Responses of tree growth to nitrogen addition in Quercus wutaishanica forests in Mount Dongling, Beijing, China
An-Long ZOU, Xiu-Ping LI, Xiao-Feng NI, Cheng-Jun JI,*Institute of Ecology, College of Urban and Environmental Sciences, Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China
Abstract Aims Temperate forest is one of the most important components of the global forests and main carbon pools. Nitrogen (N) is considered as the limiting nutrient for the forest growth. However, the heterogeneities in plant species and stem sizes were largely ignored in previous researches on the effects of N addition on plant growth. Quercus wutaishanica is one of the most common and dominant tree species in the temperate forests in North China. In this study, we investigated the responses of growth of trees and forests to N addition in the Quercus wutaishanica forests in Mt. Dongling in Beijing. Methods We conducted a 7-year N fertilization experiment in Quercus wutaishanica forests in Mt. Dongling, Beijing, since 2011. The N addition was conducted at three treatment levels, i.e., 0 kg hm -2·a -1, 50 kg hm -2·a -1 and 100 kg hm -2·a -1. Nitrogen was added at the beginning of each month from May to October each year. We used electronic vernier caliper to measure tree growth rate for each year. All trees were divided into three groups based on their diameter at breast height (DBH), namely small trees (DBH = 3-10 cm), median trees (DBH =10-20 cm) and large trees (DBH > 20 cm). Particularly, we considered growth at species level for all Quercus wutaishanica and the growth at community level for all tree species in the stands. Important findings (1) At species level, N addition enhanced the growth rate of Q. wutaishanica. (2) At community level, the growth rate showed no difference among different N addition treatments. (3) Small trees were restrained, while median and large Q. wutaishanica trees were not significantly influenced, by the N addition. Keywords:nitrogen deposition;Quercus wutaishanica;tree growth;temperate forest
PDF (1354KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 邹安龙, 李修平, 倪晓凤, 吉成均. 模拟氮沉降对北京东灵山辽东栎林树木生长的影响. 植物生态学报, 2019, 43(9): 783-792. DOI: 10.17521/cjpe.2018.0232 ZOU An-Long, LI Xiu-Ping, NI Xiao-Feng, JI Cheng-Jun. Responses of tree growth to nitrogen addition in Quercus wutaishanica forests in Mount Dongling, Beijing, China. Chinese Journal of Plant Ecology, 2019, 43(9): 783-792. DOI: 10.17521/cjpe.2018.0232
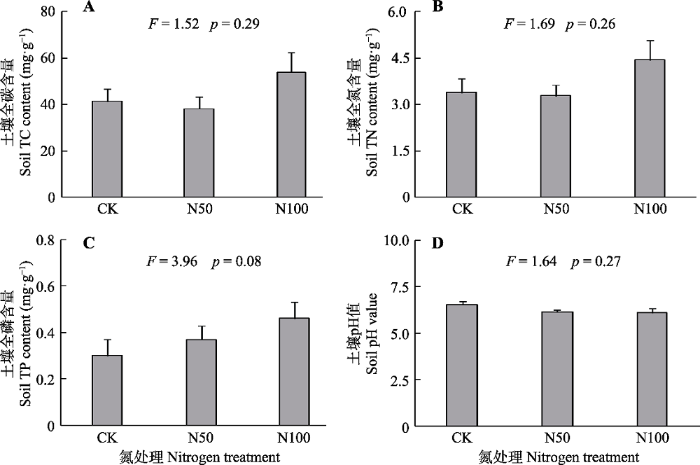
新窗口打开|下载原图ZIP|生成PPT 图1氮添加对北京东灵山辽东栎林土壤全碳(TC)、全氮(TN), 全磷(TP)含量以及pH值的影响(平均值+标准误差, n = 3)。不同小写字母表示处理间差异显著(p < 0.05)。CK、N50、N100分别表示氮添加量0、50、100 kg·hm-2·a-1。
Fig. 1Influence of nitrogen addition treatments on soil total carbon (TC), total nitrogen (TN), total phosphorus (TP) content and pH value in Quercus wutaishanica forests in Mt. Dongling, Beijing (mean + SE, n = 3). Different lowercase letters indicate significant difference among treatments (p < 0.05). CK, N50, N100 denote nitrogen addition 0, 50, 100 kg·hm-2·a-1, respectively.
新窗口打开|下载原图ZIP|生成PPT 图2氮添加对北京东灵山辽东栎叶片全碳(TC)、全氮(TN)、全磷(TP)含量的影响(平均值+标准误差, n = 3)。不同小写字母表示处理间差异显著(p < 0.05)。CK、N50、N100分别表示氮添加量0、50、100 kg·hm-2·a-1。
Fig. 2Influence of nitrogen additions on leaf total carbon (TC), total nitrogen (TN), total phosphorus (TP) content of Quercus wutaishanica in Mt. Dongling, Beijing (mean + SE, n = 3). Different lowercase letters indicate significant difference among treatments (p < 0.05). CK, N50, N100 denote nitrogen addition 0, 50, 100 kg·hm-2·a-1, respectively.
新窗口打开|下载原图ZIP|生成PPT 图3氮添加对北京东灵山辽东栎以及群落水平树木相对增长速率(RGR)变化的影响(平均值+标准误差, n = 3)。不同小写字母表示处理间差异显著(p < 0.05)。CK、N50、N100分别表示氮添加量0、50、100 kg·hm-2·a-1。
Fig. 3Influence of nitrogen addition treatments on the relative growth rate (RGR) of basal area of Quercus wutaishanica and trees at community level in Mt. Dongling, Beijing (mean + SE, n = 3). Different lowercase letters indicate significant difference among treatments (p < 0.05). CK, N50, N100 denote nitrogen addition 0, 50, 100 kg·hm-2·a-1, respectively.
Fig. 4Relationships of relative growth rate (RGR) of Quercus wutaishanica and trees at community level with soil total carbon (TC), total nitrogen (TN) and total phosphorus (TP) content in Mt. Dongling, Beijing. Each dot indicates the mean in each plot. CK, N50, N100 denote nitrogen addition 0, 50, 100 kg·hm-2·a-1, respectively.
Fig. 5Relationships of Quercus wutaishanica leaf total carbon (TC), total nitrogen (TN) and total phosphorus (TP) content with soil total carbon, total nitrogen and total phosphorus content in Mt. Dongling, Beijing. Each dot indicates the mean in each plot. CK, N50, N100 denote nitrogen addition 0, 50, 100 kg·hm-2·a-1, respectively.
新窗口打开|下载原图ZIP|生成PPT 图6氮添加对北京东灵山辽东栎以及群落水平不同径级(DBH)树木相对增长速率(RGR)的影响(平均值+标准误差, n = 3)。不同小写字母表示处理间差异显著(p < 0.05)。3-10、10-20、>20分别表示植物胸径(DBH)范围为3-10、10-20、>20 cm。CK、N50、N100分别表示氮添加量0、50、100 kg·hm-2·a-1。
Fig. 6Influence of nitrogen addition treatments on the relative growth rate (RGR) of different diameter at breast height (DBH) classes of Quercus wutaishanica and trees at community level in Mt. Dongling, Beijing (mean + SE, n = 3). Different lowercase letters indicate significant difference among treatments (p < 0.05). 3-10, 10-20, >20 denote trees diameter at breast height (DBH) range are 3-10, 10-20, >20 cm. CK, N50, N100 denote nitrogen addition 0, 50, 100 kg·hm-2·a-1, respectively.
Fig. 7Relationships of relative growth rate (RGR) of different diameter at breast height (DBH) classes of Quercus wutaishanica in Mt. Dongling, Beijing and trees at community level with soil nitrogen (TN). CK, N50, N100 denote nitrogen addition 0, 50, 100 kg·hm-2·a-1, respectively.
Alvarez-ClareS, MackMC, BrooksM ( 2013). A direct test of nitrogen and phosphorus limitation to net primary productivity in a lowland tropical wet forest Ecology, 94, 1540-1551. [本文引用: 2]
BassiriRadH, LussenhopJF, SehtiyaHL, BordenKK ( 2015). Nitrogen deposition potentially contributes to oak regeneration failure in the Midwestern temperate forests of the USA Oecologia, 177, 53-63. [本文引用: 1]
BergkvistBO, FolkesonL ( 1992). Soil acidification and element fluxes of a Fagus sylvatica forest as influenced by simulated nitrogen deposition Water, Air, & Soil Pollution, 65, 111-133. [本文引用: 1]
BurslemDFRP, GrubbPJ, TurnerIM ( 1995). Responses to nutrient addition among shade-tolerant tree seedlings of lowland tropical rain forest in Singapore Journal of Ecology, 83, 113-122. [本文引用: 2]
ClarkCM, TilmanD ( 2008). Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands Nature, 451, 712-715. [本文引用: 1]
de VriesW, DuEZ, Butterbach-BahlK ( 2014). Short and long- term impacts of nitrogen deposition on carbon sequestration by forest ecosystems Current Opinion in Environmental Sustainability, 9- 10, 90-104. [本文引用: 1]
DengMF, LiuLL, SunZZ, PiaoSL, MaYC, ChenYW, WangJ, QiaoCL, WangX, LiP ( 2016). Increased phosphate uptake but not resorption alleviates phosphorus deficiency induced by nitrogen deposition in temperate Larix principis-?rupprechtii plantations New Phytologist, 212, 1019-1029. [本文引用: 1]
DuEZ, ZhouZ, LiP, HuXY, MaYC, WangW, ZhengCY, ZhuJX, HeJS, FangJY ( 2013). NEECF: A project of nutrient enrichment experiments in China’s forests Journal of Plant Ecology, 6, 428-435. [本文引用: 1]
DuYG, GuoXW, CaoGM, LiYK ( 2016). Increased nitrous oxide emissions resulting from nitrogen addition and increased precipitation in an alpine meadow ecosystem Polish Journal of Environmental Studies, 25, 447-451. [本文引用: 1]
GilliamFS ( 2006). Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition Journal of Ecology, 94, 1176-1191. [本文引用: 1]
GundersenP, EmmettBA, Kj?naasOJ, KoopmansCJ, TietemaA ( 1998). Impact of nitrogen deposition on nitrogen cycling in forests: A synthesis of NITREX data Forest Ecology and Management, 101, 37-55. [本文引用: 2]
H?gbergP, FanHB, QuistM, BinkleyD, TammCO ( 2006). Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest Global Change Biology, 12, 489-499. [本文引用: 1]
JeftsS, FernandezIJ, RustadLE, DailDB ( 2004). Decadal responses in soil N dynamics at the Bear Brook Watershed in Maine, USA Forest Ecology and Management, 189, 189-205. [本文引用: 1]
Kj?naasOJ, StuanesAO, HuseM ( 1998). Effects of weekly nitrogen additions on N cycling in a coniferous forest catchment, G?rdsj?n, Sweden Forest Ecology and Management, 101, 227-249. [本文引用: 2]
KoerselmanW, MeulemanAFM ( 1996). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation Journal of Applied Ecology, 33, 1441-1450. [本文引用: 1]
LiY, TianDS, YangH, NiuSL ( 2018). Size-dependent nutrient limitation of tree growth from subtropical to cold temperate forests Functional Ecology, 32, 95-105. [本文引用: 4]
LuXK, MoJM, DongSF ( 2008). Effects of nitrogen deposition on forest biodiversity: A review Acta Ecologica Sinica, 28, 5532-5548. [本文引用: 1]
MagillAH, AberJD, CurrieWS, NadelhofferKJ, MartinME, McDowellWH, MelilloJM, SteudlerP ( 2004). Ecosystem response to 15 years of chronic nitrogen additions at the Harvard Forests LTER, Massachusetts, USA Forest Ecology and Management, 196, 7-28. [本文引用: 2]
MagnaniF, MencucciniM, BorghettiM, BerbigierP, BerningerF, DelzonS, GrelleA, HariP, JarvisPG, KolariP, KowalskiAS, LankreijerH, LawBE, LindrothA, LoustauD, MancaG, MoncrieffJB, RaymentM, TedeschiV, ValentiniR, GraceJ ( 2007). The human footprint in the carbon cycle of temperate and boreal forests Nature, 447, 849-851. [本文引用: 2]
MatsonP, LohseKA, HallSJ ( 2002). The globalization of nitrogen deposition: Consequences for terrestrial ecosystems AMBIO, 31, 113-119. [本文引用: 1]
McConnaughayKDM, ColemanJS ( 1999). Biomass allocation in plants: Ontogeny or optimality? A test along three resource gradients Ecology, 80, 2581-2593. [本文引用: 1]
NadelhofferKJ, EmmettBA, GundersenP, Kj?naasOJ, KoopmansCJ, SchleppiP, TietemaA, WrightRF ( 1999). Nitrogen deposition makes a minor contribution to carbon sequestration in temperate forests Nature, 398, 145-148. [本文引用: 1]
ReidsmaP, TekelenburgT, van den BergM, AlkemadeR ( 2006). Impacts of land-use change on biodiversity: An assessment of agricultural biodiversity in the European Union Agriculture, Ecosystems & Environment, 114, 86-102. [本文引用: 1]
SolbergS, DobbertinM, ReindsGJ, LangeH, AndreassenK, FernandezPG, HildingssonA, de VriesW ( 2009). Analyses of the impact of changes in atmospheric deposition and climate on forest growth in European monitoring plots: A stand growth approach Forest Ecology and Management, 258, 1735-1750. [本文引用: 1]
StevensCJ ( 2004). Impact of nitrogen deposition on the species richness of grasslands Science, 303, 1876-1879. [本文引用: 1]
SuHX, LiGQ ( 2012). Simulating the response of the Quercus mongolica forest ecosystem carbon budget to asymmetric warming Chinese Science Bulletin, 57, 1544-1552. [本文引用: 1]
SunBH, HuZY, LüJL, ZhouLN, XuCK ( 2006). The leaching solution chemistry of a broad-leaved forest red soil under simulated N deposition in southern China Acta Ecologica Sinica, 26, 1872-1881. [本文引用: 1]
TianD, LiP, FangWJ, XuJ, LuoYK, YanZB, ZhuB, WangJJ, XuXN, FangJY ( 2017). Growth responses of trees and understory plants to nitrogen fertilization in a subtropical forest in China Biogeosciences, 14, 3461-3469. [本文引用: 2]
UriarteM, CanhamCD, ThompsonJ, ZimmermanJK, BrokawN ( 2005). Seedling recruitment in a hurricane-driven tropical forest: Light limitation, density-dependence and the spatial distribution of parent trees Journal of Ecology, 93, 291-304. [本文引用: 1]
VitousekPM, HowarthRW ( 1991). Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry, 13, 87-115. [本文引用: 1]
WangQSY, ZhengCY, ZhangXY, ZengFX, XingJ ( 2016). Impacts of nitrogen addition on foliar nitrogen and phosphorus stoichiometry in a subtropical evergreen broad- leaved forest in Mount Wuyi Chinese Journal of Plant Ecology, 40, 1124-1135. [本文引用: 1]
WrightRF, RasmussenL ( 1998). Introduction to the NITREX and EXMAN projects Forest Ecology and Management, 101, 1-7. [本文引用: 1]
WrightSJ, YavittJB, WurzburgerN, TurnerBL, TannerEVJ, SayerEJ, SantiagoLS, KaspariM, HedinLO, HarmsKE, GarciaMN, CorreMD ( 2011). Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest Ecology, 92, 1616-1625. [本文引用: 1]
Forest biogeochemistry and primary production altered by nitrogen saturation 1 1995
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
Nitrogen saturation in northern forest ecosystems 1 1989
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
A direct test of nitrogen and phosphorus limitation to net primary productivity in a lowland tropical wet forest 2 2013
Short and long- term impacts of nitrogen deposition on carbon sequestration by forest ecosystems 1 2014
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
Increased phosphate uptake but not resorption alleviates phosphorus deficiency induced by nitrogen deposition in temperate Larix principis-?rupprechtii plantations 1 2016
NEECF: A project of nutrient enrichment experiments in China’s forests 1 2013
... 研究地点位于北京东灵山森林生态系统定位研究站(39.97° N, 115.43° E), 海拔1 250 m.该站点为中国森林生态系统养分添加实验网络(NEECF)平台站点之一, 为长期氮添加实验标准区域(Du et al., 2013).该地区属于温带半湿润季风气候, 夏季高温多雨, 冬季寒冷干燥.年降水量(570.3 ± 112.2) mm, 大部分集中在6-8月份; 年平均气温(4.9 ± 0.6) ℃, 最热月(7月)平均气温(18.7 ± 1.0) ℃, 最冷月(1月)平均气温(-10.4 ± 1.5) ℃.棕壤土, 土层厚度为90- 120 cm, pH值为6.3-6.9 (苏宏新和李广起, 2012). ...
Increased nitrous oxide emissions resulting from nitrogen addition and increased precipitation in an alpine meadow ecosystem 1 2016
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition 1 2006
Impact of nitrogen deposition on nitrogen cycling in forests: A synthesis of NITREX data 2 1998
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
Nutrient co-limitation of primary producer communities 1 2011
... 将辽东栎和群落植物分为小(3-10 cm)、中(10-20 cm)和大(>20 cm) 3个径级后, 发现不同径级辽东栎和群落植物对氮添加响应趋势不一致(图6): 氮添加明显抑制了小径级树木的生长, 氮添加对中径级和大径级树木的生长有促进作用, 但响应不显著(Tian et al., 2017; Li et al., 2018).通常认为, 外源元素的输入若超过了植物本身对该元素的同化作用, 不仅会影响植物对该元素的吸收, 同时也会破坏植物体内其他营养元素化学计量耦合关系的平衡, 从而抑制植物生长(Harpole et al., 2011).小径级树木对环境的适应能力以及对外源元素的同化作用和维持化学计量耦合关系的稳定性要弱于中、大径级树木, 而且元素吸收和转化能力比中、大径级树木也更弱.因此, 对元素变化更敏感的小径级树木, 很有可能因为外源元素的输入打乱了化学计量耦合关系的平衡, 从而表现为抑制生长. ...
Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest 1 2006
... 资源的可利用性影响着生态系统的存续和发展, 其中营养元素是所有资源中最重要的一项.当一种或者几种营养元素含量不足或偏低时, 植物的生长会受到抑制.通过外源元素添加能够提高土壤中该营养元素含量, 进而减缓营养元素对生态系统的限制作用(Koerselman & Meuleman, 1996).当植物或生态系统对外源元素添加呈积极响应, 比如促进植物生长或提高地上生物量, 就可认定该元素为植物或生态系统的限制元素(Vitousek & Howarth, 1991; Alvarzclare et al., 2013).我们的研究(图3)发现, 优势树种辽东栎以及群落水平植物的RGR整体上对氮添加呈积极响应(Högberg et al., 2006; BassiriRad et al., 2015), 说明外源氮添加能够促进优势树种以及辽东栎群落的生长.由此推断, 氮可能是辽东栎以及暖温带森林生态系统的限制元素, 本研究结果与传统的元素限制理论一致(Magnani et al., 2007; Vitousek et al., 2010). ...
Decadal responses in soil N dynamics at the Bear Brook Watershed in Maine, USA 1 2004
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
Nitrogenous fertilizer effects on soil structural properties under switchgrass 1 2011
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
Effects of weekly nitrogen additions on N cycling in a coniferous forest catchment, G?rdsj?n, Sweden 2 1998
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation 1 1996
... 资源的可利用性影响着生态系统的存续和发展, 其中营养元素是所有资源中最重要的一项.当一种或者几种营养元素含量不足或偏低时, 植物的生长会受到抑制.通过外源元素添加能够提高土壤中该营养元素含量, 进而减缓营养元素对生态系统的限制作用(Koerselman & Meuleman, 1996).当植物或生态系统对外源元素添加呈积极响应, 比如促进植物生长或提高地上生物量, 就可认定该元素为植物或生态系统的限制元素(Vitousek & Howarth, 1991; Alvarzclare et al., 2013).我们的研究(图3)发现, 优势树种辽东栎以及群落水平植物的RGR整体上对氮添加呈积极响应(Högberg et al., 2006; BassiriRad et al., 2015), 说明外源氮添加能够促进优势树种以及辽东栎群落的生长.由此推断, 氮可能是辽东栎以及暖温带森林生态系统的限制元素, 本研究结果与传统的元素限制理论一致(Magnani et al., 2007; Vitousek et al., 2010). ...
Size-dependent nutrient limitation of tree growth from subtropical to cold temperate forests 4 2018
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
Ecosystem response to 15 years of chronic nitrogen additions at the Harvard Forests LTER, Massachusetts, USA 2 2004
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
The human footprint in the carbon cycle of temperate and boreal forests 2 2007
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
... 资源的可利用性影响着生态系统的存续和发展, 其中营养元素是所有资源中最重要的一项.当一种或者几种营养元素含量不足或偏低时, 植物的生长会受到抑制.通过外源元素添加能够提高土壤中该营养元素含量, 进而减缓营养元素对生态系统的限制作用(Koerselman & Meuleman, 1996).当植物或生态系统对外源元素添加呈积极响应, 比如促进植物生长或提高地上生物量, 就可认定该元素为植物或生态系统的限制元素(Vitousek & Howarth, 1991; Alvarzclare et al., 2013).我们的研究(图3)发现, 优势树种辽东栎以及群落水平植物的RGR整体上对氮添加呈积极响应(Högberg et al., 2006; BassiriRad et al., 2015), 说明外源氮添加能够促进优势树种以及辽东栎群落的生长.由此推断, 氮可能是辽东栎以及暖温带森林生态系统的限制元素, 本研究结果与传统的元素限制理论一致(Magnani et al., 2007; Vitousek et al., 2010). ...
The globalization of nitrogen deposition: Consequences for terrestrial ecosystems 1 2002
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
Biomass allocation in plants: Ontogeny or optimality? A test along three resource gradients 1 1999
Analyses of the impact of changes in atmospheric deposition and climate on forest growth in European monitoring plots: A stand growth approach 1 2009
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
Impact of nitrogen deposition on the species richness of grasslands 1 2004
Nitrogen limitation on land and in the sea: How can it occur? 1 1991
... 资源的可利用性影响着生态系统的存续和发展, 其中营养元素是所有资源中最重要的一项.当一种或者几种营养元素含量不足或偏低时, 植物的生长会受到抑制.通过外源元素添加能够提高土壤中该营养元素含量, 进而减缓营养元素对生态系统的限制作用(Koerselman & Meuleman, 1996).当植物或生态系统对外源元素添加呈积极响应, 比如促进植物生长或提高地上生物量, 就可认定该元素为植物或生态系统的限制元素(Vitousek & Howarth, 1991; Alvarzclare et al., 2013).我们的研究(图3)发现, 优势树种辽东栎以及群落水平植物的RGR整体上对氮添加呈积极响应(Högberg et al., 2006; BassiriRad et al., 2015), 说明外源氮添加能够促进优势树种以及辽东栎群落的生长.由此推断, 氮可能是辽东栎以及暖温带森林生态系统的限制元素, 本研究结果与传统的元素限制理论一致(Magnani et al., 2007; Vitousek et al., 2010). ...
Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions 2 2010
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
... 资源的可利用性影响着生态系统的存续和发展, 其中营养元素是所有资源中最重要的一项.当一种或者几种营养元素含量不足或偏低时, 植物的生长会受到抑制.通过外源元素添加能够提高土壤中该营养元素含量, 进而减缓营养元素对生态系统的限制作用(Koerselman & Meuleman, 1996).当植物或生态系统对外源元素添加呈积极响应, 比如促进植物生长或提高地上生物量, 就可认定该元素为植物或生态系统的限制元素(Vitousek & Howarth, 1991; Alvarzclare et al., 2013).我们的研究(图3)发现, 优势树种辽东栎以及群落水平植物的RGR整体上对氮添加呈积极响应(Högberg et al., 2006; BassiriRad et al., 2015), 说明外源氮添加能够促进优势树种以及辽东栎群落的生长.由此推断, 氮可能是辽东栎以及暖温带森林生态系统的限制元素, 本研究结果与传统的元素限制理论一致(Magnani et al., 2007; Vitousek et al., 2010). ...
氮添加对武夷山亚热带常绿阔叶林植物叶片氮磷化学计量特征的影响 1 2016
... 在2015年8月底(距离上次氮添加时间间隔近一个月), 在每个样方随机取5个点, 用内径为5 cm的土钻采集0-10 cm表层土壤, 带回实验室风干, 在剔除石块和植物残根之后, 粉碎并过2 mm筛, 充分混匀.称取15 g风干土壤, 用50 mL KCl (1 mol·L-1)溶液浸提, 用酸度计(pH S-3B, 上海精科, 上海)测定pH值.另取50 g自然风干的土壤样品, 研磨充分后, 使用元素分析仪(Elementar Vario EL III, Hanau, Germany)测定土壤全氮(TN)和全碳(TC)含量, 用钼锑抗比色法测定土壤全磷(TP)含量(王乔姝怡等, 2016). ...
氮添加对武夷山亚热带常绿阔叶林植物叶片氮磷化学计量特征的影响 1 2016
... 在2015年8月底(距离上次氮添加时间间隔近一个月), 在每个样方随机取5个点, 用内径为5 cm的土钻采集0-10 cm表层土壤, 带回实验室风干, 在剔除石块和植物残根之后, 粉碎并过2 mm筛, 充分混匀.称取15 g风干土壤, 用50 mL KCl (1 mol·L-1)溶液浸提, 用酸度计(pH S-3B, 上海精科, 上海)测定pH值.另取50 g自然风干的土壤样品, 研磨充分后, 使用元素分析仪(Elementar Vario EL III, Hanau, Germany)测定土壤全氮(TN)和全碳(TC)含量, 用钼锑抗比色法测定土壤全磷(TP)含量(王乔姝怡等, 2016). ...
Introduction to the NITREX and EXMAN projects 1 1998
... 元素限制模型是预测全球植被生产力变化的重要工具, 该模型认为氮(N)和磷(P)是限制植物生长的重要元素, 外源氮和磷添加能促进植物生长, 提高植物生产力(Aber et al., 1989; Jung et al., 2011; Du et al., 2016).传统观点认为, 在热带和亚热带等低纬度地区, 树木生长受磷限制, 而暖温带和寒温带等中高纬度区域, 树木生长主要受氮限制(Magnani et al., 2007; Vitousek et al., 2010).在模拟施肥实验中, 前人主要通过监测树木生物量的变化来探究限制元素对树木生长的影响(Gundersen et al., 1998; Wright & Rasmussen, 1998; Jefts et al., 2004; Magill et al., 2004).由于大径级树木的地上生物量对群落生物量的贡献度要大于小径级树木, 以前的研究往往忽视了限制元素对小径级树木生长的影响(Li et al., 2018).其次, 传统限制模型主要从群落水平研究树木生长对限制元素的响应情况, 而忽略了不同物种的响应(Aber et al., 1995; Matson et al., 2002; Solberg et al., 2009; de Vries et al., 2014).Li等(2018)在暖温带和寒温带地区进行的施肥实验表明, 元素的限制作用对小径级树木生长表现明显, 对大径级树木以及整体群落植物生长无显著影响.Magill等(2004)对温带森林的研究指出, 氮添加促进了硬木林植物的生长, 抑制了红松林植物的生长.因此, 需要分径级和分树种探究限制元素对树木生长的影响, 以期对传统的元素限制模型进行补充和完善. ...
Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest 1 2011
 ,*北京大学城市与环境学院, 北京大学生态研究中心, 地表过程分析与模拟教育部重点实验室, 北京 100871
,*北京大学城市与环境学院, 北京大学生态研究中心, 地表过程分析与模拟教育部重点实验室, 北京 100871
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT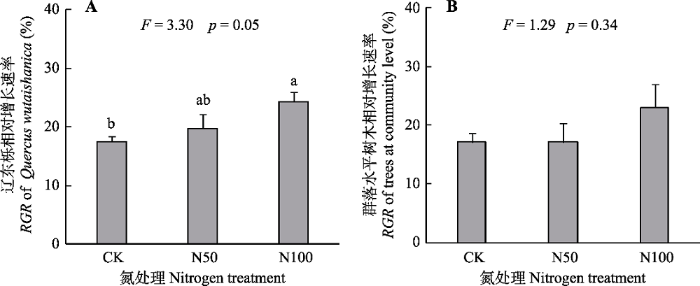 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT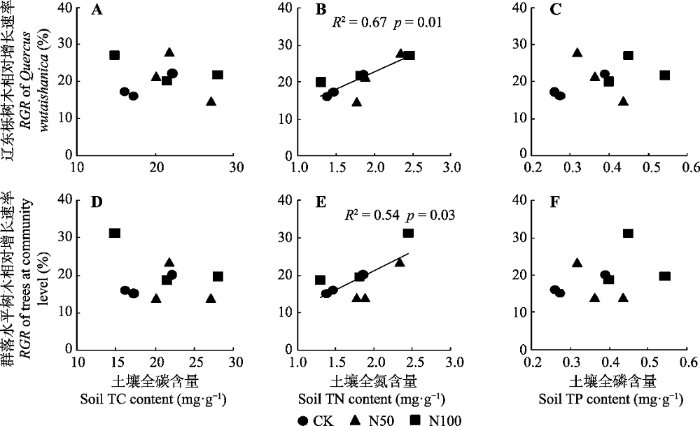 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT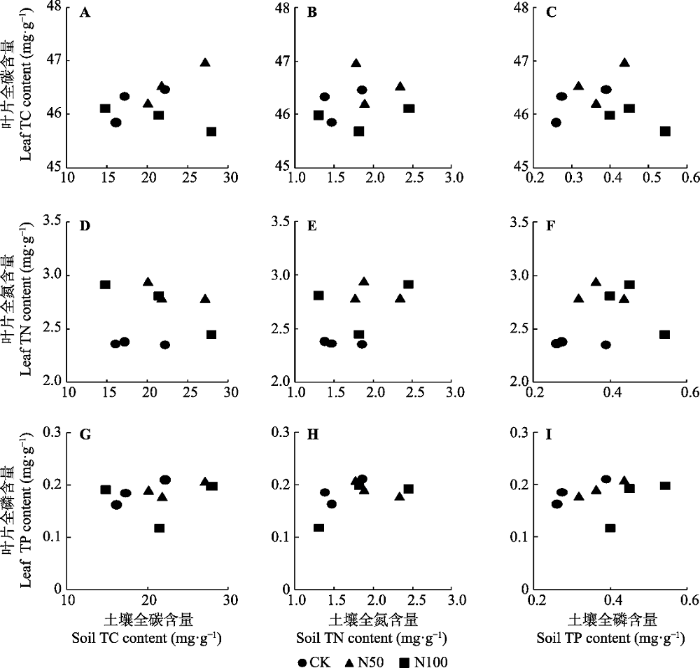 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT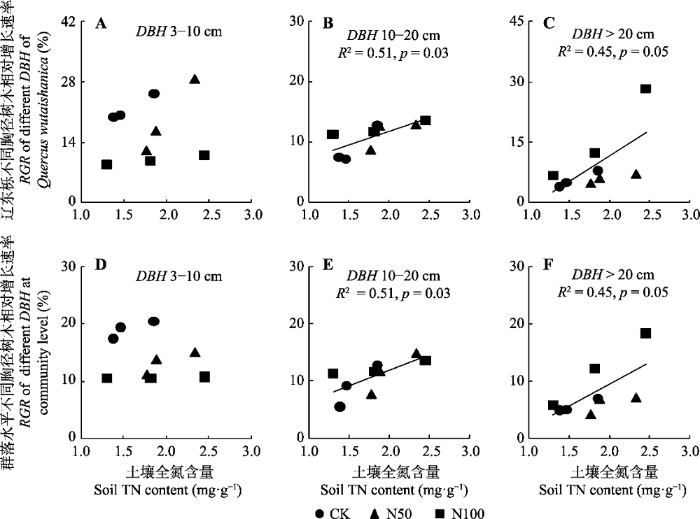 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}