 ,*东北林业大学林学院, 东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040
,*东北林业大学林学院, 东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040Carbon storage and its allocation in Betula platyphylla forests of different ages in cold temperate zone of China
Hong WEI, Xiu-Ling MAN,*School of Forestry, Northeast Forestry University, Key Laboratory of Sustainable Forest Ecosystem Management-Ministry of Education, Northeast Forestry University, Harbin 150040, China通讯作者: 通讯作者Author for correspondence (E-mail:jshe@pku.edu.cn)
编委: 李敏
收稿日期:2019-05-28接受日期:2019-10-6网络出版日期:2019-10-20
| 基金资助: |
Received:2019-05-28Accepted:2019-10-6Online:2019-10-20
| Fund supported: |

摘要
为了解中国寒温带地区不同林龄白桦林生态系统碳储量及固碳能力, 在样地调查基础上, 以大兴安岭地区25、40与61年白桦(Betula platyphylla)林生态系统为研究对象, 对其乔木层、林下地被物层(灌木层、草本层、凋落物层)、土壤层(0-100 cm)碳储量与分配特征进行调查研究。结果表明白桦林乔木层各器官碳含量在440.7-506.7 g·kg -1之间, 各器官碳含量随着林龄的增长而降低; 灌木层、草本层碳含量随林龄的增加呈先降后升的变化趋势; 凋落物层碳含量随林龄增加而降低; 土壤层(0-100 cm)碳含量随林龄增加而显著升高, 随着土层深度的增加而降低。白桦林生态系统各层次碳储量均随林龄的增加而明显升高。25、40与61年白桦林乔木层碳储量分别为11.9、19.1和34.2 t·hm -2, 各器官碳储量大小顺序表现为树干>树根>树枝>树叶, 树干碳储量分配比例随林龄增加而升高。25、40与61年白桦林生态系统碳储量分别为77.4、180.9和271.4 t·hm -2, 其中土壤层占生态系统总碳储量的81.6%、87.7%和85.9%, 是白桦林生态系统的主要碳库。随林龄增加, 白桦林年净生产力(2.0-4.4 t·hm -2·a -1)、年净固碳量(1.0-2.1 t·hm -2·a -1)均出现增长, 老龄白桦林仍具有较强的碳汇作用。
关键词:
Abstract
Aims The objective of this study was to estimate the carbon storage and its allocation in the Betula platyphylla forests of three different ages (25, 40, 61-year-old) in the cold temperate zone, NE China. Methods Through analyzing the field data, we estimated the carbon storage and sequestration rates of tree, understory layer (shrub layer, herb layer, litter layer) and soil layer (0-100 cm) of the 25, 40 and 61-year-old Betula platyphylla ecosystems in the north section of the Da Hinggan Ling Mountains. Important findings The results showed that the carbon content of each organ in the tree layer of the forests ranged from 440.7 to 506.7 g·kg-1, that decreased as the forest age increases. Carbon content in the shrub and herb layers decreased first and then increased as the forest aged, while that in the litter layer decreased with the increase of forest age. The carbon content in soil layer (0-100 cm) increased significantly with the forest age (p < 0.05), and decreased as soil drought intensified. The carbon storage of B. platyphylla ecosystem at all levels increased significantly with the increase of forest age. The carbon storage of tree layer in the forests of 25, 40 and 61-year-old were 11.9, 19.1 and 34.2 t·hm-2, respectively. The carbon storage of the organs follow the order of: trunk > root > branch > leaf, and the allocation ratio of trunk carbon increased as the forest aged. The carbon storage in the Betula platyphylla forest ecosystems of 25, 40 and 61-year-old were 77.4, 180.9 and 271.4 t·hm -2, respectively. Soil layer, the main carbon pool of the ecosystems, accounted for 81.6%, 87.7% and 85.9% of the total carbon storage. The annual net productivity (2.0-4.4 t·hm -2·a -1) and annual net carbon sequestration (1.0-2.1 t·hm -2·a -1) of the forests increased with the age increase of the forest, and the old-growth B. platyphylla forests hold a strong carbon sequestration capacity.
Keywords:
PDF (1082KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
魏红, 满秀玲. 中国寒温带不同林龄白桦林碳储量及分配特征. 植物生态学报, 2019, 43(10): 843-852. DOI: 10.17521/cjpe.2019.0127
WEI Hong, MAN Xiu-Ling.
由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(Wang et al., 2012), 实施碳减排、增加碳汇来应对气候变化的影响已成为国际社会的共识(吴贤荣等, 2014)。森林生态系统作为陆地上最大的碳库, 对全球碳循环及碳平衡至关重要(Pan et al., 2011), 因此森林碳储量的估算成为适应全球变化和增汇减排机制研究的重要内容(方精云等, 2001)。森林生态系统碳储量受气候因素、立地条件、森林类型、森林经营管理、林龄结构的影响较大(Fang et al., 2001; Chen & Luo, 2015), 其中林龄对森林碳储量的影响尤为重要(Zhu et al., 2017a)。目前已有很多****针对森林碳储量及分配格局随林龄的变化进行了研究(明安刚等, 2014; 胡海清等, 2015; 张毅等, 2015; Zhu et al., 2017b), 结果表明, 在一定年龄范围内随着林龄的增加, 植被碳储量通常呈增加状态, 土壤碳储量由于更易受到土地利用方式、气候及植物种类等因素的影响, 因此随着林龄的增加土壤碳储量的变化较为不确定(Li et al., 2011; 韩营营等, 2015)。但目前研究多集中于我国亚热带及温带地区(周序力等, 2018), 有关我国寒温带森林生态系统碳储量及分配格局随林龄变化特征的研究鲜有报道。
寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(鲁博权等, 2019), 对该区域植被生长与养分循环存在巨大影响, 白桦(Betula platyphylla)是我国寒温带地区主要树种, 占本区森林总面积和总蓄积量的41.15%和41.59% (刘双娜等, 2012; Girardin et al., 2016; Zhou et al., 2019), 具有重要的生态地位。目前有关白桦林的研究多集中于其生长发育(兰士波, 2018)、土壤特性(李文影等, 2009)、水化学(刘茜等, 2015)、生物量(李建强等, 2010)、植被层碳储量(胡海清等, 2015)和土壤碳通量(韩营营等, 2015)等, 研究地点多位于温带地区, 且多以中龄白桦林为研究对象(李娜娜等, 2015)。本文选择我国寒温带地区25、40和61年白桦林为研究对象, 通过野外调查与室内试验相结合的方法, 研究白桦林生态系统碳储量及分配特征, 既可以丰富我国寒温带地区森林生态系统碳汇研究, 也可为区域尺度精确测算白桦林生态系统碳储量及碳汇功能提供基础数据和科学参考。
1 材料和方法
1.1 研究区概况与样地设置
研究区位于黑龙江漠河森林生态系统国家定位观测研究站(122.10°-122.45° E, 53.28°-53.50° N)内, 该区属于寒温带大陆性季风气候, 冬季气候寒冷、干燥而漫长; 夏季气候湿热, 降水集中, 雨量充沛。该区年平均气温-4.9 ℃, 平均无霜期为89天, 年降水量为350-500 mm, 是多年冻土分布区。主要森林类型有兴安落叶松(Larix gmelinii)林、白桦林、樟子松(Pinus sylvestris var. mongolica) 林和混交林等。林下灌木主要有兴安杜鹃(Rhododendron dauricum)、杜香(Ledum palustre)、越桔(Vaccinium vitis- idaea)等, 草本植物有小叶章(Deyeuxia angustifolia)、北柴胡(Bupleurum chinense)、肾叶鹿蹄草(Pyrola renifolia)、地榆(Sanguisorba officinalis)等。地带性土壤为棕色针叶林土, 还分布有草甸土和沼泽土等土壤类型。在踏查基础上, 选取该区域占比较高的(25 ± 6)、(40 ± 4)和(61 ± 7)年白桦林为研究对象(以下简称25、40和61 a), 按照中华人民共和国林业行业标准(LY/T2908-2017)并结合大兴安岭白桦林生长特点(胡海清等, 2015), 将这3种不同林龄白桦林划归为幼龄林、中龄林和老龄林。在25、40和61 a白桦林内选择立地条件基本一致的典型区域, 各随机布设3块20 m × 20 m样地, 对每个样地进行每木检尺, 样地基本概况见表1。
1.2 白桦林生态系统生物量调查
1.2.1 乔木层乔木生物量的估算采用相对生长法, 对样地内胸径≥4 cm乔木进行每木检尺, 将树高、胸径的实测结果代入表2马林(2012)研究出的白桦单木生物量模型, 计算各器官生物量, 汇总得出单位面积乔木生物量。
1.2.2 林下地被物层
1.2.2.1 灌木层 在每个样地内, 沿对角线随机设置3个2 m × 2 m小样方, 记录小样方内灌木的种类与株数, 采用“全收获法”按照地上、地下两部分采集小样方内全部灌木, 将同种植物相同部分混合, 在野外称量各部分鲜质量。随后选取部分样品带回实验室, 85 ℃烘干至恒质量(间隔6 h两次称量所得的含水率差小于0.1%即被视为达到恒质量), 用精度为0.01 g的电子天平称干质量, 计算地上、地下部分的生物量, 两部分生物量相加计算出单位面积生物量(张治军等, 2009)。
Table 1
表1
表1大兴安岭北部不同林龄白桦林样地基本概况(平均值±标准偏差)
Table 1
| 林龄 Stand age (a) | 坡度 Slope (°) | 密度 Stand density (No.·hm-2) | 郁闭度 Canopy density | 平均树高 Mean tree height (m) | 平均胸径 Mean DBH (cm) | 林下主要植物 Main understory plants |
|---|---|---|---|---|---|---|
| 25 ± 6 | 3 ± 1 | 1 825 ± 225 | 0.7 ± 0.1 | 8.6 ± 1.6 | 7.1 ± 1.8 | 1, 2, 3, 5, 6, 8 |
| 40 ± 4 | 2 ± 1 | 2 075 ± 300 | 0.9 ± 0.1 | 11.1 ± 3.0 | 9.5 ± 1.8 | 1, 2, 4, 5, 7, 9 |
| 61 ± 7 | 6 ± 2 | 1 820 ± 250 | 0.8 ± 0.1 | 15.2 ± 3.5 | 12.4 ± 2.2 | 1, 2, 4, 5, 6, 7, 8 |
新窗口打开|下载CSV
Table 2
表2
表2白桦生物量方程
Table 2
| 器官 Organ | 回归方程 Regression equation | R2 |
|---|---|---|
| 树干 Stem | W = 0.046D1.55H1.163 | 0.969 |
| 树枝 Branch | W = 5.921D1.925H-1.54 | 0.793 |
| 树叶 Leaf | W = 0.012D1.662H0.49 | 0.711 |
| 树根 Root | W = 0.016D2.171H0.281 | 0.933 |
新窗口打开|下载CSV
1.2.2.2 草本层 在每个样地内, 沿对角线随机设置5个1 m × 1 m小样方, 调查样方内草本植物的盖度、种类和平均高度, 按照地上、地下两部分采集小样方内全部草本植物, 将同种植物相同部分混合, 在野外称量各部分鲜质量。测定方法与灌木层生物量测定方法相同。
1.2.2.3 凋落物层 在每一样地内, 按对角线设置5个50 cm × 50 cm凋落物样方, 按照未分解层与半分解层收集样方内凋落物, 相同层凋落物混合, 野外测定湿质量。测定方法同灌木生物量测定方法。
1.3 白桦林生态系统碳储量测定
1.3.1 乔木层在各样地内选取白桦标准木, 共选取标准木10株(25 a林分4株, 40 a林分3株, 61 a林分3株), 采集标准木的树干、树枝、树叶和根, 其中树干分别从树干基部、胸径和梢头3个部位进行取样, 枝带皮从粗枝到小枝按比例取样, 叶亦分别从不同部位取样, 根带皮从粗根到小根按比例分别取样。带回实验室, 放置于85 ℃烘箱, 烘干至恒质量, 粉碎, 过筛, 用于碳含量的测定。将表1所得到的白桦林每木检尺数据带入表2回归方程中可得到白桦乔木层干、枝、叶、根每hm2的生物量, 再乘以各器官碳含量, 即为乔木层干、枝、叶、根每hm2的碳储量。
1.3.2 林下地被物层
灌木层每hm2碳储量为各部分碳含量与对应部分每hm2生物量的乘积。草本层与凋落物层每hm2碳储量计算方法同上。
林下地被物层碳储量为各层次每hm2碳储量之和。
1.3.3 土壤层
在各样地内随机挖取3个土壤剖面, 按照0-10、10-20、20-30、30-50、50-100 cm分层取样, 并分层取环刀, 测定容重。将土壤样品带回实验室, 自然风干, 研磨过筛, 使用Elemental Vario TOC cube型Hanau, 元素分析仪(Elemental, Hanau, Germany)测定土壤内有机碳含量。土壤剖面有机碳储量的计算模型如下:
式中, SOCD为土壤剖面有机碳储量(t·hm-2); θi为第i层砾石含量(体积%); ρi为第i层土壤容重(g·cm-3); Ci为第i层土壤有机碳含量(C g·kg-1); Ti为第i层土层厚度(cm)。
1.4 数据处理
叶片的净生产量按照现存量直接转换(郑瞳等, 2016), 其他部分均按照白桦林龄, 结合年净生产量公式计算:式中, ΔW为年净生产量(t·hm-2·a-1), Wm为m年时单位面积现存生物量(t·hm-2), Wm-n为(m-n)年时单位面积现存生物量(t·hm-2), n为年数(a)(丁贵杰和王鹏程, 2002)。
采用SPSS 19.0软件据进行统计分析、方差分析和差异性显著检验(ɑ = 0.05), 采用Excel 2016软件作图, 图表中数据为平均值±标准偏差。
2 结果和分析
2.1 不同林龄白桦林生态系统各组分碳含量
2.1.1 乔木层碳含量由表3可知, 不同林龄白桦林乔木层各器官碳含量在440.7-506.7 g·kg-1, 林龄对乔木层各器官碳含量均存在显著影响(p < 0.05), 随着林龄的增加乔木层各器官碳含量均呈现下降趋势。其中树干碳含量降幅最大, 从490.3减至440.7 g·kg-1, 树枝碳含量降幅最小, 从485.9减至453.6 g·kg-1。不同器官碳含量及大小顺序存在差异, 但均以树叶最高。
2.1.2 林下地被物层碳含量
林下地被物包括活地被物与死地被物, 本研究主要测定了白桦林林下灌木层、草本层与凋落物层的碳含量。由表3可知, 随着林龄的增加, 灌木层与草本层碳含量先降低后升高, 表现为61 a > 25 a > 40 a, 凋落物层的碳含量随着林龄的增加而下降, 表现为25 a (515.2 g·kg-1) > 40 a (483.1 g·kg-1) > 61 a (426.4 g·kg-1), 不同林龄间差异显著(p < 0.05)。25 a与40 a白桦林林下地被物层的碳含量表现为凋落物层>灌木层>草本层, 61 a白桦林表现为灌木层>草本层>凋落物层。灌木层、草本层地上部分碳含量高于地下部分, 凋落物未分解层的碳含量高于半分解层。
2.1.3 土壤层碳含量
从图1可知, 不同林龄白桦林土壤层碳含量为1.3-77.0 g·kg-1, 同一土层不同林龄间碳含量存在显著差异(p < 0.05), 随着林龄的增加土壤碳含量升高, 表现为: 61 a > 40 a > 25 a。不同林龄白桦林土壤层碳含量具有明显垂直分布特征, 表层(0-10 cm)土壤碳含量最高, 分别为19.9、42.9和77.0 g·kg-1, 随着土层深度的增加土壤碳含量明显下降。0-30 cm土层碳含量显著高于下层土壤, 随着土层深度的继续增加, 土壤碳含量继续下降, 但不同土层间差异逐渐减小。
2.2 白桦林生态系统各组分碳储量及分配特征
2.2.1 乔木层生物量、碳储量及分配特征由表3可知, 随着林龄的增加, 乔木层各器官生物量均呈显著增加趋势(p < 0.05), 25、40与61 a白桦林乔木层生物量分别为24.1、41.5和76.5 t·hm-2, 树干、树根生物量分配比例随着林龄的增加而增加(48.5%-70.6%和8.7%-10.6%), 树枝生物量分配比例随着林龄的增加而降低(39.0%-14.9%), 树叶生物量分配比例较为稳定, 随林龄的增加未表现出明显变化(3.7%-4.1%)。乔木层各器官碳储量随林龄变化趋势与生物量较为接近, 随着林龄的增加各器官碳储量呈现增加趋势, 25、40和61 a白桦林乔木层碳储量分别为11.9、19.1和34.2 t·hm-2。各器官碳储量排序为树干(48.7%-69.6%) >树枝(38.6%-15.2%) >树叶(3.8%-4.2%) >树根(8.4%-11.1%)。林龄对树干、树枝、树根碳储量分配比例存在显著影响, 随着林龄增加树干碳储量分配比例逐渐增加, 树枝和树根碳储量分配比例随着林龄的增加而降低, 随着林龄增加树叶碳储量分配比例未发现明显变化。
2.2.2 林下地被物碳储量及分配特征
由表3可知, 林下地被物层碳储量较低, 不同林龄白桦林下地被物层碳储量分别为2.3 t·hm-2、3.2 t·hm-2和4.0 t·hm-2, 随着林龄增加而升高。灌木层、草本层碳储量随着林龄的增加先降低后升高, 凋落物层碳储量随着林龄的增加而升高。
25a白桦林林下地被物层碳储量表现为灌木层>凋落物层>草本层, 40 a与61 a白桦林表现为凋落物层>灌木层>草本层。灌木层与草本层地上部分碳储量高于地下部分, 凋落物半分解层碳储量高于未分解层, 差异均达到显著水平(p < 0.05)。
2.2.3 土壤层碳储量及分配特征
由图2可知, 白桦林土壤碳储量随林龄的增加呈增长趋势, 25、40和61 a白桦林0-100 cm土层碳储量分别为63.2、158.6和233.2 t·hm-2, 林龄对不同土层土壤碳储量均存在显著影响(p < 0.05)。各林龄白桦林土壤碳储量均随着土层深度的增加表现出下降趋势, 土壤碳主要集中于0-30 cm土层, 其碳分配比例分别为71.1%、78.9%和65.3%, 30-100 cm土层土壤碳储量较低。
2.3 白桦林固碳能力
由表4可知, 白桦林年净生产力、年固碳量均随着林龄的增加而显著升高, 25、40和61 a白桦林的年净生产力分别为2.0、3.0和4.4 t·hm-2·a-1, 年固碳量分别为1.0、1.8和2.1 t·hm-2·a-1。其中乔木层固碳能力最强, 且随着林龄增长乔木层固碳量逐渐增多, 分别为0.9、1.3和2.0 t·hm-2·a-1, 占生态系统总固碳量的90.0%、72.2%和95.2%, 且由中龄至老龄阶段 的增长幅度(53.8%)高于幼龄至中龄阶段(44.4%)。不同林龄白桦林乔木层各组分年固碳量的排序大致为树叶(55.6%-70.0%) >树干(20.0%-23.1%) >树枝(5.0%-22.2%) >树根(0-7.7%)。随着林龄的增加, 乔木层树叶年固碳量占比逐渐增大, 其余组分年固碳量占比均逐渐减小。林下植被层的年固碳量分别为0.2、0.5和0.1 t·hm-2·a-1, 即40 a白桦林林下地被物层固碳能力最强, 61 a最弱。Table 3
表3
表3大兴安岭北部不同林龄白桦林各组分的生物量(t·hm-2)、碳含量(g·kg-1)和碳储量(t·hm-2)(平均值±标准偏差)
Table 3
| 组分 Component | 25 a | 40 a | 61 a | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 生物量 Biomass (t·hm-2) | 碳含量 Carbon Content (g·kg-1) | 碳储量 Carbon Storage (t·hm-2) | 生物量 Biomass (t·hm-2) | 碳含量 Carbon content (g·kg-1) | 碳储量 Carbon storage (t·hm-2) | 生物量 Biomass (t·hm-2) | 碳含量 Carbon content (g·kg-1) | 碳储量 Carbon storage (t·hm-2) | ||
| 乔木层 Arbor layer | 树干 Stem | 11.7 ± 0.6Ac | 490.3 ± 8.8 Ba | 5.8 ± 0.5Ac | 24.8 ± 1.2Ab | 453.3 ± 8.7Bb | 11.2 ± 0.6Ab | 54.0 ± 2.7Aa | 440.7 ± 4.7Db | 23.8 ± 0.7Aa |
| 树枝 Branch | 9.4 ± 0.5Bb | 485.9 ± 8.4Ca | 4.6 ± 0.4Bb | 11.1 ± 0.5Ba | 473.7 ± 3.6Aa | 5.3 ± 0.3Ba | 11.4 ± 0.1Ba | 453.6 ± 6.8Cb | 5.2 ± 0.2Ba | |
| 树叶 Leaf | 0.9 ± 0.1Fc | 506.7 ± 7.3Ba | 0.5 ± 0.0Gc | 1.7 ± 0.1Db | 484.2 ± 16.0Ab | 0.8 ± 0.0Eb | 3.0 ± 0.2Da | 467.9 ± 9.7Cb | 1.4 ± 0.0Ea | |
| 树根 Root | 2.1 ± 0.1Cc | 505.9 ± 11.0Ba | 1.0 ± 0.1Cc | 3.9 ± 0.2Cb | 471.6 ± 12.5Ab | 1.8 ± 0.1Cb | 8.1 ± 0.5Ca | 465.6 ± 12.6Cb | 3.8 ± 0.1Ca | |
| 总计 Sum | 24.1 ± 1.1c | 11.9 ± 1.0c | 41.5 ± 2.0b | 19.1 ± 1.0b | 76.5 ± 3.5a | 34.2 ± 1.0a | ||||
| 灌木层 Shrub layer | 地上部分 Aboveground | 1.5 ± 0.1Da | 505.3 ± 1.7Bb | 0.8 ± 0.1Db | 1.0 ± 0.1Eb | 494.3 ± 14.9Ab | 0.5 ± 0.1Fc | 1.5 ± 0.1Ea | 575.3 ± 26.6Aa | 0.9 ± 0.2Fa |
| 地下部分 Underground | 1.2 ± 0.1Ea | 483.7 ± 1.9Cb | 0.6 ± 0.1Fb | 0.9 ± 0.0Fb | 465.7 ± 1.3Bc | 0.4 ± 0.1Gc | 1.3 ± 0.1Fa | 551.7 ± 7.5Aa | 0.7 ± 0.1Ga | |
| 总计 Sum | 2.8 ± 0.2a | 1.4 ± 0.2b | 1.8 ± 0.1b | 0.9 ± 0.2c | 2.8 ± 0.2a | 1.6 ± 0.3a | ||||
| 草本层 Herb layer | 地上部分 Aboveground | 0.2 ± 0.0Gb | 426.1 ± 2.9Db | 0.1 ± 0.0Ib | 0.7 ± 0.1Ga | 412.8 ± 2.7Cc | 0.3 ± 0.1Ja | 0.7 ± 0.0Ga | 462.4 ± 7.1ABc | 0.1 ± 0.0Ib |
| 地下部分 Underground | 0.1 ± 0.0Hb | 424.8 ± 1.5Db | 0.0 ± 0.0Jc | 0.5 ± 0.0Ha | 399.8 ± 9.1Cc | 0.2 ± 0.0Ja | 0.5 ± 0.0Ha | 471.5 ± 8.3Ba | 0.0 ± 0.0Jb | |
| 总计 Sum | 0.2 ± 0.0b | 0.1 ± 0.0b | 1.1 ± 0.1a | 0.5 ± 0.1a | 0.2 ± 0.0b | 0.1 ± 0.0b | ||||
| 凋落 物层 Litter layer | 未分解层 Undecomposed layer | 544.0 ± 63.0Aa | 0.1 ± 0.0Hc | 478.1 ± 21.8Ab | 0.3 ± 0.1Hb | 433.0 ± 17.8Dc | 0.6 ± 0.1Ha | |||
| 半分解层 partially- decomposed layer | 486.3 ± 11.7Ca | 0.7 ± 0.1Eb | 488.1 ± 13.3Aa | 1.5 ± 0.2Da | 419.9 ± 2.4Eb | 1.8 ± 0.2Da | ||||
| 总计 Sum | 515.2 | 0.8 ± 0.1c | 483.1 | 1.8 ± 0.3b | 426.4 | 2.3 ± 0.3a | ||||
新窗口打开|下载CSV
图1
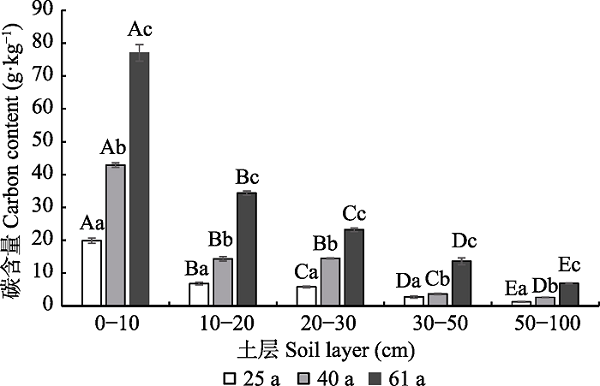 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1大兴安岭北部不同林龄白桦林土壤层碳含量(平均值±标准偏差)。不同大写字母表示同一林龄不同土层间差异显著, 不同小写字母表示同一土层不同林龄间差异显著(p < 0.05)。
Fig. 1Carbon content of soil layer in the Betula platyphylla forests of different ages in the north section of the Da Hinggan Ling Mountains, NE China (mean ± SD). Different capital letters indicate significant differences among different levels of the components of the forest of same age, and different lowercase letters showed significant differences among the same type of component, but different forest ages (p < 0.05).
2.4 白桦林生态系统碳储量的分配特征
由表5可知, 随着林龄增加, 白桦林生态系统碳储量出现明显增长, 25、40与61 a白桦林生态系统碳储量分别为77.4、180.9和271.4 t·hm-2, 不同林龄间差异显著(p < 0.05)。乔木层、林下地被物层、土壤层碳储量均随着林龄的增加而增长, 乔木层碳储量分配比例随着林龄的增加先降低后升高, 分别为15.4%、10.6%和12.6%, 林下地被物层碳储量分配比例随着林龄的增加而降低, 分别为3.0%、1.8%和1.5%, 白桦林生态系统碳储量主要集中于土壤层(81.6%-87.7%), 随着林龄的增加土壤层碳储量分配比例先升高后降低, 分配比例随林龄的变化特征与乔木层相反。图2
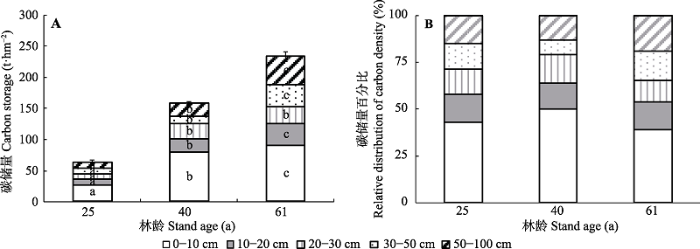 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2大兴安岭北部不同林龄白桦林土壤碳储量绝对值(平均值±标准偏差)(A)和相对值(B)。
Fig. 2Absolute (mean ± SD)(A) and relative distribution (B) of soil carbon density by soil depth in the Betula platyphylla forests of different ages in the north section of the Da Hinggan Ling Mountains, NE China.
Table 4
表4
表4大兴安岭北部白桦林各组分年净生产力和年净固碳量(t·hm-2·a-1)(平均值±标准偏差)
Table 4
| 林龄 Age (a) | 乔木层 Arbor layer | 灌木层 Shrub layer | 草本层 Herb layer | 总计 Total | |||||
|---|---|---|---|---|---|---|---|---|---|
| 树干 Trunk | 树枝 Bark | 树叶 Leaf | 树根 Root | 总计 Total | |||||
| 25 | 年净生产力 Annual net productivity | 0.4 ± 0.0 | 0.3 ± 0.0 | 0.9 ± 0.1 | 0.1 ± 0.0 | 1.7 ± 0.1 | 0.1 ± 0.0 | 0.2 ± 0.0 | 2.0 ± 0.1 |
| 年固碳量 Annual carbon storage | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.5 ± 0.0 | 0.0 ± 0.0 | 0.9 ± 0.1 | 0.1 ± 0.0 | 0.1 ± 0.0 | 1.0 ± 0.1 | |
| 40 | 年净生产力 Annual net productivity | 0.6 ± 0.0 | 0.3 ± 0.0 | 1.7 ± 0.1 | 0.1 ± 0.0 | 1.9 ± 0.1 | 0.0 ± 0.0 | 1.0 ± 0.1 | 3.0 ± 0.3 |
| 年固碳量 Annual carbon storage | 0.3 ± 0.0 | 0.1 ± 0.0 | 0.8 ± 0.0 | 0.1 ± 0.0 | 1.3 ± 0.1 | 0.0 ± 0.0 | 0.5 ± 0.1 | 1.8 ± 0.1 | |
| 61 | 年净生产力 Annual net productivity | 0.9 ± 0.0 | 0.2 ± 0.0 | 3.0 ± 0.2 | 0.1 ± 0.0 | 4.2 ± 0.2 | 0.0 ± 0.0 | 0.2 ± 0.0 | 4.4 ± 0.2 |
| 年固碳量 Annual carbon storage | 0.4 ± 0.0 | 0.1 ± 0.0 | 1.4 ± 0.1 | 0.1 ± 0.0 | 2.0 ± 0.1 | 0.0 ± 0.0 | 0.1 ± 0.0 | 2.1 ± 0.1 | |
新窗口打开|下载CSV
Table 5
表5
表5大兴安岭北部白桦林碳储量(t·hm-2)及分配格局(平均值±标准偏差)
Table 5
| 项目 Item | 林龄 Age (a) | |||||
|---|---|---|---|---|---|---|
| 25 | 40 | 61 | ||||
| 碳储量 Carbon storage | 分配比例 Allocation pattern | 碳储量 Carbon storage | 分配比例 Allocation pattern | 碳储量 Carbon storage | 分配比例 Allocation pattern | |
| 乔木层 Arbor layer | 11.9 ± 1.0 | 15.4% ± 1.4% | 19.1 ± 1.0 | 10.5% ± 0.6% | 34.2 ± 1.0 | 12.6% ± 0.4% |
| 林下地被物层 Understory ground cover layer | 2.3 ± 0.3 | 3.0% ± 0.4% | 3.2 ± 0.5 | 1.8% ± 0.3% | 4.0 ± 0.6 | 1.5% ± 0.2% |
| 土壤层 Soil layer | 63.2 ± 4.1 | 81.6% ± 5.3% | 158.6 ± 3.0 | 87.7% ± 1.7% | 233.2 ± 7.6 | 85.9% ± 2.8% |
| 生态系统 Ecosystem | 77.4 ± 5.4 | 180.9 ± 4.5 | 271.4 ± 9.2 | |||
新窗口打开|下载CSV
3 讨论
3.1 乔木层碳储量
林龄是影响森林生态系统碳储量的重要因素(Pregitzer & Euskirchen, 2004), 我国温带及热带地区森林生态系统中普遍存在着随林龄增加其碳储量升高的变化趋势(Zhu et al., 2017b; 周序力等, 2018)。本研究结果发现我国寒温带地区白桦林生态系统乔木层碳储量亦随着林龄的增加而显著升高(p < 0.05), 25、40和61 a白桦林乔木层碳储量分别为11.9、19.1和34.2 t·hm-2。本研究结果低于六盘山51 a白桦林(41.80 t·hm-2)(杨丽丽等, 2015)和长白山地区35 a白桦林(40.81-80.08 t·hm-2)(李娜娜等, 2015), 老龄林(61 a)与小兴安岭30 a白桦林(29.2-42.6 t·hm-2)(张毅, 2015)相近, 这是由于北方森林多属于中营养、贫营养地区, 树木生长受到养分供应限制(吕超群和孙书存, 2004; 解宪丽等, 2004), 且本研究区位于寒温带, 土壤有永冻层存在, 年平均气温-4.9 ℃, 不利于植被生长, 使得该区域白桦生长速度较慢, 合成碳物质的累积量较小, 乔木层碳含量较低(van Huysen et al., 2016), 即白桦林乔木层碳储量受到林龄与立地条件的显著影响, 这与该区域其他树种的研究结果(黄国胜等, 2014)相类似。3.2 林下地被物层碳储量
25、40与61 a白桦林的林下地被物层碳储量分别为2.3、3.2和4.0 t·hm-2, 随着林龄的增加呈升高趋势。林下生物量随林龄的增加未发生明显变化, 其平均碳含量随林龄的增加先升高后降低, 这可能与林内郁闭度有关, 40 a白桦林内郁闭度高(表1), 林下植物接受光照相对较少, 不利于植物体碳的累积(齐光等, 2011), 因此活地被物层碳储量随林龄的增加呈现先降低后升高的变化趋势。有关凋落物随林龄增加的变化趋势目前已有研究结果, 但尚未得出定论(Bradford & Kastendick, 2010; Zhu et al., 2017a), 本研究发现寒温带地区白桦林凋落物碳储量随林龄的增长呈现升高趋势, 这是因为随着林龄的增加植被碳储量显著增加, 导致凋落物的产量也随之增加(周序力等, 2018)。3.3 土壤碳储量
本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b)。不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似。一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(向云西等, 2019)。寒温带地区白桦林碳储量低于其他区域研究结果(韩营营等, 2015; 杨丽丽等, 2015), 主要是由于该区域存在冻融现象, 冻融作用导致土壤可溶性有机碳流失, 在很大程度上影响了土壤碳的累积(鲁博权等, 2019); 此外该地区年平均气温较低, 不利于凋落物分解, 土壤碳输入较少, 且土层较薄, 也使得其土壤碳储量较低(刘林馨等, 2018)。3.4 生态系统碳储量及其分配
本研究中我国寒温带地区白桦林生态系统碳储量从25 a的77.4 t·hm-2增加至61 a的271.4 t·hm-2, 主要由土壤碳储量贡献, 这与小兴安岭地区白桦林(张毅等, 2015)及国内外很多高纬度地区森林的研究结果(吕超群和孙书存, 2004)相一致。本研究中寒温带地区25、40与61 a白桦林生态系统碳储量均表现为土壤>乔木层>林下地被物层, 地上>地下, 这与国内外许多温带、亚热带森林类型的研究结果相一致, 如大兴安岭南段华北落叶松林(舒洋等, 2016)和亚热带日本落叶松林(马丰丰等, 2016)等。寒温带地区白桦林生态系统碳储量主要由土壤碳储量贡献, 随着林龄增长, 土壤碳储量比重先升高后降低, 乔木层碳储量比重与之相反, 先降低后升高。林下植被层的碳储量比重随林龄的增加而降低, 由25 a的3.0%减少至61 a的1.5%。3.5 白桦林植被净初级生产力与年固碳量
本研究发现寒温带地区白桦林在生长季内植被净初级生产力与年固碳量随着林龄的增加而升高, 其年固碳量从25 a的1.0 t·hm-2增加至61 a的2.1 t·hm-2, 但低于其他地区的白桦林(李娜娜等, 2015; 郑瞳等, 2016), 这是由于本研究区纬度较高, 年平均气温较低, 而森林净初级生产力与年平均气温之间存在正相关关系(Raich et al., 2006)。不同林龄白桦林植被年固碳量均以乔木层占主体地位(72.2%- 95.2%), 其中乔木层各器官以树叶最大, 且随着林龄增加树叶占乔木层比例逐渐增大, 由25 a的55.6%增加至61 a的70.0%。灌木层与草本层占其植被净初级生产力(4.5%-33.3%)和年固碳量(4.8%- 27.8%)的次要地位, 不同林龄白桦林灌木层与草本层生物量相差较大, 对植被净初级生产力与年固碳量的贡献率存在显著差异, 因此计算寒温带地区白桦林植被净初级生产力与年固碳量时应区分龄组并考虑灌木层与草本层对其的影响, 否则所得数值将与实际值间存在较大偏差。3.6 不确定性
本研究调查了我国寒温带地区不同林龄白桦林生态系统乔木层、林下地被物层与土壤层碳储量及分配特征。以下因素可能会为本研究结果带来一定的不确定性: 一是由于本实验区域位于保护区内, 该区严禁采伐树木,因此未能建立本研究区白桦的相对生长方程。只能采用相近气候条件下同一树种的相对生长方程进行乔木层生物量计算,会对本研究结果造成一定的不确定性。二是本研究采用空间代替时间的方法, 尽管所选的不同林龄白桦林立地条件基本一致, 但不排除空间异质性的存在, 也会为本研究结果带来一定的不确定性。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1139/X10-002URL
DOI:10.1111/gcb.12994URLPMID:26136379 [本文引用: 1]

Biomass change of the world's forests is critical to the global carbon cycle. Despite storing nearly half of global forest carbon, the boreal biome of diverse forest types and ages is a poorly understood component of the carbon cycle. Using data from 871 permanent plots in the western boreal forest of Canada, we examined net annual aboveground biomass change (ΔAGB) of four major forest types between 1958 and 2011. We found that ΔAGB was higher for deciduous broadleaf (DEC) (1.44?Mg?ha(-1) ?year(-1) , 95% Bayesian confidence interval (CI), 1.22-1.68) and early-successional coniferous forests (ESC) (1.42, CI, 1.30-1.56) than mixed forests (MIX) (0.80, CI, 0.50-1.11) and late-successional coniferous (LSC) forests (0.62, CI, 0.39-0.88). ΔAGB declined with forest age as well as calendar year. After accounting for the effects of forest age, ΔAGB declined by 0.035, 0.021, 0.032 and 0.069?Mg?ha(-1) ?year(-1) per calendar year in DEC, ESC, MIX and LSC forests, respectively. The ΔAGB declines resulted from increased tree mortality and reduced growth in all forest types except DEC, in which a large biomass loss from mortality was accompanied with a small increase in growth. With every degree of annual temperature increase, ΔAGB decreased by 1.00, 0.20, 0.55 and 1.07 Mg?ha(-1) ?year(-1) in DEC, ESC, MIX and LSC forests, respectively. With every cm decrease of annual climatic moisture availability, ΔAGB decreased 0.030, 0.045 and 0.17?Mg?ha(-1) ?year(-1) in ESC, MIX and LSC forests, but changed little in DEC forests. Our results suggest that persistent warming and decreasing water availability have profound negative effects on forest biomass in the boreal forests of western Canada. Furthermore, our results indicate that forest responses to climate change are strongly dependent on forest composition with late-successional coniferous forests being most vulnerable to climate changes in terms of aboveground biomass.
[本文引用: 1]
[本文引用: 1]
DOI:10.1126/science.1058629URLPMID:11423660 [本文引用: 1]
The location and mechanisms responsible for the carbon sink in northern mid-latitude lands are uncertain. Here, we used an improved estimation method of forest biomass and a 50-year national forest resource inventory in China to estimate changes in the storage of living biomass between 1949 and 1998. Our results suggest that Chinese forests released about 0.68 petagram of carbon between 1949 and 1980, for an annual emission rate of 0.022 petagram of carbon. Carbon storage increased significantly after the late 1970s from 4.38 to 4.75 petagram of carbon by 1998, for a mean accumulation rate of 0.021 petagram of carbon per year, mainly due to forest expansion and regrowth. Since the mid-1970s, planted forests (afforestation and reforestation) have sequestered 0.45 petagram of carbon, and their average carbon density increased from 15.3 to 31.1 megagrams per hectare, while natural forests have lost an additional 0.14 petagram of carbon, suggesting that carbon sequestration through forest management practices addressed in the Kyoto Protocol could help offset industrial carbon dioxide emissions.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 3]
[本文引用: 3]
DOI:10.11707/j.1001-7488.20140622URL [本文引用: 1]
Based on the data of the national continuous forest inventory from 2008 to 2013, single tree biomass of larch forest in northeastern China was estimate using biomass compatibility model of different tree species, and summed to plot biomass. Biomass expansion factor (BEF) was calculated to estimate forest biomass, and then biomass was transformed into carbon storage with carbon concentration. With the inventory statistics, carbon storages of provinces at different stages were summarized. The results indicated that: 1) vertical distribution of biomass for larch organs was: stem (48.11%)> root (23.04%)> branch (16.15%)> bark (7.2%)> foliage (4.76%), aboveground biomass made up 76.9% of the total, and root/shoot ratios were raised by age groups. 2) BEF values of larch field plot were reducing by age groups, around an average level of 0.95. 3) total carbon storage of northeastern China was 167.5×106t, based on mid-aged forest, which accounting for 44.22%, the maximum carbon density of mature forest reached to 55.22 t·hm-2, and the mean value of all age groups was only 37.06 t·hm-2, both were far below to the national average. 4) Heilongjiang province carbon storage as 128.35×106 t took the largest percentage of 76.63%, all three provinces had virtually equal carbon density, which increased with age group, but grew slower after near-mature stage.
DOI:10.11707/j.1001-7488.20140622URL [本文引用: 1]
Based on the data of the national continuous forest inventory from 2008 to 2013, single tree biomass of larch forest in northeastern China was estimate using biomass compatibility model of different tree species, and summed to plot biomass. Biomass expansion factor (BEF) was calculated to estimate forest biomass, and then biomass was transformed into carbon storage with carbon concentration. With the inventory statistics, carbon storages of provinces at different stages were summarized. The results indicated that: 1) vertical distribution of biomass for larch organs was: stem (48.11%)> root (23.04%)> branch (16.15%)> bark (7.2%)> foliage (4.76%), aboveground biomass made up 76.9% of the total, and root/shoot ratios were raised by age groups. 2) BEF values of larch field plot were reducing by age groups, around an average level of 0.95. 3) total carbon storage of northeastern China was 167.5×106t, based on mid-aged forest, which accounting for 44.22%, the maximum carbon density of mature forest reached to 55.22 t·hm-2, and the mean value of all age groups was only 37.06 t·hm-2, both were far below to the national average. 4) Heilongjiang province carbon storage as 128.35×106 t took the largest percentage of 76.63%, all three provinces had virtually equal carbon density, which increased with age group, but grew slower after near-mature stage.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 3]
The effect of site types on the ecosystem carbon storage (vegetation and soil), net primary productivity (NPP) and annual net carbon sequestration (ANCS) of natural white birch (Betula platyphylla Suk.) forests were measured on seven site types (the top, middle, and bottom of the sunny slope and shady slope, and the valley floor) using relative growth equations and carbon/nitrogen analytical approach in Changbai Mountains, Northeast China. The results are as follows. (1) The vegetation carbon storage (45.61 87.22 t·hm-2) of natural white birch forests took on the upper and middle of shady slope and the valley > the upper and middle of sunny slope, and the lower of shady slope > the lower of sunny slope, which the high site types were significantly higher (50.8% 91.2%, P<0.05) than the low site type, and the medium site types were 20.4% 44.4% higher than the low site type (P>0.05). (2) The soil organic carbon storage (66.71 158.51 t·hm-2) took on the upper of sunny slope, the middle of shady slope and the valley > the lower and middle of sunny slope, and the lower of shady slope > the upper of shady slope, which the high site types were significantly higher (99.3% 137.6%, P<0.05) than the low site type, and the medium site types were 40.7% 67.0% higher than the low site type (P>0.05). (3) The ecosystem carbon storage (139.44 231.12 t·hm-2) took on the middle of shady slope and the valley > the upper of sunny slope and the lower of shady slope > the lower and the middle of sunny slope, and the upper of shady slope, which the high site types were significantly higher (35.6% 65.7%, P<0.05) than the low site type, and the medium site types were 5.8% 34.7% higher (P>0.05) than the low site type. (4) The NPP and ANCS of the white birch forests(4.92 11.25 t·hm-2·a-1 and 2.32 5.32 t·hm-2·a-1) all took on the upper and middle of shady slope > the upper and lower of sunny slope, the lower of shady slope, and the valley > the middle of sunny, which the high site types were significantly higher (42.5% 128.7% or 45.2% 129.3%, P<0.05) than the low and medium site type, and the medium site types were 10.6% 56.3% or 14.2% 53.4% higher than the low site type, but only the lower of shady slope had significant difference with it. Therefore, the ecosystem carbon storage and carbon sequestration capacity of natural birch forests were strongly influenced by the site types in Changbai Mountains, and the site differentiation regularity should be considered in the evaluation of the carbon sink function.
URL [本文引用: 3]
The effect of site types on the ecosystem carbon storage (vegetation and soil), net primary productivity (NPP) and annual net carbon sequestration (ANCS) of natural white birch (Betula platyphylla Suk.) forests were measured on seven site types (the top, middle, and bottom of the sunny slope and shady slope, and the valley floor) using relative growth equations and carbon/nitrogen analytical approach in Changbai Mountains, Northeast China. The results are as follows. (1) The vegetation carbon storage (45.61 87.22 t·hm-2) of natural white birch forests took on the upper and middle of shady slope and the valley > the upper and middle of sunny slope, and the lower of shady slope > the lower of sunny slope, which the high site types were significantly higher (50.8% 91.2%, P<0.05) than the low site type, and the medium site types were 20.4% 44.4% higher than the low site type (P>0.05). (2) The soil organic carbon storage (66.71 158.51 t·hm-2) took on the upper of sunny slope, the middle of shady slope and the valley > the lower and middle of sunny slope, and the lower of shady slope > the upper of shady slope, which the high site types were significantly higher (99.3% 137.6%, P<0.05) than the low site type, and the medium site types were 40.7% 67.0% higher than the low site type (P>0.05). (3) The ecosystem carbon storage (139.44 231.12 t·hm-2) took on the middle of shady slope and the valley > the upper of sunny slope and the lower of shady slope > the lower and the middle of sunny slope, and the upper of shady slope, which the high site types were significantly higher (35.6% 65.7%, P<0.05) than the low site type, and the medium site types were 5.8% 34.7% higher (P>0.05) than the low site type. (4) The NPP and ANCS of the white birch forests(4.92 11.25 t·hm-2·a-1 and 2.32 5.32 t·hm-2·a-1) all took on the upper and middle of shady slope > the upper and lower of sunny slope, the lower of shady slope, and the valley > the middle of sunny, which the high site types were significantly higher (42.5% 128.7% or 45.2% 129.3%, P<0.05) than the low and medium site type, and the medium site types were 10.6% 56.3% or 14.2% 53.4% higher than the low site type, but only the lower of shady slope had significant difference with it. Therefore, the ecosystem carbon storage and carbon sequestration capacity of natural birch forests were strongly influenced by the site types in Changbai Mountains, and the site differentiation regularity should be considered in the evaluation of the carbon sink function.
URL [本文引用: 1]
Soil characteristics,water holding capacity of litter and soil water storage properties of four different ages of Betula platyphylla secondary forest in Xiaoxing'an Mountain were analyzed.The results showed that: the volume and maximum water holding capacity of litter in 38 years old of Betula platyphylla secondary forest were biggest,while that in 70 years old were lower,relatively. There were significant positive correlation between the volume and maximum water holding capacity of litter.And the non-capillary porosity was fluctuating changes with ages.It was beneficial to rainfall infiltration through the 0-30 cm soil layer in 38 years old of the Betula platyphylla secondary forest,while that in 25 years old with the smallest non-capillary porosity is not beneficial to rainfall infiltration according to the soil bulk density and non-capillary characteristics analysis.Based on the investigated data,the decreasing order of soil water conservation function of the four ages of the Betula platyphyllasecondary forest was 70 a(3 628.445 t/hm2),56 a(3 524.015 t/hm2),25 a(3 433.626 t/hm2),38 a(3 275.820 t/hm2).
URL [本文引用: 1]
Soil characteristics,water holding capacity of litter and soil water storage properties of four different ages of Betula platyphylla secondary forest in Xiaoxing'an Mountain were analyzed.The results showed that: the volume and maximum water holding capacity of litter in 38 years old of Betula platyphylla secondary forest were biggest,while that in 70 years old were lower,relatively. There were significant positive correlation between the volume and maximum water holding capacity of litter.And the non-capillary porosity was fluctuating changes with ages.It was beneficial to rainfall infiltration through the 0-30 cm soil layer in 38 years old of the Betula platyphylla secondary forest,while that in 25 years old with the smallest non-capillary porosity is not beneficial to rainfall infiltration according to the soil bulk density and non-capillary characteristics analysis.Based on the investigated data,the decreasing order of soil water conservation function of the four ages of the Betula platyphyllasecondary forest was 70 a(3 628.445 t/hm2),56 a(3 524.015 t/hm2),25 a(3 433.626 t/hm2),38 a(3 275.820 t/hm2).
DOI:10.1007/s12374-010-9140-9URL [本文引用: 1]
This study examined the biomass and carbon pools of the main ecosystem components in an age sequence of five Korean pine plantation forest stands in central Korea. The C contents in the tree and ground vegetation biomass, coarse woody debris, forest floor, and mineral soil were estimated by analyzing the C concentration of each component. The aboveground and total tree biomass increased with increasing stand age. The highest C concentration across this chronosequence was found in the tree branch while the lowest C concentration was found in the ground vegetation. The observed C contents for tree components, ground vegetation, and coarse woody debris were generally lower than the predicted C contents estimated from a biomass C factor of 0.5. Forest floor C content was age-independent. Total mineral soil C content appeared to decline initially after establishing Korean pine plantations and recover by the stand age of 35 years. Although aboveground tree biomass C content showed considerable accumulation with increasing age, the relative contribution of below ground C to total ecosystem C content varied substantially. These results suggest that successional development as temporal factor has a key role in estimating the C storage in Korean pine plantation forests.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
Soil samples were collected from depths of 0~10,10~20 and 20~30 cm in two typical forest types of Pinus sylvestris var.mongolica and Betula platyphylla in Daxing'anling region in October 2017. Indoor experiment on freeze-thaw simulation was conducted to investigate the trends of change in soil active organic carbon and characteristics of nitrogen mineralization under freeze-thaw cycles. Results showed that:①soil dissolved organic carbon (DOC) and soil microbial biomass carbon (MBC) both showed increasing trends firstly and then were followed by declined tendencies with the increase of freeze-thaw cycles. Freeze-thaw significantly increased soil DOC, but with decreased soil MBC content (p<0.05). The difference in temperature of freeze-thaw had no obvious effect on activated carbon (p>0.05). Besides, we found the DOC content of Pinus sylvestris forest was higher than birch forest in 0~10 cm soil and lower in 10~20 cm and 20~30 cm soils, respectively. However, the MBC content in each soil layer of Pinus sylvestris forest was consistently higher than that of birch forest. ②Freeze-thaw significantly increased soil ammonium nitrogen and nitrate nitrogen content (p<0.05), which presented different trends of change. Soil ammonium nitrogen experienced a trend of decreasing before the notable increase. However, nitrate nitrogen noted an opposite change of trend compared with soil ammonium nitrogen. The large difference of temperature in freeze-thaw accelerated nitrogen mineralization (p<0.05). Meantime, the ammonium nitrogen and nitrate nitrogen in birch forest soil were persistently larger than that in Pinus sylvestris forest during freezing and thawing. ③The contents of DOC, MBC, ammonium nitrogen and nitrate nitrogen decreased with the increase of soil depth in both the forest types (p<0.05). The above results show that the loss of DOC content in forest soil caused by freeze-thaw affects the accumulation of soil carbon pool and inorganic nitrogen, which provides nitrogen fertility for plant growth. Moreover, the response to freeze-thaw action of soil carbon and nitrogen from different vegetation types vary to some extent.
URL [本文引用: 2]
Soil samples were collected from depths of 0~10,10~20 and 20~30 cm in two typical forest types of Pinus sylvestris var.mongolica and Betula platyphylla in Daxing'anling region in October 2017. Indoor experiment on freeze-thaw simulation was conducted to investigate the trends of change in soil active organic carbon and characteristics of nitrogen mineralization under freeze-thaw cycles. Results showed that:①soil dissolved organic carbon (DOC) and soil microbial biomass carbon (MBC) both showed increasing trends firstly and then were followed by declined tendencies with the increase of freeze-thaw cycles. Freeze-thaw significantly increased soil DOC, but with decreased soil MBC content (p<0.05). The difference in temperature of freeze-thaw had no obvious effect on activated carbon (p>0.05). Besides, we found the DOC content of Pinus sylvestris forest was higher than birch forest in 0~10 cm soil and lower in 10~20 cm and 20~30 cm soils, respectively. However, the MBC content in each soil layer of Pinus sylvestris forest was consistently higher than that of birch forest. ②Freeze-thaw significantly increased soil ammonium nitrogen and nitrate nitrogen content (p<0.05), which presented different trends of change. Soil ammonium nitrogen experienced a trend of decreasing before the notable increase. However, nitrate nitrogen noted an opposite change of trend compared with soil ammonium nitrogen. The large difference of temperature in freeze-thaw accelerated nitrogen mineralization (p<0.05). Meantime, the ammonium nitrogen and nitrate nitrogen in birch forest soil were persistently larger than that in Pinus sylvestris forest during freezing and thawing. ③The contents of DOC, MBC, ammonium nitrogen and nitrate nitrogen decreased with the increase of soil depth in both the forest types (p<0.05). The above results show that the loss of DOC content in forest soil caused by freeze-thaw affects the accumulation of soil carbon pool and inorganic nitrogen, which provides nitrogen fertility for plant growth. Moreover, the response to freeze-thaw action of soil carbon and nitrogen from different vegetation types vary to some extent.
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
Carbon storage and its allocation of 7-, 29- and 32-year-old Erythrophleum fordii plantation ecosystems in Guangxi were studied on the basis of biomass survey. The results showed that the carbon contents in different organs of E. fordii, ranging from 509.0 to 572.4 g·kg-1, were in the order of stem > branch > root > bark > leaf. No significant differences in carbon content were observed among the shrub, herb and litter layers of the E. fordii plantations with different ages. Carbon content in the soil layer (0-100 cm) decreased with increasing the soil depth, but increased with increasing the stand age. The carbon storage of the arbor layer was 21.8, 100.0 and 121.6 t·hm-2 for 7-, 29- and 32-year-old stands, respectively, and the order of carbon storage allocation in different organs was same as the order of carbon content. The 7-, 29- and 32-year-old E. fordii plantation ecosystems stored carbon at 132.6, 220.2 and 242.6 t·hm-2, respectively. The arbor layer and soil layer were the main carbon pools, accounting for more than 97% of carbon storage in the ecosystem. Carbon storage allocation increased in arbor layer but decreased in soil layer with increasing the stand age. The influence of stand age on carbon storage allocation in shrub, herb and litter layers did not show a obvious regular pattern.
URL [本文引用: 1]
Carbon storage and its allocation of 7-, 29- and 32-year-old Erythrophleum fordii plantation ecosystems in Guangxi were studied on the basis of biomass survey. The results showed that the carbon contents in different organs of E. fordii, ranging from 509.0 to 572.4 g·kg-1, were in the order of stem > branch > root > bark > leaf. No significant differences in carbon content were observed among the shrub, herb and litter layers of the E. fordii plantations with different ages. Carbon content in the soil layer (0-100 cm) decreased with increasing the soil depth, but increased with increasing the stand age. The carbon storage of the arbor layer was 21.8, 100.0 and 121.6 t·hm-2 for 7-, 29- and 32-year-old stands, respectively, and the order of carbon storage allocation in different organs was same as the order of carbon content. The 7-, 29- and 32-year-old E. fordii plantation ecosystems stored carbon at 132.6, 220.2 and 242.6 t·hm-2, respectively. The arbor layer and soil layer were the main carbon pools, accounting for more than 97% of carbon storage in the ecosystem. Carbon storage allocation increased in arbor layer but decreased in soil layer with increasing the stand age. The influence of stand age on carbon storage allocation in shrub, herb and litter layers did not show a obvious regular pattern.
DOI:10.1126/science.1204588URL [本文引用: 1]
Immune clearance and resource limitation (via red blood cell depletion) shape the peaks and troughs of malaria parasitemia, which in turn affect disease severity and transmission. Quantitatively partitioning the relative roles of these effects through time is challenging. Using data from rodent malaria, we estimated the effective propagation number, which reflects the relative importance of contrasting within-host control mechanisms through time and is sensitive to the inoculating parasite dose. Our analysis showed that the capacity of innate responses to restrict initial parasite growth saturates with parasite dose and that experimentally enhanced innate immunity can affect parasite density indirectly via resource depletion. Such a statistical approach offers a tool to improve targeting of drugs or vaccines for human therapy by revealing the dynamics and interactions of within-host regulatory mechanisms.
DOI:10.1111/gcb.2004.10.issue-12URL [本文引用: 1]
URL [本文引用: 1]
Through sampling site investigation, this paper studied the carbon storage of arbor, herb, and whole vegetation in 10-, 12-, 15-, 26-, and 61-year old Larix gmelinii plantations in Huzhong Forestry Bureau of Great Xing’an Mountains, Northeast China, and ‘temporal for spatial’ method was employed to approach the variations of the vegetation carbon storage during the growth of the plantations. The results revealed that the vegetation carbon storage in the plantations increased with stand age, and reached 105.69 t·hm-2 at age of 61 years, representing a marked role as a carbon sink. The L. gmelinii plantations at the ages from 15 to 26 years had the strongest capability in carbon sequestration, in which, the carbon storage in trunk occupied 54.3%-73.9% of the total carbon storage of arbor, and, with the increase of stand age, the trunk’s carbon storage to the total carbon storage of arbor as well as the trunk’s carbon density increased. As for the other organs, the rate of their carbon storage to the total carbon storage of arbor decreased with stand age, while their carbon density increased first but eventually leveled off or had a slight decrease till at age of 61 years. Based on these results, the rotation age for the L. gmelinii plantations in Great Xing’an Mountains would properly be lengthened to at least 60years.
URL [本文引用: 1]
Through sampling site investigation, this paper studied the carbon storage of arbor, herb, and whole vegetation in 10-, 12-, 15-, 26-, and 61-year old Larix gmelinii plantations in Huzhong Forestry Bureau of Great Xing’an Mountains, Northeast China, and ‘temporal for spatial’ method was employed to approach the variations of the vegetation carbon storage during the growth of the plantations. The results revealed that the vegetation carbon storage in the plantations increased with stand age, and reached 105.69 t·hm-2 at age of 61 years, representing a marked role as a carbon sink. The L. gmelinii plantations at the ages from 15 to 26 years had the strongest capability in carbon sequestration, in which, the carbon storage in trunk occupied 54.3%-73.9% of the total carbon storage of arbor, and, with the increase of stand age, the trunk’s carbon storage to the total carbon storage of arbor as well as the trunk’s carbon density increased. As for the other organs, the rate of their carbon storage to the total carbon storage of arbor decreased with stand age, while their carbon density increased first but eventually leveled off or had a slight decrease till at age of 61 years. Based on these results, the rotation age for the L. gmelinii plantations in Great Xing’an Mountains would properly be lengthened to at least 60years.
DOI:10.1890/05-0023URLPMID:16634298 [本文引用: 1]
Evergreen broad-leaved tropical forests can have high rates of productivity and large accumulations of carbon in plant biomass and soils. They can therefore play an important role in the global carbon cycle, influencing atmospheric CO2 concentrations if climate warms. We applied meta-analyses to published data to evaluate the apparent effects of temperature on carbon fluxes and storages in mature, moist tropical evergreen forest ecosystems. Among forests, litter production, tree growth, and belowground carbon allocation all increased significantly with site mean annual temperature (MAT); total net primary productivity (NPP) increased by an estimated 0.2-0.7 Mg C x ha(-1) x yr(-1) x degrees C(-1). Temperature had no discernible effect on the turnover rate of aboveground forest biomass, which averaged 0.014 yr(-1) among sites. Consistent with these findings, forest biomass increased with site MAT at a rate of 5-13 Mg C x ha(-1) x degrees C(-1). Despite greater productivity in warmer forests, soil organic matter accumulations decreased with site MAT, with a slope of -8 Mg C x ha(-1) x degrees C(-1), indicating that decomposition rates of soil organic matter increased with MAT faster than did rates of NPP. Turnover rates of surface litter also increased with temperature among forests. We found no detectable effect of temperature on total carbon storage among moist-tropical evergreen forests, but rather a shift in ecosystem structure, from low-biomass forests with relatively large accumulations of detritus in cooler sites, to large-biomass forests with relatively smaller detrital stocks in warmer locations. These results imply that, in a warmer climate, conservation of forest biomass will be critical to the maintenance of carbon stocks in moist tropical forests.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11104-016-2857-6URL [本文引用: 1]
DOI:10.1016/j.catena.2012.07.003URL [本文引用: 1]
Land use change is one of the major factors that affect soil organic carbon (SOC) variation and global carbon balance. Since the late 1970s. a significant area of the hilly Loess Plateau has undergone major land use changes during several revegetation programs, especially in the Grain for Green Project. However, so far there has not been a comprehensive study to determine temporal and spatial variations in SOC due to revegetation in this region, which hampers accurate predictions of the SOC sequestration potential and the land use change impacts. In this study, slope cropland and five typical revegetation types in the hilly Loess Plateau were selected, and then classified into 36 groups according to different revegetation years and landforms to investigate temporal and spatial variations of SOC and the impacts of relevant factors following revegetation. The results showed that the SOC concentration in the top soil horizon (0-5 cm) increased most significantly with revegetation and that increases in SOC slowed with increasing soil depth, but coefficient of variations in different soil horizons indicated that revegetation could cause SOC concentration differences at up to a 30 cm depth. Temporal variation in SOC occurred in two main phases in replanted cropland: in the first 10 or 15 yr, the profile mass of SOC (SOC density, SOCD) increased slightly; after this phase. SOCD increased significantly, with sequestration rates of 0.69, 0.55 and 0.24 t.ha(-1).yr(-1) (revegetation 10 to 35 yr) for planted woodland, planted shrubland and abandoned cropland, respectively. The SOC sequestration rate in wild grassland was 0.23 t.ha(-1).yr(-1) (revegetation 10 to 35 yr), which was similar to that in abandoned cropland. In contrast, the SOCD in wild shrubland increased rapidly in the first 10 yr, with a rate of 0.93 t.ha(-1).yr(-1), and then by 0.56 t.ha(-1).yr(-1) over the next 25 years. SOCD spatial differences in different landforms had reached several times the annual SOCD increment and followed a new trend after revegetation: SOC sequestration in shady slope areas was significantly higher than in sunny slope areas, but no significant difference was found between gentle slopes and steep slopes after revegetation. A general linear model was used to identify the factors that were most relevant to SOC variation. Revegetation years accounted for nearly half of the total contribution to SOC variation (42%), and land use type was responsible for 33% of SOC variation. SOCD distribution in a watershed confirmed that variables of land use and years after revegetation dominated SOC variation. Landforms had a small influence on SOC variation at the regional scale, but the influence of the slope aspect was still large. The SOC sequestration rate was about 0.21 to 0.64 t.ha(-1).yr(-1) with revegetation in the hilly Loess Plateau, which was highest in wild shrubland, followed by planted woodland, planted shrubland, abandoned cropland and wild grassland. Considering the large area of revegetation and relatively high SOC sequestration rate, SOC sequestration in this region should contribute significantly to decreasing the carbon concentration in atmosphere. (C) 2012 Elsevier B.V.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.11707/j.1001-7488.20090507URL [本文引用: 1]
The biomass, carbon storage and spatial distribution of 46-old-years (42-old-years to 51-old-years) Pinus massoniana nature secondary forest were studied in Tieshanping of ChongQing. The results showed that the total biomass is 142.06 t·hm-2, the distributing sequence of the biomass of the forest ecosystem is: arbor layer(89.91%) > shrub layer(5.61%)>Standing litters(2.98%)> >herb(1.50%). The total organic carbon storage in forest ecosystem amounted to 197.78 t C·hm-2, of which trees layer stored 76.06 t C·hm-2 and accounted for 38.45%, shrub layer stored 3.55 t C·hm-2 and accounted for 1.79%, herb layer stored 0.88 t C·hm-2 and accounted to 0.44%, standing litters mass layer stored 2.34 t C·hm-2 and accounted to 1.17%, and soil stored 114.96 t C·hm-2 and accounted to 58.13%. The carbon storage in different organs was positively related to the biomass of corresponding organs. Stem accumulated the highest biomass, which also stored the highest carbon storage and comprised 75.06% of carbon storage in arbor layer. The annual net productivity of Pinus Massoniana trees was 9.01 t·hm-2a-1, and annual carbon storage was up to 4.49 t C·hm-2a-1, amounted CO2 to 16.46 t CO2·hm-2a-1.
DOI:10.11707/j.1001-7488.20090507URL [本文引用: 1]
The biomass, carbon storage and spatial distribution of 46-old-years (42-old-years to 51-old-years) Pinus massoniana nature secondary forest were studied in Tieshanping of ChongQing. The results showed that the total biomass is 142.06 t·hm-2, the distributing sequence of the biomass of the forest ecosystem is: arbor layer(89.91%) > shrub layer(5.61%)>Standing litters(2.98%)> >herb(1.50%). The total organic carbon storage in forest ecosystem amounted to 197.78 t C·hm-2, of which trees layer stored 76.06 t C·hm-2 and accounted for 38.45%, shrub layer stored 3.55 t C·hm-2 and accounted for 1.79%, herb layer stored 0.88 t C·hm-2 and accounted to 0.44%, standing litters mass layer stored 2.34 t C·hm-2 and accounted to 1.17%, and soil stored 114.96 t C·hm-2 and accounted to 58.13%. The carbon storage in different organs was positively related to the biomass of corresponding organs. Stem accumulated the highest biomass, which also stored the highest carbon storage and comprised 75.06% of carbon storage in arbor layer. The annual net productivity of Pinus Massoniana trees was 9.01 t·hm-2a-1, and annual carbon storage was up to 4.49 t C·hm-2a-1, amounted CO2 to 16.46 t CO2·hm-2a-1.
[本文引用: 2]
[本文引用: 2]
DOI:10.17521/cjpe.2018.0064URL [本文引用: 3]
DOI:10.17521/cjpe.2018.0064URL [本文引用: 3]
DOI:10.1016/j.scitotenv.2018.08.016URLPMID:30180331 [本文引用: 1]
Soil is recognized as the largest carbon reservoir in the terrestrial ecosystem. Soil organic carbon (SOC) is vulnerable to changes in land use and climate. For a better understanding of the SOC dynamics and its driving factors, we collected data of the 1980s and 2000s in the North and Northeast China and conducted the digital soil mapping for spatial variation of SOC for the respective period. In the 1980s, 585 soils were sampled and the area was resampled in 2003 and 2004 (1062 samples) in a 30-km grid. The main land use in the area was cropland, forest and grassland. The random forest was used to predict the SOC concentration and its temporal change using land use, terrain factors, vegetation index, vis-NIR spectra and climate factors as predictors. The average SOC concentration in 1985 was 10.0?g?kg-1 compared to 12.5?g?kg-1 in 2004. The SOC variation was similar over the two periods, and levels increased from south to north. The estimated SOC stock was 1.68 Pg in 1985 and 1.66 Pg in 2004, but the SOC changes were different under different land uses. Over the twenty-year period, average temperatures increased and large areas of forests and grassland were converted to cropland. SOC under cropland was increased by 0.094 Pg (+9%) whereas 0.089 Pg SOC was lost under forests (-25%) and 0.037 Pg in the soils under grassland (-25%). It is concluded that land use is the main drivers for SOC changes in this area while climate change had different contributions in different regions. SOC loss was remarkable under the land use conversion while cropland has considerable potential to sequester SOC.
DOI:10.1038/s41467-017-00207-1URLPMID:28751686 [本文引用: 2]
Forests play an important role in global carbon cycles. However, the lack of available information on carbon stocks in dead organic matter, including woody debris and litter, reduces the reliability of assessing the carbon cycles in entire forest ecosystems. Here we estimate that the national DOM carbon stock in the period of 2004-2008 is 925?±?54?Tg, with an average density of 5.95?±?0.35?Mg C?ha-1. Over the past two decades from periods of 1984-1988 to 2004-2008, the national dead organic matter carbon stock has increased by 6.7?±?2.2?Tg carbon per year, primarily due to increasing forest area. Temperature and precipitation increase the carbon density of woody debris, but decrease that of litter. Additionally, the woody debris increases significantly with above ground biomass and forest age. Our results can improve estimates of the carbon budget in China's forests and for better understanding of effects of climate and stand characteristics on dead organic matter distribution.Reliable estimates of the total forest carbon (C) pool are lacking due to insufficient information on dead organic matter (DOM). Here, the authors estimate that the current DOM C stock in China is 925?±?54?Tg and that it grew by 6.7?±?2.2?Tg C/yr over the past two decades primarily due to increasing forest area.
DOI:10.3390/f8060191URL [本文引用: 2]
Age-related patterns of forest complexity and carbon storage in pine and aspen- birch ecosystems of northern Minnesota, USA
2010
Net aboveground biomass declines of four major forest types with forest ageing and climate change in western Canada’s boreal forests
1
2015
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
马尾松人工林生物量及生产力变化规律研究II. 不同林龄生物量及生产力
1
2002
... 式中, ΔW为年净生产量(t·hm-2·a-1), Wm为m年时单位面积现存生物量(t·hm-2), Wm-n为(m-n)年时单位面积现存生物量(t·hm-2), n为年数(a)(
马尾松人工林生物量及生产力变化规律研究II. 不同林龄生物量及生产力
1
2002
... 式中, ΔW为年净生产量(t·hm-2·a-1), Wm为m年时单位面积现存生物量(t·hm-2), Wm-n为(m-n)年时单位面积现存生物量(t·hm-2), n为年数(a)(
Changes in forest biomass carbon storage in China between 1949 and 1998
1
2001
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
CO2失汇与北半球中高纬度陆地生态系统的碳汇
1
2001
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
CO2失汇与北半球中高纬度陆地生态系统的碳汇
1
2001
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
No growth stimulation of Canada’s boreal forest under half-century of combined warming and CO2 fertilization
1
2016
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
不同林龄白桦天然次生林土壤碳通量和有机碳储量
3
2015
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
... 本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b).不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似.一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(
不同林龄白桦天然次生林土壤碳通量和有机碳储量
3
2015
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
... 本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b).不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似.一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(
大兴安岭5种典型林型森林生物碳储量
3
2015
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
... 在踏查基础上, 选取该区域占比较高的(25 ± 6)、(40 ± 4)和(61 ± 7)年白桦林为研究对象(以下简称25、40和61 a), 按照中华人民共和国林业行业标准(LY/T2908-2017)并结合大兴安岭白桦林生长特点(
大兴安岭5种典型林型森林生物碳储量
3
2015
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
... 在踏查基础上, 选取该区域占比较高的(25 ± 6)、(40 ± 4)和(61 ± 7)年白桦林为研究对象(以下简称25、40和61 a), 按照中华人民共和国林业行业标准(LY/T2908-2017)并结合大兴安岭白桦林生长特点(
东北地区落叶松林碳储量估算
1
2014
... 林龄是影响森林生态系统碳储量的重要因素(
东北地区落叶松林碳储量估算
1
2014
... 林龄是影响森林生态系统碳储量的重要因素(
中国北方白桦自然群体比较选择及子代测定
1
2018
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
中国北方白桦自然群体比较选择及子代测定
1
2018
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
白桦天然林生物量模型的研究
1
2010
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
白桦天然林生物量模型的研究
1
2010
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
立地类型对长白山天然白桦林生态系统碳储量的影响
3
2015
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
... 林龄是影响森林生态系统碳储量的重要因素(
... 本研究发现寒温带地区白桦林在生长季内植被净初级生产力与年固碳量随着林龄的增加而升高, 其年固碳量从25 a的1.0 t·hm-2增加至61 a的2.1 t·hm-2, 但低于其他地区的白桦林(
立地类型对长白山天然白桦林生态系统碳储量的影响
3
2015
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
... 林龄是影响森林生态系统碳储量的重要因素(
... 本研究发现寒温带地区白桦林在生长季内植被净初级生产力与年固碳量随着林龄的增加而升高, 其年固碳量从25 a的1.0 t·hm-2增加至61 a的2.1 t·hm-2, 但低于其他地区的白桦林(
不同林龄白桦次生林土壤特性及其水源涵养功能
1
2009
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
不同林龄白桦次生林土壤特性及其水源涵养功能
1
2009
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
Biomass and carbon storage in an age-sequence of Korean pine (Pinus koraiensis) plantation forests in central Korea.
1
2011
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
大兴安岭不同森林群落植被多样性对土壤有机碳密度的影响
1
2018
... 本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b).不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似.一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(
大兴安岭不同森林群落植被多样性对土壤有机碳密度的影响
1
2018
... 本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b).不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似.一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(
中国森林植被的碳汇/源空间分布格局
1
2012
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
中国森林植被的碳汇/源空间分布格局
1
2012
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
白桦次生林降雨水化学及养分输入特征
1
2015
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
白桦次生林降雨水化学及养分输入特征
1
2015
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
冻融作用对大兴安岭典型森林土壤活性有机碳和氮矿化的影响
2
2019
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
... 本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b).不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似.一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(
冻融作用对大兴安岭典型森林土壤活性有机碳和氮矿化的影响
2
2019
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
... 本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b).不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似.一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(
陆地生态系统碳密度格局研究概述
2
2004
... 林龄是影响森林生态系统碳储量的重要因素(
... 本研究中我国寒温带地区白桦林生态系统碳储量从25 a的77.4 t·hm-2增加至61 a的271.4 t·hm-2, 主要由土壤碳储量贡献, 这与小兴安岭地区白桦林(
陆地生态系统碳密度格局研究概述
2
2004
... 林龄是影响森林生态系统碳储量的重要因素(
... 本研究中我国寒温带地区白桦林生态系统碳储量从25 a的77.4 t·hm-2增加至61 a的271.4 t·hm-2, 主要由土壤碳储量贡献, 这与小兴安岭地区白桦林(
亚热带日本落叶松人工林生态系统碳密度及其分配特征
1
2016
... 本研究中我国寒温带地区白桦林生态系统碳储量从25 a的77.4 t·hm-2增加至61 a的271.4 t·hm-2, 主要由土壤碳储量贡献, 这与小兴安岭地区白桦林(
亚热带日本落叶松人工林生态系统碳密度及其分配特征
1
2016
... 本研究中我国寒温带地区白桦林生态系统碳储量从25 a的77.4 t·hm-2增加至61 a的271.4 t·hm-2, 主要由土壤碳储量贡献, 这与小兴安岭地区白桦林(
小兴安岭白桦天然林生态系统碳储量的研究
1
2012
... 乔木生物量的估算采用相对生长法, 对样地内胸径≥4 cm乔木进行每木检尺, 将树高、胸径的实测结果代入
小兴安岭白桦天然林生态系统碳储量的研究
1
2012
... 乔木生物量的估算采用相对生长法, 对样地内胸径≥4 cm乔木进行每木检尺, 将树高、胸径的实测结果代入
不同林龄格木人工林碳储量及其分配特征
1
2014
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
不同林龄格木人工林碳储量及其分配特征
1
2014
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
A large and persistent carbon sink in the world’s forests
1
2011
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
Carbon cycling and storage in world forests: Biome patterns related to forest age
1
2004
... 林龄是影响森林生态系统碳储量的重要因素(
大兴安岭林区兴安落叶松人工林植被碳贮量
1
2011
... 25、40与61 a白桦林的林下地被物层碳储量分别为2.3、3.2和4.0 t·hm-2, 随着林龄的增加呈升高趋势.林下生物量随林龄的增加未发生明显变化, 其平均碳含量随林龄的增加先升高后降低, 这可能与林内郁闭度有关, 40 a白桦林内郁闭度高(
大兴安岭林区兴安落叶松人工林植被碳贮量
1
2011
... 25、40与61 a白桦林的林下地被物层碳储量分别为2.3、3.2和4.0 t·hm-2, 随着林龄的增加呈升高趋势.林下生物量随林龄的增加未发生明显变化, 其平均碳含量随林龄的增加先升高后降低, 这可能与林内郁闭度有关, 40 a白桦林内郁闭度高(
Temperature influences carbon accumulation in moist tropical forests
1
2006
... 本研究发现寒温带地区白桦林在生长季内植被净初级生产力与年固碳量随着林龄的增加而升高, 其年固碳量从25 a的1.0 t·hm-2增加至61 a的2.1 t·hm-2, 但低于其他地区的白桦林(
大兴安岭南段华北落叶松人工林碳储量及分配特征研究
1
2016
... 本研究中我国寒温带地区白桦林生态系统碳储量从25 a的77.4 t·hm-2增加至61 a的271.4 t·hm-2, 主要由土壤碳储量贡献, 这与小兴安岭地区白桦林(
大兴安岭南段华北落叶松人工林碳储量及分配特征研究
1
2016
... 本研究中我国寒温带地区白桦林生态系统碳储量从25 a的77.4 t·hm-2增加至61 a的271.4 t·hm-2, 主要由土壤碳储量贡献, 这与小兴安岭地区白桦林(
Decomposition drives convergence of forest litter nutrient stoichiometry following phosphorus addition
1
2016
... 林龄是影响森林生态系统碳储量的重要因素(
Temporal and spatial variations in soil organic carbon sequestration following revegetation in the hilly Loess Plateau, China
1
2012
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
中国省域农业碳排放: 测算、效率变动及影响因素研究——基于DEA-Malmquist指数分解方法与Tobit模型运用
1
2014
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
中国省域农业碳排放: 测算、效率变动及影响因素研究——基于DEA-Malmquist指数分解方法与Tobit模型运用
1
2014
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
马尾松叶片-凋落物-土壤的碳氮磷化学计量特征
1
2019
... 本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b).不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似.一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(
马尾松叶片-凋落物-土壤的碳氮磷化学计量特征
1
2019
... 本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b).不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似.一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(
不同植被下中国土壤有机碳的储量与影响因子
1
2004
... 林龄是影响森林生态系统碳储量的重要因素(
不同植被下中国土壤有机碳的储量与影响因子
1
2004
... 林龄是影响森林生态系统碳储量的重要因素(
六盘山四种森林生态系统的碳氮储量、组成及分布特征
2
2015
... 林龄是影响森林生态系统碳储量的重要因素(
... 本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b).不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似.一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(
六盘山四种森林生态系统的碳氮储量、组成及分布特征
2
2015
... 林龄是影响森林生态系统碳储量的重要因素(
... 本研究结果显示林龄对我国寒温带地区白桦林土壤(0-100 cm)碳储量存在显著影响(p < 0.05), 25、40与61 a白桦林土壤层碳储量分别为63.2、158.6和233.2 t·hm-2, 随着林龄的增长显著增加, 表现为碳积累过程(Zhu et al., 2017b).不同林龄白桦林土壤层(0-100 cm)平均碳含量表现为61 a (31.6 g·kg-1) > 40 a (15.9 g·kg-1) > 25 a (7.6 g·kg-1), 随着林龄的增加而显著升高(p < 0.05), 与其他地区白桦林的研究结果(韩营营等, 2015)相似.一方面是因为随着林龄增加土壤碳蓄积能力增强, 另一方面是随着林龄的增加乔木层碳含量降低, 干物质合成累积量逐渐减少, 降低了对土壤养分量的需求, 土壤碳消耗量小于累积量(
立地类型对小兴安岭天然白桦次生林生态系统碳储量的影响
1
2015
... 林龄是影响森林生态系统碳储量的重要因素(
立地类型对小兴安岭天然白桦次生林生态系统碳储量的影响
1
2015
... 林龄是影响森林生态系统碳储量的重要因素(
小兴安岭天然白桦林生态系统碳储量
2
2015
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
... 本研究中我国寒温带地区白桦林生态系统碳储量从25 a的77.4 t·hm-2增加至61 a的271.4 t·hm-2, 主要由土壤碳储量贡献, 这与小兴安岭地区白桦林(
小兴安岭天然白桦林生态系统碳储量
2
2015
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
... 本研究中我国寒温带地区白桦林生态系统碳储量从25 a的77.4 t·hm-2增加至61 a的271.4 t·hm-2, 主要由土壤碳储量贡献, 这与小兴安岭地区白桦林(
重庆铁山坪马尾松林生态系统碳贮量及其分配特征
1
2009
... 1.2.2.1 灌木层 在每个样地内, 沿对角线随机设置3个2 m × 2 m小样方, 记录小样方内灌木的种类与株数, 采用“全收获法”按照地上、地下两部分采集小样方内全部灌木, 将同种植物相同部分混合, 在野外称量各部分鲜质量.随后选取部分样品带回实验室, 85 ℃烘干至恒质量(间隔6 h两次称量所得的含水率差小于0.1%即被视为达到恒质量), 用精度为0.01 g的电子天平称干质量, 计算地上、地下部分的生物量, 两部分生物量相加计算出单位面积生物量(
重庆铁山坪马尾松林生态系统碳贮量及其分配特征
1
2009
... 1.2.2.1 灌木层 在每个样地内, 沿对角线随机设置3个2 m × 2 m小样方, 记录小样方内灌木的种类与株数, 采用“全收获法”按照地上、地下两部分采集小样方内全部灌木, 将同种植物相同部分混合, 在野外称量各部分鲜质量.随后选取部分样品带回实验室, 85 ℃烘干至恒质量(间隔6 h两次称量所得的含水率差小于0.1%即被视为达到恒质量), 用精度为0.01 g的电子天平称干质量, 计算地上、地下部分的生物量, 两部分生物量相加计算出单位面积生物量(
立地类型对张广才岭天然白桦林生态系统碳储量的影响
2
2016
... 叶片的净生产量按照现存量直接转换(
... 本研究发现寒温带地区白桦林在生长季内植被净初级生产力与年固碳量随着林龄的增加而升高, 其年固碳量从25 a的1.0 t·hm-2增加至61 a的2.1 t·hm-2, 但低于其他地区的白桦林(
立地类型对张广才岭天然白桦林生态系统碳储量的影响
2
2016
... 叶片的净生产量按照现存量直接转换(
... 本研究发现寒温带地区白桦林在生长季内植被净初级生产力与年固碳量随着林龄的增加而升高, 其年固碳量从25 a的1.0 t·hm-2增加至61 a的2.1 t·hm-2, 但低于其他地区的白桦林(
贵州月亮山不同演替阶段亮叶水青冈林碳储量及其分配格局
3
2018
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
... 林龄是影响森林生态系统碳储量的重要因素(
... 25、40与61 a白桦林的林下地被物层碳储量分别为2.3、3.2和4.0 t·hm-2, 随着林龄的增加呈升高趋势.林下生物量随林龄的增加未发生明显变化, 其平均碳含量随林龄的增加先升高后降低, 这可能与林内郁闭度有关, 40 a白桦林内郁闭度高(
贵州月亮山不同演替阶段亮叶水青冈林碳储量及其分配格局
3
2018
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
... 林龄是影响森林生态系统碳储量的重要因素(
... 25、40与61 a白桦林的林下地被物层碳储量分别为2.3、3.2和4.0 t·hm-2, 随着林龄的增加呈升高趋势.林下生物量随林龄的增加未发生明显变化, 其平均碳含量随林龄的增加先升高后降低, 这可能与林内郁闭度有关, 40 a白桦林内郁闭度高(
Land use and climate change effects on soil organic carbon in North and Northeast China
1
2019
... 寒温带林区土壤中分布着大面积连续多年冻土, 存在冻融循环现象(
a). Carbon stocks and changes of dead organic matter in China’s forests
2
2017
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
... 25、40与61 a白桦林的林下地被物层碳储量分别为2.3、3.2和4.0 t·hm-2, 随着林龄的增加呈升高趋势.林下生物量随林龄的增加未发生明显变化, 其平均碳含量随林龄的增加先升高后降低, 这可能与林内郁闭度有关, 40 a白桦林内郁闭度高(
. Plant debris and its contribution to ecosystem carbon storage in successional Larix gmelinii forests in northeastern China.
2
2017
... 由大气中温室气体排放量增加所导致的温室效应、气候变暖等问题已影响到全人类的生存和发展(
... 林龄是影响森林生态系统碳储量的重要因素(

{kind=link}
{kind=link}
{kind=link}
{kind=link}