Traits of shrubs in forests and bushes reveal different life strategies
Jia-Yu CAO1, Jian-Feng LIU1, Quan YUAN1, De-Yu XU1, Hai-Dong FAN1, Hai-Yan CHEN1, Bin TAN1, Li-Bin LIU1,2, Duo YE1,2, Jian NI,,1,2,*1College of Chemistry and Life Sciences, Zhejiang Normal University, Jinhua, Zhejiang 321004, China 2Jinhua Mountain Observation and Research Station for Subtropical Forest Ecosystems, Jinhua, Zhejiang 321004, China
Abstract Aims Shrubs play important roles in both forest and bushland ecosystems. This study aims to identify the adaptative strategies of shrubs in different habitats by analyzing the differences in functional traits of shrubs grown in understory of forest communities and in two bushlands. Methods Nine functional traits for leaves and twigs were measured on samples collected from the dominant shrub species in 24 plots distributed in three contrasting habitats: forest understory, low mountain bushes, and bushes on the mountaintop, in Beishan Mountain of Jinhua, Zhejiang Province. The overall differences among habitats, inter- and intra-specific variations, and differences between life forms in the functional traits of shrubs were tested by statistical analysis. Important findings The nine plant traits differed for shrubs grown in the three habitats. The shrubs in forest understory had higher leaf area (LA) and specific leaf area (SLA), lower leaf dry-matter content (LDMC), leaf tissue density (LTD) and twig tissue density (TTD), while those in low mountain bushes had greater leaf thickness (LT) and LTD, smaller SLA and twig dry-matter content (TDMC), compared with shrubs from bushes on the mountaintop. The inter- and intra-specific variation coefficients of SLA, twig diameter (TD), TTD, and TDMC were greatest in shrubs of the forest understory, whereas the inter- and intra-specific variation coefficients of SLA, LDMC, TDMC, and TTD were smallest in ??shrubs of low mountain bushes. Among different life forms, the understory evergreen shrubs had significantly higher LT, LTD, and LDMC, and lower SLA, than that of deciduous shrubs. The differences in LT and SLA between evergreen and deciduous shrubs of the mountaintop bushes were the same as the understory shrubs, but the differences in LTD and LDMC were reversed. Species and its interaction with habitat are the major factors affecting the shrub traits. In short, compared to the shrubs from bushes, the understory shrubs in forest communities form a series of trait combinations with greater LA and SLA, and smaller LTD, TTD and LDMC for faster growth in order to adapt to the understory environment with less light and stronger competition; this is a quick investment-return (resource acquisitive) strategy. Shrubs from low mountain bushes and the mountaintop bushes are associated with a series trait combinations with greater LT, LTD, LDMC and TTD, and smaller LA, SLA for storing more nutrients and growing slower; this is a slow investment-return (resource conservative) strategy. Different combinations of shrub functional traits and their various life strategies can provide guidance to the ecological restoration of degraded vegetation in the subtropical region of China. Keywords��shrub;plant functional traits;subtropical forests;shrubland;habitat;life history strategy
PDF (2077KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر��� ���ñ��� �ܼ��, ������, ԬȪ, �����, ������, �º���, ̷��, ������, Ҷ��, �߽�. ɭ�����ԵĹ�ľ��״��ʾ��ͬ���������. ֲ����̬ѧ��, 2020, 44(7): 715-729. DOI: 10.17521/cjpe.2020.0024 CAO Jia-Yu, LIU Jian-Feng, YUAN Quan, XU De-Yu, FAN Hai-Dong, CHEN Hai-Yan, TAN Bin, LIU Li-Bin, YE Duo, NI Jian. Traits of shrubs in forests and bushes reveal different life strategies. Chinese Journal of Plant Ecology, 2020, 44(7): 715-729. DOI: 10.17521/cjpe.2020.0024
��2017��2018���ļ�, �ڽ�ɽ����ѡ����Ҷ�֡������콻�֡�������Ҷ��Ҷ�콻�֡�������Ҷ�֡���ɽ��Ժ�ɽ�����6��ֲ������, ������24���̶�����(��1), ����ɭ����������ֱ�Ϊ30 m �� 30 m, 12����Ҷ�ֺ������콻�־�Ϊ������, 9����Ҷ�־�Ϊ�˹���; 1����ɽ����������Ϊ30 m �� 10 m, ����Ҷ�ֿ�����Ĵ�����ֲ��; 2��ɽ�������������ֱ�Ϊ10 m �� 20 m, Ϊɽ����������Խ�С�Ĵ�����ֲ����������ɭ�ֺ���������ؾ�(DBH) �� 1 cm, �Լ����������DBH < 1 cm, �߶ȡ�10 cm��ľ��ֲ������Ʊ��, ����¼��Ӧ���������ؾ�(��ľΪ����)�����ߡ��ڷ��Լ���������ָ�ꡣ������Զ�ȡ����Ƶ�Ⱥ�������ƶȼ����ľ����Ҫֵ(��������, 2019)��
Table 1 ��1 ��1��ɽ24�����ػ� �����������ƹ�ľ Table 1Site characteristics and dominant shrubs for the 24 plots in Beishan Mountain of Jinhua, Zhejiang Province
AckerlyDD, CornwellWK (2007). A trait-based approach to community assembly: partitioning of species trait values into within- and among-community components Ecology Letters, 10, 135-145. DOI:10.1111/j.1461-0248.2006.01006.xURLPMID:17257101 Plant functional traits vary both along environmental gradients and among species occupying similar conditions, creating a challenge for the synthesis of functional and community ecology. We present a trait-based approach that provides an additive decomposition of species' trait values into alpha and beta components: beta values refer to a species' position along a gradient defined by community-level mean trait values; alpha values are the difference between a species' trait values and the mean of co-occurring taxa. In woody plant communities of coastal California, beta trait values for specific leaf area, leaf size, wood density and maximum height all covary strongly, reflecting species distributions across a gradient of soil moisture availability. Alpha values, on the other hand, are generally not significantly correlated, suggesting several independent axes of differentiation within communities. This trait-based framework provides a novel approach to integrate functional ecology and gradient analysis with community ecology and coexistence theory.
AugerS, ShipleyB (2013). Inter-specific and intra-specific trait variation along short environmental gradients in an old-growth temperate forest Journal of Vegetation Science, 24, 419-428. [��������: 1]
ChaveJ, CoomesD, JansenS, LewisSL, SwensonNG, ZanneAE (2009). Towards a worldwide wood economics spectrum Ecology Letters, 12, 351-366. DOI:10.1111/j.1461-0248.2009.01285.xURLPMID:19243406 [��������: 3] Wood performs several essential functions in plants, including mechanically supporting aboveground tissue, storing water and other resources, and transporting sap. Woody tissues are likely to face physiological, structural and defensive trade-offs. How a plant optimizes among these competing functions can have major ecological implications, which have been under-appreciated by ecologists compared to the focus they have given to leaf function. To draw together our current understanding of wood function, we identify and collate data on the major wood functional traits, including the largest wood density database to date (8412 taxa), mechanical strength measures and anatomical features, as well as clade-specific features such as secondary chemistry. We then show how wood traits are related to one another, highlighting functional trade-offs, and to ecological and demographic plant features (growth form, growth rate, latitude, ecological setting). We suggest that, similar to the manifold that tree species leaf traits cluster around the 'leaf economics spectrum', a similar 'wood economics spectrum' may be defined. We then discuss the biogeography, evolution and biogeochemistry of the spectrum, and conclude by pointing out the major gaps in our current knowledge of wood functional traits.
ChenW, WangJH, MaRJ, QiW, LiuK, ZhangLN, ChenXL (2016). Variance in leaf functional traits of 89 species from the eastern Guangdong of China Chinese Journal of Ecology, 35, 2101-2109. [��������: 1]
CornelissenJHC, LavorelS, GarnierE, D��azS, BuchmannN, GurvichDE, ReichPB, terSteege H, MorganHD, vander Heijden MGA, PausasJG, PoorterH (2003). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide Australian Journal of Botany, 51, 335-380. [��������: 3]
dela Riva EG, TostoA, P��rez-RamosIM, Navarro-Fern��ndezCM, OlmoM, AntenNPR, Mara?��nT, VillarR (2016). A plant economics spectrum in Mediterranean forests along environmental gradients: Is there coordination among leaf, stem and root traits? Journal of Vegetation Science, 27, 187-199. [��������: 1]
deSmedt P, OttavianiG, Wardell-JohnsonG, SykoraKV, MucinaL (2018). Habitat heterogeneity promotes intraspecific trait variability of shrub species in Australian granite inselbergs Folia Geobotanica, 53, 133-145. [��������: 3]
D��azS, KattgeJ, CornelissenJHC, WrightIJ, LavorelS, DrayS, ReuB, KleyerM, WirthC, ColinPrentice I, GarnierE, B?nischG, WestobyM, PoorterH, ReichPB, MolesAT, DickieJ, GillisonAN, ZanneAE, ChaveJ, JosephWright S, Sheremet��evSN, JactelH, BaralotoC, CeraboliniB, PierceS, ShipleyB, KirkupD, CasanovesF, JoswigJS, G��ntherA, FalczukV, R��gerN, MahechaMD, Gorn��LD (2016). The global spectrum of plant form and function Nature, 529, 167-171. URLPMID:26700811 [��������: 2]
DlugosDM, CollinsH, BartelmeEM, DrenovskyRE (2015). The non-native plant Rosa multiflora expresses shade avoidance traits under low light availability American Journal of Botany, 102, 1323-1331. URLPMID:26290555 [��������: 1]
FanHD, ChenHY, WuYN, LiuJF, XuDY, CaoJY, YuanQ, TanB, LiuXT, XuJ, WangGM, HanWJ, LiuLB, NiJ (2019). Community characteristics of main vegetation types on the southern slope of Beishan Mountain in Jinhua, Zhejiang, China Chinese Journal of Plant Ecology, 43, 921-928. DOI:10.17521/cjpe.2019.0114URL [��������: 1] The Beishan Mountain of Jinhua is located in the northern part of the mid-subtropical region. In vegetation division, this region belongs to the northern sub-zone of mid-subtropical evergreen broad-leaved forest vegetation belt. The vegetation in this mountain is a secondary forest restored from a severe deforestation. It is currently in a rapid process of positive succession. Thus, this region is suitable for studying the dynamics and succession mechanisms of plant communities and the restoration of degraded ecosystems. To further understand the community characteristics of the main vegetation types in Beishan Mountain, we surveyed the main plant communities on its southern slope with a fixed plot method (the plot area is 30 m × 30 m for forests and 20 m × 10 m, 30 m × 10 m for shrubs) based on the internationally accepted forest survey approach used in the survey of dynamic forest plots. The species composition, quantitative characteristics and habitat information of all plant communities were investigated and recorded. The importance values of trees and shrubs were calculated. The community types and their characteristics were analyzed. The spatial distribution maps of the major trees and shrubs in each plot were also drawn. In the present study, detailed community data for 24 sampled plots representing 11 formations were presented (consisting of 21 forest plots and 3 shrub plots). [ ������, �º���, ������, ������, �����, �ܼ��, ԬȪ, ̷��, ����ͮ, ���, ������, ���ľ�, ������, �߽� (2019). ��ɽ������Ҫֲ�����͵�Ⱥ������ ֲ����̬ѧ��, 43, 921-928.] [��������: 1]
FortF, JouanyC, CruzP (2013). Root and leaf functional trait relations in Poaceae species: implications of differing resource-acquisition strategies Journal of Plant Ecology, 6, 211-219. DOI:10.1093/jpe/rts034URL [��������: 1] Aims Root systems play an essential role in grassland functioning in both acquisition and storage of resources. Nevertheless, root functional traits have not received as much attention as those measured on above-ground organs, and little is known about their relations. Our objectives were to test whether morphological and root system traits allowed identification of grass species' functional strategies and to determine whether a relation exists between above- and below-ground traits.Methods Functional traits of root tissues (specific root length, diameter, tissue density and nitrogen concentration), whole root systems (root mass, root length density, root mass percentage below a depth of 20cm and fine root %) and two major leaf traits (specific leaf area and leaf dry matter content) were determined under field conditions and their relations were analysed in eleven perennial temperate Poaceae species.Important findings Canonical correspondence analysis along Axis 1 revealed a gradient of species, from those with deep, dense and coarse root systems with a large root mass to those with shallow root systems, thin roots and high specific root length; this suggests strong correlations among root traits. Correlations between specific root length and specific leaf area reveal two groups of species, which probably indicates different drought-tolerance capacities. Root trait syndromes enable ranking grasses along a gradient from conservative-strategy species (from stressful habitats), which display a deep and coarse root system, to acquisitive species (from rich and moist meadows), which display a shallow and thin root system. Although both types display similar above-ground strategies, drought-tolerant species have lower specific root lengths than drought-sensitive species, revealing more conservative root strategies.]]>
FunkJL, CornwellWK (2013). Leaf traits within communities: context may affect the mapping of traits to function Ecology, 94, 1893-1897. URLPMID:24279259 [��������: 1]
GaoJ, XuB, WangJN, ZhouHY, WangYX, WuY (2015). Correlations among leaf traits of typical shrubs and their responses to different light environments in shrub-grassland of southern China Chinese Journal of Ecology, 34, 2424-2431. URL [��������: 1] i.e.�� ��Salix etosia, Rubus setchuenensis ��and�� Hydrangea aspera) in shrub��grassland of southern China. Field sampling survey was carried out to collect leaves under different light environments (inside and outside the forests) in Yunyang County. The results showed that the correlations among leaf traits of three shrubs vary one to another, ��i.e��. species��specific. There was no significant difference in leaf traits of ��S. etosia�� under different light environments, while significant differences in leaf traits were found for ��R. setchuenensi�� and ��H. aspera��. Both SLA and LNC of ��R. setchuenensi ��and ��H. aspera ��increased inside the forests for adapting to lower illuminance. In addition, LCC of�� H. aspera�� increased significantly inside the forests, which indicated a relatively strong carbon fixation capacity. Leaf traits of ��R. setchuenensis�� and ��H. aspera ��were mainly affected by soil temperature under different light environments. Our study demonstrated that there existed the phenotypic plasticity for plant leaf traits, and different plants had species��specific responses as well as strategies to adapt to different environments.]]> [ �߾�, �첨, ����ţ, �ܺ���, ������, ���� (2015). �Ϸ���ݴԵ���ľ��ͬҶƬ��״������Լ���Բ�ͬ�������Ӧ ��̬ѧ��־, 34, 2424-2431.] [��������: 1]
GaoSH, GeYX, ZhouLY, ZhuBL, GeXY, LiK, NiJ (2018). What is the optimal number of leaves when measuring leaf area of tree species in a forest community? Chinese Journal of Plant Ecology, 42, 917-925. DOI:10.17521/cjpe.2018.0087URL [��������: 1] Aims Leaf size, as one of the easier measured plant morphological traits, reflects response and adaptation of plants to environment and indicates functions and processes of ecosystem. When measuring leaf size (the leaf area, LA) on the field, the common accepted practice considers that the number of leaves picked off is often 10-20. However, what is the optimal number of leaves remains unknown. In this study, we attempt to determine how many leaves should be investigated when the leaf size of a tree is measured. Methods This study selected two dominant tree species (Schima superba, Ss and Liquidambar formosana, Lf) from a broadleaved evergreen and deciduous mixed forest in Jinhua Mountain of Zhejiang Province, eastern China. On the basis of sampling (>2 500 leaves for each tree) in five classes of the diameter of breast height (DBH) of tree species and at six directions for each individual, variations of LA in the two tree species are statistically analyzed. The optimal number of leaves, which can mostly represent the common leaf size feature, is further investigated. Important findings Mean LAof the evergreen tree Ss was smaller than that of the deciduous tree Lf. The former was (41.60 ± 10.88) cm 2 (16.74-100.80 cm 2) and the latter was (57.65 ± 19.35) cm 2 (11.31-129.51 cm 2). LA of Lf was significantly related to the DBH, but LAof Ss was not. LA of both trees in the middle DBH class (15-20 cm) was not significantly different from their means. LA of two trees have no significant correlations with the sampling directions, but LA at the east, west and bottom had no significant relationships with their means. Considering the representativeness and practicality in the field sampling, the priority of selecting leaves can target the bottom direction of middle diameter mature trees. Random sampling analysis indicated that, the optimal number of leaves for tree LA measurement is species specific. The optimal number of leaves for Ss is 40 and for Lf is at least 170, respectively. Therefore, when measuring leaf area in a forest community, the optimal sampling number of leaves should not be limited to 10-20 leaves. Under sufficient labor, material and time, more leaves should be measured. ]]> [ ��˼��, ����ϣ, ������, �챦��, ������, �, �߽� (2018). �ⶨɭ����ľҶ���������ҶƬ���Ƕ���? ֲ����̬ѧ��, 42, 917-925.] [��������: 1]
GarnierE, VileD, RoumetC, LavorelS, GrigulisK, NavasM-L, LloretF (2019). Inter- and intra-specific trait shifts among sites differing in drought conditions at the north western edge of the Mediterranean Region Flora, 254, 147-160. [��������: 1]
GrataniL, BombelliA (2001). Differences in leaf traits among Mediterranean broad-leaved evergreen shrubs Annales Botanici Fennici, 38, 15-24. [��������: 2]
GuoSL, LiuP, ChenG, LuX (1993). Observations on the vegetation of Mount Bei of Jinhua in Zhejiang Province Journal of Zhejiang Normal University (Nature Sciences), 16, 59-67. [��������: 1]
HeberlingJM, FridleyJD (2013). Resource-use strategies of native and invasive plants in Eastern North American forests New Phytologist, 200, 523-533. URLPMID:23815090 [��������: 1]
JungV, ViolleC, MondyC, HoffmannL, MullerS (2010). Intraspecific variability and trait-based community assembly Journal of Ecology, 98, 1134-1140. [��������: 1]
KattgeJ, B?nischG, D��azS, LavorelS, PrenticeIC, WirthC (2020). TRY plant trait database-enhanced coverage and open access Global Change Biology, 26, 119-188. DOI:10.1111/gcb.14904URLPMID:31891233 [��������: 2] Plant traits-the morphological, anatomical, physiological, biochemical and phenological characteristics of plants-determine how plants respond to environmental factors, affect other trophic levels, and influence ecosystem properties and their benefits and detriments to people. Plant trait data thus represent the basis for a vast area of research spanning from evolutionary biology, community and functional ecology, to biodiversity conservation, ecosystem and landscape management, restoration, biogeography and earth system modelling. Since its foundation in 2007, the TRY database of plant traits has grown continuously. It now provides unprecedented data coverage under an open access data policy and is the main plant trait database used by the research community worldwide. Increasingly, the TRY database also supports new frontiers of trait-based plant research, including the identification of data gaps and the subsequent mobilization or measurement of new data. To support this development, in this article we evaluate the extent of the trait data compiled in TRY and analyse emerging patterns of data coverage and representativeness. Best species coverage is achieved for categorical traits-almost complete coverage for 'plant growth form'. However, most traits relevant for ecology and vegetation modelling are characterized by continuous intraspecific variation and trait-environmental relationships. These traits have to be measured on individual plants in their respective environment. Despite unprecedented data coverage, we observe a humbling lack of completeness and representativeness of these continuous traits in many aspects. We, therefore, conclude that reducing data gaps and biases in the TRY database remains a key challenge and requires a coordinated approach to data mobilization and trait measurements. This can only be achieved in collaboration with other initiatives.
KumordziBB, AubinI, CardouF, ShipleyB, ViolleC, JohnstoneJ, AnandM, ArsenaultA, BellFW, BergeronY, BoulangeatI, BrousseauM, DeGrandpr�� L, DelagrangeS, FentonNJ, GravelD, EllenMacdonald S, HamelB, HigelinM, H��bertF, IsabelN, MallikA, MclntoshACS, McLarenJR, MessierC, MorrisD, ThiffaultN, TremblayJ-P, MunsonAD (2019). Geographic scale and disturbance influence intraspecific trait variability in leaves and roots of North American understorey plants Functional Ecology, 33, 1771-1784. [��������: 1]
Lebrija-TrejosE, P��rez-Garc��aEA, MeaveJA, BongersF, PoorterL (2010). Functional traits and environmental filtering drive community assembly in a species-rich tropical system Ecology, 91, 386-398. URLPMID:20392004 [��������: 1]
LiGY, YangDM, SunSC (2008). Allometric relationships between lamina area, lamina mass and petiole mass of 93 temperate woody species vary with leaf habit, leaf form and altitude Functional Ecology, 22, 557-564. DOI:10.1111/fec.2008.22.issue-4URL [��������: 1]
LiL, LiXY, XuXW, LinLS, ZengFJ, ChenFL (2014). Assimilative branches and leaves of the desert plant Alhagi sparsifolia Shap. possesses a different adaptation mechanism to shade Plant Physiology and Biochemistry, 74, 239-245. URLPMID:24316582 [��������: 1]
LiuXJ, MaKP (2015). Plant functional traits��Concepts, applications and future directions Scientia Sinica Vitae, 45, 325-339. [��������: 3]
LuoYK, HuHF, ZhaoMY, LiH, LiuSS, FangJY (2019). Latitudinal pattern and the driving factors of leaf functional traits in 185 shrub species across eastern China Journal of Plant Ecology, 12, 67-77. [��������: 4]
MaXL (2014). A Study on Twig and Leaf Traits of the Shrub Species in Secondary Forest Succession Process Master degree dissertation, Northwest Normal University, Lanzhou. [��������: 1]
MaZ, GuoD, XuX, LuM, BardgettRD, EissenstatDM, LukeMcCormack M, HedinLO (2018). Evolutionary history resolves global organization of root functional traits Nature, 555, 94-97. DOI:10.1038/nature25783URLPMID:29466331 [��������: 1] Plant roots have greatly diversified in form and function since the emergence of the first land plants, but the global organization of functional traits in roots remains poorly understood. Here we analyse a global dataset of 10 functionally important root traits in metabolically active first-order roots, collected from 369 species distributed across the natural plant communities of 7 biomes. Our results identify a high degree of organization of root traits across species and biomes, and reveal a pattern that differs from expectations based on previous studies of leaf traits. Root diameter exerts the strongest influence on root trait variation across plant species, growth forms and biomes. Our analysis suggests that plants have evolved thinner roots since they first emerged in land ecosystems, which has enabled them to markedly improve their efficiency of soil exploration per unit of carbon invested and to reduce their dependence on symbiotic mycorrhizal fungi. We also found that diversity in root morphological traits is greatest in the tropics, where plant diversity is highest and many ancestral phylogenetic groups are preserved. Diversity in root morphology declines sharply across the sequence of tropical, temperate and desert biomes, presumably owing to changes in resource supply caused by seasonally inhospitable abiotic conditions. Our results suggest that root traits have evolved along a spectrum bounded by two contrasting strategies of root life: an ancestral 'conservative' strategy in which plants with thick roots depend on symbiosis with mycorrhizal fungi for soil resources and a more-derived 'opportunistic' strategy in which thin roots enable plants to more efficiently leverage photosynthetic carbon for soil exploration. These findings imply that innovations of belowground traits have had an important role in preparing plants to colonize new habitats, and in generating biodiversity within and across biomes.
McDonaldPG, FonsecaCR, OvertonJMcC, WestobyM (2003). Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades? Functional Ecology, 17, 50-57. [��������: 1]
PausasJG, BradstockRA (2007). Fire persistence traits of plants along a productivity and disturbance gradient in mediterranean shrublands of south-east Australia Global Ecology and Biogeography, 16, 330-340. [��������: 1]
RolhauserAG, PuchetaE (2016). Annual plant functional traits explain shrub facilitation in a desert community Journal of Vegetation Science, 27, 60-68. [��������: 3]
RosellJA, GleasonS, M��ndez-AlonzoR, ChangY, WestobyM (2014). Bark functional ecology: evidence for tradeoffs, functional coordination, and environment producing bark diversity New Phytologist, 201, 486-497. URLPMID:24117609 [��������: 1]
ScheepensJF, FreiES, St?cklinJ (2010). Genotypic and environmental variation in specific leaf area in a widespread Alpine plant after transplantation to different altitudes Oecologia, 164, 141-150. URLPMID:20461412 [��������: 1]
ShahS, ShresthaKK, ScheideggerC (2019). Variation in plant functional traits along altitudinal gradient and land use types in Sagarmatha National Park and buffer zone, Nepal American Journal of Plant Sciences, 10, 595-614. [��������: 1]
SterckFJ, vanGelder HA, PoorterL (2006). Mechanical branch constraints contribute to life-history variation across tree species in a Bolivian forest Journal of Ecology, 94, 1192-1200. [��������: 1]
TangQQ, HuangYT, DingY, ZangRG (2016). Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broad-leaved mixed forests Biodiversity Science, 24, 262-270. [��������: 1]
TianD, YanZB, NiklasKJ, HanWX, KattgeJ, ReichPB, LuoYK, ChenYH, TangZY, HuHF, WrightIJ, SchmidB, FangJY (2018). Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent National Science Review, 5, 728-739. DOI:10.1093/nsr/nwx142URL [��������: 1]
VannesteT, Vald��sA, VerheyenK, PerringMP, Bernhardt-R?mermannM, AndrieuE, BrunetJ, CousinsSAO, DeconchatM, deSmedt P, DiekmannM, EhrmannS, HeinkenT, HermyM, KolbA, LenoirJ, LiiraJ, NaafT, PaalT, WulfM, DecocqG, deFrenne P (2019). Functional trait variation of forest understorey plant communities across Europe Basic and Applied Ecology, 34, 02356900. DOI: 10.1016/j.baae.2018.09.004. [��������: 1]
ViolleC, NavasML, VileD, KazakouE, FortunelC, HummelI, GarnierE (2007). Let the concept of trait be functional! Oikos, 116, 882-892. [��������: 1]
WangCY, XiaoHG, LiuJ, ZhouJW (2017a). Differences in leaf functional traits between red and green leaves of two evergreen shrubs Photinia �� fraseri and Osmanthus fragrans Journal of Forestry Research, 28, 473-479. [��������: 1]
WangM, WanPC, GuoJC, XuJS, ChaiYF, YueM (2017b). Relationships among leaf, stem and root traits of the dominant shrubs from four vegetation zones in Shaanxi Province, China Israel Journal of Ecology & Evolution, 63, 25-32. [��������: 3]
WangY, WangJ, LaiL, JiangL, ZhuangP, ZhangL, ZhengY, BaskinJM, BaskinCC (2014). Geographic variation in seed traits within and among forty-two species of Rhododendron(Ericaceae) on the Tibetan Plateau: relationships with altitude, habitat, plant height, and phylogeny Ecology and Evolution, 4, 1913-1923. DOI:10.1002/ece3.1067URLPMID:24963385 [��������: 1] Seed mass and morphology are plant life history traits that influence seed dispersal ability, seeding establishment success, and population distribution pattern. Southeastern Tibet is a diversity center for Rhododendron species, which are distributed from a few hundred meters to 5500 m above sea level. We examined intra- and interspecific variation in seed mass and morphology in relation to altitude, habitat, plant height, and phylogeny. Seed mass decreased significantly with the increasing altitude and increased significantly with increasing plant height among populations of the same species. Seed mass differed significantly among species and subsections, but not among sections and subgenera. Seed length, width, surface area, and wing length were significantly negative correlated with altitude and significantly positive correlated with plant height. Further, these traits differed significantly among habitats and varied among species and subsection, but not among sections and subgenera. Species at low elevation had larger seeds with larger wings, and seeds became smaller and the wings of seeds tended to be smaller with the increasing altitude. Morphology of the seed varied from flat round to long cylindrical with increasing altitude. We suggest that seed mass and morphology have evolved as a result of both long-term adaptation and constraints of the taxonomic group over their long evolutionary history.
WestobyM, ReichPB, WrightIJ (2013). Understanding ecological variation across species: area-based vs mass-based expression of leaf traits New Phytologist, 199, 322-323. URLPMID:23692294 [��������: 1]
WrightIJ, AckerlyDD, BongersF, HarmsKE, Ibarra-ManriquezG, Martinez-RamosM, MazerSJ, Muller-LandauHC, PazH, PitmanNCA, PoorterL, SilmanMR, VriesendorpCF, WebbCO, WestobyM, WrightSJ (2007). Relationships among ecologically important dimensions of plant trait variation in seven Neotropical forests Annals of Botany, 99, 1003-1015. DOI:10.1093/aob/mcl066URLPMID:16595553 [��������: 1] BACKGROUND AND AIMS: When ecologically important plant traits are correlated they may be said to constitute an ecological 'strategy' dimension. Through identifying these dimensions and understanding their inter-relationships we gain insight into why particular trait combinations are favoured over others and into the implications of trait differences among species. Here we investigated relationships among several traits, and thus the strategy dimensions they represented, across 2134 woody species from seven Neotropical forests. METHODS: Six traits were studied: specific leaf area (SLA), the average size of leaves, seed and fruit, typical maximum plant height, and wood density (WD). Trait relationships were quantified across species at each individual forest as well as across the dataset as a whole. 'Phylogenetic' analyses were used to test for correlations among evolutionary trait-divergences and to ascertain whether interspecific relationships were biased by strong taxonomic patterning in the traits. KEY RESULTS: The interspecific and phylogenetic analyses yielded congruent results. Seed and fruit size were expected, and confirmed, to be tightly related. As expected, plant height was correlated with each of seed and fruit size, albeit weakly. Weak support was found for an expected positive relationship between leaf and fruit size. The prediction that SLA and WD would be negatively correlated was not supported. Otherwise the traits were predicted to be largely unrelated, being representatives of putatively independent strategy dimensions. This was indeed the case, although WD was consistently, negatively related to leaf size. CONCLUSIONS: The dimensions represented by SLA, seed/fruit size and leaf size were essentially independent and thus conveyed largely independent information about plant strategies. To a lesser extent the same was true for plant height and WD. Our tentative explanation for negative WD-leaf size relationships, now also known from other habitats, is that the traits are indirectly linked via plant hydraulics.
XiangL, ChenFQ, GengMY, WangYB, L��K, YangSL (2019). Response of leaf functional traits of shrubs to altitude in Rhododendron latoucheae communities in Mt. Jinggangshan, Jiangxi, China Journal of Tropical and Subtropical Botany, 27, 129-138. [��������: 2]
YangDM, ZhangJJ, ZhouD, QianMJ, ZhengY, JinLM (2012). Leaf and twig functional traits of woody plants and their relationships with environmental change: a review Chinese Journal of Ecology, 31, 702-713. [��������: 2]
YangWG, ZiHB, ChenKY, AdeLJ, HuL, WangX, WangGX, WangCT (2019). Ecological stoichiometric characteristics of shrubs and soils in different forest types in Qinghai, China Chinese Journal of Plant Ecology, 43, 352-364. [��������: 1]
ZhongQL, LiuLB, XuX, YangY, GuoYM, XuHY, CaiXL, NiJ (2018). Variations of plant functional traits and adaptive strategy of woody species in a karst forest of central Guizhou Province, southwestern China Chinese Journal of Plant Ecology, 42, 562-572. [��������: 1]
The non-native plant Rosa multiflora expresses shade avoidance traits under low light availability 1 2015
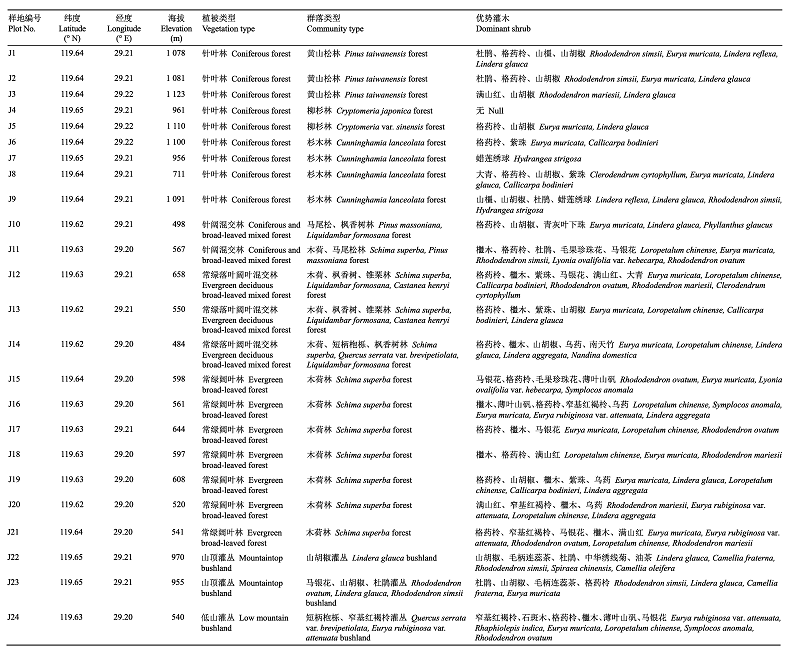
... ���ķ���, 20�ֹ�ľֲ���9��֦Ҷ������״�����¹�ľ�㡢��ɽ�����ɽ�������3�ֲ�ͬ�����¾�������������, ���¹�ľ��LA��SLA������2�ֹ�Թ�ľ, ��LTD��LDMC��TTD��С��2�ֹ�Թ�ľ, TDMCҲԶС�ڵ�ɽ��Զ��Դ���ɽ����Թ�ľ(ͼ1), ��ӳ�˹�ľ�Բ�ͬ��������ԴȨ����Դ��ڲ���.����ɭ��Ⱥ�����¹�ľ���������ȹ�Թ�ľ���ս���, ��Ҷ������Ҷ����ϴ�, ���������¹�ľ�Թ�IJ��������; �ϵ͵�Ҷ��֯�ܶ�������CO2��ϸ����϶���ƶ�(Gratani & Bombelli, 2001), ��������ʽϸ�(�ݲⶨ, ���¹�ľ�ľ��������ƽ��ֵΪ4.02 ��mol��m-2��s-1, ��Թ�ľƽ��ֵΪ3.74 ��mol��m-2��s-1), �Ӷ������ղ������������Ӱ��, ͬʱ�Ϻñ������ڵ�Ӫ��(Li et al., 2014; Rolhauser & Pucheta, 2016).���, ���¹�ľ���ڡ�����Ͷ��-���桱��.��һ����, �ϵ͵�ҶƬ�����ʺ�����ҶƬ��ȡ�Ҷ��С֦��֯�ܶ��Լ���Ƥ��ȵ�, ���������¹�ľ��������������ά�����ƽ��(Chave et al., 2009; ��������, 2018; Luo et al., 2019).ҶƬ��Сͨ����С֦��֯�ܶȵļ�С������(Wright et al., 2007), ����֮���Ȩ���ϵ��ֲ��ˮ��ѧ����, �ϴ�ҶƬ�������ո�������, ��˾��нϴ�ҶƬ�����ֿ��Ը���ؽ����������, �����нϵ͵�ľ���ܶ�(Chave et al., 2009; Wang et al., 2017b).ɭ��Ⱥ�����־���ǿ�ȴ��ڹ��, ���¹�ľ����λ����»�ı�����̬�Դٽ��ڼ��쳤�ͼ��ٷ�֦, ��С֦��֯�ܶ�С, ��Ȼ����еǿ��������, ������������ʸ�, ��������Դ��������֦, �Ի�ȡ����������ٽ�����(Sterck et al., 2006; Dlugos et al., 2015; ��������, 2018).���, ���¹�ľ�γɽϴ�LA��SLA�ͽ�СLTD��LDMC��TTD�ȹ�����״�������ǿ����Դ��ȡ�����ո������ֺ����������ʶ���������, �Ӷ���Ӧ�����١�����������»���, �ǵ��͵���Դ��ȡ�Ͳ���. ...
��ɽ������Ҫֲ�����͵�Ⱥ������ 1 2019
... ��2017��2018���ļ�, �ڽ�ɽ����ѡ����Ҷ�֡������콻�֡�������Ҷ��Ҷ�콻�֡�������Ҷ�֡���ɽ��Ժ�ɽ�����6��ֲ������, ������24���̶�����(��1), ����ɭ����������ֱ�Ϊ30 m �� 30 m, 12����Ҷ�ֺ������콻�־�Ϊ������, 9����Ҷ�־�Ϊ�˹���; 1����ɽ����������Ϊ30 m �� 10 m, ����Ҷ�ֿ�����Ĵ�����ֲ��; 2��ɽ�������������ֱ�Ϊ10 m �� 20 m, Ϊɽ����������Խ�С�Ĵ�����ֲ��.������ɭ�ֺ���������ؾ�(DBH) �� 1 cm, �Լ����������DBH < 1 cm, �߶ȡ�10 cm��ľ��ֲ������Ʊ��, ����¼��Ӧ���������ؾ�(��ľΪ����)�����ߡ��ڷ��Լ���������ָ��.������Զ�ȡ����Ƶ�Ⱥ�������ƶȼ����ľ����Ҫֵ(��������, 2019). ...
��ɽ������Ҫֲ�����͵�Ⱥ������ 1 2019
... ��2017��2018���ļ�, �ڽ�ɽ����ѡ����Ҷ�֡������콻�֡�������Ҷ��Ҷ�콻�֡�������Ҷ�֡���ɽ��Ժ�ɽ�����6��ֲ������, ������24���̶�����(��1), ����ɭ����������ֱ�Ϊ30 m �� 30 m, 12����Ҷ�ֺ������콻�־�Ϊ������, 9����Ҷ�־�Ϊ�˹���; 1����ɽ����������Ϊ30 m �� 10 m, ����Ҷ�ֿ�����Ĵ�����ֲ��; 2��ɽ�������������ֱ�Ϊ10 m �� 20 m, Ϊɽ����������Խ�С�Ĵ�����ֲ��.������ɭ�ֺ���������ؾ�(DBH) �� 1 cm, �Լ����������DBH < 1 cm, �߶ȡ�10 cm��ľ��ֲ������Ʊ��, ����¼��Ӧ���������ؾ�(��ľΪ����)�����ߡ��ڷ��Լ���������ָ��.������Զ�ȡ����Ƶ�Ⱥ�������ƶȼ����ľ����Ҫֵ(��������, 2019). ...
Root and leaf functional trait relations in Poaceae species: implications of differing resource-acquisition strategies 1 2013
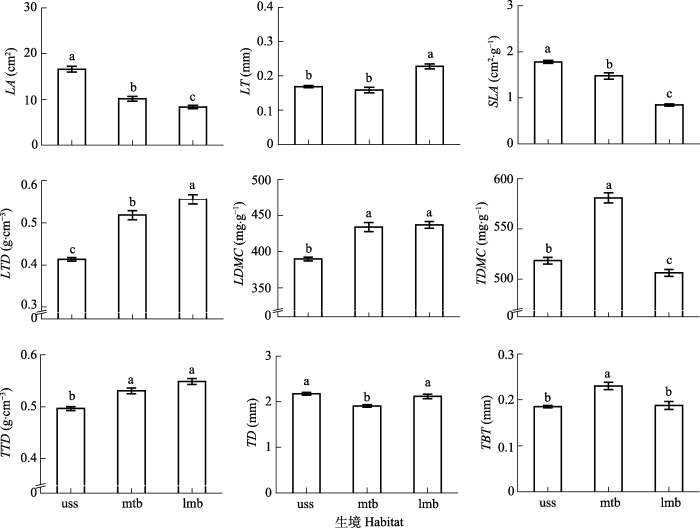
... ��ɽ���ƹ�ľ�Ĺ�����״��Ҫ�ܵ����֡����ֺ������������õ�Ӱ��, ���������������(��3), ��Ҫ�����ڱ��о���3��Ҷ��״(LA��LTD��LDMC)��4��֦��״(TD��TBT��TTD��TDMC), ����2�ֱ��ֲ�����.��ǰ����, ֲ����״��Ҫ�����ַ����λ���������˺���Դ������ͬӰ��(Jung et al., 2010; Auger & Shipley, 2013; Siefert et al., 2015; �������, 2016).����, �����ڳ��ڵĽ�����ʷ��, ��ȡһϵ����ѹ�����״��Ϸ�ӳ���ǶԻ����仯����Ӧ, �Ӷ�ʹ�����ַ����λ��ֲ�﹦����״������ԶӰ��, ��������Ӱ������Զ���ڻ�������(Scheepens et al., 2010; ���ĵ�, 2016).Ҳ����˵, ���ַ��ͬ����ֲ����Ӧ�������ԵIJ�ͬ, �����;�����һ��������ֲ��ԵIJ���(Lebrija- Trejos et al., 2010; Fort et al., 2013), ����Դ����Ҳ�������ֻ��ı���֮һ.���, Ӱ�챾�о��Ĺ�ľ��״��Ҫ�����ֺ��������佻������.ͬһ�����ڵIJ�ͬ���ֹ��淴ӳ������������Ե��ȶ����, ��ֲ���ڲ�ͬ�����µ���״�仯Ҳ���Խ��ͻ����ĸ���, �Ӷ��ٽ����ֹ���, ά����̬ϵͳ���ܵ��ȶ���(Wang et al., 2017b; ��������, 2018; Luo et al., 2019). ...
Leaf traits within communities: context may affect the mapping of traits to function 1 2013
Geographic scale and disturbance influence intraspecific trait variability in leaves and roots of North American understorey plants 1 2019
... ��ֲ�﹦����״�о���, ���ǶԵش��Ե�ֲ��������ɭ�֡���ԭ����Į����ԭ�ȹ�ע�϶�, ���Էǵش��ԵĹ��ֲ���Լ�ɭ���²�Ĺ�ľֲ���ע����.�������еĹ�ľ��״�о���, �������Ե��к������¹�ľֲ����ͻ��뿹����״(Gratani & Bombelli, 2001; Pausas & Bradstock, 2007; de la Riva et al., 2016; de Smedt et al., 2018; Garnier et al., 2019; Nielsen et al., 2019), �Լ���Į��ľ������Բ�(Rolhauser & Pucheta, 2016), �����о����ݶ�������ֲ�����״����, ��ŷ��(Vanneste et al., 2019)�ͱ�����(Kumordzi et al., 2019).���ҹ����Ĺ�ľ������״�뻷����Ӧ�о���, ���о�̽���˶���������ľ��״��γ�ȸ�ּ�����������(Luo et al., 2019), �Լ���ͬ����������(��Զ����, 2018), ���θ߶�(���յ�, 2019), ����(�߾���, 2015), ��ˮ�ݶ���ֲ������(��оع, 2018), �Լ������ؽ�������(Wang et al., 2014)�Թ��ֲ����״��Ӱ��.���Ƕ�ɭ��ֲ�����¹�ľ����״�о�Ҳ����, �漰��ľ��ֲ����̬��״(Wang et al., 2017b)�뻯ѧ��������(���ĸߵ�, 2019), ��ͬ�����(��С��, 2014)�����±���(��������, 2018)��ľС֦��Ҷ��״��Ȩ���ϵ, �Լ����¹�ľ��ͬ����Ҷ��״�IJ���(Wang et al., 2017a).Ȼ��, ��ľ��״���о���ֻ�����漰���ֲ���Ĺ�ľֲ��, �������¹�ľ��Ĺ�ľ��״, ���߶��漰������, �������ص㲻һ��(Luo et al., 2019), ����ͬһ���������¹�ľ���ľ����ֲ���Ĺ�ľ������״���бȽ��о�, �Ӷ���ʾ����״�����IJ���, �Լ���Ӧ����Ӧ����������ʷ���ԵIJ�ͬ, ��δ������. ...
Functional traits and environmental filtering drive community assembly in a species-rich tropical system 1 2010
... ��ɽ���ƹ�ľ�Ĺ�����״��Ҫ�ܵ����֡����ֺ������������õ�Ӱ��, ���������������(��3), ��Ҫ�����ڱ��о���3��Ҷ��״(LA��LTD��LDMC)��4��֦��״(TD��TBT��TTD��TDMC), ����2�ֱ��ֲ�����.��ǰ����, ֲ����״��Ҫ�����ַ����λ���������˺���Դ������ͬӰ��(Jung et al., 2010; Auger & Shipley, 2013; Siefert et al., 2015; �������, 2016).����, �����ڳ��ڵĽ�����ʷ��, ��ȡһϵ����ѹ�����״��Ϸ�ӳ���ǶԻ����仯����Ӧ, �Ӷ�ʹ�����ַ����λ��ֲ�﹦����״������ԶӰ��, ��������Ӱ������Զ���ڻ�������(Scheepens et al., 2010; ���ĵ�, 2016).Ҳ����˵, ���ַ��ͬ����ֲ����Ӧ�������ԵIJ�ͬ, �����;�����һ��������ֲ��ԵIJ���(Lebrija- Trejos et al., 2010; Fort et al., 2013), ����Դ����Ҳ�������ֻ��ı���֮һ.���, Ӱ�챾�о��Ĺ�ľ��״��Ҫ�����ֺ��������佻������.ͬһ�����ڵIJ�ͬ���ֹ��淴ӳ������������Ե��ȶ����, ��ֲ���ڲ�ͬ�����µ���״�仯Ҳ���Խ��ͻ����ĸ���, �Ӷ��ٽ����ֹ���, ά����̬ϵͳ���ܵ��ȶ���(Wang et al., 2017b; ��������, 2018; Luo et al., 2019). ...
Allometric relationships between lamina area, lamina mass and petiole mass of 93 temperate woody species vary with leaf habit, leaf form and altitude 1 2008
... �������Ϳ�, ���¹�ľ��ij��̹�ľ����Ҷ��ľ���н�С��LA��SLA��TDMC��TD, �ϴ��LT��LTD��LDMC��TTD, ��ɽ����Եij��̹�ľ����Ҷ��ľ���н�С��SLA��LTD��LDMC, �ϴ��LT, ������״���첻����(ͼ3).ͨ����˵, ���̱���Ҷֲ����е͵�SLA���͵Ĺ�����ʺߵ����ֺ���, ��������Ҷ, ҶƬ��, ��֯�ܶȴ�, ���������ڹ���������֯�Ե�����ʱ��ĵ��»���; ����Ҷ��ľҶ������, �ڶ�ʱ����۹�ϲ�������ֲ��������Ҫ, ����ҶƬ��֯��������CO2��ɢ;��, ��߹������(Wright et al., 2004; Li et al., 2008; �÷��, 2012).���о�ͬһ�����µ��о������֮һ��, ���̹�ľ����Դ�����Ͳ���, ��Ҷ��ľ����Դ��ȡ�Ͳ���, �������������µ�ͬһ������ľ�������������һ���IJ���, ɽ�������Ҷ��ľ����Դ��ȡ������ǿ. ...
Assimilative branches and leaves of the desert plant Alhagi sparsifolia Shap. possesses a different adaptation mechanism to shade 1 2014
... ���ķ���, 20�ֹ�ľֲ���9��֦Ҷ������״�����¹�ľ�㡢��ɽ�����ɽ�������3�ֲ�ͬ�����¾�������������, ���¹�ľ��LA��SLA������2�ֹ�Թ�ľ, ��LTD��LDMC��TTD��С��2�ֹ�Թ�ľ, TDMCҲԶС�ڵ�ɽ��Զ��Դ���ɽ����Թ�ľ(ͼ1), ��ӳ�˹�ľ�Բ�ͬ��������ԴȨ����Դ��ڲ���.����ɭ��Ⱥ�����¹�ľ���������ȹ�Թ�ľ���ս���, ��Ҷ������Ҷ����ϴ�, ���������¹�ľ�Թ�IJ��������; �ϵ͵�Ҷ��֯�ܶ�������CO2��ϸ����϶���ƶ�(Gratani & Bombelli, 2001), ��������ʽϸ�(�ݲⶨ, ���¹�ľ�ľ��������ƽ��ֵΪ4.02 ��mol��m-2��s-1, ��Թ�ľƽ��ֵΪ3.74 ��mol��m-2��s-1), �Ӷ������ղ������������Ӱ��, ͬʱ�Ϻñ������ڵ�Ӫ��(Li et al., 2014; Rolhauser & Pucheta, 2016).���, ���¹�ľ���ڡ�����Ͷ��-���桱��.��һ����, �ϵ͵�ҶƬ�����ʺ�����ҶƬ��ȡ�Ҷ��С֦��֯�ܶ��Լ���Ƥ��ȵ�, ���������¹�ľ��������������ά�����ƽ��(Chave et al., 2009; ��������, 2018; Luo et al., 2019).ҶƬ��Сͨ����С֦��֯�ܶȵļ�С������(Wright et al., 2007), ����֮���Ȩ���ϵ��ֲ��ˮ��ѧ����, �ϴ�ҶƬ�������ո�������, ��˾��нϴ�ҶƬ�����ֿ��Ը���ؽ����������, �����нϵ͵�ľ���ܶ�(Chave et al., 2009; Wang et al., 2017b).ɭ��Ⱥ�����־���ǿ�ȴ��ڹ��, ���¹�ľ����λ����»�ı�����̬�Դٽ��ڼ��쳤�ͼ��ٷ�֦, ��С֦��֯�ܶ�С, ��Ȼ����еǿ��������, ������������ʸ�, ��������Դ��������֦, �Ի�ȡ����������ٽ�����(Sterck et al., 2006; Dlugos et al., 2015; ��������, 2018).���, ���¹�ľ�γɽϴ�LA��SLA�ͽ�СLTD��LDMC��TTD�ȹ�����״�������ǿ����Դ��ȡ�����ո������ֺ����������ʶ���������, �Ӷ���Ӧ�����١�����������»���, �ǵ��͵���Դ��ȡ�Ͳ���. ...
ֲ�﹦����״�о���չ 3 2015
... ������֪, ֲ�﹦����״��ֲ���ڽ����������뻷����������γɵ�, ����Ӧ��绷���ı仯, ҲӰ����̬ϵͳ�����빦�ܵ�һϵ����̬�����ʡ���������ѧ������(Cornelissen et al., 2003; �����õ�, 2007; Violle et al., 2007; �����������ƽ, 2015; Kattge et al., 2020).���������˽�ֲ����Ӧ����Ӧ��������̬����, ������̬ϵͳ���̺���, �Ӷ���Ч��չ��̬ϵͳ������ֲ���ָ�����Ҫ����, ͬʱҲ��Ԥ�������ȫ��߶��ϵĻ������ű仯����̬ϵͳ�����������Ӱ��ļ�ʵ����, �ڵ�ǰ����̬ѧ�о��д���ǰ�ص�λ(�����õ�, 2007; �����������ƽ, 2015).��ȫ��������߶���, ��ѧ��̽����ֲ��ҶƬ�����ɺ�ϵ��״�ػ����ݶȵķֲ���ּ��价������(Wright et al., 2004; Chave et al., 2009; D��az et al., 2016; ��������, 2018), �Լ�ֲ��ҶƬ��������̬��ѧ����ѧ(Tian et al., 2018)��ϵ������״����̬���������(Ma et al., 2018), �����ڶ�����ˮƽ����״������������̬�Բ߷���.��ͬ������ֲ�﹦����״�ı���, �Լ���״Ȩ������ϵIJ�ͬ, ��ӳֲ����弰��Ⱥ���ȡ������Դ�ķ�ʽ��ͬ, ���ֲ���ȡ���������Ҳ���ڲ���.��ֲ�﹦����״�ĽǶ������ľֲ�������ʷ�Բ�, �������ڽ�һ�����ӹ�ľֲ�����̬����.����, ֲ��ҶƬ��С֦��״��ֲ���ȡ�����û�����Դ�������, ͬʱ����ȡ�����ⶨ���������, ��ֱ�ӷ�ӳֲ����Ӧ����������ʷ�Բ�, �����ֲ����״�о��з����ŹǸɹ���(Kattge et al., 2020). ...
... ; �����������ƽ, 2015).��ȫ��������߶���, ��ѧ��̽����ֲ��ҶƬ�����ɺ�ϵ��״�ػ����ݶȵķֲ���ּ��价������(Wright et al., 2004; Chave et al., 2009; D��az et al., 2016; ��������, 2018), �Լ�ֲ��ҶƬ��������̬��ѧ����ѧ(Tian et al., 2018)��ϵ������״����̬���������(Ma et al., 2018), �����ڶ�����ˮƽ����״������������̬�Բ߷���.��ͬ������ֲ�﹦����״�ı���, �Լ���״Ȩ������ϵIJ�ͬ, ��ӳֲ����弰��Ⱥ���ȡ������Դ�ķ�ʽ��ͬ, ���ֲ���ȡ���������Ҳ���ڲ���.��ֲ�﹦����״�ĽǶ������ľֲ�������ʷ�Բ�, �������ڽ�һ�����ӹ�ľֲ�����̬����.����, ֲ��ҶƬ��С֦��״��ֲ���ȡ�����û�����Դ�������, ͬʱ����ȡ�����ⶨ���������, ��ֱ�ӷ�ӳֲ����Ӧ����������ʷ�Բ�, �����ֲ����״�о��з����ŹǸɹ���(Kattge et al., 2020). ...
Mechanical branch constraints contribute to life-history variation across tree species in a Bolivian forest 1 2006
... ���ķ���, 20�ֹ�ľֲ���9��֦Ҷ������״�����¹�ľ�㡢��ɽ�����ɽ�������3�ֲ�ͬ�����¾�������������, ���¹�ľ��LA��SLA������2�ֹ�Թ�ľ, ��LTD��LDMC��TTD��С��2�ֹ�Թ�ľ, TDMCҲԶС�ڵ�ɽ��Զ��Դ���ɽ����Թ�ľ(ͼ1), ��ӳ�˹�ľ�Բ�ͬ��������ԴȨ����Դ��ڲ���.����ɭ��Ⱥ�����¹�ľ���������ȹ�Թ�ľ���ս���, ��Ҷ������Ҷ����ϴ�, ���������¹�ľ�Թ�IJ��������; �ϵ͵�Ҷ��֯�ܶ�������CO2��ϸ����϶���ƶ�(Gratani & Bombelli, 2001), ��������ʽϸ�(�ݲⶨ, ���¹�ľ�ľ��������ƽ��ֵΪ4.02 ��mol��m-2��s-1, ��Թ�ľƽ��ֵΪ3.74 ��mol��m-2��s-1), �Ӷ������ղ������������Ӱ��, ͬʱ�Ϻñ������ڵ�Ӫ��(Li et al., 2014; Rolhauser & Pucheta, 2016).���, ���¹�ľ���ڡ�����Ͷ��-���桱��.��һ����, �ϵ͵�ҶƬ�����ʺ�����ҶƬ��ȡ�Ҷ��С֦��֯�ܶ��Լ���Ƥ��ȵ�, ���������¹�ľ��������������ά�����ƽ��(Chave et al., 2009; ��������, 2018; Luo et al., 2019).ҶƬ��Сͨ����С֦��֯�ܶȵļ�С������(Wright et al., 2007), ����֮���Ȩ���ϵ��ֲ��ˮ��ѧ����, �ϴ�ҶƬ�������ո�������, ��˾��нϴ�ҶƬ�����ֿ��Ը���ؽ����������, �����нϵ͵�ľ���ܶ�(Chave et al., 2009; Wang et al., 2017b).ɭ��Ⱥ�����־���ǿ�ȴ��ڹ��, ���¹�ľ����λ����»�ı�����̬�Դٽ��ڼ��쳤�ͼ��ٷ�֦, ��С֦��֯�ܶ�С, ��Ȼ����еǿ��������, ������������ʸ�, ��������Դ��������֦, �Ի�ȡ����������ٽ�����(Sterck et al., 2006; Dlugos et al., 2015; ��������, 2018).���, ���¹�ľ�γɽϴ�LA��SLA�ͽ�СLTD��LDMC��TTD�ȹ�����״�������ǿ����Դ��ȡ�����ո������ֺ����������ʶ���������, �Ӷ���Ӧ�����١�����������»���, �ǵ��͵���Դ��ȡ�Ͳ���. ...
���ȴ�������Ҷ��Ҷ�콻��ֲ�﹦����״���ּ�����ڱ��� 1 2016
... ��ɽ���ƹ�ľ�Ĺ�����״��Ҫ�ܵ����֡����ֺ������������õ�Ӱ��, ���������������(��3), ��Ҫ�����ڱ��о���3��Ҷ��״(LA��LTD��LDMC)��4��֦��״(TD��TBT��TTD��TDMC), ����2�ֱ��ֲ�����.��ǰ����, ֲ����״��Ҫ�����ַ����λ���������˺���Դ������ͬӰ��(Jung et al., 2010; Auger & Shipley, 2013; Siefert et al., 2015; �������, 2016).����, �����ڳ��ڵĽ�����ʷ��, ��ȡһϵ����ѹ�����״��Ϸ�ӳ���ǶԻ����仯����Ӧ, �Ӷ�ʹ�����ַ����λ��ֲ�﹦����״������ԶӰ��, ��������Ӱ������Զ���ڻ�������(Scheepens et al., 2010; ���ĵ�, 2016).Ҳ����˵, ���ַ��ͬ����ֲ����Ӧ�������ԵIJ�ͬ, �����;�����һ��������ֲ��ԵIJ���(Lebrija- Trejos et al., 2010; Fort et al., 2013), ����Դ����Ҳ�������ֻ��ı���֮һ.���, Ӱ�챾�о��Ĺ�ľ��״��Ҫ�����ֺ��������佻������.ͬһ�����ڵIJ�ͬ���ֹ��淴ӳ������������Ե��ȶ����, ��ֲ���ڲ�ͬ�����µ���״�仯Ҳ���Խ��ͻ����ĸ���, �Ӷ��ٽ����ֹ���, ά����̬ϵͳ���ܵ��ȶ���(Wang et al., 2017b; ��������, 2018; Luo et al., 2019). ...
���ȴ�������Ҷ��Ҷ�콻��ֲ�﹦����״���ּ�����ڱ��� 1 2016
... ��ɽ���ƹ�ľ�Ĺ�����״��Ҫ�ܵ����֡����ֺ������������õ�Ӱ��, ���������������(��3), ��Ҫ�����ڱ��о���3��Ҷ��״(LA��LTD��LDMC)��4��֦��״(TD��TBT��TTD��TDMC), ����2�ֱ��ֲ�����.��ǰ����, ֲ����״��Ҫ�����ַ����λ���������˺���Դ������ͬӰ��(Jung et al., 2010; Auger & Shipley, 2013; Siefert et al., 2015; �������, 2016).����, �����ڳ��ڵĽ�����ʷ��, ��ȡһϵ����ѹ�����״��Ϸ�ӳ���ǶԻ����仯����Ӧ, �Ӷ�ʹ�����ַ����λ��ֲ�﹦����״������ԶӰ��, ��������Ӱ������Զ���ڻ�������(Scheepens et al., 2010; ���ĵ�, 2016).Ҳ����˵, ���ַ��ͬ����ֲ����Ӧ�������ԵIJ�ͬ, �����;�����һ��������ֲ��ԵIJ���(Lebrija- Trejos et al., 2010; Fort et al., 2013), ����Դ����Ҳ�������ֻ��ı���֮һ.���, Ӱ�챾�о��Ĺ�ľ��״��Ҫ�����ֺ��������佻������.ͬһ�����ڵIJ�ͬ���ֹ��淴ӳ������������Ե��ȶ����, ��ֲ���ڲ�ͬ�����µ���״�仯Ҳ���Խ��ͻ����ĸ���, �Ӷ��ٽ����ֹ���, ά����̬ϵͳ���ܵ��ȶ���(Wang et al., 2017b; ��������, 2018; Luo et al., 2019). ...
Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent 1 2018
... ������֪, ֲ�﹦����״��ֲ���ڽ����������뻷����������γɵ�, ����Ӧ��绷���ı仯, ҲӰ����̬ϵͳ�����빦�ܵ�һϵ����̬�����ʡ���������ѧ������(Cornelissen et al., 2003; �����õ�, 2007; Violle et al., 2007; �����������ƽ, 2015; Kattge et al., 2020).���������˽�ֲ����Ӧ����Ӧ��������̬����, ������̬ϵͳ���̺���, �Ӷ���Ч��չ��̬ϵͳ������ֲ���ָ�����Ҫ����, ͬʱҲ��Ԥ�������ȫ��߶��ϵĻ������ű仯����̬ϵͳ�����������Ӱ��ļ�ʵ����, �ڵ�ǰ����̬ѧ�о��д���ǰ�ص�λ(�����õ�, 2007; �����������ƽ, 2015).��ȫ��������߶���, ��ѧ��̽����ֲ��ҶƬ�����ɺ�ϵ��״�ػ����ݶȵķֲ���ּ��价������(Wright et al., 2004; Chave et al., 2009; D��az et al., 2016; ��������, 2018), �Լ�ֲ��ҶƬ��������̬��ѧ����ѧ(Tian et al., 2018)��ϵ������״����̬���������(Ma et al., 2018), �����ڶ�����ˮƽ����״������������̬�Բ߷���.��ͬ������ֲ�﹦����״�ı���, �Լ���״Ȩ������ϵIJ�ͬ, ��ӳֲ����弰��Ⱥ���ȡ������Դ�ķ�ʽ��ͬ, ���ֲ���ȡ���������Ҳ���ڲ���.��ֲ�﹦����״�ĽǶ������ľֲ�������ʷ�Բ�, �������ڽ�һ�����ӹ�ľֲ�����̬����.����, ֲ��ҶƬ��С֦��״��ֲ���ȡ�����û�����Դ�������, ͬʱ����ȡ�����ⶨ���������, ��ֱ�ӷ�ӳֲ����Ӧ����������ʷ�Բ�, �����ֲ����״�о��з����ŹǸɹ���(Kattge et al., 2020). ...
Functional trait variation of forest understorey plant communities across Europe 1 2019
... ��ֲ�﹦����״�о���, ���ǶԵش��Ե�ֲ��������ɭ�֡���ԭ����Į����ԭ�ȹ�ע�϶�, ���Էǵش��ԵĹ��ֲ���Լ�ɭ���²�Ĺ�ľֲ���ע����.�������еĹ�ľ��״�о���, �������Ե��к������¹�ľֲ����ͻ��뿹����״(Gratani & Bombelli, 2001; Pausas & Bradstock, 2007; de la Riva et al., 2016; de Smedt et al., 2018; Garnier et al., 2019; Nielsen et al., 2019), �Լ���Į��ľ������Բ�(Rolhauser & Pucheta, 2016), �����о����ݶ�������ֲ�����״����, ��ŷ��(Vanneste et al., 2019)�ͱ�����(Kumordzi et al., 2019).���ҹ����Ĺ�ľ������״�뻷����Ӧ�о���, ���о�̽���˶���������ľ��״��γ�ȸ�ּ�����������(Luo et al., 2019), �Լ���ͬ����������(��Զ����, 2018), ���θ߶�(���յ�, 2019), ����(�߾���, 2015), ��ˮ�ݶ���ֲ������(��оع, 2018), �Լ������ؽ�������(Wang et al., 2014)�Թ��ֲ����״��Ӱ��.���Ƕ�ɭ��ֲ�����¹�ľ����״�о�Ҳ����, �漰��ľ��ֲ����̬��״(Wang et al., 2017b)�뻯ѧ��������(���ĸߵ�, 2019), ��ͬ�����(��С��, 2014)�����±���(��������, 2018)��ľС֦��Ҷ��״��Ȩ���ϵ, �Լ����¹�ľ��ͬ����Ҷ��״�IJ���(Wang et al., 2017a).Ȼ��, ��ľ��״���о���ֻ�����漰���ֲ���Ĺ�ľֲ��, �������¹�ľ��Ĺ�ľ��״, ���߶��漰������, �������ص㲻һ��(Luo et al., 2019), ����ͬһ���������¹�ľ���ľ����ֲ���Ĺ�ľ������״���бȽ��о�, �Ӷ���ʾ����״�����IJ���, �Լ���Ӧ����Ӧ����������ʷ���ԵIJ�ͬ, ��δ������. ...
Let the concept of trait be functional! 1 2007
... ������֪, ֲ�﹦����״��ֲ���ڽ����������뻷����������γɵ�, ����Ӧ��绷���ı仯, ҲӰ����̬ϵͳ�����빦�ܵ�һϵ����̬�����ʡ���������ѧ������(Cornelissen et al., 2003; �����õ�, 2007; Violle et al., 2007; �����������ƽ, 2015; Kattge et al., 2020).���������˽�ֲ����Ӧ����Ӧ��������̬����, ������̬ϵͳ���̺���, �Ӷ���Ч��չ��̬ϵͳ������ֲ���ָ�����Ҫ����, ͬʱҲ��Ԥ�������ȫ��߶��ϵĻ������ű仯����̬ϵͳ�����������Ӱ��ļ�ʵ����, �ڵ�ǰ����̬ѧ�о��д���ǰ�ص�λ(�����õ�, 2007; �����������ƽ, 2015).��ȫ��������߶���, ��ѧ��̽����ֲ��ҶƬ�����ɺ�ϵ��״�ػ����ݶȵķֲ���ּ��价������(Wright et al., 2004; Chave et al., 2009; D��az et al., 2016; ��������, 2018), �Լ�ֲ��ҶƬ��������̬��ѧ����ѧ(Tian et al., 2018)��ϵ������״����̬���������(Ma et al., 2018), �����ڶ�����ˮƽ����״������������̬�Բ߷���.��ͬ������ֲ�﹦����״�ı���, �Լ���״Ȩ������ϵIJ�ͬ, ��ӳֲ����弰��Ⱥ���ȡ������Դ�ķ�ʽ��ͬ, ���ֲ���ȡ���������Ҳ���ڲ���.��ֲ�﹦����״�ĽǶ������ľֲ�������ʷ�Բ�, �������ڽ�һ�����ӹ�ľֲ�����̬����.����, ֲ��ҶƬ��С֦��״��ֲ���ȡ�����û�����Դ�������, ͬʱ����ȡ�����ⶨ���������, ��ֱ�ӷ�ӳֲ����Ӧ����������ʷ�Բ�, �����ֲ����״�о��з����ŹǸɹ���(Kattge et al., 2020). ...
Differences in leaf functional traits between red and green leaves of two evergreen shrubs Photinia �� fraseri and Osmanthus fragrans 1 2017
... ��ֲ�﹦����״�о���, ���ǶԵش��Ե�ֲ��������ɭ�֡���ԭ����Į����ԭ�ȹ�ע�϶�, ���Էǵش��ԵĹ��ֲ���Լ�ɭ���²�Ĺ�ľֲ���ע����.�������еĹ�ľ��״�о���, �������Ե��к������¹�ľֲ����ͻ��뿹����״(Gratani & Bombelli, 2001; Pausas & Bradstock, 2007; de la Riva et al., 2016; de Smedt et al., 2018; Garnier et al., 2019; Nielsen et al., 2019), �Լ���Į��ľ������Բ�(Rolhauser & Pucheta, 2016), �����о����ݶ�������ֲ�����״����, ��ŷ��(Vanneste et al., 2019)�ͱ�����(Kumordzi et al., 2019).���ҹ����Ĺ�ľ������״�뻷����Ӧ�о���, ���о�̽���˶���������ľ��״��γ�ȸ�ּ�����������(Luo et al., 2019), �Լ���ͬ����������(��Զ����, 2018), ���θ߶�(���յ�, 2019), ����(�߾���, 2015), ��ˮ�ݶ���ֲ������(��оع, 2018), �Լ������ؽ�������(Wang et al., 2014)�Թ��ֲ����״��Ӱ��.���Ƕ�ɭ��ֲ�����¹�ľ����״�о�Ҳ����, �漰��ľ��ֲ����̬��״(Wang et al., 2017b)�뻯ѧ��������(���ĸߵ�, 2019), ��ͬ�����(��С��, 2014)�����±���(��������, 2018)��ľС֦��Ҷ��״��Ȩ���ϵ, �Լ����¹�ľ��ͬ����Ҷ��״�IJ���(Wang et al., 2017a).Ȼ��, ��ľ��״���о���ֻ�����漰���ֲ���Ĺ�ľֲ��, �������¹�ľ��Ĺ�ľ��״, ���߶��漰������, �������ص㲻һ��(Luo et al., 2019), ����ͬһ���������¹�ľ���ľ����ֲ���Ĺ�ľ������״���бȽ��о�, �Ӷ���ʾ����״�����IJ���, �Լ���Ӧ����Ӧ����������ʷ���ԵIJ�ͬ, ��δ������. ...
Relationships among leaf, stem and root traits of the dominant shrubs from four vegetation zones in Shaanxi Province, China 3 2017
... ��ֲ�﹦����״�о���, ���ǶԵش��Ե�ֲ��������ɭ�֡���ԭ����Į����ԭ�ȹ�ע�϶�, ���Էǵش��ԵĹ��ֲ���Լ�ɭ���²�Ĺ�ľֲ���ע����.�������еĹ�ľ��״�о���, �������Ե��к������¹�ľֲ����ͻ��뿹����״(Gratani & Bombelli, 2001; Pausas & Bradstock, 2007; de la Riva et al., 2016; de Smedt et al., 2018; Garnier et al., 2019; Nielsen et al., 2019), �Լ���Į��ľ������Բ�(Rolhauser & Pucheta, 2016), �����о����ݶ�������ֲ�����״����, ��ŷ��(Vanneste et al., 2019)�ͱ�����(Kumordzi et al., 2019).���ҹ����Ĺ�ľ������״�뻷����Ӧ�о���, ���о�̽���˶���������ľ��״��γ�ȸ�ּ�����������(Luo et al., 2019), �Լ���ͬ����������(��Զ����, 2018), ���θ߶�(���յ�, 2019), ����(�߾���, 2015), ��ˮ�ݶ���ֲ������(��оع, 2018), �Լ������ؽ�������(Wang et al., 2014)�Թ��ֲ����״��Ӱ��.���Ƕ�ɭ��ֲ�����¹�ľ����״�о�Ҳ����, �漰��ľ��ֲ����̬��״(Wang et al., 2017b)�뻯ѧ��������(���ĸߵ�, 2019), ��ͬ�����(��С��, 2014)�����±���(��������, 2018)��ľС֦��Ҷ��״��Ȩ���ϵ, �Լ����¹�ľ��ͬ����Ҷ��״�IJ���(Wang et al., 2017a).Ȼ��, ��ľ��״���о���ֻ�����漰���ֲ���Ĺ�ľֲ��, �������¹�ľ��Ĺ�ľ��״, ���߶��漰������, �������ص㲻һ��(Luo et al., 2019), ����ͬһ���������¹�ľ���ľ����ֲ���Ĺ�ľ������״���бȽ��о�, �Ӷ���ʾ����״�����IJ���, �Լ���Ӧ����Ӧ����������ʷ���ԵIJ�ͬ, ��δ������. ...
... ���ķ���, 20�ֹ�ľֲ���9��֦Ҷ������״�����¹�ľ�㡢��ɽ�����ɽ�������3�ֲ�ͬ�����¾�������������, ���¹�ľ��LA��SLA������2�ֹ�Թ�ľ, ��LTD��LDMC��TTD��С��2�ֹ�Թ�ľ, TDMCҲԶС�ڵ�ɽ��Զ��Դ���ɽ����Թ�ľ(ͼ1), ��ӳ�˹�ľ�Բ�ͬ��������ԴȨ����Դ��ڲ���.����ɭ��Ⱥ�����¹�ľ���������ȹ�Թ�ľ���ս���, ��Ҷ������Ҷ����ϴ�, ���������¹�ľ�Թ�IJ��������; �ϵ͵�Ҷ��֯�ܶ�������CO2��ϸ����϶���ƶ�(Gratani & Bombelli, 2001), ��������ʽϸ�(�ݲⶨ, ���¹�ľ�ľ��������ƽ��ֵΪ4.02 ��mol��m-2��s-1, ��Թ�ľƽ��ֵΪ3.74 ��mol��m-2��s-1), �Ӷ������ղ������������Ӱ��, ͬʱ�Ϻñ������ڵ�Ӫ��(Li et al., 2014; Rolhauser & Pucheta, 2016).���, ���¹�ľ���ڡ�����Ͷ��-���桱��.��һ����, �ϵ͵�ҶƬ�����ʺ�����ҶƬ��ȡ�Ҷ��С֦��֯�ܶ��Լ���Ƥ��ȵ�, ���������¹�ľ��������������ά�����ƽ��(Chave et al., 2009; ��������, 2018; Luo et al., 2019).ҶƬ��Сͨ����С֦��֯�ܶȵļ�С������(Wright et al., 2007), ����֮���Ȩ���ϵ��ֲ��ˮ��ѧ����, �ϴ�ҶƬ�������ո�������, ��˾��нϴ�ҶƬ�����ֿ��Ը���ؽ����������, �����нϵ͵�ľ���ܶ�(Chave et al., 2009; Wang et al., 2017b).ɭ��Ⱥ�����־���ǿ�ȴ��ڹ��, ���¹�ľ����λ����»�ı�����̬�Դٽ��ڼ��쳤�ͼ��ٷ�֦, ��С֦��֯�ܶ�С, ��Ȼ����еǿ��������, ������������ʸ�, ��������Դ��������֦, �Ի�ȡ����������ٽ�����(Sterck et al., 2006; Dlugos et al., 2015; ��������, 2018).���, ���¹�ľ�γɽϴ�LA��SLA�ͽ�СLTD��LDMC��TTD�ȹ�����״�������ǿ����Դ��ȡ�����ո������ֺ����������ʶ���������, �Ӷ���Ӧ�����١�����������»���, �ǵ��͵���Դ��ȡ�Ͳ���. ...
... ��ɽ���ƹ�ľ�Ĺ�����״��Ҫ�ܵ����֡����ֺ������������õ�Ӱ��, ���������������(��3), ��Ҫ�����ڱ��о���3��Ҷ��״(LA��LTD��LDMC)��4��֦��״(TD��TBT��TTD��TDMC), ����2�ֱ��ֲ�����.��ǰ����, ֲ����״��Ҫ�����ַ����λ���������˺���Դ������ͬӰ��(Jung et al., 2010; Auger & Shipley, 2013; Siefert et al., 2015; �������, 2016).����, �����ڳ��ڵĽ�����ʷ��, ��ȡһϵ����ѹ�����״��Ϸ�ӳ���ǶԻ����仯����Ӧ, �Ӷ�ʹ�����ַ����λ��ֲ�﹦����״������ԶӰ��, ��������Ӱ������Զ���ڻ�������(Scheepens et al., 2010; ���ĵ�, 2016).Ҳ����˵, ���ַ��ͬ����ֲ����Ӧ�������ԵIJ�ͬ, �����;�����һ��������ֲ��ԵIJ���(Lebrija- Trejos et al., 2010; Fort et al., 2013), ����Դ����Ҳ�������ֻ��ı���֮һ.���, Ӱ�챾�о��Ĺ�ľ��״��Ҫ�����ֺ��������佻������.ͬһ�����ڵIJ�ͬ���ֹ��淴ӳ������������Ե��ȶ����, ��ֲ���ڲ�ͬ�����µ���״�仯Ҳ���Խ��ͻ����ĸ���, �Ӷ��ٽ����ֹ���, ά����̬ϵͳ���ܵ��ȶ���(Wang et al., 2017b; ��������, 2018; Luo et al., 2019). ...
Geographic variation in seed traits within and among forty-two species of Rhododendron(Ericaceae) on the Tibetan Plateau: relationships with altitude, habitat, plant height, and phylogeny 1 2014
... ��ֲ�﹦����״�о���, ���ǶԵش��Ե�ֲ��������ɭ�֡���ԭ����Į����ԭ�ȹ�ע�϶�, ���Էǵش��ԵĹ��ֲ���Լ�ɭ���²�Ĺ�ľֲ���ע����.�������еĹ�ľ��״�о���, �������Ե��к������¹�ľֲ����ͻ��뿹����״(Gratani & Bombelli, 2001; Pausas & Bradstock, 2007; de la Riva et al., 2016; de Smedt et al., 2018; Garnier et al., 2019; Nielsen et al., 2019), �Լ���Į��ľ������Բ�(Rolhauser & Pucheta, 2016), �����о����ݶ�������ֲ�����״����, ��ŷ��(Vanneste et al., 2019)�ͱ�����(Kumordzi et al., 2019).���ҹ����Ĺ�ľ������״�뻷����Ӧ�о���, ���о�̽���˶���������ľ��״��γ�ȸ�ּ�����������(Luo et al., 2019), �Լ���ͬ����������(��Զ����, 2018), ���θ߶�(���յ�, 2019), ����(�߾���, 2015), ��ˮ�ݶ���ֲ������(��оع, 2018), �Լ������ؽ�������(Wang et al., 2014)�Թ��ֲ����״��Ӱ��.���Ƕ�ɭ��ֲ�����¹�ľ����״�о�Ҳ����, �漰��ľ��ֲ����̬��״(Wang et al., 2017b)�뻯ѧ��������(���ĸߵ�, 2019), ��ͬ�����(��С��, 2014)�����±���(��������, 2018)��ľС֦��Ҷ��״��Ȩ���ϵ, �Լ����¹�ľ��ͬ����Ҷ��״�IJ���(Wang et al., 2017a).Ȼ��, ��ľ��״���о���ֻ�����漰���ֲ���Ĺ�ľֲ��, �������¹�ľ��Ĺ�ľ��״, ���߶��漰������, �������ص㲻һ��(Luo et al., 2019), ����ͬһ���������¹�ľ���ľ����ֲ���Ĺ�ľ������״���бȽ��о�, �Ӷ���ʾ����״�����IJ���, �Լ���Ӧ����Ӧ����������ʷ���ԵIJ�ͬ, ��δ������. ...
Understanding ecological variation across species: area-based vs mass-based expression of leaf traits 1 2013
 ,
, ,1,2,*
,1,2,*

 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT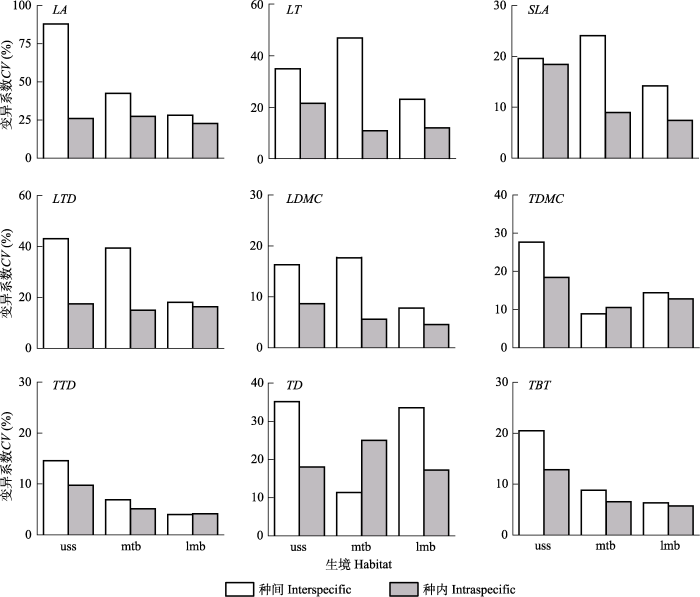 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}