曾加芹, 2017)。凋落物分解是生态系统中碳周转和矿质养分循环的重要过程(Fu et al., 2009)。据统计, 全球每年因凋落物分解释放的CO2约占全球年碳通量的70% (Raich & Schlesinger, 1992)。在全球陆地生态系统中, 有80%的有机碳储存于土壤中(Eswaran et al., 1993), 而凋落物又是土壤碳输入的重要来源, 凋落物分解以可溶性有机碳和稳定态腐殖质的形式向土壤输入碳(Rubino et al., 2010)。因此, 了解凋落物分解的影响因素对于理解全球碳循环至关重要。
Fig. 2Ratio of remaining litter mass at different times of decomposition in extremely arid region (mean ± SE). A, Karelinia caspia. B, Alhagi sparsifolia. C, Populus euphratica. Different lowercase letters indicate that the significant differences between treatments (p < 0.05).
Fig. 3Values of the litter decomposition constant (k) for different burial treatments in extremely arid region (mean ± SE). Different lowercase letters indicate significant differences between species under the same treatments (p < 0.05).
Fig. 4Ratio of litter water-soluble salt residue at different times of decomposition in extremely arid region (mean ± SE). A, Karelinia caspia. B, Alhagi sparsifolia. C, Populus euphratica. Different lowercase letters indicate significant differences between treatments (p < 0.05).
Fig. 5Relationships between litter mass and water-soluble salt residue in extremely arid region. A, Surface. B, 2 cm depth. C, 15 cm depth. The solid line represents the fitting curve of the relationship, and the dotted line represents y = x (1:1).
Fig. 6Litter dissolved water-soluble salt at different times of decomposition in extremely arid region (mean ± SE). A, Karelinia caspia. B, Alhagi sparsifolia. C, Populus euphratica. Different lowercase letters indicate significant differences between treatments (p < 0.05).
Fig. 7Ratio of litter water-soluble salt residue of three species under different treatments in extremely arid region (mean ± SE). Different lowercase letters indicate significant differences between treatments (p < 0.05).
AnGX, ZengFJ, SunXW, LiuB, LiuZ, ZhangXL (2011). Soil water conditions under various vegetations in southern fringe of Takelamakan Desert Bulletin of Soil and Water Conservation, 31,63-67. [本文引用: 1]
AustinAT, AraujoPI, LevaPE (2009). Interaction of position, litter type, and water pulses on decomposition of grasses from the semiarid Patagonian steppe Ecology, 90,2642-2647. PMID:19769141 [本文引用: 1] Litter lignin and nutrient content, annual rainfall, and biotic activity are not good predictors of litter decomposition in arid and semiarid ecosystems, suggesting that other factors may be important in controlling carbon turnover. We explored the relative importance of litter position (above- vs. belowground), litter type (leaf vs. root), and pulsed water events (large vs. small) on mass loss with grass species of the semiarid Patagonian steppe. In a factorial experiment of mesocosms, we incubated leaf and root litter simultaneously above- and belowground and manipulated water availability with large and small pulses. Significant interactions between position and litter type and position and pulse sizes demonstrated interactive controls on organic mass loss. Aboveground decomposition showed no response to pulse size or litter type, as roots and leaves decomposed equally rapidly under all circumstances. In contrast, belowground decomposition was significantly altered by litter type and water pulses, with roots decomposing significantly slower and small water pulses reducing belowground decomposition. The results of this mesocosm experiment support the idea that controls other than water availability may dominate aboveground mass loss, while a combination of recalcitrant litter and water penetration in the soil profile are critical factors determining belowground decomposition, which is ultimately mediated by biotic degradation.
AustinAT, BallareCL (2010). Dual role of lignin in plant litter decomposition in terrestrial ecosystems Proceedings of the National Academy of Sciences of the United States of America, 107,4618-4622. DOI:10.1073/pnas.0909396107PMID:20176940 [本文引用: 1] Plant litter decomposition is a critical step in the formation of soil organic matter, the mineralization of organic nutrients, and the carbon balance in terrestrial ecosystems. Biotic decomposition in mesic ecosystems is generally negatively correlated with the concentration of lignin, a group of complex aromatic polymers present in plant cell walls that is recalcitrant to enzymatic degradation and serves as a structural barrier impeding microbial access to labile carbon compounds. Although photochemical mineralization of carbon has recently been shown to be important in semiarid ecosystems, litter chemistry controls on photodegradative losses are not understood. We evaluated the importance of litter chemistry on photodegradation of grass litter and cellulose substrates with varying levels of lignin [cellulose-lignin (CL) substrates] under field conditions. Using wavelength-specific light attenuation filters, we found that light-driven mass loss was promoted by both UV and visible radiation. The spectral dependence of photodegradation correlated with the absorption spectrum of lignin but not of cellulose. Field incubations demonstrated that increasing lignin concentration reduced biotic decomposition, as expected, but linearly increased photodegradation. In addition, lignin content in CL substrates consistently decreased in photodegradative incubations. We conclude that lignin has a dual role affecting litter decomposition, depending on the dominant driver (biotic or abiotic) controlling carbon turnover. Under photodegradative conditions, lignin is preferentially degraded because it acts as an effective light-absorbing compound over a wide range of wavelengths. This mechanistic understanding of the role of lignin in plant litter decomposition will allow for more accurate predictions of carbon dynamics in terrestrial ecosystems.
AustinAT, VivancoL (2006). Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation Nature, 442,555-558. PMID:16885982 [本文引用: 1] The carbon balance in terrestrial ecosystems is determined by the difference between inputs from primary production and the return of carbon to the atmosphere through decomposition of organic matter. Our understanding of the factors that control carbon turnover in water-limited ecosystems is limited, however, as studies of litter decomposition have shown contradictory results and only a modest correlation with precipitation. Here we evaluate the influence of solar radiation, soil biotic activity and soil resource availability on litter decomposition in the semi-arid Patagonian steppe using the results of manipulative experiments carried out under ambient conditions of rainfall and temperature. We show that intercepted solar radiation was the only factor that had a significant effect on the decomposition of organic matter, with attenuation of ultraviolet-B and total radiation causing a 33 and 60 per cent reduction in decomposition, respectively. We conclude that photodegradation is a dominant control on above-ground litter decomposition in this semi-arid ecosystem. Losses through photochemical mineralization may represent a short-circuit in the carbon cycle, with a substantial fraction of carbon fixed in plant biomass being lost directly to the atmosphere without cycling through soil organic matter pools. Furthermore, future changes in radiation interception due to decreased cloudiness, increased stratospheric ozone depletion, or reduced vegetative cover may have a more significant effect on the carbon balance in these water-limited ecosystems than changes in temperature or precipitation.
BaoSD (2000). Soil and Agricultural Chemistry Analysis. China Agriculture Press, Beijing.257-263. [本文引用: 1]
BergB (2000). Litter decomposition and organic matter turnover in northern forest soils Forest Ecology and Management, 133(1-2),13-22. DOI:10.1016/S0378-1127(99)00292-3URL [本文引用: 1]
BergB, LaskowskiR (2006). Litter decomposition: a guide to carbon and nutrient turnover //Luo Y. Advances in Ecological Research. Academic Press, London. [本文引用: 1]
DayTA, ZhangET, RuhlandCT (2007). Exposure to solar UV-B radiation accelerates mass and lignin loss of Larrea tridentata litter in the Sonoran Desert Plant Ecology, 193,185-194. DOI:10.1007/s11258-006-9257-6URL [本文引用: 1]
DongZW, ZhaoY, LeiJQ, XiYQ (2018). Distribution pattern and influencing factors of soil salinity at Tamarix cones in the Taklimakan Desert Chinese Journal of Plant Ecology, 42,873-884. DOI:10.17521/cjpe.2018.0060URL [本文引用: 1]
FuS, ZouX, ColemanD (2009). Highlights and perspectives of soil biology and ecology research in China Soil Biology & Biochemistry, 41,868-876. DOI:10.1016/j.soilbio.2008.10.014URL [本文引用: 1]
GallardoA, MerinoJ (1993). Leaf decomposition in two Mediterranean ecosystems of southwest Spain: influence of substrate quality Ecology, 74,152-161. DOI:10.2307/1939510URL [本文引用: 1]
GalloME, SinsabaughRL, CabanissSE (2006). The role of ultraviolet radiation in litter decomposition in arid ecosystems Applied Soil Ecology, 34,82-91. DOI:10.1016/j.apsoil.2005.12.006URL [本文引用: 1]
GeorgiouCD, SunHJ, McKayCP, GrintzalisK, PapapostolouI, ZisimopoulosD, PanagiotidisK, ZhangG, KoutsopoulouE, ChristidisGE, MargiolakiI (2015). Evidence for photochemical production of reactive oxygen species in desert soils Nature Communications, 6,7100. DOI: 10.1038/ncomms8100. DOI:10.1038/ncomms8100URL [本文引用: 1]
GholzHL, WedinDA, SmithermanSM, HarmonME, PartonWJ (2000). Long-term dynamics of pine and hardwood litter in contrasting environments: toward a global model of decomposition Global Change Biology, 6,751-765. DOI:10.1046/j.1365-2486.2000.00349.xURL [本文引用: 1]
HeYH, LiuXP, XieZK (2015). Enrichment of soil salinity and nutrients under desertification shrub Reaumuria soongorica Journal of Arid Land Resources and Environment, 29(3),115-119. [本文引用: 1]
JiaoJ, ZouH, JiaY, WangN (2009). Research progress on the effects of soil erosion on vegetation Acta Ecologica Sinica, 29(2),85-91. DOI:10.1016/j.chnaes.2009.05.001URL [本文引用: 1]
LiCD, LIXY, SunHJ, LIL, LinLS (2019). Decomposition characteristics of Karelinia caspia, Alhagi sparsifolia and Populus euphratica leaves in extremely arid areas Journal of Desert Research, 39(2),193-201. [本文引用: 1]
LiuGF, CornwellWK, PanX, YeD, LiuFH, HuangZY, DongM, CornelissenJHC (2015). Decomposition of 51 semidesert species from wide-ranging phylogeny is faster in standing and sand-buried than in surface leaf litters: implications for carbon and nutrient dynamics Plant and Soil, 396,175-187. DOI:10.1007/s11104-015-2595-1URL [本文引用: 2]
LiuJH, WangXQ, MaY, TanFZ (2016). Spatial variation of soil salinity on Tamarix ramosissima nebkhas and iterdune in oasis-desert ecotone Journal of Desert Research, 36(1),181-189. [本文引用: 1]
LiuQ (2014). The Environmental Change Revealed by the Material Composition of the Tamarix cone Sedimentary in the Southern Region of the Taklimakan Desert PhD dissertation, Hebei Normal University, Shijiazhuang. [本文引用: 1]
LiuSR, HuRG, CaiGC (2012). Effects of enhanced UV-B radiation on terrestrial ecosystem carbon cycle: a review Chinese Journal of Applied Ecology, 23,1992-1998. [本文引用: 1]
LuoY, HuSJ, WangXF, TianCY, YinCH (2012). A new method to determine soil soluble salt using electrical conductivity index Acta Pedologica Sinica, 49,1257-1261. [本文引用: 1]
MaY, WangXQ, ZhangB, LiuJH, HanZY, TangGL (2014). Effects of wind erosion and sand burial on water relations and photosynthesis in Alhagi sparsifolia in the southern edge of the Taklimakan Desert Chinese Journal of Plant Ecology, 38,491-498. DOI:10.3724/SP.J.1258.2014.00045URL [本文引用: 1]
MaoDL, LeiJQ, LiSY, LuiGJ, ZhengZH, XueJie (2015). Spatial differentiation of physico-chemical properties of surface sand materials in the oasis-desert ecotone in Cele, Xinjiang, China Journal of Desert Research, 35(1),136-144. [本文引用: 1]
Martínez-YrízarA, Nú?ezS, BúrquezA (2007). Leaf litter decomposition in a southern Sonoran Desert ecosystem, northwestern Mexico: effects of habitat and litter quality Acta Oecologica, 32,291-300. DOI:10.1016/j.actao.2007.05.010URL [本文引用: 1]
MooshammerM, WanekW, SchneckerJ, WildB, LeitnerS, HofhanslF, Bl?chlA, H?mmerleI, FrankAH, FuchsluegerL, KeiblingerKM, Zechmeister-BoltensternS, RichterA (2012). Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter Ecology, 93,770-782. PMID:22690628 [本文引用: 1] Resource stoichiometry (C:N:P) is an important determinant of litter decomposition. However, the effect of elemental stoichiometry on the gross rates of microbial N and P cycling processes during litter decomposition is unknown. In a mesocosm experiment, beech (Fagus sylvatica L.) litter with natural differences in elemental stoichiometry (C:N:P) was incubated under constant environmental conditions. After three and six months, we measured various aspects of nitrogen and phosphorus cycling. We found that gross protein depolymerization, N mineralization (ammonification), and nitrification rates were negatively related to litter C:N. Rates of P mineralization were negatively correlated with litter C:P. The negative correlations with litter C:N were stronger for inorganic N cycling processes than for gross protein depolymerization, indicating that the effect of resource stoichiometry on intracellular processes was stronger than on processes catalyzed by extracellular enzymes. Consistent with this, extracellular protein depolymerization was mainly limited by substrate availability and less so by the amount of protease. Strong positive correlations between the interconnected N and P pools and the respective production and consumption processes pointed to feed-forward control of microbial litter N and P cycling. A negative relationship between litter C:N and phosphatase activity (and between litter C:P and protease activity) demonstrated that microbes tended to allocate carbon and nutrients in ample supply into the production of extracellular enzymes to mine for the nutrient that is more limiting. Overall, the study demonstrated a strong effect of litter stoichiometry (C:N:P) on gross processes of microbial N and P cycling in decomposing litter; mineralization of N and P were tightly coupled to assist in maintaining cellular homeostasis of litter microbial communities.
OlsonJS (1963). Energy storage and the balance of producers and decomposition in ecological systems Ecology, 44,322-331. DOI:10.2307/1932179URL [本文引用: 1]
PauliF (1964). Soil fertility problem in arid and semi-arid lands Nature, 204,1286-1288. DOI:10.1038/2041286a0URL
PuchetaE, LlanosM, MeglioliC, GaviornoM, RuizM, PareraC (2006). Litter decomposition in a sandy Monte desert of western Argentina: influences of vegetation patches and summer rainfall Austral Ecology, 31,808-816. DOI:10.1111/aec.2006.31.issue-7URL [本文引用: 1]
RaichJW, SchlesingerWH (1992). The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate Tellus B: Chemical and Physical Meteorology, 44(2),81-99. DOI:10.3402/tellusb.v44i2.15428URL [本文引用: 1]
RathKM, RouskJ (2015). Salt effects on the soil microbial decomposer community and their role in organic carbon cycling: a review Soil Biology & Biochemistry, 81,108-123. DOI:10.1016/j.soilbio.2014.11.001URL [本文引用: 1]
RubinoM, DungaitJAJ, EvershedRP, BertoliniT, deAngelis P, D'OnofrioA, LagomarsinoA, LubrittoC, MerolaA, TerrasiF, CotrufoMF (2010). Carbon input belowground is the major C flux contributing to leaf litter mass loss: evidences from a13C labelled-leaf litter experiment Soil Biology & Biochemistry, 42,1009-1016. DOI:10.1016/j.soilbio.2010.02.018URL [本文引用: 1]
SchreegLA, MackMC, TurnerBL (2013). Nutrient-specific solubility patterns of leaf litter across 41 lowland tropical woody species Ecology, 94,94-105. PMID:23600244 [本文引用: 1] Leaching is a mechanism for the release of nutrients from litter or senesced leaves that can drive interactions among plants, microbes, and soil. Although leaching is well established in conceptual models of litter decomposition, potential nutrient solubility of mineral elements from recently senesced litter has seldom been quantified. Using a standardized extraction (1:50 litter-to-water ratio and four-hour extraction) and recently senesced leaf litter of 41 tropical tree and liana species, we investigated how solubility varies among elements, and whether the solubility of elements could be predicted by litter traits (e.g., lignin, total element concentrations). In addition, we investigated nutrient forms (i.e., inorganic and organic) and ratios in leachate. Water-soluble elements per unit litter mass were strongly predicted by total initial litter element concentrations for potassium (K; r2 = 0.79), sodium (Na; r2 = 0.51) and phosphorus (P; r2 = 0.66), while a significant but weaker positive relationship was found for nitrogen (N; r2 = 0.36). There was no significant relationship for carbon (C) or calcium (Ca). Element-specific solubility varied markedly. On average 100% of total K, 35% of total P, 28% of total Na, 5% of total N, 4% of total Ca, and 3% of total C were soluble. For soluble P, 90% was inorganic orthophosphate. The high solubility of K, Na, and P as inorganic orthophosphate suggests that these nutrients can become rapidly available to litter microbes with no metabolic cost. Few common predictors of decomposition rates were correlated with element solubility, although soluble C (milligrams per gram of litter) was negatively related to lignin content (r2 = 0.19; P < 0.004). Solubility of elements was linked within a species: when a species ranked high in the soluble fraction of one element, it also ranked high in the solubility of other elements. Overall nutrient-specific patterns of solubility from recently senesced litter emphasize that litter elements cannot be treated equally in our conceptual and empirical models of decomposition. The relatively high potential solubility of P as orthophosphate from fresh litter advances our understanding of ecological stoichiometric ratios and nutrient bioavailability in tropical forests.
TaylorBR, ParkinsonD, ParsonsWFJ (1989). Nitrogen and lignin content as predictors of litter decay rates: a microcosm test Ecology, 70,97-104. DOI:10.2307/1938416URL [本文引用: 1]
ThroopHL, ArcherSR (2007). Interrelationships among shrub encroachment, land management, and litter decomposition in a semidesert grassland Ecological Applications, 17,1809-1823. DOI:10.1890/06-0889.1URL [本文引用: 1]
ThroopHL, ArcherSR (2009). Resolving the dryland decomposition conundrum: some new perspectives on potential drivers //Lüttge U, Beyschlag W, Büdel B, Francis D. Progress in Botany. Springer-Verlag, Berlin, Heidelberg.171-194. [本文引用: 2]
VivancoL, AustinAT (2006). Intrinsic effects of species on leaf litter and root decomposition: a comparison of temperate grasses from North and South America Oecologia, 150,97-107. PMID:16917779 [本文引用: 1] Plant species affect natural ecosystems through interactions between environmental and genetic factors. The importance of plant species in controlling decomposition is now well-established through its influence on litter quality, which affects mass loss and nutrient release. At the same time, direct species effects are often confounded with indirect site effects due to the ecophysiological responses of plants to environmental variability. We evaluated the intrinsic effects of species on litter quality and decomposition, comparing 14 native perennial grass species from three different grassland ecosystems in North and South America. Plants were grown under controlled greenhouse conditions to eliminate any indirect effects of climate on litter quality, and senescent material of leaf litter and roots were collected. The initial litter nutrient quality and the carbon quality were assessed, and decomposition was determined over a period of one year by placing litterbags in a common grassland site. In spite of constant growth conditions, species' litter showed broad and significant differences in N, P and lignin concentration, as well as C:N ratio, with the greatest differences occurring between C(3) and C(4) species and leaf litter and root material. In addition, decomposition was significantly different among species and between leaf litter and roots within species, with constants (k) ranging from 1.50 to 3.65 year(-1) for leaf litter, and 0.51-1.82 year(-1) for roots. These results highlight the fact that, independent of climate or edaphic changes due to human activity, changes in plant species or in allocation patterns among plant organs in grassland ecosystems could have a large effect on carbon turnover. At the same time, the way in which intrinsic species characteristics affect decomposition demonstrates a large degree of functional convergence among species from grasslands of North and South America.
XiaXC, CaoQY, WangFB, LeiJQ, ZhaoYJ (2005). Significance of studying age layers of Tamarix ramosissima sand-hillock in Lop Nur region, Xinjiang Arid Land Geography, 28,565-568. [本文引用: 1]
XuJ, ChenYJ, LiuJZ (2020). Research progress of the effects of halophyte shrubs on spatial distribution of soil nutrients and salts and their mechanisms Journal of Anhui Agricultural Sciences, 48(1),19-23. [本文引用: 1]
YinCH, FengG, TianCY, BaiDS, ZhangFS (2007). Influence of tamarisk shrub on the distribution of soil salinity and moisture on the edge of Taklamakan desert China Environmental Science, 27,670-675. [本文引用: 1]
ZhaoYJ, CheGH, LiuH, ZengJ, XiaXC (2016). C and N content in organic matter of Tamarix cone and climatic and environmental change in southern region of Taklimakan desert Arid Land Geography, 39,461-467. [本文引用: 1]
Decomposition of 51 semidesert species from wide-ranging phylogeny is faster in standing and sand-buried than in surface leaf litters: implications for carbon and nutrient dynamics 2 2015
 ,
, ,1,3,*, Henry J. SUN5, 林丽莎1,2,3, 刘波6
,1,3,*, Henry J. SUN5, 林丽莎1,2,3, 刘波6

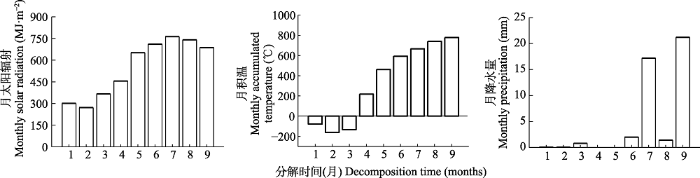 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT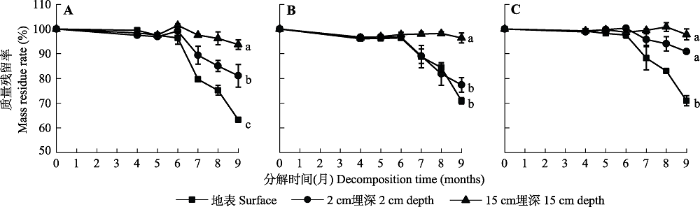 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT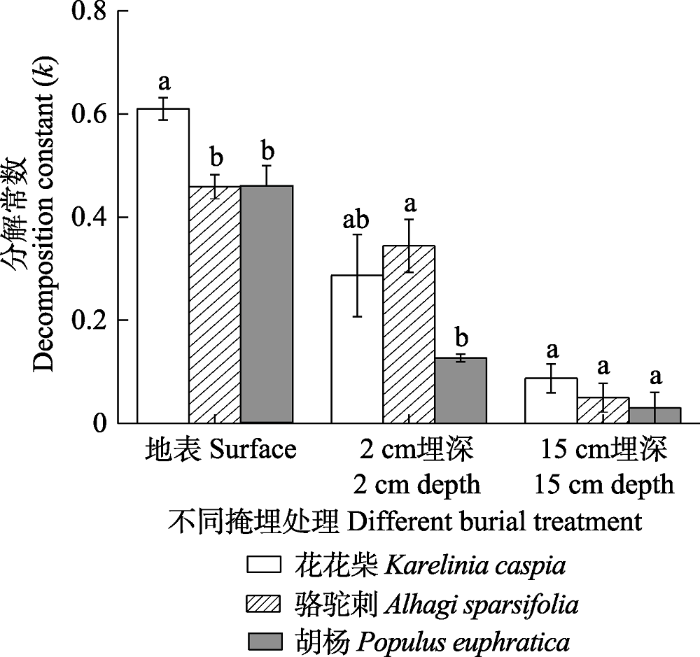 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT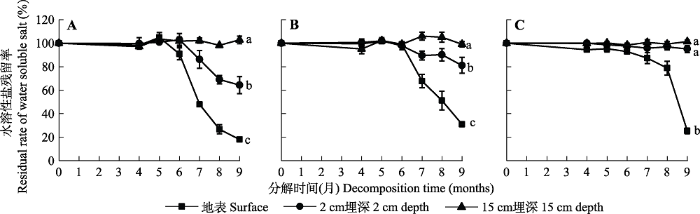 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT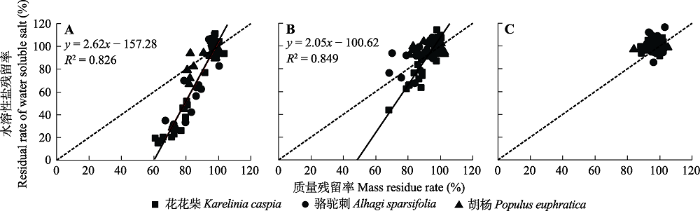 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT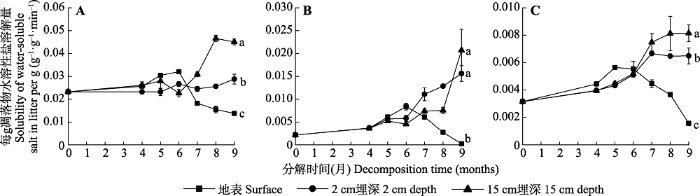 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT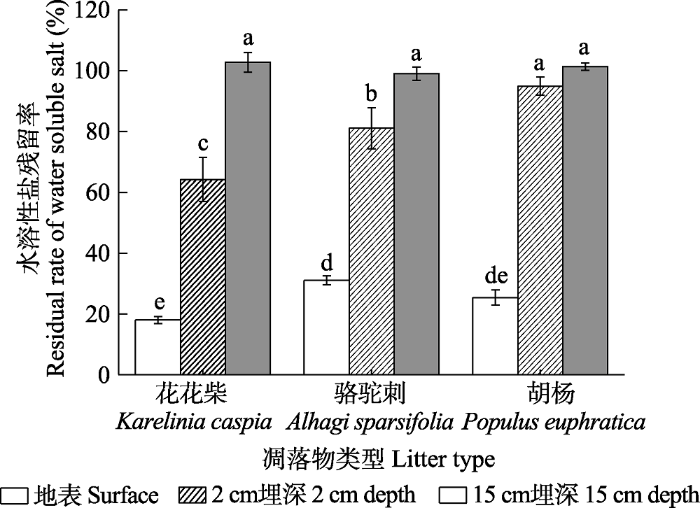 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}