Growth and decomposition characteristics of Sphagnum in a subalpine wetland, southwestern Hubei, China
Xue-Fei LIU,1,2, Lin WU,1,2,*, Han WANG1,2, Liu HONG1,2, Li-Jun XIONG31Hubei Key Laboratory of Biological Resources Protection and Utilization, Enshi, Hubei 445000, China 2College of Forestry and Horticulture, Hubei Minzu University, Enshi, Hubei 445000, China 3Meteorological Bureau of Enshi Tujia and Miao Autonomous Prefecture, Enshi, Hubei 445000, China
Abstract Aims We investigated the growth and decomposition of Sphagnum, a principal plant involved in peat accumulation in wetlands, in subtropical wetlands. Methods We selected a Sphagnum-dominated wetland in southwestern Hubei Province as the study area. In situ growth and litter decomposition experiments in different microhabitats were carried out, and the chemical composition of litter was measured in laboratory to examine the processes of its growth and decomposition. Important findings The average height of Sphagnum in hummock and hollow were 2.9 and 2.7 cm, and the net production was 270.94 and 370.88 g·m -2, respectively, after 12 months. Growth time and microhabitats significantly affected on the height growth and the net production, and there was an interaction between time and microhabitats. However, the growth trend in two microhabitats was significantly different. The average growth rate of Sphagnum under the two microhabitats (July to October 2017) was 0.33 mm·d-1, which was higher than that in the cold temperate zone. In addition, the decomposition time had an significant effect on the decomposition amount of Sphagnum, and the residual rate decreased first and then increased with time. By the end 12-month period, the final residual rates in hollow, hummock and pool were 100.67%, 90.54% and 85.63%, respectively. The content of carbon (C), carbon nitrogen (N) ratio (C:N) and polyphenols in the litters decreased, while the nitrogen content increased. During the first 3 months of decomposition, a significant difference of decomposed litter was found between the microhabitats, but no significant difference later on. Keywords:subtropical;Sphagnum;microhabitats;net production;decomposition
PDF (1544KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 刘雪飞, 吴林, 王涵, 洪柳, 熊莉军. 鄂西南亚高山湿地泥炭藓的生长与分解. 植物生态学报, 2020, 44(3): 228-235. DOI: 10.17521/cjpe.2019.0316 LIU Xue-Fei, WU Lin, WANG Han, HONG Liu, XIONG Li-Jun. Growth and decomposition characteristics of Sphagnum in a subalpine wetland, southwestern Hubei, China. Chinese Journal of Plant Ecology, 2020, 44(3): 228-235. DOI: 10.17521/cjpe.2019.0316
新窗口打开|下载原图ZIP|生成PPT 图1鄂西南亚高山泥炭藓湿地中的3种微生境(从左至右依次为: 丘上、丘间、水坑)。 Fig. 1Three types of microhabitats in Sphagnum-dominated subalpine wetland in southwestern Hubei (From left to right: hummock, hollow, and pool).
新窗口打开|下载原图ZIP|生成PPT 图2鄂西南亚高山泥炭藓湿地丘间与丘上两种微生境下泥炭藓高度增长量(A)及净生产量(B)变化(平均值±标准误差, n = 3)。*表示在同一时间不同微生境下有显著性差异(p < 0.05)。
Fig. 2Increment of height growth (A) and net production (B) of Sphagnum in hollow and hummock of Sphagnum-dominated subalpine wetlands in southwestern Hubei (mean ± SE, n = 3). * refers a significant difference between the two microhabitats at the same time (p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图3鄂西南亚高山泥炭藓湿地3种微生境下泥炭藓凋落物残留率变化规律(平均值±标准误差, n = 3)。*表示在同一时间不同微生境下有显著性差异(p < 0.05), 不同小写字母表示在同一微生境不同时间有显著性差异(p < 0.05)。
Fig. 3Changes in litters residual rate of Sphagnum under three microhabitats of Sphagnum-dominated subalpine wetlands in southwestern Hubei (mean ± SE, n = 3). * refers a significant difference among the three microhabitats at the same time (p < 0.05); different lowercase letters indicate significant differences at different time periods in the same microhabitat (p < 0.05).
Fig. 4Contents of C (A), N (B), C:N (C) and polyphenols (D) in the litters of Sphagnum of the three microhabitats of Sphagnum- dominated subalpine wetlands in southwestern Hubei (mean ± SE, n = 3). * refers a significant difference among the three microhabitats at the same time (p < 0.05); different lowercase letters indicate significant differences at different time periods in the same microhabitat(p < 0.05)
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. S. ang, 小叶泥炭藓; S. cap, 尖叶泥炭鲜; S. fal, 假泥炭藓; S. fus, 锈色泥炭藓; S. mag, 中位泥炭藓; S. pal, 泥炭藓; S. rec, 喙叶泥炭藓。泥炭藓属植物学名及拉丁名参照王利松等(2018)。泥炭藓属植物的生长数据均为采用Clymo (1970)方法进行野外原位测定所得, 测定时间均为生长季节, 且生长速率及净生产力统一转换为mm·d-1和g·m-2·d-1。
AsadaT, WarnerBG, BannerA (2003). Growth of mosses in relation to climate factors in a hypermaritime coastal peatland in British Columbia, Canada The Bryologist, 106, 516-527. DOI:10.1639/0007-2745(2003)106[516:GOMIRT]2.0.CO;2URL [本文引用: 2]
BellMC, RitsonJP, VerhoefA, BrazierRE, TempletonMR, GrahamNJD, FreemanC, ClarkJM (2018). Sensitivity of peatland litter decomposition to changes in temperature and rainfall Geoderma, 331, 29-37. DOI:10.1016/j.geoderma.2018.06.002URL [本文引用: 1]
BengtssonF, GranathG, RydinH (2016). Photosynthesis, growth, and decay traits in Sphagnum—A multispecies comparison Ecology and Evolution, 6, 3325-3341. DOI:10.1002/ece3.2016.6.issue-10URL [本文引用: 4]
BengtssonF, RydinH, HájekT (2018). Biochemical determinants of litter quality in 15 species of Sphagnum Plant and Soil, 425, 161-176. DOI:10.1007/s11104-018-3579-8URL [本文引用: 1]
BragazzaL, SiffiC, IacuminP, GerdolR (2007). Mass loss and nutrient release during litter decay in peatland: the role of microbial adaptability to litter chemistry Soil Biology & Biochemistry, 39, 257-267. DOI:10.1016/j.soilbio.2006.07.014URL [本文引用: 2]
BreeuwerA, HeijmansM, RobroekBJM, LimpensJ, BerendseF (2008). The effect of increased temperature and nitrogen deposition on decomposition in bogs Oikos, 117, 1258-1268. DOI:10.1111/oik.2008.117.issue-8URL [本文引用: 5]
ClymoRS (1970). The growth of Sphagnum: methods of measurement Journal of Ecology, 58, 13-49. DOI:10.2307/2258168URL [本文引用: 3]
ClymoRS, HaywardPM (1982). The ecology of Sphagnum// Smith AJE Bryophyte Ecology. Chapman and Hall, London. DOI:10.1890/07-1822.1URLPMID:19449698 [本文引用: 2] Landscape heterogeneity causes spatial variation in disturbance regimes and resilience. We asked whether the resilience of bryophyte (liverwort and moss) assemblages to clear-cutting differs between streamside and upland boreal forests in northern Sweden. We hypothesized that bryophyte survival and recolonization rates are higher in streamside areas, thus raising resilience. Conversely, disturbance-intolerant but also invading species should be more frequent here, potentially reducing resilience. In each of 18 sites, we compared two 0.1-ha plots (one streamside and one upland) located in old forest that had never been clear-cut with two matching plots in young stands established after clear-cutting of old forests 30-50 years earlier. We used the magnitude of the difference in assemblages between old and young stands as a measure of change and, therefore, resilience (large difference implying low resilience). Species assemblages were more resilient in streamside than in upland forests. Species composition changed significantly in upland but not in streamside forests. Reductions in species richness were more pronounced in upland forests for total richness and for eight subgroups of species. Two results indicated lower survival/recolonization in upland forests: (1) species had a stronger association with old stands in upland areas, and (2) among species present in both the old streamside and old upland plot in a site, fewer appeared in the young upland than in the corresponding streamside plot. Simultaneously, a higher proportion of species invaded streamside areas; 40 of the 262 species encountered in streamside forests increased their occupancy by two or more sites compared to only two of 134 species in uplands. We suggest that in boreal forests spatial variation in resilience of assemblages of forest organisms intolerant of canopy removal is related to factors governed mainly by topography. More generally, we argue that landscape-scale variation in resilience of assemblages is influenced by spatial variation in (1) stress and resource availability, (2) number of disturbance-intolerant species, and (3) magnitude of environmental changes brought about by a disturbance with a specific intensity. We also suggest that rapid recovery in the short term does not necessarily imply higher long-term ability to return to the pre-disturbance state.
DorrepaalE, AertsR, CornelissenJHC, CallaghanTV, van LogtestijnRSP (2003). Summer warming and increased winter snow cover affect Sphagnum fuscum growth, structure and production in a sub-arctic bog Global Change Biology, 10, 93-104. DOI:10.1111/gcb.2004.10.issue-1URL [本文引用: 1]
Editorial Board of Wetland Vegetation in China (1999). Wetland Vegetation in China. Science Press, Beijing. [本文引用: 1]
FurnessSB, GrimeJP (1982). Growth rate and temperature responses in bryophytes: I. An investigation of Brachythecium rutabulum Journal of Ecology, 70, 513-523. DOI:10.2307/2259919URL [本文引用: 1]
GenetH, OberbauerSF, ColbySJ, StaudhammerCL, StarrG (2013). Growth responses of Sphagnum hollows to a growing season lengthening manipulation in Alaskan Arctic tundra Polar Biology, 36, 41-50. DOI:10.1007/s00300-012-1236-xURL [本文引用: 1]
GerdolR (1995). The growth dynamics of Sphagnum based on field measurements in a temperate bog and on laboratory cultures Journal of Ecology, 83, 431-437. DOI:10.2307/2261596URL [本文引用: 2]
GerdolR, PetragliaA, BragazzaL, IacuminP, BrancaleoniL (2007). Nitrogen deposition interacts with climate in affecting production and decomposition rates in Sphagnum mosses Global Change Biology, 13, 1810-1821. DOI:10.1111/gcb.2007.13.issue-8URL [本文引用: 1]
GorhamE (1991). Northern peatlands: role in the carbon cycle and probable responses to climatic warming Ecological Applications, 1, 182-195. DOI:10.2307/1941811URL [本文引用: 1]
HoYS, MckayG (2000). The kinetics of sorption of divalent metal ions onto Sphagnum moss peat Water Research, 34, 735-742. DOI:10.1016/S0043-1354(99)00232-8URL [本文引用: 1]
JasseyVEJ, ChiapusioG, GilbertD, ButtlerA, ToussaintML, BinetP (2011). Experimental climate effect on seasonal variability of polyphenol/phenoloxidase interplay along a narrow fen-bog ecological gradient in Sphagnum fallax Global Change Biology, 17, 2945-2957. DOI:10.1111/gcb.v17.9URL [本文引用: 1]
JohnsonLC, DammanAWH (1993). Decay and its regulation in Sphagnum peatlands Advances in Bryology, 5, 249-296. [本文引用: 1]
KosykhNP, KoronatovaNG, GranathG (2017). Effect of temperature and precipitation on linear increment of Sphagnum fuscum and S. magellanicum in Western Siberia Russian Journal of Ecology,2017,48, 203-211. DOI:10.1134/S1067413617030080URL [本文引用: 2]
LangSI, CornelissenJHC, KlahnT, van LogtestijnRSP, BroekmanR, SchweikertW, AertsR (2009). An experimental comparison of chemical traits and litter decomposition rates in a diverse range of subarctic bryophyte, lichen and vascular plant species Journal of Ecology, 97, 886-900. DOI:10.1111/jec.2009.97.issue-5URL [本文引用: 1]
LeroyF, GogoS, GuimbaudC, Bernard-JanninL, HuZ, Laggoun-?DéfargeF (2017). Vegetation composition controls temperature sensitivity of CO2 and CH4 emissions and DOC concentration in peatlands Soil Biology & Biochemistry, 107, 164-167. DOI:10.1016/j.soilbio.2017.01.005URL [本文引用: 1]
LiH, ParentLE, KaramA, TremblayC (2004). Potential of Sphagnum peat for improving soil organic matter, water holding capacity, bulk density and potato yield in a sandy soil Plant and Soil, 265, 355-365. DOI:10.1007/s11104-005-0753-6URL [本文引用: 1]
LiW, BuZJ, ZhangBJ, LongC, TangRJ, CuiQW (2013). Decomposition of Sphagnum litter in 4 peatlands of the Changbai Mountains along an altitudinal gradient Journal of Mountain Science, 31, 442-447. [本文引用: 7]
LiuYY, MaJZ, BuZJ, WangSZ, ZhangXB, ZhangTY, LiuSS, FuB, KangY (2018). Effect of geographical sources and biochemical traits on plant litter decomposition in a peatland Chinese Journal of Plant Ecology, 42, 713-722. DOI:10.17521/cjpe.2018.0029URL [本文引用: 5]
LoiselJ, Gallego-SalaAV, YuZ (2012). Global-scale pattern of peatland Sphagnum growth driven by photosynthetically active radiation and growing season length Biogeosciences, 9, 2737-2746. DOI:10.5194/bg-9-2737-2012URL [本文引用: 3]
MaJZ (2018). The Simulation Study on Effects of Climate Warming on Plant Litter Decomposition in Peatlands Basing on Three Experimental Modes Master degree dissertation, Northeast Normal University, Changchun. [本文引用: 2]
MironovVL, KondratevAY (2017). Peat moss Sphagnum riparium follows a circatrigintan growth rhythm in situ: a case report Chronobiology International, 34, 981-984. DOI:10.1080/07420528.2017.1329208URL [本文引用: 3]
MooreTR (1989). Growth and net production of Sphagnum at five fen sites, subarctic eastern Canada Canadian Journal of Botany, 67, 1203-1207. DOI:10.1139/b89-156URL [本文引用: 3]
NewmanTR, WrightN, WrightB, Sj?gerstenS (2018). Interacting effects of elevated atmospheric CO2 and hydrology on the growth and carbon sequestration of Sphagnum moss Wetlands Ecology and Management, 26, 763-774. DOI:10.1007/s11273-018-9607-xURL [本文引用: 1]
OrwinKH, OstleNJ (2012). Moss species effects on peatland carbon cycling after fire Functional Ecology, 26, 829-836. DOI:10.1111/fec.2012.26.issue-4URL [本文引用: 1]
PhilbenM, HolmquistJ, MacDonaldG, DuanDD, Kaiser,K, BennerR (2015). Temperature, oxygen, and vegetation controls on decomposition in a James Bay peatland Global Biogeochemical Cycles, 29, 729-743. DOI:10.1002/2014GB004989URL [本文引用: 1]
RochefortL, VittDH, BayleySE (1990). Growth, production, and decomposition dynamics of Sphagnum under natural and experimentally acidified conditions Ecology, 71, 1986-2000. DOI:10.2307/1937607URL [本文引用: 2]
SingletonVL, OrthoferR, Lamuela-RaventósRM (1999). Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent Methods in Enzymology, 299, 152-178. [本文引用: 1]
TahvanainenT, HaraguchiA (2013). Effect of pH on phenol oxidase activity on decaying Sphagnum mosses European Journal of Soil Biology, 54, 41-47. DOI:10.1016/j.ejsobi.2012.10.005URL [本文引用: 1]
ThormannMN, BayleySE (1997). Aboveground net primary production along a bog-fen-marsh gradient in southern boreal Alberta, Canada écoscience, 4, 374-384. DOI:10.1080/11956860.1997.11682416URL [本文引用: 1]
WaddingtonJM, RochefortL, CampeauS (2003). Sphagnum production and decomposition in a restored cutover peatland Wetlands Ecology & Management, 11, 85-95. [本文引用: 1]
WangLS, JiaY, ZhangXC, QinHN (2018). Species Catalogue of China. Volume 1 Plants A Synoptic Checklist (I). Science Press, Beijing. [本文引用: 2]
WiederRK, LangGE (1983). Net primary production of the dominant bryophytes in a Sphagnum-dominated wetland in West Virginia The Bryologist, 86, 280-286. DOI:10.2307/3242723URL [本文引用: 2]
WiederRK, VittDH, Burke-ScollM, ScottKD, HouseM, VileMA (2010). Nitrogen and sulphur deposition and the growth of Sphagnum fuscum in bogs of the Athabasca Oil Sands Region, Alberta Journal of Limnology, 69, 161-170. DOI:10.4081/jlimnol.2010.s1.161URL [本文引用: 3]
YazakiT, YabeK (2012). Effects of snow-load and shading by vascular plants on the vertical growth of hummocks formed by Sphagnum papillosum in a mire of northern Japan Plant Ecology, 213, 1055-1067. DOI:10.1007/s11258-012-0065-xURL [本文引用: 1]
YuZC, LoiselJ, BrosseauDP, BeilmanDW, HuntSJ (2010). Global peatland dynamics since the last glacial maximum Geophysical Research Letters, 37, L13402. DOI: 10.1029/2010gl043584. [本文引用: 1]
ZhangXH, SunXX, MaoR (2017). Effects of litter evenness, nitrogen enrichment and temperature on short-term litter decomposition in freshwater marshes of Northeast China Wetlands, 37, 145-152. DOI:10.1007/s13157-016-0855-3URL [本文引用: 1]
Growth of mosses in relation to climate factors in a hypermaritime coastal peatland in British Columbia, Canada 2 2003
Sensitivity of peatland litter decomposition to changes in temperature and rainfall 1 2018
... 近年来, 国内外****对泥炭藓属植物的生长及凋落物的分解已开展了大量的研究(Moore, 1989; Rochefort et al., 1990; Gunnarsson, 2005; Breeuwer et al., 2008; Loisel et al., 2012; 李伟等, 2013; Philben et al., 2015; Bengtsson et al., 2016; Mironov & Kondratev, 2017; Bell et al., 2018; 刘媛媛等, 2018; Newman et al., 2018).然而这些研究主要集中在高纬度的寒温带地区, 在低纬度的亚热带地区却鲜有报道(马进泽, 2018), 有关亚热带地区泥炭藓的生长及凋落物分解的原位研究更是欠缺.事实上, 泥炭藓属植物的生长及凋落物的分解主要受年平均气温、降水、植被类型、氮沉降等因素的影响(Breeuwer et al., 2008; 李伟等, 2013; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
Photosynthesis, growth, and decay traits in Sphagnum—A multispecies comparison 4 2016
... 近年来, 国内外****对泥炭藓属植物的生长及凋落物的分解已开展了大量的研究(Moore, 1989; Rochefort et al., 1990; Gunnarsson, 2005; Breeuwer et al., 2008; Loisel et al., 2012; 李伟等, 2013; Philben et al., 2015; Bengtsson et al., 2016; Mironov & Kondratev, 2017; Bell et al., 2018; 刘媛媛等, 2018; Newman et al., 2018).然而这些研究主要集中在高纬度的寒温带地区, 在低纬度的亚热带地区却鲜有报道(马进泽, 2018), 有关亚热带地区泥炭藓的生长及凋落物分解的原位研究更是欠缺.事实上, 泥炭藓属植物的生长及凋落物的分解主要受年平均气温、降水、植被类型、氮沉降等因素的影响(Breeuwer et al., 2008; 李伟等, 2013; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
... ; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
... Growth rate and net primary productivity of Sphagnum spp. in areas of different latitude Table 1
纬度 Latitude (°)
经度 Longitude (°)
年平均气温 Annual mean temperature (℃)
年降水量 Precipitation (mm)
物种 Species
生长速率 Growth rate (mm·d-1)
净生产力 Net primary productivity (g·m-2·d-1)
参考文献 Reference
68.35
18.82
0.5
320
S. fus
0.04
1.51
Dorrepaal et al., 2003
60.52
17.92
3.1
544
S. fus
0.10
0.64
Bengtsson et al., 2016
59.90
15.83
1.4
515
S. fus, S. mag
0.13
1.12
Bengtsson et al., 2016
57.15
-111.98
2.1
387
S. fus
0.09
1.68
Wieder et al., 2010
56.63
-110.20
1.1
420
S. fus
0.09
1.72
Wieder et al., 2010
55.68
-111.83
2.1
421
S. fus
0.10
1.96
Wieder et al., 2010
54.68
-113.47
1.7
500
S. fus
0.03
1.15
Thormann & Bayley, 1997
49.67
-93.72
2.6
714
S. mag, S. fus, S. ang
0.12
0.86
Rochefort et al., 1990
46.32
11.67
5.0
1 100
S. cap, S. mag, S. fal
0.26
1.70
Gerdol, 1995
39.12
-79.58
7.9
1 330
S. mag, S. rec
0.42
3.44
Wieder & Lang, 1983
30.17
109.73
7.2
1 590
S. pal
0.33
3.84
本研究 This study
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
... Bengtsson et al., 2016
57.15
-111.98
2.1
387
S. fus
0.09
1.68
Wieder et al., 2010
56.63
-110.20
1.1
420
S. fus
0.09
1.72
Wieder et al., 2010
55.68
-111.83
2.1
421
S. fus
0.10
1.96
Wieder et al., 2010
54.68
-113.47
1.7
500
S. fus
0.03
1.15
Thormann & Bayley, 1997
49.67
-93.72
2.6
714
S. mag, S. fus, S. ang
0.12
0.86
Rochefort et al., 1990
46.32
11.67
5.0
1 100
S. cap, S. mag, S. fal
0.26
1.70
Gerdol, 1995
39.12
-79.58
7.9
1 330
S. mag, S. rec
0.42
3.44
Wieder & Lang, 1983
30.17
109.73
7.2
1 590
S. pal
0.33
3.84
本研究 This study
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
Biochemical determinants of litter quality in 15 species of Sphagnum 1 2018
... 泥炭藓的分解相比生长更为复杂, 其分解过程不仅受地表植被类型、生境、气候条件及氮沉降等环境因素的影响(Gerdol et al., 2007; Breeuwer et al., 2008; 李伟等, 2013), 同时也受不同物种间本身属性的差异(李伟等, 2013; Bengtsson et al., 2018; 刘媛媛等, 2018)和土壤养分含量的差异(Bragazza et al., 2007)所影响.本研究3种微生境下泥炭藓凋落物野外分解12个月后最终残留率平均为92.28%, 略高于刘媛媛等(2018)在长白山哈泥泥炭地的研究结果(90%).同样是泥炭藓湿地, 李伟等(2013)在长白山的研究发现泥炭藓凋落物经历14个月的分解后最终残留率为80.8%, 显著低于本研究结果.这可能与不同凋落物本身的化学属性及研究区的环境有关.研究表明, 凋落物C:N越高, 分解越快(Limpens & Berendse, 2003).与李伟等(2013)及刘媛媛等(2018)在长白山的研究相比, 本研究中泥炭藓初始C:N较低, 而分解却更慢, 这可能与本研究区凋落物本身多酚含量更高、土壤pH较低及土壤碳氮含量更低有一定关系.研究表明, 较高的多酚含量(Jassey et al., 2011)、较低的pH (Tahvanainen & Haraguchi, 2013)及较低的土壤养分含量(Bragazza et al., 2007)会抑制凋落物的分解.另外, 温度也是影响凋落物分解的一个重要因素.以往的研究表明, 较高的温度会促进凋落物的分解(Zhang et al., 2017), 也有研究认为较高的温度对凋落物的分解无显著影响(Breeuwer et al., 2008).本研究区温度显著高于寒温带地区, 然而分解率却低于寒温带地区, 可能与研究区其他环境因子对凋落物分解的抑制作用较强有关, 比如研究区土壤酸性高于寒温带地区, 凋落物多酚含量也更高. ...
Mass loss and nutrient release during litter decay in peatland: the role of microbial adaptability to litter chemistry 2 2007
... 泥炭藓的分解相比生长更为复杂, 其分解过程不仅受地表植被类型、生境、气候条件及氮沉降等环境因素的影响(Gerdol et al., 2007; Breeuwer et al., 2008; 李伟等, 2013), 同时也受不同物种间本身属性的差异(李伟等, 2013; Bengtsson et al., 2018; 刘媛媛等, 2018)和土壤养分含量的差异(Bragazza et al., 2007)所影响.本研究3种微生境下泥炭藓凋落物野外分解12个月后最终残留率平均为92.28%, 略高于刘媛媛等(2018)在长白山哈泥泥炭地的研究结果(90%).同样是泥炭藓湿地, 李伟等(2013)在长白山的研究发现泥炭藓凋落物经历14个月的分解后最终残留率为80.8%, 显著低于本研究结果.这可能与不同凋落物本身的化学属性及研究区的环境有关.研究表明, 凋落物C:N越高, 分解越快(Limpens & Berendse, 2003).与李伟等(2013)及刘媛媛等(2018)在长白山的研究相比, 本研究中泥炭藓初始C:N较低, 而分解却更慢, 这可能与本研究区凋落物本身多酚含量更高、土壤pH较低及土壤碳氮含量更低有一定关系.研究表明, 较高的多酚含量(Jassey et al., 2011)、较低的pH (Tahvanainen & Haraguchi, 2013)及较低的土壤养分含量(Bragazza et al., 2007)会抑制凋落物的分解.另外, 温度也是影响凋落物分解的一个重要因素.以往的研究表明, 较高的温度会促进凋落物的分解(Zhang et al., 2017), 也有研究认为较高的温度对凋落物的分解无显著影响(Breeuwer et al., 2008).本研究区温度显著高于寒温带地区, 然而分解率却低于寒温带地区, 可能与研究区其他环境因子对凋落物分解的抑制作用较强有关, 比如研究区土壤酸性高于寒温带地区, 凋落物多酚含量也更高. ...
... )及较低的土壤养分含量(Bragazza et al., 2007)会抑制凋落物的分解.另外, 温度也是影响凋落物分解的一个重要因素.以往的研究表明, 较高的温度会促进凋落物的分解(Zhang et al., 2017), 也有研究认为较高的温度对凋落物的分解无显著影响(Breeuwer et al., 2008).本研究区温度显著高于寒温带地区, 然而分解率却低于寒温带地区, 可能与研究区其他环境因子对凋落物分解的抑制作用较强有关, 比如研究区土壤酸性高于寒温带地区, 凋落物多酚含量也更高. ...
The effect of increased temperature and nitrogen deposition on decomposition in bogs 5 2008
... 近年来, 国内外****对泥炭藓属植物的生长及凋落物的分解已开展了大量的研究(Moore, 1989; Rochefort et al., 1990; Gunnarsson, 2005; Breeuwer et al., 2008; Loisel et al., 2012; 李伟等, 2013; Philben et al., 2015; Bengtsson et al., 2016; Mironov & Kondratev, 2017; Bell et al., 2018; 刘媛媛等, 2018; Newman et al., 2018).然而这些研究主要集中在高纬度的寒温带地区, 在低纬度的亚热带地区却鲜有报道(马进泽, 2018), 有关亚热带地区泥炭藓的生长及凋落物分解的原位研究更是欠缺.事实上, 泥炭藓属植物的生长及凋落物的分解主要受年平均气温、降水、植被类型、氮沉降等因素的影响(Breeuwer et al., 2008; 李伟等, 2013; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
... ), 有关亚热带地区泥炭藓的生长及凋落物分解的原位研究更是欠缺.事实上, 泥炭藓属植物的生长及凋落物的分解主要受年平均气温、降水、植被类型、氮沉降等因素的影响(Breeuwer et al., 2008; 李伟等, 2013; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
... S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
... S. ang, 小叶泥炭藓; S. cap, 尖叶泥炭鲜; S. fal, 假泥炭藓; S. fus, 锈色泥炭藓; S. mag, 中位泥炭藓; S. pal, 泥炭藓; S. rec, 喙叶泥炭藓.泥炭藓属植物学名及拉丁名参照王利松等(2018).泥炭藓属植物的生长数据均为采用Clymo (1970)方法进行野外原位测定所得, 测定时间均为生长季节, 且生长速率及净生产力统一转换为mm·d-1和g·m-2·d-1. ...
The ecology of Sphagnum// Smith AJE 2 1982
... 泥炭藓湿地是由适应能力强和耐周期性干旱的泥炭藓属(Sphagnum)植物以及草本植物沼泽化形成的一种特殊湿地类型(中国湿地植被编辑委员会, 1999).由于其净初级生产力(NPP)大于分解力, 持续吸收大气中的CO2, 并积累了大量的泥炭(Gorham, 1991; Yu, 2012), 因此, 在全球碳循环中发挥着重要作用.泥炭藓属植物作为泥炭藓湿地的优势植物, 在部分地区其盖度可达100%, 其植物残体能为湿地提供巨大的生物量(Clymo & Hayward, 1982; Orwin & Ostle, 2012), 储存在泥炭藓属植物及其残体中的碳甚至高于每年固定在所有陆地植物体中的碳量(Clymo & Hayward, 1982).泥炭藓属植物是湿地土壤碳的重要来源, 其固定的碳占据了全球土壤碳库的30% (Yu et al., 2010), 在土壤碳累积过程中发挥着关键作用(Li et al., 2004; Lang et al., 2009).因此, 研究泥炭藓的生长及凋落物的分解对于揭示湿地碳循环具有重要意义. ...
... ), 储存在泥炭藓属植物及其残体中的碳甚至高于每年固定在所有陆地植物体中的碳量(Clymo & Hayward, 1982).泥炭藓属植物是湿地土壤碳的重要来源, 其固定的碳占据了全球土壤碳库的30% (Yu et al., 2010), 在土壤碳累积过程中发挥着关键作用(Li et al., 2004; Lang et al., 2009).因此, 研究泥炭藓的生长及凋落物的分解对于揭示湿地碳循环具有重要意义. ...
Summer warming and increased winter snow cover affect Sphagnum fuscum growth, structure and production in a sub-arctic bog 1 2003
... Growth rate and net primary productivity of Sphagnum spp. in areas of different latitude Table 1
纬度 Latitude (°)
经度 Longitude (°)
年平均气温 Annual mean temperature (℃)
年降水量 Precipitation (mm)
物种 Species
生长速率 Growth rate (mm·d-1)
净生产力 Net primary productivity (g·m-2·d-1)
参考文献 Reference
68.35
18.82
0.5
320
S. fus
0.04
1.51
Dorrepaal et al., 2003
60.52
17.92
3.1
544
S. fus
0.10
0.64
Bengtsson et al., 2016
59.90
15.83
1.4
515
S. fus, S. mag
0.13
1.12
Bengtsson et al., 2016
57.15
-111.98
2.1
387
S. fus
0.09
1.68
Wieder et al., 2010
56.63
-110.20
1.1
420
S. fus
0.09
1.72
Wieder et al., 2010
55.68
-111.83
2.1
421
S. fus
0.10
1.96
Wieder et al., 2010
54.68
-113.47
1.7
500
S. fus
0.03
1.15
Thormann & Bayley, 1997
49.67
-93.72
2.6
714
S. mag, S. fus, S. ang
0.12
0.86
Rochefort et al., 1990
46.32
11.67
5.0
1 100
S. cap, S. mag, S. fal
0.26
1.70
Gerdol, 1995
39.12
-79.58
7.9
1 330
S. mag, S. rec
0.42
3.44
Wieder & Lang, 1983
30.17
109.73
7.2
1 590
S. pal
0.33
3.84
本研究 This study
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
1 1999
... 泥炭藓湿地是由适应能力强和耐周期性干旱的泥炭藓属(Sphagnum)植物以及草本植物沼泽化形成的一种特殊湿地类型(中国湿地植被编辑委员会, 1999).由于其净初级生产力(NPP)大于分解力, 持续吸收大气中的CO2, 并积累了大量的泥炭(Gorham, 1991; Yu, 2012), 因此, 在全球碳循环中发挥着重要作用.泥炭藓属植物作为泥炭藓湿地的优势植物, 在部分地区其盖度可达100%, 其植物残体能为湿地提供巨大的生物量(Clymo & Hayward, 1982; Orwin & Ostle, 2012), 储存在泥炭藓属植物及其残体中的碳甚至高于每年固定在所有陆地植物体中的碳量(Clymo & Hayward, 1982).泥炭藓属植物是湿地土壤碳的重要来源, 其固定的碳占据了全球土壤碳库的30% (Yu et al., 2010), 在土壤碳累积过程中发挥着关键作用(Li et al., 2004; Lang et al., 2009).因此, 研究泥炭藓的生长及凋落物的分解对于揭示湿地碳循环具有重要意义. ...
1 1999
... 泥炭藓湿地是由适应能力强和耐周期性干旱的泥炭藓属(Sphagnum)植物以及草本植物沼泽化形成的一种特殊湿地类型(中国湿地植被编辑委员会, 1999).由于其净初级生产力(NPP)大于分解力, 持续吸收大气中的CO2, 并积累了大量的泥炭(Gorham, 1991; Yu, 2012), 因此, 在全球碳循环中发挥着重要作用.泥炭藓属植物作为泥炭藓湿地的优势植物, 在部分地区其盖度可达100%, 其植物残体能为湿地提供巨大的生物量(Clymo & Hayward, 1982; Orwin & Ostle, 2012), 储存在泥炭藓属植物及其残体中的碳甚至高于每年固定在所有陆地植物体中的碳量(Clymo & Hayward, 1982).泥炭藓属植物是湿地土壤碳的重要来源, 其固定的碳占据了全球土壤碳库的30% (Yu et al., 2010), 在土壤碳累积过程中发挥着关键作用(Li et al., 2004; Lang et al., 2009).因此, 研究泥炭藓的生长及凋落物的分解对于揭示湿地碳循环具有重要意义. ...
Growth rate and temperature responses in bryophytes: I. An investigation of Brachythecium rutabulum 1 1982
The growth dynamics of Sphagnum based on field measurements in a temperate bog and on laboratory cultures 2 1995
... Growth rate and net primary productivity of Sphagnum spp. in areas of different latitude Table 1
纬度 Latitude (°)
经度 Longitude (°)
年平均气温 Annual mean temperature (℃)
年降水量 Precipitation (mm)
物种 Species
生长速率 Growth rate (mm·d-1)
净生产力 Net primary productivity (g·m-2·d-1)
参考文献 Reference
68.35
18.82
0.5
320
S. fus
0.04
1.51
Dorrepaal et al., 2003
60.52
17.92
3.1
544
S. fus
0.10
0.64
Bengtsson et al., 2016
59.90
15.83
1.4
515
S. fus, S. mag
0.13
1.12
Bengtsson et al., 2016
57.15
-111.98
2.1
387
S. fus
0.09
1.68
Wieder et al., 2010
56.63
-110.20
1.1
420
S. fus
0.09
1.72
Wieder et al., 2010
55.68
-111.83
2.1
421
S. fus
0.10
1.96
Wieder et al., 2010
54.68
-113.47
1.7
500
S. fus
0.03
1.15
Thormann & Bayley, 1997
49.67
-93.72
2.6
714
S. mag, S. fus, S. ang
0.12
0.86
Rochefort et al., 1990
46.32
11.67
5.0
1 100
S. cap, S. mag, S. fal
0.26
1.70
Gerdol, 1995
39.12
-79.58
7.9
1 330
S. mag, S. rec
0.42
3.44
Wieder & Lang, 1983
30.17
109.73
7.2
1 590
S. pal
0.33
3.84
本研究 This study
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
Experimental climate effect on seasonal variability of polyphenol/phenoloxidase interplay along a narrow fen-bog ecological gradient in Sphagnum fallax 1 2011
... 泥炭藓的分解相比生长更为复杂, 其分解过程不仅受地表植被类型、生境、气候条件及氮沉降等环境因素的影响(Gerdol et al., 2007; Breeuwer et al., 2008; 李伟等, 2013), 同时也受不同物种间本身属性的差异(李伟等, 2013; Bengtsson et al., 2018; 刘媛媛等, 2018)和土壤养分含量的差异(Bragazza et al., 2007)所影响.本研究3种微生境下泥炭藓凋落物野外分解12个月后最终残留率平均为92.28%, 略高于刘媛媛等(2018)在长白山哈泥泥炭地的研究结果(90%).同样是泥炭藓湿地, 李伟等(2013)在长白山的研究发现泥炭藓凋落物经历14个月的分解后最终残留率为80.8%, 显著低于本研究结果.这可能与不同凋落物本身的化学属性及研究区的环境有关.研究表明, 凋落物C:N越高, 分解越快(Limpens & Berendse, 2003).与李伟等(2013)及刘媛媛等(2018)在长白山的研究相比, 本研究中泥炭藓初始C:N较低, 而分解却更慢, 这可能与本研究区凋落物本身多酚含量更高、土壤pH较低及土壤碳氮含量更低有一定关系.研究表明, 较高的多酚含量(Jassey et al., 2011)、较低的pH (Tahvanainen & Haraguchi, 2013)及较低的土壤养分含量(Bragazza et al., 2007)会抑制凋落物的分解.另外, 温度也是影响凋落物分解的一个重要因素.以往的研究表明, 较高的温度会促进凋落物的分解(Zhang et al., 2017), 也有研究认为较高的温度对凋落物的分解无显著影响(Breeuwer et al., 2008).本研究区温度显著高于寒温带地区, 然而分解率却低于寒温带地区, 可能与研究区其他环境因子对凋落物分解的抑制作用较强有关, 比如研究区土壤酸性高于寒温带地区, 凋落物多酚含量也更高. ...
Decay and its regulation in Sphagnum peatlands 1 1993
An experimental comparison of chemical traits and litter decomposition rates in a diverse range of subarctic bryophyte, lichen and vascular plant species 1 2009
... 泥炭藓湿地是由适应能力强和耐周期性干旱的泥炭藓属(Sphagnum)植物以及草本植物沼泽化形成的一种特殊湿地类型(中国湿地植被编辑委员会, 1999).由于其净初级生产力(NPP)大于分解力, 持续吸收大气中的CO2, 并积累了大量的泥炭(Gorham, 1991; Yu, 2012), 因此, 在全球碳循环中发挥着重要作用.泥炭藓属植物作为泥炭藓湿地的优势植物, 在部分地区其盖度可达100%, 其植物残体能为湿地提供巨大的生物量(Clymo & Hayward, 1982; Orwin & Ostle, 2012), 储存在泥炭藓属植物及其残体中的碳甚至高于每年固定在所有陆地植物体中的碳量(Clymo & Hayward, 1982).泥炭藓属植物是湿地土壤碳的重要来源, 其固定的碳占据了全球土壤碳库的30% (Yu et al., 2010), 在土壤碳累积过程中发挥着关键作用(Li et al., 2004; Lang et al., 2009).因此, 研究泥炭藓的生长及凋落物的分解对于揭示湿地碳循环具有重要意义. ...
Vegetation composition controls temperature sensitivity of CO2 and CH4 emissions and DOC concentration in peatlands 1 2017
... 近年来, 国内外****对泥炭藓属植物的生长及凋落物的分解已开展了大量的研究(Moore, 1989; Rochefort et al., 1990; Gunnarsson, 2005; Breeuwer et al., 2008; Loisel et al., 2012; 李伟等, 2013; Philben et al., 2015; Bengtsson et al., 2016; Mironov & Kondratev, 2017; Bell et al., 2018; 刘媛媛等, 2018; Newman et al., 2018).然而这些研究主要集中在高纬度的寒温带地区, 在低纬度的亚热带地区却鲜有报道(马进泽, 2018), 有关亚热带地区泥炭藓的生长及凋落物分解的原位研究更是欠缺.事实上, 泥炭藓属植物的生长及凋落物的分解主要受年平均气温、降水、植被类型、氮沉降等因素的影响(Breeuwer et al., 2008; 李伟等, 2013; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
Potential of Sphagnum peat for improving soil organic matter, water holding capacity, bulk density and potato yield in a sandy soil 1 2004
... 泥炭藓湿地是由适应能力强和耐周期性干旱的泥炭藓属(Sphagnum)植物以及草本植物沼泽化形成的一种特殊湿地类型(中国湿地植被编辑委员会, 1999).由于其净初级生产力(NPP)大于分解力, 持续吸收大气中的CO2, 并积累了大量的泥炭(Gorham, 1991; Yu, 2012), 因此, 在全球碳循环中发挥着重要作用.泥炭藓属植物作为泥炭藓湿地的优势植物, 在部分地区其盖度可达100%, 其植物残体能为湿地提供巨大的生物量(Clymo & Hayward, 1982; Orwin & Ostle, 2012), 储存在泥炭藓属植物及其残体中的碳甚至高于每年固定在所有陆地植物体中的碳量(Clymo & Hayward, 1982).泥炭藓属植物是湿地土壤碳的重要来源, 其固定的碳占据了全球土壤碳库的30% (Yu et al., 2010), 在土壤碳累积过程中发挥着关键作用(Li et al., 2004; Lang et al., 2009).因此, 研究泥炭藓的生长及凋落物的分解对于揭示湿地碳循环具有重要意义. ...
长白山不同海拔泥炭地泥炭藓残体的分解 7 2013
... 近年来, 国内外****对泥炭藓属植物的生长及凋落物的分解已开展了大量的研究(Moore, 1989; Rochefort et al., 1990; Gunnarsson, 2005; Breeuwer et al., 2008; Loisel et al., 2012; 李伟等, 2013; Philben et al., 2015; Bengtsson et al., 2016; Mironov & Kondratev, 2017; Bell et al., 2018; 刘媛媛等, 2018; Newman et al., 2018).然而这些研究主要集中在高纬度的寒温带地区, 在低纬度的亚热带地区却鲜有报道(马进泽, 2018), 有关亚热带地区泥炭藓的生长及凋落物分解的原位研究更是欠缺.事实上, 泥炭藓属植物的生长及凋落物的分解主要受年平均气温、降水、植被类型、氮沉降等因素的影响(Breeuwer et al., 2008; 李伟等, 2013; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
... ; 李伟等, 2013; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
Interacting effects of elevated atmospheric CO2 and hydrology on the growth and carbon sequestration of Sphagnum moss 1 2018
... 近年来, 国内外****对泥炭藓属植物的生长及凋落物的分解已开展了大量的研究(Moore, 1989; Rochefort et al., 1990; Gunnarsson, 2005; Breeuwer et al., 2008; Loisel et al., 2012; 李伟等, 2013; Philben et al., 2015; Bengtsson et al., 2016; Mironov & Kondratev, 2017; Bell et al., 2018; 刘媛媛等, 2018; Newman et al., 2018).然而这些研究主要集中在高纬度的寒温带地区, 在低纬度的亚热带地区却鲜有报道(马进泽, 2018), 有关亚热带地区泥炭藓的生长及凋落物分解的原位研究更是欠缺.事实上, 泥炭藓属植物的生长及凋落物的分解主要受年平均气温、降水、植被类型、氮沉降等因素的影响(Breeuwer et al., 2008; 李伟等, 2013; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
Moss species effects on peatland carbon cycling after fire 1 2012
... 泥炭藓湿地是由适应能力强和耐周期性干旱的泥炭藓属(Sphagnum)植物以及草本植物沼泽化形成的一种特殊湿地类型(中国湿地植被编辑委员会, 1999).由于其净初级生产力(NPP)大于分解力, 持续吸收大气中的CO2, 并积累了大量的泥炭(Gorham, 1991; Yu, 2012), 因此, 在全球碳循环中发挥着重要作用.泥炭藓属植物作为泥炭藓湿地的优势植物, 在部分地区其盖度可达100%, 其植物残体能为湿地提供巨大的生物量(Clymo & Hayward, 1982; Orwin & Ostle, 2012), 储存在泥炭藓属植物及其残体中的碳甚至高于每年固定在所有陆地植物体中的碳量(Clymo & Hayward, 1982).泥炭藓属植物是湿地土壤碳的重要来源, 其固定的碳占据了全球土壤碳库的30% (Yu et al., 2010), 在土壤碳累积过程中发挥着关键作用(Li et al., 2004; Lang et al., 2009).因此, 研究泥炭藓的生长及凋落物的分解对于揭示湿地碳循环具有重要意义. ...
Temperature, oxygen, and vegetation controls on decomposition in a James Bay peatland 1 2015
... 近年来, 国内外****对泥炭藓属植物的生长及凋落物的分解已开展了大量的研究(Moore, 1989; Rochefort et al., 1990; Gunnarsson, 2005; Breeuwer et al., 2008; Loisel et al., 2012; 李伟等, 2013; Philben et al., 2015; Bengtsson et al., 2016; Mironov & Kondratev, 2017; Bell et al., 2018; 刘媛媛等, 2018; Newman et al., 2018).然而这些研究主要集中在高纬度的寒温带地区, 在低纬度的亚热带地区却鲜有报道(马进泽, 2018), 有关亚热带地区泥炭藓的生长及凋落物分解的原位研究更是欠缺.事实上, 泥炭藓属植物的生长及凋落物的分解主要受年平均气温、降水、植被类型、氮沉降等因素的影响(Breeuwer et al., 2008; 李伟等, 2013; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
Growth, production, and decomposition dynamics of Sphagnum under natural and experimentally acidified conditions 2 1990
... 近年来, 国内外****对泥炭藓属植物的生长及凋落物的分解已开展了大量的研究(Moore, 1989; Rochefort et al., 1990; Gunnarsson, 2005; Breeuwer et al., 2008; Loisel et al., 2012; 李伟等, 2013; Philben et al., 2015; Bengtsson et al., 2016; Mironov & Kondratev, 2017; Bell et al., 2018; 刘媛媛等, 2018; Newman et al., 2018).然而这些研究主要集中在高纬度的寒温带地区, 在低纬度的亚热带地区却鲜有报道(马进泽, 2018), 有关亚热带地区泥炭藓的生长及凋落物分解的原位研究更是欠缺.事实上, 泥炭藓属植物的生长及凋落物的分解主要受年平均气温、降水、植被类型、氮沉降等因素的影响(Breeuwer et al., 2008; 李伟等, 2013; Bengtsson et al., 2016; Leroy et al., 2017).比如Breeuwer等(2008)在西欧的研究表明, 维管植物的分解速率随气温升高而增加, 而泥炭藓残体的分解速率随气温升高变化不明显, 甚至有减小趋势.相反, 我国****在长白山的研究结果表明增温能够促进泥炭藓凋落物的分解(李伟等, 2013).不同地区的泥炭藓植物生长及凋落物分解的规律并不一致, 这与研究区气候条件、微生境、植物本身属性等有关(Loisel et al., 2012; 刘媛媛等, 2018).亚热带地区年平均气温明显高于寒温带地区, 较高的有效积温和充足的降水更有利于泥炭藓的生长.因此, 我们推测亚热带地区泥炭藓的生长及凋落物的分解将不同于寒温带地区, 甚至亚热带地区泥炭藓的生长速率高于寒温带地区. ...
... Growth rate and net primary productivity of Sphagnum spp. in areas of different latitude Table 1
纬度 Latitude (°)
经度 Longitude (°)
年平均气温 Annual mean temperature (℃)
年降水量 Precipitation (mm)
物种 Species
生长速率 Growth rate (mm·d-1)
净生产力 Net primary productivity (g·m-2·d-1)
参考文献 Reference
68.35
18.82
0.5
320
S. fus
0.04
1.51
Dorrepaal et al., 2003
60.52
17.92
3.1
544
S. fus
0.10
0.64
Bengtsson et al., 2016
59.90
15.83
1.4
515
S. fus, S. mag
0.13
1.12
Bengtsson et al., 2016
57.15
-111.98
2.1
387
S. fus
0.09
1.68
Wieder et al., 2010
56.63
-110.20
1.1
420
S. fus
0.09
1.72
Wieder et al., 2010
55.68
-111.83
2.1
421
S. fus
0.10
1.96
Wieder et al., 2010
54.68
-113.47
1.7
500
S. fus
0.03
1.15
Thormann & Bayley, 1997
49.67
-93.72
2.6
714
S. mag, S. fus, S. ang
0.12
0.86
Rochefort et al., 1990
46.32
11.67
5.0
1 100
S. cap, S. mag, S. fal
0.26
1.70
Gerdol, 1995
39.12
-79.58
7.9
1 330
S. mag, S. rec
0.42
3.44
Wieder & Lang, 1983
30.17
109.73
7.2
1 590
S. pal
0.33
3.84
本研究 This study
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent 1 1999
... 将取回的尼龙网袋于室内, 清除表面上的泥土和植物根系后, 置于100目筛(孔径0.15 mm)中, 用蒸馏水缓慢冲洗, 冲洗干净后于75 ℃烘干至恒质量, 称量记录凋落物的残留量.将残留的凋落物用粉碎机粉碎, 过50目筛后, 用元素分析仪(Elementar Vario EL III, Elementar, Langenselbold, Germany)测定残留物中的总碳(C)和总氮(N)含量, 用Folin- Ciocalteu法测多酚含量(Singleton et al., 1999). ...
Effect of pH on phenol oxidase activity on decaying Sphagnum mosses 1 2013
... 泥炭藓的分解相比生长更为复杂, 其分解过程不仅受地表植被类型、生境、气候条件及氮沉降等环境因素的影响(Gerdol et al., 2007; Breeuwer et al., 2008; 李伟等, 2013), 同时也受不同物种间本身属性的差异(李伟等, 2013; Bengtsson et al., 2018; 刘媛媛等, 2018)和土壤养分含量的差异(Bragazza et al., 2007)所影响.本研究3种微生境下泥炭藓凋落物野外分解12个月后最终残留率平均为92.28%, 略高于刘媛媛等(2018)在长白山哈泥泥炭地的研究结果(90%).同样是泥炭藓湿地, 李伟等(2013)在长白山的研究发现泥炭藓凋落物经历14个月的分解后最终残留率为80.8%, 显著低于本研究结果.这可能与不同凋落物本身的化学属性及研究区的环境有关.研究表明, 凋落物C:N越高, 分解越快(Limpens & Berendse, 2003).与李伟等(2013)及刘媛媛等(2018)在长白山的研究相比, 本研究中泥炭藓初始C:N较低, 而分解却更慢, 这可能与本研究区凋落物本身多酚含量更高、土壤pH较低及土壤碳氮含量更低有一定关系.研究表明, 较高的多酚含量(Jassey et al., 2011)、较低的pH (Tahvanainen & Haraguchi, 2013)及较低的土壤养分含量(Bragazza et al., 2007)会抑制凋落物的分解.另外, 温度也是影响凋落物分解的一个重要因素.以往的研究表明, 较高的温度会促进凋落物的分解(Zhang et al., 2017), 也有研究认为较高的温度对凋落物的分解无显著影响(Breeuwer et al., 2008).本研究区温度显著高于寒温带地区, 然而分解率却低于寒温带地区, 可能与研究区其他环境因子对凋落物分解的抑制作用较强有关, 比如研究区土壤酸性高于寒温带地区, 凋落物多酚含量也更高. ...
Aboveground net primary production along a bog-fen-marsh gradient in southern boreal Alberta, Canada 1 1997
... Growth rate and net primary productivity of Sphagnum spp. in areas of different latitude Table 1
纬度 Latitude (°)
经度 Longitude (°)
年平均气温 Annual mean temperature (℃)
年降水量 Precipitation (mm)
物种 Species
生长速率 Growth rate (mm·d-1)
净生产力 Net primary productivity (g·m-2·d-1)
参考文献 Reference
68.35
18.82
0.5
320
S. fus
0.04
1.51
Dorrepaal et al., 2003
60.52
17.92
3.1
544
S. fus
0.10
0.64
Bengtsson et al., 2016
59.90
15.83
1.4
515
S. fus, S. mag
0.13
1.12
Bengtsson et al., 2016
57.15
-111.98
2.1
387
S. fus
0.09
1.68
Wieder et al., 2010
56.63
-110.20
1.1
420
S. fus
0.09
1.72
Wieder et al., 2010
55.68
-111.83
2.1
421
S. fus
0.10
1.96
Wieder et al., 2010
54.68
-113.47
1.7
500
S. fus
0.03
1.15
Thormann & Bayley, 1997
49.67
-93.72
2.6
714
S. mag, S. fus, S. ang
0.12
0.86
Rochefort et al., 1990
46.32
11.67
5.0
1 100
S. cap, S. mag, S. fal
0.26
1.70
Gerdol, 1995
39.12
-79.58
7.9
1 330
S. mag, S. rec
0.42
3.44
Wieder & Lang, 1983
30.17
109.73
7.2
1 590
S. pal
0.33
3.84
本研究 This study
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
Sphagnum production and decomposition in a restored cutover peatland 1 2003
... S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
... S. ang, 小叶泥炭藓; S. cap, 尖叶泥炭鲜; S. fal, 假泥炭藓; S. fus, 锈色泥炭藓; S. mag, 中位泥炭藓; S. pal, 泥炭藓; S. rec, 喙叶泥炭藓.泥炭藓属植物学名及拉丁名参照王利松等(2018).泥炭藓属植物的生长数据均为采用Clymo (1970)方法进行野外原位测定所得, 测定时间均为生长季节, 且生长速率及净生产力统一转换为mm·d-1和g·m-2·d-1. ...
2 2018
... S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
... S. ang, 小叶泥炭藓; S. cap, 尖叶泥炭鲜; S. fal, 假泥炭藓; S. fus, 锈色泥炭藓; S. mag, 中位泥炭藓; S. pal, 泥炭藓; S. rec, 喙叶泥炭藓.泥炭藓属植物学名及拉丁名参照王利松等(2018).泥炭藓属植物的生长数据均为采用Clymo (1970)方法进行野外原位测定所得, 测定时间均为生长季节, 且生长速率及净生产力统一转换为mm·d-1和g·m-2·d-1. ...
Net primary production of the dominant bryophytes in a Sphagnum-dominated wetland in West Virginia 2 1983
... Growth rate and net primary productivity of Sphagnum spp. in areas of different latitude Table 1
纬度 Latitude (°)
经度 Longitude (°)
年平均气温 Annual mean temperature (℃)
年降水量 Precipitation (mm)
物种 Species
生长速率 Growth rate (mm·d-1)
净生产力 Net primary productivity (g·m-2·d-1)
参考文献 Reference
68.35
18.82
0.5
320
S. fus
0.04
1.51
Dorrepaal et al., 2003
60.52
17.92
3.1
544
S. fus
0.10
0.64
Bengtsson et al., 2016
59.90
15.83
1.4
515
S. fus, S. mag
0.13
1.12
Bengtsson et al., 2016
57.15
-111.98
2.1
387
S. fus
0.09
1.68
Wieder et al., 2010
56.63
-110.20
1.1
420
S. fus
0.09
1.72
Wieder et al., 2010
55.68
-111.83
2.1
421
S. fus
0.10
1.96
Wieder et al., 2010
54.68
-113.47
1.7
500
S. fus
0.03
1.15
Thormann & Bayley, 1997
49.67
-93.72
2.6
714
S. mag, S. fus, S. ang
0.12
0.86
Rochefort et al., 1990
46.32
11.67
5.0
1 100
S. cap, S. mag, S. fal
0.26
1.70
Gerdol, 1995
39.12
-79.58
7.9
1 330
S. mag, S. rec
0.42
3.44
Wieder & Lang, 1983
30.17
109.73
7.2
1 590
S. pal
0.33
3.84
本研究 This study
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
Nitrogen and sulphur deposition and the growth of Sphagnum fuscum in bogs of the Athabasca Oil Sands Region, Alberta 3 2010
... Growth rate and net primary productivity of Sphagnum spp. in areas of different latitude Table 1
纬度 Latitude (°)
经度 Longitude (°)
年平均气温 Annual mean temperature (℃)
年降水量 Precipitation (mm)
物种 Species
生长速率 Growth rate (mm·d-1)
净生产力 Net primary productivity (g·m-2·d-1)
参考文献 Reference
68.35
18.82
0.5
320
S. fus
0.04
1.51
Dorrepaal et al., 2003
60.52
17.92
3.1
544
S. fus
0.10
0.64
Bengtsson et al., 2016
59.90
15.83
1.4
515
S. fus, S. mag
0.13
1.12
Bengtsson et al., 2016
57.15
-111.98
2.1
387
S. fus
0.09
1.68
Wieder et al., 2010
56.63
-110.20
1.1
420
S. fus
0.09
1.72
Wieder et al., 2010
55.68
-111.83
2.1
421
S. fus
0.10
1.96
Wieder et al., 2010
54.68
-113.47
1.7
500
S. fus
0.03
1.15
Thormann & Bayley, 1997
49.67
-93.72
2.6
714
S. mag, S. fus, S. ang
0.12
0.86
Rochefort et al., 1990
46.32
11.67
5.0
1 100
S. cap, S. mag, S. fal
0.26
1.70
Gerdol, 1995
39.12
-79.58
7.9
1 330
S. mag, S. rec
0.42
3.44
Wieder & Lang, 1983
30.17
109.73
7.2
1 590
S. pal
0.33
3.84
本研究 This study
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
... Wieder et al., 2010
55.68
-111.83
2.1
421
S. fus
0.10
1.96
Wieder et al., 2010
54.68
-113.47
1.7
500
S. fus
0.03
1.15
Thormann & Bayley, 1997
49.67
-93.72
2.6
714
S. mag, S. fus, S. ang
0.12
0.86
Rochefort et al., 1990
46.32
11.67
5.0
1 100
S. cap, S. mag, S. fal
0.26
1.70
Gerdol, 1995
39.12
-79.58
7.9
1 330
S. mag, S. rec
0.42
3.44
Wieder & Lang, 1983
30.17
109.73
7.2
1 590
S. pal
0.33
3.84
本研究 This study
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
... Wieder et al., 2010
54.68
-113.47
1.7
500
S. fus
0.03
1.15
Thormann & Bayley, 1997
49.67
-93.72
2.6
714
S. mag, S. fus, S. ang
0.12
0.86
Rochefort et al., 1990
46.32
11.67
5.0
1 100
S. cap, S. mag, S. fal
0.26
1.70
Gerdol, 1995
39.12
-79.58
7.9
1 330
S. mag, S. rec
0.42
3.44
Wieder & Lang, 1983
30.17
109.73
7.2
1 590
S. pal
0.33
3.84
本研究 This study
S. ang, S. angustifolium; S. cap, S. capillifolium; S. fal, S. fallax; S. fus, S. fuscum; S. mag, S. magellanicum; S. pal, S. palustre; S. rec, S. recurvum. Scientific and Latin names of Sphagnum spp. refer to Wang et al. (2018). The growth data of Sphagnum spp. in the table were all obtained by field measurement using Clymo (1970) method. Measuring time covers whole growing season; the growth rate and net primary productivity were converted to mm·d-1 and g·m-2·d-1. ...
Effects of snow-load and shading by vascular plants on the vertical growth of hummocks formed by Sphagnum papillosum in a mire of northern Japan 1 2012
 ,1,2, 吴林
,1,2, 吴林 ,1,2,*, 王涵1,2, 洪柳1,2, 熊莉军3
,1,2,*, 王涵1,2, 洪柳1,2, 熊莉军3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT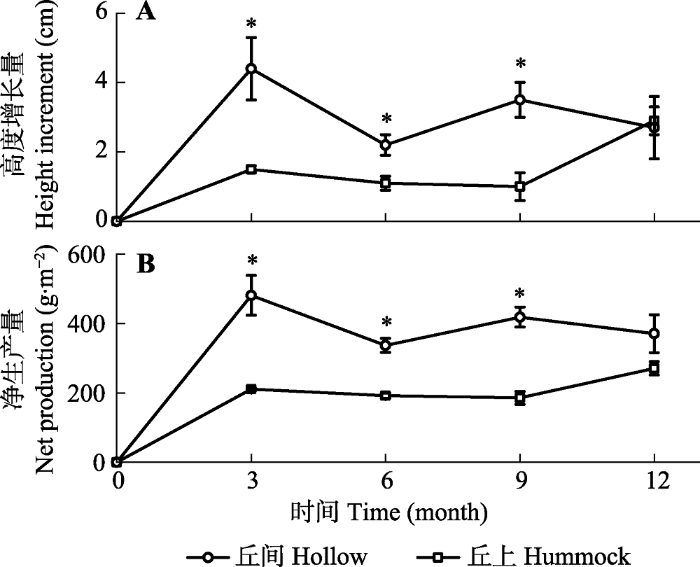 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT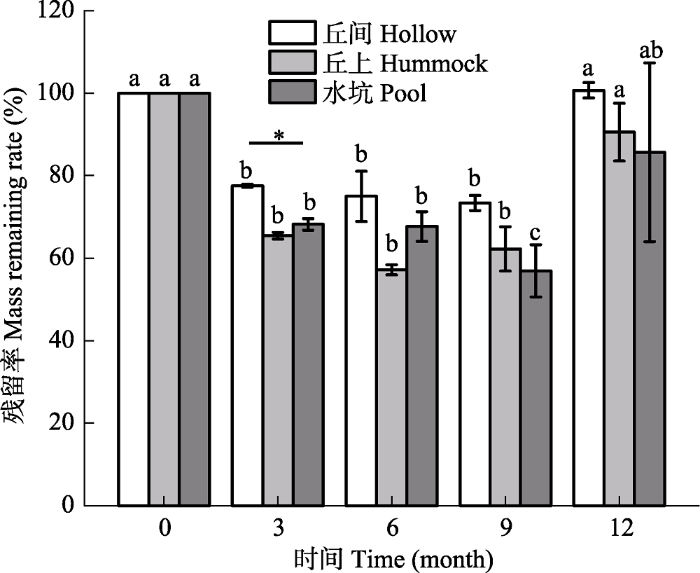 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT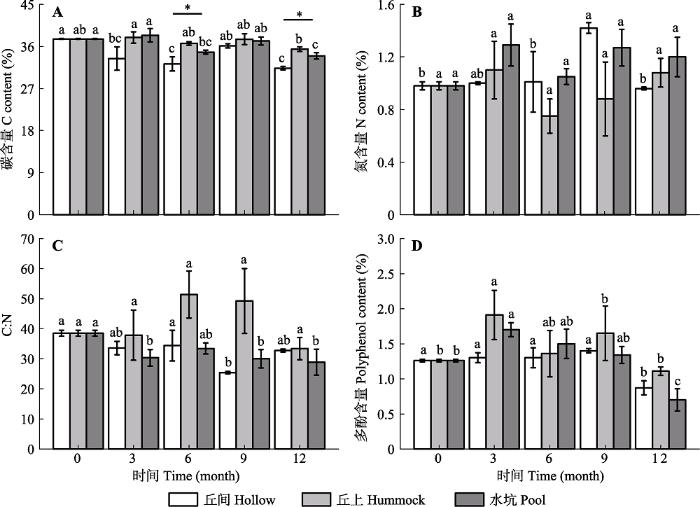 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT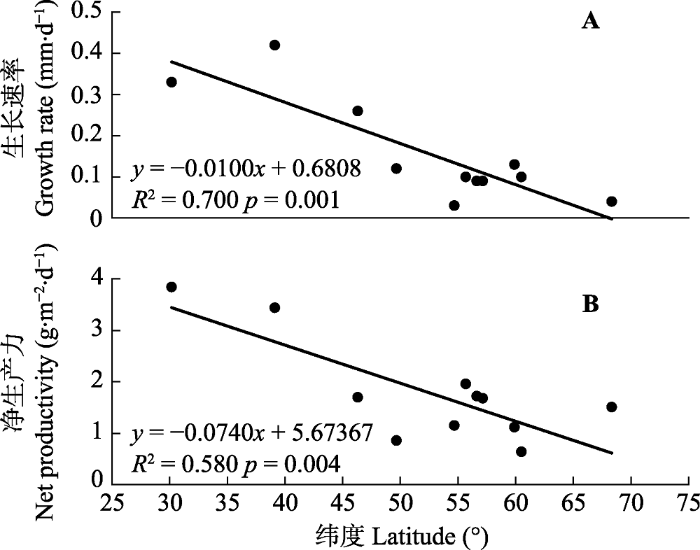 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}