Effects of simulated changes in precipitation pattern on sap flux in two tree species in subtropical region
Zhen-Zhen ZHANG,1,*, Ke-Jia YANG1, Yu-Lu GU1, Ping ZHAO2, Lei OUYANG21College of Geography and Environmental Sciences, Zhejiang Normal University, Jinhua, Zhejiang 321000, China 2South China Botanical Garden, Chinese Academy of Sciences, Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, Chinese Academy of Sciences, Guangdong Provincial Key Laboratory of Applied Botany, Guangzhou 510650, China
Supported by the National Natural Science Foundation of China(41630752) Supported by the National Natural Science Foundation of China(41701226) the Zhejiang Province Public Welfare Technology Application Research Project(LGF19C030002)
Abstract Aims Over the past decades, the precipitation patterns in subtropical regions markedly changed in response to global climate change. The impacts of changing precipitation patterns on plant growths and forest water balance remain unclear. In this study, the effects of varying precipitation patterns on whole tree water were tested in a natural forest in South China.Methods The study was conducted in the Heshan National Field Research Station of Forest Ecosystem in Guangdong Province, from September 2012 to December 2014. Throughfalls were intercepted by installing rain-shelters underneath the tree canopy and transferred to a nearby water reservior in dry season (from October to March of the next year), which were then reapplied to the field plots in equal quantity of the interception in wet season (from April to September), to simulate changed rainfall pattern to drier dry season and wetter wet season (DD). Sap flux density was continually measured on two tree species, Schima superba and Michelia macclurei. Student t-test was used to determine the significance of differences in mean maximum sap flux density (ˉJS) between the two species in the control plots (AC), and between AC and DD treatments during the experiment.Important findings The average ˉJS was (49.5 ± 1.7) mL?m -2?s -1in M. macclurei and (43.6 ± 2.0) mL?m -2?s -1 in S. superba in the AC treatment when the active photosynthetic radiation (PAR) was greater than 1 100 μmol·m -2·s -1. M. macclurei showed higher sensitivity to increasing PAR. The ˉJS ratio (DD/AC) in both species initially increased significantly, followed by a short-term decrease. In S. superba, the ratio decreased from 0.74 to 0.68 in DD from October 2012 to March 2013, and then increased to 0.93 in March 2014 and 1.04 in November 2014. In M. macclurei, the ratio decreased from 1.00 to 0.94 in DD from October 2012 to March 2013, and then increased to 1.06 in March 2014. We found that S. superba could maintain higher ˉJS in response to the increasing PAR and vapor pressure deficit (VPD) in the DD treatment. Our results showed that the short-term drought would lead to a decline in tree transpiration; but in the long run, plants tended to compensate for the drought induced growth loss by elevating the ˉJS. Compared to M. macclurei, S. superba could maintain higher water transport capacity due to its more extensive ˉJS plasticity in response to the extended drought. Keywords:changed precipitation patterns;water use strategy;sap flux density;water stress
PDF (1900KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张振振, 杨轲嘉, 顾宇璐, 赵平, 欧阳磊. 模拟降雨格局变化对亚热带地区两树种液流特征的影响. 植物生态学报, 2019, 43(11): 988-998. DOI: 10.17521/cjpe.2019.0128 ZHANG Zhen-Zhen, YANG Ke-Jia, GU Yu-Lu, ZHAO Ping, OUYANG Lei. Effects of simulated changes in precipitation pattern on sap flux in two tree species in subtropical region. Chinese Journal of Plant Ecology, 2019, 43(11): 988-998. DOI: 10.17521/cjpe.2019.0128
森林植被受降雨格局变化的影响程度往往与不同树种的干旱响应策略密切相关(Attia et al., 2015; Liu et al., 2015)。例如同一地区,非等渗植物(anisohydric species)的生长受干旱加剧的影响相对较小(Attia et al., 2015; Liu et al., 2015)。在我国半干旱地区, 李亮等(2011)发现不耐旱植物核桃楸(Juglans mandshurica)的生物量在长期干旱影响下并没有降低, 而耐旱植物辽东栎(Quercus liaotungensis)受长期干旱影响时生物量显著下降。Attia等(2015)认为, 干旱发生时, 在不同的环境及资源条件下, 植物往往会采取3种不同的响应策略来应对水分胁迫: (1)高存活率, 低生长率; (2)高耐受性, 适度生长; (3)低存活率, 高生长率。这种种间差异会导致不同森林类型间对降雨格局变化的响应的异质性。如地中海常绿针叶林对干旱的耐受性远低于湿润的温带落叶阔叶林(Gazol et al., 2018)。因此, 降雨变化对森林生态系统影响的长期模拟研究应该考虑物种对干旱的不同响应策略。
植物的生长速率往往取决于冠层叶片蒸腾耦合的气体交换的速率(Catovsky et al., 2002; Skelton et al., 2017)。因此在干旱条件下, 植物蒸腾的响应能够在较大程度上反映干旱对植物生长的影响。树干液流作为植物最重要的一种生理活动, 反映了植物水分传输能力, 因而经常被用来研究环境变化对植物生长的影响(Graham et al., 2003; Steppe et al., 2006)。为了检验降雨格局变化对我国亚热带常绿阔叶林的影响,我们在亚热带地区典型演替早期天然次生林中选择木荷(Schima superba)和火力楠(Michelia macclurei)为研究对象, 开展人工隔除林下降雨实验模拟降雨格局变化。木荷(SS)为亚热带地区演替早期优势树种, 火力楠(MM)则为伴生其中的亚优势种。在木材解剖结构上, 两者均属于散孔材树种和深根系植物(蔡锡安等, 2012; 张振振等, 2014, 2019), 且生长迅速, 为理想的造林树种。通过对两者树干液流的监测, 我们探讨加剧的季节性干旱对演替早期优势树种水分传输的影响。具体来讲,我们主要关注以下两个问题: (1)在降雨量较为丰沛的亚热带地区, 林下降雨隔除对该地区的树干液流是否会带来负效应? (2)不同树种树干液流对干旱的响应方式是否存在差异? 这一研究将给气候变化背景下森林群落演替的变化规律研究提供参考。
1 材料和方法
1.1 研究地概况
本研究位于广东鹤山森林生态系统国家野外科学观测研究站(22.68° N, 112.90° E, 海拔47 m)。属亚热带季风气候, 年降水量为1 580.4 mm, 年平均气温21.92 ℃。历史最低气温0 ℃。降水分布可划分为明显的干季和雨季。干季从10月持续到次年3月, 而雨季从4月持续至9月。干季降水量在一年中所占的比例为17.72%, 是雨季降水量的21.54% (1985-2014年的年平均值)。地带性植被以亚热带季风常绿阔叶林为主。实验样地位于试验站内东南面一个坡度为15°的斜坡上。土壤是在砂岩上发育并受强烈淋溶的红土, pH为4.26。
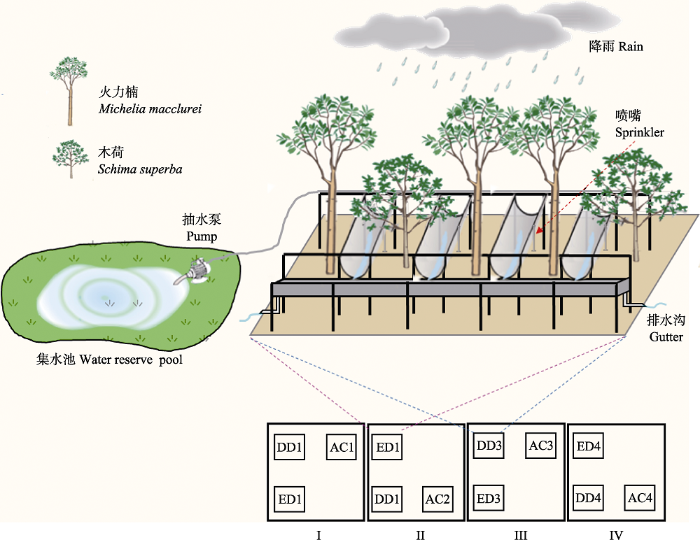
Fig. 1Experimental designs for the “simulated seasonal changes in precipitation”. Throughfall exclusion from October to March of the following year (DD) and from April to May (ED). Reapplication of throughfall rainwater from June to September. AC, control; DD, drier dry season and wetter wet season; ED, extended dry season.
Table 1 表1 表1截留降雨处理和对照组不同树种在鹤山实验站各区组中的个体数量、胸径及树高(平均值±标准偏差) Table 1The number of trees and tree diameter at breast height (DBH), tree height for different species in the drier dry season and wetter wet season (DD) and control (AC) treatments (mean ± SD) in each block in Heshan experimental station
DD
AC
物种 Species
I
II
III
IV
I
II
III
IV
数量 N
SS
3
1
3
0
3
1
2
3
MM
0
3
3
6
3
5
4
3
胸径 DBH (cm)
SS
14.02 ± 0.93 (0.13)
12.93 ± 1.17 (0.15)
MM
19.02 ± 1.43 (0.09)
17.53 ± 1.35 (0.11)
树高 H (m)
SS
7.02 ± 0.48 (0.11)
7.04 ± 0.5 (0.10)
MM
13.59 ± 0.51 (0.12)
9.43 ± 0.64 (0.13)
边材厚度 Sapwood depth (cm)
SS
5.5-6.3
MM
3.5-4.2
边材密度 Sapwood density (g·cm-3)
SS
0.61 ± 0.03 (0.05)
MM
0.53 ± 0.03 (0.06)
Data in brackets indicate the coefficient of variation. MM, Michelia macclurei; SS, Schima superba. AC, control; DD, drier dry season and wetter wet season. 括号中为变异系数。MM, 火力楠; SS, 木荷。AC, 对照; DD, 干季更干, 湿季更湿。
Table 2 表2 表2遮雨处理和对照组湿/干季降雨的灌溉/排除量及相应的土壤含水量 Table 2The excluded/irrigated precipitation in wet/dry season and the corresponding soil water content in the control and drier dry season and wetter wet season treatments
补水量 Water input (mm)
土壤含水量 Soil water content (%)
0-20 cm
50 cm
湿季增雨 Irrigated in wet season
DD1
370.56
35.12
39.34
DD2
370.56
33.85
38.69
DD3
370.56
DD4
370.56
AC1
0
34.98
37.79
AC2
0
34.13
38.55
AC3
0
AC4
0
干季减雨 Excluded in dry season
DD1
-533.25
22.15
31.04
DD2
-496.89
21.64
30.61
DD3
-499.37
DD4
-513.80
AC1
0
29.59
33.91
AC2
0
27.54
35.68
AC3
0
AC4
0
AC, control; DD, drier dry season and wetter wet season. AC, 对照; DD, 干季更干, 湿季更湿。
Fig. 2Annual dynamics of mean maximum sap flux density (¯JS)(average values between 11:00-13:00, mean ± SD) in Michelia macclurei and Schima superba, and the fitted relationship of ¯JS as a function of the photosynthetically active radiation (PAR) from the upper boundary of the boundary line analysis during 1 October 2013 and 31 March 2014.
Fig. 3Annual variations of mean maximum sap flux density (¯JS)(mean ± SD) for trees in DD (red dot) and AC (black dot) groups in Michelia macclurei (n = 15) and Schima superba (n = 9). The shaded sections indicate the period of throughfall exclusion, and other sections the period of enhanced rainfall. AC, control; DD, drier dry season and wetter wet season.
Fig. 4Linear fitting of the relationship of mean maximum sap flux density (¯JS)(mean ± SD) between control (AC) and drier dry season and wetter wet season (DD) treatments during different drought periods in Michelia macclurei (n = 15) and Schima superba (n = 9). The inserted figures show the ¯JS ratios (JS(DD)/JS(AC)) during different periods.
Fig. 5Daily dynamics of mean maximum sap flux density (¯JS) and environmental drivers (VPD and PAR) in drier dry season and wetter wet season (DD) treatments during the second phase of dry season (from 14 to 18 October 2013) in Michelia macclurei and Schima superba (mean ± SD). The arrow lines indicate the direction of time series. PAR, photosynthetically active radiation; VPD, water vapor deficit.
AndradeJL, MeinzerFC, GoldsteinG, HolbrookNM, CavelierJ, JacksonP, SilveraK ( 1998). Regulation of water flux through trunks, branches, and leaves in trees of a lowland tropical forest Oecologia, 115, 463-471. [本文引用: 1]
AttiaZ, DomecJC, OrenR, WayDA, MoshelionM ( 2015). Growth and physiological responses of isohydric and anisohydric poplars to drought Journal of Experimental Botany, 66, 4373-4381. [本文引用: 5]
BabstF, PoulterB, TrouetV, TanK, NeuwirthB, WilsonR, CarrerM, GrabnerM, TegelW, LevanicT, PanayotovM, UrbinatiC, BouriaudO, CiaisP, FrankD ( 2013). Site- and species-specific responses of forest growth to climate across the European continent Global Ecology and Biogeography, 22, 706-717. [本文引用: 1]
BonalD, BurbanB, StahlC, WagnerF, HéraultB ( 2016). The response of tropical rainforests to drought—Lessons from recent research and future prospects Annals of Forest Science, 73, 27-44. [本文引用: 1]
BrunnerI, HerzogC, DawesMA, ArendM, SperisenC ( 2015). How tree roots respond to drought Frontiers in Plant Science, 6, 547. DOI: 10.3389/fpls.2015.00547. [本文引用: 1]
CaiXA, ZhaoP, ZengXP, ZhuLW, ZouLL, MeiTT, YuMH ( 2012). Dynamics of xylem sap flow of two Magnoliaceae tree species in relation to environmental factors Chinese Journal of Ecology, 31, 2163-2169. [本文引用: 1]
CatovskyS, HolbrookNM, BazzazFA ( 2002). Coupling whole-tree transpiration and canopy photosynthesis in coniferous and broad-leaved tree species Canadian Journal of Forest Research, 32, 295-309. [本文引用: 1]
GaoJG, ZhaoP, ShenWJ, RaoXQ, HuYT ( 2017). Physiological homeostasis and morphological plasticity of two tree species subjected to precipitation seasonal distribution changes Perspectives in Plant Ecology, Evolution and Systematics, 25, 1-19. [本文引用: 3]
GazolA, CamareroJJ, Vicente-SerranoSM, Sánchez-SalgueroR, GutiérrezE, de LuisM, Sangüesa-BarredaG, NovakK, RozasV, TíscarPA, LinaresJC, Martín-HernándezN, Martínez Del CastilloE, RibasM, García-GonzálezI, SillaF, CamisónA, GénovaM, OlanoJM, LongaresLA, HeviaA, Tomás-BurgueraM, GalvánJD ( 2018). Forest resilience to drought varies across biomes Global Change Biology, 24, 2143-2158. [本文引用: 1]
GrahamEA, MulkeySS, KitajimaK, PhillipsNG, WrightSJ ( 2003). Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons Proceedings of the National Academy of Sciences of the United States of America, 100, 572-576. [本文引用: 1]
GranierA ( 1987). Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements Tree Physiology, 3, 309-320. [本文引用: 1]
HertelD, StreckerT, Müller-HauboldH, LeuschnerC ( 2013). Fine root biomass and dynamics in beech forests across a precipitation gradient—Is optimal resource partitioning theory applicable to water-limited mature trees? Journal of Ecology, 101, 1183-1200. [本文引用: 1]
HoeberS, LeuschnerC, K?hlerL, Arias-AguilarD, SchuldtB ( 2014). The importance of hydraulic conductivity and wood density to growth performance in eight tree species from a tropical semi-dry climate Forest Ecology and Management, 330, 126-136. [本文引用: 1]
HoffmannWA, MarchinRM, AbitP, LauOL ( 2011). Hydraulic failure and tree dieback are associated with high wood density in a temperate forest under extreme drought Global Change Biology, 17, 2731-2742. [本文引用: 1]
HuY, HuTX, HuHL, ChenH, WangB, LiH ( 2014). Effects of drought stress on growth and photosynthetic characteristics of Cinnamomum camphora saplings Chinese Journal of Applied and Environmental Biology, 20, 675-682. [本文引用: 1]
KlosRJ, WangGG, BauerleWL, RieckJR ( 2009). Drought impact on forest growth and mortality in the southeast USA: An analysis using forest health and monitoring data Ecological Applications, 19, 699-708. [本文引用: 1]
KuangYW, XuYM, ZhangLL, HouEQ, ShenWJ ( 2017). Dominant trees in a subtropical forest respond to drought mainly via adjusting tissue soluble sugar and proline content Frontiers in Plant Science, 8, 802. DOI: 10.3389/?fpls.2017.00802. [本文引用: 2]
LiL, SuHX, SangWG ( 2011). Simulating impacts of summer drought on forest dynamics in Dongling Mountain Chinese Journal of Plant Ecology, 35, 147-158. [本文引用: 2]
LiuDJ, OgayaR, BarbetaA, YangXH, Pe?uelasJ ( 2015). Contrasting impacts of continuous moderate drought and episodic severe droughts on the aboveground-biomass increment and litterfall of three coexisting Mediterranean woody species Global Change Biology, 21, 4196-4209. [本文引用: 2]
MalhiY, AragaoLEOC, GalbraithD, HuntingfordC, FisherR, ZelazowskiP, SitchS, McSweeneyC, MeirP ( 2009). Exploring the likelihood and mechanism of a climate-?change-induced dieback of the Amazon rainforest Proceedings of the National Academy of Sciences of the United States of America, 106, 20610-20615. [本文引用: 1]
ParryML, CanzianiOF, PalutikofJP, LindenPVD, HansonCE ( 2007). Intergovernmental Panel on Climate Change Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. [本文引用: 1]
PashoE, CamareroJJ, de LuisM, Vicente-SerranoSM ( 2011). Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain Agricultural and Forest Meteorology, 151, 1800-1811. [本文引用: 1]
SamantaA, GangulyS, HashimotoH, DevadigaS, VermoteE, KnyazikhinY, NemaniRR, MyneniRB ( 2010). Amazon forests did not green-up during the 2005 drought Geophysical Research Letters, 37, 1-5. [本文引用: 1]
SantiagoLS, GoldsteinG, MeinzerFC, FisherJB, MacHadoK, WoodruffD, JonesT ( 2004). Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees Oecologia, 140, 543-550. [本文引用: 1]
SeneviratneSI, LüthiD, LitschiM, Sch?rC ( 2006). Land-atmosphere coupling and climate change in Europe Nature, 443, 205-209. [本文引用: 1]
SkeltonRP, BrodribbTJ, McAdamSAM, MitchellPJ ( 2017). Gas exchange recovery following natural drought is rapid unless limited by loss of leaf hydraulic conductance: Evidence from an evergreen woodland New Phytologist, 215, 1399-1412. [本文引用: 1]
SteppeK, de PauwDJW, LemeurR, VanrolleghemPA ( 2006). A mathematical model linking tree sap flow dynamics to daily stem diameter fluctuations and radial stem growth Tree Physiology, 26, 257-273. [本文引用: 1]
SunGC, ZhaoP, ZengXP, CaiXA ( 2009). Hydraulic responses of stomatal conductance in leaves of successional tree species in subtropical forest to environmental moisture Acta Ecologica Sinica, 29, 698-708. [本文引用: 3]
Vicente-SerranoSM, GouveiaC, CamareroJJ, BegueríaS, TrigoR, López-MorenoJI, Azorín-MolinaC, PashoE, Lorenzo-LacruzJ, RevueltoJ, Morán-TejedaE, Sanchez- LorenzoA ( 2013). Response of vegetation to drought time-scales across global land biomes Proceedings of the National Academy of Sciences of the United States of America, 110, 52-57. [本文引用: 1]
WangWX, VinocurB, AltmanA ( 2003). Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance Planta, 218, 1-14. [本文引用: 1]
WangXY, ChenDP, XuGZ, LiY, LuJM ( 2017). Difference analysis of anatomic structure of Fraxinus mandschurica Rupr in different ecological environments Journal of Anhui Agricultural Sciences, 45(21), 1-3, 8. [本文引用: 1]
WeltzinJF, LoikME, SchwinningS, WilliamsDG, FayPA, HaddadBM, HarteJ, HuxmanTE, KnappAK, LinGH, PockmanWT, ShawMR, SmallEE, SmithMD, SmithSD, TissueDT, ZakJC ( 2003). Assessing the response of terrestrial ecosystems to potential changes in precipitation BioScience, 53, 941-952. [本文引用: 2]
WuQ, DingJ, YanH, ZhangSR, FangT, MaKP ( 2011). Effects of simulated precipitation and nitrogen addition on seedling growth and biomass in five tree species in Gutian Mountain, Zhejiang Province, China Chinese Journal of Plant Ecology, 35, 256-267. [本文引用: 4]
WuX, ChenYM, TangYK ( 2015). Sap flow characteristics and its responses to precipitation in Robinia pseudoacacia and Platycladus orientalis plantations Chinese Journal of Plant Ecology, 39, 1176-1187. [本文引用: 3]
XuF, GuoWH, XuWH, WangRQ ( 2010). Effects of water stress on morphology, biomass allocation and photosynthesis in Robinia pseudoacacia seedlings Journal of Beijing Forestry University, 32(1), 24-30. [本文引用: 2]
ZangU, GoisserM, H?berleKH, MatyssekR, MatznerE, BorkenW ( 2014). Effects of drought stress on photosynthesis, rhizosphere respiration, and fine-root characteristics of beech saplings: A rhizotron field study Journal of Plant Nutrition and Soil Science, 177, 168-177. [本文引用: 1]
ZhangZF, YouYM, HuangYQ, LiXK, ZhangJC, ZhangDN, HeCX ( 2012). Effects of drought stress on the photosynthesis and growth of Cyclobalanopsis glauca seedlings: A study with simulated hierarchical karst water supply Chinese Journal of Ecology, 31, 2197-2202. [本文引用: 1]
ZhangZZ, ZhaoP, McCarthyHR, ZhaoXH, NiuJF, ZhuLW, NiGY, OuyangL, HuangYQ ( 2016). Influence of the decoupling degree on the estimation of canopy stomatal conductance for two broadleaf tree species Agricultural and Forest Meteorology, 221, 230-241. [本文引用: 1]
ZhangZZ, ZhaoP, NiGY, ZhuLW, ZhaoXH, ZhaoPQ, NiuJF ( 2014). Water use of re-vegetation pioneer tree species Schima superba and Acacia mangium in hilly land of South China Chinese Journal of Applied Ecology, 25, 931-939. [本文引用: 1]
ZhaoP ( 2011). On the coordinated regulation of forest transpiration by hydraulic conductance and canopy stomatal conductance Acta Ecologica Sinica, 31, 1164-1173. [本文引用: 1]
ZhouGY, PengCH, LiYL, LiuSZ, ZhangQM, TangXL, LiuJX, YanJH, ZhangDQ, ChuGW ( 2013). A climate change-induced threat to the ecological resilience of a subtropical monsoon evergreen broad-leaved forest in Southern China Global Change Biology, 19, 1197-1210. [本文引用: 1]
ZhouJ, ZhangZQ, SunG, FangXR, ZhaTG, ZhangY, WangXP, ChenJQ, ChenJQ ( 2013). Environmental controls on water use efficiency of a poplar plantation under different soil water conditions Acta Ecologica Sinica, 33, 1465-1474. [本文引用: 1]
ZlatevZ, LidonFC ( 2012). An overview on drought induced changes in plant growth, water relations and photosynthesis Emirates Journal of Food and Agriculture, 24, 57-72. [本文引用: 1]
Regulation of water flux through trunks, branches, and leaves in trees of a lowland tropical forest 1 1998
... 一般来讲, 干旱加剧会引起叶片水势和气孔导度下降(Wang et al., 2003), 并造成乔木生长速率大幅度降低(Klos et al., 2009; Zlatev & Lidon, 2012), 这种现象普遍存在于干旱半干旱区(李亮等, 2011;吴旭等, 2015)、半湿润区(Pasho et al., 2011), 甚至湿润的热带亚热带地区(Samanta et al., 2010; Vicente- Serrano et al., 2013).例如在我国西南地区, 张中峰等(2012)发现, 在干旱胁迫下青冈(Cyclobalanopsis glauca)二年生幼苗光合速率和生长速率均显著降低.胡义等(2014)发现, 干旱胁迫下樟(Cinnamomum camphora)幼树的地径、树高生长量受到了抑制.然而这一结论在其他研究中并没有得到证实.例如最近研究发现, 当降水减少30%时, 古田山地区5个演替早期树种的生长均没有受到显著影响(吴茜等, 2011).孙谷畴等(2009)认为, 在我国亚热带地区, 演替早期树种能够在较强的水分胁迫下保持较高的气孔导度, 从而保证植物依然能够维持较高的光合速率(Attia et al., 2015).而演替后期树种, 即使叶片水势相对较高, 光合速率依然会大幅度下降(孙谷畴等, 2009).然而以上研究的实验对象往往是幼苗和幼树, 不能表征成熟森林的响应方式(吴茜等, 2011).而叶片尺度的植物生理测定由于具有较大的局限性, 很有可能会得到片面的结论(Andrade et al., 1998), 因此必须在整树尺度上对其进行验证(赵平, 2011; 吴旭等, 2015). ...
Growth and physiological responses of isohydric and anisohydric poplars to drought 5 2015
... 一般来讲, 干旱加剧会引起叶片水势和气孔导度下降(Wang et al., 2003), 并造成乔木生长速率大幅度降低(Klos et al., 2009; Zlatev & Lidon, 2012), 这种现象普遍存在于干旱半干旱区(李亮等, 2011;吴旭等, 2015)、半湿润区(Pasho et al., 2011), 甚至湿润的热带亚热带地区(Samanta et al., 2010; Vicente- Serrano et al., 2013).例如在我国西南地区, 张中峰等(2012)发现, 在干旱胁迫下青冈(Cyclobalanopsis glauca)二年生幼苗光合速率和生长速率均显著降低.胡义等(2014)发现, 干旱胁迫下樟(Cinnamomum camphora)幼树的地径、树高生长量受到了抑制.然而这一结论在其他研究中并没有得到证实.例如最近研究发现, 当降水减少30%时, 古田山地区5个演替早期树种的生长均没有受到显著影响(吴茜等, 2011).孙谷畴等(2009)认为, 在我国亚热带地区, 演替早期树种能够在较强的水分胁迫下保持较高的气孔导度, 从而保证植物依然能够维持较高的光合速率(Attia et al., 2015).而演替后期树种, 即使叶片水势相对较高, 光合速率依然会大幅度下降(孙谷畴等, 2009).然而以上研究的实验对象往往是幼苗和幼树, 不能表征成熟森林的响应方式(吴茜等, 2011).而叶片尺度的植物生理测定由于具有较大的局限性, 很有可能会得到片面的结论(Andrade et al., 1998), 因此必须在整树尺度上对其进行验证(赵平, 2011; 吴旭等, 2015). ...
... 森林植被受降雨格局变化的影响程度往往与不同树种的干旱响应策略密切相关(Attia et al., 2015; Liu et al., 2015).例如同一地区,非等渗植物(anisohydric species)的生长受干旱加剧的影响相对较小(Attia et al., 2015; Liu et al., 2015).在我国半干旱地区, 李亮等(2011)发现不耐旱植物核桃楸(Juglans mandshurica)的生物量在长期干旱影响下并没有降低, 而耐旱植物辽东栎(Quercus liaotungensis)受长期干旱影响时生物量显著下降.Attia等(2015)认为, 干旱发生时, 在不同的环境及资源条件下, 植物往往会采取3种不同的响应策略来应对水分胁迫: (1)高存活率, 低生长率; (2)高耐受性, 适度生长; (3)低存活率, 高生长率.这种种间差异会导致不同森林类型间对降雨格局变化的响应的异质性.如地中海常绿针叶林对干旱的耐受性远低于湿润的温带落叶阔叶林(Gazol et al., 2018).因此, 降雨变化对森林生态系统影响的长期模拟研究应该考虑物种对干旱的不同响应策略. ...
... ).例如同一地区,非等渗植物(anisohydric species)的生长受干旱加剧的影响相对较小(Attia et al., 2015; Liu et al., 2015).在我国半干旱地区, 李亮等(2011)发现不耐旱植物核桃楸(Juglans mandshurica)的生物量在长期干旱影响下并没有降低, 而耐旱植物辽东栎(Quercus liaotungensis)受长期干旱影响时生物量显著下降.Attia等(2015)认为, 干旱发生时, 在不同的环境及资源条件下, 植物往往会采取3种不同的响应策略来应对水分胁迫: (1)高存活率, 低生长率; (2)高耐受性, 适度生长; (3)低存活率, 高生长率.这种种间差异会导致不同森林类型间对降雨格局变化的响应的异质性.如地中海常绿针叶林对干旱的耐受性远低于湿润的温带落叶阔叶林(Gazol et al., 2018).因此, 降雨变化对森林生态系统影响的长期模拟研究应该考虑物种对干旱的不同响应策略. ...
... 在自然状态下, 当PAR > 1 100 μmol·m-2·s-1时, 火力楠的¯JS ((49.5 ± 1.7) mL∙m-2∙s-1)显著高于木荷((43.6 ± 2.0) mL∙m-2∙s-1), 并且当光照强度低时, 前者对PAR的增加具有更强的敏感性(图2), 这种差异性可能与两者的边材导水能力有关(吴旭等, 2015).尽管木荷和火力楠均为典型的散孔材, 但木荷的边材密度显著高于火力楠(Gao et al., 2017).以往的研究表明, 边材密度与最大导水率以及光合速率之间存在显著的负相关关系(Santiago et al., 2004; Hoeber et al., 2014).这种差异性会导致不同的干旱响应策略(Hoffmann et al., 2011).在我们的结果中, 尽管木荷与火力楠在受水分限制时的¯JS均具有相似的变化模式, 但火力楠¯JS的下降(干季1下降6%)和回升(干季2增加6%)幅度均小于木荷(干季1下降8.1%, 干季2增加25.7%, 干季3增加40.5%), 这表明木荷相对于火力楠具有更强的形态可塑性.事实上, Kuang等(2017)在对我们样地中这两种植物的研究中已经发现, 木荷在遮雨处理下, 在形态结构等方面表现出更强的可塑性和适应性, 尤其是根系长度和根系生物量.而我们的结果也表明, 在长期的干旱处理后, 木荷的¯JS对PAR和VPD的敏感性都高于火力楠(图5), 这与自然状态下两者的¯JS敏感性截然相反(图5).这说明与火力楠相比, 干旱发生时, 木荷更加倾向于非等渗行为(Attia et al., 2015), 这一水力特征保证植物在水分胁迫条件下依然能够维持较高的光合速率(孙谷畴等, 2009).因此, 我们在研究环境变化对森林植被的影响时, 即使是相同生境条件下, 不同树种间的水力响应策略差异也可能会对研究结果产生影响(Babst et al., 2013). ...
Forest resilience to drought varies across biomes 1 2018
... 森林植被受降雨格局变化的影响程度往往与不同树种的干旱响应策略密切相关(Attia et al., 2015; Liu et al., 2015).例如同一地区,非等渗植物(anisohydric species)的生长受干旱加剧的影响相对较小(Attia et al., 2015; Liu et al., 2015).在我国半干旱地区, 李亮等(2011)发现不耐旱植物核桃楸(Juglans mandshurica)的生物量在长期干旱影响下并没有降低, 而耐旱植物辽东栎(Quercus liaotungensis)受长期干旱影响时生物量显著下降.Attia等(2015)认为, 干旱发生时, 在不同的环境及资源条件下, 植物往往会采取3种不同的响应策略来应对水分胁迫: (1)高存活率, 低生长率; (2)高耐受性, 适度生长; (3)低存活率, 高生长率.这种种间差异会导致不同森林类型间对降雨格局变化的响应的异质性.如地中海常绿针叶林对干旱的耐受性远低于湿润的温带落叶阔叶林(Gazol et al., 2018).因此, 降雨变化对森林生态系统影响的长期模拟研究应该考虑物种对干旱的不同响应策略. ...
Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons 1 2003
... 植物的生长速率往往取决于冠层叶片蒸腾耦合的气体交换的速率(Catovsky et al., 2002; Skelton et al., 2017).因此在干旱条件下, 植物蒸腾的响应能够在较大程度上反映干旱对植物生长的影响.树干液流作为植物最重要的一种生理活动, 反映了植物水分传输能力, 因而经常被用来研究环境变化对植物生长的影响(Graham et al., 2003; Steppe et al., 2006).为了检验降雨格局变化对我国亚热带常绿阔叶林的影响,我们在亚热带地区典型演替早期天然次生林中选择木荷(Schima superba)和火力楠(Michelia macclurei)为研究对象, 开展人工隔除林下降雨实验模拟降雨格局变化.木荷(SS)为亚热带地区演替早期优势树种, 火力楠(MM)则为伴生其中的亚优势种.在木材解剖结构上, 两者均属于散孔材树种和深根系植物(蔡锡安等, 2012; 张振振等, 2014, 2019), 且生长迅速, 为理想的造林树种.通过对两者树干液流的监测, 我们探讨加剧的季节性干旱对演替早期优势树种水分传输的影响.具体来讲,我们主要关注以下两个问题: (1)在降雨量较为丰沛的亚热带地区, 林下降雨隔除对该地区的树干液流是否会带来负效应? (2)不同树种树干液流对干旱的响应方式是否存在差异? 这一研究将给气候变化背景下森林群落演替的变化规律研究提供参考. ...
Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements 1 1987
Fine root biomass and dynamics in beech forests across a precipitation gradient—Is optimal resource partitioning theory applicable to water-limited mature trees? 1 2013
Drought impact on forest growth and mortality in the southeast USA: An analysis using forest health and monitoring data 1 2009
... 一般来讲, 干旱加剧会引起叶片水势和气孔导度下降(Wang et al., 2003), 并造成乔木生长速率大幅度降低(Klos et al., 2009; Zlatev & Lidon, 2012), 这种现象普遍存在于干旱半干旱区(李亮等, 2011;吴旭等, 2015)、半湿润区(Pasho et al., 2011), 甚至湿润的热带亚热带地区(Samanta et al., 2010; Vicente- Serrano et al., 2013).例如在我国西南地区, 张中峰等(2012)发现, 在干旱胁迫下青冈(Cyclobalanopsis glauca)二年生幼苗光合速率和生长速率均显著降低.胡义等(2014)发现, 干旱胁迫下樟(Cinnamomum camphora)幼树的地径、树高生长量受到了抑制.然而这一结论在其他研究中并没有得到证实.例如最近研究发现, 当降水减少30%时, 古田山地区5个演替早期树种的生长均没有受到显著影响(吴茜等, 2011).孙谷畴等(2009)认为, 在我国亚热带地区, 演替早期树种能够在较强的水分胁迫下保持较高的气孔导度, 从而保证植物依然能够维持较高的光合速率(Attia et al., 2015).而演替后期树种, 即使叶片水势相对较高, 光合速率依然会大幅度下降(孙谷畴等, 2009).然而以上研究的实验对象往往是幼苗和幼树, 不能表征成熟森林的响应方式(吴茜等, 2011).而叶片尺度的植物生理测定由于具有较大的局限性, 很有可能会得到片面的结论(Andrade et al., 1998), 因此必须在整树尺度上对其进行验证(赵平, 2011; 吴旭等, 2015). ...
Dominant trees in a subtropical forest respond to drought mainly via adjusting tissue soluble sugar and proline content 2 2017
... 人类活动正深远地影响着全球的水文过程, 导致降水格局发生剧烈的变化(Seneviratne et al., 2006; 吴茜等, 2011; Kuang et al., 2017).在中国亚热带地区, 这种趋势主要表现为干季降水频率减少、雨季强降雨事件频发(Parry et al., 2007; 董思言和高学杰, 2014).其后果主要在于改变土壤含水量(Weltzin et al., 2003; 周洁等, 2013), 影响与植物水分利用相关的形态、结构和功能(吴茜等, 2011).尽管亚热带地区降水丰沛, 土壤含水量较高, 但干湿分明的特点使得植物仍然会受到周期性季节干旱的扰动.而在降水格局变化影响下, 这种干湿差异性可能会更加明显(Zhou et al., 2013).亚热带森林是中国森林生态系统的主要组成部分, 其微弱变化也可能对区域气候带来巨大的影响.然而目前对于当前这种降水格局变化究竟会对亚热带森林带来怎样的影响, 我们知之甚少. ...
... 在自然状态下, 当PAR > 1 100 μmol·m-2·s-1时, 火力楠的¯JS ((49.5 ± 1.7) mL∙m-2∙s-1)显著高于木荷((43.6 ± 2.0) mL∙m-2∙s-1), 并且当光照强度低时, 前者对PAR的增加具有更强的敏感性(图2), 这种差异性可能与两者的边材导水能力有关(吴旭等, 2015).尽管木荷和火力楠均为典型的散孔材, 但木荷的边材密度显著高于火力楠(Gao et al., 2017).以往的研究表明, 边材密度与最大导水率以及光合速率之间存在显著的负相关关系(Santiago et al., 2004; Hoeber et al., 2014).这种差异性会导致不同的干旱响应策略(Hoffmann et al., 2011).在我们的结果中, 尽管木荷与火力楠在受水分限制时的¯JS均具有相似的变化模式, 但火力楠¯JS的下降(干季1下降6%)和回升(干季2增加6%)幅度均小于木荷(干季1下降8.1%, 干季2增加25.7%, 干季3增加40.5%), 这表明木荷相对于火力楠具有更强的形态可塑性.事实上, Kuang等(2017)在对我们样地中这两种植物的研究中已经发现, 木荷在遮雨处理下, 在形态结构等方面表现出更强的可塑性和适应性, 尤其是根系长度和根系生物量.而我们的结果也表明, 在长期的干旱处理后, 木荷的¯JS对PAR和VPD的敏感性都高于火力楠(图5), 这与自然状态下两者的¯JS敏感性截然相反(图5).这说明与火力楠相比, 干旱发生时, 木荷更加倾向于非等渗行为(Attia et al., 2015), 这一水力特征保证植物在水分胁迫条件下依然能够维持较高的光合速率(孙谷畴等, 2009).因此, 我们在研究环境变化对森林植被的影响时, 即使是相同生境条件下, 不同树种间的水力响应策略差异也可能会对研究结果产生影响(Babst et al., 2013). ...
模拟夏季干旱对东灵山森林植被动态的影响 2 2011
... 一般来讲, 干旱加剧会引起叶片水势和气孔导度下降(Wang et al., 2003), 并造成乔木生长速率大幅度降低(Klos et al., 2009; Zlatev & Lidon, 2012), 这种现象普遍存在于干旱半干旱区(李亮等, 2011;吴旭等, 2015)、半湿润区(Pasho et al., 2011), 甚至湿润的热带亚热带地区(Samanta et al., 2010; Vicente- Serrano et al., 2013).例如在我国西南地区, 张中峰等(2012)发现, 在干旱胁迫下青冈(Cyclobalanopsis glauca)二年生幼苗光合速率和生长速率均显著降低.胡义等(2014)发现, 干旱胁迫下樟(Cinnamomum camphora)幼树的地径、树高生长量受到了抑制.然而这一结论在其他研究中并没有得到证实.例如最近研究发现, 当降水减少30%时, 古田山地区5个演替早期树种的生长均没有受到显著影响(吴茜等, 2011).孙谷畴等(2009)认为, 在我国亚热带地区, 演替早期树种能够在较强的水分胁迫下保持较高的气孔导度, 从而保证植物依然能够维持较高的光合速率(Attia et al., 2015).而演替后期树种, 即使叶片水势相对较高, 光合速率依然会大幅度下降(孙谷畴等, 2009).然而以上研究的实验对象往往是幼苗和幼树, 不能表征成熟森林的响应方式(吴茜等, 2011).而叶片尺度的植物生理测定由于具有较大的局限性, 很有可能会得到片面的结论(Andrade et al., 1998), 因此必须在整树尺度上对其进行验证(赵平, 2011; 吴旭等, 2015). ...
... 森林植被受降雨格局变化的影响程度往往与不同树种的干旱响应策略密切相关(Attia et al., 2015; Liu et al., 2015).例如同一地区,非等渗植物(anisohydric species)的生长受干旱加剧的影响相对较小(Attia et al., 2015; Liu et al., 2015).在我国半干旱地区, 李亮等(2011)发现不耐旱植物核桃楸(Juglans mandshurica)的生物量在长期干旱影响下并没有降低, 而耐旱植物辽东栎(Quercus liaotungensis)受长期干旱影响时生物量显著下降.Attia等(2015)认为, 干旱发生时, 在不同的环境及资源条件下, 植物往往会采取3种不同的响应策略来应对水分胁迫: (1)高存活率, 低生长率; (2)高耐受性, 适度生长; (3)低存活率, 高生长率.这种种间差异会导致不同森林类型间对降雨格局变化的响应的异质性.如地中海常绿针叶林对干旱的耐受性远低于湿润的温带落叶阔叶林(Gazol et al., 2018).因此, 降雨变化对森林生态系统影响的长期模拟研究应该考虑物种对干旱的不同响应策略. ...
模拟夏季干旱对东灵山森林植被动态的影响 2 2011
... 一般来讲, 干旱加剧会引起叶片水势和气孔导度下降(Wang et al., 2003), 并造成乔木生长速率大幅度降低(Klos et al., 2009; Zlatev & Lidon, 2012), 这种现象普遍存在于干旱半干旱区(李亮等, 2011;吴旭等, 2015)、半湿润区(Pasho et al., 2011), 甚至湿润的热带亚热带地区(Samanta et al., 2010; Vicente- Serrano et al., 2013).例如在我国西南地区, 张中峰等(2012)发现, 在干旱胁迫下青冈(Cyclobalanopsis glauca)二年生幼苗光合速率和生长速率均显著降低.胡义等(2014)发现, 干旱胁迫下樟(Cinnamomum camphora)幼树的地径、树高生长量受到了抑制.然而这一结论在其他研究中并没有得到证实.例如最近研究发现, 当降水减少30%时, 古田山地区5个演替早期树种的生长均没有受到显著影响(吴茜等, 2011).孙谷畴等(2009)认为, 在我国亚热带地区, 演替早期树种能够在较强的水分胁迫下保持较高的气孔导度, 从而保证植物依然能够维持较高的光合速率(Attia et al., 2015).而演替后期树种, 即使叶片水势相对较高, 光合速率依然会大幅度下降(孙谷畴等, 2009).然而以上研究的实验对象往往是幼苗和幼树, 不能表征成熟森林的响应方式(吴茜等, 2011).而叶片尺度的植物生理测定由于具有较大的局限性, 很有可能会得到片面的结论(Andrade et al., 1998), 因此必须在整树尺度上对其进行验证(赵平, 2011; 吴旭等, 2015). ...
... 森林植被受降雨格局变化的影响程度往往与不同树种的干旱响应策略密切相关(Attia et al., 2015; Liu et al., 2015).例如同一地区,非等渗植物(anisohydric species)的生长受干旱加剧的影响相对较小(Attia et al., 2015; Liu et al., 2015).在我国半干旱地区, 李亮等(2011)发现不耐旱植物核桃楸(Juglans mandshurica)的生物量在长期干旱影响下并没有降低, 而耐旱植物辽东栎(Quercus liaotungensis)受长期干旱影响时生物量显著下降.Attia等(2015)认为, 干旱发生时, 在不同的环境及资源条件下, 植物往往会采取3种不同的响应策略来应对水分胁迫: (1)高存活率, 低生长率; (2)高耐受性, 适度生长; (3)低存活率, 高生长率.这种种间差异会导致不同森林类型间对降雨格局变化的响应的异质性.如地中海常绿针叶林对干旱的耐受性远低于湿润的温带落叶阔叶林(Gazol et al., 2018).因此, 降雨变化对森林生态系统影响的长期模拟研究应该考虑物种对干旱的不同响应策略. ...
Contrasting impacts of continuous moderate drought and episodic severe droughts on the aboveground-biomass increment and litterfall of three coexisting Mediterranean woody species 2 2015
... 森林植被受降雨格局变化的影响程度往往与不同树种的干旱响应策略密切相关(Attia et al., 2015; Liu et al., 2015).例如同一地区,非等渗植物(anisohydric species)的生长受干旱加剧的影响相对较小(Attia et al., 2015; Liu et al., 2015).在我国半干旱地区, 李亮等(2011)发现不耐旱植物核桃楸(Juglans mandshurica)的生物量在长期干旱影响下并没有降低, 而耐旱植物辽东栎(Quercus liaotungensis)受长期干旱影响时生物量显著下降.Attia等(2015)认为, 干旱发生时, 在不同的环境及资源条件下, 植物往往会采取3种不同的响应策略来应对水分胁迫: (1)高存活率, 低生长率; (2)高耐受性, 适度生长; (3)低存活率, 高生长率.这种种间差异会导致不同森林类型间对降雨格局变化的响应的异质性.如地中海常绿针叶林对干旱的耐受性远低于湿润的温带落叶阔叶林(Gazol et al., 2018).因此, 降雨变化对森林生态系统影响的长期模拟研究应该考虑物种对干旱的不同响应策略. ...
... ; Liu et al., 2015).在我国半干旱地区, 李亮等(2011)发现不耐旱植物核桃楸(Juglans mandshurica)的生物量在长期干旱影响下并没有降低, 而耐旱植物辽东栎(Quercus liaotungensis)受长期干旱影响时生物量显著下降.Attia等(2015)认为, 干旱发生时, 在不同的环境及资源条件下, 植物往往会采取3种不同的响应策略来应对水分胁迫: (1)高存活率, 低生长率; (2)高耐受性, 适度生长; (3)低存活率, 高生长率.这种种间差异会导致不同森林类型间对降雨格局变化的响应的异质性.如地中海常绿针叶林对干旱的耐受性远低于湿润的温带落叶阔叶林(Gazol et al., 2018).因此, 降雨变化对森林生态系统影响的长期模拟研究应该考虑物种对干旱的不同响应策略. ...
Exploring the likelihood and mechanism of a climate-?change-induced dieback of the Amazon rainforest 1 2009
... 人类活动正深远地影响着全球的水文过程, 导致降水格局发生剧烈的变化(Seneviratne et al., 2006; 吴茜等, 2011; Kuang et al., 2017).在中国亚热带地区, 这种趋势主要表现为干季降水频率减少、雨季强降雨事件频发(Parry et al., 2007; 董思言和高学杰, 2014).其后果主要在于改变土壤含水量(Weltzin et al., 2003; 周洁等, 2013), 影响与植物水分利用相关的形态、结构和功能(吴茜等, 2011).尽管亚热带地区降水丰沛, 土壤含水量较高, 但干湿分明的特点使得植物仍然会受到周期性季节干旱的扰动.而在降水格局变化影响下, 这种干湿差异性可能会更加明显(Zhou et al., 2013).亚热带森林是中国森林生态系统的主要组成部分, 其微弱变化也可能对区域气候带来巨大的影响.然而目前对于当前这种降水格局变化究竟会对亚热带森林带来怎样的影响, 我们知之甚少. ...
Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain 1 2011
... 一般来讲, 干旱加剧会引起叶片水势和气孔导度下降(Wang et al., 2003), 并造成乔木生长速率大幅度降低(Klos et al., 2009; Zlatev & Lidon, 2012), 这种现象普遍存在于干旱半干旱区(李亮等, 2011;吴旭等, 2015)、半湿润区(Pasho et al., 2011), 甚至湿润的热带亚热带地区(Samanta et al., 2010; Vicente- Serrano et al., 2013).例如在我国西南地区, 张中峰等(2012)发现, 在干旱胁迫下青冈(Cyclobalanopsis glauca)二年生幼苗光合速率和生长速率均显著降低.胡义等(2014)发现, 干旱胁迫下樟(Cinnamomum camphora)幼树的地径、树高生长量受到了抑制.然而这一结论在其他研究中并没有得到证实.例如最近研究发现, 当降水减少30%时, 古田山地区5个演替早期树种的生长均没有受到显著影响(吴茜等, 2011).孙谷畴等(2009)认为, 在我国亚热带地区, 演替早期树种能够在较强的水分胁迫下保持较高的气孔导度, 从而保证植物依然能够维持较高的光合速率(Attia et al., 2015).而演替后期树种, 即使叶片水势相对较高, 光合速率依然会大幅度下降(孙谷畴等, 2009).然而以上研究的实验对象往往是幼苗和幼树, 不能表征成熟森林的响应方式(吴茜等, 2011).而叶片尺度的植物生理测定由于具有较大的局限性, 很有可能会得到片面的结论(Andrade et al., 1998), 因此必须在整树尺度上对其进行验证(赵平, 2011; 吴旭等, 2015). ...
Amazon forests did not green-up during the 2005 drought 1 2010
... 一般来讲, 干旱加剧会引起叶片水势和气孔导度下降(Wang et al., 2003), 并造成乔木生长速率大幅度降低(Klos et al., 2009; Zlatev & Lidon, 2012), 这种现象普遍存在于干旱半干旱区(李亮等, 2011;吴旭等, 2015)、半湿润区(Pasho et al., 2011), 甚至湿润的热带亚热带地区(Samanta et al., 2010; Vicente- Serrano et al., 2013).例如在我国西南地区, 张中峰等(2012)发现, 在干旱胁迫下青冈(Cyclobalanopsis glauca)二年生幼苗光合速率和生长速率均显著降低.胡义等(2014)发现, 干旱胁迫下樟(Cinnamomum camphora)幼树的地径、树高生长量受到了抑制.然而这一结论在其他研究中并没有得到证实.例如最近研究发现, 当降水减少30%时, 古田山地区5个演替早期树种的生长均没有受到显著影响(吴茜等, 2011).孙谷畴等(2009)认为, 在我国亚热带地区, 演替早期树种能够在较强的水分胁迫下保持较高的气孔导度, 从而保证植物依然能够维持较高的光合速率(Attia et al., 2015).而演替后期树种, 即使叶片水势相对较高, 光合速率依然会大幅度下降(孙谷畴等, 2009).然而以上研究的实验对象往往是幼苗和幼树, 不能表征成熟森林的响应方式(吴茜等, 2011).而叶片尺度的植物生理测定由于具有较大的局限性, 很有可能会得到片面的结论(Andrade et al., 1998), 因此必须在整树尺度上对其进行验证(赵平, 2011; 吴旭等, 2015). ...
Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees 1 2004
... 在自然状态下, 当PAR > 1 100 μmol·m-2·s-1时, 火力楠的¯JS ((49.5 ± 1.7) mL∙m-2∙s-1)显著高于木荷((43.6 ± 2.0) mL∙m-2∙s-1), 并且当光照强度低时, 前者对PAR的增加具有更强的敏感性(图2), 这种差异性可能与两者的边材导水能力有关(吴旭等, 2015).尽管木荷和火力楠均为典型的散孔材, 但木荷的边材密度显著高于火力楠(Gao et al., 2017).以往的研究表明, 边材密度与最大导水率以及光合速率之间存在显著的负相关关系(Santiago et al., 2004; Hoeber et al., 2014).这种差异性会导致不同的干旱响应策略(Hoffmann et al., 2011).在我们的结果中, 尽管木荷与火力楠在受水分限制时的¯JS均具有相似的变化模式, 但火力楠¯JS的下降(干季1下降6%)和回升(干季2增加6%)幅度均小于木荷(干季1下降8.1%, 干季2增加25.7%, 干季3增加40.5%), 这表明木荷相对于火力楠具有更强的形态可塑性.事实上, Kuang等(2017)在对我们样地中这两种植物的研究中已经发现, 木荷在遮雨处理下, 在形态结构等方面表现出更强的可塑性和适应性, 尤其是根系长度和根系生物量.而我们的结果也表明, 在长期的干旱处理后, 木荷的¯JS对PAR和VPD的敏感性都高于火力楠(图5), 这与自然状态下两者的¯JS敏感性截然相反(图5).这说明与火力楠相比, 干旱发生时, 木荷更加倾向于非等渗行为(Attia et al., 2015), 这一水力特征保证植物在水分胁迫条件下依然能够维持较高的光合速率(孙谷畴等, 2009).因此, 我们在研究环境变化对森林植被的影响时, 即使是相同生境条件下, 不同树种间的水力响应策略差异也可能会对研究结果产生影响(Babst et al., 2013). ...
Land-atmosphere coupling and climate change in Europe 1 2006
... 人类活动正深远地影响着全球的水文过程, 导致降水格局发生剧烈的变化(Seneviratne et al., 2006; 吴茜等, 2011; Kuang et al., 2017).在中国亚热带地区, 这种趋势主要表现为干季降水频率减少、雨季强降雨事件频发(Parry et al., 2007; 董思言和高学杰, 2014).其后果主要在于改变土壤含水量(Weltzin et al., 2003; 周洁等, 2013), 影响与植物水分利用相关的形态、结构和功能(吴茜等, 2011).尽管亚热带地区降水丰沛, 土壤含水量较高, 但干湿分明的特点使得植物仍然会受到周期性季节干旱的扰动.而在降水格局变化影响下, 这种干湿差异性可能会更加明显(Zhou et al., 2013).亚热带森林是中国森林生态系统的主要组成部分, 其微弱变化也可能对区域气候带来巨大的影响.然而目前对于当前这种降水格局变化究竟会对亚热带森林带来怎样的影响, 我们知之甚少. ...
Gas exchange recovery following natural drought is rapid unless limited by loss of leaf hydraulic conductance: Evidence from an evergreen woodland 1 2017
... 植物的生长速率往往取决于冠层叶片蒸腾耦合的气体交换的速率(Catovsky et al., 2002; Skelton et al., 2017).因此在干旱条件下, 植物蒸腾的响应能够在较大程度上反映干旱对植物生长的影响.树干液流作为植物最重要的一种生理活动, 反映了植物水分传输能力, 因而经常被用来研究环境变化对植物生长的影响(Graham et al., 2003; Steppe et al., 2006).为了检验降雨格局变化对我国亚热带常绿阔叶林的影响,我们在亚热带地区典型演替早期天然次生林中选择木荷(Schima superba)和火力楠(Michelia macclurei)为研究对象, 开展人工隔除林下降雨实验模拟降雨格局变化.木荷(SS)为亚热带地区演替早期优势树种, 火力楠(MM)则为伴生其中的亚优势种.在木材解剖结构上, 两者均属于散孔材树种和深根系植物(蔡锡安等, 2012; 张振振等, 2014, 2019), 且生长迅速, 为理想的造林树种.通过对两者树干液流的监测, 我们探讨加剧的季节性干旱对演替早期优势树种水分传输的影响.具体来讲,我们主要关注以下两个问题: (1)在降雨量较为丰沛的亚热带地区, 林下降雨隔除对该地区的树干液流是否会带来负效应? (2)不同树种树干液流对干旱的响应方式是否存在差异? 这一研究将给气候变化背景下森林群落演替的变化规律研究提供参考. ...
A mathematical model linking tree sap flow dynamics to daily stem diameter fluctuations and radial stem growth 1 2006
... 植物的生长速率往往取决于冠层叶片蒸腾耦合的气体交换的速率(Catovsky et al., 2002; Skelton et al., 2017).因此在干旱条件下, 植物蒸腾的响应能够在较大程度上反映干旱对植物生长的影响.树干液流作为植物最重要的一种生理活动, 反映了植物水分传输能力, 因而经常被用来研究环境变化对植物生长的影响(Graham et al., 2003; Steppe et al., 2006).为了检验降雨格局变化对我国亚热带常绿阔叶林的影响,我们在亚热带地区典型演替早期天然次生林中选择木荷(Schima superba)和火力楠(Michelia macclurei)为研究对象, 开展人工隔除林下降雨实验模拟降雨格局变化.木荷(SS)为亚热带地区演替早期优势树种, 火力楠(MM)则为伴生其中的亚优势种.在木材解剖结构上, 两者均属于散孔材树种和深根系植物(蔡锡安等, 2012; 张振振等, 2014, 2019), 且生长迅速, 为理想的造林树种.通过对两者树干液流的监测, 我们探讨加剧的季节性干旱对演替早期优势树种水分传输的影响.具体来讲,我们主要关注以下两个问题: (1)在降雨量较为丰沛的亚热带地区, 林下降雨隔除对该地区的树干液流是否会带来负效应? (2)不同树种树干液流对干旱的响应方式是否存在差异? 这一研究将给气候变化背景下森林群落演替的变化规律研究提供参考. ...
亚热带森林演替树种叶片气孔导度对环境水分的水力响应 3 2009
... 一般来讲, 干旱加剧会引起叶片水势和气孔导度下降(Wang et al., 2003), 并造成乔木生长速率大幅度降低(Klos et al., 2009; Zlatev & Lidon, 2012), 这种现象普遍存在于干旱半干旱区(李亮等, 2011;吴旭等, 2015)、半湿润区(Pasho et al., 2011), 甚至湿润的热带亚热带地区(Samanta et al., 2010; Vicente- Serrano et al., 2013).例如在我国西南地区, 张中峰等(2012)发现, 在干旱胁迫下青冈(Cyclobalanopsis glauca)二年生幼苗光合速率和生长速率均显著降低.胡义等(2014)发现, 干旱胁迫下樟(Cinnamomum camphora)幼树的地径、树高生长量受到了抑制.然而这一结论在其他研究中并没有得到证实.例如最近研究发现, 当降水减少30%时, 古田山地区5个演替早期树种的生长均没有受到显著影响(吴茜等, 2011).孙谷畴等(2009)认为, 在我国亚热带地区, 演替早期树种能够在较强的水分胁迫下保持较高的气孔导度, 从而保证植物依然能够维持较高的光合速率(Attia et al., 2015).而演替后期树种, 即使叶片水势相对较高, 光合速率依然会大幅度下降(孙谷畴等, 2009).然而以上研究的实验对象往往是幼苗和幼树, 不能表征成熟森林的响应方式(吴茜等, 2011).而叶片尺度的植物生理测定由于具有较大的局限性, 很有可能会得到片面的结论(Andrade et al., 1998), 因此必须在整树尺度上对其进行验证(赵平, 2011; 吴旭等, 2015). ...
 ,1,*, 杨轲嘉1, 顾宇璐1, 赵平2, 欧阳磊2
,1,*, 杨轲嘉1, 顾宇璐1, 赵平2, 欧阳磊2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT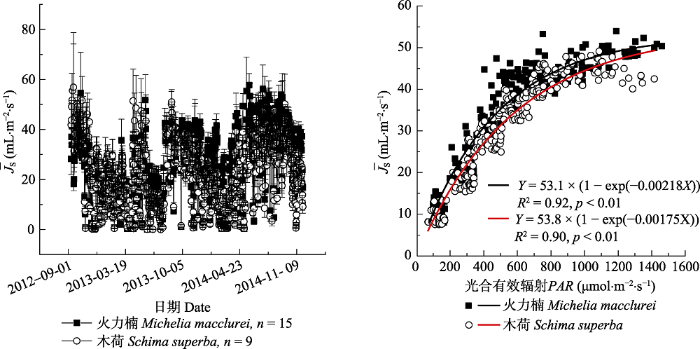 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT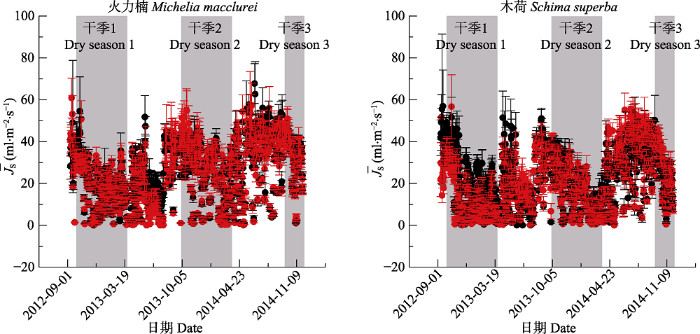 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT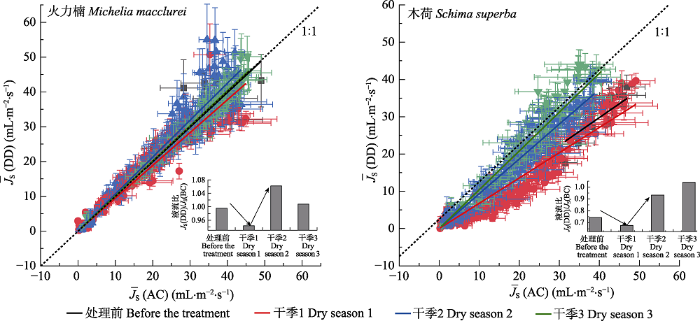 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT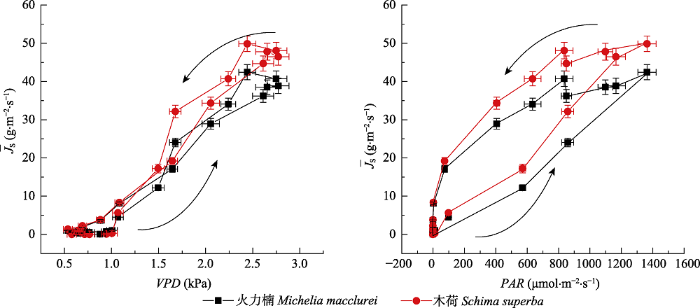 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}