Effects of functional diversity and phylogenetic diversity on the tropical cloud forest community assembly
Yi-Kang CHENG1, Hui ZHANG1, Xu WANG1, Wen-Xing LONG,,1,2,*, Chao LI1, Yan-Shan FANG3, Ming-Qi FU3, Kong-Xin ZHU41 Institute of Tropical Agriculture and Forestry, Hainan University, Haikou 570228, China 2 National Positioning Observation and Research Station of Forest Ecosystem, Wuzhishan, Hainan 572200, China 3 Administration of Limushan Provincial Natural Reserve, Qiongzhong, Hainan 572900, China 4 Forestry Bureau of Bawangling, Changjiang, Hainan 572700, China
Supported by the National Natural Science Foundation of China(31660163) Supported by the National Natural Science Foundation of China(31870508) The Natural Science Foundation of Hainan Province, China(312064) The Natural Science Foundation of Hainan Province, China(2016CXTD003D)
Abstract Aims Phylogenetic and functional diversity are important components of species biodiversity, and are thought to play key roles in the mechanisms of community assembly. In this study, we explore the mechanisms of community assembly in tropical cloud forest plant community in Hainan Island, in southern China, using phylogenetic and functional diversity based methods. Methods We constructed a species pool of 186 woody plant species from three tropical cloud forest sites in Hainan Island. For these species, we measured 13 functional traits and assessed their phylogenetic signals. In addition, we measured seven environmental factors and assessed their relationships using Principal component analysis (PCA). Then we chose Rao’s quadratic entropy (RaoQ) and mean pairwise distance (MPD) indices to examine the effects of functional diversity and phylogenetic diversity on tropical cloud forest community assembly. To do this we compared these indices to expectations under null models that assume neutral community assembly. We used standard effect sizes to evaluate the influence of environmental factors on community assembly. Important findings Canopy openness, soil total phosphorus content and slope were significant environmental predictors in tropical cloud forest. The phylogenetic signals of most functional traits were very low and not significant, indicating that the phylogenetic relationship and functional traits were not consistent with the change of historical process. The observed values of RaoQ and MPD were significantly lower than expected, and their standard effect sizes were significantly correlated with soil phosphorus content, which suggested that habitat filtering driven by soil phosphorus was the key factor driving the community assembly in tropical cloud forest. Keywords:community assembly;functional diversity;phylogenetic diversity;environment factor;tropical cloud forest
PDF (7311KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 程毅康, 张辉, 王旭, 龙文兴, 李超, 方燕山, 符明期, 朱孔新. 功能多样性和谱系多样性对热带云雾林群落构建的影响. 植物生态学报, 2019, 43(3): 217-226. DOI: 10.17521/cjpe.2019.0003 CHENG Yi-Kang, ZHANG Hui, WANG Xu, LONG Wen-Xing, LI Chao, FANG Yan-Shan, FU Ming-Qi, ZHU Kong-Xin. Effects of functional diversity and phylogenetic diversity on the tropical cloud forest community assembly. Chinese Journal of Plant Ecology, 2019, 43(3): 217-226. DOI: 10.17521/cjpe.2019.0003
Table 2 表2 表2热带云雾林环境因子的主成分分析 Table 2Principle component analysis (PCA) among environment factors in tropical cloud forest
环境变量 Environmental variable
PCA1
PCA2
PCA3
PCA4
林冠开阔度 CO (%)
-0.33
-
0.89
-0.24
土壤有机质含量 SOM (g·kg-1)
-0.35
0.47
-0.23
-0.34
全磷含量 TP (g·kg-1)
-0.50
-
-
0.31
全氮含量 TN (g·kg-1)
-0.47
-0.11
-0.32
-0.15
有效氮含量 AN (mg·kg-1)
-0.47
-0.19
-0.19
-0.19
有效磷含量 AP (mg·kg-1)
0.24
0.59
-
-0.48
坡度 SP (o)
0.13
-0.61
-
-0.66
特征值 Characteristic value
3.32
1.87
0.48
0.17
解释方差比例 Explained variance proportion
0.52
0.27
0.10
0.07
累积解释方差比例 Cumulative explained variance proportion
0.52
0.79
0.89
0.96
Data in bold letters indicate that the absolute loadings of environmental variables are over 0.5. CO, canopy openness; SOM, soil organic matter content; TP, total phosphorus content; TN, total nitrogen content; AN, available nitrogen content; AP, available phosphorus content; SP, slope. 加粗数字表示该变量的负荷绝对值大于0.5。
新窗口打开|下载原图ZIP|生成PPT 图2海南岛热带云雾林群落环境因子相关性分析。CO, 林冠开阔度; SOM, 土壤有机质含量; TP, 全磷含量; TN, 全氮含量; AN, 有效氮含量; AP, 有效磷含量; SP, 坡度。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 2Correlation analysis among environmental factors of Hainan tropical cloud forests. CO, canopy openness; SOM, soil organic matter content; TP, total phosphorus content; TN, total nitrogen content; AN, available nitrogen content; AP, available phosphorus content; SP, slope. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Fig. 3Comparison of functional diversity and phylogenetic diversity expectations with observed values in tropical cloud forest. The null distribution is the grey histogram and the observed values are the black vertical bars. MPD, mean pairwise distance; RaoQ, Rao’s quadratic entropy.
2.4 环境因子对群落构建的影响
只有土壤全磷含量对SES.RaoQ和NRI有显著影响(图4A, p = 0.03; 图5A, p = 0.03), 其他环境因子对标准效应值影响都不显著。
Fig. 4Patterns of standard effect size of Rao’s quadratic entropy (SES.RaoQ) along environmental gradients in tropical cloud forest. CO, canopy openness; TP, total phosphorus content; SP, slope.
Fig. 5Patterns of net relatedness index (NRI) along environmental gradients in tropical cloud forest. CO, canopy openness; TP, total phosphorus content; SP, slope.
Agricultural Chemistry Committee of Soil Society of China ( 1983). Agricultural chemical Routine Analysis Method of Soil. Science Press, Beijing. 186-194. [本文引用: 1]
AndersenKM, EndaraMJ, TurnerBL, DallingJW ( 2012). Trait-based community assembly of understory palms along a soil nutrient gradient in a lower montane tropical forest Oecologia, 168, 519-531.
BlombergSP, GarlandT, IvesAR ( 2003). Testing for phylogenetic signal in comparative data: Behavioral traits are more labile Evolution, 57, 717-745. [本文引用: 1]
Botta-DukátZ, CzúczB ( 2016). Testing the ability of functional diversity indices to detect trait convergence and divergence using individual-based simulation Methods in Ecology and Evolution, 7, 114-126. [本文引用: 1]
BuWS, ZangRG, DingY ( 2014). Field observed relationships between biodiversity and ecosystem functioning during secondary succession in a tropical lowland rainforest Acta Oecologica, 55, 1-7. [本文引用: 1]
CadotteMW, CardinaleBJ, OakleyTH ( 2008). Evolutionary history and the effect of biodiversity on plant productivity Proceedings of the National Academy of Sciences of the United States of America, 105, 17012-17017. [本文引用: 2]
Cavender-BaresJ, KozakKH, FinePVA, KembelSW ( 2009). The merging of community ecology and phylogenetic biology Ecology Letters, 12, 693-715. [本文引用: 1]
CBOL Plant WordingGroup ( 2009). A DNA barcode for land plants Proceedings of the National Academy of Sciences of the United States of America, 106, 12794-12797. [本文引用: 1]
CornelissenJHC, LavorelS, GarnierE, DiazS, BuchmannN, GurvichDE, ReichPB, SteegeH, MorganHD, HeijdenA, PausasJG, PoorterH ( 2003). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide Australian Journal of Botany, 51, 335-380. [本文引用: 1]
CornellHV ( 1985). Local and regional richness of cynipine gall wasps on California oaks Ecology, 66, 1247-1260. [本文引用: 1]
CornellHV, HarrisonSP ( 2014). What are species pools and when are they important? Annual Review of Ecology, Evolution,and Systematics, 45, 45-67. [本文引用: 2]
de BelloF, PriceJN, MünkemüllerT, LiiraJ, ZobelM, ThuillerW, GerholdP, G?tzenbergerL, LavergneS, Lep?S, ZobelK, P?rtelM ( 2012). Functional species pool framework to test for biotic effects on community assembly Ecology, 93, 2263-2273.
EllerCB, BurgessSSO, OliveiraRS ( 2015). Environmental controls in the water use patterns of a tropical cloud forest tree species, Drimys brasiliensis (Winteraceae) Tree Physiology, 35, 387-399. [本文引用: 1]
ErikssonO ( 1993). The species-pool hypothesis and plant community diversity Oikos, 68, 371-374. [本文引用: 1]
GoldsmithGR, MatzkeNJ, DawsonTE ( 2013). The incidence and implications of clouds for cloud forest plant water relations Ecology Letters, 16, 307-314. [本文引用: 1]
Gonzalez-CaroS, UmanaMN, AlvarezE, StevensonPR, SwensonNG ( 2014). Phylogenetic alpha and beta diversity in tropical tree assemblages along regional-scale environmental gradients in northwest South America Journal of Plant Ecology, 7, 145-153.
HardyOJ ( 2008). Testing the spatial phylogenetic structure of local communities: Statistical performances of different null models and test statistics on a locally neutral community Journal of Ecology, 96, 914-926. [本文引用: 1]
HarrisonS, CornellH ( 2008). Toward a better understanding of the regional causes of local community richness Ecology Letters, 11, 969-979. [本文引用: 1]
HeJS, WangX, FlynnDFB, WangL, SchmidB, FangJ ( 2009). Taxonomic, phylogenetic, and environmental trade-offs between leaf productivity and persistence Ecology, 90, 2779-2791. [本文引用: 1]
KangY, DengZ, ZangR, LongW ( 2017). DNA barcoding analysis and phylogenetic relationships of tree species in tropical cloud forests Scientific Reports, 7, 12564. DOI: 10.1038/s41598-017-13057-0. [本文引用: 2]
KraftNJB, AckerlyDD ( 2010). Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest Ecological Monographs, 80, 401-422.
KraftNJB, ValenciaR, AckerlyDD ( 2008). Functional traits and niche-based tree community assembly in an Amazonia forest Science, 322, 580-582. [本文引用: 2]
LongWX, SchampBS, ZangRG, DingY, HuangYF, XiangYZ ( 2015 a). Community assembly in a tropical cloud forest related to specific leaf area and maximum species height Journal of Vegetation Science, 47, 416-423. [本文引用: 4]
LongWX, XiongMH, ZangRG, SchampBS, YangXB, DingY, HuangYF, XiangYZ ( 2015 b). Changes in patterns of species co-occurrence across two tropical cloud forests differing in soil nutrients and air temperature Biotropica, 47, 416-423. [本文引用: 2]
LongWX, ZangRG, DingY ( 2011 a). Air temperature and soil phosphorus availability correlate with trait differences between two types of tropical cloud forests Flora, 206, 896-903.
LongWX, ZangRG, SchampBS, DingY ( 2011 b). Within- and among-species variation in specific leaf area drive community assembly in a tropical cloud forest Oecologia, 167, 1103-1113. [本文引用: 2]
LososJB ( 2008). Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species Ecology Letters, 11, 995-1007. [本文引用: 1]
Luna-VegaI, MagallónS ( 2010). Phylogenetic composition of angiosperm diversity in the cloud forests of Mexico Biotropica, 42, 444-454. [本文引用: 1]
MasonNWH, RichardsonSJ, PeltzerDA, BelloFD, WardleDA, AllenRB ( 2012). Changes in coexistence mechanisms along a long-term soil chronosequence revealed by functional trait diversity Journal of Ecology, 100, 678-689. [本文引用: 1]
McGillBJ, EnquistBJ, WeiherE, WestobyM ( 2006). Rebuilding community ecology from functional traits Trends in Ecology & Evolution, 21, 178-185. [本文引用: 1]
McIntireEJB, FajardoA ( 2014). Facilitation as a ubiquitous driver of biodiversity New Phytologist, 201, 403-416. [本文引用: 1]
MoriAS, ShionoT, KoideD, KitagawaR, OtaAT, MizumachiE ( 2013). Community assembly processes shape an altitudinal gradient of forest biodiversity Global Ecology and Biogeography, 22, 878-888.
NaeemS, WrightJP ( 2003). Disentangling biodiversity effects on ecosystem functioning: deriving solutions to a seemingly insurmountable problem Ecology Letters, 6, 567-579. [本文引用: 1]
NiuKC, LiuYN, ShenZH, HeFL, FangJY ( 2009). Community assemble: The relative important of neutral theory and niche theory Biodiversity Science, 17, 579-593. [本文引用: 1]
PaineCET, BaralotoC, ChaveJ, HéraultB ( 2011). Functional traits of individual trees reveal ecological constraints on community assembly in tropical rain forests Oikos, 120, 720-727. [本文引用: 1]
P?rtelM, Szava-KovatsR, ZobelM ( 2011). Dark diversity: Shedding light on absent species Trends in Ecology & Evolution, 26, 124-128.
PearseWD, JonesA, PurvisA ( 2013). Barro Colorado Island’s phylogenetic assemblage structure across fine spatial scales and among clades of different ages Ecology, 94, 2861-2872. [本文引用: 1]
RicklefsRE ( 1987). Community diversity: Relative roles of local and regional processes Science, 235, 206-207. [本文引用: 1]
SpasojevicMJ, SudingKN ( 2012). Inferring community assembly mechanisms from functional diversity patterns: The importance of multiple assembly processes Journal of Ecology, 100, 652-661. [本文引用: 2]
SrivastavaDS ( 1999). Using local-regional richness plots to test for species saturation: Pitfalls and potentials Journal of Animal Ecology, 68, 1-16 [本文引用: 1]
SwensonNG ( 2011). Phylogenetic beta diversity metrics, trait evolution and inferring the functional beta diversity of communities PLOS ONE, 6, e21264. DOI: 10.137/journal.pone.0021264. [本文引用: 1]
SwensonNG ( 2013). The assembly of tropical tree communities-?the advances and shortcomings of phylogenetic and functional trait analyses Ecography, 36, 264-276. [本文引用: 2]
SwensonNG, EnquistBJ ( 2009). Opposing assembly mechanisms in a Neotropical dry forest: Implications for phylogenetic and functional community ecology Ecology, 90, 2161-2170. [本文引用: 1]
SwensonNG, EnquistBJ, PitherJ, ThompsonJ, ZimmermanJK ( 2006). The problem and promise of scale dependency in community phylogenetics Ecology, 87, 2418-2424. [本文引用: 1]
SwensonNG, EricksonDL, MiX, BourgNA, Forero-Monta?aJ, GeXJ, HoweR, LakeJK, LiuXJ, MaKP, PeiNC, ThomsonJ, UriarteM, WolfA, WrightSJ, YeWH, ZhangJL, ZimmermanJK, KressWJ ( 2012). Phylogenetic and functional alpha and beta diversity in temperate and tropical tree communities Ecology, 93, S112-S125. [本文引用: 2]
VamosiSM, HeardSB, VamosiJC, WebbCO ( 2009). Emerging patterns in the comparative analysis of phylogenetic community structure Molecular Ecology, 18, 572-592. [本文引用: 1]
WangXX, LongWX, YangXB, XiongMH, KangY, HuangJ, WangX, HongXJ, ZhouZL, LuYQ, FangJ, LiSX ( 2016). Patterns of plant diversity within and among three tropical cloud forest communities in Hainan Island Chinese Journal of Plant Ecology, 40, 469-479. [本文引用: 1]
WebbCO, AckerlyDD, KembelSW ( 2008). Phylocom: Software for the analysis of phylogenetic community structure and trait evolution Bioinformatics, 24, 2098-2100. [本文引用: 1]
WebbCO, AckerlyDD, McpeekMA, DonoghueMJ ( 2002). Phylogenies and community ecology Annual Review of Ecology and Systematics, 33, 475-505. [本文引用: 5]
WeiherE, KeddyCPA ( 1998). Community assembly rules, morphological dispersion, and the coexistence of plant species Oikos, 81, 309-322.
YangJ, ZhangG, CiX, SwensonNG, CaoM, ShaLQ, LiJ, BaskinCC, SlikJWF, LinLX ( 2014). Functional and phylogenetic assembly in a Chinese tropical tree community across size classes, spatial scales and habitats Functional Ecology, 28, 520-529. [本文引用: 1]
ZobelM, ScheinerS ( 2016). The species pool concept as a framework for studying patterns of plant diversity: Official organ of the International Association for Vegetation Science Journal of Vegetation Science, 27, 8-18. [本文引用: 1]
Field observed relationships between biodiversity and ecosystem functioning during secondary succession in a tropical lowland rainforest 1 2014
... 每个物种采集3株个体, 每株个体采集2-3片成熟的、受太阳光直射的、健康的冠层叶片(Cornelissen et al., 2003), 叶片厚度用数显游标卡尺(SF2000, 广陆, 广西桂林)测量; 叶绿素含量用便携式叶绿素仪(SPAD-502 Plus, Konica Minolta, Tokyo, Japan)测量; 叶面积用叶面积仪(LI-COR 3100C Area Meter, LI-COR, Lincoln, USA)测定.然后将叶片装入信封放于80 ℃烘箱烘干至恒质量, 用电子天平(AR2140, Ohaus, Parsippany, USA)称量干质量.比叶面积由叶面积与干质量的比值算得.在采集叶片样品的同时, 从每个植株上截取4根直径为2-3 cm、长约10 cm的枝条并采取根部样品, 去皮后用排水法测量枝条体积.然后将枝条装入信封放于80 ℃烘箱烘干至恒质量, 称量其干质量, 根据枝条干质量与体积之比计算木材密度.研究发现热带森林树木枝条密度与木材密度显著相关(硬木: R2 = 0.95, p < 0.001; 软木: R2 = 0.93, p < 0.001; Bu et al., 2014), 所以可用植物枝条密度表示木材密度, 如此可避免用生长锥钻取年轮条对树木的伤害.氮含量用凯氏定氮法测定, 磷含量用HClO4-H2SO4消化法分解样品, 然后用钼蓝比色法测量, 可溶性糖含量用蒽酮比色法测定. ...
Evolutionary history and the effect of biodiversity on plant productivity 2 2008
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
... 正是因为基于系统发育的方法存在很大局限性, 很多的生态学家认为基于系统发育关系检验群落构建机制的方法并不合适(Losos, 2008), 也更倾向于使用基于功能性状的检验方法, 甚至提出在功能性状可以测定的条件下, 群落系统发育就不需要考虑了.但是基于功能性状的群落构建机制方法通常就是选择关于群落中物种生态策略或者生理过程相关的一些功能性状(例如形态和结构特征、营养成分等) (Yang et al., 2014), 包括本文也是利用一些营养元素和结构特征的功能性状.但是我们不可能把所有与植物生理生态相关的功能性状全部进行测定, 往往功能性状的选择只是根据一些常用的标准或者在其他研究中得到的结果, 但这些可能并不适用于所有的群落类型; 再者大多数的研究都主要集中于物种水平的功能性状变异, 往往忽略了种内变异情况.由于群落物种间的系统发育关系携带了大量群落历史进化信息, 这些不能由简简单单的功能性状表现出来(Cadotte et al., 2008).因而研究群落构建机制的时候, 同时兼顾群落物种的功能性状和系统发育关系非常有必要. ...
The merging of community ecology and phylogenetic biology 1 2009
Functional species pool framework to test for biotic effects on community assembly 2012
Environmental controls in the water use patterns of a tropical cloud forest tree species, Drimys brasiliensis (Winteraceae) 1 2015
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
The species-pool hypothesis and plant community diversity 1 1993
The incidence and implications of clouds for cloud forest plant water relations 1 2013
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Phylogenetic alpha and beta diversity in tropical tree assemblages along regional-scale environmental gradients in northwest South America 2014
Testing the spatial phylogenetic structure of local communities: Statistical performances of different null models and test statistics on a locally neutral community 1 2008
... 选择平均成对谱系距离MPD谱系多样性指数, MPD指数可以预估在聚类树上所有成对分类单元之间的平均谱系距离(Webb et al., 2008), 可以说明进化树的整体聚类情况(Webb et al., 2002), 而且MPD指数常常是基于物种多度或者基面积加权, 这种特性可以更好地反映生物和非生物因素相互作用的相对重要性(Hardy, 2008; Vamosi et al., 2009).以物种库数据为基础, 构建系统发育树, 保证物种数量和个体数目不变, 系统发育树末端的物种随机置换9 999次, 得到群落MPD的期望值; 用霸王岭样地的功能性状和多度数据计算MPD观测值; 基于R语言中的randomizeMatrix和MPD函数进行计算.运用Wilcoxon test检验期望值与观测值的差异性. ...
Toward a better understanding of the regional causes of local community richness 1 2008
Taxonomic, phylogenetic, and environmental trade-offs between leaf productivity and persistence 1 2009
... 群落构建研究一直是生态学研究的热点, 准确理解和预测驱动群落构建的关键因子并采取针对性的行动可以缓解全球气候变化所带来的问题(Naeem & Wright, 2003; McGill et al., 2006).群落构建机制主要分为两类: 侧重物种生态策略差异的生态位过程和侧重物种相同适合度的中性过程(牛克昌等, 2009).前者包括竞争作用和促进作用等; 后者为随机过程.近年来发现受非生物环境胁迫(如低温、土壤养分缺乏、干旱等)的影响, 促进作用和竞争作用能同时影响同一群落物种共存, 且影响大小随环境胁迫强弱而变化(Kraft et al., 2008; McIntire & Fajardo, 2014; Long et al., 2015a, 2015b).非生物环境不仅可以作为环境筛影响植物形态结构、生理功能和系统发育关系等, 决定群落中共存物种的组成特征; 也可以通过给植物体提供不同数量和质量的资源而影响其生态位, 促进群落物种共存(He et al., 2009). ...
DNA barcoding analysis and phylogenetic relationships of tree species in tropical cloud forests 2 2017
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest 2010
Functional traits and niche-based tree community assembly in an Amazonia forest 2 2008
... 群落构建研究一直是生态学研究的热点, 准确理解和预测驱动群落构建的关键因子并采取针对性的行动可以缓解全球气候变化所带来的问题(Naeem & Wright, 2003; McGill et al., 2006).群落构建机制主要分为两类: 侧重物种生态策略差异的生态位过程和侧重物种相同适合度的中性过程(牛克昌等, 2009).前者包括竞争作用和促进作用等; 后者为随机过程.近年来发现受非生物环境胁迫(如低温、土壤养分缺乏、干旱等)的影响, 促进作用和竞争作用能同时影响同一群落物种共存, 且影响大小随环境胁迫强弱而变化(Kraft et al., 2008; McIntire & Fajardo, 2014; Long et al., 2015a, 2015b).非生物环境不仅可以作为环境筛影响植物形态结构、生理功能和系统发育关系等, 决定群落中共存物种的组成特征; 也可以通过给植物体提供不同数量和质量的资源而影响其生态位, 促进群落物种共存(He et al., 2009). ...
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Community assembly in a tropical cloud forest related to specific leaf area and maximum species height 4 2015
... 群落构建研究一直是生态学研究的热点, 准确理解和预测驱动群落构建的关键因子并采取针对性的行动可以缓解全球气候变化所带来的问题(Naeem & Wright, 2003; McGill et al., 2006).群落构建机制主要分为两类: 侧重物种生态策略差异的生态位过程和侧重物种相同适合度的中性过程(牛克昌等, 2009).前者包括竞争作用和促进作用等; 后者为随机过程.近年来发现受非生物环境胁迫(如低温、土壤养分缺乏、干旱等)的影响, 促进作用和竞争作用能同时影响同一群落物种共存, 且影响大小随环境胁迫强弱而变化(Kraft et al., 2008; McIntire & Fajardo, 2014; Long et al., 2015a, 2015b).非生物环境不仅可以作为环境筛影响植物形态结构、生理功能和系统发育关系等, 决定群落中共存物种的组成特征; 也可以通过给植物体提供不同数量和质量的资源而影响其生态位, 促进群落物种共存(He et al., 2009). ...
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
... ).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
... ), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Changes in patterns of species co-occurrence across two tropical cloud forests differing in soil nutrients and air temperature 2 2015
... 群落构建研究一直是生态学研究的热点, 准确理解和预测驱动群落构建的关键因子并采取针对性的行动可以缓解全球气候变化所带来的问题(Naeem & Wright, 2003; McGill et al., 2006).群落构建机制主要分为两类: 侧重物种生态策略差异的生态位过程和侧重物种相同适合度的中性过程(牛克昌等, 2009).前者包括竞争作用和促进作用等; 后者为随机过程.近年来发现受非生物环境胁迫(如低温、土壤养分缺乏、干旱等)的影响, 促进作用和竞争作用能同时影响同一群落物种共存, 且影响大小随环境胁迫强弱而变化(Kraft et al., 2008; McIntire & Fajardo, 2014; Long et al., 2015a, 2015b).非生物环境不仅可以作为环境筛影响植物形态结构、生理功能和系统发育关系等, 决定群落中共存物种的组成特征; 也可以通过给植物体提供不同数量和质量的资源而影响其生态位, 促进群落物种共存(He et al., 2009). ...
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Air temperature and soil phosphorus availability correlate with trait differences between two types of tropical cloud forests 2011
Within- and among-species variation in specific leaf area drive community assembly in a tropical cloud forest 2 2011
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
... ).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species 1 2008
... 正是因为基于系统发育的方法存在很大局限性, 很多的生态学家认为基于系统发育关系检验群落构建机制的方法并不合适(Losos, 2008), 也更倾向于使用基于功能性状的检验方法, 甚至提出在功能性状可以测定的条件下, 群落系统发育就不需要考虑了.但是基于功能性状的群落构建机制方法通常就是选择关于群落中物种生态策略或者生理过程相关的一些功能性状(例如形态和结构特征、营养成分等) (Yang et al., 2014), 包括本文也是利用一些营养元素和结构特征的功能性状.但是我们不可能把所有与植物生理生态相关的功能性状全部进行测定, 往往功能性状的选择只是根据一些常用的标准或者在其他研究中得到的结果, 但这些可能并不适用于所有的群落类型; 再者大多数的研究都主要集中于物种水平的功能性状变异, 往往忽略了种内变异情况.由于群落物种间的系统发育关系携带了大量群落历史进化信息, 这些不能由简简单单的功能性状表现出来(Cadotte et al., 2008).因而研究群落构建机制的时候, 同时兼顾群落物种的功能性状和系统发育关系非常有必要. ...
Phylogenetic composition of angiosperm diversity in the cloud forests of Mexico 1 2010
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Changes in coexistence mechanisms along a long-term soil chronosequence revealed by functional trait diversity 1 2012
Rebuilding community ecology from functional traits 1 2006
... 群落构建研究一直是生态学研究的热点, 准确理解和预测驱动群落构建的关键因子并采取针对性的行动可以缓解全球气候变化所带来的问题(Naeem & Wright, 2003; McGill et al., 2006).群落构建机制主要分为两类: 侧重物种生态策略差异的生态位过程和侧重物种相同适合度的中性过程(牛克昌等, 2009).前者包括竞争作用和促进作用等; 后者为随机过程.近年来发现受非生物环境胁迫(如低温、土壤养分缺乏、干旱等)的影响, 促进作用和竞争作用能同时影响同一群落物种共存, 且影响大小随环境胁迫强弱而变化(Kraft et al., 2008; McIntire & Fajardo, 2014; Long et al., 2015a, 2015b).非生物环境不仅可以作为环境筛影响植物形态结构、生理功能和系统发育关系等, 决定群落中共存物种的组成特征; 也可以通过给植物体提供不同数量和质量的资源而影响其生态位, 促进群落物种共存(He et al., 2009). ...
Facilitation as a ubiquitous driver of biodiversity 1 2014
... 群落构建研究一直是生态学研究的热点, 准确理解和预测驱动群落构建的关键因子并采取针对性的行动可以缓解全球气候变化所带来的问题(Naeem & Wright, 2003; McGill et al., 2006).群落构建机制主要分为两类: 侧重物种生态策略差异的生态位过程和侧重物种相同适合度的中性过程(牛克昌等, 2009).前者包括竞争作用和促进作用等; 后者为随机过程.近年来发现受非生物环境胁迫(如低温、土壤养分缺乏、干旱等)的影响, 促进作用和竞争作用能同时影响同一群落物种共存, 且影响大小随环境胁迫强弱而变化(Kraft et al., 2008; McIntire & Fajardo, 2014; Long et al., 2015a, 2015b).非生物环境不仅可以作为环境筛影响植物形态结构、生理功能和系统发育关系等, 决定群落中共存物种的组成特征; 也可以通过给植物体提供不同数量和质量的资源而影响其生态位, 促进群落物种共存(He et al., 2009). ...
Community assembly processes shape an altitudinal gradient of forest biodiversity 2013
Disentangling biodiversity effects on ecosystem functioning: deriving solutions to a seemingly insurmountable problem 1 2003
... 群落构建研究一直是生态学研究的热点, 准确理解和预测驱动群落构建的关键因子并采取针对性的行动可以缓解全球气候变化所带来的问题(Naeem & Wright, 2003; McGill et al., 2006).群落构建机制主要分为两类: 侧重物种生态策略差异的生态位过程和侧重物种相同适合度的中性过程(牛克昌等, 2009).前者包括竞争作用和促进作用等; 后者为随机过程.近年来发现受非生物环境胁迫(如低温、土壤养分缺乏、干旱等)的影响, 促进作用和竞争作用能同时影响同一群落物种共存, 且影响大小随环境胁迫强弱而变化(Kraft et al., 2008; McIntire & Fajardo, 2014; Long et al., 2015a, 2015b).非生物环境不仅可以作为环境筛影响植物形态结构、生理功能和系统发育关系等, 决定群落中共存物种的组成特征; 也可以通过给植物体提供不同数量和质量的资源而影响其生态位, 促进群落物种共存(He et al., 2009). ...
群落构建的中性理论和生态位理论 1 2009
... 群落构建研究一直是生态学研究的热点, 准确理解和预测驱动群落构建的关键因子并采取针对性的行动可以缓解全球气候变化所带来的问题(Naeem & Wright, 2003; McGill et al., 2006).群落构建机制主要分为两类: 侧重物种生态策略差异的生态位过程和侧重物种相同适合度的中性过程(牛克昌等, 2009).前者包括竞争作用和促进作用等; 后者为随机过程.近年来发现受非生物环境胁迫(如低温、土壤养分缺乏、干旱等)的影响, 促进作用和竞争作用能同时影响同一群落物种共存, 且影响大小随环境胁迫强弱而变化(Kraft et al., 2008; McIntire & Fajardo, 2014; Long et al., 2015a, 2015b).非生物环境不仅可以作为环境筛影响植物形态结构、生理功能和系统发育关系等, 决定群落中共存物种的组成特征; 也可以通过给植物体提供不同数量和质量的资源而影响其生态位, 促进群落物种共存(He et al., 2009). ...
群落构建的中性理论和生态位理论 1 2009
... 群落构建研究一直是生态学研究的热点, 准确理解和预测驱动群落构建的关键因子并采取针对性的行动可以缓解全球气候变化所带来的问题(Naeem & Wright, 2003; McGill et al., 2006).群落构建机制主要分为两类: 侧重物种生态策略差异的生态位过程和侧重物种相同适合度的中性过程(牛克昌等, 2009).前者包括竞争作用和促进作用等; 后者为随机过程.近年来发现受非生物环境胁迫(如低温、土壤养分缺乏、干旱等)的影响, 促进作用和竞争作用能同时影响同一群落物种共存, 且影响大小随环境胁迫强弱而变化(Kraft et al., 2008; McIntire & Fajardo, 2014; Long et al., 2015a, 2015b).非生物环境不仅可以作为环境筛影响植物形态结构、生理功能和系统发育关系等, 决定群落中共存物种的组成特征; 也可以通过给植物体提供不同数量和质量的资源而影响其生态位, 促进群落物种共存(He et al., 2009). ...
Functional traits of individual trees reveal ecological constraints on community assembly in tropical rain forests 1 2011
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Dark diversity: Shedding light on absent species 2011
Barro Colorado Island’s phylogenetic assemblage structure across fine spatial scales and among clades of different ages 1 2013
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Community diversity: Relative roles of local and regional processes 1 1987
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Using local-regional richness plots to test for species saturation: Pitfalls and potentials 1 1999
Emerging patterns in the comparative analysis of phylogenetic community structure 1 2009
... 选择平均成对谱系距离MPD谱系多样性指数, MPD指数可以预估在聚类树上所有成对分类单元之间的平均谱系距离(Webb et al., 2008), 可以说明进化树的整体聚类情况(Webb et al., 2002), 而且MPD指数常常是基于物种多度或者基面积加权, 这种特性可以更好地反映生物和非生物因素相互作用的相对重要性(Hardy, 2008; Vamosi et al., 2009).以物种库数据为基础, 构建系统发育树, 保证物种数量和个体数目不变, 系统发育树末端的物种随机置换9 999次, 得到群落MPD的期望值; 用霸王岭样地的功能性状和多度数据计算MPD观测值; 基于R语言中的randomizeMatrix和MPD函数进行计算.运用Wilcoxon test检验期望值与观测值的差异性. ...
海南岛3个林区热带云雾林植物多样性变化 1 2016
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
海南岛3个林区热带云雾林植物多样性变化 1 2016
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
Phylocom: Software for the analysis of phylogenetic community structure and trait evolution 1 2008
... 选择平均成对谱系距离MPD谱系多样性指数, MPD指数可以预估在聚类树上所有成对分类单元之间的平均谱系距离(Webb et al., 2008), 可以说明进化树的整体聚类情况(Webb et al., 2002), 而且MPD指数常常是基于物种多度或者基面积加权, 这种特性可以更好地反映生物和非生物因素相互作用的相对重要性(Hardy, 2008; Vamosi et al., 2009).以物种库数据为基础, 构建系统发育树, 保证物种数量和个体数目不变, 系统发育树末端的物种随机置换9 999次, 得到群落MPD的期望值; 用霸王岭样地的功能性状和多度数据计算MPD观测值; 基于R语言中的randomizeMatrix和MPD函数进行计算.运用Wilcoxon test检验期望值与观测值的差异性. ...
... 很多生态学家把热带森林选作验证物种共存机制的理想场所.从空间格局、功能性状、谱系结构等角度阐明了物种多样性的维持机制(Kraft et al., 2008; Paine et al., 2011; Pearse et al., 2013).但这些研究多数局限于低海拔森林, 对高海拔森林(如热带云雾林)较少涉及.国内外热带云雾林研究集中于群落结构、生物多样性、植物对水分利用和养分循环等方面(Goldsmith et al., 2013; Eller et al., 2015), 部分****开展了功能性状特征和系统发育的研究(Luna-Vega & Magallón, 2010; Long et al., 2011b, 2015a; 王茜茜等, 2016; Kang et al., 2017).海南岛的热带云雾林分布在霸王岭、五指山、尖峰岭、黎母山、吊罗山等山顶或山脊, 群落空气温度低, 云雾出现频率高, 风力强劲, 土壤含水量常处于饱和状态; 树木高度和径级小, 树干常弯曲, 小叶革质, 单叶比例高(Long et al., 2011b).在前期研究中, 发现比叶面积小、高度大的物种在热带云雾林有生存优势(Long et al., 2015a), 在热带云雾林低温和低磷环境中, 竞争作用和促进作用同时影响物种共存; 而基于物种比叶面积功能性状数据时发现生境过滤在较大空间尺度上主导了热带云雾林群落构建过程(Long et al., 2015a, 2015b).但是, 功能多样性能根据植物在资源获取、更新繁衍、环境耐受等多个功能性状轴信息, 检验多个生态过程共同作用于群落构建(Spasojevic & Suding, 2012); 而谱系多样性能够利用物种的系统发育状况推测历史因素对现有群落的影响(Webb et al., 2002; Cadotte et al., 2008).因此通过功能性状和谱系多样性探究热带云雾林群落构建机制, 可能会有新的研究发现.在海南霸王岭热带云雾林设置固定样地, 测定物种的功能性状、DNA条形码和群落环境因子, 分析功能多样性和谱系多样性对群落构建的影响.我们假设: 受环境因子影响, 热带云雾林的功能多样性和谱系多样性呈聚集状态. ...
... 选择平均成对谱系距离MPD谱系多样性指数, MPD指数可以预估在聚类树上所有成对分类单元之间的平均谱系距离(Webb et al., 2008), 可以说明进化树的整体聚类情况(Webb et al., 2002), 而且MPD指数常常是基于物种多度或者基面积加权, 这种特性可以更好地反映生物和非生物因素相互作用的相对重要性(Hardy, 2008; Vamosi et al., 2009).以物种库数据为基础, 构建系统发育树, 保证物种数量和个体数目不变, 系统发育树末端的物种随机置换9 999次, 得到群落MPD的期望值; 用霸王岭样地的功能性状和多度数据计算MPD观测值; 基于R语言中的randomizeMatrix和MPD函数进行计算.运用Wilcoxon test检验期望值与观测值的差异性. ...
Community assembly rules, morphological dispersion, and the coexistence of plant species 1998
Functional and phylogenetic assembly in a Chinese tropical tree community across size classes, spatial scales and habitats 1 2014
... 正是因为基于系统发育的方法存在很大局限性, 很多的生态学家认为基于系统发育关系检验群落构建机制的方法并不合适(Losos, 2008), 也更倾向于使用基于功能性状的检验方法, 甚至提出在功能性状可以测定的条件下, 群落系统发育就不需要考虑了.但是基于功能性状的群落构建机制方法通常就是选择关于群落中物种生态策略或者生理过程相关的一些功能性状(例如形态和结构特征、营养成分等) (Yang et al., 2014), 包括本文也是利用一些营养元素和结构特征的功能性状.但是我们不可能把所有与植物生理生态相关的功能性状全部进行测定, 往往功能性状的选择只是根据一些常用的标准或者在其他研究中得到的结果, 但这些可能并不适用于所有的群落类型; 再者大多数的研究都主要集中于物种水平的功能性状变异, 往往忽略了种内变异情况.由于群落物种间的系统发育关系携带了大量群落历史进化信息, 这些不能由简简单单的功能性状表现出来(Cadotte et al., 2008).因而研究群落构建机制的时候, 同时兼顾群落物种的功能性状和系统发育关系非常有必要. ...
The species pool concept as a framework for studying patterns of plant diversity: Official organ of the International Association for Vegetation Science 1 2016
 ,
, ,1,2,*, 李超1, 方燕山3, 符明期3, 朱孔新41
,1,2,*, 李超1, 方燕山3, 符明期3, 朱孔新41 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT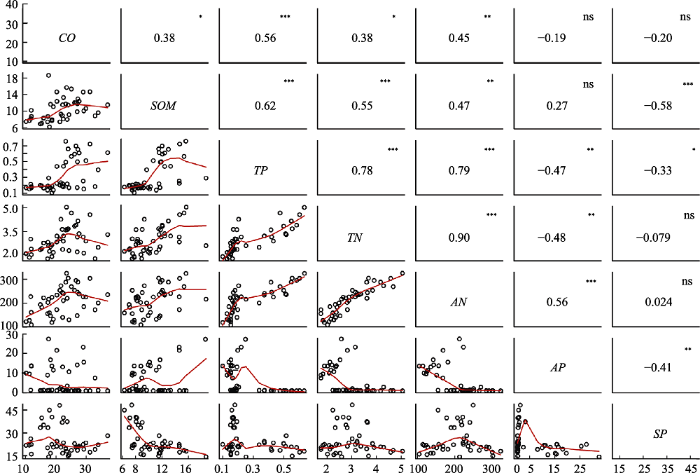 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT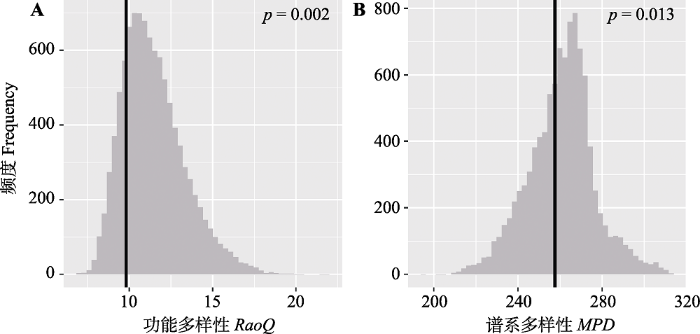 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT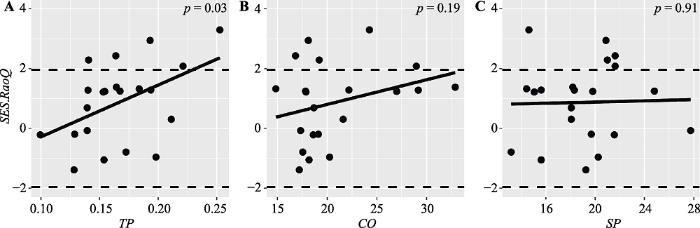 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT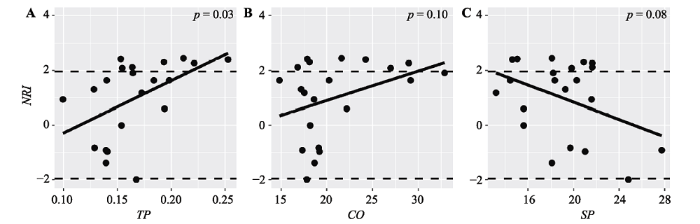 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}