Variations of leaf construction cost and leaf traits within the species of Artemisia ordosica along a precipitation gradient in the Mau Us sandy land
Zhi-Guo ZHANG1, Hai-Xia WEI,1,2,3,*1Department of Tourism and Resource Environment, Zaozhuang University, Zaozhuang, Shandong 277160, China 2Key Laboratory of Alpine Ecology and Biodiversity, Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing 100101, China 3Institute of Desertification Studies, Chinese Academy of Forestry, Beijing 100091, China
Abstract Aims There would be a trade-off between enhancing water use efficiency and decreasing leaf construction cost in arid plants. We hypothesized that plants in lower precipitation areas could have higher water use efficiency at a given leaf construction cost compared to plants in higher precipitation areas by altering relationships between leaf functional traits (i.e. increasing area-based leaf nitrogen concentration, Narea).Methods Artemisia ordosica is a widespread species in the Mau Us sandy land. Variations of leaf construction cost of A. ordosica along a precipitation gradient and its relationships to other leaf traits such as specific leaf area (SLA), mass-based leaf nitrogen concentration (Nmass), Narea and leaf stable carbon isotopes ratio (δ13C) value were analyzed.Important findings There were no significant differences in mass-based leaf construction cost (CCm) among different precipitation areas. Although area-based leaf construction cost (CCa) showed significant differences among different precipitation areas, they did not increase significantly with decreasing precipitation. The relationship between CCm and SLA was not significant while CCm positively correlated with leaf δ13C value. There was a strategy shift in the positive relationship of leaf construction cost to Narea between plant-groups in the lowest precipitation (264 mm) areas and other higher precipitation (310-370 mm) areas, i.e. plants from low precipitation areas have higher Narea at a given leaf construction cost. Our results indicated that although high water use efficiency was accompanied by high leaf construction cost, the high Narea for plants from lower precipitation areas did not increase their leaf construction cost. Keywords:Mau Us sandy land;Artemisia ordosica;rainfall gradient;leaf construction cost;strategy shift
PDF (1256KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张治国, 魏海霞. 毛乌素沙地油蒿叶建成成本及相关叶性状沿降水梯度的变化. 植物生态学报, 2019, 43(11): 979-987. DOI: 10.17521/cjpe.2019.0149 ZHANG Zhi-Guo, WEI Hai-Xia. Variations of leaf construction cost and leaf traits within the species of Artemisia ordosica along a precipitation gradient in the Mau Us sandy land. Chinese Journal of Plant Ecology, 2019, 43(11): 979-987. DOI: 10.17521/cjpe.2019.0149
在毛乌素沙地, 沿着油蒿自然分布区的降水梯度, 从东南到西北一共选择了4个气温差异不大, 但降水量明显不同的研究点。以GPS定位各研究点的经纬度和海拔(表1), 基于我国北方142个气象站点的气象数据(1983-2007年), 通过克里金插值法获得各个研究点的实际降水量和气温数据。4个研究点多年平均降水量分别为370、353、310和264 mm, 基本上覆盖了毛乌素沙地的降水范围, 反映了油蒿自然分布区降水的梯度变化(表1)。
Table 1 表1 表1毛乌素沙地研究点气候和土壤特征 Table 1Climatic and soil characteristics of study sites in the Mau Us sandy land
经度 Longitude
纬度 Latitude
海拔 Altitude (m)
年降水量 Annual precipitation (mm)
年蒸发量 Annual pan-evaporation (mm)
年平均气温 Annual mean temperature (℃)
土壤全氮含量 Soil total N concentration (mg·g-1)
固定沙地 Fixed sandy land
半固定沙地 Semi-fixed sandy land
榆林 Yulin
109.87°
38.62°
1 210
370
1 890
8.6
0.28 ± 0.049aBC
0.18 ± 0.075bA
乌审旗 Uxin Qi
108.64°
38.16°
1 270
353
2 322
8.7
0.24 ± 0.012aC
0.07 ± 0.016bB
鄂托克前旗 Otog Qianqi
108.46°
37.70°
1 320
310
2 133
8.8
0.32 ± 0.052aAB
0.19 ± 0.038bA
鄂托克旗 Otog Qi
108.06°
39.06°
1 420
264
2 450
7.6
0.37 ± 0.039aA
0.19 ± 0.018bA
Different uppercase letters indicate significant differences in soil total N concentration among study sites; different lowercase letters indicate significant differences in soil total N concentration between two sandy land habitats. 不同大写字母表示土壤全氮含量在不同研究点差异显著(p < 0.05); 不同小写字母表示土壤全氮含量在沙地生境间差异显著(p < 0.05)。
在每个研究点, 选择两类不同退化程度的沙地生境(固定沙地和半固定沙地)进行植物和土壤样品的采集。在本研究中, 对固定和半固定沙地生境的界定主要依据植被盖度(>40% vs. 20%-40%)和土壤结皮厚度(灌丛下和灌丛间结皮厚度>1 cm vs. 灌丛下结皮厚度0.5-1 cm)(Wu & Ci, 2002)。对于两类沙地生境, 分别设置3个5 m × 5 m的样方进行植物和土壤样品的采集, 共设置了24个采样样方。
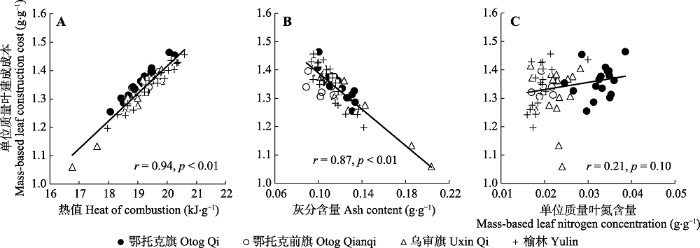
Fig. 1Relationships between mass-based leaf construction cost and heat of combustion (A), ash content (B) and mass-based leaf nitrogen concentration (C) of leaves within species of Artemisia ordosica along a precipitation gradient.
Fig. 2Comparisons of mass-based leaf construction cost (A) and area-based leaf construction cost (B) of Artemisia ordosica between two sandy land habitats and among different precipitation areas (mean + SD). Different uppercase letters indicate significant differences in leaf construction cost among different precipitation areas; different lowercase letters indicate significant differences in leaf construction cost between two sandy land habitats.
Table 2 表2 表2沿降水梯度油蒿叶片单位质量建成成本(CCm)和单位面积建成成本(CCa)与降水和土壤全氮含量的偏相关分析 Table 2Partial correlation coefficients for relationships of mass-based leaf construction cost (CCm) and area-based leaf construction cost (CCa) to annual precipitation and total soil nitrogen content within species of Artemisia ordosica along a precipitation gradient
Fig. 3Relationships of mass-based leaf construction cost and area-based leaf construction cost to specific leaf area (A, B), area-based leaf nitrogen concentration (C, D) and leaf stable carbon isotopes ratio (δ13C) value (E, F) within species of Artemisia ordosica along a precipitation gradient.
Fig. 4Relationships of mass-based leaf construction cost (A) and area-based leaf construction cost (B) to area-based leaf nitrogen concentration within species of Artemisia ordosica along a precipitation gradient in the sandy lands of the northern Qinghai-Xizang Plateau.
AmthorJS ( 1989). Respiration and Crop Productivity Springer-Verlgag, New York. [本文引用: 1]
AtkinOK, VillarR, CumminsWR ( 1993). The ability of several high arctic plant species to utilize nitrate nitrogen under field conditions Oecologia, 96, 239-245. [本文引用: 1]
CavattePC, Rodríguez-LópezNF, MartinsSCV, MattosMS, SanglardLMVP, DaMattaFM ( 2012). Functional analysis of the relative growth rate, chemical composition, construction and maintenance costs, and the payback time of Coffea arabica L. leaves in response to light and water availability Journal of Experimental Botany, 63, 3071-3082. [本文引用: 1]
ChenFY, LuoTX, ZhangL, DengKM, TianXY ( 2006). Comparison of leaf construction cost in dominant tree species of the evergreen broad-leaved forest in Jiulian Mountain, Jiangxi Province Acta Ecologica Sinica, 26, 2485-2493. [本文引用: 1]
DongZY, BaiXF, HouYP, BuQM ( 2015). Leaf calorific value of 8 tree species in the coastal areas of Jiaodong and cost of construction of leaf biomass and its adaptability Scientia Silvae Sinicae, 51(3), 8-15. [本文引用: 1]
Falc?oHM, MedeirosCD, Almeida-CortezJ, SantosMG ( 2017). Leaf construction cost is related to water availability in three species of different growth forms in a Brazilian tropical dry forest Theoretical and Experimental Plant Physiology, 29, 95-108. [本文引用: 3]
FortunelC, FinePVA, BaralotoC ( 2012). Leaf, stem and root tissue strategies across 758 neotropical tree species Functional Ecology, 26, 1153-1161. [本文引用: 1]
GriffinKL ( 1994). Calorimetric estimates of construction cost and their use in ecological studies Functional Ecology, 8, 551-562. [本文引用: 1]
GriffinKL, ThomasRB, StrainBR ( 1993). Effects of nitrogen supply and elevated carbon dioxide on construction cost in leaves of Pinus taeda ( L.) seedlings Oecologia, 95, 575-580. [本文引用: 1]
GriffinKL, WinnerWE, StrainBR ( 1996). Construction cost of loblolly and ponderosa pine leaves grown with varying carbon and nitrogen availability Plant, Cell & Environment, 19, 729-738. [本文引用: 2]
LiFL, YangL, ZanQJ, ShinPKS, CheungSG, WongYS, TamNoraFY, LeiAP ( 2017). Does energetic cost for leaf construction in Sonneratia change after introduce to another mangrove wetland and differ from native mangrove plants in South China? Marine Pollution Bulletin, 124, 1071-1077. [本文引用: 1]
LiYH, LuoTX, LuQ, TianXY, WuB, YangHH ( 2005). Comparisons of leaf traits among 17 major plant species in shazhuyu sand control experimental station of Qinghai Province Acta Ecologica Sinica, 25, 994-999. [本文引用: 1]
MartinezF, LazoYO, Fernandez-GalianoRM, MerinoJA ( 2002). Chemical composition and construction cost for roots of Mediterranean trees, shrub species and grassland communities Plant, Cell & Environment, 25, 601-608. [本文引用: 1]
MatíasL, QueroJL, ZamoraR, CastroJ ( 2012). Evidence for plant traits driving specific drought resistance. A community field experiment Environmental and Experimental Botany, 81, 55-61. [本文引用: 1]
NagelJM, GriffinKL ( 2001). Construction cost and invasive potential: Comparing Lythrum salicaria ( Lythraceae) with co-occurring native species along pond banks American Journal of Botany, 88, 2252-2258. [本文引用: 1]
NagelJM, WangXZ, LewisJD, FungHA, TissueDT, GriffinKL ( 2005). Atmospheric CO2 enrichment alters energy assimilation, investment and allocation in Xanthium strumarium New Phytologist, 166, 513-523. [本文引用: 1]
Penning de VriesFWT, BrunstingAHM, van LaarHH ( 1974). Products, requirements and efficiency of biosynthesis: A quantitative approach Journal of Theoretical Biology, 45, 339-377. [本文引用: 1]
PitelkaLF ( 1978). Variation in caloric values of annual and perennial lupines (Lupinus: Leguminosae) American Midland Naturalist, 99, 454-462. [本文引用: 1]
PoorterH ( 1994). Construction costs and payback time of biomass: A whole plant perspective In: Roy J, Gariner E eds. A Whole Plant Perspective on Carbon-Nitrogen Interactions. SPB Academic Publishing, Hague, The Netherlands. 11-127. [本文引用: 1]
PoorterH, de JongR ( 1999). A comparison of specific leaf area, chemical composition and leaf construction costs of field plants from 15 habitats differing in productivity New Phytologist, 143, 163-176. [本文引用: 2]
PoorterH, PepinS, RijkersT, de JongY, EvansJR, K?rnerC ( 2006). Construction costs, chemical composition and payback time of high- and low-irradiance leaves Journal of Experimental Botany, 57, 355-371. [本文引用: 1]
PoorterH, VillarR ( 1997). The fate of acquired carbon in plants: Chemical composition and construction costs In: Bazzaz FA, Grace J eds. Plant Resource Allocation. Elsevier, New York.39-72. [本文引用: 2]
PrenticeIC, DongN, GleasonSM, MaireV, WrightIJ ( 2014). Balancing the costs of carbon gain and water transport: Testing a new theoretical framework for plant functional ecology Ecology Letters, 17, 82-91. [本文引用: 1]
P'yankovVI, IvanovLA, LambersH ( 2001). Plant construction cost in the boreal species differing in their ecological strategies Russian Journal of Plant Physiology, 48, 67-73. [本文引用: 1]
RonzhinaDA, IvanovLA ( 2014). Construction costs and mesostructure of leaves in hydrophytes Russian Journal of Plant Physiology, 61, 776-783. [本文引用: 1]
ShanBQ, HeXL ( 2007). Leaf anatomic characteristics of 12 species of Artemisia in Mu Us desert Journal of Northwest A & F University (Natural Science Edition), 35, 211-217. [本文引用: 1]
SmithWK, VogelmannTC, DeLuciaEH, BellDT, ShepherdKA ( 1997). Leaf form and photosynthesis BioScience, 47, 785-793. [本文引用: 1]
SongLY, PengCL, PengSL ( 2009). Comparison of leaf construction costs between three invasive species and three native species in South China Biodiversity Science, 17, 378-384. [本文引用: 1]
SuárezN ( 2005). Leaf construction cost in Avicennia germinans as affected by salinity under field conditions Biologia Plantarum, 49, 111-116. [本文引用: 1]
TangSB, ZhangLL, KuangYW, YanJH, ChenFL ( 2017). Leaf construction costs of 34 dominant species in karst forest, Guizhou Earth and Environment, 45, 18-24. [本文引用: 1]
TuCY, HuangfuCH, JiangN, GaoSB, YangDL ( 2013). Comparison of leaf construction cost between invasive plant Flaveria bidentis and its five co-occuring plants Chinese Journal of Ecology, 32, 2985-2991. [本文引用: 1]
TurnerNC, SchulzeED, NicolleD, SchumacherJ, KuhlmannI ( 2008). Annual rainfall does not directly determine the carbon isotope ratio of leaves of Eucalyptus species Physiologia Plantarum, 132, 440-445. [本文引用: 2]
VillarR, MerinoJ ( 2001). Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems New Phytologist, 151, 213-226. [本文引用: 5]
WeiH, WuB, YangW, LuoT ( 2011). Low rainfall-induced shift in leaf trait relationship within species along a semi-arid sandy land transect in northern China Plant Biology, 13, 85-92. [本文引用: 4]
WeiHX, LuoTX, WuB ( 2016). Optimal balance of water use efficiency and leaf construction cost with a link to the drought threshold of the desert steppe ecotone in northern China Annals of Botany, 118, 541-553. [本文引用: 3]
WilliamsK, PercivalF, MerinoJ, MooneyHA ( 1987). Estimation of tissue construction cost from heat of combustion and organic nitrogen content Plant, Cell & Environment, 10, 725-734. [本文引用: 3]
WrightIJ, ReichPB, WestobyM ( 2001). Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats Functional Ecology, 15, 423-434. [本文引用: 2]
WrightIJ, ReichPB, WestobyM ( 2003). Least-cost input mixtures of water and nitrogen for photosynthesis The American Naturalist, 161, 98-111. [本文引用: 1]
WrightIJ, WestobyM ( 2002). Leaves at low versus high rainfall: Coordination of structure, lifespan and physiology New Phytologist, 155, 403-416. [本文引用: 1]
WuB, CiLJ ( 2002). Landscape change and desertification development in the Mu Us sandy land, northern China Journal of Arid Environments, 50, 429-444. [本文引用: 1]
ZhangJ, HuangYM, JiaoHJ, XiaXD ( 2007). Ecophysiological characteristics of Artemisia ordosica community succession in Mu Us sandy grassland Journal of Desert Research, 27, 977-983. [本文引用: 1]
ZhangJZ, BaiXF, HouYP, DongZY, BuQM ( 2016). Comparison on the competitiveness of the invaded pokeweed with its accompanying species in the coastal protection forest of Shandong Peninsula Scientia Silvae Sinicae, 52(3), 23-29. [本文引用: 1]
ZhangL, LuoTX, LiuXS, WangY ( 2012). Altitudinal variation in leaf construction cost and energy content of Bergenia purpurascens Acta Oecologica, 43, 72-79. [本文引用: 2]
ZhuSD, LiRH, SongJ, HePC, LiuH, BerningerF, YeQ ( 2016). Different leaf cost-benefit strategies of ferns distributed in contrasting light habitats of sub-tropical forests Annals of Botany, 117, 497-506. [本文引用: 1]
Respiration and Crop Productivity 1 1989
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
The ability of several high arctic plant species to utilize nitrate nitrogen under field conditions 1 1993
Functional analysis of the relative growth rate, chemical composition, construction and maintenance costs, and the payback time of Coffea arabica L. leaves in response to light and water availability 1 2012
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
江西九连山常绿阔叶林主要树种叶建成消耗的比较 1 2006
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
江西九连山常绿阔叶林主要树种叶建成消耗的比较 1 2006
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
胶东滨海8种树木叶片热值、建成成本及其适应能力 1 2015
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
胶东滨海8种树木叶片热值、建成成本及其适应能力 1 2015
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
Leaf construction cost is related to water availability in three species of different growth forms in a Brazilian tropical dry forest 3 2017
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
Leaf, stem and root tissue strategies across 758 neotropical tree species 1 2012
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
Calorimetric estimates of construction cost and their use in ecological studies 1 1994
Does energetic cost for leaf construction in Sonneratia change after introduce to another mangrove wetland and differ from native mangrove plants in South China? 1 2017
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
青海省沙珠玉治沙站17种主要植物叶性因子的比较 1 2005
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
青海省沙珠玉治沙站17种主要植物叶性因子的比较 1 2005
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
Chemical composition and construction cost for roots of Mediterranean trees, shrub species and grassland communities 1 2002
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
Evidence for plant traits driving specific drought resistance. A community field experiment 1 2012
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
Construction cost and invasive potential: Comparing Lythrum salicaria ( Lythraceae) with co-occurring native species along pond banks 1 2001
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
Atmospheric CO2 enrichment alters energy assimilation, investment and allocation in Xanthium strumarium 1 2005
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
Products, requirements and efficiency of biosynthesis: A quantitative approach 1 1974
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
Variation in caloric values of annual and perennial lupines (Lupinus: Leguminosae) 1 1978
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
Construction costs and payback time of biomass: A whole plant perspective 1 1994
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
A comparison of specific leaf area, chemical composition and leaf construction costs of field plants from 15 habitats differing in productivity 2 1999
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
... 本研究中, 不同沙地生境和降水条件下土壤全氮含量均存在显著差异(表1), 偏相关分析表明, 油蒿CCm随土壤全氮含量的增加而增加(表2), 这与以往的研究结果(Griffin et al., 1993, 1996; Poorter & de Jong, 1999)一致.Griffin等(1996)研究认为, 土壤中全氮含量的增加导致了叶片中氮含量的增加, 因叶片中的氮主要用于蛋白质的构建, 叶片蛋白质含量的增加降低了总体的非结构碳骨架的比率, 增加了高能量投资组分的比例, 从而增加了CCm.我们前期的研究发现, 土壤全氮含量的增加并没有导致油蒿叶片中氮含量的增加(Wei et al., 2011), 油蒿单位质量叶氮含量与CCm相关性不显著, 因此, 土壤全氮含量对CCm的影响可能并不是通过影响叶氮含量实现的. ...
Construction costs, chemical composition and payback time of high- and low-irradiance leaves 1 2006
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
The fate of acquired carbon in plants: Chemical composition and construction costs 2 1997
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
Balancing the costs of carbon gain and water transport: Testing a new theoretical framework for plant functional ecology 1 2014
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
Plant construction cost in the boreal species differing in their ecological strategies 1 2001
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
Construction costs and mesostructure of leaves in hydrophytes 1 2014
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
华南地区3种入侵植物与本地植物叶片建成成本的比较 1 2009
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
Leaf construction cost in Avicennia germinans as affected by salinity under field conditions 1 2005
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
贵州喀斯特森林34个优势种叶片构建成本特征 1 2017
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
贵州喀斯特森林34个优势种叶片构建成本特征 1 2017
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
入侵植物黄顶菊与5种共生植物叶片建成成本的比较 1 2013
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
入侵植物黄顶菊与5种共生植物叶片建成成本的比较 1 2013
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
Annual rainfall does not directly determine the carbon isotope ratio of leaves of Eucalyptus species 2 2008
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
... 植物对环境的适应往往表现为几个叶性状的组合, 而叶性状关系格局的变化体现了植物对环境变化的适应策略(Wright et al., 2001; Wright & Westoby, 2002).本研究中, 油蒿CCa-Narea正相关关系在低降水量区(鄂托克旗, 264 mm)植物和其他3个高降水量区(榆林、乌审旗和鄂托克前旗, 310-370 mm)植物之间出现了策略位移现象(图3D), 在相同CCa下, 低降水量区植物比高降水量区植物具有更高的Narea, 而Narea的增加有助于提高植物光合生产力和水分利用效率(Turner et al., 2008; Wei et al., 2011, 2016), 这一生理生态机制在一定程度上解释了油蒿在毛乌素沙地的广泛分布特性及其生态系统的相对稳定性.青藏高原北部沙地两个不同降水地区(都兰, 210 mm; 青海湖, 405 mm)的12个样方测定数据进一步证实了油蒿叶建成成本与Narea关系沿降水梯度发生策略位移现象的普遍性(图4A, 斜率6.39 vs. 3.54, p > 0.05, 截距p < 0.05; 图4B, 斜率0.015 vs. 0.004, p > 0.05, 截距p < 0.05). ...
Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems 5 2001
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
... 叶建成成本作为衡量植物能量利用效率的重要指标, 反映了植物适应环境的能量策略(Penning de Vries et al., 1974; Villar & Merino, 2001).关于植物叶建成成本对环境因子的响应仍存在争议.一些研究者认为, 植物在受到环境胁迫时, 叶建成成本会增加(Amthor, 1989; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
... ; Villar & Merino, 2001; Fortunel et al., 2012; Matías et al., 2012); 但也有研究者认为, 植物在受到环境胁迫时会降低叶建成成本以增强对环境的适应能力(Pitelka, 1978; Suárez, 2005; Falcão et al., 2017).与以往研究不同, 本研究中, 随着降水的减少, 油蒿CCm并没有表现出增加或降低的变化规律(图2A), 而偏相关分析结果也进一步表明降水对CCm没有显著影响(表2).通常CCm是由叶片化学特性决定的, 因此, 植物CCm沿着降水梯度的变化规律可能与叶片在不同降水条件下具有不同的生化组分有关.研究表明, 植物叶片中高能量投资组分与低能量投资组分呈正相关关系, 如蛋白质和矿质元素(Poorter, 1994; Poorter & Villar, 1997), 高能量投资组分之间呈负相关关系, 如蛋白质和油脂(Martínez et al., 2002).因此, 这种补偿机制有可能导致不同环境条件下植物具有相似的CCm.在本研究中, 油蒿热值(与高能物质含量呈正相关关系)与灰分含量(与矿质元素含量呈正相关关系)沿着降水梯度较为一致的变化规律(数据没有列出)可能是导致不同降水条件下油蒿CCm没有显著差异的主要原因. ...
Low rainfall-induced shift in leaf trait relationship within species along a semi-arid sandy land transect in northern China 4 2011
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
... ; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
... 本研究中, 不同沙地生境和降水条件下土壤全氮含量均存在显著差异(表1), 偏相关分析表明, 油蒿CCm随土壤全氮含量的增加而增加(表2), 这与以往的研究结果(Griffin et al., 1993, 1996; Poorter & de Jong, 1999)一致.Griffin等(1996)研究认为, 土壤中全氮含量的增加导致了叶片中氮含量的增加, 因叶片中的氮主要用于蛋白质的构建, 叶片蛋白质含量的增加降低了总体的非结构碳骨架的比率, 增加了高能量投资组分的比例, 从而增加了CCm.我们前期的研究发现, 土壤全氮含量的增加并没有导致油蒿叶片中氮含量的增加(Wei et al., 2011), 油蒿单位质量叶氮含量与CCm相关性不显著, 因此, 土壤全氮含量对CCm的影响可能并不是通过影响叶氮含量实现的. ...
... 植物对环境的适应往往表现为几个叶性状的组合, 而叶性状关系格局的变化体现了植物对环境变化的适应策略(Wright et al., 2001; Wright & Westoby, 2002).本研究中, 油蒿CCa-Narea正相关关系在低降水量区(鄂托克旗, 264 mm)植物和其他3个高降水量区(榆林、乌审旗和鄂托克前旗, 310-370 mm)植物之间出现了策略位移现象(图3D), 在相同CCa下, 低降水量区植物比高降水量区植物具有更高的Narea, 而Narea的增加有助于提高植物光合生产力和水分利用效率(Turner et al., 2008; Wei et al., 2011, 2016), 这一生理生态机制在一定程度上解释了油蒿在毛乌素沙地的广泛分布特性及其生态系统的相对稳定性.青藏高原北部沙地两个不同降水地区(都兰, 210 mm; 青海湖, 405 mm)的12个样方测定数据进一步证实了油蒿叶建成成本与Narea关系沿降水梯度发生策略位移现象的普遍性(图4A, 斜率6.39 vs. 3.54, p > 0.05, 截距p < 0.05; 图4B, 斜率0.015 vs. 0.004, p > 0.05, 截距p < 0.05). ...
Optimal balance of water use efficiency and leaf construction cost with a link to the drought threshold of the desert steppe ecotone in northern China 3 2016
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
... , 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
... 植物对环境的适应往往表现为几个叶性状的组合, 而叶性状关系格局的变化体现了植物对环境变化的适应策略(Wright et al., 2001; Wright & Westoby, 2002).本研究中, 油蒿CCa-Narea正相关关系在低降水量区(鄂托克旗, 264 mm)植物和其他3个高降水量区(榆林、乌审旗和鄂托克前旗, 310-370 mm)植物之间出现了策略位移现象(图3D), 在相同CCa下, 低降水量区植物比高降水量区植物具有更高的Narea, 而Narea的增加有助于提高植物光合生产力和水分利用效率(Turner et al., 2008; Wei et al., 2011, 2016), 这一生理生态机制在一定程度上解释了油蒿在毛乌素沙地的广泛分布特性及其生态系统的相对稳定性.青藏高原北部沙地两个不同降水地区(都兰, 210 mm; 青海湖, 405 mm)的12个样方测定数据进一步证实了油蒿叶建成成本与Narea关系沿降水梯度发生策略位移现象的普遍性(图4A, 斜率6.39 vs. 3.54, p > 0.05, 截距p < 0.05; 图4B, 斜率0.015 vs. 0.004, p > 0.05, 截距p < 0.05). ...
Estimation of tissue construction cost from heat of combustion and organic nitrogen content 3 1987
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
... 一般为0.89 (Williams et al., 1987). 本研究假设植物的氮来源均为硝态氮, 因为在大多数生境下硝态氮是高等植物最主要的氮源(Villar & Merino, 2001), 而铵态氮仅在苔原地区是植物的主要氮来源(Atkin et al., 1993).单位面积叶建成成本(CCa)为CCm与SLA的比值. ...
Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats 2 2001
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
... 植物对环境的适应往往表现为几个叶性状的组合, 而叶性状关系格局的变化体现了植物对环境变化的适应策略(Wright et al., 2001; Wright & Westoby, 2002).本研究中, 油蒿CCa-Narea正相关关系在低降水量区(鄂托克旗, 264 mm)植物和其他3个高降水量区(榆林、乌审旗和鄂托克前旗, 310-370 mm)植物之间出现了策略位移现象(图3D), 在相同CCa下, 低降水量区植物比高降水量区植物具有更高的Narea, 而Narea的增加有助于提高植物光合生产力和水分利用效率(Turner et al., 2008; Wei et al., 2011, 2016), 这一生理生态机制在一定程度上解释了油蒿在毛乌素沙地的广泛分布特性及其生态系统的相对稳定性.青藏高原北部沙地两个不同降水地区(都兰, 210 mm; 青海湖, 405 mm)的12个样方测定数据进一步证实了油蒿叶建成成本与Narea关系沿降水梯度发生策略位移现象的普遍性(图4A, 斜率6.39 vs. 3.54, p > 0.05, 截距p < 0.05; 图4B, 斜率0.015 vs. 0.004, p > 0.05, 截距p < 0.05). ...
Least-cost input mixtures of water and nitrogen for photosynthesis 1 2003
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
Leaves at low versus high rainfall: Coordination of structure, lifespan and physiology 1 2002
... 植物对环境的适应往往表现为几个叶性状的组合, 而叶性状关系格局的变化体现了植物对环境变化的适应策略(Wright et al., 2001; Wright & Westoby, 2002).本研究中, 油蒿CCa-Narea正相关关系在低降水量区(鄂托克旗, 264 mm)植物和其他3个高降水量区(榆林、乌审旗和鄂托克前旗, 310-370 mm)植物之间出现了策略位移现象(图3D), 在相同CCa下, 低降水量区植物比高降水量区植物具有更高的Narea, 而Narea的增加有助于提高植物光合生产力和水分利用效率(Turner et al., 2008; Wei et al., 2011, 2016), 这一生理生态机制在一定程度上解释了油蒿在毛乌素沙地的广泛分布特性及其生态系统的相对稳定性.青藏高原北部沙地两个不同降水地区(都兰, 210 mm; 青海湖, 405 mm)的12个样方测定数据进一步证实了油蒿叶建成成本与Narea关系沿降水梯度发生策略位移现象的普遍性(图4A, 斜率6.39 vs. 3.54, p > 0.05, 截距p < 0.05; 图4B, 斜率0.015 vs. 0.004, p > 0.05, 截距p < 0.05). ...
Landscape change and desertification development in the Mu Us sandy land, northern China 1 2002
... 在每个研究点, 选择两类不同退化程度的沙地生境(固定沙地和半固定沙地)进行植物和土壤样品的采集.在本研究中, 对固定和半固定沙地生境的界定主要依据植被盖度(>40% vs. 20%-40%)和土壤结皮厚度(灌丛下和灌丛间结皮厚度>1 cm vs. 灌丛下结皮厚度0.5-1 cm)(Wu & Ci, 2002).对于两类沙地生境, 分别设置3个5 m × 5 m的样方进行植物和土壤样品的采集, 共设置了24个采样样方. ...
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
山东半岛沿海防护林入侵植物美洲商陆及其伴生种生长竞争力的比较 1 2016
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
Altitudinal variation in leaf construction cost and energy content of Bergenia purpurascens 2 2012
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
... 在干旱和半干旱生态系统中, 水分是决定植物叶性状变化的主要因子(李永华等, 2005; Wei et al., 2011, 2016).目前关于水分对叶建成成本影响的研究较少, 植物在受到干旱胁迫时, 叶建成成本更高或更低依然不明确(Villar & Merino, 2001; Falcão et al., 2017).理论上, 在干旱和半干旱地区, 干旱胁迫可能会导致一些化合物的累积, 这些化合物(脂质)可能有助于植物组织防止水分的散失等干旱适应, 但由于这些物质的合成需要较高的能量, 植物叶建成成本可能增加.此外, 以往研究表明, 随着降水的减少, 植物可通过降低SLA或提高单位质量叶氮含量(Nmass)的方式来提高水分利用效率(Wright et al., 2001; Turner et al., 2008; Wei et al., 2011, 2016).具有较低SLA和较高Nmass的叶片往往具有高的建成成本(Nagel & Griffin, 2001; 陈飞宇等, 2006; Zhang et al., 2012).综上所述, 干旱区植物在提高水分利用效率(高的单位面积叶氮含量, Narea)与降低叶建成成本之间可能存在一种权衡(Wright et al., 2003; Prentice et al., 2014), 与湿润区植物相比, 干旱区植物可能通过调节叶功能性状之间的关系(如通过提高Narea), 实现在相同的叶建成成本下具有更高的水分利用效率.但是, 这种预测仍有待验证, 尤其需要同一物种沿降水梯度的观测数据加以验证. ...
Different leaf cost-benefit strategies of ferns distributed in contrasting light habitats of sub-tropical forests 1 2016
... 叶建成成本是植物在碳收获过程中所必需的成本投资, 常以构建单位质量(面积)叶片所需要的葡萄糖当量表示(Williams et al., 1987).不同物种间和不同环境条件下植物叶建成成本的变化反映了植物的能量利用策略和对环境的适应能力(P’yankov et al., 2001; Villar & Merino, 2001; 董周焱等, 2015; 汤松波等, 2017).研究表明, 具有较低叶建成成本的植物往往具有较高的生长速率(Pooter & Villar, 1997; Cavatte et al., 2012)和较强的入侵能力(宋莉英等, 2009; 屠臣阳等, 2013; 张靖梓等, 2016; Li et al., 2017).植物叶建成成本主要受到光照强度(Poorter et al., 2006; Zhu et al., 2016)、CO2浓度(Nagel et al., 2005)、土壤氮含量(Poorter & de Jong, 1999)以及水分(Falcão et al., 2017)等环境因子的影响.此外, 植物叶片的其他关键性状如叶寿命、比叶面积(SLA)和叶氮含量等也会影响叶建成成本(Zhang et al., 2012; Ronzhina & Ivanov, 2014). ...
 ,1,2,3,*
,1,2,3,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT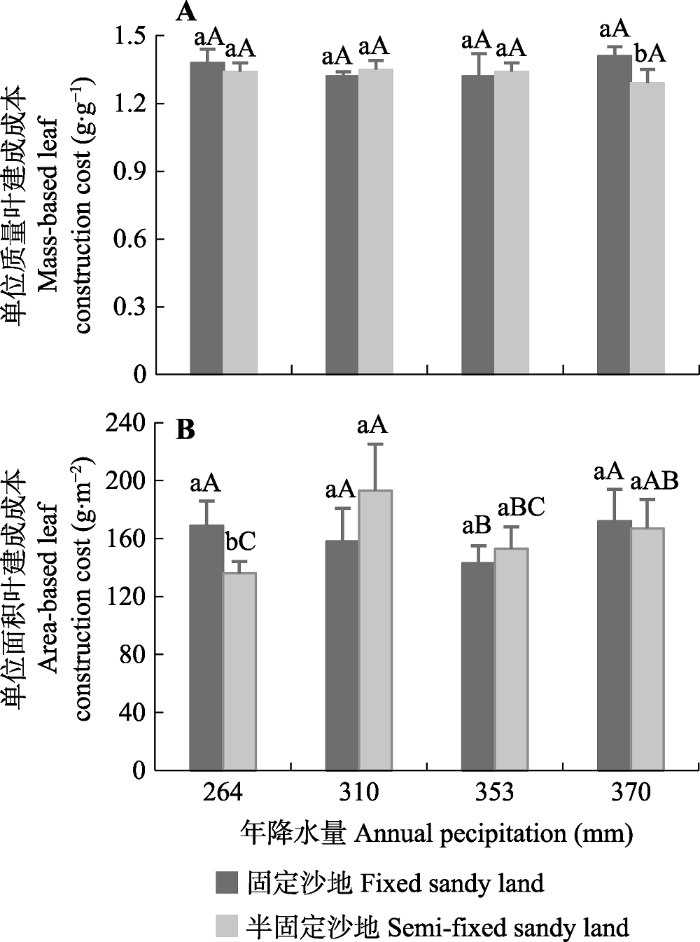 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT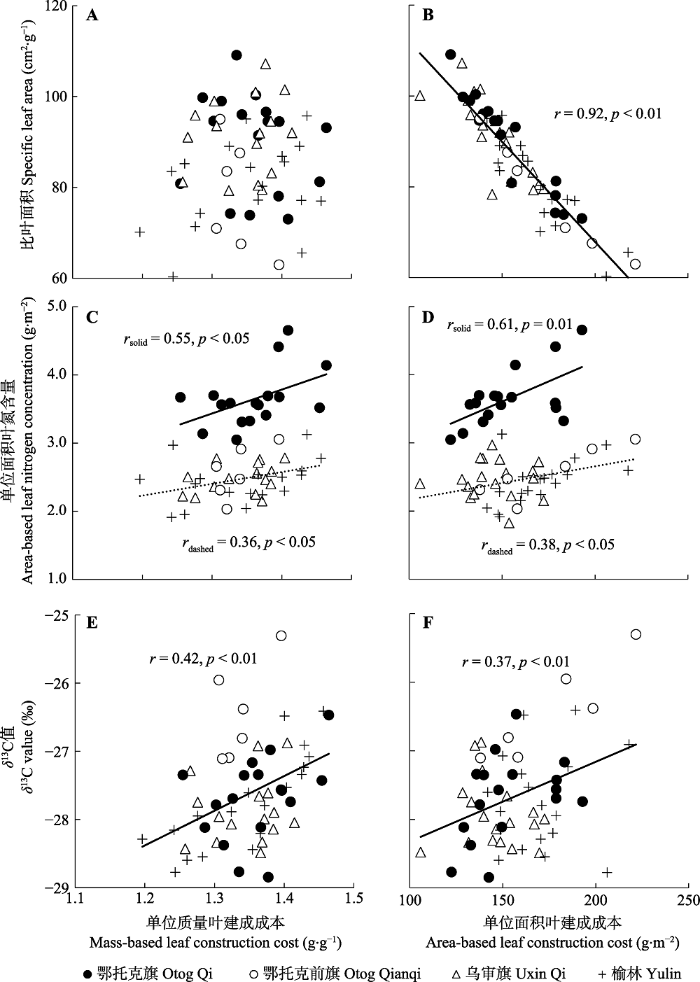 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT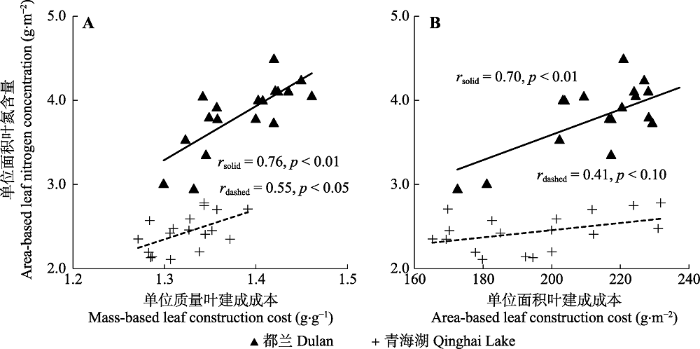 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}