A review on the effects of nitrogen and phosphorus addition on tree growth and productivity in forest ecosystems
Ji-Guang FENG,, Biao ZHU,,*Institute of Ecology, College of Urban and Environmental Sciences, and Key Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China
Abstract Nitrogen (N) and phosphorus (P) inputs induced by anthropogenic activities and atmospheric N and P deposition have largely increased the availability of soil N and P in terrestrial ecosystems, which have considerably affected terrestrial carbon cycling processes. Tree growth and productivity in forest ecosystems play an important role in global carbon cycling, and determine the magnitude and direction of terrestrial carbon sequestration. Currently, a large number of field manipulation experiments have been conducted to investigate the effects of N and/or P addition on tree growth and forest productivity, but the results from these studies were inconsistent. Such inconsistent results might be affected by multiple factors, including biological, environmental and experimental variables. Here, we reviewed the present research status of the effects of N and P addition on tree growth and forest productivity in forest ecosystems based on three aspects, including the number of publications and experiments with field N and P addition, and the global distributions of these experiments. Then, we summarized the methods for assessing tree growth and forest productivity at ecosystem level in forest ecosystems, including relative growth rate and absolute increment. According to the related results, we reviewed the regulating factors that affect tree growth and productivity, and the potential mechanisms for such factors, including climate, tree size and stand age, plant functional traits (including type of tree-associated mycorrhizal fungi, N-fixation property of trees, and conservative and acquisitive functional traits), plant-microbe interaction, ambient nutrient (i.e., N and P) deposition rate, and experimental variables. Finally, we summarized the current studies, and pointed out five aspects that are urgently needed to provide further insights in future studies, including the physiological mechanism of how tree growth responds to N and P addition, the tradeoff and allocation among growth of various parts of tree under N and P addition, the role of plant functional traits in regulating and predicting the responses of tree growth to N and P addition, how the competition among trees regulates the responses of tree growth to N and P addition, and conducting long-term and coordinated distributed field experiments investigating the effects of N and P addition on tree growth and forest productivity at the global scale. Keywords:nutrient limitation;tree growth;ecosystem productivity;plant functional traits;plant-microbe interaction;nitrogen deposition;phosphorus deposition
PDF (2242KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 冯继广, 朱彪. 氮磷添加对树木生长和森林生产力影响的研究进展. 植物生态学报, 2020, 44(6): 583-597. DOI: 10.17521/cjpe.2019.0176 FENG Ji-Guang, ZHU Biao. A review on the effects of nitrogen and phosphorus addition on tree growth and productivity in forest ecosystems. Chinese Journal of Plant Ecology, 2020, 44(6): 583-597. DOI: 10.17521/cjpe.2019.0176
A, 1970-2019年每年的论文数量及累积论文数量。B, 不同N、P添加处理在不同气候区森林的实验数量。N表示该研究中只有N添加; P表示该研究中只有P添加; N, P表示该研究中有单独的N和单独的P添加; N, P, NP表明该研究中包含单独的N、单独的P和N、P共同添加。 Fig. 1Number of papers and experiments investigating the effects of nitrogen (N) and/or phosphorus (P) addition on tree growth and productivity in forest ecosystems.
A, The number of papers in each year and the accumulated number of papers between the year 1970 and 2019. B, The number of experiments with different treatment of nitrogen and phosphorus addition. N represents only N addition in the case study; P represents only P addition in the case study; N, P represent only N and only P addition in the case study; N, P, NP represents only N, only P and N plus P addition in the case study.
N表示该研究中只有N添加; P表示该研究中只有P添加; N, P表示该研究中有单独的N和单独的P添加; N, P, NP表明该研究中包含单独的N、单独的P和N、P共同添加。该图使用ArcGIS 10.2.2绘制。 Fig. 2Global distribution of case studies investigating the effects of nitrogen (N) and/or phosphorus (P) addition on tree growth and productivity in forest ecosystems. The horizontal lines in the figure indicate the distribution boundaries of forests in different climatic regions based on latitude, including tropical forest (23.5° S-23.5° N), boreal forest (46°-66° N), and temperate forest (between the tropical and boreal latitudes).
N represents only N addition in the case study; P represents only P addition in the case study; N, P represent only N and only P addition in the case study; N, P, NP represents only N, only P and N plus P addition in the case study. This map was drawn using the software ArcGIS 10.2.2.
BA, 胸高断面积; BP, 生物量生产; DBH, 胸径; t1和t2分别代表起始和最后的测量时间。 BA, basal area; BP, biomass production; DBH, diameter at breast height; t1 and t2 represent the starting and last measurement time, respectively.
AM, 丛枝菌根; ECM, 外生菌根。 Fig. 3Summary diagram for studying the effects of nitrogen and/or phosphorus addition on tree growth and productivity in forest ecosystems.
AM, arbuscular mycorrhizal; ECM, ectomycorrhizal.
(1)加强树木生长响应N、P添加的生理学机制研究。一项在N富集的热带森林中的N添加实验表明, 10年左右的N添加使土壤发生了酸化, 降低了土壤中的盐基离子(钙和镁)的可利用性, 但是树木能够通过增强叶片蒸腾作用提高对盐基离子的吸收来维持自身的养分平衡(Lu et al., 2018)。N和P的可利用性提高后, 植物的生理活动会发生相应的响应, 这些响应可能会对树木的生长产生影响, 并最终反应到DBH和生物量变化上。目前, 仅有少数研究(Zhu et al., 2014; Li et al., 2018; Lu et al., 2018)对N、P添加后的树木生理水平上的响应(光合作用、蒸腾作用、叶片养分重吸收效率等)进行了测定。因此, 后续研究可以从植物的生理角度开展, 如光合作用、水分传输、根系寿命等, 以期能够提供深入的机理解释。
Alvarez-ClareS, MackMC, BrooksM (2013). A direct test of nitrogen and phosphorus limitation to net primary productivity in a lowland tropical wet forest Ecology, 94, 1540-1551. DOI:10.1890/12-2128.1URL [本文引用: 5] 30 cm dbh). Phosphorus additions also increased the percentage of seedling survival from 59% to 78% (P
AndersenKM, CorreMD, TurnerBL, DallingJW (2010). Plant-soil associations in a lower montane tropical forest: physiological acclimation and herbivore-mediated responses to nitrogen addition Functional Ecology, 24, 1171-1180. DOI:10.1111/j.1365-2435.2010.01731.xURL [本文引用: 1]
Arag?oLEOC, MalhiY, MetcalfeDB, Silva-EspejoJE, JiménezE, NavarreteD, AlmeidaS, CostaACL, SalinasN, PhillipsOL, AndersonLO, AlvarezE, BakerTR, GoncalvezPH, Huamán-OvalleJ, Mamani-SolórzanoM, MeirP, MonteagudoA, Pati?oS, Pe?uelaMC, PrietoA, QuesadaCA, Rozas-DávilaA, RudasA, Silva JrJA, VásquezR (2009). Above- and below-ground net primary productivity across ten Amazonian forests on contrasting soils Biogeosciences, 6, 2759-2778. DOI:10.5194/bg-6-2759-2009URL [本文引用: 1]
AverillC, DietzeMC, BhatnagarJM (2018). Continental-scale nitrogen pollution is shifting forest mycorrhizal associations and soil carbon stocks Global Change Biology, 24, 4544-4553. DOI:10.1111/gcb.14368URLPMID:30051940 [本文引用: 4] Most tree roots on Earth form a symbiosis with either ecto- or arbuscular mycorrhizal fungi. Nitrogen fertilization is hypothesized to favor arbuscular mycorrhizal tree species at the expense of ectomycorrhizal species due to differences in fungal nitrogen acquisition strategies, and this may alter soil carbon balance, as differences in forest mycorrhizal associations are linked to differences in soil carbon pools. Combining nitrogen deposition data with continental-scale US forest data, we show that nitrogen pollution is spatially associated with a decline in ectomycorrhizal vs. arbuscular mycorrhizal trees. Furthermore, nitrogen deposition has contrasting effects on arbuscular vs. ectomycorrhizal demographic processes, favoring arbuscular mycorrhizal trees at the expense of ectomycorrhizal trees, and is spatially correlated with reduced soil carbon stocks. This implies future changes in nitrogen deposition may alter the capacity of forests to sequester carbon and offset climate change via interactions with the forest microbiome.
BáezS, HomeierJ (2018). Functional traits determine tree growth and ecosystem productivity of a tropical montane forest: insights from a long-term nutrient manipulation experiment Global Change Biology, 24, 399-409. DOI:10.1111/gcb.13905URLPMID:28921844 [本文引用: 4] Trait-response effects are critical to forecast community structure and biomass production in highly diverse tropical forests. Ecological theory and few observation studies indicate that trees with acquisitive functional traits would respond more strongly to higher resource availability than those with conservative traits. We assessed how long-term tree growth in experimental nutrient addition plots (N, P, and N + P) varied as a function of morphological traits, tree size, and species identity. We also evaluated how trait-based responses affected stand scale biomass production considering the community structure. We found that tree growth depended on interactions between functional traits and the type or combination of nutrients added. Common species with acquisitive functional traits responded more strongly to nutrient addition, mainly to N + P. Phosphorous enhanced the growth rates of species with acquisitive and conservative traits, had mostly positive effects on common species and neutral or negative effects in rare species. Moreover, trees receiving N + P grew faster irrespective of their initial size relative to trees in control or to trees in other treatment plots. Finally, species responses were highly idiosyncratic suggesting that community processes including competition and niche dimensionality may be altered under increased resource availability. We found no statistically significant effects of nutrient additions on aboveground biomass productivity because acquisitive species had a limited potential to increase their biomass, possibly due to their generally lower wood density. In contrast, P addition increased the growth rates of species characterized by more conservative resource strategies (with higher wood density) that were poorly represented in the plant community. We provide the first long-term experimental evidence that trait-based responses, community structure, and community processes modulate the effects of increased nutrient availability on biomass productivity in a tropical forest.
BardgettRD, MommerL, de VriesFT (2014). Going underground: root traits as drivers of ecosystem processes Trends in Ecology & Evolution, 29, 692-699. DOI:10.1016/j.tree.2014.10.006URLPMID:25459399 [本文引用: 1] Ecologists are increasingly adopting trait-based approaches to understand how community change influences ecosystem processes. However, most of this research has focussed on aboveground plant traits, whereas it is becoming clear that root traits are important drivers of many ecosystem processes, such as carbon (C) and nutrient cycling, and the formation and structural stability of soil. Here, we synthesise emerging evidence that illustrates how root traits impact ecosystem processes, and propose a pathway to unravel the complex roles of root traits in driving ecosystem processes and their response to global change. Finally, we identify research challenges and novel technologies to address them.
BassiriRadH, LussenhopJF, SehtiyaHL, BordenKK (2015). Nitrogen deposition potentially contributes to oak regeneration failure in the Midwestern temperate forests of the USA Oecologia, 177, 53-63. DOI:10.1007/s00442-014-3119-zURLPMID:25407618 [本文引用: 1] We conducted a 7-year field study at two oak-dominated forest sites which differ in their atmospheric N deposition to test the hypothesis that red oak regeneration failure in the upper Midwestern US forests, at least in part, results from increased N load. The sites are located in Swallow Cliffs (SC) in Cook County, Illinois, and Indiana Dunes National Lakeshore (IDNL) in Porter County, Indiana. Annual wet NO3(-) deposition for the 22 years immediately prior to the experiments was significantly higher in IDNL than in the SC site. Results from common garden experiments showed that oak seedling biomass was 60% lower at IDNL compared with SC, but there was little site effect on growth of maple seedlings. Experimental N addition also resulted in a 45% decrease in the total biomass of the oak seedlings at SC, but had no significant effect on the biomass at IDNL. Maple seedlings responded little to experimental fertilization. The growth rate of mature oak trees was also lower at IDNL but to a much smaller extent than that of seedlings. Maple trees did not significantly differ between sites. We conclude that: (1) chronic N load adversely affects seedling performance of red oak, but not sugar maple, in these temperate forests; and (2) the seedling establishment phase rather than the adult tree is the likely target stage for this adverse effect of N loading. The exact mechanisms for the differential effects of N on these co-occurring species are not clear, but different plasticity in fractional biomass and N allocation to the leaves might be involved.
BorerET, HarpoleWS, AdlerPB, LindEM, OrrockJL, SeabloomEW, SmithMD (2014). Finding generality in ecology: a model for globally distributed experiments Methods in Ecology and Evolution, 5, 65-73. DOI:10.1111/2041-210X.12125URL [本文引用: 1] Advancing the field of ecology relies on understanding generalities and developing theories based on empirical and functional relationships that integrate across organismal to global spatial scales and span temporal scales. Significant advances in predicting responses of ecological communities to globally extensive anthropogenic perturbations, for example, require understanding the role of environmental context in determining outcomes, which in turn requires standardized experiments across sites and regions. Distributed collaborative experiments can lead to high-impact advances that would otherwise be unachievable. Here, we provide specific advice and considerations relevant to researchers interested in employing this emerging approach using as a case study our experience developing and running the Nutrient Network, a globally distributed experimental network (currently >75 sites in 17 countries) that arose from a grassroots, cooperative research effort. We clarify the design, goals and function of the Nutrient Network as a model to empower others in the scientific community to employ distributed experiments to advance our predictive understanding of global-scale ecological trends and responses. Our experiences to date demonstrate that globally distributed experimental science need not be prohibitively expensive or time-consuming on a per capita basis and is not limited to senior scientists or countries where science is well funded. While distributed experiments are not a panacea for understanding ecological systems, they can substantially complement existing approaches.
BraunS, SchindlerC, RihmB (2017). Growth trends of beech and Norway spruce in Switzerland: the role of nitrogen deposition, ozone, mineral nutrition and climate Science of the Total Environment, 599-600, 637-646. [本文引用: 1]
CamareroJJ, CarrerM (2017). Bridging long-term wood functioning and nitrogen deposition to better understand changes in tree growth and forest productivity Tree Physiology, 37, 1-3. DOI:10.1093/treephys/tpw111URLPMID:28175915 [本文引用: 1]
?apekP, ManzoniS, Ka?tovskáE, WildB, DiákováK, BártaJ, SchneckerJ, BiasiC, MartikainenPJ, AlvesRJE, GuggenbergerG, GentschN, HugeliusG, PalmtagJ, MikuttaR, ShibistovaO, UrichT, SchleperC, RichterA, ?antr??kováH (2018). A plant-microbe interaction framework explaining nutrient effects on primary production Nature Ecology & Evolution, 2, 1588-1596. DOI:10.1038/s41559-018-0662-8URLPMID:30201963 [本文引用: 4] In most terrestrial ecosystems, plant growth is limited by nitrogen and phosphorus. Adding either nutrient to soil usually affects primary production, but their effects can be positive or negative. Here we provide a general stoichiometric framework for interpreting these contrasting effects. First, we identify nitrogen and phosphorus limitations on plants and soil microorganisms using their respective nitrogen to phosphorus critical ratios. Second, we use these ratios to show how soil microorganisms mediate the response of primary production to limiting and non-limiting nutrient addition along a wide gradient of soil nutrient availability. Using a meta-analysis of 51 factorial nitrogen-phosphorus fertilization experiments conducted across multiple ecosystems, we demonstrate that the response of primary production to nitrogen and phosphorus additions is accurately predicted by our stoichiometric framework. The only pattern that could not be predicted by our original framework suggests that nitrogen has not only a structural function in growing organisms, but also a key role in promoting plant and microbial nutrient acquisition. We conclude that this stoichiometric framework offers the most parsimonious way to interpret contrasting and, until now, unresolved responses of primary production to nutrient addition in terrestrial ecosystems.
CarlsonCA, BurkhartHE, Lee AllenH, FoxTR (2008). Absolute and relative changes in tree growth rates and changes to the stand diameter distribution of Pinus taeda as a result of midrotation fertilizer applications Canadian Journal of Forest Research, 38, 2063-2071. DOI:10.1139/X08-050URL [本文引用: 1]
ChalotM, BrunA (1998). Physiology of organic nitrogen acquisition by ectomycorrhizal fungi and ectomycorrhizas FEMS Microbiology Reviews, 22, 21-44. DOI:10.1111/j.1574-6976.1998.tb00359.xURLPMID:9640645 [本文引用: 1] Ectomycorrhizal fungi are symbiotically associated microorganisms which ecological importance has been repeatedly demonstrated. There has been a considerable amount of research aimed at assessing the ability of ectomycorrhizal fungi and ectomycorrhizas to utilize organic nitrogen sources. The fate of soil proteins, peptides and amino acids has been studied from a number of perspectives. Exocellular hydrolytic enzymes have been detected and characterized in a number of ectomycorrhizal and ericoid fungi. Studies on amino acid transport through the plasma membrane have demonstrated the ability of ectomycorrhizal fungi to take up the products of proteolytic activities. Investigations on intracellular metabolism of amino acids have allowed the identification of the metabolic pathways involved. Possible intracellular compartmentation of amino acids will be examined by immunocytochemistry. Further translocation of amino acids in symbiotic tissues has been established by experiments using isotopic tracers, although the exact nature of the nitrogenous compounds transferred at the symbiotic interface remained unclear. One of the main future challenges in the physiology of organic nitrogen acquisition is to determine the nature, the regulation and the location of N-compound transporters at the soil-fungus and fungus-plant interfaces. The molecular approach which is just emerging in this particular research area will greatly improve our knowledge. Future research should also address the extent of competition between different ectomycorrhizal species and between different microbial populations for organic nitrogen.
Chapin IIIFS, VitousekPM, van CleveK (1986). The nature of nutrient limitation in plant communities The American Naturalist, 127, 48-58. DOI:10.1086/284466URL [本文引用: 1]
ChenD, LiJ, LanZ, HuS, BaiY (2016). Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment Functional Ecology, 30, 658-669. DOI:10.1111/fec.2016.30.issue-4URL [本文引用: 1]
ClevelandCC, TownsendAR, SchimelDS, FisherH, HowarthRW, HedinLO, PerakisSS, LattyEF, von FischerJC, ElseroadA, WassonMF (1999). Global patterns of terrestrial biological nitrogen (N2) fixation in natural ecosystems Global Biogeochemical Cycles, 13, 623-645. DOI:10.1029/1999GB900014URL [本文引用: 1]
ColeyPD, BryantJP, Chapin IIIFS (1985). Resource availability and plant antiherbivore defense Science, 230, 895-899. DOI:10.1126/science.230.4728.895URLPMID:17739203 [本文引用: 1] The degree of herbivory and the effectiveness of defense varies widely among plant species. Resource availability in the environment is proposed as the major determinant of both the amount and type of plant defense. When resource are limited, plants with inherently slow growth are favored over those with fast growth rates; slow rates in turn favor large investments in antiherbivore defenses. Leaf lifetime, also determined by resource availability, affects the relative advantages of defenses with different turnover rates. Relative limitation of different resources also constrains the types of defenses. The proposals are compared with other theories on the evolution of plant defenses.
CornelissenJHC, LavorelS, GarnierE, DiazS, BuchmannN, GurvichDE, ReichPB, ter SteegeH, MorganHD, van der HeijdenMGA, PausasJG, PoorterH (2003). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide Australian Journal of Botany, 51, 335-380. DOI:10.1071/BT02124URL [本文引用: 1]
CrowleyKF, McNeilBE, LovettGM, CanhamCD, DriscollCT, RustadLE, DennyE, HallettRA, ArthurMA, BoggsJL, GoodaleCL, KahlJS, McNultySG, OllingerSV, PardoLH, SchabergPG, StoddardJL, WeandMP, WeathersKC (2012). Do nutrient limitation patterns shift from nitrogen toward phosphorus with increasing nitrogen deposition across the northeastern United States? Ecosystems, 15, 940-957. DOI:10.1007/s10021-012-9550-2URL [本文引用: 1] Atmospheric nitrogen (N) deposition is altering biogeochemical cycling in forests and interconnected lakes of the northeastern US, and may shift nutrient limitation from N toward other essential elements, such as phosphorus (P). Whether this shift is occurring relative to N deposition gradients across the northeastern US has not been investigated. We used datasets for the northeastern US and the Adirondack sub-region to evaluate whether P limitation is increasing where N deposition is high at two geographic scales, based on N:P mass ratios. Using a model-selection approach, we determined that foliar N for dominant tree species and lake dissolved inorganic N (DIN) increased coincident with increasing N deposition, independent of relationships between foliar N or lake DIN and precipitation or temperature. Foliar P also increased with N deposition across the northeastern US for seven of eight deciduous species, but changed less across the Adirondacks. Foliar N:P therefore declined at the highest levels of N deposition for most deciduous species across the region (remaining nearly constant for most conifers and increasing only for black cherry and hemlock), but increased across all species in the Adirondacks. Ratios between DIN and total P (DIN:TP) in lakes were unrelated to N deposition regionally but increased across the Adirondacks. Thus, nutrient limitation patterns shifted from N toward P for dominant trees, and further toward P for predominantly P-limited lakes, at the sub-regional but not regional scale. For the northeastern US overall, accumulated N deposition may be insufficient to drive nutrient limitation from N toward P; alternatively, elements other than P (for example, calcium, magnesium) may become limiting as N accumulates. The consistent Adirondack foliar and lake response could provide early indication of shifts toward P limitation within the northeastern US, and together with regional patterns, suggests that foliar chemistry could be a predictor of lake chemistry in the context of N deposition across the region.
DaiZ, SuW, ChenH, BarberánA, ZhaoH, YuM, YuL, BrookesPC, SchadtCW, ChangSX, XuJ (2018). Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of Actinobacteria and Proteobacteria in agro-ecosystems across the globe Global Change Biology, 24, 3452-3461. DOI:10.1111/gcb.14163URLPMID:29645398 [本文引用: 1] Long-term elevated nitrogen (N) input from anthropogenic sources may cause soil acidification and decrease crop yield, yet the response of the belowground microbial community to long-term N input alone or in combination with phosphorus (P) and potassium (K) is poorly understood. We explored the effect of long-term N and NPK fertilization on soil bacterial diversity and community composition using meta-analysis of a global dataset. Nitrogen fertilization decreased soil pH, and increased soil organic carbon (C) and available N contents. Bacterial taxonomic diversity was decreased by N fertilization alone, but was increased by NPK fertilization. The effect of N fertilization on bacterial diversity varied with soil texture and water management, but was independent of crop type or N application rate. Changes in bacterial diversity were positively related to both soil pH and organic C content under N fertilization alone, but only to soil organic C under NPK fertilization. Microbial biomass C decreased with decreasing bacterial diversity under long-term N fertilization. Nitrogen fertilization increased the relative abundance of Proteobacteria and Actinobacteria, but reduced the abundance of Acidobacteria, consistent with the general life history strategy theory for bacteria. The positive correlation between N application rate and the relative abundance of Actinobacteria indicates that increased N availability favored the growth of Actinobacteria. This first global analysis of long-term N and NPK fertilization that differentially affects bacterial diversity and community composition provides a reference for nutrient management strategies for maintaining belowground microbial diversity in agro-ecosystems worldwide.
DeForestJL, SnellRS (2020). Tree growth response to shifting soil nutrient economy depends on mycorrhizal associations New Phytologist, 225, 2557-2566. DOI:10.1111/nph.16299URLPMID:31677163 [本文引用: 8] The mycorrhizal-associated nutrient economy hypothesis proposes a strong connection between plant and fungal traits and the dominant form of soil nutrients. If true, then shifting from an organic to an inorganic nutrient economy should benefit arbuscular mycorrhizal (AM) trees because they are more suited to acquiring inorganic forms of nutrients and have limited decomposing capabilities when compared with ectomycorrhizal (ECM) trees. An inorganic nutrient economy was experimentally promoted by applying inorganic phosphorus (P) fertiliser and/or elevating soil pH with lime in three Allegheny Plateau mixed-mesophytic forests. Trees were measured over seven growing seasons to determine how growth responded to the treatments based on mycorrhizal association. AM-associated trees showed increased growth in response to increased inorganic nutrients, but ECM tree growth was suppressed when compared with the control. We also observed that understory and mid-story trees responded to the treatments, but large overstory trees showed no significant growth response. Results support the hypothesis that AM trees respond positively to an inorganic nutrient economy. While raising pH in acidic soils can be detrimental to ECM tree growth, the exact mechanism for this response is unclear.
DengM, LiuL, JiangL, LiuW, WangX, LiS, YangS, WangB (2018). Ecosystem scale trade-off in nitrogen acquisition pathways Nature Ecology & Evolution, 2, 1724-1734. DOI:10.1038/s41559-018-0677-1URLPMID:30250156 [本文引用: 2] The nitrogen (N) cycle in terrestrial ecosystems is strongly influenced by resorption before litter fall and by mineralization after litter fall. Although both resorption and mineralization make N available to plants and are influenced by climate, their linkage in a changing environment remains largely unknown. Here, our synthesis study shows that, at the global scale, increasing N-resorption efficiency negatively affects the N-mineralization rate. As temperature and precipitation increase, the increasing rates of N cycling closely correspond to a shift from the more conservative resorption pathway to the mineralization pathway. Furthermore, ecosystems with faster N-cycle rates support plant species that have higher foliar N:P ratios and microbial communities with lower fungi:bacteria ratios. Our study shows an ecosystem scale trade-off in N-acquisition pathways. We propose that incorporating the dynamic interaction between N resorption and N mineralization into Earth system models will improve the simulation of nutrient constraints on ecosystem productivity.
DuE, FangJ (2014). Weak growth response to nitrogen deposition in an old-growth boreal forest Ecosphere, 5, 109. DOI: 10.1890/es14-00109.1. [本文引用: 4]
DuEZ, TerrerC, PellegriniAFA, Ahlstr?mA, van LissaCJ, ZhaoX, XiaN, WuXH, JacksonRB (2020). Global patterns of terrestrial nitrogen and phosphorus limitation Nature Geoscience, 13, 221-226. DOI:10.1038/s41561-019-0530-4URL [本文引用: 1]
DynarskiKA, HoultonBZ (2018). Nutrient limitation of terrestrial free-living nitrogen fixation New Phytologist, 217, 1050-1061. DOI:10.1111/nph.14905URLPMID:29165820 [本文引用: 1] Nitrogen (N) fixation by free-living bacteria is a primary N input pathway in many ecosystems and sustains global plant productivity. Uncertainty exists over the importance of N, phosphorus (P) and molybdenum (Mo) availability in controlling free-living N fixation rates. Here, we investigate the geographic occurrence and variability of nutrient constraints to free-living N fixation in the terrestrial biosphere. We compiled data from studies measuring free-living N fixation in response to N, P and Mo fertilizers. We used meta-analysis to quantitatively determine the extent to which N, P and Mo stimulate or suppress N fixation, and if environmental variables influence the degree of nutrient limitation of N fixation. Across our compiled dataset, free-living N fixation is suppressed by N fertilization and stimulated by Mo fertilization. Additionally, free-living N fixation is stimulated by P additions in tropical forests. These findings suggest that nutrient limitation is an intrinsic property of the biochemical demands of N fixation, constraining free-living N fixation in the terrestrial biosphere. These findings have implications for understanding the causes and consequences of N limitation in coupled nutrient cycles, as well as modeling and forecasting nutrient controls over carbon-climate feedbacks.
ElserJJ, BrackenMES, ClelandEE, GrunerDS, HarpoleWS, HillebrandH, NgaiJT, SeabloomEW, ShurinJB, SmithJE (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems Ecology Letters, 10, 1135-1142. DOI:10.1111/j.1461-0248.2007.01113.xURLPMID:17922835 [本文引用: 4] The cycles of the key nutrient elements nitrogen (N) and phosphorus (P) have been massively altered by anthropogenic activities. Thus, it is essential to understand how photosynthetic production across diverse ecosystems is, or is not, limited by N and P. Via a large-scale meta-analysis of experimental enrichments, we show that P limitation is equally strong across these major habitats and that N and P limitation are equivalent within both terrestrial and freshwater systems. Furthermore, simultaneous N and P enrichment produces strongly positive synergistic responses in all three environments. Thus, contrary to some prevailing paradigms, freshwater, marine and terrestrial ecosystems are surprisingly similar in terms of N and P limitation.
FengJG, ZhuB (2019). A global meta-analysis of soil respiration and its components in response to phosphorus addition Soil Biology & Biochemistry, 135, 38-47. DOI:10.1016/j.soilbio.2019.04.008URL [本文引用: 3]
FisherJB, MalhiY, TorresIC, MetcalfeDB, van de WegMJ, MeirP, Silva-EspejoJE, HuascoWH (2013). Nutrient limitation in rainforests and cloud forests along a 3000-m elevation gradient in the Peruvian Andes Oecologia, 172, 889-902. DOI:10.1007/s00442-012-2522-6URL [本文引用: 2] We report results from a large-scale nutrient fertilization experiment along a "megadiverse" (154 unique species were included in the study) 3,000-m elevation transect in the Peruvian Andes and adjacent lowland Amazonia. Our objectives were to test if nitrogen (N) and phosphorus (P) limitation shift along this elevation gradient, and to determine how an alleviation of nutrient limitation would manifest in ecosystem changes. Tree height decreased with increasing elevation, but leaf area index (LAI) and diameter at breast height (DBH) did not vary with elevation. Leaf N:P decreased with increasing elevation (from 24 at 200 m to 11 at 3,000 m), suggesting increased N limitation and decreased P limitation with increasing elevation. After 4 years of fertilization (N, P, N + P), plots at the lowland site (200 m) fertilized with N + P showed greater relative growth rates in DBH than did the control plots; no significant differences were evident at the 1,000 m site, and plots fertilized with N at the highest elevation sites (1,500, 3,000 m) showed greater relative growth rates in DBH than did the control plots, again suggesting increased N constraint with elevation. Across elevations in general N fertilization led to an increase in microbial respiration, while P and N + P addition led to an increase in root respiration and corresponding decrease in hyphal respiration. There was no significant canopy response (LAI, leaf nutrients) to fertilization, suggesting that photosynthetic capacity was not N or P limited in these ecosystems. In sum, our study significantly advances ecological understanding of nutrient cycling and ecosystem response in a region where our collective knowledge and data are sparse: we demonstrate N limitation in high elevation tropical montane forests, N and P co-limitation in lowland Amazonia, and a nutrient limitation response manifested not in canopy changes, but rather in stem and belowground changes.
FleischerK, RebelKT, van der MolenMK, ErismanJW, WassenMJ, van LoonEE, MontagnaniL, GoughCM, HerbstM, JanssensIA, GianelleD, DolmanAJ (2013). The contribution of nitrogen deposition to the photosynthetic capacity of forests Global Biogeochemical Cycles, 27, 187-199. DOI:10.1002/gbc.20026URL [本文引用: 1] Global terrestrial carbon (C) sequestration has increased over the last few decades. The drivers of carbon sequestration, the geographical spread and magnitude of this sink are however hotly debated. Photosynthesis determines the total C uptake of terrestrial ecosystems and is a major flux of the global C balance. We contribute to the discussion on enhanced C sequestration by analyzing the influence of nitrogen (N) deposition on photosynthetic capacity (A(max)) of forest canopies. Eddy covariance measurements of net exchange of carbon provide estimates of gross primary production, from which A(max) is derived with a novel approach. Canopy A(max) is combined with modeled N deposition, environmental variables and stand characteristics to study the relative effects on A(max) for a unique global data set of 80 forest FLUXNET sites. Canopy A(max) relates positively to N deposition for evergreen needleleaf forests below an observed critical load of similar to 8 kgN ha(-1) yr(-1), with a slope of 2.0 +/- 0.4 (S.E.) mu mol CO2 m(-2) s(-1) per 1 kgN ha(-1) yr(-1). Above this threshold canopy A(max) levels off, exhibiting a saturating response in line with the N saturation hypothesis. Climate effects on canopy A(max) cannot be separated from the effect of N deposition due to considerable covariation. For deciduous broadleaf forests and forests in the temperate (-continental) climate zones, the analysis shows the N deposition effect to be either small or absent. Leaf area index and foliar N concentration are positively but weakly related to A(max). We conclude that flux tower measurements of C fluxes provide valuable data to study physiological processes at the canopy scale. Future efforts need to be directed toward standardizing measures N cycling and pools within C monitoring networks to gain a better understanding of C and N interactions, and to disentangle the role of climate and N deposition in forest ecosystems.
GoswamiS, FiskMC, VadeboncoeurMA, Garrison-JohnstonM, YanaiRD, FaheyTJ (2018). Phosphorus limitation of aboveground production in northern hardwood forests Ecology, 99, 438-449. DOI:10.1002/ecy.2100URLPMID:29205288 [本文引用: 4] Forest productivity on glacially derived soils with weatherable phosphorus (P) is expected to be limited by nitrogen (N), according to theories of long-term ecosystem development. However, recent studies and model simulations based on resource optimization theory indicate that productivity can be co-limited by N and P. We conducted a full factorial N x P fertilization experiment in 13 northern hardwood forest stands of three age classes in central New Hampshire, USA, to test the hypothesis that forest productivity is co-limited by N and P. We also asked whether the response of productivity to N and P addition differs among species and whether differential species responses contribute to community-level co-limitation. Plots in each stand were fertilized with 30 kg N.ha(-1) .yr(-1) , 10 kg P.ha(-1) .yr(-1) , N + P, or neither nutrient (control) for four growing seasons. The productivity response to treatments was assessed using per-tree annual relative basal area increment (RBAI) as an index of growth. RBAI responded significantly to P (P = 0.02) but not to N (P = 0.73). However, evidence for P limitation was not uniform among stands. RBAI responded to P fertilization in mid-age (P = 0.02) and mature (P = 0.07) stands, each taken as a group, but was greatest in N-fertilized plots of two stands in these age classes, and there was no significant effect of P in the young stands. Both white birch (Betula papyrifera Marsh.) and beech (Fagus grandifolia Ehrh.) responded significantly to P; no species responded significantly to N. We did not find evidence for N and P co-limitation of tree growth. The response to N + P did not differ from that to P alone, and there was no significant N x P interaction (P = 0.68). Our P limitation results support neither the N limitation prediction of ecosystem theory nor the N and P co-limitation prediction of resource optimization theory, but could be a consequence of long-term anthropogenic N deposition in these forests. Inconsistencies in response to P suggest that successional status and variation in site conditions influence patterns of nutrient limitation and recycling across the northern hardwood forest landscape.
HarpoleWS, NgaiJT, ClelandEE, SeabloomEW, BorerET, BrackenMES, ElserJJ, GrunerDS, HillebrandH, ShurinJB, SmithJE (2011). Nutrient co-limitation of primary producer communities Ecology Letters, 14, 852-862. DOI:10.1111/j.1461-0248.2011.01651.xURL [本文引用: 2] Synergistic interactions between multiple limiting resources are common, highlighting the importance of co-limitation as a constraint on primary production. Our concept of resource limitation has shifted over the past two decades from an earlier paradigm of single-resource limitation towards concepts of co-limitation by multiple resources, which are predicted by various theories. Herein, we summarise multiple-resource limitation responses in plant communities using a dataset of 641 studies that applied factorial addition of nitrogen (N) and phosphorus (P) in freshwater, marine and terrestrial systems. We found that more than half of the studies displayed some type of synergistic response to N and P addition. We found support for strict definitions of co-limitation in 28% of the studies: i.e. community biomass responded to only combined N and P addition, or to both N and P when added separately. Our results highlight the importance of interactions between N and P in regulating primary producer community biomass and point to the need for future studies that address the multiple mechanisms that could lead to different types of co-limitation.
HeNP, ZhangJH, LiuCC, XuL, ChenZ, LiuY, WangRL, ZhaoN, XuZW, TianJ, WangQ, ZhuJX, LiY, HouJH, YuGR (2018). Patterns and influencing factors of traits in forest ecosystems: synthesis and perspectives on the synthetic investigation from the north-east transect of eastern China (NETEC) Acta Ecologica Sinica, 38, 6359-6382. [本文引用: 1]
HedinLO (2004). Global organization of terrestrial plant- nutrient interactions Proceedings of the National Academy of Sciences of the United States of America, 101, 10849-10850. DOI:10.1073/pnas.0404222101URLPMID:15263081 [本文引用: 1]
HerbertDA, FownesJH (1995). Phosphorus limitation of forest leaf-area and net primary production on a highly weathered soil Biogeochemistry, 29, 223-235. [本文引用: 1]
H?gbergP, FanH, QuistM, BinkleyD, TammCO (2006). Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest Global Change Biology, 12, 489-499. [本文引用: 3]
HopmansP, ElmsSR (2013). Impact of defoliation by Essigella californica on the growth of mature Pinus radiata and response to N, P and S fertilizer Forest Ecology and Management, 289, 190-200. [本文引用: 1]
HouEQ, LuoYQ, KuangYW, ChenCR, LuXK, JiangLF, LuoXZ, WenDZ (2020). Global meta-analysis shows pervasive phosphorus limitation of aboveground plant production in natural terrestrial ecosystems Nature Communications, 11, 637. DOI: 10.1038/s41467-020-14492-w. URLPMID:32005808 [本文引用: 2]
HoultonBZ, WangYP, VitousekPM, FieldCB (2008). A unifying framework for dinitrogen fixation in the terrestrial biosphere Nature, 454, 327-334. DOI:10.1038/nature07028URLPMID:18563086 [本文引用: 1] Dinitrogen (N(2)) fixation is widely recognized as an important process in controlling ecosystem responses to global environmental change, both today and in the past; however, significant discrepancies exist between theory and observations of patterns of N(2) fixation across major sectors of the land biosphere. A question remains as to why symbiotic N(2)-fixing plants are more abundant in vast areas of the tropics than in many of the mature forests that seem to be nitrogen-limited in the temperate and boreal zones. Here we present a unifying framework for terrestrial N(2) fixation that can explain the geographic occurrence of N(2) fixers across diverse biomes and at the global scale. By examining trade-offs inherent in plant carbon, nitrogen and phosphorus capture, we find a clear advantage to symbiotic N(2) fixers in phosphorus-limited tropical savannas and lowland tropical forests. The ability of N(2) fixers to invest nitrogen into phosphorus acquisition seems vital to sustained N(2) fixation in phosphorus-limited tropical ecosystems. In contrast, modern-day temperatures seem to constrain N(2) fixation rates and N(2)-fixing species from mature forests in the high latitudes. We propose that an analysis that couples biogeochemical cycling and biophysical mechanisms is sufficient to explain the principal geographical patterns of symbiotic N(2) fixation on land, thus providing a basis for predicting the response of nutrient-limited ecosystems to climate change and increasing atmospheric CO(2).
Ibá?ezI, ZakDR, BurtonAJ, PregitzerKS (2016). Chronic nitrogen deposition alters tree allometric relationships: implications for biomass production and carbon storage Ecological Applications, 26, 913-925. DOI:10.1890/15-0883URLPMID:27411260 [本文引用: 1] As increasing levels of nitrogen (N) deposition impact many terrestrial ecosystems, understanding the potential effects of higher N availability is critical for forecasting tree carbon allocation patterns and thus future forest productivity. Most regional estimates of forest biomass apply allometric equations, with parameters estimated from a limited number of studies, to forest inventory data (i.e., tree diameter). However most of these allometric equations cannot account for potential effects of increased N availability on biomass allocation patterns. Using 18 yr of tree diameter, height, and mortality data collected for a dominant tree species (Acer saccharum) in an atmospheric N deposition experiment, we evaluated how greater N availability affects allometric relationships in this species. After taking into account site and individual variability, our results reveal significant differences in allometric parameters between ambient and experimental N deposition treatments. Large trees under experimental N deposition reached greater heights at a given diameter; moreover, their estimated maximum height (mean +/- standard deviation: 33.7 +/- 0.38 m) was significantly higher than that estimated under the ambient condition (31.3 +/- 0.31 m). Within small tree sizes (5-10 cm diameter) there was greater mortality under experimental N deposition, whereas the relative growth rates of small trees were greater under experimental N deposition. Calculations of stemwood biomass using our parameter estimates for the diameter-height relationship indicated the potential for significant biases in these estimates (~2.5%), with under predictions of stemwood biomass averaging 4 Mg/ha lower if ambient parameters were to be used to estimate stem biomass of trees in the experimental N deposition treatment. As atmospheric N deposition continues to increase into the future, ignoring changes in tree allometry will contribute to the uncertainty associated with aboveground carbon storage estimates across a forest with a large geographic distribution in eastern North America.
JiangJ, WangY, YangY, YuM, WangC, YanJ (2019) Interactive effects of nitrogen and phosphorus additions on plant growth vary with ecosystem type Plant and Soil, 440, 523-537. [本文引用: 2]
JiangL, TianD, MaS, ZhouX, XuL, ZhuJ, JingX, ZhengC, ShenH, ZhouZ, LiY, ZhuB, FangJ (2018). The response of tree growth to nitrogen and phosphorus additions in a tropical montane rainforest Science of the Total Environment, 618, 1064-1070. [本文引用: 7]
LeBauerDS, TresederKK (2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed Ecology, 89, 371-379. DOI:10.1890/06-2057.1URLPMID:18409427 [本文引用: 4] Our meta-analysis of 126 nitrogen addition experiments evaluated nitrogen (N) limitation of net primary production (NPP) in terrestrial ecosystems. We tested the hypothesis that N limitation is widespread among biomes and influenced by geography and climate. We used the response ratio (R approximately equal ANPP(N)/ANPP(ctrl)) of aboveground plant growth in fertilized to control plots and found that most ecosystems are nitrogen limited with an average 29% growth response to nitrogen (i.e., R = 1.29). The response ratio was significant within temperate forests (R = 1.19), tropical forests (R = 1.60), temperate grasslands (R = 1.53), tropical grasslands (R = 1.26), wetlands (R = 1.16), and tundra (R = 1.35), but not deserts. Eight tropical forest studies had been conducted on very young volcanic soils in Hawaii, and this subgroup was strongly N limited (R = 2.13), which resulted in a negative correlation between forest R and latitude. The degree of N limitation in the remainder of the tropical forest studies (R = 1.20) was comparable to that of temperate forests, and when the young Hawaiian subgroup was excluded, forest R did not vary with latitude. Grassland response increased with latitude, but was independent of temperature and precipitation. These results suggest that the global N and C cycles interact strongly and that geography can mediate ecosystem response to N within certain biome types.
LiY, NiuS, YuG (2016). Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: a meta-analysis Global Change Biology, 22, 934-943. DOI:10.1111/gcb.13125URLPMID:26463578 [本文引用: 5] Nitrogen (N) and phosphorus (P), either individually or in combination, have been demonstrated to limit biomass production in terrestrial ecosystems. Field studies have been extensively synthesized to assess global patterns of N impacts on terrestrial ecosystem processes. However, to our knowledge, no synthesis has been done so far to reveal global patterns of P impacts on terrestrial ecosystems, especially under different nitrogen (N) levels. Here, we conducted a meta-analysis of impacts of P addition, either alone or with N addition, on aboveground (AGB) and belowground biomass production (BGB), plant and soil P concentrations, and N : P ratio in terrestrial ecosystems. Overall, our meta-analysis quantitatively confirmed existing notions: (i) colimitation of N and P on biomass production and (ii) more P limitation in tropical forest than other ecosystems. More importantly, our analysis revealed new findings: (i) P limitation on biomass production was aggravated by N enrichment and (ii) plant P concentration was a better indicator of P limitation than soil P availability. Specifically, P addition increased AGB and BGB by 34% and 13%, respectively. The effect size of P addition on biomass production was larger in tropical forest than grassland, wetland, and tundra and varied with P fertilizer forms, P addition rates, or experimental durations. The P-induced increase in biomass production and plant P concentration was larger under elevated than ambient N. Our findings suggest that the global limitation of P on biomass production will become severer under increasing N fertilizer and deposition in the future.
LiY, TianD, YangH, NiuS (2018). Size-dependent nutrient limitation of tree growth from subtropical to cold temperate forests Functional Ecology, 32, 95-105. [本文引用: 9]
LiuXY, DuEZ, XuLC, ShenHH, FangJY, HuHF (2015). Response of tree growth to nitrogen addition in aLarix gmelinii primitive forest Chinese Journal of Plant Ecology, 39, 433-441. [本文引用: 4]
LuM, YangY, LuoY, FangC, ZhouX, ChenJ, YangX, LiB (2011a). Responses of ecosystem nitrogen cycle to nitrogen addition: a meta-analysis New Phytologist, 189, 1040-1050. DOI:10.1111/j.1469-8137.2010.03563.xURLPMID:21138438 [本文引用: 1] * Anthropogenic nitrogen (N) addition may substantially alter the terrestrial N cycle. However, a comprehensive understanding of how the ecosystem N cycle responds to external N input remains elusive. * Here, we evaluated the central tendencies of the responses of 15 variables associated with the ecosystem N cycle to N addition, using data extracted from 206 peer-reviewed papers. * Our results showed that the largest changes in the ecosystem N cycle caused by N addition were increases in soil inorganic N leaching (461%), soil NO(3)(-) concentration (429%), nitrification (154%), nitrous oxide emission (134%), and denitrification (84%). N addition also substantially increased soil NH(4)+ concentration (47%), and the N content in belowground (53%) and aboveground (44%) plant pools, leaves (24%), litter (24%) and dissolved organic N (21%). Total N content in the organic horizon (6.1%) and mineral soil (6.2%) slightly increased in response to N addition. However, N addition induced a decrease in microbial biomass N by 5.8%. * The increases in N effluxes caused by N addition were much greater than those in plant and soil pools except soil NO(3)(-), suggesting a leaky terrestrial N system.
LuM, ZhouX, LuoY, YangY, FangC, ChenJ, LiB (2011b). Minor stimulation of soil carbon storage by nitrogen addition: a meta-analysis Agriculture, Ecosystems & Environment, 140, 234-244. [本文引用: 2]
LuX, VitousekPM, MaoQ, GilliamFS, LuoY, ZhouG, ZouX, BaiE, ScanlonTM, HouE, MoJ (2018). Plant acclimation to long-term high nitrogen deposition in an N-rich tropical forest Proceedings of the National Academy of Sciences of the United States of America, 115, 5187-5192. DOI:10.1073/pnas.1720777115URLPMID:29717039 [本文引用: 4] Anthropogenic nitrogen (N) deposition has accelerated terrestrial N cycling at regional and global scales, causing nutrient imbalance in many natural and seminatural ecosystems. How added N affects ecosystems where N is already abundant, and how plants acclimate to chronic N deposition in such circumstances, remains poorly understood. Here, we conducted an experiment employing a decade of N additions to examine ecosystem responses and plant acclimation to added N in an N-rich tropical forest. We found that N additions accelerated soil acidification and reduced biologically available cations (especially Ca and Mg) in soils, but plants maintained foliar nutrient supply at least in part by increasing transpiration while decreasing soil water leaching below the rooting zone. We suggest a hypothesis that cation-deficient plants can adjust to elevated N deposition by increasing transpiration and thereby maintaining nutrient balance. This result suggests that long-term elevated N deposition can alter hydrological cycling in N-rich forest ecosystems.
LuXK, MoJM, ZhangW, MaoQG, LiuRZ, WangC, WangSH, ZhengMH, MoriT, MaoJH, ZhangYQ, WangYF, HuangJ (2019). Effects of simulated atmospheric nitrogen deposition on forest ecosystems in China: an overview Journal of Tropical and Subtropical Botany, 27, 500-522. [本文引用: 1]
MagnaniF, MencucciniM, BorghettiM, BerbigierP, BerningerF, DelzonS, GrelleA, HariP, JarvisPG, KolariP, KowalskiAS, LankreijerH, LawBE, LindrothA, LoustauD, MancaG, MoncrieffJB, RaymentM, TedeschiV, ValentiniR, GraceJ (2007). The human footprint in the carbon cycle of temperate and boreal forests Nature, 447, 848-850. DOI:10.1038/nature05847URLPMID:17568744 [本文引用: 2] Temperate and boreal forests in the Northern Hemisphere cover an area of about 2 x 10(7) square kilometres and act as a substantial carbon sink (0.6-0.7 petagrams of carbon per year). Although forest expansion following agricultural abandonment is certainly responsible for an important fraction of this carbon sink activity, the additional effects on the carbon balance of established forests of increased atmospheric carbon dioxide, increasing temperatures, changes in management practices and nitrogen deposition are difficult to disentangle, despite an extensive network of measurement stations. The relevance of this measurement effort has also been questioned, because spot measurements fail to take into account the role of disturbances, either natural (fire, pests, windstorms) or anthropogenic (forest harvesting). Here we show that the temporal dynamics following stand-replacing disturbances do indeed account for a very large fraction of the overall variability in forest carbon sequestration. After the confounding effects of disturbance have been factored out, however, forest net carbon sequestration is found to be overwhelmingly driven by nitrogen deposition, largely the result of anthropogenic activities. The effect is always positive over the range of nitrogen deposition covered by currently available data sets, casting doubts on the risk of widespread ecosystem nitrogen saturation under natural conditions. The results demonstrate that mankind is ultimately controlling the carbon balance of temperate and boreal forests, either directly (through forest management) or indirectly (through nitrogen deposition).
MainwaringDB, MaguireDA, PerakisSS (2014). Three-year growth response of young Douglas-fir to nitrogen, calcium, phosphorus, and blended fertilizers in Oregon and Washington Forest Ecology and Management, 327, 178-188. [本文引用: 1]
MaoHR, JinGZ (2017). Impacts of nitrogen addition on net primary productivity in the typical mixed broadleaved-?Korean pine forest Journal of Beijing Forestry University, 39(8), 42-49. [本文引用: 1]
MarkewitzD, FigueiredoRdO, de CarvalhoCJR, DavidsonEA (2012). Soil and tree response to P fertilization in a secondary tropical forest supported by an Oxisol Biology and Fertility of Soils, 48, 665-678. [本文引用: 4]
MarkleinAR, HoultonBZ (2012). Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems New Phytologist, 193, 696-704. DOI:10.1111/j.1469-8137.2011.03967.xURLPMID:22122515 [本文引用: 1] * Biologically essential elements--especially nitrogen (N) and phosphorus (P)--constrain plant growth and microbial functioning; however, human activities are drastically altering the magnitude and pattern of such nutrient limitations on land. Here we examine interactions between N and P cycles of P mineralizing enzyme activities (phosphatase enzymes) across a wide variety of terrestrial biomes. * We synthesized results from 34 separate studies and used meta-analysis to evaluate phosphatase activity with N, P, or NxP fertilization. * Our results show that N fertilization enhances phosphatase activity, from the tropics to the extra-tropics, both on plant roots and in bulk soils. By contrast, P fertilization strongly suppresses rates of phosphatase activity. * These results imply that phosphatase enzymes are strongly responsive to changes in local nutrient cycle conditions. We also show that plant phosphatases respond more strongly to fertilization than soil phosphatases. The tight coupling between N and P provides a mechanism for recent observations of N and P co-limitation on land. Moreover, our results suggest that terrestrial plants and microbes can allocate excess N to phosphatase enzymes, thus delaying the onset of single P limitation to plant productivity as can occur via human modifications to the global N cycle.
McGrathJF, CopelandB, DumbrellIC (2003). Magnitude and duration of growth and wood quality responses to phosphorus and nitrogen in thinned Pinus radiata in southern Western Australia Australian Forestry, 66, 223-230. [本文引用: 1]
NastoMK, Alvarez-ClareS, LekbergY, SullivanBW, TownsendAR, ClevelandCC (2014). Interactions among nitrogen fixation and soil phosphorus acquisition strategies in lowland tropical rain forests Ecology Letters, 17, 1282-1289. URLPMID:25070023 [本文引用: 3]
NastoMK, WinterK, TurnerBL, ClevelandCC (2019). Nutrient acquisition strategies augment growth in tropical N2-fixing trees in nutrient-poor soil and under elevated CO2 Ecology, 100, e02646. DOI: 10.1002/ecy.2646. DOI:10.1002/ecy.2646URLPMID:30714149 [本文引用: 5] Tropical forests play a dominant role in the global carbon (C) cycle, and models predict increases in tropical net primary productivity (NPP) and C storage in response to rising atmospheric carbon dioxide (CO2 ) concentrations. The extent to which increasing CO2 will enhance NPP depends in part on the availability of nitrogen (N) and phosphorus (P) to support growth. Some tropical trees can potentially overcome nutrient limitation by acquiring N via symbiotic dinitrogen (N2 ) fixation, which may provide a benefit in acquiring P via investment in N-rich phosphatase enzymes or arbuscular mycorrhizal (AM) fungi. We conducted a seedling experiment to investigate the effects of elevated CO2 and soil nutrient availability on the growth of two N2 -fixing and two non-N2 -fixing tropical tree species. We hypothesized that under elevated CO2 and at low nutrient availability (i.e., low N and P), N2 fixers would have higher growth rates than non-N2 fixers because N2 fixers have a greater capacity to acquire both N and P. We also hypothesized that differences in growth rates between N2 fixers and non-N2 fixers would decline as nutrient availability increases because N2 fixers no longer have an advantage in nutrient acquisition. We found that the N2 fixers had higher growth rates than the non-N2 fixers under elevated CO2 and at low nutrient availability, and that the difference in growth rates between the N2 and non-N2 fixers declined as nutrient availability increased, irrespective of CO2 . Overall, N2 fixation, root phosphatase activity, and AM colonization decreased with increasing nutrient availability, and increased under elevated CO2 at low nutrient availability. Further, AM colonization was positively related to the growth of the non-N2 fixers, whereas both N2 fixation and root phosphatase activity were positively related to the growth of the N2 fixers. Though our results indicate all four tree species have the capacity to up- or down-regulate nutrient acquisition to meet their stoichiometric demands, the greater capacity for the N2 fixers to acquire both N and P may enable them to overcome nutritional constraints to NPP under elevated CO2 , with implications for the response of tropical forests to future environmental change.
NoguchiK, NagakuraJ, Kon?pkaB, SakataT, KanekoS, TakahashiM (2013). Fine-root dynamics in sugi (Cryptomeria japonica) under manipulated soil nitrogen conditions Plant and Soil, 364, 159-169. [本文引用: 1]
NorbyRJ, de KauweMG, DominguesTF, DuursmaRA, EllsworthDS, GollDS, LapolaDM, LuusKA, MacKenzieAR, MedlynBE, PavlickR, RammigA, SmithB, ThomasR, ThonickeK, WalkerAP, YangX, ZaehleS (2016). Model-data synthesis for the next generation of forest free-air CO2 enrichment (FACE) experiments New Phytologist, 209, 17-28. DOI:10.1111/nph.13593URLPMID:26249015 [本文引用: 1] The first generation of forest free-air CO2 enrichment (FACE) experiments has successfully provided deeper understanding about how forests respond to an increasing CO2 concentration in the atmosphere. Located in aggrading stands in the temperate zone, they have provided a strong foundation for testing critical assumptions in terrestrial biosphere models that are being used to project future interactions between forest productivity and the atmosphere, despite the limited inference space of these experiments with regards to the range of global ecosystems. Now, a new generation of FACE experiments in mature forests in different biomes and over a wide range of climate space and biodiversity will significantly expand the inference space. These new experiments are: EucFACE in a mature Eucalyptus stand on highly weathered soil in subtropical Australia; AmazonFACE in a highly diverse, primary rainforest in Brazil; BIFoR-FACE in a 150-yr-old deciduous woodland stand in central England; and SwedFACE proposed in a hemiboreal, Pinus sylvestris stand in Sweden. We now have a unique opportunity to initiate a model-data interaction as an integral part of experimental design and to address a set of cross-site science questions on topics including responses of mature forests; interactions with temperature, water stress, and phosphorus limitation; and the influence of biodiversity.
PanY, BirdseyRA, FangJ, HoughtonR, KauppiPE, KurzWA, PhillipsOL, ShvidenkoA, LewisSL, CanadellJG, CiaisP, JacksonRB, PacalaSW, McGuireAD, PiaoS, RautiainenA, SitchS, HayesD (2011). A large and persistent carbon sink in the world?s forests Science, 333, 988-993. DOI:10.1126/science.1201609URLPMID:21764754 [本文引用: 1] The terrestrial carbon sink has been large in recent decades, but its size and location remain uncertain. Using forest inventory data and long-term ecosystem carbon studies, we estimate a total forest sink of 2.4 +/- 0.4 petagrams of carbon per year (Pg C year(-1)) globally for 1990 to 2007. We also estimate a source of 1.3 +/- 0.7 Pg C year(-1) from tropical land-use change, consisting of a gross tropical deforestation emission of 2.9 +/- 0.5 Pg C year(-1) partially compensated by a carbon sink in tropical forest regrowth of 1.6 +/- 0.5 Pg C year(-1). Together, the fluxes comprise a net global forest sink of 1.1 +/- 0.8 Pg C year(-1), with tropical estimates having the largest uncertainties. Our total forest sink estimate is equivalent in magnitude to the terrestrial sink deduced from fossil fuel emissions and land-use change sources minus ocean and atmospheric sinks.
PengYF, GuoDL, YangYH (2017). Global patterns of root dynamics under nitrogen enrichment Global Ecology and Biogeography, 26, 102-114. [本文引用: 1]
PengYF, YangYH (2016). Allometric biomass partitioning under nitrogen enrichment: evidence from manipulative experiments around the world Scientific Reports, 6, 28918. DOI: 10.1038/srep28918. DOI:10.1038/srep28918URLPMID:27349584 [本文引用: 1] Allometric and optimal hypotheses have been widely used to explain biomass partitioning in response to resource changes for individual plants; however, little evidence has been reported from measurements at the community level across a broad geographic scale. This study assessed the nitrogen (N) effect on community-level root to shoot (R/S) ratios and biomass partitioning functions by synthesizing global manipulative experiments. Results showed that, in aggregate, N addition decreased the R/S ratios in various biomes. However, the scaling slopes of the allometric equations were not significantly altered by the N enrichment, possibly indicating that N-induced reduction of the R/S ratio is a consequence of allometric allocation as a function of increasing plant size rather than an optimal partitioning model. To further illustrate this point, we developed power function models to explore the relationships between aboveground and belowground biomass for various biomes; then, we generated the predicted root biomass from the observed shoot biomass and predicted R/S ratios. The comparison of predicted and observed N-induced changes of the R/S ratio revealed no significant differences between each other, supporting the allometric allocation hypothesis. These results suggest that allometry, rather than optimal allocation, explains the N-induced reduction in the R/S ratio across global biomes.
Pe?uelasJ, PoulterB, SardansJ, CiaisP, van der VeldeM, BoppL, BoucherO, GodderisY, HinsingerP, LlusiaJ, NardinE, ViccaS, ObersteinerM, JanssensIA (2013). Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe Nature Communications, 4, 2934. DOI: 10.1038/ncomms3934. DOI:10.1038/ncomms3934URLPMID:24343268 [本文引用: 2] The availability of carbon from rising atmospheric carbon dioxide levels and of nitrogen from various human-induced inputs to ecosystems is continuously increasing; however, these increases are not paralleled by a similar increase in phosphorus inputs. The inexorable change in the stoichiometry of carbon and nitrogen relative to phosphorus has no equivalent in Earth's history. Here we report the profound and yet uncertain consequences of the human imprint on the phosphorus cycle and nitrogen:phosphorus stoichiometry for the structure, functioning and diversity of terrestrial and aquatic organisms and ecosystems. A mass balance approach is used to show that limited phosphorus and nitrogen availability are likely to jointly reduce future carbon storage by natural ecosystems during this century. Further, if phosphorus fertilizers cannot be made increasingly accessible, the crop yields projections of the Millennium Ecosystem Assessment imply an increase of the nutrient deficit in developing regions.
Rappe-GeorgeMO, G?rden?sAI, KlejaDB (2013). The impact of four decades of annual nitrogen addition on dissolved organic matter in a boreal forest soil Biogeosciences, 10, 1365-1377. [本文引用: 4]
ReichPB, HungateBA, LuoY (2006). Carbon-nitrogen interactions in terrestrial ecosystems in response to rising atmospheric carbon dioxide Annual Review of Ecology, Evolution, and Systematics, 37, 611-636. [本文引用: 1]
RoslingA, MidgleyMG, CheekeT, UrbinaH, FranssonP, PhillipsRP (2016). Phosphorus cycling in deciduous forest soil differs between stands dominated by ecto- and arbuscular mycorrhizal trees New Phytologist, 209, 1184-1195. DOI:10.1111/nph.13720URLPMID:26510093 [本文引用: 1] Although much is known about how trees and their associated microbes influence nitrogen cycling in temperate forest soils, less is known about biotic controls over phosphorus (P) cycling. Given that mycorrhizal fungi are instrumental for P acquisition and that the two dominant associations - arbuscular mycorrhizal (AM) and ectomycorrhizal (ECM) fungi - possess different strategies for acquiring P, we hypothesized that P cycling would differ in stands dominated by trees associated with AM vs ECM fungi. We quantified soil solution P, microbial biomass P, and sequentially extracted inorganic and organic P pools from May to November in plots dominated by trees forming either AM or ECM associations in south-central Indiana, USA. Overall, fungal communities in AM and ECM plots were functionally different and soils exhibited fundamental differences in P cycling. Organic forms of P were more available in ECM plots than in AM plots. Yet inorganic P decreased and organic P accumulated over the growing season in both ECM and AM plots, resulting in increasingly P-limited microbial biomass. Collectively, our results suggest that P cycling in hardwood forests is strongly influenced by biotic processes in soil and that these are driven by plant-associated fungal communities.
SaarsalmiA, SmolanderA, KukkolaM, MoilanenM, Saram?kiJ (2012). 30-year effects of wood ash and nitrogen fertilization on soil chemical properties, soil microbial processes and stand growth in a Scots pine stand Forest Ecology and Management, 278, 63-70. [本文引用: 3]
Schulte-UebbingL, de VriesW (2018). Global-scale impacts of nitrogen deposition on tree carbon sequestration in tropical, temperate, and boreal forests: a meta-analysis Global Change Biology, 24, 416-431. [本文引用: 9]
TalbotJM, AllisonSD, TresederKK (2008). Decomposers in disguise: mycorrhizal fungi as regulators of soil C dynamics in ecosystems under global change Functional Ecology, 22, 955-963. [本文引用: 1]
TaylorBN, MengeDNL (2018). Light regulates tropical symbiotic nitrogen fixation more strongly than soil nitrogen Nature Plants, 4, 655-661. DOI:10.1038/s41477-018-0231-9URLPMID:30127409 [本文引用: 1] Nitrogen limits primary production in almost every biome on Earth(1,2). Symbiotic nitrogen fixation, conducted by certain angiosperms and their endosymbiotic bacteria, is the largest potential natural source of new nitrogen into the biosphere(3), influencing global primary production, carbon sequestration and element cycling. Because symbiotic nitrogen fixation represents an alternative to soil nitrogen uptake, much of the work on symbiotic nitrogen fixation regulation has focused on soil nitrogen availability(4-8). However, because symbiotic nitrogen fixation is an energetically expensive process(9), light availability to the plant may also regulate symbiotic nitrogen fixation rates(10,11). Despite the importance of symbiotic nitrogen fixation to biosphere functioning, the environmental factors that most strongly regulate this process remain unresolved. Here we show that light regulates symbiotic nitrogen fixation more strongly than does soil nitrogen and that light mediates the response of symbiotic nitrogen fixation to soil nitrogen availability. In a shadehouse experiment, low light levels (comparable with forest understories) completely shut down symbiotic nitrogen fixation, whereas soil nitrogen levels that far exceeded plant demand did not fully downregulate symbiotic nitrogen fixation at high light. For in situ forest seedlings, light was a notable predictor of symbiotic nitrogen fixation activity, but soil-extractable nitrogen was not. Light as a primary regulator of symbiotic nitrogen fixation is a departure from decades of focus on soil nitrogen availability. This shift in our understanding of symbiotic nitrogen fixation regulation can resolve a long-standing biogeochemical paradox(12), and it will improve our ability to predict how symbiotic nitrogen fixation will fuel the global forest carbon sink and respond to human alteration of the global nitrogen cycle.
ThomasRQ, CanhamCD, WeathersKC, GoodaleCL (2010). Increased tree carbon storage in response to nitrogen deposition in the US Nature Geoscience, 3, 13-17. [本文引用: 3]
TianD, LiP, FangW, XuJ, LuoY, YanZ, ZhuB, WangJ, XuX, FangJ (2017). Growth responses of trees and understory plants to nitrogen fertilization in a subtropical forest in China Biogeosciences, 14, 3461-3469. [本文引用: 7]
TianD, NiuS (2015). A global analysis of soil acidification caused by nitrogen addition Environmental Research Letters, 10, 024019. DOI: 10.1088/1748-9326/10/2/024019. [本文引用: 2]
TianD, WangH, SunJ, NiuS (2016). Global evidence on nitrogen saturation of terrestrial ecosystem net primary productivity Environmental Research Letters, 11, 024012. DOI: 10.1088/1748-9326/11/2/024012. [本文引用: 1]
VadeboncoeurMA (2010). Meta-analysis of fertilization experiments indicates multiple limiting nutrients in northeastern deciduous forests Canadian Journal of Forest Research, 40, 1766-1780. [本文引用: 5]
ViolleC, NavasML, VileD, KazakouE, FortunelC, HummelI, GarnierE (2007). Let the concept of trait be functional! Oikos, 116, 882-892. DOI:10.1111/oik.2007.116.issue-5URL [本文引用: 1]
VitousekPM (1984). Litterfall, nutrient cycling, and nutrient limitation in tropical forests Ecology, 65, 285-298. DOI:10.2307/1939481URL [本文引用: 4]
VitousekPM, FarringtonH (1997). Nutrient limitation and soil development: experimental test of a biogeochemical theory Biogeochemistry, 37, 63-75. [本文引用: 2]
VitousekPM, PorderS, HoultonBZ, ChadwickOA (2010). Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions Ecological Applications, 20, 5-15. DOI:10.1890/08-0127.1URLPMID:20349827 [本文引用: 6] Nutrient limitation to primary productivity and other biological processes is widespread in terrestrial ecosystems, and nitrogen (N) and phosphorus (P) are the most common limiting elements, both individually and in combination. Mechanisms that drive P limitation, and their interactions with the N cycle, have received less attention than mechanisms causing N limitation. We identify and discuss six mechanisms that could drive P limitation in terrestrial ecosystems. The best known of these is depletion-driven limitation, in which accumulated P losses during long-term soil and ecosystem development contribute to what Walker and Syers termed a
VitousekPM, WalkerLR, WhiteakerLD, MatsonPA (1993). Nutrient limitations to plant growth during primary succession in Hawaii Volcanoes National Park Biogeochemistry, 23, 197-215. [本文引用: 1]
WalkerTW, SyersJK (1976). The fate of phosphorus during pedogenesis Geoderma, 15, 1-19. [本文引用: 1]
WaringBG, Pérez-AvilesD, MurrayJG, PowersJS (2019). Plant community responses to stand-level nutrient fertilization in a secondary tropical dry forest Ecology, 100, e02691. DOI: 10.1002/ecy.2691. DOI:10.1002/ecy.2691URLPMID:30989648 [本文引用: 3] The size of the terrestrial carbon (C) sink is mediated by the availability of nutrients that limit plant growth. However, nutrient controls on primary productivity are poorly understood in the geographically extensive yet understudied tropical dry forest biome. To examine how nutrients influence above- and belowground biomass production in a secondary, seasonally dry tropical forest, we conducted a replicated, fully factorial nitrogen (N) and phosphorus (P) fertilization experiment at the stand scale in Guanacaste, Costa Rica. The production of leaves, wood, and fine roots was monitored through time; root colonization by mycorrhizal fungi and the abundance of N-fixing root nodules were also quantified. In this seasonal forest, interannual variation in rainfall had the largest influence on stand-level productivity, with lower biomass growth under drought. By contrast, aboveground productivity was generally not increased by nutrient addition, although fertilization enhanced growth of individual tree stems in a wet year. However, root growth increased markedly and consistently under P addition, significantly altering patterns of stand-level biomass allocation to above- vs. belowground compartments. Although nutrients did not stimulate total biomass production at the community scale, N-fixing legumes exhibited a twofold increase in woody growth in response to added P, accompanied by a dramatic increase in the abundance of root nodules. These data suggest that the relationship between nutrient availability and primary production in tropical dry forest is contingent on both water availability and plant functional diversity.
WooliverRC, MarionZH, PetersonCR, PottsBM, SeniorJK, BaileyJK, SchweitzerJA (2017). Phylogeny is a powerful tool for predicting plant biomass responses to nitrogen enrichment Ecology, 98, 2120-2132. DOI:10.1002/ecy.1896URLPMID:28500764 [本文引用: 1] Increasing rates of anthropogenic nitrogen (N) enrichment to soils often lead to the dominance of nitrophilic plant species and reduce plant diversity in natural ecosystems. Yet, we lack a framework to predict which species will be winners or losers in soil N enrichment scenarios, a framework that current literature suggests should integrate plant phylogeny, functional tradeoffs, and nutrient co-limitation. Using a controlled fertilization experiment, we quantified biomass responses to N enrichment for 23 forest tree species within the genus Eucalyptus that are native to Tasmania, Australia. Based on previous work with these species' responses to global change factors and theory on the evolution of plant resource-use strategies, we hypothesized that (1) growth responses to N enrichment are phylogenetically structured, (2) species with more resource-acquisitive functional traits have greater growth responses to N enrichment, and (3) phosphorus (P) limits growth responses to N enrichment differentially across species, wherein P enrichment increases growth responses to N enrichment more in some species than others. We built a hierarchical Bayesian model estimating effects of functional traits (specific leaf area, specific stem density, and specific root length) and P fertilization on species' biomass responses to N, which we then compared between lineages to determine whether phylogeny explains variation in responses to N. In concordance with literature on N limitation, a majority of species responded strongly and positively to N enrichment. Mean responses ranged three-fold, from 6.21 (E. pulchella) to 16.87 (E. delegatensis) percent increases in biomass per g N.m(-2) .yr(-1) added. We identified a strong difference in responses to N between two phylogenetic lineages in the Eucalyptus subgenus Symphyomyrtus, suggesting that shared ancestry explains variation in N limitation. However, our model indicated that after controlling for phylogenetic non-independence, eucalypt responses to N were not associated with functional traits (although post-hoc analyses show a phylogenetic pattern in specific root length similar to that of responses to N), nor were responses differentially limited by P. Overall, our model results suggest that phylogeny is a powerful predictor of winners and losers in anthropogenic N enrichment scenarios in Tasmanian eucalypts, which may have implications for other species.
WrightSJ (2019). Plant responses to nutrient addition experiments conducted in tropical forests Ecological Monographs, 89, e01382. DOI: 10.1002/ecm.1382. [本文引用: 3]
WrightSJ, TurnerBL, YavittJB, HarmsKE, KaspariM, TannerEVJ, BujanJ, GriffinEA, MayorJR, PasquiniSC, SheldrakeM, GarciaMN (2018). Plant responses to fertilization experiments in lowland, species-rich, tropical forests Ecology, 99, 1129-1138. DOI:10.1002/ecy.2193URLPMID:29460277 [本文引用: 3] We present a meta-analysis of plant responses to fertilization experiments conducted in lowland, species-rich, tropical forests. We also update a key result and present the first species-level analyses of tree growth rates for a 15-yr factorial nitrogen (N), phosphorus (P), and potassium (K) experiment conducted in central Panama. The update concerns community-level tree growth rates, which responded significantly to the addition of N and K together after 10 yr of fertilization but not after 15 yr. Our experimental soils are infertile for the region, and species whose regional distributions are strongly associated with low soil P availability dominate the local tree flora. Under these circumstances, we expect muted responses to fertilization, and we predicted species associated with low-P soils would respond most slowly. The data did not support this prediction, species-level tree growth responses to P addition were unrelated to species-level soil P associations. The meta-analysis demonstrated that nutrient limitation is widespread in lowland tropical forests and evaluated two directional hypotheses concerning plant responses to N addition and to P addition. The meta-analysis supported the hypothesis that tree (or biomass) growth rate responses to fertilization are weaker in old growth forests and stronger in secondary forests, where rapid biomass accumulation provides a nutrient sink. The meta-analysis found no support for the long-standing hypothesis that plant responses are stronger for P addition and weaker for N addition. We do not advocate discarding the latter hypothesis. There are only 14 fertilization experiments from lowland, species-rich, tropical forests, 13 of the 14 experiments added nutrients for five or fewer years, and responses vary widely among experiments. Potential fertilization responses should be muted when the species present are well adapted to nutrient-poor soils, as is the case in our experiment, and when pest pressure increases with fertilization, as it does in our experiment. The statistical power and especially the duration of fertilization experiments conducted in old growth, tropical forests might be insufficient to detect the slow, modest growth responses that are to be expected.
XuXF, ThorntonPE, PostWM (2013). A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems Global Ecology and Biogeography, 22, 737-749. [本文引用: 2]
YuanZY, ChenHYH (2012). A global analysis of fine root production as affected by soil nitrogen and phosphorus Proceedings of the Royal Society B, 279, 3796-3802. DOI:10.1098/rspb.2012.0955URLPMID:22764168 [本文引用: 4]
YueK, FornaraDA, YangW, PengY, PengC, LiuZ, WuF (2017). Influence of multiple global change drivers on terrestrial carbon storage: additive effects are common Ecology Letters, 20, 663-672. DOI:10.1111/ele.12767URLPMID:28370812 [本文引用: 1] The interactive effects of multiple global change drivers on terrestrial carbon (C) storage remain poorly understood. Here, we synthesise data from 633 published studies to show how the interactive effects of multiple drivers are generally additive (i.e. not differing from the sum of their individual effects) rather than synergistic or antagonistic. We further show that (1) elevated CO2 , warming, N addition, P addition and increased rainfall, all exerted positive individual effects on plant C pools at both single-plant and plant-community levels; (2) plant C pool responses to individual or combined effects of multiple drivers are seldom scale-dependent (i.e. not differing from single-plant to plant-community levels) and (3) soil and microbial biomass C pools are significantly less sensitive than plant C pools to individual or combined effects. We provide a quantitative basis for integrating additive effects of multiple global change drivers into future assessments of the C storage ability of terrestrial ecosystems.
ZhaoQ, ZengDH (2019). Nitrogen addition effects on tree growth and soil properties mediated by soil phosphorus availability and tree species identity Forest Ecology and Management, 449, 117478. DOI: 10.1016/j.foreco.2019.117478. [本文引用: 1]
ZhengM, ZhouZ, LuoY, ZhaoP, MoJ (2019). Global pattern and controls of biological nitrogen fixation under nutrient enrichment: a meta-analysis Global Change Biology, 25, 3018-3030. DOI:10.1111/gcb.14705URLPMID:31120621 [本文引用: 2] /=30 degrees ) biomes (32.9%-61.3%), and the sensitivity (defined as the absolute value of response ratios) of BNF to nutrients in mid-/high-latitude biomes decreased with decreasing latitude (p 0.05), but environmental factors did affect it (p
ZhouXL, CaiQ, XiongXY, FangWJ, ZhuJX, ZhuJL, FangJY, JiCJ (2018). Ecosystem carbon stock and within-?system distribution in successionalFagus lucida forests in Mt. Yueliang, Guizhou, China Chinese Journal of Plant Ecology, 42, 703-712. [本文引用: 1]
ZhuF, LuX, MoJ (2014). Phosphorus limitation on photosynthesis of two dominant understory species in a lowland tropical forest Journal of Plant Ecology, 7, 526-534. [本文引用: 1]
ZouAL, LiXP, NiXF, JiCJ (2019). Responses of tree growth to nitrogen addition inQuercus wutaishanica forests in Mt. Dongling, Beijing, China Chinese Journal of Plant Ecology, 43, 783-792. [本文引用: 2]
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
... 在生态系统内的某类植物个体或群体水平上(如小树与大树), N、P添加对树木生长的影响因树木径级的不同而存在差异.海南热带山地森林的一项为期5年的研究发现, P添加对小树(DBH ≤ 10 cm)和中树(10 cm < DBH ≤ 20 cm)的生长表现为显著的促进作用; 但对大树(DBH > 20 cm)的生长没有显著影响(Jiang et al., 2018).其他一些在热带森林中开展的实验也表明, N、P添加对树木生长的影响也因树木径级的不同而存在明显的差异(Alvarez-Clare et al., 2013; Fisher et al., 2013).在鸡公山温带森林的研究表明, N添加对小树和大树生长都没有显著的影响, 而P添加在小树中表现为显著的促进作用(Li et al., 2018).也有研究发现, N、P添加对树木生长的促进作用与树木的初始大小具有密切的正相关关系, 即在N、P添加下大树的生长要快于小树(Báez & Homeier, 2018).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... ).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... ); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
Plant-soil associations in a lower montane tropical forest: physiological acclimation and herbivore-mediated responses to nitrogen addition 1 2010
... 在生态系统尺度上, N、P添加对树木生长和森林生产力的影响在不同的林龄中也不尽相同, 受到林龄的调控.Goswami等(2018)在温带森林的研究发现, N添加对幼龄林生长的促进作用强于中龄林和成熟林; P添加在中龄林中的促进作用最为明显, 而在幼龄林和成熟林中均没有明显的作用.然而, 也有研究发现, N、P添加对次生林(林龄较小)的生长表现为促进, 而对成熟林没有显著影响, 即次生林的生长响应明显高于成熟林(Wright et al., 2018; Wright, 2019).也有定量研究发现, N添加对树木生长的影响随着年龄的增大而显著降低(Vadeboncoeur, 2010), 表明幼龄林对N添加的响应更为强烈.总体而言, 目前的研究结果表明, N添加对幼龄林的促进作用一般高于中龄林和老龄林(Vadeboncoeur, 2010; Rappe-George et al., 2013; Schulte-Uebbing & de Vries, 2018), 因为幼龄林树木在吸收添加的养分之后, 能够快速地进行光合作用, 合成自身新陈代谢所需要的有机物, 生长相对较快.Wright等(2018)认为老龄林生长对N、P添加响应较弱的原因可能存在于两个方面: 一方面, 生活在养分可利用性较低环境中的物种已经适应了这种养分缺乏的条件, 在面对较好的养分条件时其增加生长的潜力有限(Coley et al., 1985), 群落生长的响应可能需要等到物种组成发生变化以适应较高养分可用性时才能体现出来(Chapin III et al., 1986); 另一方面, 植物在施肥条件下其组织中的N含量升高, 会增加其受到天敌(食草动物和其他害虫)危害的风险并消耗其初级生产力, 从而对树木生长的响应产生不利影响(Andersen et al., 2010). ...
Above- and below-ground net primary productivity across ten Amazonian forests on contrasting soils 1 2009
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
Continental-scale nitrogen pollution is shifting forest mycorrhizal associations and soil carbon stocks 4 2018
... 在菌根类型上, 不同研究发现, 与丛枝菌根(AM)真菌共生的树种(AM树种)和与外生菌根(ECM)真菌共生的树种(ECM树种)在N、P沉降下的响应存在差异, 表明菌根类型会调控N、P添加对树木生长和森林生产力的影响(Thomas et al., 2010; Averill et al., 2018; DeForest & Snell, 2020).在N添加研究方面, 基于美国森林资源清查数据的研究表明, N沉降促进了11个物种的生长, 抑制了3个物种的生长, 而且AM树种全部表现为促进作用(Thomas et al., 2010).最新的一项研究表明, 在N水平低的时候, ECM树种的生长对N的响应要高于AM树种, 而在N水平高的时候, AM树种的生长要高于ECM树种(Averill et al., 2018).AM树种和ECM树种对N可利用性增加的差异性响应, 可能与它们对N的获取与利用策略有关系.与ECM真菌相比, AM真菌缺少分解土壤有机质的酶, 主要获取土壤中的无机N来满足新陈代谢的需要和向植物输送养分(Chalot & Brun, 1998; Talbot et al., 2008).因此, 当活性N沉降导致土壤N的可利用性提高时, AM树种能够比ECM树种获得更多的益处使生长加快. ...
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
... (3)深度研究植物功能性状在调节与预测树木生长对N、P添加响应中的作用.尽管已有相关研究从植物功能性状的角度研究了N、P添加如何影响树木生长, 但这些研究还依旧十分匮乏, 而且现有的研究也仅仅考虑了单一的植物功能性状(Thomas et al., 2010; Averill et al., 2018; Nasto et al., 2019).植物的功能性状具有多个维度, 而且这些性状常常是交织在一起, 比如植物可以同时具备固氮和共生菌根的特性, 固氮植物具有较高的AM菌根侵染率等(Nasto et al., 2014).在这种情况下, 考虑单一的功能性状可能会使研究结果受到其他因素的干扰.因此, 在未来研究中需要综合考虑植物各个部分的性状, 并采用较为复杂全面的统计方法定量分析植物功能性状在调控树木生长响应N、P添加中的作用. ...
Functional traits determine tree growth and ecosystem productivity of a tropical montane forest: insights from a long-term nutrient manipulation experiment 4 2018
... 在生态系统内的某类植物个体或群体水平上(如小树与大树), N、P添加对树木生长的影响因树木径级的不同而存在差异.海南热带山地森林的一项为期5年的研究发现, P添加对小树(DBH ≤ 10 cm)和中树(10 cm < DBH ≤ 20 cm)的生长表现为显著的促进作用; 但对大树(DBH > 20 cm)的生长没有显著影响(Jiang et al., 2018).其他一些在热带森林中开展的实验也表明, N、P添加对树木生长的影响也因树木径级的不同而存在明显的差异(Alvarez-Clare et al., 2013; Fisher et al., 2013).在鸡公山温带森林的研究表明, N添加对小树和大树生长都没有显著的影响, 而P添加在小树中表现为显著的促进作用(Li et al., 2018).也有研究发现, N、P添加对树木生长的促进作用与树木的初始大小具有密切的正相关关系, 即在N、P添加下大树的生长要快于小树(Báez & Homeier, 2018).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... 多数研究发现, 不同物种树木的生长对N、P添加的响应不尽相同(Báez & Homeier, 2018; Goswami et al., 2018; DeForest & Snell, 2020), 这种差异在很大程度上可能取决于植物功能性状的异同.植物功能性状是植物对外界环境长期响应与适应后所呈现出来的可量度的特征(Violle et al., 2007), 如叶片特征(大小、厚度、韧性、比叶面积和元素含量等), 根系特征(根系构型、根系呼吸、根系分泌物、直径、长度和元素含量等)以及生物性状(植物共生菌根、固氮等)等(Cornelissen et al., 2003; Bardgett et al., 2014; 何念鹏等, 2018).目前, 植物功能性状调控树木生长响应N、P添加方面的研究主要集中在共生菌根类型和树木固氮特性两个方面. ...
Absolute and relative changes in tree growth rates and changes to the stand diameter distribution of Pinus taeda as a result of midrotation fertilizer applications 1 2008
... Methods for assessing the effects of nitrogen and/or phosphorus addition on plant growth and productivity Table 1
Physiology of organic nitrogen acquisition by ectomycorrhizal fungi and ectomycorrhizas 1 1998
... 在菌根类型上, 不同研究发现, 与丛枝菌根(AM)真菌共生的树种(AM树种)和与外生菌根(ECM)真菌共生的树种(ECM树种)在N、P沉降下的响应存在差异, 表明菌根类型会调控N、P添加对树木生长和森林生产力的影响(Thomas et al., 2010; Averill et al., 2018; DeForest & Snell, 2020).在N添加研究方面, 基于美国森林资源清查数据的研究表明, N沉降促进了11个物种的生长, 抑制了3个物种的生长, 而且AM树种全部表现为促进作用(Thomas et al., 2010).最新的一项研究表明, 在N水平低的时候, ECM树种的生长对N的响应要高于AM树种, 而在N水平高的时候, AM树种的生长要高于ECM树种(Averill et al., 2018).AM树种和ECM树种对N可利用性增加的差异性响应, 可能与它们对N的获取与利用策略有关系.与ECM真菌相比, AM真菌缺少分解土壤有机质的酶, 主要获取土壤中的无机N来满足新陈代谢的需要和向植物输送养分(Chalot & Brun, 1998; Talbot et al., 2008).因此, 当活性N沉降导致土壤N的可利用性提高时, AM树种能够比ECM树种获得更多的益处使生长加快. ...
The nature of nutrient limitation in plant communities 1 1986
... 在生态系统尺度上, N、P添加对树木生长和森林生产力的影响在不同的林龄中也不尽相同, 受到林龄的调控.Goswami等(2018)在温带森林的研究发现, N添加对幼龄林生长的促进作用强于中龄林和成熟林; P添加在中龄林中的促进作用最为明显, 而在幼龄林和成熟林中均没有明显的作用.然而, 也有研究发现, N、P添加对次生林(林龄较小)的生长表现为促进, 而对成熟林没有显著影响, 即次生林的生长响应明显高于成熟林(Wright et al., 2018; Wright, 2019).也有定量研究发现, N添加对树木生长的影响随着年龄的增大而显著降低(Vadeboncoeur, 2010), 表明幼龄林对N添加的响应更为强烈.总体而言, 目前的研究结果表明, N添加对幼龄林的促进作用一般高于中龄林和老龄林(Vadeboncoeur, 2010; Rappe-George et al., 2013; Schulte-Uebbing & de Vries, 2018), 因为幼龄林树木在吸收添加的养分之后, 能够快速地进行光合作用, 合成自身新陈代谢所需要的有机物, 生长相对较快.Wright等(2018)认为老龄林生长对N、P添加响应较弱的原因可能存在于两个方面: 一方面, 生活在养分可利用性较低环境中的物种已经适应了这种养分缺乏的条件, 在面对较好的养分条件时其增加生长的潜力有限(Coley et al., 1985), 群落生长的响应可能需要等到物种组成发生变化以适应较高养分可用性时才能体现出来(Chapin III et al., 1986); 另一方面, 植物在施肥条件下其组织中的N含量升高, 会增加其受到天敌(食草动物和其他害虫)危害的风险并消耗其初级生产力, 从而对树木生长的响应产生不利影响(Andersen et al., 2010). ...
Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment 1 2016
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
Global patterns of terrestrial biological nitrogen (N2) fixation in natural ecosystems 1 1999
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
Resource availability and plant antiherbivore defense 1 1985
... 在生态系统尺度上, N、P添加对树木生长和森林生产力的影响在不同的林龄中也不尽相同, 受到林龄的调控.Goswami等(2018)在温带森林的研究发现, N添加对幼龄林生长的促进作用强于中龄林和成熟林; P添加在中龄林中的促进作用最为明显, 而在幼龄林和成熟林中均没有明显的作用.然而, 也有研究发现, N、P添加对次生林(林龄较小)的生长表现为促进, 而对成熟林没有显著影响, 即次生林的生长响应明显高于成熟林(Wright et al., 2018; Wright, 2019).也有定量研究发现, N添加对树木生长的影响随着年龄的增大而显著降低(Vadeboncoeur, 2010), 表明幼龄林对N添加的响应更为强烈.总体而言, 目前的研究结果表明, N添加对幼龄林的促进作用一般高于中龄林和老龄林(Vadeboncoeur, 2010; Rappe-George et al., 2013; Schulte-Uebbing & de Vries, 2018), 因为幼龄林树木在吸收添加的养分之后, 能够快速地进行光合作用, 合成自身新陈代谢所需要的有机物, 生长相对较快.Wright等(2018)认为老龄林生长对N、P添加响应较弱的原因可能存在于两个方面: 一方面, 生活在养分可利用性较低环境中的物种已经适应了这种养分缺乏的条件, 在面对较好的养分条件时其增加生长的潜力有限(Coley et al., 1985), 群落生长的响应可能需要等到物种组成发生变化以适应较高养分可用性时才能体现出来(Chapin III et al., 1986); 另一方面, 植物在施肥条件下其组织中的N含量升高, 会增加其受到天敌(食草动物和其他害虫)危害的风险并消耗其初级生产力, 从而对树木生长的响应产生不利影响(Andersen et al., 2010). ...
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide 1 2003
... 多数研究发现, 不同物种树木的生长对N、P添加的响应不尽相同(Báez & Homeier, 2018; Goswami et al., 2018; DeForest & Snell, 2020), 这种差异在很大程度上可能取决于植物功能性状的异同.植物功能性状是植物对外界环境长期响应与适应后所呈现出来的可量度的特征(Violle et al., 2007), 如叶片特征(大小、厚度、韧性、比叶面积和元素含量等), 根系特征(根系构型、根系呼吸、根系分泌物、直径、长度和元素含量等)以及生物性状(植物共生菌根、固氮等)等(Cornelissen et al., 2003; Bardgett et al., 2014; 何念鹏等, 2018).目前, 植物功能性状调控树木生长响应N、P添加方面的研究主要集中在共生菌根类型和树木固氮特性两个方面. ...
Do nutrient limitation patterns shift from nitrogen toward phosphorus with increasing nitrogen deposition across the northeastern United States? 1 2012
... 养分限制模式对于生态系统生产力及其对全球变化的响应至关重要(Reich et al., 2006; Crowley et al., 2012).在全球尺度上, 氮(N)和磷(P)对植物生产力的限制被认为普遍存在于陆地、淡水和海洋生态系统中(Elser et al., 2007).在大气CO2浓度持续上升背景下, 植物生长受到促进, 从而会从土壤中摄取更多的养分, 进而导致其生长逐渐受到养分可利用性的限制(Luo et al., 2004).模型预测结果表明, 在考虑N、P可利用性限制的情况下, 净初级生产力的增加将低于N和P不受限制状态下的增加量, 而陆地生态系统也将由碳吸收转变为碳排放(Wieder et al., 2015).由此可见, 养分可利用性在调节陆地净初级生产力和陆地碳库中发挥着至关重要的作用. ...
Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of Actinobacteria and Proteobacteria in agro-ecosystems across the globe 1 2018
... 尽管有很多研究发现, N、P添加能够促进生态系统尺度上的树木生长和森林生产力, 但是N、P添加对它们的抑制作用在实验中也经常被观测到(Harpole et al., 2011; Li et al., 2018; Nasto et al., 2019).因此, 这对根据N、P添加是否促进生长从而判断生态系统或树木N、P的限制提出了挑战.以往的研究表明, 长期的N、P添加能够导致土壤pH值下降, 使得土壤酸化(Tian & Niu, 2015; Dai et al., 2018).尽管N、P添加所产生的负效应能够从过量的养分添加所产生的土壤酸化等毒害作用进行解释, 但是当没有毒害作用发生时, N、P添加依旧对树木生长产生负效应(?apek et al., 2018).针对这种情况, ?apek等(2018)提出了一个基于植物和微生物相互作用(plant-microbe interaction)的新的概念框架模型来解释并预测在生态系统尺度上N、P添加对初级生产力的影响.在该化学计量框架模型中, 植物与微生物之间的相互作用包括两种情况: 一是竞争关系; 二是协作关系, 如植物通过根系向土壤中分泌活性有机碳, 促进微生物的生长, 而微生物矿化有机质为植物提供N或P.该概念框架模型认为, 在一个生态系统中存在3种N、P的限制模式, 一是对于植物和土壤微生物都是N限制; 二是对于植物为N限制, 但是对于土壤微生物是P限制; 三是对于植物和土壤微生物都是P限制.简单来说, 如果一个系统中, 植物和微生物之间是协作关系占主导, 添加对植物生长非限制性的养分可以通过促进受该养分限制的微生物生长而间接提高植物的营养和生长; 相反, 在植物和微生物间是竞争作用占主导的系统内, 植物会受到多种养分的共同限制(?apek et al., 2018).该概念框架模型能够很好地解释生态系统生产力对N、P添加的响应, 并对以往研究中难以解释的响应提供了解决思路. ...
Tree growth response to shifting soil nutrient economy depends on mycorrhizal associations 8 2020
... 在各种类型的生态系统中, 森林占到全球陆地面积的30%, 其每年的碳吸收量占到全球陆地碳汇总量的82%, 在全球碳循环中占据着重要地位(Pan et al., 2011).自工业革命以来, 由于人为活动, 如施用矿质肥料、生物质燃烧等, 使得输入到生物圈的N和P呈现快速上升的趋势.N和P的沉降量在2000年的时候分别达到220和12 Tg·a-1 (Pe?uelas et al., 2013), 预计在未来将呈持续上升的趋势.N和P的持续输入会使得生态系统中的N、P可利用性提高(Lu et al., 2011a; Feng & Zhu, 2019), 并对生态系统碳循环过程产生显著影响, 如树木生长、生产力及土壤碳库(Lu et al., 2011b; Yue et al., 2017).森林树木生长及其生产力的变化对于预测全球变化背景下的碳循环过程和陆地碳汇潜力至关重要.目前, N、P沉降如何影响树木生长和森林生产力成为全球变化领域的研究热点(Elser et al., 2007; Camarero & Carrer, 2017; DeForest & Snell, 2020). ...
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
... 多数研究发现, 不同物种树木的生长对N、P添加的响应不尽相同(Báez & Homeier, 2018; Goswami et al., 2018; DeForest & Snell, 2020), 这种差异在很大程度上可能取决于植物功能性状的异同.植物功能性状是植物对外界环境长期响应与适应后所呈现出来的可量度的特征(Violle et al., 2007), 如叶片特征(大小、厚度、韧性、比叶面积和元素含量等), 根系特征(根系构型、根系呼吸、根系分泌物、直径、长度和元素含量等)以及生物性状(植物共生菌根、固氮等)等(Cornelissen et al., 2003; Bardgett et al., 2014; 何念鹏等, 2018).目前, 植物功能性状调控树木生长响应N、P添加方面的研究主要集中在共生菌根类型和树木固氮特性两个方面. ...
... 在菌根类型上, 不同研究发现, 与丛枝菌根(AM)真菌共生的树种(AM树种)和与外生菌根(ECM)真菌共生的树种(ECM树种)在N、P沉降下的响应存在差异, 表明菌根类型会调控N、P添加对树木生长和森林生产力的影响(Thomas et al., 2010; Averill et al., 2018; DeForest & Snell, 2020).在N添加研究方面, 基于美国森林资源清查数据的研究表明, N沉降促进了11个物种的生长, 抑制了3个物种的生长, 而且AM树种全部表现为促进作用(Thomas et al., 2010).最新的一项研究表明, 在N水平低的时候, ECM树种的生长对N的响应要高于AM树种, 而在N水平高的时候, AM树种的生长要高于ECM树种(Averill et al., 2018).AM树种和ECM树种对N可利用性增加的差异性响应, 可能与它们对N的获取与利用策略有关系.与ECM真菌相比, AM真菌缺少分解土壤有机质的酶, 主要获取土壤中的无机N来满足新陈代谢的需要和向植物输送养分(Chalot & Brun, 1998; Talbot et al., 2008).因此, 当活性N沉降导致土壤N的可利用性提高时, AM树种能够比ECM树种获得更多的益处使生长加快. ...
Ecosystem scale trade-off in nitrogen acquisition pathways 2 2018
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
... ; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
Weak growth response to nitrogen deposition in an old-growth boreal forest 4 2014
... 目前, 在N、P添加实验中, 树木生长和森林生产力的常用评估方法主要有两种: 一是相对生长速率, 二是绝对增长量.相对生长速率(表1)一般是用一段时间N、P添加处理之后树木胸径(DBH)、胸高断面积或生物量与处理之前的差值除以所对应的时间(Markewitz et al., 2012; Tian et al., 2017; Li et al., 2018).此外, 也有研究使用一段时间N、P添加处理之后DBH或胸高断面积与处理之前的差值除以处理之前的DBH或胸高断面积来表示树木的相对生长(Jiang et al., 2018).绝对增长量(表1)一般是用N、P添加处理一段时间内的DBH、胸高断面积、生物量(或生产力, 以碳储量计算)的绝对变化量来表示.生物量的变化是基于胸径-树高曲线、树木其他器官生物量与DBH和树高之间混合回归模型、异速生长方程等计算林分尺度总生物量的变化(Herbert & Fownes, 1995; Markewitz et al., 2012; Du & Fang, 2014; 周序力等, 2018).在森林根系生产力的估测中, 也有很多研究利用内生长环法(ingrowth core method)进行实测(Waring et al., 2019).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
... Methods for assessing the effects of nitrogen and/or phosphorus addition on plant growth and productivity Table 1
Nutrient limitation in rainforests and cloud forests along a 3000-m elevation gradient in the Peruvian Andes 2 2013
... 目前, 大部分的研究集中在单独的N添加和单独的P添加对树木生长和森林生产力的影响, 仅少数研究探讨了二者交互作用的影响(图1B).在N × P交互作用的影响方面, 部分研究发现, N和P添加之间不存在交互作用(Fisher et al., 2013; Goswami et al., 2018; Wright, 2019; Zhao & Zeng, 2019), 即N × P交互作用对树木生长和森林生产力不产生显著影响.也有研究表明, N和P添加之间存在交互作用, 这种交互作用在热带森林中表现为协同作用(Jiang et al., 2018, 2019; Li et al., 2018), 即N × P交互作用对树木生长和森林生产力表现为促进作用.N和P协同作用的产生机制可能与单一的N或P添加会加剧另一元素的限制有关(Elser et al., 2007; Li et al., 2016; Jiang et al., 2019).目前, N × P交互作用对树木生长和森林生产力影响的研究主要分布在热带森林中, 在温带森林和北方森林中的研究还十分有限(图1B), 相关机制需要进一步探讨. ...
... 在生态系统内的某类植物个体或群体水平上(如小树与大树), N、P添加对树木生长的影响因树木径级的不同而存在差异.海南热带山地森林的一项为期5年的研究发现, P添加对小树(DBH ≤ 10 cm)和中树(10 cm < DBH ≤ 20 cm)的生长表现为显著的促进作用; 但对大树(DBH > 20 cm)的生长没有显著影响(Jiang et al., 2018).其他一些在热带森林中开展的实验也表明, N、P添加对树木生长的影响也因树木径级的不同而存在明显的差异(Alvarez-Clare et al., 2013; Fisher et al., 2013).在鸡公山温带森林的研究表明, N添加对小树和大树生长都没有显著的影响, 而P添加在小树中表现为显著的促进作用(Li et al., 2018).也有研究发现, N、P添加对树木生长的促进作用与树木的初始大小具有密切的正相关关系, 即在N、P添加下大树的生长要快于小树(Báez & Homeier, 2018).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
The contribution of nitrogen deposition to the photosynthetic capacity of forests 1 2013
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
Phosphorus limitation of aboveground production in northern hardwood forests 4 2018
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
... 目前, 大部分的研究集中在单独的N添加和单独的P添加对树木生长和森林生产力的影响, 仅少数研究探讨了二者交互作用的影响(图1B).在N × P交互作用的影响方面, 部分研究发现, N和P添加之间不存在交互作用(Fisher et al., 2013; Goswami et al., 2018; Wright, 2019; Zhao & Zeng, 2019), 即N × P交互作用对树木生长和森林生产力不产生显著影响.也有研究表明, N和P添加之间存在交互作用, 这种交互作用在热带森林中表现为协同作用(Jiang et al., 2018, 2019; Li et al., 2018), 即N × P交互作用对树木生长和森林生产力表现为促进作用.N和P协同作用的产生机制可能与单一的N或P添加会加剧另一元素的限制有关(Elser et al., 2007; Li et al., 2016; Jiang et al., 2019).目前, N × P交互作用对树木生长和森林生产力影响的研究主要分布在热带森林中, 在温带森林和北方森林中的研究还十分有限(图1B), 相关机制需要进一步探讨. ...
... 在生态系统尺度上, N、P添加对树木生长和森林生产力的影响在不同的林龄中也不尽相同, 受到林龄的调控.Goswami等(2018)在温带森林的研究发现, N添加对幼龄林生长的促进作用强于中龄林和成熟林; P添加在中龄林中的促进作用最为明显, 而在幼龄林和成熟林中均没有明显的作用.然而, 也有研究发现, N、P添加对次生林(林龄较小)的生长表现为促进, 而对成熟林没有显著影响, 即次生林的生长响应明显高于成熟林(Wright et al., 2018; Wright, 2019).也有定量研究发现, N添加对树木生长的影响随着年龄的增大而显著降低(Vadeboncoeur, 2010), 表明幼龄林对N添加的响应更为强烈.总体而言, 目前的研究结果表明, N添加对幼龄林的促进作用一般高于中龄林和老龄林(Vadeboncoeur, 2010; Rappe-George et al., 2013; Schulte-Uebbing & de Vries, 2018), 因为幼龄林树木在吸收添加的养分之后, 能够快速地进行光合作用, 合成自身新陈代谢所需要的有机物, 生长相对较快.Wright等(2018)认为老龄林生长对N、P添加响应较弱的原因可能存在于两个方面: 一方面, 生活在养分可利用性较低环境中的物种已经适应了这种养分缺乏的条件, 在面对较好的养分条件时其增加生长的潜力有限(Coley et al., 1985), 群落生长的响应可能需要等到物种组成发生变化以适应较高养分可用性时才能体现出来(Chapin III et al., 1986); 另一方面, 植物在施肥条件下其组织中的N含量升高, 会增加其受到天敌(食草动物和其他害虫)危害的风险并消耗其初级生产力, 从而对树木生长的响应产生不利影响(Andersen et al., 2010). ...
... 多数研究发现, 不同物种树木的生长对N、P添加的响应不尽相同(Báez & Homeier, 2018; Goswami et al., 2018; DeForest & Snell, 2020), 这种差异在很大程度上可能取决于植物功能性状的异同.植物功能性状是植物对外界环境长期响应与适应后所呈现出来的可量度的特征(Violle et al., 2007), 如叶片特征(大小、厚度、韧性、比叶面积和元素含量等), 根系特征(根系构型、根系呼吸、根系分泌物、直径、长度和元素含量等)以及生物性状(植物共生菌根、固氮等)等(Cornelissen et al., 2003; Bardgett et al., 2014; 何念鹏等, 2018).目前, 植物功能性状调控树木生长响应N、P添加方面的研究主要集中在共生菌根类型和树木固氮特性两个方面. ...
Nutrient co-limitation of primary producer communities 2 2011
... 尽管有很多研究发现, N、P添加能够促进生态系统尺度上的树木生长和森林生产力, 但是N、P添加对它们的抑制作用在实验中也经常被观测到(Harpole et al., 2011; Li et al., 2018; Nasto et al., 2019).因此, 这对根据N、P添加是否促进生长从而判断生态系统或树木N、P的限制提出了挑战.以往的研究表明, 长期的N、P添加能够导致土壤pH值下降, 使得土壤酸化(Tian & Niu, 2015; Dai et al., 2018).尽管N、P添加所产生的负效应能够从过量的养分添加所产生的土壤酸化等毒害作用进行解释, 但是当没有毒害作用发生时, N、P添加依旧对树木生长产生负效应(?apek et al., 2018).针对这种情况, ?apek等(2018)提出了一个基于植物和微生物相互作用(plant-microbe interaction)的新的概念框架模型来解释并预测在生态系统尺度上N、P添加对初级生产力的影响.在该化学计量框架模型中, 植物与微生物之间的相互作用包括两种情况: 一是竞争关系; 二是协作关系, 如植物通过根系向土壤中分泌活性有机碳, 促进微生物的生长, 而微生物矿化有机质为植物提供N或P.该概念框架模型认为, 在一个生态系统中存在3种N、P的限制模式, 一是对于植物和土壤微生物都是N限制; 二是对于植物为N限制, 但是对于土壤微生物是P限制; 三是对于植物和土壤微生物都是P限制.简单来说, 如果一个系统中, 植物和微生物之间是协作关系占主导, 添加对植物生长非限制性的养分可以通过促进受该养分限制的微生物生长而间接提高植物的营养和生长; 相反, 在植物和微生物间是竞争作用占主导的系统内, 植物会受到多种养分的共同限制(?apek et al., 2018).该概念框架模型能够很好地解释生态系统生产力对N、P添加的响应, 并对以往研究中难以解释的响应提供了解决思路. ...
... 多数研究发现, 不同物种树木的生长对N、P添加的响应不尽相同(Báez & Homeier, 2018; Goswami et al., 2018; DeForest & Snell, 2020), 这种差异在很大程度上可能取决于植物功能性状的异同.植物功能性状是植物对外界环境长期响应与适应后所呈现出来的可量度的特征(Violle et al., 2007), 如叶片特征(大小、厚度、韧性、比叶面积和元素含量等), 根系特征(根系构型、根系呼吸、根系分泌物、直径、长度和元素含量等)以及生物性状(植物共生菌根、固氮等)等(Cornelissen et al., 2003; Bardgett et al., 2014; 何念鹏等, 2018).目前, 植物功能性状调控树木生长响应N、P添加方面的研究主要集中在共生菌根类型和树木固氮特性两个方面. ...
森林生态系统性状的空间格局与影响因素研究进展——基于中国东部样带的整合分析 1 2018
... 多数研究发现, 不同物种树木的生长对N、P添加的响应不尽相同(Báez & Homeier, 2018; Goswami et al., 2018; DeForest & Snell, 2020), 这种差异在很大程度上可能取决于植物功能性状的异同.植物功能性状是植物对外界环境长期响应与适应后所呈现出来的可量度的特征(Violle et al., 2007), 如叶片特征(大小、厚度、韧性、比叶面积和元素含量等), 根系特征(根系构型、根系呼吸、根系分泌物、直径、长度和元素含量等)以及生物性状(植物共生菌根、固氮等)等(Cornelissen et al., 2003; Bardgett et al., 2014; 何念鹏等, 2018).目前, 植物功能性状调控树木生长响应N、P添加方面的研究主要集中在共生菌根类型和树木固氮特性两个方面. ...
Global organization of terrestrial plant- nutrient interactions 1 2004
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
Phosphorus limitation of forest leaf-area and net primary production on a highly weathered soil 1 1995
... 目前, 在N、P添加实验中, 树木生长和森林生产力的常用评估方法主要有两种: 一是相对生长速率, 二是绝对增长量.相对生长速率(表1)一般是用一段时间N、P添加处理之后树木胸径(DBH)、胸高断面积或生物量与处理之前的差值除以所对应的时间(Markewitz et al., 2012; Tian et al., 2017; Li et al., 2018).此外, 也有研究使用一段时间N、P添加处理之后DBH或胸高断面积与处理之前的差值除以处理之前的DBH或胸高断面积来表示树木的相对生长(Jiang et al., 2018).绝对增长量(表1)一般是用N、P添加处理一段时间内的DBH、胸高断面积、生物量(或生产力, 以碳储量计算)的绝对变化量来表示.生物量的变化是基于胸径-树高曲线、树木其他器官生物量与DBH和树高之间混合回归模型、异速生长方程等计算林分尺度总生物量的变化(Herbert & Fownes, 1995; Markewitz et al., 2012; Du & Fang, 2014; 周序力等, 2018).在森林根系生产力的估测中, 也有很多研究利用内生长环法(ingrowth core method)进行实测(Waring et al., 2019).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest 3 2006
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
... ), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
... (5)加强开展长期的和联网的N、P添加对树木生长影响的研究.一项长达30年的N添加研究表明, N添加对树木生长的影响随时间波动, 在前10年里, N添加显著促进了树木生长, 而随后的20年里N添加抑制了树木生长(Saarsalmi et al., 2012); 这种变化可能与植物对土壤N的周期性响应有关系.树木的生长周期较长, 短时间内小树对N、P添加的响应可能较为迅速, 而较长时期内才能观测到大树的响应; 因此, 需要长时间的观测才能得到更加精确的结果.然而, 当前大部分的养分添加实验持续时间都较短, 仅有个别长期实验(H?gberg et al., 2006; Saarsalmi et al., 2012; Rappe-George et al., 2013), 这限制了我们对自然长期N、P沉降背景下树木生长响应的认识.后续研究需要在全球范围内尤其是北方森林和热带森林不同类型森林中开展长期的N、P添加实验, 以提高我们对不同演替阶段森林的树木生长响应N、P沉降的认识.此外, 虽然当前在全球森林中已有一定数量的N、P添加实验, 但是这些实验的处理条件存在很大差异, 如养分添加量、养分种类、养分添加方式(溶液或者固体).这些不一致的实验处理条件会对实验结果产生一些影响(Schulte-?Uebbing & de Vries, 2018), 限制了我们在N、P添加对不同森林间树木生长及生产力影响的机制比较.目前, 草地中分布式协作性的实验(coordinated distributed experiment)养分添加网络(Nutrient Network, NutNet)已经开展(Borer et al., 2014), 它采用基本相同的实验设计、观测与采样手段, 已经取得了良好的成效, 这为森林等其他生态系统联网实验的开展提供了良好的借鉴.因此, 森林中的养分添加实验网络也亟待开展, 这将有助于我们加深对养分富集调控树木生长机制的认识与理解. ...
Impact of defoliation by Essigella californica on the growth of mature Pinus radiata and response to N, P and S fertilizer 1 2013
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
Global meta-analysis shows pervasive phosphorus limitation of aboveground plant production in natural terrestrial ecosystems 2 2020
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
A unifying framework for dinitrogen fixation in the terrestrial biosphere 1 2008
... 植物的固氮属性会调控N、P添加对树木生长的影响, 原因可能是固氮植物与非固氮植物具有不同的养分获取策略.Nasto等(2019)研究发现, 在土壤养分可利用性较低的热带森林土壤中, N和P共同添加(该研究中土壤P含量较低, 可能P添加起主要作用)促进了非固氮植物的生长; 但对固氮植物的生长影响不大.固氮植物在养分有限的情况下, 可以通过固氮作用满足自身对N的需求, 同时通过固氮还可以促进根系磷酸酶的产生, 增强植物对P的获取能力(Houlton et al., 2008; Marklein & Houlton, 2012; Nasto et al., 2014).在这种情况下, 与非固氮植物相比, N和P共同添加对固氮植物的生长并不表现出明显的促进作用(Nasto et al., 2019).此外, 有研究发现, 在光资源受限的时候, N添加对树木的共生固氮作用及其生长均没有显著影响; 而在光资源充足的时候, N添加抑制了树木共生固氮, 促进了其生长; 该研究结果揭示了N添加对固氮树木生长的影响也会受到光资源的调控(Taylor & Menge, 2018).与固氮植物相比, 在N和P添加下, 非固氮植物的养分限制得到缓解, 表现为N和P添加促进其生长.此外, 也有研究表明, P添加可以通过促进固氮植物的结瘤和共生固氮作用来促进植物的生长(Waring et al., 2019; Zheng et al., 2019).然而, 对于非固氮植物, P添加对其生长的影响可能比较复杂, 这是因为P添加虽然可以提高P的可利用性, 但会降低游离固氮作用(Zheng et al., 2019).因此, P添加对非固氮植物生长的影响可能取决于N的可利用性状态.总体而言, 在P可利用性较低时, 单独的P添加对固氮树木(或固氮树木占优势的森林)生长的促进作用较非固氮树木(或非固氮树木占优势的森林)大, 而单独的N添加对固氮与非固氮植物(或固氮与非固氮植物分别占优势的森林)生长的促进作用与单独的P添加相反. ...
Chronic nitrogen deposition alters tree allometric relationships: implications for biomass production and carbon storage 1 2016
... (2)研究树木各部分生长对N、P添加之后的分配与权衡.N、P添加常常会改变树木各部分(如凋落物、叶片、粗根、细根、枝干)的生产力比例(Du & Fang, 2014; BassiriRad et al., 2015; 毛宏蕊和金光泽, 2017), 即N、P添加影响树木生产力向各部分的分配策略.整合分析研究表明, N、P添加倾向于降低植物的地下生产力(Janssens et al., 2010; Peng et al., 2017)和地下部分与地上部分生物量之比(Lu et al., 2011b; Li et al., 2016; Peng & Yang, 2016).这种变化可能与植物在面对N、P可利用性提高时, 不需要将更多的光合作用产物分配到地下来获取养分, 而是产生更多的地上生物量有关.在这种情况下, 以胸径的变化衡量树木生长对养分添加的响应会存在一定的偏颇.然而, 即使是基于生产力的测定研究树木生长的响应, 绝大多数研究还是用一个不变的相对生长方程估测对照和施肥处理组的生产力变化, 同样也会使得结果与真实情况存在差异(Ibá?ez et al., 2016).因此, 后续的研究需要重视养分添加之后树木各部分生长之间的权衡与分配策略, 以提高对树木生长和森林生产力估测的精度. ...
Reduction of forest soil respiration in response to nitrogen deposition 2 2010
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
... (2)研究树木各部分生长对N、P添加之后的分配与权衡.N、P添加常常会改变树木各部分(如凋落物、叶片、粗根、细根、枝干)的生产力比例(Du & Fang, 2014; BassiriRad et al., 2015; 毛宏蕊和金光泽, 2017), 即N、P添加影响树木生产力向各部分的分配策略.整合分析研究表明, N、P添加倾向于降低植物的地下生产力(Janssens et al., 2010; Peng et al., 2017)和地下部分与地上部分生物量之比(Lu et al., 2011b; Li et al., 2016; Peng & Yang, 2016).这种变化可能与植物在面对N、P可利用性提高时, 不需要将更多的光合作用产物分配到地下来获取养分, 而是产生更多的地上生物量有关.在这种情况下, 以胸径的变化衡量树木生长对养分添加的响应会存在一定的偏颇.然而, 即使是基于生产力的测定研究树木生长的响应, 绝大多数研究还是用一个不变的相对生长方程估测对照和施肥处理组的生产力变化, 同样也会使得结果与真实情况存在差异(Ibá?ez et al., 2016).因此, 后续的研究需要重视养分添加之后树木各部分生长之间的权衡与分配策略, 以提高对树木生长和森林生产力估测的精度. ...
Interactive effects of nitrogen and phosphorus additions on plant growth vary with ecosystem type 2 2019
... 目前, 大部分的研究集中在单独的N添加和单独的P添加对树木生长和森林生产力的影响, 仅少数研究探讨了二者交互作用的影响(图1B).在N × P交互作用的影响方面, 部分研究发现, N和P添加之间不存在交互作用(Fisher et al., 2013; Goswami et al., 2018; Wright, 2019; Zhao & Zeng, 2019), 即N × P交互作用对树木生长和森林生产力不产生显著影响.也有研究表明, N和P添加之间存在交互作用, 这种交互作用在热带森林中表现为协同作用(Jiang et al., 2018, 2019; Li et al., 2018), 即N × P交互作用对树木生长和森林生产力表现为促进作用.N和P协同作用的产生机制可能与单一的N或P添加会加剧另一元素的限制有关(Elser et al., 2007; Li et al., 2016; Jiang et al., 2019).目前, N × P交互作用对树木生长和森林生产力影响的研究主要分布在热带森林中, 在温带森林和北方森林中的研究还十分有限(图1B), 相关机制需要进一步探讨. ...
... ; Jiang et al., 2019).目前, N × P交互作用对树木生长和森林生产力影响的研究主要分布在热带森林中, 在温带森林和北方森林中的研究还十分有限(图1B), 相关机制需要进一步探讨. ...
The response of tree growth to nitrogen and phosphorus additions in a tropical montane rainforest 7 2018
... 目前, 在N、P添加实验中, 树木生长和森林生产力的常用评估方法主要有两种: 一是相对生长速率, 二是绝对增长量.相对生长速率(表1)一般是用一段时间N、P添加处理之后树木胸径(DBH)、胸高断面积或生物量与处理之前的差值除以所对应的时间(Markewitz et al., 2012; Tian et al., 2017; Li et al., 2018).此外, 也有研究使用一段时间N、P添加处理之后DBH或胸高断面积与处理之前的差值除以处理之前的DBH或胸高断面积来表示树木的相对生长(Jiang et al., 2018).绝对增长量(表1)一般是用N、P添加处理一段时间内的DBH、胸高断面积、生物量(或生产力, 以碳储量计算)的绝对变化量来表示.生物量的变化是基于胸径-树高曲线、树木其他器官生物量与DBH和树高之间混合回归模型、异速生长方程等计算林分尺度总生物量的变化(Herbert & Fownes, 1995; Markewitz et al., 2012; Du & Fang, 2014; 周序力等, 2018).在森林根系生产力的估测中, 也有很多研究利用内生长环法(ingrowth core method)进行实测(Waring et al., 2019).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
... ).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
... Methods for assessing the effects of nitrogen and/or phosphorus addition on plant growth and productivity Table 1
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
... 目前, 大部分的研究集中在单独的N添加和单独的P添加对树木生长和森林生产力的影响, 仅少数研究探讨了二者交互作用的影响(图1B).在N × P交互作用的影响方面, 部分研究发现, N和P添加之间不存在交互作用(Fisher et al., 2013; Goswami et al., 2018; Wright, 2019; Zhao & Zeng, 2019), 即N × P交互作用对树木生长和森林生产力不产生显著影响.也有研究表明, N和P添加之间存在交互作用, 这种交互作用在热带森林中表现为协同作用(Jiang et al., 2018, 2019; Li et al., 2018), 即N × P交互作用对树木生长和森林生产力表现为促进作用.N和P协同作用的产生机制可能与单一的N或P添加会加剧另一元素的限制有关(Elser et al., 2007; Li et al., 2016; Jiang et al., 2019).目前, N × P交互作用对树木生长和森林生产力影响的研究主要分布在热带森林中, 在温带森林和北方森林中的研究还十分有限(图1B), 相关机制需要进一步探讨. ...
... 在生态系统内的某类植物个体或群体水平上(如小树与大树), N、P添加对树木生长的影响因树木径级的不同而存在差异.海南热带山地森林的一项为期5年的研究发现, P添加对小树(DBH ≤ 10 cm)和中树(10 cm < DBH ≤ 20 cm)的生长表现为显著的促进作用; 但对大树(DBH > 20 cm)的生长没有显著影响(Jiang et al., 2018).其他一些在热带森林中开展的实验也表明, N、P添加对树木生长的影响也因树木径级的不同而存在明显的差异(Alvarez-Clare et al., 2013; Fisher et al., 2013).在鸡公山温带森林的研究表明, N添加对小树和大树生长都没有显著的影响, 而P添加在小树中表现为显著的促进作用(Li et al., 2018).也有研究发现, N、P添加对树木生长的促进作用与树木的初始大小具有密切的正相关关系, 即在N、P添加下大树的生长要快于小树(Báez & Homeier, 2018).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... 目前, 关于N、P添加对树木生长和森林生产力的影响已经有很多相关研究, 一些新出现的机制或新发现, 拓展了我们对N、P沉降背景下森林生态系统生产力变化的认识.以前的观点认为, 在纬度梯度上热带森林生产力主要受P限制, 北方森林和温带森林生产力主要受N限制(Vitousek, 1984; Vitousek et al., 2010).然而, 也有研究指出在亚热带和温带的一些区域, 主要是小树而不是大树受到P的限制(Jiang et al., 2018; Li et al., 2018); 启示我们当评价森林生产力养分限制的时候需要考虑树木大小依赖的养分限制.从基于单一的树木径级和树木的某一生物性状(如固氮属性、共生菌根), 到基于定量的植物功能性状、植物和微生物之间的相互作用关系, 我们对树木生长响应N、P添加的理解逐渐深入.基于此, 我们对N、P添加对树木生长和森林生产力影响的研究进行了总结与归纳(图3), 该图涵盖了树木生长和森林生产力的评估方法、植物功能性状以及植物和微生物之间的关系, 为理解N、P添加影响树木生长提供了研究思路.尽管如此, 目前关于树木生长和生产力如何响应N、P添加仍然存在很多不清楚的方面, 限制了我们的深层认识, 未来需要在以下几个方面加强研究. ...
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed 4 2008
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: a meta-analysis 5 2016
... 目前, 大部分的研究集中在单独的N添加和单独的P添加对树木生长和森林生产力的影响, 仅少数研究探讨了二者交互作用的影响(图1B).在N × P交互作用的影响方面, 部分研究发现, N和P添加之间不存在交互作用(Fisher et al., 2013; Goswami et al., 2018; Wright, 2019; Zhao & Zeng, 2019), 即N × P交互作用对树木生长和森林生产力不产生显著影响.也有研究表明, N和P添加之间存在交互作用, 这种交互作用在热带森林中表现为协同作用(Jiang et al., 2018, 2019; Li et al., 2018), 即N × P交互作用对树木生长和森林生产力表现为促进作用.N和P协同作用的产生机制可能与单一的N或P添加会加剧另一元素的限制有关(Elser et al., 2007; Li et al., 2016; Jiang et al., 2019).目前, N × P交互作用对树木生长和森林生产力影响的研究主要分布在热带森林中, 在温带森林和北方森林中的研究还十分有限(图1B), 相关机制需要进一步探讨. ...
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
... ); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
... (2)研究树木各部分生长对N、P添加之后的分配与权衡.N、P添加常常会改变树木各部分(如凋落物、叶片、粗根、细根、枝干)的生产力比例(Du & Fang, 2014; BassiriRad et al., 2015; 毛宏蕊和金光泽, 2017), 即N、P添加影响树木生产力向各部分的分配策略.整合分析研究表明, N、P添加倾向于降低植物的地下生产力(Janssens et al., 2010; Peng et al., 2017)和地下部分与地上部分生物量之比(Lu et al., 2011b; Li et al., 2016; Peng & Yang, 2016).这种变化可能与植物在面对N、P可利用性提高时, 不需要将更多的光合作用产物分配到地下来获取养分, 而是产生更多的地上生物量有关.在这种情况下, 以胸径的变化衡量树木生长对养分添加的响应会存在一定的偏颇.然而, 即使是基于生产力的测定研究树木生长的响应, 绝大多数研究还是用一个不变的相对生长方程估测对照和施肥处理组的生产力变化, 同样也会使得结果与真实情况存在差异(Ibá?ez et al., 2016).因此, 后续的研究需要重视养分添加之后树木各部分生长之间的权衡与分配策略, 以提高对树木生长和森林生产力估测的精度. ...
Size-dependent nutrient limitation of tree growth from subtropical to cold temperate forests 9 2018
... 目前, 在N、P添加实验中, 树木生长和森林生产力的常用评估方法主要有两种: 一是相对生长速率, 二是绝对增长量.相对生长速率(表1)一般是用一段时间N、P添加处理之后树木胸径(DBH)、胸高断面积或生物量与处理之前的差值除以所对应的时间(Markewitz et al., 2012; Tian et al., 2017; Li et al., 2018).此外, 也有研究使用一段时间N、P添加处理之后DBH或胸高断面积与处理之前的差值除以处理之前的DBH或胸高断面积来表示树木的相对生长(Jiang et al., 2018).绝对增长量(表1)一般是用N、P添加处理一段时间内的DBH、胸高断面积、生物量(或生产力, 以碳储量计算)的绝对变化量来表示.生物量的变化是基于胸径-树高曲线、树木其他器官生物量与DBH和树高之间混合回归模型、异速生长方程等计算林分尺度总生物量的变化(Herbert & Fownes, 1995; Markewitz et al., 2012; Du & Fang, 2014; 周序力等, 2018).在森林根系生产力的估测中, 也有很多研究利用内生长环法(ingrowth core method)进行实测(Waring et al., 2019).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
... Methods for assessing the effects of nitrogen and/or phosphorus addition on plant growth and productivity Table 1
... 目前, 大部分的研究集中在单独的N添加和单独的P添加对树木生长和森林生产力的影响, 仅少数研究探讨了二者交互作用的影响(图1B).在N × P交互作用的影响方面, 部分研究发现, N和P添加之间不存在交互作用(Fisher et al., 2013; Goswami et al., 2018; Wright, 2019; Zhao & Zeng, 2019), 即N × P交互作用对树木生长和森林生产力不产生显著影响.也有研究表明, N和P添加之间存在交互作用, 这种交互作用在热带森林中表现为协同作用(Jiang et al., 2018, 2019; Li et al., 2018), 即N × P交互作用对树木生长和森林生产力表现为促进作用.N和P协同作用的产生机制可能与单一的N或P添加会加剧另一元素的限制有关(Elser et al., 2007; Li et al., 2016; Jiang et al., 2019).目前, N × P交互作用对树木生长和森林生产力影响的研究主要分布在热带森林中, 在温带森林和北方森林中的研究还十分有限(图1B), 相关机制需要进一步探讨. ...
... 在生态系统内的某类植物个体或群体水平上(如小树与大树), N、P添加对树木生长的影响因树木径级的不同而存在差异.海南热带山地森林的一项为期5年的研究发现, P添加对小树(DBH ≤ 10 cm)和中树(10 cm < DBH ≤ 20 cm)的生长表现为显著的促进作用; 但对大树(DBH > 20 cm)的生长没有显著影响(Jiang et al., 2018).其他一些在热带森林中开展的实验也表明, N、P添加对树木生长的影响也因树木径级的不同而存在明显的差异(Alvarez-Clare et al., 2013; Fisher et al., 2013).在鸡公山温带森林的研究表明, N添加对小树和大树生长都没有显著的影响, 而P添加在小树中表现为显著的促进作用(Li et al., 2018).也有研究发现, N、P添加对树木生长的促进作用与树木的初始大小具有密切的正相关关系, 即在N、P添加下大树的生长要快于小树(Báez & Homeier, 2018).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... ; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... ; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... 尽管有很多研究发现, N、P添加能够促进生态系统尺度上的树木生长和森林生产力, 但是N、P添加对它们的抑制作用在实验中也经常被观测到(Harpole et al., 2011; Li et al., 2018; Nasto et al., 2019).因此, 这对根据N、P添加是否促进生长从而判断生态系统或树木N、P的限制提出了挑战.以往的研究表明, 长期的N、P添加能够导致土壤pH值下降, 使得土壤酸化(Tian & Niu, 2015; Dai et al., 2018).尽管N、P添加所产生的负效应能够从过量的养分添加所产生的土壤酸化等毒害作用进行解释, 但是当没有毒害作用发生时, N、P添加依旧对树木生长产生负效应(?apek et al., 2018).针对这种情况, ?apek等(2018)提出了一个基于植物和微生物相互作用(plant-microbe interaction)的新的概念框架模型来解释并预测在生态系统尺度上N、P添加对初级生产力的影响.在该化学计量框架模型中, 植物与微生物之间的相互作用包括两种情况: 一是竞争关系; 二是协作关系, 如植物通过根系向土壤中分泌活性有机碳, 促进微生物的生长, 而微生物矿化有机质为植物提供N或P.该概念框架模型认为, 在一个生态系统中存在3种N、P的限制模式, 一是对于植物和土壤微生物都是N限制; 二是对于植物为N限制, 但是对于土壤微生物是P限制; 三是对于植物和土壤微生物都是P限制.简单来说, 如果一个系统中, 植物和微生物之间是协作关系占主导, 添加对植物生长非限制性的养分可以通过促进受该养分限制的微生物生长而间接提高植物的营养和生长; 相反, 在植物和微生物间是竞争作用占主导的系统内, 植物会受到多种养分的共同限制(?apek et al., 2018).该概念框架模型能够很好地解释生态系统生产力对N、P添加的响应, 并对以往研究中难以解释的响应提供了解决思路. ...
... 目前, 关于N、P添加对树木生长和森林生产力的影响已经有很多相关研究, 一些新出现的机制或新发现, 拓展了我们对N、P沉降背景下森林生态系统生产力变化的认识.以前的观点认为, 在纬度梯度上热带森林生产力主要受P限制, 北方森林和温带森林生产力主要受N限制(Vitousek, 1984; Vitousek et al., 2010).然而, 也有研究指出在亚热带和温带的一些区域, 主要是小树而不是大树受到P的限制(Jiang et al., 2018; Li et al., 2018); 启示我们当评价森林生产力养分限制的时候需要考虑树木大小依赖的养分限制.从基于单一的树木径级和树木的某一生物性状(如固氮属性、共生菌根), 到基于定量的植物功能性状、植物和微生物之间的相互作用关系, 我们对树木生长响应N、P添加的理解逐渐深入.基于此, 我们对N、P添加对树木生长和森林生产力影响的研究进行了总结与归纳(图3), 该图涵盖了树木生长和森林生产力的评估方法、植物功能性状以及植物和微生物之间的关系, 为理解N、P添加影响树木生长提供了研究思路.尽管如此, 目前关于树木生长和生产力如何响应N、P添加仍然存在很多不清楚的方面, 限制了我们的深层认识, 未来需要在以下几个方面加强研究. ...
... (1)加强树木生长响应N、P添加的生理学机制研究.一项在N富集的热带森林中的N添加实验表明, 10年左右的N添加使土壤发生了酸化, 降低了土壤中的盐基离子(钙和镁)的可利用性, 但是树木能够通过增强叶片蒸腾作用提高对盐基离子的吸收来维持自身的养分平衡(Lu et al., 2018).N和P的可利用性提高后, 植物的生理活动会发生相应的响应, 这些响应可能会对树木的生长产生影响, 并最终反应到DBH和生物量变化上.目前, 仅有少数研究(Zhu et al., 2014; Li et al., 2018; Lu et al., 2018)对N、P添加后的树木生理水平上的响应(光合作用、蒸腾作用、叶片养分重吸收效率等)进行了测定.因此, 后续研究可以从植物的生理角度开展, 如光合作用、水分传输、根系寿命等, 以期能够提供深入的机理解释. ...
落叶松原始林树木生长对氮添加的响应 4 2015
... 在生态系统内的某类植物个体或群体水平上(如小树与大树), N、P添加对树木生长的影响因树木径级的不同而存在差异.海南热带山地森林的一项为期5年的研究发现, P添加对小树(DBH ≤ 10 cm)和中树(10 cm < DBH ≤ 20 cm)的生长表现为显著的促进作用; 但对大树(DBH > 20 cm)的生长没有显著影响(Jiang et al., 2018).其他一些在热带森林中开展的实验也表明, N、P添加对树木生长的影响也因树木径级的不同而存在明显的差异(Alvarez-Clare et al., 2013; Fisher et al., 2013).在鸡公山温带森林的研究表明, N添加对小树和大树生长都没有显著的影响, 而P添加在小树中表现为显著的促进作用(Li et al., 2018).也有研究发现, N、P添加对树木生长的促进作用与树木的初始大小具有密切的正相关关系, 即在N、P添加下大树的生长要快于小树(Báez & Homeier, 2018).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... ), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
... ), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
落叶松原始林树木生长对氮添加的响应 4 2015
... 在生态系统内的某类植物个体或群体水平上(如小树与大树), N、P添加对树木生长的影响因树木径级的不同而存在差异.海南热带山地森林的一项为期5年的研究发现, P添加对小树(DBH ≤ 10 cm)和中树(10 cm < DBH ≤ 20 cm)的生长表现为显著的促进作用; 但对大树(DBH > 20 cm)的生长没有显著影响(Jiang et al., 2018).其他一些在热带森林中开展的实验也表明, N、P添加对树木生长的影响也因树木径级的不同而存在明显的差异(Alvarez-Clare et al., 2013; Fisher et al., 2013).在鸡公山温带森林的研究表明, N添加对小树和大树生长都没有显著的影响, 而P添加在小树中表现为显著的促进作用(Li et al., 2018).也有研究发现, N、P添加对树木生长的促进作用与树木的初始大小具有密切的正相关关系, 即在N、P添加下大树的生长要快于小树(Báez & Homeier, 2018).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... ), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
... ), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
Responses of ecosystem nitrogen cycle to nitrogen addition: a meta-analysis 1 2011a
... 在各种类型的生态系统中, 森林占到全球陆地面积的30%, 其每年的碳吸收量占到全球陆地碳汇总量的82%, 在全球碳循环中占据着重要地位(Pan et al., 2011).自工业革命以来, 由于人为活动, 如施用矿质肥料、生物质燃烧等, 使得输入到生物圈的N和P呈现快速上升的趋势.N和P的沉降量在2000年的时候分别达到220和12 Tg·a-1 (Pe?uelas et al., 2013), 预计在未来将呈持续上升的趋势.N和P的持续输入会使得生态系统中的N、P可利用性提高(Lu et al., 2011a; Feng & Zhu, 2019), 并对生态系统碳循环过程产生显著影响, 如树木生长、生产力及土壤碳库(Lu et al., 2011b; Yue et al., 2017).森林树木生长及其生产力的变化对于预测全球变化背景下的碳循环过程和陆地碳汇潜力至关重要.目前, N、P沉降如何影响树木生长和森林生产力成为全球变化领域的研究热点(Elser et al., 2007; Camarero & Carrer, 2017; DeForest & Snell, 2020). ...
Minor stimulation of soil carbon storage by nitrogen addition: a meta-analysis 2 2011b
... 在各种类型的生态系统中, 森林占到全球陆地面积的30%, 其每年的碳吸收量占到全球陆地碳汇总量的82%, 在全球碳循环中占据着重要地位(Pan et al., 2011).自工业革命以来, 由于人为活动, 如施用矿质肥料、生物质燃烧等, 使得输入到生物圈的N和P呈现快速上升的趋势.N和P的沉降量在2000年的时候分别达到220和12 Tg·a-1 (Pe?uelas et al., 2013), 预计在未来将呈持续上升的趋势.N和P的持续输入会使得生态系统中的N、P可利用性提高(Lu et al., 2011a; Feng & Zhu, 2019), 并对生态系统碳循环过程产生显著影响, 如树木生长、生产力及土壤碳库(Lu et al., 2011b; Yue et al., 2017).森林树木生长及其生产力的变化对于预测全球变化背景下的碳循环过程和陆地碳汇潜力至关重要.目前, N、P沉降如何影响树木生长和森林生产力成为全球变化领域的研究热点(Elser et al., 2007; Camarero & Carrer, 2017; DeForest & Snell, 2020). ...
... (2)研究树木各部分生长对N、P添加之后的分配与权衡.N、P添加常常会改变树木各部分(如凋落物、叶片、粗根、细根、枝干)的生产力比例(Du & Fang, 2014; BassiriRad et al., 2015; 毛宏蕊和金光泽, 2017), 即N、P添加影响树木生产力向各部分的分配策略.整合分析研究表明, N、P添加倾向于降低植物的地下生产力(Janssens et al., 2010; Peng et al., 2017)和地下部分与地上部分生物量之比(Lu et al., 2011b; Li et al., 2016; Peng & Yang, 2016).这种变化可能与植物在面对N、P可利用性提高时, 不需要将更多的光合作用产物分配到地下来获取养分, 而是产生更多的地上生物量有关.在这种情况下, 以胸径的变化衡量树木生长对养分添加的响应会存在一定的偏颇.然而, 即使是基于生产力的测定研究树木生长的响应, 绝大多数研究还是用一个不变的相对生长方程估测对照和施肥处理组的生产力变化, 同样也会使得结果与真实情况存在差异(Ibá?ez et al., 2016).因此, 后续的研究需要重视养分添加之后树木各部分生长之间的权衡与分配策略, 以提高对树木生长和森林生产力估测的精度. ...
Plant acclimation to long-term high nitrogen deposition in an N-rich tropical forest 4 2018
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
... (1)加强树木生长响应N、P添加的生理学机制研究.一项在N富集的热带森林中的N添加实验表明, 10年左右的N添加使土壤发生了酸化, 降低了土壤中的盐基离子(钙和镁)的可利用性, 但是树木能够通过增强叶片蒸腾作用提高对盐基离子的吸收来维持自身的养分平衡(Lu et al., 2018).N和P的可利用性提高后, 植物的生理活动会发生相应的响应, 这些响应可能会对树木的生长产生影响, 并最终反应到DBH和生物量变化上.目前, 仅有少数研究(Zhu et al., 2014; Li et al., 2018; Lu et al., 2018)对N、P添加后的树木生理水平上的响应(光合作用、蒸腾作用、叶片养分重吸收效率等)进行了测定.因此, 后续研究可以从植物的生理角度开展, 如光合作用、水分传输、根系寿命等, 以期能够提供深入的机理解释. ...
... ; Lu et al., 2018)对N、P添加后的树木生理水平上的响应(光合作用、蒸腾作用、叶片养分重吸收效率等)进行了测定.因此, 后续研究可以从植物的生理角度开展, 如光合作用、水分传输、根系寿命等, 以期能够提供深入的机理解释. ...
Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide 1 2004
... 养分限制模式对于生态系统生产力及其对全球变化的响应至关重要(Reich et al., 2006; Crowley et al., 2012).在全球尺度上, 氮(N)和磷(P)对植物生产力的限制被认为普遍存在于陆地、淡水和海洋生态系统中(Elser et al., 2007).在大气CO2浓度持续上升背景下, 植物生长受到促进, 从而会从土壤中摄取更多的养分, 进而导致其生长逐渐受到养分可利用性的限制(Luo et al., 2004).模型预测结果表明, 在考虑N、P可利用性限制的情况下, 净初级生产力的增加将低于N和P不受限制状态下的增加量, 而陆地生态系统也将由碳吸收转变为碳排放(Wieder et al., 2015).由此可见, 养分可利用性在调节陆地净初级生产力和陆地碳库中发挥着至关重要的作用. ...
The human footprint in the carbon cycle of temperate and boreal forests 2 2007
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
Three-year growth response of young Douglas-fir to nitrogen, calcium, phosphorus, and blended fertilizers in Oregon and Washington 1 2014
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
氮添加对典型阔叶红松林净初级生产力的影响 1 2017
... (2)研究树木各部分生长对N、P添加之后的分配与权衡.N、P添加常常会改变树木各部分(如凋落物、叶片、粗根、细根、枝干)的生产力比例(Du & Fang, 2014; BassiriRad et al., 2015; 毛宏蕊和金光泽, 2017), 即N、P添加影响树木生产力向各部分的分配策略.整合分析研究表明, N、P添加倾向于降低植物的地下生产力(Janssens et al., 2010; Peng et al., 2017)和地下部分与地上部分生物量之比(Lu et al., 2011b; Li et al., 2016; Peng & Yang, 2016).这种变化可能与植物在面对N、P可利用性提高时, 不需要将更多的光合作用产物分配到地下来获取养分, 而是产生更多的地上生物量有关.在这种情况下, 以胸径的变化衡量树木生长对养分添加的响应会存在一定的偏颇.然而, 即使是基于生产力的测定研究树木生长的响应, 绝大多数研究还是用一个不变的相对生长方程估测对照和施肥处理组的生产力变化, 同样也会使得结果与真实情况存在差异(Ibá?ez et al., 2016).因此, 后续的研究需要重视养分添加之后树木各部分生长之间的权衡与分配策略, 以提高对树木生长和森林生产力估测的精度. ...
氮添加对典型阔叶红松林净初级生产力的影响 1 2017
... (2)研究树木各部分生长对N、P添加之后的分配与权衡.N、P添加常常会改变树木各部分(如凋落物、叶片、粗根、细根、枝干)的生产力比例(Du & Fang, 2014; BassiriRad et al., 2015; 毛宏蕊和金光泽, 2017), 即N、P添加影响树木生产力向各部分的分配策略.整合分析研究表明, N、P添加倾向于降低植物的地下生产力(Janssens et al., 2010; Peng et al., 2017)和地下部分与地上部分生物量之比(Lu et al., 2011b; Li et al., 2016; Peng & Yang, 2016).这种变化可能与植物在面对N、P可利用性提高时, 不需要将更多的光合作用产物分配到地下来获取养分, 而是产生更多的地上生物量有关.在这种情况下, 以胸径的变化衡量树木生长对养分添加的响应会存在一定的偏颇.然而, 即使是基于生产力的测定研究树木生长的响应, 绝大多数研究还是用一个不变的相对生长方程估测对照和施肥处理组的生产力变化, 同样也会使得结果与真实情况存在差异(Ibá?ez et al., 2016).因此, 后续的研究需要重视养分添加之后树木各部分生长之间的权衡与分配策略, 以提高对树木生长和森林生产力估测的精度. ...
Soil and tree response to P fertilization in a secondary tropical forest supported by an Oxisol 4 2012
... 目前, 在N、P添加实验中, 树木生长和森林生产力的常用评估方法主要有两种: 一是相对生长速率, 二是绝对增长量.相对生长速率(表1)一般是用一段时间N、P添加处理之后树木胸径(DBH)、胸高断面积或生物量与处理之前的差值除以所对应的时间(Markewitz et al., 2012; Tian et al., 2017; Li et al., 2018).此外, 也有研究使用一段时间N、P添加处理之后DBH或胸高断面积与处理之前的差值除以处理之前的DBH或胸高断面积来表示树木的相对生长(Jiang et al., 2018).绝对增长量(表1)一般是用N、P添加处理一段时间内的DBH、胸高断面积、生物量(或生产力, 以碳储量计算)的绝对变化量来表示.生物量的变化是基于胸径-树高曲线、树木其他器官生物量与DBH和树高之间混合回归模型、异速生长方程等计算林分尺度总生物量的变化(Herbert & Fownes, 1995; Markewitz et al., 2012; Du & Fang, 2014; 周序力等, 2018).在森林根系生产力的估测中, 也有很多研究利用内生长环法(ingrowth core method)进行实测(Waring et al., 2019).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
... ; Markewitz et al., 2012; Du & Fang, 2014; 周序力等, 2018).在森林根系生产力的估测中, 也有很多研究利用内生长环法(ingrowth core method)进行实测(Waring et al., 2019).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
... Methods for assessing the effects of nitrogen and/or phosphorus addition on plant growth and productivity Table 1
... 植物的固氮属性会调控N、P添加对树木生长的影响, 原因可能是固氮植物与非固氮植物具有不同的养分获取策略.Nasto等(2019)研究发现, 在土壤养分可利用性较低的热带森林土壤中, N和P共同添加(该研究中土壤P含量较低, 可能P添加起主要作用)促进了非固氮植物的生长; 但对固氮植物的生长影响不大.固氮植物在养分有限的情况下, 可以通过固氮作用满足自身对N的需求, 同时通过固氮还可以促进根系磷酸酶的产生, 增强植物对P的获取能力(Houlton et al., 2008; Marklein & Houlton, 2012; Nasto et al., 2014).在这种情况下, 与非固氮植物相比, N和P共同添加对固氮植物的生长并不表现出明显的促进作用(Nasto et al., 2019).此外, 有研究发现, 在光资源受限的时候, N添加对树木的共生固氮作用及其生长均没有显著影响; 而在光资源充足的时候, N添加抑制了树木共生固氮, 促进了其生长; 该研究结果揭示了N添加对固氮树木生长的影响也会受到光资源的调控(Taylor & Menge, 2018).与固氮植物相比, 在N和P添加下, 非固氮植物的养分限制得到缓解, 表现为N和P添加促进其生长.此外, 也有研究表明, P添加可以通过促进固氮植物的结瘤和共生固氮作用来促进植物的生长(Waring et al., 2019; Zheng et al., 2019).然而, 对于非固氮植物, P添加对其生长的影响可能比较复杂, 这是因为P添加虽然可以提高P的可利用性, 但会降低游离固氮作用(Zheng et al., 2019).因此, P添加对非固氮植物生长的影响可能取决于N的可利用性状态.总体而言, 在P可利用性较低时, 单独的P添加对固氮树木(或固氮树木占优势的森林)生长的促进作用较非固氮树木(或非固氮树木占优势的森林)大, 而单独的N添加对固氮与非固氮植物(或固氮与非固氮植物分别占优势的森林)生长的促进作用与单独的P添加相反. ...
... (3)深度研究植物功能性状在调节与预测树木生长对N、P添加响应中的作用.尽管已有相关研究从植物功能性状的角度研究了N、P添加如何影响树木生长, 但这些研究还依旧十分匮乏, 而且现有的研究也仅仅考虑了单一的植物功能性状(Thomas et al., 2010; Averill et al., 2018; Nasto et al., 2019).植物的功能性状具有多个维度, 而且这些性状常常是交织在一起, 比如植物可以同时具备固氮和共生菌根的特性, 固氮植物具有较高的AM菌根侵染率等(Nasto et al., 2014).在这种情况下, 考虑单一的功能性状可能会使研究结果受到其他因素的干扰.因此, 在未来研究中需要综合考虑植物各个部分的性状, 并采用较为复杂全面的统计方法定量分析植物功能性状在调控树木生长响应N、P添加中的作用. ...
Nutrient acquisition strategies augment growth in tropical N2-fixing trees in nutrient-poor soil and under elevated CO2 5 2019
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
... 植物的固氮属性会调控N、P添加对树木生长的影响, 原因可能是固氮植物与非固氮植物具有不同的养分获取策略.Nasto等(2019)研究发现, 在土壤养分可利用性较低的热带森林土壤中, N和P共同添加(该研究中土壤P含量较低, 可能P添加起主要作用)促进了非固氮植物的生长; 但对固氮植物的生长影响不大.固氮植物在养分有限的情况下, 可以通过固氮作用满足自身对N的需求, 同时通过固氮还可以促进根系磷酸酶的产生, 增强植物对P的获取能力(Houlton et al., 2008; Marklein & Houlton, 2012; Nasto et al., 2014).在这种情况下, 与非固氮植物相比, N和P共同添加对固氮植物的生长并不表现出明显的促进作用(Nasto et al., 2019).此外, 有研究发现, 在光资源受限的时候, N添加对树木的共生固氮作用及其生长均没有显著影响; 而在光资源充足的时候, N添加抑制了树木共生固氮, 促进了其生长; 该研究结果揭示了N添加对固氮树木生长的影响也会受到光资源的调控(Taylor & Menge, 2018).与固氮植物相比, 在N和P添加下, 非固氮植物的养分限制得到缓解, 表现为N和P添加促进其生长.此外, 也有研究表明, P添加可以通过促进固氮植物的结瘤和共生固氮作用来促进植物的生长(Waring et al., 2019; Zheng et al., 2019).然而, 对于非固氮植物, P添加对其生长的影响可能比较复杂, 这是因为P添加虽然可以提高P的可利用性, 但会降低游离固氮作用(Zheng et al., 2019).因此, P添加对非固氮植物生长的影响可能取决于N的可利用性状态.总体而言, 在P可利用性较低时, 单独的P添加对固氮树木(或固氮树木占优势的森林)生长的促进作用较非固氮树木(或非固氮树木占优势的森林)大, 而单独的N添加对固氮与非固氮植物(或固氮与非固氮植物分别占优势的森林)生长的促进作用与单独的P添加相反. ...
Model-data synthesis for the next generation of forest free-air CO2 enrichment (FACE) experiments 1 2016
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
A large and persistent carbon sink in the world?s forests 1 2011
... 在各种类型的生态系统中, 森林占到全球陆地面积的30%, 其每年的碳吸收量占到全球陆地碳汇总量的82%, 在全球碳循环中占据着重要地位(Pan et al., 2011).自工业革命以来, 由于人为活动, 如施用矿质肥料、生物质燃烧等, 使得输入到生物圈的N和P呈现快速上升的趋势.N和P的沉降量在2000年的时候分别达到220和12 Tg·a-1 (Pe?uelas et al., 2013), 预计在未来将呈持续上升的趋势.N和P的持续输入会使得生态系统中的N、P可利用性提高(Lu et al., 2011a; Feng & Zhu, 2019), 并对生态系统碳循环过程产生显著影响, 如树木生长、生产力及土壤碳库(Lu et al., 2011b; Yue et al., 2017).森林树木生长及其生产力的变化对于预测全球变化背景下的碳循环过程和陆地碳汇潜力至关重要.目前, N、P沉降如何影响树木生长和森林生产力成为全球变化领域的研究热点(Elser et al., 2007; Camarero & Carrer, 2017; DeForest & Snell, 2020). ...
Global patterns of root dynamics under nitrogen enrichment 1 2017
... (2)研究树木各部分生长对N、P添加之后的分配与权衡.N、P添加常常会改变树木各部分(如凋落物、叶片、粗根、细根、枝干)的生产力比例(Du & Fang, 2014; BassiriRad et al., 2015; 毛宏蕊和金光泽, 2017), 即N、P添加影响树木生产力向各部分的分配策略.整合分析研究表明, N、P添加倾向于降低植物的地下生产力(Janssens et al., 2010; Peng et al., 2017)和地下部分与地上部分生物量之比(Lu et al., 2011b; Li et al., 2016; Peng & Yang, 2016).这种变化可能与植物在面对N、P可利用性提高时, 不需要将更多的光合作用产物分配到地下来获取养分, 而是产生更多的地上生物量有关.在这种情况下, 以胸径的变化衡量树木生长对养分添加的响应会存在一定的偏颇.然而, 即使是基于生产力的测定研究树木生长的响应, 绝大多数研究还是用一个不变的相对生长方程估测对照和施肥处理组的生产力变化, 同样也会使得结果与真实情况存在差异(Ibá?ez et al., 2016).因此, 后续的研究需要重视养分添加之后树木各部分生长之间的权衡与分配策略, 以提高对树木生长和森林生产力估测的精度. ...
Allometric biomass partitioning under nitrogen enrichment: evidence from manipulative experiments around the world 1 2016
... (2)研究树木各部分生长对N、P添加之后的分配与权衡.N、P添加常常会改变树木各部分(如凋落物、叶片、粗根、细根、枝干)的生产力比例(Du & Fang, 2014; BassiriRad et al., 2015; 毛宏蕊和金光泽, 2017), 即N、P添加影响树木生产力向各部分的分配策略.整合分析研究表明, N、P添加倾向于降低植物的地下生产力(Janssens et al., 2010; Peng et al., 2017)和地下部分与地上部分生物量之比(Lu et al., 2011b; Li et al., 2016; Peng & Yang, 2016).这种变化可能与植物在面对N、P可利用性提高时, 不需要将更多的光合作用产物分配到地下来获取养分, 而是产生更多的地上生物量有关.在这种情况下, 以胸径的变化衡量树木生长对养分添加的响应会存在一定的偏颇.然而, 即使是基于生产力的测定研究树木生长的响应, 绝大多数研究还是用一个不变的相对生长方程估测对照和施肥处理组的生产力变化, 同样也会使得结果与真实情况存在差异(Ibá?ez et al., 2016).因此, 后续的研究需要重视养分添加之后树木各部分生长之间的权衡与分配策略, 以提高对树木生长和森林生产力估测的精度. ...
Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe 2 2013
... 在各种类型的生态系统中, 森林占到全球陆地面积的30%, 其每年的碳吸收量占到全球陆地碳汇总量的82%, 在全球碳循环中占据着重要地位(Pan et al., 2011).自工业革命以来, 由于人为活动, 如施用矿质肥料、生物质燃烧等, 使得输入到生物圈的N和P呈现快速上升的趋势.N和P的沉降量在2000年的时候分别达到220和12 Tg·a-1 (Pe?uelas et al., 2013), 预计在未来将呈持续上升的趋势.N和P的持续输入会使得生态系统中的N、P可利用性提高(Lu et al., 2011a; Feng & Zhu, 2019), 并对生态系统碳循环过程产生显著影响, 如树木生长、生产力及土壤碳库(Lu et al., 2011b; Yue et al., 2017).森林树木生长及其生产力的变化对于预测全球变化背景下的碳循环过程和陆地碳汇潜力至关重要.目前, N、P沉降如何影响树木生长和森林生产力成为全球变化领域的研究热点(Elser et al., 2007; Camarero & Carrer, 2017; DeForest & Snell, 2020). ...
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
The impact of four decades of annual nitrogen addition on dissolved organic matter in a boreal forest soil 4 2013
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
... 在生态系统尺度上, N、P添加对树木生长和森林生产力的影响在不同的林龄中也不尽相同, 受到林龄的调控.Goswami等(2018)在温带森林的研究发现, N添加对幼龄林生长的促进作用强于中龄林和成熟林; P添加在中龄林中的促进作用最为明显, 而在幼龄林和成熟林中均没有明显的作用.然而, 也有研究发现, N、P添加对次生林(林龄较小)的生长表现为促进, 而对成熟林没有显著影响, 即次生林的生长响应明显高于成熟林(Wright et al., 2018; Wright, 2019).也有定量研究发现, N添加对树木生长的影响随着年龄的增大而显著降低(Vadeboncoeur, 2010), 表明幼龄林对N添加的响应更为强烈.总体而言, 目前的研究结果表明, N添加对幼龄林的促进作用一般高于中龄林和老龄林(Vadeboncoeur, 2010; Rappe-George et al., 2013; Schulte-Uebbing & de Vries, 2018), 因为幼龄林树木在吸收添加的养分之后, 能够快速地进行光合作用, 合成自身新陈代谢所需要的有机物, 生长相对较快.Wright等(2018)认为老龄林生长对N、P添加响应较弱的原因可能存在于两个方面: 一方面, 生活在养分可利用性较低环境中的物种已经适应了这种养分缺乏的条件, 在面对较好的养分条件时其增加生长的潜力有限(Coley et al., 1985), 群落生长的响应可能需要等到物种组成发生变化以适应较高养分可用性时才能体现出来(Chapin III et al., 1986); 另一方面, 植物在施肥条件下其组织中的N含量升高, 会增加其受到天敌(食草动物和其他害虫)危害的风险并消耗其初级生产力, 从而对树木生长的响应产生不利影响(Andersen et al., 2010). ...
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
... (5)加强开展长期的和联网的N、P添加对树木生长影响的研究.一项长达30年的N添加研究表明, N添加对树木生长的影响随时间波动, 在前10年里, N添加显著促进了树木生长, 而随后的20年里N添加抑制了树木生长(Saarsalmi et al., 2012); 这种变化可能与植物对土壤N的周期性响应有关系.树木的生长周期较长, 短时间内小树对N、P添加的响应可能较为迅速, 而较长时期内才能观测到大树的响应; 因此, 需要长时间的观测才能得到更加精确的结果.然而, 当前大部分的养分添加实验持续时间都较短, 仅有个别长期实验(H?gberg et al., 2006; Saarsalmi et al., 2012; Rappe-George et al., 2013), 这限制了我们对自然长期N、P沉降背景下树木生长响应的认识.后续研究需要在全球范围内尤其是北方森林和热带森林不同类型森林中开展长期的N、P添加实验, 以提高我们对不同演替阶段森林的树木生长响应N、P沉降的认识.此外, 虽然当前在全球森林中已有一定数量的N、P添加实验, 但是这些实验的处理条件存在很大差异, 如养分添加量、养分种类、养分添加方式(溶液或者固体).这些不一致的实验处理条件会对实验结果产生一些影响(Schulte-?Uebbing & de Vries, 2018), 限制了我们在N、P添加对不同森林间树木生长及生产力影响的机制比较.目前, 草地中分布式协作性的实验(coordinated distributed experiment)养分添加网络(Nutrient Network, NutNet)已经开展(Borer et al., 2014), 它采用基本相同的实验设计、观测与采样手段, 已经取得了良好的成效, 这为森林等其他生态系统联网实验的开展提供了良好的借鉴.因此, 森林中的养分添加实验网络也亟待开展, 这将有助于我们加深对养分富集调控树木生长机制的认识与理解. ...
Carbon-nitrogen interactions in terrestrial ecosystems in response to rising atmospheric carbon dioxide 1 2006
... 养分限制模式对于生态系统生产力及其对全球变化的响应至关重要(Reich et al., 2006; Crowley et al., 2012).在全球尺度上, 氮(N)和磷(P)对植物生产力的限制被认为普遍存在于陆地、淡水和海洋生态系统中(Elser et al., 2007).在大气CO2浓度持续上升背景下, 植物生长受到促进, 从而会从土壤中摄取更多的养分, 进而导致其生长逐渐受到养分可利用性的限制(Luo et al., 2004).模型预测结果表明, 在考虑N、P可利用性限制的情况下, 净初级生产力的增加将低于N和P不受限制状态下的增加量, 而陆地生态系统也将由碳吸收转变为碳排放(Wieder et al., 2015).由此可见, 养分可利用性在调节陆地净初级生产力和陆地碳库中发挥着至关重要的作用. ...
Phosphorus cycling in deciduous forest soil differs between stands dominated by ecto- and arbuscular mycorrhizal trees 1 2016
30-year effects of wood ash and nitrogen fertilization on soil chemical properties, soil microbial processes and stand growth in a Scots pine stand 3 2012
... Methods for assessing the effects of nitrogen and/or phosphorus addition on plant growth and productivity Table 1
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
... (5)加强开展长期的和联网的N、P添加对树木生长影响的研究.一项长达30年的N添加研究表明, N添加对树木生长的影响随时间波动, 在前10年里, N添加显著促进了树木生长, 而随后的20年里N添加抑制了树木生长(Saarsalmi et al., 2012); 这种变化可能与植物对土壤N的周期性响应有关系.树木的生长周期较长, 短时间内小树对N、P添加的响应可能较为迅速, 而较长时期内才能观测到大树的响应; 因此, 需要长时间的观测才能得到更加精确的结果.然而, 当前大部分的养分添加实验持续时间都较短, 仅有个别长期实验(H?gberg et al., 2006; Saarsalmi et al., 2012; Rappe-George et al., 2013), 这限制了我们对自然长期N、P沉降背景下树木生长响应的认识.后续研究需要在全球范围内尤其是北方森林和热带森林不同类型森林中开展长期的N、P添加实验, 以提高我们对不同演替阶段森林的树木生长响应N、P沉降的认识.此外, 虽然当前在全球森林中已有一定数量的N、P添加实验, 但是这些实验的处理条件存在很大差异, 如养分添加量、养分种类、养分添加方式(溶液或者固体).这些不一致的实验处理条件会对实验结果产生一些影响(Schulte-?Uebbing & de Vries, 2018), 限制了我们在N、P添加对不同森林间树木生长及生产力影响的机制比较.目前, 草地中分布式协作性的实验(coordinated distributed experiment)养分添加网络(Nutrient Network, NutNet)已经开展(Borer et al., 2014), 它采用基本相同的实验设计、观测与采样手段, 已经取得了良好的成效, 这为森林等其他生态系统联网实验的开展提供了良好的借鉴.因此, 森林中的养分添加实验网络也亟待开展, 这将有助于我们加深对养分富集调控树木生长机制的认识与理解. ...
Decomposers in disguise: mycorrhizal fungi as regulators of soil C dynamics in ecosystems under global change 1 2008
... 在菌根类型上, 不同研究发现, 与丛枝菌根(AM)真菌共生的树种(AM树种)和与外生菌根(ECM)真菌共生的树种(ECM树种)在N、P沉降下的响应存在差异, 表明菌根类型会调控N、P添加对树木生长和森林生产力的影响(Thomas et al., 2010; Averill et al., 2018; DeForest & Snell, 2020).在N添加研究方面, 基于美国森林资源清查数据的研究表明, N沉降促进了11个物种的生长, 抑制了3个物种的生长, 而且AM树种全部表现为促进作用(Thomas et al., 2010).最新的一项研究表明, 在N水平低的时候, ECM树种的生长对N的响应要高于AM树种, 而在N水平高的时候, AM树种的生长要高于ECM树种(Averill et al., 2018).AM树种和ECM树种对N可利用性增加的差异性响应, 可能与它们对N的获取与利用策略有关系.与ECM真菌相比, AM真菌缺少分解土壤有机质的酶, 主要获取土壤中的无机N来满足新陈代谢的需要和向植物输送养分(Chalot & Brun, 1998; Talbot et al., 2008).因此, 当活性N沉降导致土壤N的可利用性提高时, AM树种能够比ECM树种获得更多的益处使生长加快. ...
Light regulates tropical symbiotic nitrogen fixation more strongly than soil nitrogen 1 2018
... 植物的固氮属性会调控N、P添加对树木生长的影响, 原因可能是固氮植物与非固氮植物具有不同的养分获取策略.Nasto等(2019)研究发现, 在土壤养分可利用性较低的热带森林土壤中, N和P共同添加(该研究中土壤P含量较低, 可能P添加起主要作用)促进了非固氮植物的生长; 但对固氮植物的生长影响不大.固氮植物在养分有限的情况下, 可以通过固氮作用满足自身对N的需求, 同时通过固氮还可以促进根系磷酸酶的产生, 增强植物对P的获取能力(Houlton et al., 2008; Marklein & Houlton, 2012; Nasto et al., 2014).在这种情况下, 与非固氮植物相比, N和P共同添加对固氮植物的生长并不表现出明显的促进作用(Nasto et al., 2019).此外, 有研究发现, 在光资源受限的时候, N添加对树木的共生固氮作用及其生长均没有显著影响; 而在光资源充足的时候, N添加抑制了树木共生固氮, 促进了其生长; 该研究结果揭示了N添加对固氮树木生长的影响也会受到光资源的调控(Taylor & Menge, 2018).与固氮植物相比, 在N和P添加下, 非固氮植物的养分限制得到缓解, 表现为N和P添加促进其生长.此外, 也有研究表明, P添加可以通过促进固氮植物的结瘤和共生固氮作用来促进植物的生长(Waring et al., 2019; Zheng et al., 2019).然而, 对于非固氮植物, P添加对其生长的影响可能比较复杂, 这是因为P添加虽然可以提高P的可利用性, 但会降低游离固氮作用(Zheng et al., 2019).因此, P添加对非固氮植物生长的影响可能取决于N的可利用性状态.总体而言, 在P可利用性较低时, 单独的P添加对固氮树木(或固氮树木占优势的森林)生长的促进作用较非固氮树木(或非固氮树木占优势的森林)大, 而单独的N添加对固氮与非固氮植物(或固氮与非固氮植物分别占优势的森林)生长的促进作用与单独的P添加相反. ...
Increased tree carbon storage in response to nitrogen deposition in the US 3 2010
... 在菌根类型上, 不同研究发现, 与丛枝菌根(AM)真菌共生的树种(AM树种)和与外生菌根(ECM)真菌共生的树种(ECM树种)在N、P沉降下的响应存在差异, 表明菌根类型会调控N、P添加对树木生长和森林生产力的影响(Thomas et al., 2010; Averill et al., 2018; DeForest & Snell, 2020).在N添加研究方面, 基于美国森林资源清查数据的研究表明, N沉降促进了11个物种的生长, 抑制了3个物种的生长, 而且AM树种全部表现为促进作用(Thomas et al., 2010).最新的一项研究表明, 在N水平低的时候, ECM树种的生长对N的响应要高于AM树种, 而在N水平高的时候, AM树种的生长要高于ECM树种(Averill et al., 2018).AM树种和ECM树种对N可利用性增加的差异性响应, 可能与它们对N的获取与利用策略有关系.与ECM真菌相比, AM真菌缺少分解土壤有机质的酶, 主要获取土壤中的无机N来满足新陈代谢的需要和向植物输送养分(Chalot & Brun, 1998; Talbot et al., 2008).因此, 当活性N沉降导致土壤N的可利用性提高时, AM树种能够比ECM树种获得更多的益处使生长加快. ...
... ).在N添加研究方面, 基于美国森林资源清查数据的研究表明, N沉降促进了11个物种的生长, 抑制了3个物种的生长, 而且AM树种全部表现为促进作用(Thomas et al., 2010).最新的一项研究表明, 在N水平低的时候, ECM树种的生长对N的响应要高于AM树种, 而在N水平高的时候, AM树种的生长要高于ECM树种(Averill et al., 2018).AM树种和ECM树种对N可利用性增加的差异性响应, 可能与它们对N的获取与利用策略有关系.与ECM真菌相比, AM真菌缺少分解土壤有机质的酶, 主要获取土壤中的无机N来满足新陈代谢的需要和向植物输送养分(Chalot & Brun, 1998; Talbot et al., 2008).因此, 当活性N沉降导致土壤N的可利用性提高时, AM树种能够比ECM树种获得更多的益处使生长加快. ...
... (3)深度研究植物功能性状在调节与预测树木生长对N、P添加响应中的作用.尽管已有相关研究从植物功能性状的角度研究了N、P添加如何影响树木生长, 但这些研究还依旧十分匮乏, 而且现有的研究也仅仅考虑了单一的植物功能性状(Thomas et al., 2010; Averill et al., 2018; Nasto et al., 2019).植物的功能性状具有多个维度, 而且这些性状常常是交织在一起, 比如植物可以同时具备固氮和共生菌根的特性, 固氮植物具有较高的AM菌根侵染率等(Nasto et al., 2014).在这种情况下, 考虑单一的功能性状可能会使研究结果受到其他因素的干扰.因此, 在未来研究中需要综合考虑植物各个部分的性状, 并采用较为复杂全面的统计方法定量分析植物功能性状在调控树木生长响应N、P添加中的作用. ...
Growth responses of trees and understory plants to nitrogen fertilization in a subtropical forest in China 7 2017
... 目前, 在N、P添加实验中, 树木生长和森林生产力的常用评估方法主要有两种: 一是相对生长速率, 二是绝对增长量.相对生长速率(表1)一般是用一段时间N、P添加处理之后树木胸径(DBH)、胸高断面积或生物量与处理之前的差值除以所对应的时间(Markewitz et al., 2012; Tian et al., 2017; Li et al., 2018).此外, 也有研究使用一段时间N、P添加处理之后DBH或胸高断面积与处理之前的差值除以处理之前的DBH或胸高断面积来表示树木的相对生长(Jiang et al., 2018).绝对增长量(表1)一般是用N、P添加处理一段时间内的DBH、胸高断面积、生物量(或生产力, 以碳储量计算)的绝对变化量来表示.生物量的变化是基于胸径-树高曲线、树木其他器官生物量与DBH和树高之间混合回归模型、异速生长方程等计算林分尺度总生物量的变化(Herbert & Fownes, 1995; Markewitz et al., 2012; Du & Fang, 2014; 周序力等, 2018).在森林根系生产力的估测中, 也有很多研究利用内生长环法(ingrowth core method)进行实测(Waring et al., 2019).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
... Methods for assessing the effects of nitrogen and/or phosphorus addition on plant growth and productivity Table 1
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
... 在生态系统内的某类植物个体或群体水平上(如小树与大树), N、P添加对树木生长的影响因树木径级的不同而存在差异.海南热带山地森林的一项为期5年的研究发现, P添加对小树(DBH ≤ 10 cm)和中树(10 cm < DBH ≤ 20 cm)的生长表现为显著的促进作用; 但对大树(DBH > 20 cm)的生长没有显著影响(Jiang et al., 2018).其他一些在热带森林中开展的实验也表明, N、P添加对树木生长的影响也因树木径级的不同而存在明显的差异(Alvarez-Clare et al., 2013; Fisher et al., 2013).在鸡公山温带森林的研究表明, N添加对小树和大树生长都没有显著的影响, 而P添加在小树中表现为显著的促进作用(Li et al., 2018).也有研究发现, N、P添加对树木生长的促进作用与树木的初始大小具有密切的正相关关系, 即在N、P添加下大树的生长要快于小树(Báez & Homeier, 2018).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
... ; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
A global analysis of soil acidification caused by nitrogen addition 2 2015
... 尽管有很多研究发现, N、P添加能够促进生态系统尺度上的树木生长和森林生产力, 但是N、P添加对它们的抑制作用在实验中也经常被观测到(Harpole et al., 2011; Li et al., 2018; Nasto et al., 2019).因此, 这对根据N、P添加是否促进生长从而判断生态系统或树木N、P的限制提出了挑战.以往的研究表明, 长期的N、P添加能够导致土壤pH值下降, 使得土壤酸化(Tian & Niu, 2015; Dai et al., 2018).尽管N、P添加所产生的负效应能够从过量的养分添加所产生的土壤酸化等毒害作用进行解释, 但是当没有毒害作用发生时, N、P添加依旧对树木生长产生负效应(?apek et al., 2018).针对这种情况, ?apek等(2018)提出了一个基于植物和微生物相互作用(plant-microbe interaction)的新的概念框架模型来解释并预测在生态系统尺度上N、P添加对初级生产力的影响.在该化学计量框架模型中, 植物与微生物之间的相互作用包括两种情况: 一是竞争关系; 二是协作关系, 如植物通过根系向土壤中分泌活性有机碳, 促进微生物的生长, 而微生物矿化有机质为植物提供N或P.该概念框架模型认为, 在一个生态系统中存在3种N、P的限制模式, 一是对于植物和土壤微生物都是N限制; 二是对于植物为N限制, 但是对于土壤微生物是P限制; 三是对于植物和土壤微生物都是P限制.简单来说, 如果一个系统中, 植物和微生物之间是协作关系占主导, 添加对植物生长非限制性的养分可以通过促进受该养分限制的微生物生长而间接提高植物的营养和生长; 相反, 在植物和微生物间是竞争作用占主导的系统内, 植物会受到多种养分的共同限制(?apek et al., 2018).该概念框架模型能够很好地解释生态系统生产力对N、P添加的响应, 并对以往研究中难以解释的响应提供了解决思路. ...
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
Global evidence on nitrogen saturation of terrestrial ecosystem net primary productivity 1 2016
... 除环境与生物因素外, 树木生长及生产力对养分添加的响应程度也会受到实验处理条件的影响, 如养分添加量、实验持续时间、养分种类等.在养分添加量方面, 不同的研究结果并不统一.一些研究发现, 树木生长及生产力对养分添加的响应随添加量的升高而增大(Magnani et al., 2007; Du & Fang, 2014; 刘修元等, 2015).然而, 也有研究表明单位N、P添加量所导致的地上生产力增加量随养分添加量的增加而降低(H?gberg et al., 2006; Tian et al., 2016; Schulte-Uebbing & de Vries, 2018).目前, 多数研究结果显示树木生长速率与N添加量之间是非线性关系(一般为凸形抛物线), 表现为中水平N添加的促进作用强于低水平N和高水平N添加(Braun et al., 2017; Averill et al., 2018), 这可能是因为N添加量过高时会暂时超过微生物对N的需求, 从而导致硝化作用增强和土壤酸化(H?gberg et al., 2006; Tian & Niu, 2015; Chen et al., 2016), 对植物根系产生毒害作用以及降低植物的光合作用能力.研究发现, 常绿针叶林的光合作用能力与N添加量存在凸形抛物线关系(Fleischer et al., 2013).对于P添加而言, 当前的研究结果发现, 植物地上和地下的生物量和生产力的响应基本都随着P添加量的增加而增强(Li et al., 2016), 或者达到一定添加量时树木生长不再变化(McGrath et al., 2003).此外, 养分添加对树木生长和生产力的影响会随实验处理时间而发生变化.有研究发现, N添加对森林生产力的促进作用随时间的持续而逐渐减弱(Rappe-George et al., 2013), 但也有研究表明随着施肥时间的持续树木生长对N添加的响应逐渐增强(刘修元等, 2015); 这种差异可能和森林所处的林龄阶段、树种以及施肥的时间跨度有关.养分种类也会影响森林生产力对N、P添加的响应, 研究发现, 过磷酸钙(或重过磷酸钙)对森林生产力的促进作用强于磷酸二氢钠等P肥(Li et al., 2016; Hou et al., 2020), 这可能与过磷酸钙中含有的额外的钙和硫元素对植物生长的促进作用和对土壤环境的调节作用有关(Vadeboncoeur, 2010; Hopmans & Elms, 2013; Mainwaring et al., 2014). ...
Meta-analysis of fertilization experiments indicates multiple limiting nutrients in northeastern deciduous forests 5 2010
... 在生态系统尺度上, N、P添加对树木生长和森林生产力的影响在不同的林龄中也不尽相同, 受到林龄的调控.Goswami等(2018)在温带森林的研究发现, N添加对幼龄林生长的促进作用强于中龄林和成熟林; P添加在中龄林中的促进作用最为明显, 而在幼龄林和成熟林中均没有明显的作用.然而, 也有研究发现, N、P添加对次生林(林龄较小)的生长表现为促进, 而对成熟林没有显著影响, 即次生林的生长响应明显高于成熟林(Wright et al., 2018; Wright, 2019).也有定量研究发现, N添加对树木生长的影响随着年龄的增大而显著降低(Vadeboncoeur, 2010), 表明幼龄林对N添加的响应更为强烈.总体而言, 目前的研究结果表明, N添加对幼龄林的促进作用一般高于中龄林和老龄林(Vadeboncoeur, 2010; Rappe-George et al., 2013; Schulte-Uebbing & de Vries, 2018), 因为幼龄林树木在吸收添加的养分之后, 能够快速地进行光合作用, 合成自身新陈代谢所需要的有机物, 生长相对较快.Wright等(2018)认为老龄林生长对N、P添加响应较弱的原因可能存在于两个方面: 一方面, 生活在养分可利用性较低环境中的物种已经适应了这种养分缺乏的条件, 在面对较好的养分条件时其增加生长的潜力有限(Coley et al., 1985), 群落生长的响应可能需要等到物种组成发生变化以适应较高养分可用性时才能体现出来(Chapin III et al., 1986); 另一方面, 植物在施肥条件下其组织中的N含量升高, 会增加其受到天敌(食草动物和其他害虫)危害的风险并消耗其初级生产力, 从而对树木生长的响应产生不利影响(Andersen et al., 2010). ...
... ), 表明幼龄林对N添加的响应更为强烈.总体而言, 目前的研究结果表明, N添加对幼龄林的促进作用一般高于中龄林和老龄林(Vadeboncoeur, 2010; Rappe-George et al., 2013; Schulte-Uebbing & de Vries, 2018), 因为幼龄林树木在吸收添加的养分之后, 能够快速地进行光合作用, 合成自身新陈代谢所需要的有机物, 生长相对较快.Wright等(2018)认为老龄林生长对N、P添加响应较弱的原因可能存在于两个方面: 一方面, 生活在养分可利用性较低环境中的物种已经适应了这种养分缺乏的条件, 在面对较好的养分条件时其增加生长的潜力有限(Coley et al., 1985), 群落生长的响应可能需要等到物种组成发生变化以适应较高养分可用性时才能体现出来(Chapin III et al., 1986); 另一方面, 植物在施肥条件下其组织中的N含量升高, 会增加其受到天敌(食草动物和其他害虫)危害的风险并消耗其初级生产力, 从而对树木生长的响应产生不利影响(Andersen et al., 2010). ...
... 目前, 关于N、P添加对树木生长和森林生产力的影响已经有很多相关研究, 一些新出现的机制或新发现, 拓展了我们对N、P沉降背景下森林生态系统生产力变化的认识.以前的观点认为, 在纬度梯度上热带森林生产力主要受P限制, 北方森林和温带森林生产力主要受N限制(Vitousek, 1984; Vitousek et al., 2010).然而, 也有研究指出在亚热带和温带的一些区域, 主要是小树而不是大树受到P的限制(Jiang et al., 2018; Li et al., 2018); 启示我们当评价森林生产力养分限制的时候需要考虑树木大小依赖的养分限制.从基于单一的树木径级和树木的某一生物性状(如固氮属性、共生菌根), 到基于定量的植物功能性状、植物和微生物之间的相互作用关系, 我们对树木生长响应N、P添加的理解逐渐深入.基于此, 我们对N、P添加对树木生长和森林生产力影响的研究进行了总结与归纳(图3), 该图涵盖了树木生长和森林生产力的评估方法、植物功能性状以及植物和微生物之间的关系, 为理解N、P添加影响树木生长提供了研究思路.尽管如此, 目前关于树木生长和生产力如何响应N、P添加仍然存在很多不清楚的方面, 限制了我们的深层认识, 未来需要在以下几个方面加强研究. ...
Nutrient limitations to plant growth during primary succession in Hawaii Volcanoes National Park 1 1993
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
The fate of phosphorus during pedogenesis 1 1976
... 全球整合分析结果表明, 单独的N和P添加整体上提高了森林地上部分的生物量和生产力, 以及地上地下的总生物量和生产力(Janssens et al., 2010; Yuan & Chen, 2012; Li et al., 2016; ?apek et al., 2018; Schulte-Uebbing & de Vries, 2018; Hou et al., 2020).虽然, N和P的作用整体上以促进为主, 但这种N、P的促进作用在不同气候区的森林中存在差异.N的促进作用在北方森林和温带森林中最为明显, 但是在热带森林中较弱或不明显(Yuan & Chen, 2012; Tian et al., 2017; Lu et al., 2018; Schulte-Uebbing & de Vries, 2018); 而P的促进作用在热带森林中稍强于温带森林, 这些相异性的结果可能与N、P养分限制的纬度格局有关(Vitousek, 1984; Vitousek et al., 2010; Norby et al., 2016; Du et al., 2020).生态系统的N输入主要来源于生物固氮, 而P输入主要来源于岩石风化(Vitousek et al., 2010; Pe?uelas et al., 2013).由于温度和降水量较高, 土壤发育时间较长, 生物固氮作用强且积累时间较长, 土壤碳氮比较小, N矿化速率以及N循环速率较高(Cleveland et al., 1999; Xu et al., 2013; Deng et al., 2018; Dynarski & Houlton, 2018), 热带森林被认为是N充足的生态系统类型(Vitousek & Farrington, 1997; Vitousek et al., 2010); 但是, 由于土壤发育时间较长, 风化程度较高, 土壤P淋溶流失严重, 热带森林的P较为缺乏而表现为植物生长受到P的限制(Vitousek, 1984; Elser et al., 2007; Vitousek et al., 2010).与之相反, 在北方森林中, 由于温度较低, 土壤发育时间较短, 土壤碳氮比较大, 生物固氮作用以及N矿化作用较弱(Xu et al., 2013; Deng et al., 2018; Schulte-Uebbing & de Vries, 2018), 北方森林生态系统生产力更多地受到N的限制(Magnani et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010).在温带森林或中纬度区域, 生态系统生产力应该受到N和P的共同限制(Hedin, 2004).因此, 在北半球纬度梯度上, 从南到北P的限制程度逐渐减弱, 而N的限制程度逐渐增强.与此类似, 在热带森林地区, 沿海拔梯度也存在与纬度梯度相似的N和P养分限制的格局, 这种格局的产生可能主要和海拔梯度上气候和土壤条件的异同有关系; 一般表现为低海拔的热带低地雨林主要受P的限制, 而高海拔的热带山地森林主要受N的限制(Walker & Syers, 1976; Vitousek, 1984; Vitousek & Farrington, 1997; Arag?o et al., 2009).然而, 最新的一项整合研究发现, 在热带森林中并不存在海拔梯度上的N、P限制格局(Wright, 2019).因此, 在热带森林区域, 海拔梯度上的养分限制格局是否存在以及能否用这种格局解释热带低地雨林和山地雨林树木生长和生产力响应N、P添加的差异仍存在争议, 还需要进一步的研究. ...
Plant community responses to stand-level nutrient fertilization in a secondary tropical dry forest 3 2019
... 目前, 在N、P添加实验中, 树木生长和森林生产力的常用评估方法主要有两种: 一是相对生长速率, 二是绝对增长量.相对生长速率(表1)一般是用一段时间N、P添加处理之后树木胸径(DBH)、胸高断面积或生物量与处理之前的差值除以所对应的时间(Markewitz et al., 2012; Tian et al., 2017; Li et al., 2018).此外, 也有研究使用一段时间N、P添加处理之后DBH或胸高断面积与处理之前的差值除以处理之前的DBH或胸高断面积来表示树木的相对生长(Jiang et al., 2018).绝对增长量(表1)一般是用N、P添加处理一段时间内的DBH、胸高断面积、生物量(或生产力, 以碳储量计算)的绝对变化量来表示.生物量的变化是基于胸径-树高曲线、树木其他器官生物量与DBH和树高之间混合回归模型、异速生长方程等计算林分尺度总生物量的变化(Herbert & Fownes, 1995; Markewitz et al., 2012; Du & Fang, 2014; 周序力等, 2018).在森林根系生产力的估测中, 也有很多研究利用内生长环法(ingrowth core method)进行实测(Waring et al., 2019).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
... ; Waring et al., 2019). ...
... 植物的固氮属性会调控N、P添加对树木生长的影响, 原因可能是固氮植物与非固氮植物具有不同的养分获取策略.Nasto等(2019)研究发现, 在土壤养分可利用性较低的热带森林土壤中, N和P共同添加(该研究中土壤P含量较低, 可能P添加起主要作用)促进了非固氮植物的生长; 但对固氮植物的生长影响不大.固氮植物在养分有限的情况下, 可以通过固氮作用满足自身对N的需求, 同时通过固氮还可以促进根系磷酸酶的产生, 增强植物对P的获取能力(Houlton et al., 2008; Marklein & Houlton, 2012; Nasto et al., 2014).在这种情况下, 与非固氮植物相比, N和P共同添加对固氮植物的生长并不表现出明显的促进作用(Nasto et al., 2019).此外, 有研究发现, 在光资源受限的时候, N添加对树木的共生固氮作用及其生长均没有显著影响; 而在光资源充足的时候, N添加抑制了树木共生固氮, 促进了其生长; 该研究结果揭示了N添加对固氮树木生长的影响也会受到光资源的调控(Taylor & Menge, 2018).与固氮植物相比, 在N和P添加下, 非固氮植物的养分限制得到缓解, 表现为N和P添加促进其生长.此外, 也有研究表明, P添加可以通过促进固氮植物的结瘤和共生固氮作用来促进植物的生长(Waring et al., 2019; Zheng et al., 2019).然而, 对于非固氮植物, P添加对其生长的影响可能比较复杂, 这是因为P添加虽然可以提高P的可利用性, 但会降低游离固氮作用(Zheng et al., 2019).因此, P添加对非固氮植物生长的影响可能取决于N的可利用性状态.总体而言, 在P可利用性较低时, 单独的P添加对固氮树木(或固氮树木占优势的森林)生长的促进作用较非固氮树木(或非固氮树木占优势的森林)大, 而单独的N添加对固氮与非固氮植物(或固氮与非固氮植物分别占优势的森林)生长的促进作用与单独的P添加相反. ...
Future productivity and carbon storage limited by terrestrial nutrient availability 1 2015
... 养分限制模式对于生态系统生产力及其对全球变化的响应至关重要(Reich et al., 2006; Crowley et al., 2012).在全球尺度上, 氮(N)和磷(P)对植物生产力的限制被认为普遍存在于陆地、淡水和海洋生态系统中(Elser et al., 2007).在大气CO2浓度持续上升背景下, 植物生长受到促进, 从而会从土壤中摄取更多的养分, 进而导致其生长逐渐受到养分可利用性的限制(Luo et al., 2004).模型预测结果表明, 在考虑N、P可利用性限制的情况下, 净初级生产力的增加将低于N和P不受限制状态下的增加量, 而陆地生态系统也将由碳吸收转变为碳排放(Wieder et al., 2015).由此可见, 养分可利用性在调节陆地净初级生产力和陆地碳库中发挥着至关重要的作用. ...
Phylogeny is a powerful tool for predicting plant biomass responses to nitrogen enrichment 1 2017
... 目前, 在N、P添加实验中, 树木生长和森林生产力的常用评估方法主要有两种: 一是相对生长速率, 二是绝对增长量.相对生长速率(表1)一般是用一段时间N、P添加处理之后树木胸径(DBH)、胸高断面积或生物量与处理之前的差值除以所对应的时间(Markewitz et al., 2012; Tian et al., 2017; Li et al., 2018).此外, 也有研究使用一段时间N、P添加处理之后DBH或胸高断面积与处理之前的差值除以处理之前的DBH或胸高断面积来表示树木的相对生长(Jiang et al., 2018).绝对增长量(表1)一般是用N、P添加处理一段时间内的DBH、胸高断面积、生物量(或生产力, 以碳储量计算)的绝对变化量来表示.生物量的变化是基于胸径-树高曲线、树木其他器官生物量与DBH和树高之间混合回归模型、异速生长方程等计算林分尺度总生物量的变化(Herbert & Fownes, 1995; Markewitz et al., 2012; Du & Fang, 2014; 周序力等, 2018).在森林根系生产力的估测中, 也有很多研究利用内生长环法(ingrowth core method)进行实测(Waring et al., 2019).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
贵州月亮山不同演替阶段亮叶水青冈林碳储量及其分配格局 1 2018
... 目前, 在N、P添加实验中, 树木生长和森林生产力的常用评估方法主要有两种: 一是相对生长速率, 二是绝对增长量.相对生长速率(表1)一般是用一段时间N、P添加处理之后树木胸径(DBH)、胸高断面积或生物量与处理之前的差值除以所对应的时间(Markewitz et al., 2012; Tian et al., 2017; Li et al., 2018).此外, 也有研究使用一段时间N、P添加处理之后DBH或胸高断面积与处理之前的差值除以处理之前的DBH或胸高断面积来表示树木的相对生长(Jiang et al., 2018).绝对增长量(表1)一般是用N、P添加处理一段时间内的DBH、胸高断面积、生物量(或生产力, 以碳储量计算)的绝对变化量来表示.生物量的变化是基于胸径-树高曲线、树木其他器官生物量与DBH和树高之间混合回归模型、异速生长方程等计算林分尺度总生物量的变化(Herbert & Fownes, 1995; Markewitz et al., 2012; Du & Fang, 2014; 周序力等, 2018).在森林根系生产力的估测中, 也有很多研究利用内生长环法(ingrowth core method)进行实测(Waring et al., 2019).尽管相对生长速率和绝对增长量这两种方法都可以用来评估树木生长和森林生产力的变化, 但是二者之间的结果会存在差异(Jiang et al., 2018; Waring et al., 2019). ...
Phosphorus limitation on photosynthesis of two dominant understory species in a lowland tropical forest 1 2014
... (1)加强树木生长响应N、P添加的生理学机制研究.一项在N富集的热带森林中的N添加实验表明, 10年左右的N添加使土壤发生了酸化, 降低了土壤中的盐基离子(钙和镁)的可利用性, 但是树木能够通过增强叶片蒸腾作用提高对盐基离子的吸收来维持自身的养分平衡(Lu et al., 2018).N和P的可利用性提高后, 植物的生理活动会发生相应的响应, 这些响应可能会对树木的生长产生影响, 并最终反应到DBH和生物量变化上.目前, 仅有少数研究(Zhu et al., 2014; Li et al., 2018; Lu et al., 2018)对N、P添加后的树木生理水平上的响应(光合作用、蒸腾作用、叶片养分重吸收效率等)进行了测定.因此, 后续研究可以从植物的生理角度开展, 如光合作用、水分传输、根系寿命等, 以期能够提供深入的机理解释. ...
模拟氮沉降对北京东灵山辽东栎林树木生长的影响 2 2019
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
... 在生态系统内的某类植物个体或群体水平上(如小树与大树), N、P添加对树木生长的影响因树木径级的不同而存在差异.海南热带山地森林的一项为期5年的研究发现, P添加对小树(DBH ≤ 10 cm)和中树(10 cm < DBH ≤ 20 cm)的生长表现为显著的促进作用; 但对大树(DBH > 20 cm)的生长没有显著影响(Jiang et al., 2018).其他一些在热带森林中开展的实验也表明, N、P添加对树木生长的影响也因树木径级的不同而存在明显的差异(Alvarez-Clare et al., 2013; Fisher et al., 2013).在鸡公山温带森林的研究表明, N添加对小树和大树生长都没有显著的影响, 而P添加在小树中表现为显著的促进作用(Li et al., 2018).也有研究发现, N、P添加对树木生长的促进作用与树木的初始大小具有密切的正相关关系, 即在N、P添加下大树的生长要快于小树(Báez & Homeier, 2018).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
模拟氮沉降对北京东灵山辽东栎林树木生长的影响 2 2019
... 国内外****关于N、P添加对树木生长和森林生产力的影响进行了大量的研究, 但是不同研究得到的结果存在较大的差异, 存在3种影响: 促进作用(Vitousek et al., 1993; Rappe-George et al., 2013; Goswami et al., 2018; Jiang et al., 2018)、抑制作用(Alvarez-Clare et al., 2013; Nasto et al., 2019; DeForest & Snell, 2020)或没有显著影响(Tian et al., 2017; Lu et al., 2018; 邹安龙等, 2019).根据以往的研究, 这种不一致的结果可能受到多种因素的调控与影响, 包括气候因素、树种特性或植物功能属性(固氮特性、共生菌根)、树木大小、林龄、光资源水平、植物和微生物之间的关系、土壤属性, 以及实验处理条件(养分添加量、养分种类、实验处理时间等). ...
... 在生态系统内的某类植物个体或群体水平上(如小树与大树), N、P添加对树木生长的影响因树木径级的不同而存在差异.海南热带山地森林的一项为期5年的研究发现, P添加对小树(DBH ≤ 10 cm)和中树(10 cm < DBH ≤ 20 cm)的生长表现为显著的促进作用; 但对大树(DBH > 20 cm)的生长没有显著影响(Jiang et al., 2018).其他一些在热带森林中开展的实验也表明, N、P添加对树木生长的影响也因树木径级的不同而存在明显的差异(Alvarez-Clare et al., 2013; Fisher et al., 2013).在鸡公山温带森林的研究表明, N添加对小树和大树生长都没有显著的影响, 而P添加在小树中表现为显著的促进作用(Li et al., 2018).也有研究发现, N、P添加对树木生长的促进作用与树木的初始大小具有密切的正相关关系, 即在N、P添加下大树的生长要快于小树(Báez & Homeier, 2018).总体而言, 在同一林分中, 小树(幼树)对N、P添加的响应具有一定的不确定性.N、P添加对小树生长的影响可表现为促进作用(Alvarez-Clare et al., 2013; 刘修元等, 2015; Li et al., 2018), 也可能产生抑制作用(Tian et al., 2017; Li et al., 2018; 邹安龙等, 2019), 这种迥异的结果可能取决于小树与大树之间的资源竞争关系.小树生长的负响应可能主要与植物生态生理过程的光可利用性有关系.小树通常为林下植物或被压木, 在对光的竞争中处于劣势地位(刘修元等, 2015; Tian et al., 2017); 由于光可利用性的限制, 林下植物不能吸收所添加的养分来提升自身的光合速率(Alvarez-Clare et al., 2013). ...
 ,, 朱彪
,, 朱彪 ,*北京大学生态研究中心, 北京大学城市与环境学院, 北京大学地表过程分析与模拟教育部重点实验室, 北京 100871
,*北京大学生态研究中心, 北京大学城市与环境学院, 北京大学地表过程分析与模拟教育部重点实验室, 北京 100871
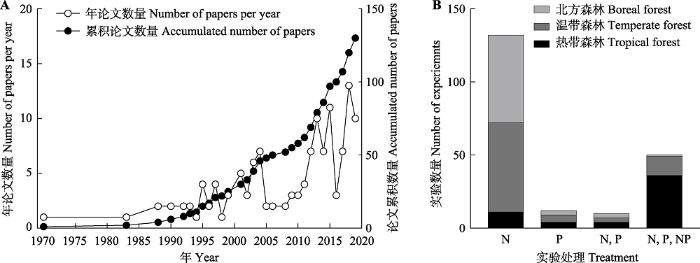 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT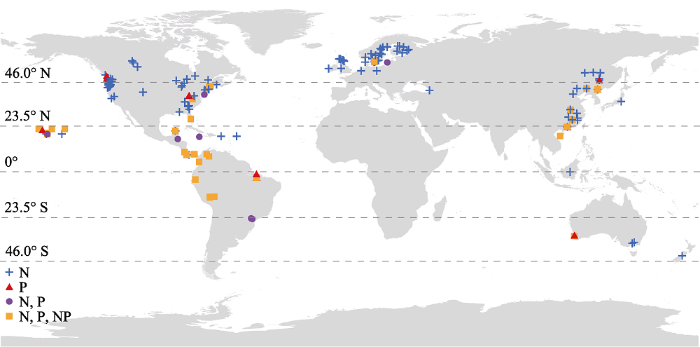 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT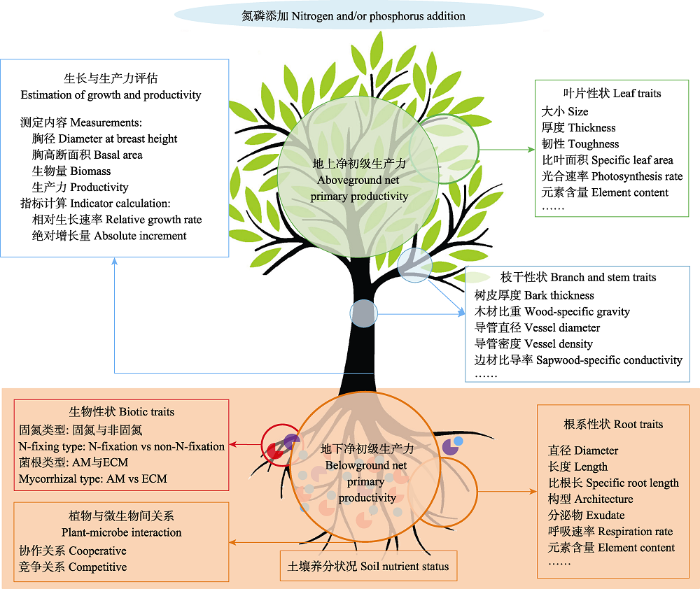 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}