季倩雯
1 , 2 , 郑成洋
, 1 , 2 , 3 , * , 张磊
4 , 曾发旭
1 , 3 1 北京大学生态研究中心, 北京 1008712 北京大学城市与环境学院地表过程与模拟教育部重点实验室, 北京 1008713 北京大学生态与环境观测系统塞罕坝实验站, 河北承德 0684664 河北省塞罕坝机械林场, 河北承德 068466Stem radial growth dynamics of Pinus sylvestris var. mongolica and their relationship with meteorological factor in Saihanba, Hebei, China Qian-Wen JI
1 , 2 , Cheng-Yang ZHENG
, 1 , 2 , 3 , * , Lei ZHANG
4 , Fa-Xu ZENG
1 , 3 1 Institute of Ecology, Peking University, Beijing 100871, China2 Key Laboratory of Surface Processes and Simulation, Ministry of Education, College of Urban and Environmental Science, Peking University, Beijing 100871, China3 Saihanba Station, Peking University Observatory System for Ecology and the Environment, Chengde, Hebei 068466, China4 Saihanba Machinery Forestry Centre, Chengde, Hebei 068466, China通讯作者: *
zhengcy@pku.edu.cn 编委: 黄建国
责任编辑: 李敏
收稿日期: 2019-12-6
接受日期: 2020-02-1
网络出版日期: 2020-03-20
基金资助: 国家重点研发计划 (2017YFC0503903 )国家自然科学基金 (31621091 )
Corresponding authors: *
zhengcy@pku.edu.cn Received: 2019-12-6
Accepted: 2020-02-1
Online: 2020-03-20
Fund supported: National Key R&D Program of China (2017YFC0503903 )Natural Science Foundation of China (31621091 )
摘要 树木是森林生态系统的基本组成, 其生长受气象因子的影响, 基于此, 该研究通过监测樟子松(
Pinus sylvestris var.
mongolica )的径向生长, 研究樟子松生长日动态规律、季节动态规律及其与气象因子的关系, 探讨河北塞罕坝地区樟子松森林生态系统对气候变化的响应机制。此外, 以往研究树木生长大多数基于树轮年代学, 缺少短期树木径向生长动态的研究。该研究利用径向生长记录仪监测河北塞罕坝机械林场内樟子松连续3年(2016-2018)的树干径向动态变化。结果表明: 由于树干的水分吸收与蒸腾作用, 樟子松树干径向昼夜变化呈现季节性规律, 可划分为4个阶段: 春季萌动期、夏季生长期、秋冬交替期和冬季休眠期。塞罕坝樟子松树干径向生长开始于每年4月初; 4月初至5月中旬为水分恢复阶段; 5月中旬至7月中旬为快速生长阶段; 7月中旬至10月中旬为缓慢生长阶段; 10月中、下旬生长趋于停止, 并有树干径向收缩现象。以一天为时间尺度, 在快速生长阶段(5月初至7月中旬)樟子松径向生长主要受空气温度的影响; 缓慢生长阶段(7月中旬至10月下旬)降水量、空气温度均影响樟子松径向生长。以15天为时间尺度, 温度对樟子松径向生长的影响显著。结果显示樟子松的生长动态规律及其影响因子, 为未来樟子松生理研究提供参考时间节点, 同时在极端低温与干旱的情况下, 为半干旱地区樟子松的生长状态提供参考依据。
关键词: 径向生长测量仪 ;
樟子松 ;
生长动态 ;
气象因子 ;
塞罕坝 Abstract Aims Trees are the basic components of forest ecosystems, and their growth is affected by meteorological factors. It is important to explore the response mechanism of forest ecosystems in this region to climate change if radial growth dynamics of dominant tree species and their relationship with meteorological factors are determined. Furthermore, previous studies on tree growth were mostly based on tree-ring chronology, whereas short- term studies on the stem radial growth dynamics of trees are scare. Therefore, this study aims to investigate the daily and seasonal dynamics of radial growth in
Pinus sylvestris var.
mongolica and its relationship with meteorological factors.
Methods In this study, dendrometer increment sensor was used to monitor the stem radial dynamic changes of
P. sylvestris var
. mongolica for three years from 2016 to 2018 in the Saihanba, Hebei Province, China.
Important findings Due to the water absorption and transpiration of the stems, the diurnal variation of the stem radial can be divided into four periods: spring germination period; summer growth period; autumn-winter alternating period; winter dormant period. Stem radial growth begins at early April in each year during water recovery from early April to mid-May. The fast-growing stage occurred from mid-May to mid-July. From mid-July to mid-October, trees entered a slow-growing stage. The radial growth tended to stop during mid- and late-October. Temperature was the main meteorological factor influencing the stem radial growth. On the time scale of one day, the stem radial growth was mainly affected by temperature during the rapid growth stage, but by rainfall and air temperature during slow growing stage. On the time scale of 15 days, only temperature had a significant effect on the stem radial growth. Under the conditions of extreme low temperature and drought, our results provide important reference for the growth status of
P. sylvestris var.
mongolica in semi-arid areas.
Keywords: Dendrometer ;
Pinus sylvestris var. mongolica ;
growth dynamic ;
meteorological factor ;
Saihanba PDF (1437KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 季倩雯, 郑成洋, 张磊, 曾发旭. 河北塞罕坝樟子松径向生长动态变化及其与气象因子的关系. 植物生态学报, 2020, 44(3): 257-265. DOI:
10.17521/cjpe.2019.0340 JI Qian-Wen, ZHENG Cheng-Yang, ZHANG Lei, ZENG Fa-Xu.
Stem radial growth dynamics of Pinus sylvestris var. mongolica and their relationship with meteorological factor in Saihanba, Hebei, China .
Chinese Journal of Plant Ecology , 2020, 44(3): 257-265. DOI:
10.17521/cjpe.2019.0340 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 )。以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究。树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态。基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 )。近年来, 国内外采用树木径向生长记录仪的研究逐渐增多。
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度。
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化。
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系。
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素。在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子。然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少。塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 )。樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义。在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义。本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系。
1 材料和方法 1.1 研究区域概况 研究区域位于河北省围场满族蒙古族自治县北部塞罕坝机械林场总场千层板林场(42.41° N, 117.24° E, 海拔1 431 m), 处于内蒙古高原、大兴安岭山系与冀北山地交汇之处。属暖温带落叶阔叶林与温带草原的过渡带, 地处半干旱半湿润气候区, 冬季漫长且寒冷, 春秋季节较短, 夏季气候凉爽。研究区域昼夜温差较大, 年平均气温-1.4 ℃, 极端最高、最低气温分别为30.9和-42.8 ℃。年日照时间为2 368 h, 年无霜期60天。年降水量为438 mm, 其中6-8月占68% (
张建东等, 2019 )。塞罕坝及周边地区分布的土壤类型有灰色森林土、黑钙土、淡黑钙土、棕壤、暗栗钙土、亚高山草甸土和风沙土(
郑成洋, 2005 ), 但塞罕坝林场土壤类型以风沙土占绝大多数, 主要成土母质为风积物、残积物及冲积物。
樟子松具有耐寒、耐旱、耐贫瘠的优良特性。1963年塞罕坝地区开始营造人工林, 树种选择以落叶松为主, 但一些较为贫瘠、干旱地块落叶松不易成活。在20世纪60年代末才开始引种樟子松。现今塞罕坝林场樟子松面积约为7 500 hm
2 , 占塞罕坝总经营面积的8.0%左右(
戴继先等, 2002 )。人工樟子松林林下灌木、草本稀疏, 林下灌木物种主要有: 稠李(
Padus racemosa )、兴安胡枝子(
Lespedeza daurica )、山刺玫(
Rosa davurica )等; 草本以大披针薹草(
Carex lanceolata )、腺毛委陵菜(
Potentilla longifolia )、地榆(
Sanguisorba officinalis )、瓣蕊唐松草(
Thalictrum petaloideum )等为主。
1.2 数据收集 2015年4月在研究样地内选择3株生长良好、树干较为笔直的樟子松作为样木。在树高1.5 m处安装树木径向生长记录仪, 间隔0.5 h实时记录胸径变化。使用2016年1月至2018年12月时间段数据。树木径向生长记录仪能够原位连续记录树木生长的精确数据, 记录树干日动态的收缩和膨胀变化。气象数据来源: 样地内安装EM50 5通道数据采集器(Decagon, Pullman, USA), 记录空气温度、空气相对湿度和5、15、25 cm土壤温湿度, 数据采集时间间隔为0.5 h。此外, 选择距离实验样地500 m的标准气象站(42.42° N, 117.25° E), 收集降水量、空气温度和空气湿度等数据。
1.3 数据统计分析 樟子松径向昼夜变化采用正弦函数(
King et al ., 2013 ), 在波形的临界点处找到局部最大值(
R max )和最小值(
R min ), 计算每日最大值(
T max )和最小值(
T min )的时间, 以及振幅(峰值幅度), 计算公式为
R =
R max -
R min , 若径向变化在同一天呈现单一的波动, 则没有度量值。密集图代表性选择春夏、秋冬交替阶段的日平均树木径向变化规律, 即3月底至5月初与9月底至11月初, 观察昼夜径向变化波形特征。樟子松径向生长期的确定方法: 采用15天的树木径向日变化来确定生长季的开始与结束时期; 使用单样本
t 检验评估15天树干径向变化均值与0是否有差异, 差异显著且为正值时, 判断为树木生长(
Deslaurier et al ., 2003 )。樟子松径向日变化采用最大值法(
Bouriaud et al ., 2005 ), 从树木径向生长记录数据中, 提取樟子松每日48个生长数据中的最大值, 计算连续两日的最大值之差, 以作为树干径向日变化值(Δ
R )。日变化值参与气象因子的关系分析。本研究数据统计分析使用SPSS 17.0, 树木径向生长与气象因子的关系采用Pearson相关分析方法; 作图使用Origin 2018c。
2 结果 2.1 樟子松树干径向日变化的季节动态特征 2016-2018年樟子松的径向日变化具有相似规律, 日变化为正负振幅波动。一年内径向日变化的振幅在0.05-0.35 mm之间。樟子松树干径向变化特征可以分为4个阶段(
图1 -3): 第一阶段: 春季萌动期。4月7日前后为塞罕坝初春季节, 昼夜径向变化振幅开始减小, 4月7日至4月19日, 全天径向变化保持平缓, 无较大波动, 4月20日开始, 昼夜径向变化出现波动, 最大值与最小值出现时间与4月初的特征相反, 逐渐完成冬季到春夏季的转换。第二阶段: 夏季生长期。每年夏季5至9月, 树干径向昼夜变化振幅减小, 振幅最大值出现在8:00-10:00, 最小值出现在15:00-17:00。第三阶段: 秋冬交替期。从9月20日左右昼夜振幅开始逐渐减小, 最大振幅不超过0.08 mm, 10月中旬左右, 树干昼夜变化不稳定; 10月下旬, 树木径向昼夜振幅大幅增大, 在11月初树干昼夜振幅逐渐稳定, 完成转化且最大振幅维持在0.35 mm。第四阶段: 冬季休眠期。每年冬季11月到翌年3月, 树干径向昼夜变化振幅较大, 最大日振幅达0.35 mm, 其中振幅最大值出现在傍晚18:00-20:00, 最小值出现在10:00-12:00。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1河北塞罕坝月平均樟子松径向日变化动态。 Fig. 1Diurnal change of monthly average radial variation of Pinus sylvestris var. mongolica in Saihanba, Hebei. 图2 新窗口打开 |
下载原图ZIP |
生成PPT 图23月20日-5月20日河北塞罕坝樟子松径向日动态变化。 Fig. 2Diurnal change of stem radial growth in Pinus sylvestris var. mongolica from March 20 to May 20 in Saihanba, Hebei. 图3 新窗口打开 |
下载原图ZIP |
生成PPT 图39月20日-11月20日河北塞罕坝樟子松径向日动态变化。 Fig. 3The diurnal change of stem radial growth in Pinus sylvestris var. mongolica from September 20 to November 20 in Saihanba, Hebei. 2.2 樟子松树干径向变化的季节动态 分析2016-2018年树干径向变化数据, 树干径向变化可以大致分为4个阶段(
图4 ): (1)树干水分恢复阶段(4月初至5月中旬), 该阶段树干径向变化出现了不稳定的迅速增粗; (2)树干快速生长阶段(5月中旬至7月中旬), 树干径向出现持续增长, 树木进入了稳定持续的生长时段; (3)树干缓慢生长阶段(7月中旬至10月下旬), 树干径向变化趋于平稳不变, 进入稳定平衡期; (4)树干收缩阶段(10月下旬至翌年3月初), 生长停止, 树干各组织开始逐渐收缩。15天间隔樟子松径向生长结果显示(
图5 ), 5月初开始, 树木径向生长与0有显著差异且为正值, 树木开始迅速生长, 此后至7月初连续出现4个显著差异且为正值, 进入7月中旬, 树木径向生长与0无显著差异。综上分析, 樟子松的生长期开始于5月初, 此后树木生长迅速, 7月中旬开始树木径向变化趋于平稳不变, 10月下旬树木径向变化趋于停止, 开始出现收缩现象。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图42016-2018年河北塞罕坝樟子松年累积径向平均变化。 Fig. 4The yearly accumulation of radial growth in Pinus sylvestris var. mongolica during 2016-2018 in Saihanba, Hebei. 图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5河北塞罕坝4-10月以15天为间隔的樟子松径向变化。斜线箱图表示径向生长量与0有显著差异(单样本t 检验, p < 0.05)。 Fig. 5Stem radial growth of Pinus sylvestris var. mongolica at 15 d interval from April to October in Saihanba, Hebei. Diagonal box shows that stem radial variation is significantly different from zero. 2.3 樟子松树干径向变化的影响因子 用Pearson相关分析研究樟子松径向日变化与降水量、空气温度、土壤温度等气象因子的关系(
图6 ;
表1 )。结果显示, 樟子松径向生长在快速生长阶段(5月初至7月中旬)主要受同一时期土壤温度的显著影响, 且呈正相关关系。樟子松径向生长在稳定生长阶段(7月中旬至10下旬)与降水量和空气温度呈显著正相关关系。以15天为时间尺度, 进一步分析降水量与温度对樟子松径向变化的影响, 结果(
图6 )显示, 随着时间尺度的扩大, 樟子松的生长受温度影响显著, 降水量对樟子松径向生长有影响, 但不显著。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6河北塞罕坝樟子松径向变化与气象因子的关系。 Fig. 6Dynamic relationship between radial variation and climatic factors of Pinus sylvestris var. mongolica in Saihanba, Hebei. Table 1 表1 表1 河北塞罕坝快速生长阶段和稳定生长阶段樟子松径向变化量与气象因子的Pearson相关关系分析
Table 1
Pearson correlation analysis between the radial variation of Pinus sylvestris var. mongolica and the meteorological factors during fast growth and stable growth phases in Saihanba, Hebei 阶段 Phase 降水量 Precipitation 土壤温度 Soil temperature 空气温度 Air temperature 降水量 Precipitation (15 d) 空气温度 Air temperature (15 d) 生长 Growth 0.247 0.363** 0.241 0.755 0.897** 稳定 Stabilization 0.400** 0.169 0.218** 0.135 0.816
The numbers in the table represent Pearson correlation coefficient; **, p < 0.01. 表中数字为Pearson相关系数; **, p < 0.01。 新窗口打开 |
下载CSV 3 讨论 3.1 樟子松树干径向日变化的成因 樟子松日径向变化曲线与树木蒸腾作用和冬季温度存在相关性(
Zweifel et al ., 2001 ;
Turcotte et al ., 2011 ;
Ehrenberger et al ., 2012 )。
Chan等(2016) 利用胸径测量仪数据与气象数据, 模拟生长曲线并预测出径向日变化与茎水势的变化近似成线性比例; 本研究结果中, 樟子松径向日变化与水分有密切关系。春夏季节, 径向昼夜变化是由于水势梯度与蒸腾过程强烈耦合作用(
Zweifel & H?sler, 2000 )。春季土壤温度高于0 ℃, 夜晚根部吸水充分, 早上气温升高, 树木开始冠层蒸腾作用, 茎干组织产生水势梯度, 水分从根部输送至茎干组织, 水分的吸收大于树冠蒸腾的损失, 此时树干半径增加; 水势相同时开始, 直到傍晚期间, 树冠的水分需求超过了根部的吸水量, 导致树干木材水势为负, 引发水分从树皮弹性组织向木质部转移, 树木茎干收缩。从傍晚开始, 蒸腾作用减小, 茎干水分得到补充(
Sevanto et al ., 2011 ), 树木茎干膨胀。
秋冬季节, 蒸腾作用不再是树木茎干变化的驱动因素, 气温成为树干茎干变化的决定因素(
Sevanto et al ., 2006 ;
King et al ., 2013 )。由于冬季初期温差较大, 树木茎干在冰冻状态与解冻状态之间切换(
Deslauriers et al ., 2003 ), 当温度到达树液凝固点时, 木材细胞开始冻结, 从而导致树皮细胞液的渗透性排出, 茎干收缩(
Zweifel & H?sler, 2000 ;
Pearce, 2001 ;
Strimbeck et al ., 2008 )。当白天温度足够高时, 上述过程发生逆转, 水分开始流回细胞, 导致茎膨胀并达到每日最大值(
Turcotte et al ., 2009 )。本研究得出的昼夜变化规律与
King等(2013) 对阿尔卑斯山落叶松的研究结果基本一致, 但不同的是, 本研究冬季振幅大于夏季, 原因可能是振幅与持续时间的长短主要取决于温度与水分的转移速度(
Deslauriers et al ., 2007a )。塞罕坝每年2-3月气温升高, 昼夜温差变大, 导致振幅增大。
3.2 樟子松树干径向变化的成因 Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致。从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象。随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期。经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳。研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 )。在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 )。10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止。树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 )。
3.3 樟子松树干径向变化与气象因子的关系 樟子松树干径向变化主要受水分与温度的影响, 但在年际间不同的生长阶段, 气象因子如何影响树干径向生长的研究报道较少。河北省塞罕坝地区进入4月后, 林内积雪开始融化, 土壤含水量增加, 树干径向生长是由于空气温度升高所致, 茎干在没有达到水分平衡的情况下, 达到树木生长有效积温, 引发形成层活动与细胞分化, 因而引发茎干生长(
Gruber et al ., 2009 ;
Swidrak et al ., 2011 )。空气温度引发空气湿度的改变, 空气湿度影响树冠蒸腾, 从而改变樟子松茎干组织的水分和形成层细胞膨压。生长稳定时期主要发生在每年的7至10月, 树木完成前期新生细胞的分裂和生长, 进入新生细胞木质化阶段, 树木径向大小基本保持稳定。在生长稳定、茎干变化不明显的情况下, 树木径向日变化量主要由茎干水分输送改变, 而降水量和空气温度共同影响水分的输送。研究发现, 15天为时间尺度时, 降水不再是樟子松径向生长的显著影响因素, 温度是径向生长的显著影响因素。
王晓春等(2011) 对大兴安岭北部樟子松树轮与气候响应的研究显示, 虽然温度对樟子松径向生长的影响在不同样点的月份存在差异, 但温度是樟子松径向生长的主要限制因子。但是
牛豪阁等(2018) 对祁连山青扦2013-2015年的树轮与气象关系的研究发现, 降水对在5-7月对青扦的影响是稳定显著的。这可能与樟子松的生理特性与形成层细胞结构有关, 后续需要对树木解剖学与形成层细胞生长进行研究。
[1] Anfodillo T Rento S Carraro V Furlanetto L Urbinati C Carrer M 1998 ). Tree water relations and climatic variations at the alpine timberline: seasonal changes of sap flux and xylem water potential in Larix decidua Miller, Picea abies ( L.) Karst. and Pinus cembra LAnnales des Sciences Forestières, 159 -172 . DOI:
10.1051/forest:19980110 URL [本文引用: 1] [2] Baig MN Tranquillini W 1980 ). The effects of wind and temperature on cuticular transpiration of Picea abies and Pinus cembra and their significance in dessication damage at the alpine treelineOecologia, 252 -256 . DOI:
10.1007/BF00346828 URL [本文引用: 1] [3] Bouriaud O Leban JM Bert D Deleuze C 2005 ). Intra-annual variations in climate influence growth and wood density of Norway spruceTree Physiology, 651 -660 . DOI:
10.1093/treephys/25.6.651 URL [本文引用: 1] [4] Chan T H?ltt? T Berninger F M?kinen H N?jd P Mencuccini M Nikinmaa E 2016 ). Separating water-potential induced swelling and shrinking from measured radial stem variations reveals a cambial growth and osmotic concentration signalPlant, Cell & Environment, 233 -244 . [本文引用: 1] [5] Dai JX Yang GL Yang ZY 2002 ). Pioneer tree species for sand control and afforestation—Afforestation techniques of Pinus sylvestris varmongolica. Forest Science and Technology, 10 ), 5 -7 . [本文引用: 1] [ 戴继先 , 杨国林 , 杨战阳 (2002 ). 治沙造林先锋树种——樟子松造林技术研究林业实用技术 , ( 10 ), 5 -7 .] [本文引用: 1] [6] Deslauriers A Anfodillo T Rossi S Carraro V 2007 a). Using simple causal modeling to understand how water and temperature affect daily stem radial variation in treesTree Physiology, 1125 -1136 . DOI:
10.1093/treephys/27.8.1125 URL [本文引用: 2] [7] Deslauriers A Morin H Urbinati C 2003 ). Daily weather response of balsam fir ( Abies balsamea ( L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada)Trees, 477 -484 . DOI:
10.1007/s00468-003-0260-4 URL [本文引用: 4] [8] Deslauriers A Rossi S Anfodillo T 2007 b). Dendrometer and intra-annual tree growth: What kind of information can be inferred?Dendrochronologia, 113 -124 . DOI:
10.1016/j.dendro.2007.05.003 URL [本文引用: 2] [9] Ehrenberger W Rüger S Fitzke R Vollenweider P Günthardt- Goerg MS Kuster T Zimmermann U Arend M 2012 ). Concomitant dendrometer and leaf patch pressure probe measurements reveal the effect of microclimate and soil moisture on diurnal stem water and leaf turgor variations in young oak treesFunctional Plant Biology, 297 -305 . DOI:
10.1071/FP11206 URL [本文引用: 1] [10] Gruber A Baumgartner D Zimmermann J Oberhuber W 2009 ). Temporal dynamic of wood formation in Pinus cembra along the alpine treeline ecotone and the effect of climate variablesTrees, 623 -635 . DOI:
10.1007/s00468-008-0307-7 URL [本文引用: 1] [11] King G Fonti P Nievergelt DU Frank D 2013 ). Climatic drivers of hourly to yearly tree radius variations along a 6 °C natural warming gradientAgricultural and Forest Meteorology, 36 -46 . DOI:
10.1016/j.agrformet.2012.08.002 URL [本文引用: 4] [12] Kocher P Horna V Leuschner C 2012 ). Environmental control of daily stem growth patterns in five temperate broad-leaved tree speciesTree Physiology, 1021 -1032 . DOI:
10.1093/treephys/tps049 URL [本文引用: 1] [13] Li CY Junttila O Ernstsen A Heino P Palva ET 2003 ). Photoperiodic control of growth, cold acclimation and dormancy development in silver birch (Betula pendula ) ecotypes. Physiologia Plantarum, 206 -212 . [本文引用: 1] [14] Li T He XY Chen ZJ 2014 ). Tree-ring growth responses of Mongolian oak ( Quercus mongolica ) to climate change in southern Northeast: a case study in Qianshan MountainsChinese Journal of Applied Ecology, 1841 -1848 . [本文引用: 1] [ 李腾 , 何兴元 , 陈振举 (2014 ). 东北南部蒙古栎径向生长对气候变化的响应——以千山为例应用生态学报 , 25, 1841 -1848 .] [本文引用: 1] [15] Li XH Liu RP Mao ZJ Song Y Liu LX Sun T 2014 ). Daily stem radial variation of Pinus koraiensis and its response to meteorological parameters in Xiaoxing’an MountainActa Ecologica Sinica, 1635 -1644 . [本文引用: 3] [ 李兴欢 , 刘瑞鹏 , 毛子军 , 宋媛 , 刘林馨 , 孙涛 (2014 ). 小兴安岭红松日径向变化及其对气象因子的响应生态学报 , 34, 1635 -1644 .] [本文引用: 3] [16] Ljung K Bhalerao RP Sandberg G 2002 ). Sites and homeostatic control of auxin biosynthesis in Arabidopsis during vegetative growthThe Plant Journal, 465 -474 . DOI:
10.1046/j.1365-313X.2001.01173.x URL [本文引用: 1] [17] Marek J Miroslav B Katarína S ?ubica D 2011 ). The impact of the 2003-2008 weather variability on intra-annual stem diameter changes of beech trees at a submontane site in central slovakiaDendrochronologia, 227 -235 . DOI:
10.1016/j.dendro.2011.01.009 URL [本文引用: 1] [18] Niu HG Zhang F Yu AL Wang F Zhang JZ Gou XH 2018 ). Intra-annual stem radial growth dynamics of Picea wilsorii in response to climate in the eastern Qilian MountainsActa Ecologica Sinica, 7412 -7420 . [本文引用: 3] [ 牛豪阁 , 张芬 , 于爱灵 , 王放 , 张军周 , 勾晓华 (2018 ). 祁连山东部青杄年内径向生长动态对气候的响应生态学报 , 38, 7412 -7420 .] [本文引用: 3] [19] Partanen J Beuker E 1999 ). Effects of photoperiod and thermal time on the growth rhythm of Pinus sylvestris seedlingsScandinavian Journal of Forest Research, 487 -497 . DOI:
10.1080/02827589908540813 URL [本文引用: 1] [20] Pearce R 2001 ). Plant freezing and damageAnnals of Botany, 417 -424 . DOI:
10.1006/anbo.2000.1352 URL [本文引用: 1] [21] Pichler P Oberhuber W 2007 ). Radial growth response of coniferous forest trees in an Inner Alpine environment to heat- wave in 2003Forest Ecology and Management, 688 -699 . DOI:
10.1016/j.foreco.2007.02.007 URL [本文引用: 1] [22] Rossi S Deslauriers A Anfodillo T Morin H Saracino A Motta R Borghetti M 2006 ). Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day lengthNew Phytologist, 301 -310 . DOI:
10.1111/nph.2006.170.issue-2 URL [本文引用: 1] [23] Sevanto S Ho?ltta? T Holbrook NM 2011 ). Effects of the hydraulic coupling between xylem and phloem on diurnal phloem diameter variationPlant, Cell & Environment, 690 -703 . [本文引用: 1] [24] Sevanto S Suni T Pumpanen J Gronholm T Kolari P Nikinmaa E Hari P Vesala T 2006 ). Wintertime photosynthesis and water uptake in a boreal forestTree Physiology, 749 -757 . DOI:
10.1093/treephys/26.6.749 URL [本文引用: 2] [25] Strimbeck GR Kjellsen TD Schaberg PG MurakamiPF , (2008 ). Dynamics of low-temperature acclimation in temperate and boreal conifer foliage in a mild winter climateTree Physiology, 1365 -1374 . DOI:
10.1093/treephys/28.9.1365 URL [本文引用: 1] [26] Swidrak I Gruber A Kofler W Oberhuber W 2011 ). Effects of environmental conditions on onset of xylem growth in Pinus sylvestris under droughtTree Physiology, 483 -493 . DOI:
10.1093/treephys/tpr034 URL [本文引用: 1] [27] Turcotte A Morin H Krause C Deslauriers A Thibeault-Martel M 2009 ). The timing of spring rehydration and its relation with the onset of wood formation in black spruceAgricultural and Forest Meteorology, 1403 -1409 . DOI:
10.1016/j.agrformet.2009.03.010 URL [本文引用: 2] [28] Turcotte A Rossi S Deslauriers A Krause C Morin H 2011 ). Dynamics of depletion and replenishment of water storage in stem and roots of black spruce measured by dendrometersFrontiers in Plant Science, 1 -8 . [本文引用: 1] [29] Wang XC Song LP Zhang YD 2011 ). Climate-tree growth relationships of Pinus sylvestris var. mongolica in the northern Daxing’an Mountains, ChinaChinese Journal of Plant Ecology, 294 -302 . DOI:
10.3724/SP.J.1258.2011.00294 URL [本文引用: 1] [ 王晓春 , 宋来萍 , 张远东 (2011 ). 大兴安岭北部樟子松树木生长与气候因子的关系植物生态学报 , 35, 294 -302 .] DOI:
10.3724/SP.J.1258.2011.00294 URL [本文引用: 1] [30] Zhang JD Jiang LL Xu ZQ Ma R 2019 ). Study on growth regularity of Pinus sylvestris var. mongolica in Saihanba areaForestry and Ecological Sciences, 135 -140 . [本文引用: 2] [ 张建东 , 姜玲玲 , 许中旗 , 马荣 (2019 ). 塞罕坝地区樟子松生长规律研究林业与生态科学 , 34, 135 -140 .] [本文引用: 2] [31] Zhang LN Jiang Y Zhang SD Jiao L Wen Y 2018 ). Relationships between tree age and climate sensitivity of radial growth in different drought conditions of Qilian Mountains, northwestern ChinaForests, 135 -153 . DOI:
10.3390/f9030135 URL [本文引用: 1] [32] Zhang WT Guo YY Dong MY Jiang Y Yang HC Wang MC 2012 ). Dynamics of stem radial growth of Picea meyeri during the growing season at the treeline of Luya Mountain, ChinaChinese Journal of Plant Ecology, 956 -964 . DOI:
10.3724/SP.J.1258.2012.00956 URL [本文引用: 3] [ 张文涛 , 郭媛媛 , 董满宇 , 江源 , 杨浩春 , 王明昌 (2012 ). 芦芽山林线白杄生长季径向生长动态植物生态学报 , 36, 956 -964 .] DOI:
10.3724/SP.J.1258.2012.00956 URL [本文引用: 3] [33] Zhang Y Bergeron Y Gao LS Zhao XH Wang XM Drobyshev I 2014 ). Tree growth and regeneration dynamics at a mountain ecotone on Changbai Mountain, northeastern China: Which factors control species distributions?écoscience, 387 -404 . DOI:
10.2980/21-(3-4)-3702 URL [本文引用: 1] [34] Zheng CY 2005 ). Forest Structure and Biomass of Saihanba, Hebei Province Post-Doctorate Reports, Peking University, Beijing . [本文引用: 1] [ 郑成洋 (2005 ). 河北省塞罕坝森林结构与生物量博士后出站报告, 北京大学, 北京 .] [本文引用: 1] [35] Zhuang LW Axmacher JC Sang WG 2017 ). Different radial growth responses to climate warming by two dominant tree species at their upper altitudinal limit on Changbai MountainJournal of Forestry Research, 795 -804 . DOI:
10.1007/s11676-016-0364-5 URL [本文引用: 1] [36] Zweifel R H?sler R 2000 ). Frost-induced reversible shrinkage of bark of mature subalpine conifersAgricultural and Forest Meteorology, 213 -222 . DOI:
10.1016/S0168-1923(00)00135-0 URL [本文引用: 5] [37] Zweifel R Item H Hasler R 2001 ). Link between diurnal stem radius changes and tree water relationsTree Physiology, 869 -877 . DOI:
10.1093/treephys/21.12-13.869 URL [本文引用: 1] Tree water relations and climatic variations at the alpine timberline: seasonal changes of sap flux and xylem water potential in
Larix decidua Miller,
Picea abies ( L.) Karst. and
Pinus cembra L
1
1998
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
The effects of wind and temperature on cuticular transpiration of
Picea abies and
Pinus cembra and their significance in dessication damage at the alpine treeline
1
1980
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Intra-annual variations in climate influence growth and wood density of Norway spruce
1
2005
... 樟子松径向昼夜变化采用正弦函数(
King et al ., 2013 ), 在波形的临界点处找到局部最大值(
R max )和最小值(
R min ), 计算每日最大值(
T max )和最小值(
T min )的时间, 以及振幅(峰值幅度), 计算公式为
R =
R max -
R min , 若径向变化在同一天呈现单一的波动, 则没有度量值.密集图代表性选择春夏、秋冬交替阶段的日平均树木径向变化规律, 即3月底至5月初与9月底至11月初, 观察昼夜径向变化波形特征.樟子松径向生长期的确定方法: 采用15天的树木径向日变化来确定生长季的开始与结束时期; 使用单样本
t 检验评估15天树干径向变化均值与0是否有差异, 差异显著且为正值时, 判断为树木生长(
Deslaurier et al ., 2003 ).樟子松径向日变化采用最大值法(
Bouriaud et al ., 2005 ), 从树木径向生长记录数据中, 提取樟子松每日48个生长数据中的最大值, 计算连续两日的最大值之差, 以作为树干径向日变化值(Δ
R ).日变化值参与气象因子的关系分析.本研究数据统计分析使用SPSS 17.0, 树木径向生长与气象因子的关系采用Pearson相关分析方法; 作图使用Origin 2018c. ...
Separating water-potential induced swelling and shrinking from measured radial stem variations reveals a cambial growth and osmotic concentration signal
1
2016
... 樟子松日径向变化曲线与树木蒸腾作用和冬季温度存在相关性(
Zweifel et al ., 2001 ;
Turcotte et al ., 2011 ;
Ehrenberger et al ., 2012 ).
Chan等(2016) 利用胸径测量仪数据与气象数据, 模拟生长曲线并预测出径向日变化与茎水势的变化近似成线性比例; 本研究结果中, 樟子松径向日变化与水分有密切关系.春夏季节, 径向昼夜变化是由于水势梯度与蒸腾过程强烈耦合作用(
Zweifel & H?sler, 2000 ).春季土壤温度高于0 ℃, 夜晚根部吸水充分, 早上气温升高, 树木开始冠层蒸腾作用, 茎干组织产生水势梯度, 水分从根部输送至茎干组织, 水分的吸收大于树冠蒸腾的损失, 此时树干半径增加; 水势相同时开始, 直到傍晚期间, 树冠的水分需求超过了根部的吸水量, 导致树干木材水势为负, 引发水分从树皮弹性组织向木质部转移, 树木茎干收缩.从傍晚开始, 蒸腾作用减小, 茎干水分得到补充(
Sevanto et al ., 2011 ), 树木茎干膨胀. ...
治沙造林先锋树种——樟子松造林技术研究
1
2002
... 樟子松具有耐寒、耐旱、耐贫瘠的优良特性.1963年塞罕坝地区开始营造人工林, 树种选择以落叶松为主, 但一些较为贫瘠、干旱地块落叶松不易成活.在20世纪60年代末才开始引种樟子松.现今塞罕坝林场樟子松面积约为7 500 hm
2 , 占塞罕坝总经营面积的8.0%左右(
戴继先等, 2002 ).人工樟子松林林下灌木、草本稀疏, 林下灌木物种主要有: 稠李(
Padus racemosa )、兴安胡枝子(
Lespedeza daurica )、山刺玫(
Rosa davurica )等; 草本以大披针薹草(
Carex lanceolata )、腺毛委陵菜(
Potentilla longifolia )、地榆(
Sanguisorba officinalis )、瓣蕊唐松草(
Thalictrum petaloideum )等为主. ...
治沙造林先锋树种——樟子松造林技术研究
1
2002
... 樟子松具有耐寒、耐旱、耐贫瘠的优良特性.1963年塞罕坝地区开始营造人工林, 树种选择以落叶松为主, 但一些较为贫瘠、干旱地块落叶松不易成活.在20世纪60年代末才开始引种樟子松.现今塞罕坝林场樟子松面积约为7 500 hm
2 , 占塞罕坝总经营面积的8.0%左右(
戴继先等, 2002 ).人工樟子松林林下灌木、草本稀疏, 林下灌木物种主要有: 稠李(
Padus racemosa )、兴安胡枝子(
Lespedeza daurica )、山刺玫(
Rosa davurica )等; 草本以大披针薹草(
Carex lanceolata )、腺毛委陵菜(
Potentilla longifolia )、地榆(
Sanguisorba officinalis )、瓣蕊唐松草(
Thalictrum petaloideum )等为主. ...
Using simple causal modeling to understand how water and temperature affect daily stem radial variation in trees
2
2007
... 秋冬季节, 蒸腾作用不再是树木茎干变化的驱动因素, 气温成为树干茎干变化的决定因素(
Sevanto et al ., 2006 ;
King et al ., 2013 ).由于冬季初期温差较大, 树木茎干在冰冻状态与解冻状态之间切换(
Deslauriers et al ., 2003 ), 当温度到达树液凝固点时, 木材细胞开始冻结, 从而导致树皮细胞液的渗透性排出, 茎干收缩(
Zweifel & H?sler, 2000 ;
Pearce, 2001 ;
Strimbeck et al ., 2008 ).当白天温度足够高时, 上述过程发生逆转, 水分开始流回细胞, 导致茎膨胀并达到每日最大值(
Turcotte et al ., 2009 ).本研究得出的昼夜变化规律与
King等(2013) 对阿尔卑斯山落叶松的研究结果基本一致, 但不同的是, 本研究冬季振幅大于夏季, 原因可能是振幅与持续时间的长短主要取决于温度与水分的转移速度(
Deslauriers et al ., 2007a ).塞罕坝每年2-3月气温升高, 昼夜温差变大, 导致振幅增大. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Daily weather response of balsam fir (
Abies balsamea ( L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada)
4
2003
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
... 樟子松径向昼夜变化采用正弦函数(
King et al ., 2013 ), 在波形的临界点处找到局部最大值(
R max )和最小值(
R min ), 计算每日最大值(
T max )和最小值(
T min )的时间, 以及振幅(峰值幅度), 计算公式为
R =
R max -
R min , 若径向变化在同一天呈现单一的波动, 则没有度量值.密集图代表性选择春夏、秋冬交替阶段的日平均树木径向变化规律, 即3月底至5月初与9月底至11月初, 观察昼夜径向变化波形特征.樟子松径向生长期的确定方法: 采用15天的树木径向日变化来确定生长季的开始与结束时期; 使用单样本
t 检验评估15天树干径向变化均值与0是否有差异, 差异显著且为正值时, 判断为树木生长(
Deslaurier et al ., 2003 ).樟子松径向日变化采用最大值法(
Bouriaud et al ., 2005 ), 从树木径向生长记录数据中, 提取樟子松每日48个生长数据中的最大值, 计算连续两日的最大值之差, 以作为树干径向日变化值(Δ
R ).日变化值参与气象因子的关系分析.本研究数据统计分析使用SPSS 17.0, 树木径向生长与气象因子的关系采用Pearson相关分析方法; 作图使用Origin 2018c. ...
... 秋冬季节, 蒸腾作用不再是树木茎干变化的驱动因素, 气温成为树干茎干变化的决定因素(
Sevanto et al ., 2006 ;
King et al ., 2013 ).由于冬季初期温差较大, 树木茎干在冰冻状态与解冻状态之间切换(
Deslauriers et al ., 2003 ), 当温度到达树液凝固点时, 木材细胞开始冻结, 从而导致树皮细胞液的渗透性排出, 茎干收缩(
Zweifel & H?sler, 2000 ;
Pearce, 2001 ;
Strimbeck et al ., 2008 ).当白天温度足够高时, 上述过程发生逆转, 水分开始流回细胞, 导致茎膨胀并达到每日最大值(
Turcotte et al ., 2009 ).本研究得出的昼夜变化规律与
King等(2013) 对阿尔卑斯山落叶松的研究结果基本一致, 但不同的是, 本研究冬季振幅大于夏季, 原因可能是振幅与持续时间的长短主要取决于温度与水分的转移速度(
Deslauriers et al ., 2007a ).塞罕坝每年2-3月气温升高, 昼夜温差变大, 导致振幅增大. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Dendrometer and intra-annual tree growth: What kind of information can be inferred?
2
2007
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Concomitant dendrometer and leaf patch pressure probe measurements reveal the effect of microclimate and soil moisture on diurnal stem water and leaf turgor variations in young oak trees
1
2012
... 樟子松日径向变化曲线与树木蒸腾作用和冬季温度存在相关性(
Zweifel et al ., 2001 ;
Turcotte et al ., 2011 ;
Ehrenberger et al ., 2012 ).
Chan等(2016) 利用胸径测量仪数据与气象数据, 模拟生长曲线并预测出径向日变化与茎水势的变化近似成线性比例; 本研究结果中, 樟子松径向日变化与水分有密切关系.春夏季节, 径向昼夜变化是由于水势梯度与蒸腾过程强烈耦合作用(
Zweifel & H?sler, 2000 ).春季土壤温度高于0 ℃, 夜晚根部吸水充分, 早上气温升高, 树木开始冠层蒸腾作用, 茎干组织产生水势梯度, 水分从根部输送至茎干组织, 水分的吸收大于树冠蒸腾的损失, 此时树干半径增加; 水势相同时开始, 直到傍晚期间, 树冠的水分需求超过了根部的吸水量, 导致树干木材水势为负, 引发水分从树皮弹性组织向木质部转移, 树木茎干收缩.从傍晚开始, 蒸腾作用减小, 茎干水分得到补充(
Sevanto et al ., 2011 ), 树木茎干膨胀. ...
Temporal dynamic of wood formation in
Pinus cembra along the alpine treeline ecotone and the effect of climate variables
1
2009
... 樟子松树干径向变化主要受水分与温度的影响, 但在年际间不同的生长阶段, 气象因子如何影响树干径向生长的研究报道较少.河北省塞罕坝地区进入4月后, 林内积雪开始融化, 土壤含水量增加, 树干径向生长是由于空气温度升高所致, 茎干在没有达到水分平衡的情况下, 达到树木生长有效积温, 引发形成层活动与细胞分化, 因而引发茎干生长(
Gruber et al ., 2009 ;
Swidrak et al ., 2011 ).空气温度引发空气湿度的改变, 空气湿度影响树冠蒸腾, 从而改变樟子松茎干组织的水分和形成层细胞膨压.生长稳定时期主要发生在每年的7至10月, 树木完成前期新生细胞的分裂和生长, 进入新生细胞木质化阶段, 树木径向大小基本保持稳定.在生长稳定、茎干变化不明显的情况下, 树木径向日变化量主要由茎干水分输送改变, 而降水量和空气温度共同影响水分的输送.研究发现, 15天为时间尺度时, 降水不再是樟子松径向生长的显著影响因素, 温度是径向生长的显著影响因素.
王晓春等(2011) 对大兴安岭北部樟子松树轮与气候响应的研究显示, 虽然温度对樟子松径向生长的影响在不同样点的月份存在差异, 但温度是樟子松径向生长的主要限制因子.但是
牛豪阁等(2018) 对祁连山青扦2013-2015年的树轮与气象关系的研究发现, 降水对在5-7月对青扦的影响是稳定显著的.这可能与樟子松的生理特性与形成层细胞结构有关, 后续需要对树木解剖学与形成层细胞生长进行研究. ...
Climatic drivers of hourly to yearly tree radius variations along a 6 °C natural warming gradient
4
2013
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
... 樟子松径向昼夜变化采用正弦函数(
King et al ., 2013 ), 在波形的临界点处找到局部最大值(
R max )和最小值(
R min ), 计算每日最大值(
T max )和最小值(
T min )的时间, 以及振幅(峰值幅度), 计算公式为
R =
R max -
R min , 若径向变化在同一天呈现单一的波动, 则没有度量值.密集图代表性选择春夏、秋冬交替阶段的日平均树木径向变化规律, 即3月底至5月初与9月底至11月初, 观察昼夜径向变化波形特征.樟子松径向生长期的确定方法: 采用15天的树木径向日变化来确定生长季的开始与结束时期; 使用单样本
t 检验评估15天树干径向变化均值与0是否有差异, 差异显著且为正值时, 判断为树木生长(
Deslaurier et al ., 2003 ).樟子松径向日变化采用最大值法(
Bouriaud et al ., 2005 ), 从树木径向生长记录数据中, 提取樟子松每日48个生长数据中的最大值, 计算连续两日的最大值之差, 以作为树干径向日变化值(Δ
R ).日变化值参与气象因子的关系分析.本研究数据统计分析使用SPSS 17.0, 树木径向生长与气象因子的关系采用Pearson相关分析方法; 作图使用Origin 2018c. ...
... 秋冬季节, 蒸腾作用不再是树木茎干变化的驱动因素, 气温成为树干茎干变化的决定因素(
Sevanto et al ., 2006 ;
King et al ., 2013 ).由于冬季初期温差较大, 树木茎干在冰冻状态与解冻状态之间切换(
Deslauriers et al ., 2003 ), 当温度到达树液凝固点时, 木材细胞开始冻结, 从而导致树皮细胞液的渗透性排出, 茎干收缩(
Zweifel & H?sler, 2000 ;
Pearce, 2001 ;
Strimbeck et al ., 2008 ).当白天温度足够高时, 上述过程发生逆转, 水分开始流回细胞, 导致茎膨胀并达到每日最大值(
Turcotte et al ., 2009 ).本研究得出的昼夜变化规律与
King等(2013) 对阿尔卑斯山落叶松的研究结果基本一致, 但不同的是, 本研究冬季振幅大于夏季, 原因可能是振幅与持续时间的长短主要取决于温度与水分的转移速度(
Deslauriers et al ., 2007a ).塞罕坝每年2-3月气温升高, 昼夜温差变大, 导致振幅增大. ...
... ).本研究得出的昼夜变化规律与
King等(2013) 对阿尔卑斯山落叶松的研究结果基本一致, 但不同的是, 本研究冬季振幅大于夏季, 原因可能是振幅与持续时间的长短主要取决于温度与水分的转移速度(
Deslauriers et al ., 2007a ).塞罕坝每年2-3月气温升高, 昼夜温差变大, 导致振幅增大. ...
Environmental control of daily stem growth patterns in five temperate broad-leaved tree species
1
2012
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
Photoperiodic control of growth, cold acclimation and dormancy development in silver birch (
1
2003
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
东北南部蒙古栎径向生长对气候变化的响应——以千山为例
1
2014
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
东北南部蒙古栎径向生长对气候变化的响应——以千山为例
1
2014
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
小兴安岭红松日径向变化及其对气象因子的响应
3
2014
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
... ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
小兴安岭红松日径向变化及其对气象因子的响应
3
2014
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
... ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Sites and homeostatic control of auxin biosynthesis in
Arabidopsis during vegetative growth
1
2002
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
The impact of the 2003-2008 weather variability on intra-annual stem diameter changes of beech trees at a submontane site in central slovakia
1
2011
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
祁连山东部青杄年内径向生长动态对气候的响应
3
2018
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
... 樟子松树干径向变化主要受水分与温度的影响, 但在年际间不同的生长阶段, 气象因子如何影响树干径向生长的研究报道较少.河北省塞罕坝地区进入4月后, 林内积雪开始融化, 土壤含水量增加, 树干径向生长是由于空气温度升高所致, 茎干在没有达到水分平衡的情况下, 达到树木生长有效积温, 引发形成层活动与细胞分化, 因而引发茎干生长(
Gruber et al ., 2009 ;
Swidrak et al ., 2011 ).空气温度引发空气湿度的改变, 空气湿度影响树冠蒸腾, 从而改变樟子松茎干组织的水分和形成层细胞膨压.生长稳定时期主要发生在每年的7至10月, 树木完成前期新生细胞的分裂和生长, 进入新生细胞木质化阶段, 树木径向大小基本保持稳定.在生长稳定、茎干变化不明显的情况下, 树木径向日变化量主要由茎干水分输送改变, 而降水量和空气温度共同影响水分的输送.研究发现, 15天为时间尺度时, 降水不再是樟子松径向生长的显著影响因素, 温度是径向生长的显著影响因素.
王晓春等(2011) 对大兴安岭北部樟子松树轮与气候响应的研究显示, 虽然温度对樟子松径向生长的影响在不同样点的月份存在差异, 但温度是樟子松径向生长的主要限制因子.但是
牛豪阁等(2018) 对祁连山青扦2013-2015年的树轮与气象关系的研究发现, 降水对在5-7月对青扦的影响是稳定显著的.这可能与樟子松的生理特性与形成层细胞结构有关, 后续需要对树木解剖学与形成层细胞生长进行研究. ...
祁连山东部青杄年内径向生长动态对气候的响应
3
2018
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
... 樟子松树干径向变化主要受水分与温度的影响, 但在年际间不同的生长阶段, 气象因子如何影响树干径向生长的研究报道较少.河北省塞罕坝地区进入4月后, 林内积雪开始融化, 土壤含水量增加, 树干径向生长是由于空气温度升高所致, 茎干在没有达到水分平衡的情况下, 达到树木生长有效积温, 引发形成层活动与细胞分化, 因而引发茎干生长(
Gruber et al ., 2009 ;
Swidrak et al ., 2011 ).空气温度引发空气湿度的改变, 空气湿度影响树冠蒸腾, 从而改变樟子松茎干组织的水分和形成层细胞膨压.生长稳定时期主要发生在每年的7至10月, 树木完成前期新生细胞的分裂和生长, 进入新生细胞木质化阶段, 树木径向大小基本保持稳定.在生长稳定、茎干变化不明显的情况下, 树木径向日变化量主要由茎干水分输送改变, 而降水量和空气温度共同影响水分的输送.研究发现, 15天为时间尺度时, 降水不再是樟子松径向生长的显著影响因素, 温度是径向生长的显著影响因素.
王晓春等(2011) 对大兴安岭北部樟子松树轮与气候响应的研究显示, 虽然温度对樟子松径向生长的影响在不同样点的月份存在差异, 但温度是樟子松径向生长的主要限制因子.但是
牛豪阁等(2018) 对祁连山青扦2013-2015年的树轮与气象关系的研究发现, 降水对在5-7月对青扦的影响是稳定显著的.这可能与樟子松的生理特性与形成层细胞结构有关, 后续需要对树木解剖学与形成层细胞生长进行研究. ...
Effects of photoperiod and thermal time on the growth rhythm of
Pinus sylvestris seedlings
1
1999
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Plant freezing and damage
1
2001
... 秋冬季节, 蒸腾作用不再是树木茎干变化的驱动因素, 气温成为树干茎干变化的决定因素(
Sevanto et al ., 2006 ;
King et al ., 2013 ).由于冬季初期温差较大, 树木茎干在冰冻状态与解冻状态之间切换(
Deslauriers et al ., 2003 ), 当温度到达树液凝固点时, 木材细胞开始冻结, 从而导致树皮细胞液的渗透性排出, 茎干收缩(
Zweifel & H?sler, 2000 ;
Pearce, 2001 ;
Strimbeck et al ., 2008 ).当白天温度足够高时, 上述过程发生逆转, 水分开始流回细胞, 导致茎膨胀并达到每日最大值(
Turcotte et al ., 2009 ).本研究得出的昼夜变化规律与
King等(2013) 对阿尔卑斯山落叶松的研究结果基本一致, 但不同的是, 本研究冬季振幅大于夏季, 原因可能是振幅与持续时间的长短主要取决于温度与水分的转移速度(
Deslauriers et al ., 2007a ).塞罕坝每年2-3月气温升高, 昼夜温差变大, 导致振幅增大. ...
Radial growth response of coniferous forest trees in an Inner Alpine environment to heat- wave in 2003
1
2007
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length
1
2006
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Effects of the hydraulic coupling between xylem and phloem on diurnal phloem diameter variation
1
2011
... 樟子松日径向变化曲线与树木蒸腾作用和冬季温度存在相关性(
Zweifel et al ., 2001 ;
Turcotte et al ., 2011 ;
Ehrenberger et al ., 2012 ).
Chan等(2016) 利用胸径测量仪数据与气象数据, 模拟生长曲线并预测出径向日变化与茎水势的变化近似成线性比例; 本研究结果中, 樟子松径向日变化与水分有密切关系.春夏季节, 径向昼夜变化是由于水势梯度与蒸腾过程强烈耦合作用(
Zweifel & H?sler, 2000 ).春季土壤温度高于0 ℃, 夜晚根部吸水充分, 早上气温升高, 树木开始冠层蒸腾作用, 茎干组织产生水势梯度, 水分从根部输送至茎干组织, 水分的吸收大于树冠蒸腾的损失, 此时树干半径增加; 水势相同时开始, 直到傍晚期间, 树冠的水分需求超过了根部的吸水量, 导致树干木材水势为负, 引发水分从树皮弹性组织向木质部转移, 树木茎干收缩.从傍晚开始, 蒸腾作用减小, 茎干水分得到补充(
Sevanto et al ., 2011 ), 树木茎干膨胀. ...
Wintertime photosynthesis and water uptake in a boreal forest
2
2006
... 秋冬季节, 蒸腾作用不再是树木茎干变化的驱动因素, 气温成为树干茎干变化的决定因素(
Sevanto et al ., 2006 ;
King et al ., 2013 ).由于冬季初期温差较大, 树木茎干在冰冻状态与解冻状态之间切换(
Deslauriers et al ., 2003 ), 当温度到达树液凝固点时, 木材细胞开始冻结, 从而导致树皮细胞液的渗透性排出, 茎干收缩(
Zweifel & H?sler, 2000 ;
Pearce, 2001 ;
Strimbeck et al ., 2008 ).当白天温度足够高时, 上述过程发生逆转, 水分开始流回细胞, 导致茎膨胀并达到每日最大值(
Turcotte et al ., 2009 ).本研究得出的昼夜变化规律与
King等(2013) 对阿尔卑斯山落叶松的研究结果基本一致, 但不同的是, 本研究冬季振幅大于夏季, 原因可能是振幅与持续时间的长短主要取决于温度与水分的转移速度(
Deslauriers et al ., 2007a ).塞罕坝每年2-3月气温升高, 昼夜温差变大, 导致振幅增大. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Dynamics of low-temperature acclimation in temperate and boreal conifer foliage in a mild winter climate
1
2008
... 秋冬季节, 蒸腾作用不再是树木茎干变化的驱动因素, 气温成为树干茎干变化的决定因素(
Sevanto et al ., 2006 ;
King et al ., 2013 ).由于冬季初期温差较大, 树木茎干在冰冻状态与解冻状态之间切换(
Deslauriers et al ., 2003 ), 当温度到达树液凝固点时, 木材细胞开始冻结, 从而导致树皮细胞液的渗透性排出, 茎干收缩(
Zweifel & H?sler, 2000 ;
Pearce, 2001 ;
Strimbeck et al ., 2008 ).当白天温度足够高时, 上述过程发生逆转, 水分开始流回细胞, 导致茎膨胀并达到每日最大值(
Turcotte et al ., 2009 ).本研究得出的昼夜变化规律与
King等(2013) 对阿尔卑斯山落叶松的研究结果基本一致, 但不同的是, 本研究冬季振幅大于夏季, 原因可能是振幅与持续时间的长短主要取决于温度与水分的转移速度(
Deslauriers et al ., 2007a ).塞罕坝每年2-3月气温升高, 昼夜温差变大, 导致振幅增大. ...
Effects of environmental conditions on onset of xylem growth in
Pinus sylvestris under drought
1
2011
... 樟子松树干径向变化主要受水分与温度的影响, 但在年际间不同的生长阶段, 气象因子如何影响树干径向生长的研究报道较少.河北省塞罕坝地区进入4月后, 林内积雪开始融化, 土壤含水量增加, 树干径向生长是由于空气温度升高所致, 茎干在没有达到水分平衡的情况下, 达到树木生长有效积温, 引发形成层活动与细胞分化, 因而引发茎干生长(
Gruber et al ., 2009 ;
Swidrak et al ., 2011 ).空气温度引发空气湿度的改变, 空气湿度影响树冠蒸腾, 从而改变樟子松茎干组织的水分和形成层细胞膨压.生长稳定时期主要发生在每年的7至10月, 树木完成前期新生细胞的分裂和生长, 进入新生细胞木质化阶段, 树木径向大小基本保持稳定.在生长稳定、茎干变化不明显的情况下, 树木径向日变化量主要由茎干水分输送改变, 而降水量和空气温度共同影响水分的输送.研究发现, 15天为时间尺度时, 降水不再是樟子松径向生长的显著影响因素, 温度是径向生长的显著影响因素.
王晓春等(2011) 对大兴安岭北部樟子松树轮与气候响应的研究显示, 虽然温度对樟子松径向生长的影响在不同样点的月份存在差异, 但温度是樟子松径向生长的主要限制因子.但是
牛豪阁等(2018) 对祁连山青扦2013-2015年的树轮与气象关系的研究发现, 降水对在5-7月对青扦的影响是稳定显著的.这可能与樟子松的生理特性与形成层细胞结构有关, 后续需要对树木解剖学与形成层细胞生长进行研究. ...
The timing of spring rehydration and its relation with the onset of wood formation in black spruce
2
2009
... 秋冬季节, 蒸腾作用不再是树木茎干变化的驱动因素, 气温成为树干茎干变化的决定因素(
Sevanto et al ., 2006 ;
King et al ., 2013 ).由于冬季初期温差较大, 树木茎干在冰冻状态与解冻状态之间切换(
Deslauriers et al ., 2003 ), 当温度到达树液凝固点时, 木材细胞开始冻结, 从而导致树皮细胞液的渗透性排出, 茎干收缩(
Zweifel & H?sler, 2000 ;
Pearce, 2001 ;
Strimbeck et al ., 2008 ).当白天温度足够高时, 上述过程发生逆转, 水分开始流回细胞, 导致茎膨胀并达到每日最大值(
Turcotte et al ., 2009 ).本研究得出的昼夜变化规律与
King等(2013) 对阿尔卑斯山落叶松的研究结果基本一致, 但不同的是, 本研究冬季振幅大于夏季, 原因可能是振幅与持续时间的长短主要取决于温度与水分的转移速度(
Deslauriers et al ., 2007a ).塞罕坝每年2-3月气温升高, 昼夜温差变大, 导致振幅增大. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Dynamics of depletion and replenishment of water storage in stem and roots of black spruce measured by dendrometers
1
2011
... 樟子松日径向变化曲线与树木蒸腾作用和冬季温度存在相关性(
Zweifel et al ., 2001 ;
Turcotte et al ., 2011 ;
Ehrenberger et al ., 2012 ).
Chan等(2016) 利用胸径测量仪数据与气象数据, 模拟生长曲线并预测出径向日变化与茎水势的变化近似成线性比例; 本研究结果中, 樟子松径向日变化与水分有密切关系.春夏季节, 径向昼夜变化是由于水势梯度与蒸腾过程强烈耦合作用(
Zweifel & H?sler, 2000 ).春季土壤温度高于0 ℃, 夜晚根部吸水充分, 早上气温升高, 树木开始冠层蒸腾作用, 茎干组织产生水势梯度, 水分从根部输送至茎干组织, 水分的吸收大于树冠蒸腾的损失, 此时树干半径增加; 水势相同时开始, 直到傍晚期间, 树冠的水分需求超过了根部的吸水量, 导致树干木材水势为负, 引发水分从树皮弹性组织向木质部转移, 树木茎干收缩.从傍晚开始, 蒸腾作用减小, 茎干水分得到补充(
Sevanto et al ., 2011 ), 树木茎干膨胀. ...
大兴安岭北部樟子松树木生长与气候因子的关系
1
2011
... 樟子松树干径向变化主要受水分与温度的影响, 但在年际间不同的生长阶段, 气象因子如何影响树干径向生长的研究报道较少.河北省塞罕坝地区进入4月后, 林内积雪开始融化, 土壤含水量增加, 树干径向生长是由于空气温度升高所致, 茎干在没有达到水分平衡的情况下, 达到树木生长有效积温, 引发形成层活动与细胞分化, 因而引发茎干生长(
Gruber et al ., 2009 ;
Swidrak et al ., 2011 ).空气温度引发空气湿度的改变, 空气湿度影响树冠蒸腾, 从而改变樟子松茎干组织的水分和形成层细胞膨压.生长稳定时期主要发生在每年的7至10月, 树木完成前期新生细胞的分裂和生长, 进入新生细胞木质化阶段, 树木径向大小基本保持稳定.在生长稳定、茎干变化不明显的情况下, 树木径向日变化量主要由茎干水分输送改变, 而降水量和空气温度共同影响水分的输送.研究发现, 15天为时间尺度时, 降水不再是樟子松径向生长的显著影响因素, 温度是径向生长的显著影响因素.
王晓春等(2011) 对大兴安岭北部樟子松树轮与气候响应的研究显示, 虽然温度对樟子松径向生长的影响在不同样点的月份存在差异, 但温度是樟子松径向生长的主要限制因子.但是
牛豪阁等(2018) 对祁连山青扦2013-2015年的树轮与气象关系的研究发现, 降水对在5-7月对青扦的影响是稳定显著的.这可能与樟子松的生理特性与形成层细胞结构有关, 后续需要对树木解剖学与形成层细胞生长进行研究. ...
大兴安岭北部樟子松树木生长与气候因子的关系
1
2011
... 樟子松树干径向变化主要受水分与温度的影响, 但在年际间不同的生长阶段, 气象因子如何影响树干径向生长的研究报道较少.河北省塞罕坝地区进入4月后, 林内积雪开始融化, 土壤含水量增加, 树干径向生长是由于空气温度升高所致, 茎干在没有达到水分平衡的情况下, 达到树木生长有效积温, 引发形成层活动与细胞分化, 因而引发茎干生长(
Gruber et al ., 2009 ;
Swidrak et al ., 2011 ).空气温度引发空气湿度的改变, 空气湿度影响树冠蒸腾, 从而改变樟子松茎干组织的水分和形成层细胞膨压.生长稳定时期主要发生在每年的7至10月, 树木完成前期新生细胞的分裂和生长, 进入新生细胞木质化阶段, 树木径向大小基本保持稳定.在生长稳定、茎干变化不明显的情况下, 树木径向日变化量主要由茎干水分输送改变, 而降水量和空气温度共同影响水分的输送.研究发现, 15天为时间尺度时, 降水不再是樟子松径向生长的显著影响因素, 温度是径向生长的显著影响因素.
王晓春等(2011) 对大兴安岭北部樟子松树轮与气候响应的研究显示, 虽然温度对樟子松径向生长的影响在不同样点的月份存在差异, 但温度是樟子松径向生长的主要限制因子.但是
牛豪阁等(2018) 对祁连山青扦2013-2015年的树轮与气象关系的研究发现, 降水对在5-7月对青扦的影响是稳定显著的.这可能与樟子松的生理特性与形成层细胞结构有关, 后续需要对树木解剖学与形成层细胞生长进行研究. ...
塞罕坝地区樟子松生长规律研究
2
2019
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
... 研究区域位于河北省围场满族蒙古族自治县北部塞罕坝机械林场总场千层板林场(42.41° N, 117.24° E, 海拔1 431 m), 处于内蒙古高原、大兴安岭山系与冀北山地交汇之处.属暖温带落叶阔叶林与温带草原的过渡带, 地处半干旱半湿润气候区, 冬季漫长且寒冷, 春秋季节较短, 夏季气候凉爽.研究区域昼夜温差较大, 年平均气温-1.4 ℃, 极端最高、最低气温分别为30.9和-42.8 ℃.年日照时间为2 368 h, 年无霜期60天.年降水量为438 mm, 其中6-8月占68% (
张建东等, 2019 ).塞罕坝及周边地区分布的土壤类型有灰色森林土、黑钙土、淡黑钙土、棕壤、暗栗钙土、亚高山草甸土和风沙土(
郑成洋, 2005 ), 但塞罕坝林场土壤类型以风沙土占绝大多数, 主要成土母质为风积物、残积物及冲积物. ...
塞罕坝地区樟子松生长规律研究
2
2019
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
... 研究区域位于河北省围场满族蒙古族自治县北部塞罕坝机械林场总场千层板林场(42.41° N, 117.24° E, 海拔1 431 m), 处于内蒙古高原、大兴安岭山系与冀北山地交汇之处.属暖温带落叶阔叶林与温带草原的过渡带, 地处半干旱半湿润气候区, 冬季漫长且寒冷, 春秋季节较短, 夏季气候凉爽.研究区域昼夜温差较大, 年平均气温-1.4 ℃, 极端最高、最低气温分别为30.9和-42.8 ℃.年日照时间为2 368 h, 年无霜期60天.年降水量为438 mm, 其中6-8月占68% (
张建东等, 2019 ).塞罕坝及周边地区分布的土壤类型有灰色森林土、黑钙土、淡黑钙土、棕壤、暗栗钙土、亚高山草甸土和风沙土(
郑成洋, 2005 ), 但塞罕坝林场土壤类型以风沙土占绝大多数, 主要成土母质为风积物、残积物及冲积物. ...
Relationships between tree age and climate sensitivity of radial growth in different drought conditions of Qilian Mountains, northwestern China
1
2018
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
芦芽山林线白杄生长季径向生长动态
3
2012
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
... ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
芦芽山林线白杄生长季径向生长动态
3
2012
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
... ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Tree growth and regeneration dynamics at a mountain ecotone on Changbai Mountain, northeastern China: Which factors control species distributions?
1
2014
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
河北省塞罕坝森林结构与生物量
1
2005
... 研究区域位于河北省围场满族蒙古族自治县北部塞罕坝机械林场总场千层板林场(42.41° N, 117.24° E, 海拔1 431 m), 处于内蒙古高原、大兴安岭山系与冀北山地交汇之处.属暖温带落叶阔叶林与温带草原的过渡带, 地处半干旱半湿润气候区, 冬季漫长且寒冷, 春秋季节较短, 夏季气候凉爽.研究区域昼夜温差较大, 年平均气温-1.4 ℃, 极端最高、最低气温分别为30.9和-42.8 ℃.年日照时间为2 368 h, 年无霜期60天.年降水量为438 mm, 其中6-8月占68% (
张建东等, 2019 ).塞罕坝及周边地区分布的土壤类型有灰色森林土、黑钙土、淡黑钙土、棕壤、暗栗钙土、亚高山草甸土和风沙土(
郑成洋, 2005 ), 但塞罕坝林场土壤类型以风沙土占绝大多数, 主要成土母质为风积物、残积物及冲积物. ...
河北省塞罕坝森林结构与生物量
1
2005
... 研究区域位于河北省围场满族蒙古族自治县北部塞罕坝机械林场总场千层板林场(42.41° N, 117.24° E, 海拔1 431 m), 处于内蒙古高原、大兴安岭山系与冀北山地交汇之处.属暖温带落叶阔叶林与温带草原的过渡带, 地处半干旱半湿润气候区, 冬季漫长且寒冷, 春秋季节较短, 夏季气候凉爽.研究区域昼夜温差较大, 年平均气温-1.4 ℃, 极端最高、最低气温分别为30.9和-42.8 ℃.年日照时间为2 368 h, 年无霜期60天.年降水量为438 mm, 其中6-8月占68% (
张建东等, 2019 ).塞罕坝及周边地区分布的土壤类型有灰色森林土、黑钙土、淡黑钙土、棕壤、暗栗钙土、亚高山草甸土和风沙土(
郑成洋, 2005 ), 但塞罕坝林场土壤类型以风沙土占绝大多数, 主要成土母质为风积物、残积物及冲积物. ...
Different radial growth responses to climate warming by two dominant tree species at their upper altitudinal limit on Changbai Mountain
1
2017
... 树木生长受制于当地的气候条件, 研究树木的径向生长动态规律及其与气象因子的关系, 有利于探讨森林生态系统对气候变化的响应(
李腾等, 2014 ).以往树木径向生长的研究主要基于树轮年代学(
Zhang et al ., 2014 ,
2018 ;
Zhuang et al ., 2017 ), 其研究的时间尺度较长, 缺少短期高频树木径向生长动态的研究.树木径向生长记录仪提供了短期高频记录树木径向生长的研究方法, 能够连续监测树木的径向变化, 记录树木径向生长的昼夜和季节变化动态.基于树木径向生长与相应气候动态监测, 在短时间尺度上研究生长规律的结果为长时间尺度树干径向生长的研究提供解释和验证(
张文涛等, 2012 ).近年来, 国内外采用树木径向生长记录仪的研究逐渐增多.
Deslauriers等(2007b) 对意大利阿尔卑斯山东部不同海拔高度欧洲云杉(
Picea abies )和欧洲落叶松(
Larix decidua )进行监测, 结果显示确定树木生长期时, 采用的时间尺度与海拔有密切关系, 低海拔(1 020 m)以10天为时间尺度, 高海拔(2 080 m)以15天为时间尺度.
King等(2013) 研究了瑞士中部阿尔卑斯山56株欧洲落叶松和欧洲云杉的生长, 结果表明: 在生长季欧洲落叶松树干直径在上午最大, 傍晚最小, 反映了水分吸收和散失导致的日变化.
Deslauriers等(2003) 利用径向生长仪检测到加拿大魁北克北部
Abies balsamea 的径向生长与降水量呈正相关关系.
Kocher等(2012) 通过径向生长监测的方法研究了梣属(
Fraxinus )、槭属(
Acer )、鹅耳枥属(
Carpinus )、椴树属(
Tilia )和水青冈属(
Fagus )共生的5种温带阔叶树种在不同时期径向生长的主要环境控制因子, 结果发现: 空气相对湿度是影响径向生长的主要因素, 降水量、温度和辐射为次要因素.在国内,
李兴欢等(2014) 监测了小兴安岭凉水国家级自然保护区原始红松(
Pinus koraiensis )径向生长, 发现该树种的径向生长于5月中旬开始, 7月末趋于结束;
牛豪阁等(2018) 探究了祁连山东部青扦(
Picea wilsonii )不同时间尺度径向生长量与气候因子的关系, 结果显示时间尺度延长可以削弱树干水分昼夜变化的干扰, 但水分仍然是青扦径向生长的限制因子.然而, 作为我国北方“三北”防护林等人工林体系的主要树种, 樟子松(
Pinus sylvestris var.
mongolica )径向生长研究的甚少.塞罕坝机械林场总经营面积9.507万hm
2 , 有林地面积7.2万hm
2 (
张建东等, 2019 ).樟子松人工林径向变化规律研究对种植大面积人工林的塞罕坝地区有生产实践指导意义.在全球气候变化背景下, 研究其生长动态规律及其对气候变化的响应机制, 对提高我国北方樟子松人工林生态系统生产力与稳定性, 合理指导樟子松人工林的经营管理也具有重要意义.本研究利用树木径向生长记录仪(DRL26, EMS, Brno, Czechic)进行监测, 具体目的是为了揭示: 1)樟子松胸径日变化动态特征; 2)樟子松胸径变化的季节动态; 3)揭示樟子松树干径向变化与气象因子的关系. ...
Frost-induced reversible shrinkage of bark of mature subalpine conifers
5
2000
... 樟子松日径向变化曲线与树木蒸腾作用和冬季温度存在相关性(
Zweifel et al ., 2001 ;
Turcotte et al ., 2011 ;
Ehrenberger et al ., 2012 ).
Chan等(2016) 利用胸径测量仪数据与气象数据, 模拟生长曲线并预测出径向日变化与茎水势的变化近似成线性比例; 本研究结果中, 樟子松径向日变化与水分有密切关系.春夏季节, 径向昼夜变化是由于水势梯度与蒸腾过程强烈耦合作用(
Zweifel & H?sler, 2000 ).春季土壤温度高于0 ℃, 夜晚根部吸水充分, 早上气温升高, 树木开始冠层蒸腾作用, 茎干组织产生水势梯度, 水分从根部输送至茎干组织, 水分的吸收大于树冠蒸腾的损失, 此时树干半径增加; 水势相同时开始, 直到傍晚期间, 树冠的水分需求超过了根部的吸水量, 导致树干木材水势为负, 引发水分从树皮弹性组织向木质部转移, 树木茎干收缩.从傍晚开始, 蒸腾作用减小, 茎干水分得到补充(
Sevanto et al ., 2011 ), 树木茎干膨胀. ...
... 秋冬季节, 蒸腾作用不再是树木茎干变化的驱动因素, 气温成为树干茎干变化的决定因素(
Sevanto et al ., 2006 ;
King et al ., 2013 ).由于冬季初期温差较大, 树木茎干在冰冻状态与解冻状态之间切换(
Deslauriers et al ., 2003 ), 当温度到达树液凝固点时, 木材细胞开始冻结, 从而导致树皮细胞液的渗透性排出, 茎干收缩(
Zweifel & H?sler, 2000 ;
Pearce, 2001 ;
Strimbeck et al ., 2008 ).当白天温度足够高时, 上述过程发生逆转, 水分开始流回细胞, 导致茎膨胀并达到每日最大值(
Turcotte et al ., 2009 ).本研究得出的昼夜变化规律与
King等(2013) 对阿尔卑斯山落叶松的研究结果基本一致, 但不同的是, 本研究冬季振幅大于夏季, 原因可能是振幅与持续时间的长短主要取决于温度与水分的转移速度(
Deslauriers et al ., 2007a ).塞罕坝每年2-3月气温升高, 昼夜温差变大, 导致振幅增大. ...
...
Deslauriers等(2007a) 指出, 树木径向变化包括不可逆的径向生长和可逆的活细胞脱水-再水化, 本研究的结果与此结论相一致.从全年的树木径向生长曲线观察, 春季降水快速补充水分, 造成3月中旬树干迅速增粗的生长假象(
Sevanto et al ., 2006 ;
Turcotte et al ., 2009 ), 这一膨胀现象容易与树木生长现象相互混淆(
Zweifel & H?sler, 2000 ;
Deslauriers et al ., 2003 ), 同时, 在红松(
李兴欢等, 2014 )、青扦(
牛豪阁等, 2018 )的相关研究中, 有相似的结果, 均在早春季节出现水分回升, 树干膨胀现象.随着气温升高, 积雪开始融化, 土壤含水量随之升高, 树干所需水分恢复并达到平衡, 但不满足生长所需的环境条件, 树木未发生显著生长, 树干径向趋于稳定不变, 此期间不再有大波动, 表现出一段短暂的平稳期.经过3月底至4月底的短暂平台期后, 樟子松于5月初开始生长, 此时土壤水分条件充足且土壤温度达到5 ℃以上, 茎干韧皮部组织产生形成层细胞, 树干开始生长, 树干径向增长持续至6月下旬, 从7月份开始形成层细胞产生缓慢(
Deslauriers et al ., 2007b ;
李兴欢等, 2014 ), 树干径向变化平稳.研究发现, 树木径向生长速度最大值发生在6月下旬, 是因为植物生长和生理过程与光周期因素有关(
Partanen & Beuker, 1999 ;
Li et al ., 2003 ;
Marek et al ., 2011 ), 生长前期光、温度和水分共同作用, 树木的新叶和茎快速生长, 也产生大量生长激素(
Ljung et al ., 2002 ), 促进树木茎干形成层大部分细胞的分裂和生长(
张文涛等, 2012 ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
... ).在生长季后期, 新生的形成层细胞逐步转化为薄壁细胞和厚壁细胞, 完成细胞壁木质化过程(
Zweifel & H?sler, 2000 ;
Rossi et al ., 2006 ).10下旬开始, 气温降至0 ℃以下, 降水量与土壤温度逐渐降低, 树木生长停止.树干脱水收缩, 各组织的细胞质浓度随之升高(
Baig & Tranquillini, 1980 ;
Anfodillo et al ., 1998 ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
... ;
Zweifel & H?sler, 2000 ;
张文涛等, 2012 ), 这是樟子松抵御寒冷与干旱的生存策略(
Pichler & Oberhuber, 2007 ). ...
Link between diurnal stem radius changes and tree water relations
1
2001
... 樟子松日径向变化曲线与树木蒸腾作用和冬季温度存在相关性(
Zweifel et al ., 2001 ;
Turcotte et al ., 2011 ;
Ehrenberger et al ., 2012 ).
Chan等(2016) 利用胸径测量仪数据与气象数据, 模拟生长曲线并预测出径向日变化与茎水势的变化近似成线性比例; 本研究结果中, 樟子松径向日变化与水分有密切关系.春夏季节, 径向昼夜变化是由于水势梯度与蒸腾过程强烈耦合作用(
Zweifel & H?sler, 2000 ).春季土壤温度高于0 ℃, 夜晚根部吸水充分, 早上气温升高, 树木开始冠层蒸腾作用, 茎干组织产生水势梯度, 水分从根部输送至茎干组织, 水分的吸收大于树冠蒸腾的损失, 此时树干半径增加; 水势相同时开始, 直到傍晚期间, 树冠的水分需求超过了根部的吸水量, 导致树干木材水势为负, 引发水分从树皮弹性组织向木质部转移, 树木茎干收缩.从傍晚开始, 蒸腾作用减小, 茎干水分得到补充(
Sevanto et al ., 2011 ), 树木茎干膨胀. ...
 ,1,2,3,*, 张磊4, 曾发旭1,3
,1,2,3,*, 张磊4, 曾发旭1,3
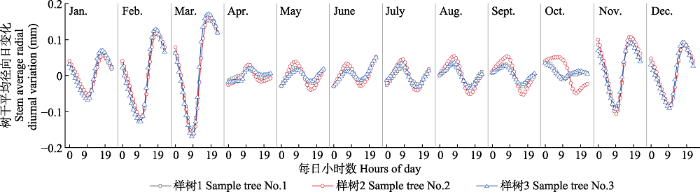 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT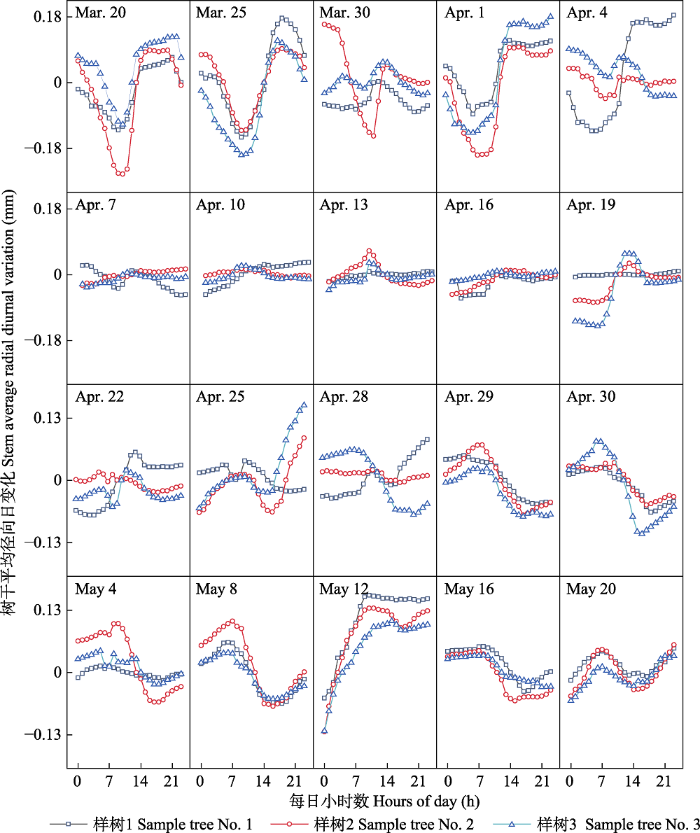 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT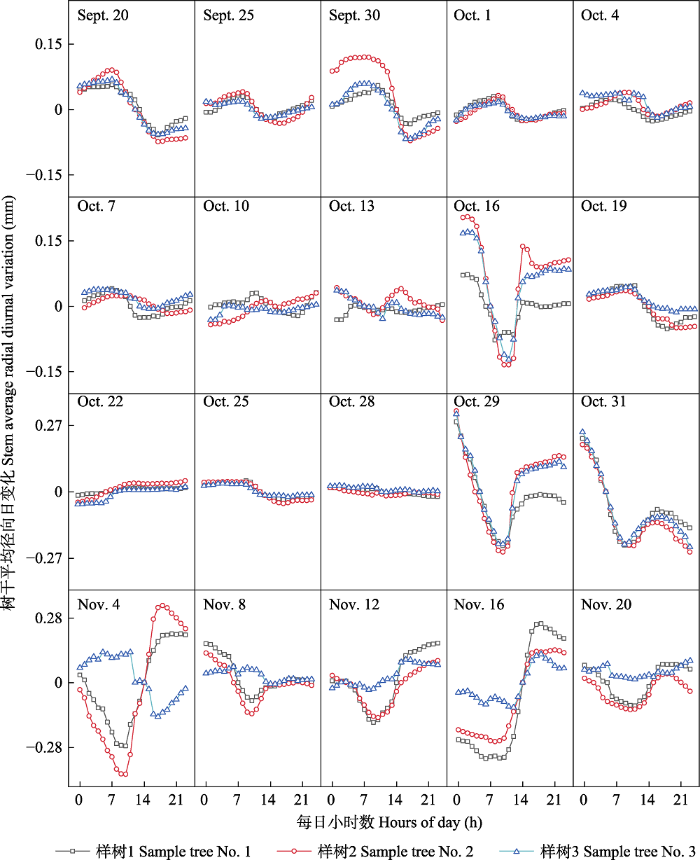 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT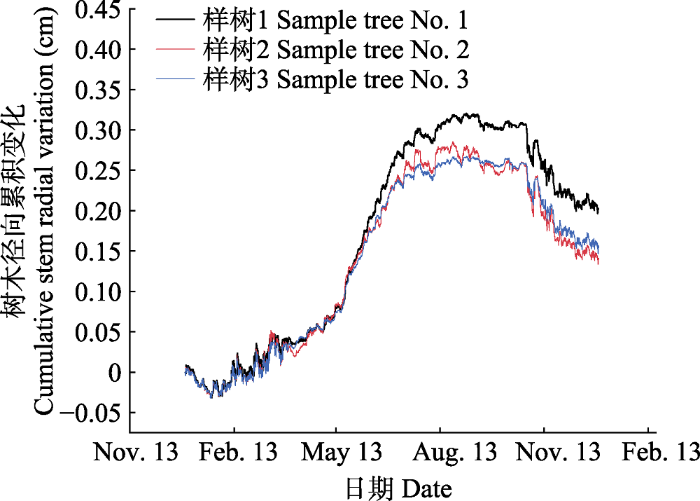 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT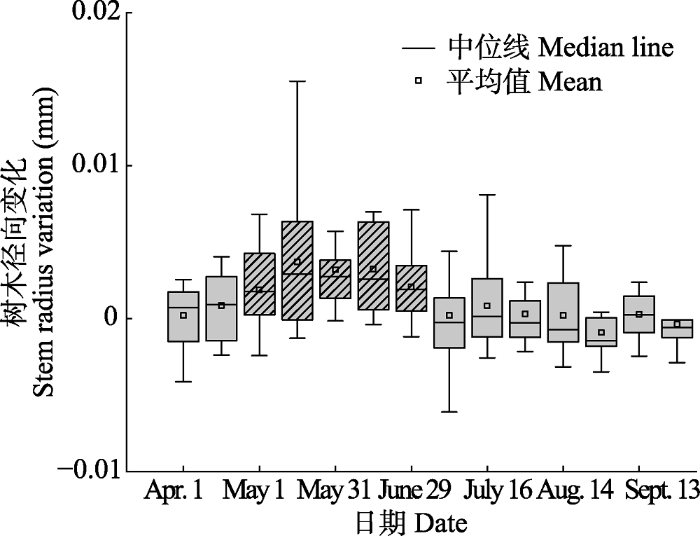 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT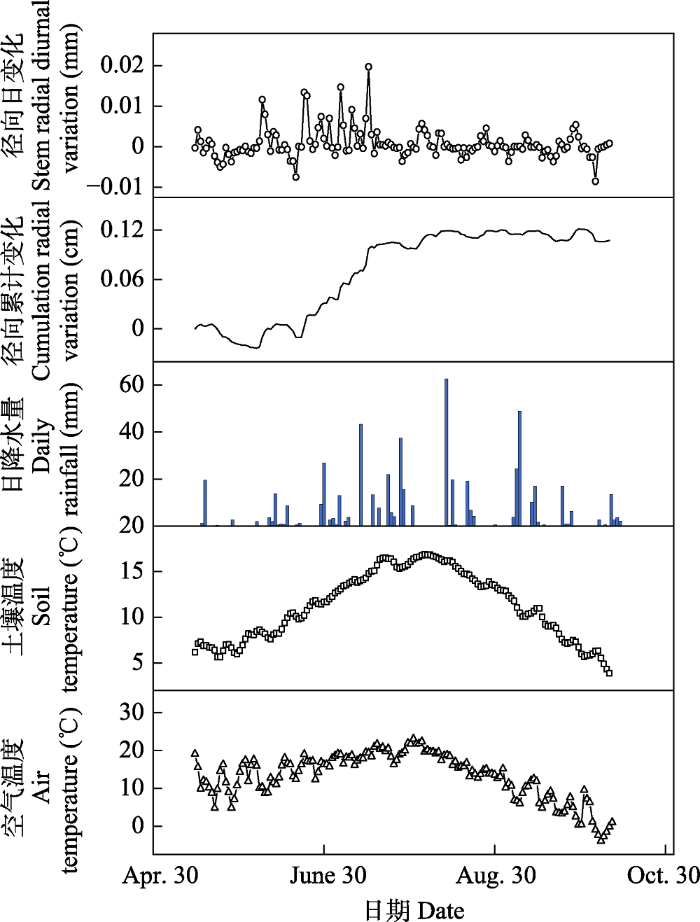 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}