温纯
1 , 金光泽
, , 1 , 2 , * 1 东北林业大学生态研究中心, 哈尔滨 150040 2 东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040 Effects of functional diversity on productivity in a typical mixed broadleaved-Korean pine forest Chun WEN
1 , Guang-Ze JIN
, , 1 , 2 , * 1 Center for Ecological Research, Northeast Forestry University, Harbin 150040, China 2 Key Laboratory of Sustainable Forest Ecosystem Management- Ministry of Education, Northeast Forestry University, Harbin 150040, China 通讯作者: taxus@126.com 编委: 李意德
责任编辑: 李敏
收稿日期: 2018-12-13
接受日期: 2019-02-13
网络出版日期: 2019-02-20
基金资助: 国家自然科学基金 (31730015 )中央高校基本科研业务费专项资金 (2572017EA02 )
Received: 2018-12-13
Accepted: 2019-02-13
Online: 2019-02-20
Fund supported: Supported by the National Natural Science Foundation of China (31730015 )the Fundamental Research Funds for the Central Universities (2572017EA02 )
摘要 为比较生物量比率假说与生态位互补假说在解释生产力变异的相对重要性, 探讨生物多样性和生产力之间的关系是否受到生物和非生物因素的影响, 该研究依托小兴安岭9 hm
2 阔叶红松(
Pinus koraiensis )林动态监测样地, 通过计算群落初始生物量、物种多样性、功能多样性、植物性状的群落加权平均值和测定环境因子, 运用线性回归模型、结构方程模型, 比较了物种多样性和功能多样性与生产力的相关性。结果表明: (1)物种多样性和功能多样性均对生产力有显著作用, 功能多样性比物种多样性与生产力的关系更为密切; (2)功能多样性指数比群落加权平均值能更好地解释生产力变异, 说明生态位互补假说更适用于解释阔叶红松林群落内生产力的变异; (3)生物多样性与生产力的关系受生物因素与非生物因素的共同作用, 相较于多样性和功能性状组成(植被质量), 初始林分生物量(植被数量)能更有效地解释生产力的变异。生物多样性与生产力关系的研究应从植被质量与植被数量同时出发, 评估生态系统过程的多种非生物和生物驱动因素, 同时维护森林功能多样性, 加强植物与土壤环境的保护, 对有效增加生产力和维持生物多样性具有重要意义。
关键词: 功能多样性 ;
森林生产力 ;
生态位互补假说 ;
生物量比率假说 ;
结构方程模型 Abstract Aims Methods 2 typical mixed broadleaved-Korean pine forest. By calculating the initial biomass, species diversity, functional diversity, community-weighted mean functional traits and measuring environmental factors, we analyzed the relationship between diversity and productivity by the linear regression model and structural equation model. Important findings Keywords: functional diversity ;
forest productivity ;
niche complementarity hypothesis ;
mass ratio hypothesis ;
structural equation model PDF (3687KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 温纯, 金光泽. 功能多样性对典型阔叶红松林生产力的影响. 植物生态学报, 2019, 43(2): 94-106. DOI:
10.17521/cjpe.2018.0312 WEN Chun, JIN Guang-Ze.
Effects of functional diversity on productivity in a typical mixed broadleaved-Korean pine forest .
Chinese Journal of Plant Ecology , 2019, 43(2): 94-106. DOI:
10.17521/cjpe.2018.0312 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 )。众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 )。关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 )。以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 )。功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 )。研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 )。
植物多样性影响生态系统功能的机理存在两种假说。一是“生态位互补假说” (
Tilman, 1997 ), 认为随着多样性的增加, 群落内树种的功能特性差异增大, 如叶面积、冠层高度、生长速度等, 能够缓解竞争。通常用多样性指数(如物种、功能多样性)来表征该假说。二是“生物量比率假说” (
Grime, 1998 ), 认为某个时间点的生态系统功能主要由植物优势物种的功能性状值决定。当该假说占主导时, 可以检测到
CWM 是解释生态系统功能的最重要因子(
Tobner et al ., 2016 ;
Ali et al ., 2017 )。尽管目前研究普遍认为生态位互补与生物量比率假说并不相互排斥, 但二者的相对重要性还存在很大争论。
Cardinale和Gonzalez (2011) 整合分析多样性种植实验结果, 发现两种假说对生产力变异的解释基本持平;
Ali等(2017) 则发现生物量比率假说在解释中国东部亚热带次生林的生产力变异中占主导地位;
Ratcliffe等(2016) 发现在欧洲温带森林中生态位互补和生物量比率效应对树木生长均有促进作用, 但在水分受限制的地中海森林中生态位互补假说能够更好地解释生产力的变异。
多样性并非生态系统生产力的唯一驱动因素, 它同时还受非生物因素和其他生物因素的影响, 如初始生物量、土壤养分等环境因子(
Lin et al ., 2012 ;
Chen et al ., 2016 )。有研究认为, 林分的初始地上生物量是森林生产力的关键驱动因素, 是植被数量(即初始生物量)而不是植被质量(即多样性)对生产力变异起着主要作用(
Lohbeck et al ., 2015 )。研究发现单纯考虑多样性与生产力二元关系时, 物种丰富度显著影响生产力, 而整合考虑土壤肥力等非生物因子的作用后, 二者关系变得不明显(
Grace et al ., 2016 ); 与此相反, 当去除其他非生物(海拔、冠层光照)和生物因子(树干密度、森林演替阶段)后, 我国亚热带森林中多样性-生产力正相关关系变弱甚至不相关(
Ouyang et al ., 2016 ); 有研究表明, 土壤因素推动了生产力的变异, 而生态位互补和生物量比率机制在天然林中的重要性却微不足道(
Van et al ., 2017 )。综上来看, 生物因素(植物多样性、初始生物量等)与非生物因素(土壤、地形等)的相对重要性仍需进一步验证。
阔叶红松(
Pinus koraiensis )林是我国东北东部山区的地带性顶极植被, 了解阔叶红松林植物多样性与生产力的关系, 可为次生林的顶极植被恢复提供科学依据。本研究以9 hm
2 典型阔叶红松林动态监测样地为研究对象, 利用线性模型和结构方程模型(SEM), 基于群落地上生产力、地上初始生物量、4个物种多样性指标、11个功能多样性指标、8种植物性状的群落加权平均值以及地形与土壤环境因子数据, 试图回答以下3个问题: (1)功能多样性是否比物种多样性能更好地解释植物群落地上生产力的变异? (2)群落加权平均值(生物量比率假说)与多样性指数(生态位互补假说)哪一个能更好地解释阔叶红松林生产力的变异? (3)多样性与生产力的关系是否受其他生物和非生物因素的共同作用?两者的相对重要性如何?
1 材料和方法 1.1 研究样地概况 调查样地设在黑龙江凉水国家级自然保护区(47.18° N, 128.89° E)。该保护区位于黑龙江省伊春市带岭区, 地处小兴安岭南坡达里带岭支脉东坡, 地形较为复杂, 海拔280-707 m, 为典型的低山丘陵地貌。该区属于温带大陆性季风气候, 年平均气温-0.3 ℃, 年平均最高气温7.5 ℃, 年平均最低气温-6.6 ℃。地带性土壤为暗棕壤, 非地带性土壤为草甸土、沼泽土和泥炭土。本地区的阔叶红松林以红松为建群种, 伴生的温性阔叶树种有水曲柳(
Fraxinus mandschurica )、白桦(
Betula platyphylla )、紫椴(
Tilia amurensis )、色木槭(
Acer mono )、花楷槭(
Acer ukurunduense )等, 寒温性树种以臭冷杉(
Abies nephrolepis )、红皮云杉(
Picea koraiensis )为主, 灌木有毛榛(
Corylus mandshurica )、东北山梅花(
Philadelphus schrenkii )和刺五加(
Acanthopanax senticosus )等。
1.2 数据调查与生产力计算 以10 m × 10 m连续样方为取样单元, 将9 hm
2 阔叶红松林动态监测样地划分为900个样方。于2005年对样地中所有胸径≥2 cm的木本植物的物种、胸径、树高、冠幅、相对位置和状态(存活、倒伏、枯立)进行记录并挂牌监测。样地每隔5年进行复查, 2010年完成第一次复查, 2015年完成第二次复查。2015年复查时样地内胸径≥2 cm的活立木共17 161株, 隶属于20科34属48种。
根据东北主要林木生物量的异速生长方程(
陈传国和朱俊凤, 1989 ), 用实测胸径计算木本植株的地上生物量。群落木本植物生产力(
CWP, Mg·hm
-2 ·a
-1 )的计算参考了
Chave等(2003) 的方法:
CWP = (
G + R )/0.01 ×
T, 式中
G 为10 m × 10 m样方内两次调查都存活个体地上生物量的增长量;
R 为10 m × 10 m样方内第一次调查胸径小于2 cm, 复查时胸径超过2 cm个体的地上生物量总和,
T 为两次调查时间的间隔(10年)。
1.3 叶片功能性状与环境变量测定 本研究测定与植物生长和群落生产力密切相关的8种功能性状: 木材密度、最大树高、比叶面积、叶面积、叶片厚度、叶片干物质含量、叶片碳含量和叶片氮含量(
Finegan et al ., 2015 ;
Van et al ., 2017 ) (
表1 )。对41种木本植物的功能性状进行了测定, 这些物种的生物量总和占群落总生物量的85%以上(
吕亭亭等, 2014 )。每个物种随机选取5株生长良好、无病虫害的植株, 每株剪下20片健康无损、被日光照射无遮阴的叶片, 依据植物性状标准化测定方法(
Cornelissen et al ., 2003 ), 测定6种叶片性状; 在样地周围对每个物种随机选择3株健康植株, 在距离地面1.3 m处钻取一个生长芯, 把生长芯分成1 cm的片段, 用水置换法测定各节段鲜体积, 103 ℃烘干72 h, 用烘干的质量除以鲜体积来测量木材密度; 最大树高来源于《中国植物志》(
Wu & Raven, 1994- 2009 )。
Table 1 表1 表1 与植物生长和群落生产力相关的8种功能性状及其意义
Table 1
Eight functional traits related to plant growth and community woody productivity and their significance 功能性状 Functional trait 功能意义 Functional significance 木材密度 Wood density (WD ) (g·mm-3 ) 木材经济谱, 生长和生存之间的权衡, 水的运输和分配 最大高度 Maximum height (MH ) (m) 植物竞争活力与策略, 光生态位, 结构多样性 比叶面积 Specific leaf area (SLA ) (mm2 ·g-1 ) 叶经济谱, 植物耐荫性 Leaf economic spectrum, plant shade tolerance 叶面积 Leaf area (LA ) (mm2 ) 光竞争, 蒸腾速率 Light acquisition, transpiration rate 叶片厚度 Leaf thickness (LT ) (mm) 储水能力, 蒸腾速率 Storage capacity, transpiration rate 叶片干物质含量 LDMC ) (mg·g-1 ) 养分吸收, 结构物质 Nutrient absorption, structural substance 叶片碳含量 LCC ) (mg·g-1 ) 结构物质, 养分吸收 Structural substance, nutrient absorption 叶片氮含量 LNC ) (mg·g-1 ) 叶经济谱, 氮吸收 Leaf economic spectrum, nitrogen acquisition
新窗口打开 |
下载CSV 地形变量包括凹凸度、海拔和坡度, 将9 hm
2 阔叶红松林动态监测样地划分为225个20 m × 20 m小样方, 样方所有变量均根据每个小样方单元的四个角的高度计算(
Chang et al ., 2013 )。在每个20 m × 20 m样方原点位置, 取1个样点, 同时随机选择一个方向, 在离原点2、5、8 m处选任意2个样点, 取土壤表层(0-10 cm)的样品, 于实验室内测定速效氮、速效磷、速效钾、全氮、全磷、有机碳含量、pH值、土壤容重、体积含水率和质量含水率。我们将每个20 m × 20 m的样方分为了16个5 m × 5 m的子样方后, 应用ArcGIS 10.1通过地统计学分析工具中的ordinary Kriging方法对地形和土壤因子共13项指标进行空间插值, 从而获得每个10 m × 10 m取样单元的生境因子指标(
表2 )。
Table 2 表2 表2 阔叶红松林9 hm
2 样地内基础环境变量信息统计表
Table 2
Summary of environment variables in a 9 hm2 mixed broadleaved-Korean pine forest plot 变量 Variables 范围 Range 平均值 Mean 标准偏差 SD 凹凸度 Convexity -1.91-2.36 0.01 0.40 海拔 Elevation (m) 425.45-505.52 463.25 18.20 坡度 Slope (°) 3.30-38.46 15.91 6.49 速效氮含量 Soil available nitrogen content (mg·kg-1 ) 605.77-1 482.85 1 103.60 161.34 速效磷含量 Soil available phosphorus content (mg·kg-1 ) 2.54-61.96 8.83 6.96 速效钾含量 Soil available potassium content (mg·kg-1 ) 190.46-516.66 332.61 59.05 全氮含量 Soil total nitrogen content (g·kg-1 ) 4.60-12.28 8.40 1.60 全磷含量 Soil total phosphorus content (g·kg-1 ) 0.40-1.19 0.82 0.15 pH值 Soil pH 5.34-6.29 5.77 0.16 有机碳含量 Soil organic carbon content (%) 31.29-216.77 75.75 31.25 土壤容重 Soil bulk density (mg·m-3 ) 0.41-0.87 0.62 0.10 体积含水率 Soil bulk moisture content (%) 13.72-48.40 30.35 5.91 质量含水率 Soil mass moisture content (%) 0.54-1.51 0.91 0.23
新窗口打开 |
下载CSV 1.4 多样性指数与群落加权平均值计算 计算每个样方的物种多样性, 包括物种丰富度(
C )、辛普森指数(
D )、香农指数(
H´ )、均匀度指数(
J ), 各指数计算公式(
孙儒泳等, 2002 )如下:
(1) C = S (2) $D=1-\sum\limits_{i\ =\ 1}^{S}{P_{i}^{2}}$ (3) $P_{i}^{2}=\frac{{{n}_{i}}({{n}_{i}}-1)}{N(N-1)}$ (4) ${H}'=-\sum\limits_{i\ =\ 1}^{S}{{{P}_{i}}\ln {{P}_{i}}}$ (5) $J=H/\ln S$ 式中,
S 为群落中物种数;
N 为所有物种的个体数之和;
Pi 为种
i 的个体在全部个体中的比例;
ni 为第
i 个物种的个体数。
基于所测定的8个植物功能性状, 计算每个小样方的功能均匀度(
FEve )、Rao二次熵指数(
FDQ )、功能分散指数(
FDis ), 计算基于8个功能性状组合的功能分散指数(
FD com )和分别基于单个功能性状的功能分散指数(
FD WD ,
FD MH ,
FD SLA ,
FD LA ,
FD LT ,
FD LDMC ,
FD LCC ), 同时计算了8个性状的群落加权平均值(
CWM )。各指数计算公式(
黄小荣, 2018 )如下:
(6) $E{{W}_{b}}=\frac{{{d}_{ij}}}{{{w}_{i}}+{{w}_{j}}}$ (7) $PE{{W}_{b}}=\frac{E{{W}_{b}}}{\sum\limits_{b-1}^{S-1}{E{{W}_{b}}}}$ (8) $P{{W}_{ve}}=\frac{\sum\limits_{b-1}^{S-1}{\min \left( PE{{W}_{b}},\frac{1}{S-1} \right)-\frac{1}{S-1}}}{1-\frac{1}{S-1}}$ (9) $F{{D}_{Q}}=\sum\limits_{i=1}^{S}{\sum\limits_{i=1}^{S}{{{d}_{ij}}{{w}_{i}}{{w}_{j}}}}$ (10) $F{{D}_{is}}=\frac{\sum{{{w}_{j}}{{z}_{j}}}}{\sum{{{w}_{j}}}}$ (11) $CWM=\sum\limits_{i\ =\ 1}^{S}{{{w}_{i}}\times trai{{t}_{i}}}$ 式中,
S 为物种数目,
PEWb 为偏加权均匀度;
EWb 为加权均匀度;
wi 和
wj 分别为物种
i 和物种
j 的相对多度,
dij 为物种
i 与物种
j 间的欧氏距离;
zj 为物种
j 到加权质心的距离;
traiti 为物种
i 的性状值。
采用R 3.5.1 vegan包的diversity函数(
Oksanen et al ., 2009 ), 计算每个样方的物种多样性指标; FD包计算功能多样性指数与群落加权平均值(
Laliberté & Legendre, 2010 )(
表3 )。
Table 3 表3 表3 阔叶红松林9 hm
2 功能多样性指数、物种多样性指数及群落加权平均值(
CWM )信息统计表
Table 3
Summary of functional diversity, species diversity indexes and community-weighted mean (CWM ) functional traits in a 9 hm2 mixed broadleaved-?Korean pine forest plot 假说 Hypothesis 指数 Index 平均值 Mean 范围 Range 生物量比率假说 木材密度的群落加权平均值 CWM of wood density (CWM WD ) (g·mm-3 ) 0.47 0.34-0.63 最大树高的群落加权平均值 CWM of maximum height (CWM MH ) (m) 17.24 4.59-43.01 比叶面积的群落加权平均值 CWM of specific leaf area (CWM SLA ) (mm2 ·g-1 ) 254.30 158.29-392.23 叶面积的群落加权平均值 CWM of leaf area (CWM LA ) (mm2 ) 33.87 8.20-74.54 叶片厚度的群落加权平均值 CWM of leaf thickness (CWM LT ) (mm) 0.18 0.08-0.44 叶干物质含量的群落加权平均值 CWM of leaf dry matter content (CWM LDMC ) ( mg·g-1 ) 0.29 0.19-0.38 叶片碳含量的群落加权平均值 CWM of leaf carbon content (CWM LCC ) ( mg·g-1 ) 499.22 446.90-563.74 叶片氮含量的群落加权平均值 CWM of leaf nitrogen content (CWM LNC ) ( mg·g-1 ) 23.56 17.49-32.38 生态位互补假说 物种丰富度 Species richness (C ) 6.42 2-13 香农指数 Shannon index (H° ) 1.45 0.41-2.24 辛普森指数 Simpson index (D ) 0.71 0.24-0.88 均匀度指数 Evenness index (J ) 0.89 0.42-1.00 多维功能均匀度指数 Multidimensional functional evenness index (FEve ) 0.72 0.21-0.99 Rao二次熵指数 Rao’s quadratic entropy index (FD Q ) 8.39 0.81-15.41 8个性状组合的功能分散指数 FD is based on eight traits combined (FD com ) 2.68 0.72-3.87 木材密度的功能分散指数 FD is based on wood density (FD WD ) 0.48 0.01-1.13 最大高度的功能分散指数 FD is based on maximum height (FD MH ) 0.78 0.07-1.71 比叶面积的功能分散指数 FD is based on specific leaf area (FD SLA ) 0.63 0.02-1.61 叶面积的功能分散指数 FD is based on leaf area (FD LA ) 0.66 0.02-1.61 叶片厚度的功能分散指数 FD is based on leaf thickness (FD LT ) 0.58 0.03-1.51 叶干物质含量的功能分散指数 FD is based on leaf dry matter content (FD LDMC ) 0.58 0.06-1.26 叶片碳含量的功能分散指数 FD is based on leaf carbon content (FD LCC ) 0.93 0.03-2.37 叶片氮含量的功能分散指数 FD is based on leaf nitrogen content (FD LNC ) 0.51 0.03-2.38
Community weight mean for each trait is the same as the unit of functional trait corresponding to Table 1 . 各个性状的群落加权平均值单位与表1 所对应的功能性状单位相同。 新窗口打开 |
下载CSV 1.5 数据分析 回归分析前, 为了使响应变量呈正态分布, 对其进行对数转换。用
Z -score方法对解释变量进行标准化处理, 使所有变量的范围在类似的尺度上具有可比性。利用转换后的响应变量和解释变量构建线性回归模型。采用R 3.5.1进行数据处理和统计分析。
1.5.1 多重共线性检验
多重共线性会影响解释变量对响应变量的解释和预测能力。利用方差膨胀因子(
VIF )检验变量间的多重共线性, 根据以往经验,
VIF < 10时认为变量间没有多重共线性(
Fox & Monette, 1992 ;
Fox, 2008 )。验证结果显示, 解释变量均匀度指数(
J )、叶片厚度的群落加权平均值(
CWM LT )和叶片厚度的功能分散指数(
FD LT )存在多重共线性, 故删去上述变量。
1.5.2 回归线性模型构建
将生产力作为响应变量, 物种多样性指数、功能多样性指数、物种多样性指数及功能多样性指数、群落加权平均值、功能多样性指数及群落加权平均值、环境变量作为解释变量, 分别与生产力构建线性回归模型, 并比较各模型中解释变量与生产力的相关性。
对于包含不同指数的6个模型, 利用决定系数(
R ²)和赤池信息量准则(AIC)判断模型优度, 比较物种多样性、功能多样性与生产力的相关性, 判断物种多样性与功能多样性对森林生产力影响的差异。
1.5.3 变量筛选
采用MuMIn包中的dredge函数(
Bartoń, 2018 ), 分别将生产力与多样性指数的模型、生产力与群落加权平均值的模型、生产力与环境变量的模型的各个变量之间所有可能的组合进行拟合, 计算出每个回归模型的变量组合中各单一子模型在模型中对响应变量解释的重要性。在拟合的模型中筛选出AIC差值小于2的所有模型, 选择变量重要性最高、与生产力最为相关的多样性指数与环境因子, 用来构建结构方程模型。
1.5.4 结构方程模型构建
利用结构方程模型分析多样性指数、环境因子与森林生产力的直接和间接关系, 并比较生物量比率假说与生态位互补假说在解释阔叶红松林生产力变异的相对重要性。在构建模型之前, 需要一个先验假设。在多样性和生产力之间已知理论的因果关系的基础上构建初始模型, 同时考虑初始林分地上生物量的影响和环境因素的影响。假设: (1)地形和土壤条件在决定生物因子中起着基础性作用; (2)地形对土壤产生影响; (3)多样性和群落加权平均值直接影响初始林分生物量和生产力; (4)初始林分生物量直接影响森林生产力。
根据变量筛选结果, 选择变量重要性最高、与生产力最为相关的多样性指数与潜在环境变量, 纳入到结构方程模型中。采用最大似然法对结构方程模型进行拟合, 利用比较拟合指数(
CFI )、标准均方根残差(
SRMR )、渐进残差均方和平方根(
RMSEA )、显著性概率值(
p )进行模型优度的评价(
Howard, 2013 )。拟合优度的临界值为
CFI > 0.9,
SRMR < 0.05,
RMSEA < 0.08,
p > 0.05。利用模型中各路径标准化系数, 定量表示不同因素对生产力的相对作用大小, 量化各个变量对森林地上生产力的直接、间接和总标准化效应(
Edwards & Lambert, 2007 ), 从而比较生态位互补假说与生物量比率假说对阔叶红松林的适用性。上述分析利用lavaan包完成(
Rosseel, 2012 )。
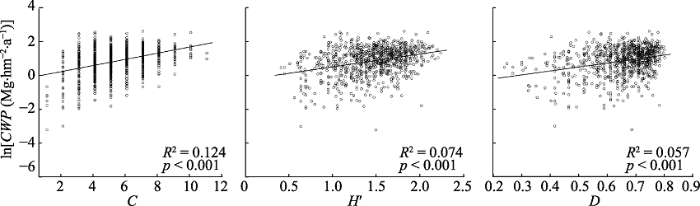
2 结果和分析 2.1 生产力与多样性、环境因子的相关性 回归分析表明, 物种丰富度、香农指数、辛普森指数与生产力极显著正相关(
p < 0.001)(
图1 ); 功能多样性指标(包括10个功能多样性指数和7个群落加权平均值)中功能均匀度指数与生产力无显著的相关关系(
p = 0.163), 其余功能多样性指数和群落加权平均值与生产力显著相关(
图2 ,
图3 ); 土壤因子中有机碳含量、全磷含量、体积含水率与生产力显著相关(
p < 0.05), 地形因子中凹凸度、海拔与生产力显著相关(
p < 0.05)(
图4 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1典型阔叶红松林群落地上生产力(ln转换后)与物种多样性指数的关系。CWP 、C 、H °、D 分别表示群落地上生产力、物种丰富度、香农指数和辛普森指数。 Fig. 1Relationships between coarse woody productivity (ln transformed) and species diversity index of communities in a typical mixed broadleaved-Korean pine forest. CWP, C, H °, D represent coarse woody productivity, species richness, Shannon index and Simpson index, respectively. 图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2典型阔叶红松林群落地上生产力(ln转换后)与功能多样性指数的关系。CWP, 群落地上生产力; FEve, 多维功能均匀度指数; FDQ , Rao二次熵指数; FD com , 8个性状组合的功能分散指数; FD WD , 木材密度的功能分散指数; FD MH , 最大高度的功能分散指数; FD SLA , 比叶面积的功能分散指数; FD LA , 叶面积的功能分散指数; FD LDMC , 叶干物质含量的功能分散指数; FD LCC , 叶片碳含量的功能分散指数; FD LNC , 叶片氮含量的功能分散指数。 Fig. 2Relationships between coarse woody productivity (ln transformed) and functional diversity index of communities in a typical mixed broadleaved-Korean pine forest. CWP, coarse woody productivity; FEve, multidimensional functional evenness index; FDQ , Rao’s quadratic entropy index; FD com , functional dispersion indices of eight trait combinations; FD WD , functional dispersion indices of wood density; FD MH , functional dispersion indices of maximum height; FD SLA , functional dispersion indices of specific leaf area; FD LA , functional dispersion indices of leaf area; FD LDMC , functional dispersion indices of leaf dry matter content; FD LCC , functional dispersion indices of leaf carbon content; FD LNC , functional dispersion indices of leaf nitrogen content. 图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3典型阔叶红松林群落地上生产力(ln转换后)与群落加权平均值的关系。CWP, 群落地上生产力; CWM WD , 木材密度的群落加权平均值; CWM MH , 最大树高的群落加权平均值; CWM SLA , 比叶面积的群落加权平均值; CWM LA , 叶面积的群落加权平均值; CWM LDMC , 叶干物质含量的群落加权平均值; CWM LCC , 叶片碳含量的群落加权平均值; CWM LNC , 叶片氮含量的群落加权平均值。 Fig. 3Relationships between coarse woody productivity (ln transformed) and community weighted mean of different traits of communities in a typical mixed broadleaved-Korean pine forest. CWP, coarse woody productivity; CWM WD , CWM of wood density; CWM MH , CWM of maximum height; CWM SLA , CWM of specific leaf area; CWM LA , CWM of leaf area; CWM LDMC , CWM of leaf dry matter content; CWM LCC , CWM of leaf carbon content; CWM LNC , CWM of leaf nitrogen content. 图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4典型阔叶红松林群落地上生产力(ln转换后, CWP )与各环境因子的关系。 Fig. 4Relationships between coarse woody productivity (ln transformed, CWP ) and environmental factors in a typical mixed broadleaved-Korean pine forest. 2.2 多样性-生产力模型评价 多样性与生产力的线性回归模型中, 功能多样性指数与群落地上生产力的多元回归模型(模型A)、群落加权平均值与群落地上生产力的多元回归模型(模型B)、功能多样性指数及群落加权平均值与群落地上生产力的多元回归模型(模型C)的决定系数(
R ² = 0.269,
R ² = 0.188,
R ² = 0.367)均高于物种多样性指数与群落地上生产力的多元回归模型的决定系数(
R ² = 0.169), 而模型A、B、C的AIC值(AIC = 2 047.73, AIC = 2 136.52, AIC = 1 932.43)均低于物种多样性指数模型的AIC值(AIC = 2 148.61)(
表4 )。
Table 4 表4 表4 典型阔叶红松林群落生产力与多样性构建的线性回归模型评价
Table 4
Evaluation of linear regression models for forest productivity and diversity in a typical mixed broadleaved-Korean pine forest 模型参数 物种多样性模型 功能多样性模型 Functional diversity models A B C R2 0.169 0.269 0.188 0.377 AIC 2 148.61 2 047.73 2 136.52 1 932.43
Functional diversity and CWM model include three independent models. Model A represents multiple regression models of functional diversity index and coarse woody productivity, Model B represents multiple regression models of community weight mean and coarse woody productivity, Model C represents multiple regression models of function diversity index, community-weight mean and coarse woody productivity. R 2 represents coefficient of determination; AIC represents the Akaike information criterion. 表中物种多样性模型包含了环境因子作为解释变量。功能多样性与群落加权平均值模型中包含A、B、C三个独立模型(包含了环境因子作为解释变量), A模型表示功能多样性指数与群落地上生产力的多元回归模型, B模型表示群落加权平均值与群落地上生产力的多元回归模型, C模型表示功能多样性指数及群落加权平均值与群落地上生产力的多元回归模型。R 2 表示决定系数; AIC表示赤池信息量准则。 新窗口打开 |
下载CSV 2.3 线性模型筛选解释生产力的最佳变量 当变量较多时, 结构方程模型不易收敛, 故逐个加入多样性指标及环境变量构建结构方程模型, 构建出最优模型。经过变量筛选比较(
表5 ), 最终以
FDQ 代表多样性, 叶片碳含量的群落加权平均值(
CWM LCC )代表群落加权平均值, 海拔代表地形变量, 土壤全磷含量代表土壤变量, 构建最优结构方程模型(
图5 ), 最优结构方程模型的
CFI = 1.000,
SRMR = 0.011,
RMSEA = 0.017,
p 值为0.260。
Table 5 表5 表5 与生产力构建的线性模型中各变量相对重要性筛选结果
Table 5
Relative importance of variables in linear model constructed with productivity 指标 解释变量 Explanatory variable CWM MH CWM LCC CWM SLA CWM LNC CWM LA CWM WD CWM LDMC 变量重要性 1.00 1.00 0.90 0.67 0.48 0.39 0.14 含变量模型数 10 10 9 6 5 4 2 指标 解释变量 Explanatory variable FD MH FD Q FD com FD SLA C D H° FD LA FD WD FD LCC FEve FD LDMC 变量重要性 1.00 1.00 0.88 0.88 0.88 0.69 0.69 0.69 0.69 0.36 0.21 0.19 含变量模型数 8 8 7 7 7 5 5 5 5 3 2 2 指标 解释变量 Explanatory variable 海拔 全磷含量 凹凸度 速效磷含量 有机碳含量 速效氮含量 体积含水率 坡度 pH 变量重要性 1.00 0.95 0.85 0.68 0.53 0.26 0.21 0.09 0.04 含变量模型数 17 16 14 11 9 4 4 2 1
Explanatory variables see Table 3 . 解释变量同表3 。 新窗口打开 |
下载CSV 图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5结构方程模型分析环境因子、功能多样性、群落加权平均值和森林初始生物量对典型阔叶红松林群落生产力影响的结果。图中实线表示作用路径显著, 虚线表示作用路径不显著。箭头旁边的值是具有相应统计意义的标准化路径系数。R ²表示由组合自变量解释的因变量的总变化。***, p < 0.001; **, p < 0.01; *, p < 0.05。 Fig. 5Structural equation model (SEM) analysis of the effects of environmental factors, functional diversity, community-weight mean of functional traits and forest initial biomass on community woody productivity in a typical mixed broadleaved-Korean pine forest. Solid lines indicate significant paths, while dashed lines indicate insignificant paths. R ² indicates the total variation in a dependent variable that is explained by the combined independent variables. ***, p < 0.001; **, p < 0.01; *, p < 0.05. 2.4 各因素对群落生产力的影响 各变量解释地上生物量变化的17.6%, 解释群落生产力变异的37.2% (
图5 )。功能多样性指数对生产力有极显著的直接影响(
p < 0.001), 反映作用大小的路径系数为0.302; 群落加权平均值对生产力具有显著的直接影响(
p = 0.002, 路径系数为0.090),且通过地上初始生物量具有极显著的间接影响(
p < 0.001, 路径系数为0.152), 总效应(直接和间接效应)极显著(
p < 0.001), 路径系数为0.242 (
表6 )。同时, 生产力还受地形因子和土壤因子的直接影响, 作用大小分别为-0.102和0.105; 生产力受地上初始生物量的极显著影响(
p < 0.001), 路径系数为0.449 (
图5 ;
表6 )。
Table 6 表6 表6 基于结构方程模型森林地上生产力的直接、间接和总标准化效应
Table 6
Direct, indirect, and total standardized effects on the forest productivity, based on the structural equation models (SEM) 解释变量 对生产力的作用途径 结构方程模型 SEM 影响效应 Effect p 地形因子 Topography factors 直接效应 Direct effect -0.102 0.003 功能多样性的间接效应 Indirect effect via functional diversity -0.027 0.009 群落加权平均值的间接效应 Indirect effect via CWMs 0.008 0.043 初始生物量的间接效应 Indirect effect via initial biomass 0.055 <0.001 土壤因子的间接效应 Indirect effect via soil factors -0.024 0.002 总效应 Total effect -0.090 <0.001 土壤因子 Edaphic factors 直接效应 Direct effect 0.105 <0.001 功能多样性的间接效应 Indirect effect via functional diversity -0.021 0.045 群落加权平均值的间接效应 Indirect effect via CWMs -0.015 0.008 总效应 Total effect 0.069 0.019 功能多样性 Functional diversity 直接效应 Direct effect 0.302 <0.001 初始生物量的间接效应 Indirect effect via initial biomass 0.081 <0.001 总效应 Total effect 0.383 <0.001 群落加权平均值 直接效应 Direct effect 0.090 0.002 初始生物量的间接效应 Indirect effect via initial biomass 0.152 <0.001 总效应 Total effect 0.242 <0.001 地上初始生物量 Initial standing biomass 直接效应 Direct effect 0.449 <0.001 总效应 Total effect 0.449 <0.001
新窗口打开 |
下载CSV 3 讨论 3.1 功能多样性比物种多样性与生产力的关系更密切 越来越多的研究表明, 生态系统过程是由物种的功能特性、物种的丰富度和物种的分布共同决定的(
McGill et al ., 2006 ;
Mouchet et al ., 2010 ), 同时森林生产力随多样性的增加而增加(
Ruiz-Benito et al ., 2014 )。本研究得出相似的结果: 群落中物种丰富度越高、功能性状差异越大, 其生产力越高(
图1 )。我们的结果同样表明, 相较于物种多样性, 功能多样性与生产力的关系更加密切(
表4 ), 功能多样性比物种多样性更好地解释了该群落生产力的变异。这可能是因为物种数量和物种多样性所包含的关于物种的特征信息少(
Petchey et al ., 2004 ), 或者忽略了物种间功能性状的相似性和差异性(
吕亭亭等, 2014 )。
Petchey等(2004) 首次证明功能多样性比物种多样性能更有效地预测生物量的变化,
Gazol和Camarero (2016) 指出功能多样性与生态系统生产力的关系非常密切, 功能多样性是解释生态系统功能变化的更有效的指标。相对于物种多样性, 功能多样性有两个优点: 首先, 各物种具有相异的功能特征, 物种多样性无法解释物种间功能特性的差异, 而功能多样性能够综合不同物种的属性(
Micheli & Halpern, 2010 ); 其次, 功能性状可以表征生物从环境中利用资源的能力(
McGill et al ., 2006 )。功能性状的多样性比物种多样性更好地反映不同物种的生态位, 因此能够更有效地揭示生产力的变异规律(
Petchey et al ., 2004 )。
3.2 生态位互补效应比生物量比率效应更适于解释阔叶红松林生产力的变异 众多研究表明, 有关多样性对木本植物生物量及其生产力的作用机理, 既有生物量比率效应也有生态位互补效应(
Ruiz-Benito et al ., 2014 ;
Mensah et al ., 2016 ), 但两种机制在生态系统过程中的作用不相等(
Mokany et al ., 2008 )。本研究中Rao二次熵指数、功能分散指数在解释该群落生产力变异的能力上明显优于群落加权平均值(
图2 ;
表4 ); 在解释多样性-生产力关系的最优结构方程模型中, 两者均与生产力显著相关, 功能多样性指数的总效应(0.383)高于群落加权平均值的总效应(0.242)(
表6 )。说明生态位互补效应和生物量比率效应共存, 前者更适于解释阔叶红松林生产力的变异。
群落加权平均值用于评估优势性状对生产力的影响, 而功能多样性指数则是量化该性状的变异性对生产力的影响(
Tobner et al ., 2016 )。在草本植物生态系统中, 群落加权平均值能更好地解释生产力的变异(
Mokany et al ., 2008 ), 而在欧洲地中海森林(
Ratcliffe et al ., 2016 )和广西马尾松(
Pinus massoniana )林(
黄小荣, 2018 )则发现功能多样性指数更好地解释了生产力的变异, 本研究结果与之一致。这种不同群落类型的多样性-生产力关系结果的不一致, 可能是方法上的差异, 包括选择不同的样地监测周期和多样性指数(
Dănescu et al ., 2016 ); 也可能是由不同生态系统类型和演替阶段造成的。当森林的木本植物存在较大的功能性状差异时, 各物种具有不同获取和利用资源的策略, 减少了生态位重叠, 生态位互补效应会减弱物种间对资源的竞争, 最大化地利用光等有限资源, 从而提高生态系统生产力(
Díaz et al ., 2007 )。而在有生境过滤和扩散限制的群落中, 优势物种的影响可能更大, 生态系统过程主要取决于群落中优势物种的功能性状, 此时生物量比率效应更为重要(
Finegan et al ., 2015 )。
3.3 生物多样性与生产力的关系受到生物和非生物因素的共同作用 生物多样性与生产力之间的关系直接和间接地受多个因素的作用(
Lohbeck et al ., 2015 )。本研究运用结构方程模型评价了功能多样性、地上初始生物量、土壤养分和地形条件对生产力的相对贡献。结果显示, 地上初始生物量对生产力作用的总效应为0.449 (
表6 ), 在各因子中与生产力的相关路径系数最大, 表明地上初始生物量是森林生产力最重要的内在驱动因素, 即植被数量(即林分生物量)而非植被质量(即功能多样性和功能性状组成)是决定森林生产力的主要因素。地形因子对土壤因子的作用效应为-0.230, 并通过多样性、初始生物量对生产力产生间接影响, 其对生产力的总效应为-0.090, 这可能是由于海拔较高生境土壤水分和养分条件较差, 导致森林生产力下降。
在相对资源受限的环境中, 各物种采取不同的资源吸收策略才能共存, 此时互补效应为生产力的主要驱动力。在生物量较高的地区, 丰富的资源导致互补效应的相对重要性降低, 多样性对生产力的影响也将减弱(
Lohbeck et al ., 2015 )。当群落中出现非生物过滤(如环境过滤)时, 只有适应相应环境条件的物种才能够生存, 导致物种间的性状差异减少, 形成性状聚集(
Schellberg & Pontes, 2012 ), 致使局域范围内群落的优势物种表现出强烈的趋同现象。因此, 生物多样性并非生态系统生产力的唯一驱动因素, 而是受非生物因素和其他生物因素的共同影响(
Lohbeck et al ., 2015 )。
在小兴安岭阔叶红松林中, 物种多样性、功能多样性对森林生产力均有一定的促进作用; 功能多样性能够更好地解释群落生产力的变异。功能多样性对生产力的效应主要通过Rao二次熵指数、基于最大树高的功能分散指数产生; 群落加权平均值效应主要由叶片碳含量、比叶面积的群落加权平均值产生; 功能多样性的效应大于群落加权平均值效应。生物因素与非生物因素共同调控多样性-生产力关系。本研究的结果为更好地了解温带森林生态系统中生物多样性与生产力之间的复杂关系提供了科学依据, 对有效增加生产力具有重要意义。
责任编委: 李意德 责任编辑: 李 敏
[1] Ali A Yan ER Chang SX Cheng JY Liu XY 2017 ). Community-weighted mean of leaf traits and divergence of wood traits predict aboveground biomass in secondary subtropical forestsScience of the Total Environment , 574, 654 -662 . DOI:
10.1016/j.scitotenv.2016.09.022 URL [本文引用: 2] [2] Bartoń K 2018 ). Package ‘MuMIn’ URL [本文引用: 1] [3] Cardinale BJ Duffy JE Gonzalez A Hooper DU Perrings C Venail P 2012 ). Biodiversity loss and its impact on humanityNature , 489, 59 -67 . [本文引用: 1] [4] Cardinale BJ Gonzalez A 2011 ). The functional role of producer diversity in ecosystemsAmerican Journal of Botany , 98, 572 -592 . DOI:
10.3732/ajb.1000364 URL [本文引用: 1] [5] Chang LW Zeleny D Li CF Chiu ST Hsieh CF 2013 ). Better environmental data may reverse conclusions about niche- and dispersal-based processes in community assemblyEcology , 94, 2145 -2151 . DOI:
10.1890/12-2053.1 URL [本文引用: 1] [6] Chave J Condit R Lao S Caspersen JP Foster RB Hubbell SP 2003 ). Spatial and temporal variation of biomass in a tropical forest: Results from a large census plot in PanamaJournal of Ecology , 91, 240 -252 . DOI:
10.1046/j.1365-2745.2003.00757.x URL [本文引用: 1] [7] Chen CG Zhu JF 1989 ). A Handbook for Main Tree Species Biomass in Northeast China . China Forestry Publishing House, Beijing. [本文引用: 1] [ 陈传国 , 朱俊凤 (1989 ). 东北主要林木生物量手册 . 中国林业出版社, 北京.] [本文引用: 1] [8] Chen DM Cheng JH Chu PF Mi J Hu SJ Xie YC Tuvshintogtokh I Bai YF 2016 ). Effect of diversity on biomass across grasslands on the Mongolian Plateau: Contrasting effects between plants and soil nematodesJournal of Biogeography , 43, 955 -966 . DOI:
10.1111/jbi.2016.43.issue-5 URL [本文引用: 1] [9] Cornelissen JHC Lavorel S Garnier E Diaz S Buchmann N Gurvich DE Reich PB ter Steege H Morgan HD van der Heijden MGA Pausas JG Poorter H 2003 ). A handbook of protocols for standardised and easy measurement of plant functional traits worldwideAustralian Journal of Botany , 51, 335 -380 . DOI:
10.1071/BT02124 URL [本文引用: 1] [10] Díaz S Lavorel S Bello FD Quétier F Grigulis K Robson TM 2007 ). Incorporating plant functional diversity effects in ecosystem service assessmentsProceedings of the National Academy of Sciences of the United States of America , 104, 20684 -20689 . DOI:
10.1073/pnas.0704716104 URL [本文引用: 2] [11] D?nescu A Albrecht AT Bauhus J 2016 ). Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern GermanyOecologia , 182, 319 -333 . DOI:
10.1007/s00442-016-3623-4 URL [本文引用: 1] [12] Edwards JR Lambert LS 2007 ). Methods for integrating moderation and mediation: A general analytical framework using moderated path analysisPsychological Methods , 12, 1 -22 . DOI:
10.1037/1082-989X.12.1.1 URL [本文引用: 1] [13] Finegan B Pe?a-Claros M de Oliveira A Ascarrunz N Bret-Harte MS Carre?o-Rocabado G Casanoves F Díaz S Eguiguren Velepucha P Fernandez F Licona JC Lorenzo L Salgado Negret B Vaz M Poorter L Canham C 2015 ). Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypothesesJournal of Ecology , 103, 191 -201 . DOI:
10.1111/1365-2745.12346 URL [本文引用: 2] [14] Fox J 2008 ). Applied Regression Analysis and Generalized Linear Models . 2nd ednSage Publications, Thousand Oaks, USA. [本文引用: 1] [15] Fox J Monette G 1992 ). Generalized collinearity diagnosticsJournal of the American Statistical Association , 87, 178 -183 . DOI:
10.1080/01621459.1992.10475190 URL [本文引用: 1] [16] Gazol A Camarero JJ 2016 ). Functional diversity enhances silver fir growth resilience to an extreme droughtJournal of Ecology , 104, 1063 -1075 . DOI:
10.1111/1365-2745.12575 URL [本文引用: 2] [17] Grace JB Anderson TM Seabloom EW Borer ET Adler PB Harpole WS Hautier Y Hillebrand H Lind EM Partel M Bakker JD Buckley YM Crawley MJ Damschen EI Davies KF Fay PA Firn J Gruner DS Hector A Knops JM MacDougall AS Melbourne BA Morgan JW Orrock JL Prober SM Smith MD 2016 ). Integrative modelling reveals mechanisms linking productivity and plant species richnessNature , 529, 390 -393 . DOI:
10.1038/nature16524 [本文引用: 1] [18] Grime JP 1998 ). Benefits of plant diversity to ecosystems: Immediate, filter and founder effectsJournal of Ecology , 86, 902 -910 . DOI:
10.1046/j.1365-2745.1998.00306.x URL [本文引用: 1] [19] Howard AL 2013 ). Handbook of structural equation modelingStructural Equation Modeling-a Multidisciplinary Journal , 20, 354 -360 . DOI:
10.1080/10705511.2013.769397 URL [本文引用: 1] [20] Huang XR 2018 ). Relationship between plant functional diversity and productivity ofPinus massoniana plantations in Guangxi. Biodiversity Science, 690 -700 . [本文引用: 3] [ 黄小荣 ( 2018 ). 广西马尾松林植物功能多样性与生产力的关系生物多样性 , 26, 690 -700 .] [本文引用: 3] [21] Laliberté E Legendre P 2010 ). A distance-based framework for measuring functional diversity from multiple traitsEcology , 91, 299 -305 . DOI:
10.1890/08-2244.1 URL [本文引用: 2] [22] Lin DM Lai JS Muller-Landau HC Mi XC Ma KP 2012 ). Topographic variation in aboveground biomass in a subtropical evergreen broad-leaved forest in ChinaPLOS ONE 10.1371/journal.pone.0048244 . [本文引用: 1] [23] Lohbeck M Poorter L Martinez-Ramos M Bongers F 2015 ). Biomass is the main driver of changes in ecosystem process rates during tropical forest successionEcology , 96, 1242 -1252 . DOI:
10.1890/14-0472.1 URL [本文引用: 4] [24] Lü TT Wang P Yan H Zhang W Liao GX Jiang HB Zou CL Sheng LX 2014 ). Relationship between functional diversity and productivity in meadow and marsh plant communitiesChinese Journal of Plant Ecology , 38, 405 -416 . [本文引用: 2] [ 吕亭亭 , 王平 , 燕红 , 张稳 , 廖桂项 , 姜海波 , 邹畅林 , 盛连喜 ( 2014 ). 草甸和沼泽植物群落功能多样性与生产力的关系植物生态学报 , 38, 405 -416 .] [本文引用: 2] [25] McGill BJ Enquist BJ Weiher E Westoby M 2006 ). Rebuilding community ecology from functional traitsTrends in Ecology and Evolution , 21, 178 -185 . DOI:
10.1016/j.tree.2006.02.002 URL [本文引用: 2] [26] Mensah S Veldtman R Assogbadjo AE Glèlè KR Seifert T 2016 ). Tree species diversity promotes aboveground carbon storage through functional diversity and functional dominanceEcology and Evolution , 6, 7546 -7557 . DOI:
10.1002/ece3.2525 URL [本文引用: 1] [27] Micheli F Halpern BS 2010 ). Low functional redundancy in coastal marine assemblagesEcology Letters , 8, 391 -400 . [本文引用: 1] [28] Mokany K Ash J Roxburgh S 2008 ). Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grasslandJournal of Ecology , 96, 884 -893 . DOI:
10.1111/j.1365-2745.2008.01395.x URL [本文引用: 3] [29] Mouchet MA Villeger S Mason NWH Mouillot D 2010 ). Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rulesFunctional Ecology , 24, 867 -876 . DOI:
10.1111/fec.2010.24.issue-4 URL [本文引用: 1] [30] Oksanen J Kindt R Legendre P O’Hara B Simpson GL Slymos P Stevens MHH Wagner H 2009 ). The vegan Package. . Cited: 2019-02-04. URL [本文引用: 3] [31] Ouyang S Xiang W Wang X Zeng Y Lei P Deng X Peng C 2016 ). Significant effects of biodiversity on forest biomass during the succession of subtropical forest in south ChinaForest Ecology and Management , 372, 291 -302 . DOI:
10.1016/j.foreco.2016.04.020 URL [本文引用: 2] [32] Petchey OL Hector A Gaston KJ 2004 ). How do different measures of functional diversity perform?Ecology , 85, 847 -857 . DOI:
10.1890/03-0226 URL [本文引用: 2] [33] Ratcliffe S Liebergesell M Ruiz-Benito P Madrigal González J Mu?oz Casta?eda JM K?ndler G Lehtonen A Dahlgren J Kattge J Pe?uelas J Zavala MA Wirth C 2016 ). Modes of functional biodiversity control on tree productivity across the European continentGlobal Ecology and Biogeography , 25, 251 -262 . DOI:
10.1111/geb.12406 URL [本文引用: 2] [34] Rosseel Y 2012 ). lavaan: An R package for structural equation modelingJournal of Statistical Software , 48, 1 -36 . [本文引用: 3] [35] Ruiz-Benito P Gomez-Aparicio L Paquette A Messier C Kattge J Zavala MA 2014 ). Diversity increases carbon storage and tree productivity in Spanish forestsGlobal Ecology and Biogeography , 23, 311 -322 . DOI:
10.1111/geb.12126 URL [本文引用: 1] [36] Schellberg J Pontes LDS 2012 ). Plant functional traits and nutrient gradients on grasslandGrass and Forage Science , 67, 305 -319 . DOI:
10.1111/gfs.2012.67.issue-3 URL [37] Sun RY Li QF Niu CJ Lou AR 2002 ). Basic Ecology . Higher Education Press, Beijing. [本文引用: 2] [ 孙儒泳 , 李庆芬 , 牛翠娟 , 娄安如 (2002 ). 基础生态学 . 高等教育出版社, 北京.] [本文引用: 2] [38] Tilman D 1997 ). Distinguishing between the effects of species diversity and species compositionOikos , 80, 185 . DOI:
10.2307/3546532 URL [本文引用: 2] [39] Tobner CM Paquette A Gravel D Reich PB Williams LJ Messier C 2016 ). Functional identity is the main driver of diversity effects in young tree communitiesEcology Letters , 19, 638 -647 . DOI:
10.1111/ele.12600 URL [本文引用: 2] [40] Van MT Pe?a-Claros M Ascarrunz N Arets EJMM Licona JC Toledo M Poorter L 2017 ). Abiotic and biotic drivers of biomass change in a neotropical forestJournal of Ecology , 105, 1223 -1234 . DOI:
10.1111/1365-2745.12756 URL [41] Wu Z Raven P 1994-2009 ). Flora of China . Science Press and Missouri Botanical Garden Press, Beijing and St Louis. [本文引用: 2] [42] Zhang Y Chen HYH 2015 ). Individual size inequality links forest diversity and above-ground biomassJournal of Ecology , 103, 1245 -1252 . DOI:
10.1111/1365-2745.12425 URL [本文引用: 1] [43] Zhang Y Chen HYH Reich PB 2012 ). Forest productivity increases with evenness, species richness and trait variation: A global meta-analysisJournal of Ecology , 100, 742 -749 . DOI:
10.1111/j.1365-2745.2011.01944.x URL Community-weighted mean of leaf traits and divergence of wood traits predict aboveground biomass in secondary subtropical forests
2
2017
... 植物多样性影响生态系统功能的机理存在两种假说.一是“生态位互补假说” (
Tilman, 1997 ), 认为随着多样性的增加, 群落内树种的功能特性差异增大, 如叶面积、冠层高度、生长速度等, 能够缓解竞争.通常用多样性指数(如物种、功能多样性)来表征该假说.二是“生物量比率假说” (
Grime, 1998 ), 认为某个时间点的生态系统功能主要由植物优势物种的功能性状值决定.当该假说占主导时, 可以检测到
CWM 是解释生态系统功能的最重要因子(
Tobner et al ., 2016 ;
Ali et al ., 2017 ).尽管目前研究普遍认为生态位互补与生物量比率假说并不相互排斥, 但二者的相对重要性还存在很大争论.
Cardinale和Gonzalez (2011) 整合分析多样性种植实验结果, 发现两种假说对生产力变异的解释基本持平;
Ali等(2017) 则发现生物量比率假说在解释中国东部亚热带次生林的生产力变异中占主导地位;
Ratcliffe等(2016) 发现在欧洲温带森林中生态位互补和生物量比率效应对树木生长均有促进作用, 但在水分受限制的地中海森林中生态位互补假说能够更好地解释生产力的变异. ...
... 整合分析多样性种植实验结果, 发现两种假说对生产力变异的解释基本持平;
Ali等(2017) 则发现生物量比率假说在解释中国东部亚热带次生林的生产力变异中占主导地位;
Ratcliffe等(2016) 发现在欧洲温带森林中生态位互补和生物量比率效应对树木生长均有促进作用, 但在水分受限制的地中海森林中生态位互补假说能够更好地解释生产力的变异. ...
Package ‘MuMIn’
1
2018
... 采用MuMIn包中的dredge函数(
Bartoń, 2018 ), 分别将生产力与多样性指数的模型、生产力与群落加权平均值的模型、生产力与环境变量的模型的各个变量之间所有可能的组合进行拟合, 计算出每个回归模型的变量组合中各单一子模型在模型中对响应变量解释的重要性.在拟合的模型中筛选出AIC差值小于2的所有模型, 选择变量重要性最高、与生产力最为相关的多样性指数与环境因子, 用来构建结构方程模型. ...
Biodiversity loss and its impact on humanity
1
2012
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
The functional role of producer diversity in ecosystems
1
2011
... 植物多样性影响生态系统功能的机理存在两种假说.一是“生态位互补假说” (
Tilman, 1997 ), 认为随着多样性的增加, 群落内树种的功能特性差异增大, 如叶面积、冠层高度、生长速度等, 能够缓解竞争.通常用多样性指数(如物种、功能多样性)来表征该假说.二是“生物量比率假说” (
Grime, 1998 ), 认为某个时间点的生态系统功能主要由植物优势物种的功能性状值决定.当该假说占主导时, 可以检测到
CWM 是解释生态系统功能的最重要因子(
Tobner et al ., 2016 ;
Ali et al ., 2017 ).尽管目前研究普遍认为生态位互补与生物量比率假说并不相互排斥, 但二者的相对重要性还存在很大争论.
Cardinale和Gonzalez (2011) 整合分析多样性种植实验结果, 发现两种假说对生产力变异的解释基本持平;
Ali等(2017) 则发现生物量比率假说在解释中国东部亚热带次生林的生产力变异中占主导地位;
Ratcliffe等(2016) 发现在欧洲温带森林中生态位互补和生物量比率效应对树木生长均有促进作用, 但在水分受限制的地中海森林中生态位互补假说能够更好地解释生产力的变异. ...
Better environmental data may reverse conclusions about niche- and dispersal-based processes in community assembly
1
2013
... 地形变量包括凹凸度、海拔和坡度, 将9 hm
2 阔叶红松林动态监测样地划分为225个20 m × 20 m小样方, 样方所有变量均根据每个小样方单元的四个角的高度计算(
Chang et al ., 2013 ).在每个20 m × 20 m样方原点位置, 取1个样点, 同时随机选择一个方向, 在离原点2、5、8 m处选任意2个样点, 取土壤表层(0-10 cm)的样品, 于实验室内测定速效氮、速效磷、速效钾、全氮、全磷、有机碳含量、pH值、土壤容重、体积含水率和质量含水率.我们将每个20 m × 20 m的样方分为了16个5 m × 5 m的子样方后, 应用ArcGIS 10.1通过地统计学分析工具中的ordinary Kriging方法对地形和土壤因子共13项指标进行空间插值, 从而获得每个10 m × 10 m取样单元的生境因子指标(
表2 ). ...
Spatial and temporal variation of biomass in a tropical forest: Results from a large census plot in Panama
1
2003
... 根据东北主要林木生物量的异速生长方程(
陈传国和朱俊凤, 1989 ), 用实测胸径计算木本植株的地上生物量.群落木本植物生产力(
CWP, Mg·hm
-2 ·a
-1 )的计算参考了
Chave等(2003) 的方法:
CWP = (
G + R )/0.01 ×
T, 式中
G 为10 m × 10 m样方内两次调查都存活个体地上生物量的增长量;
R 为10 m × 10 m样方内第一次调查胸径小于2 cm, 复查时胸径超过2 cm个体的地上生物量总和,
T 为两次调查时间的间隔(10年). ...
1
1989
... 根据东北主要林木生物量的异速生长方程(
陈传国和朱俊凤, 1989 ), 用实测胸径计算木本植株的地上生物量.群落木本植物生产力(
CWP, Mg·hm
-2 ·a
-1 )的计算参考了
Chave等(2003) 的方法:
CWP = (
G + R )/0.01 ×
T, 式中
G 为10 m × 10 m样方内两次调查都存活个体地上生物量的增长量;
R 为10 m × 10 m样方内第一次调查胸径小于2 cm, 复查时胸径超过2 cm个体的地上生物量总和,
T 为两次调查时间的间隔(10年). ...
1
1989
... 根据东北主要林木生物量的异速生长方程(
陈传国和朱俊凤, 1989 ), 用实测胸径计算木本植株的地上生物量.群落木本植物生产力(
CWP, Mg·hm
-2 ·a
-1 )的计算参考了
Chave等(2003) 的方法:
CWP = (
G + R )/0.01 ×
T, 式中
G 为10 m × 10 m样方内两次调查都存活个体地上生物量的增长量;
R 为10 m × 10 m样方内第一次调查胸径小于2 cm, 复查时胸径超过2 cm个体的地上生物量总和,
T 为两次调查时间的间隔(10年). ...
Effect of diversity on biomass across grasslands on the Mongolian Plateau: Contrasting effects between plants and soil nematodes
1
2016
... 多样性并非生态系统生产力的唯一驱动因素, 它同时还受非生物因素和其他生物因素的影响, 如初始生物量、土壤养分等环境因子(
Lin et al ., 2012 ;
Chen et al ., 2016 ).有研究认为, 林分的初始地上生物量是森林生产力的关键驱动因素, 是植被数量(即初始生物量)而不是植被质量(即多样性)对生产力变异起着主要作用(
Lohbeck et al ., 2015 ).研究发现单纯考虑多样性与生产力二元关系时, 物种丰富度显著影响生产力, 而整合考虑土壤肥力等非生物因子的作用后, 二者关系变得不明显(
Grace et al ., 2016 ); 与此相反, 当去除其他非生物(海拔、冠层光照)和生物因子(树干密度、森林演替阶段)后, 我国亚热带森林中多样性-生产力正相关关系变弱甚至不相关(
Ouyang et al ., 2016 ); 有研究表明, 土壤因素推动了生产力的变异, 而生态位互补和生物量比率机制在天然林中的重要性却微不足道(
Van et al ., 2017 ).综上来看, 生物因素(植物多样性、初始生物量等)与非生物因素(土壤、地形等)的相对重要性仍需进一步验证. ...
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide
1
2003
... 本研究测定与植物生长和群落生产力密切相关的8种功能性状: 木材密度、最大树高、比叶面积、叶面积、叶片厚度、叶片干物质含量、叶片碳含量和叶片氮含量(
Finegan et al ., 2015 ;
Van et al ., 2017 ) (
表1 ).对41种木本植物的功能性状进行了测定, 这些物种的生物量总和占群落总生物量的85%以上(
吕亭亭等, 2014 ).每个物种随机选取5株生长良好、无病虫害的植株, 每株剪下20片健康无损、被日光照射无遮阴的叶片, 依据植物性状标准化测定方法(
Cornelissen et al ., 2003 ), 测定6种叶片性状; 在样地周围对每个物种随机选择3株健康植株, 在距离地面1.3 m处钻取一个生长芯, 把生长芯分成1 cm的片段, 用水置换法测定各节段鲜体积, 103 ℃烘干72 h, 用烘干的质量除以鲜体积来测量木材密度; 最大树高来源于《中国植物志》(
Wu & Raven, 1994- 2009 ). ...
Incorporating plant functional diversity effects in ecosystem service assessments
2
2007
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
... 群落加权平均值用于评估优势性状对生产力的影响, 而功能多样性指数则是量化该性状的变异性对生产力的影响(
Tobner et al ., 2016 ).在草本植物生态系统中, 群落加权平均值能更好地解释生产力的变异(
Mokany et al ., 2008 ), 而在欧洲地中海森林(
Ratcliffe et al ., 2016 )和广西马尾松(
Pinus massoniana )林(
黄小荣, 2018 )则发现功能多样性指数更好地解释了生产力的变异, 本研究结果与之一致.这种不同群落类型的多样性-生产力关系结果的不一致, 可能是方法上的差异, 包括选择不同的样地监测周期和多样性指数(
Dănescu et al ., 2016 ); 也可能是由不同生态系统类型和演替阶段造成的.当森林的木本植物存在较大的功能性状差异时, 各物种具有不同获取和利用资源的策略, 减少了生态位重叠, 生态位互补效应会减弱物种间对资源的竞争, 最大化地利用光等有限资源, 从而提高生态系统生产力(
Díaz et al ., 2007 ).而在有生境过滤和扩散限制的群落中, 优势物种的影响可能更大, 生态系统过程主要取决于群落中优势物种的功能性状, 此时生物量比率效应更为重要(
Finegan et al ., 2015 ). ...
Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany
1
2016
... 群落加权平均值用于评估优势性状对生产力的影响, 而功能多样性指数则是量化该性状的变异性对生产力的影响(
Tobner et al ., 2016 ).在草本植物生态系统中, 群落加权平均值能更好地解释生产力的变异(
Mokany et al ., 2008 ), 而在欧洲地中海森林(
Ratcliffe et al ., 2016 )和广西马尾松(
Pinus massoniana )林(
黄小荣, 2018 )则发现功能多样性指数更好地解释了生产力的变异, 本研究结果与之一致.这种不同群落类型的多样性-生产力关系结果的不一致, 可能是方法上的差异, 包括选择不同的样地监测周期和多样性指数(
Dănescu et al ., 2016 ); 也可能是由不同生态系统类型和演替阶段造成的.当森林的木本植物存在较大的功能性状差异时, 各物种具有不同获取和利用资源的策略, 减少了生态位重叠, 生态位互补效应会减弱物种间对资源的竞争, 最大化地利用光等有限资源, 从而提高生态系统生产力(
Díaz et al ., 2007 ).而在有生境过滤和扩散限制的群落中, 优势物种的影响可能更大, 生态系统过程主要取决于群落中优势物种的功能性状, 此时生物量比率效应更为重要(
Finegan et al ., 2015 ). ...
Methods for integrating moderation and mediation: A general analytical framework using moderated path analysis
1
2007
... 根据变量筛选结果, 选择变量重要性最高、与生产力最为相关的多样性指数与潜在环境变量, 纳入到结构方程模型中.采用最大似然法对结构方程模型进行拟合, 利用比较拟合指数(
CFI )、标准均方根残差(
SRMR )、渐进残差均方和平方根(
RMSEA )、显著性概率值(
p )进行模型优度的评价(
Howard, 2013 ).拟合优度的临界值为
CFI > 0.9,
SRMR < 0.05,
RMSEA < 0.08,
p > 0.05.利用模型中各路径标准化系数, 定量表示不同因素对生产力的相对作用大小, 量化各个变量对森林地上生产力的直接、间接和总标准化效应(
Edwards & Lambert, 2007 ), 从而比较生态位互补假说与生物量比率假说对阔叶红松林的适用性.上述分析利用lavaan包完成(
Rosseel, 2012 ). ...
Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses
2
2015
... 本研究测定与植物生长和群落生产力密切相关的8种功能性状: 木材密度、最大树高、比叶面积、叶面积、叶片厚度、叶片干物质含量、叶片碳含量和叶片氮含量(
Finegan et al ., 2015 ;
Van et al ., 2017 ) (
表1 ).对41种木本植物的功能性状进行了测定, 这些物种的生物量总和占群落总生物量的85%以上(
吕亭亭等, 2014 ).每个物种随机选取5株生长良好、无病虫害的植株, 每株剪下20片健康无损、被日光照射无遮阴的叶片, 依据植物性状标准化测定方法(
Cornelissen et al ., 2003 ), 测定6种叶片性状; 在样地周围对每个物种随机选择3株健康植株, 在距离地面1.3 m处钻取一个生长芯, 把生长芯分成1 cm的片段, 用水置换法测定各节段鲜体积, 103 ℃烘干72 h, 用烘干的质量除以鲜体积来测量木材密度; 最大树高来源于《中国植物志》(
Wu & Raven, 1994- 2009 ). ...
... 群落加权平均值用于评估优势性状对生产力的影响, 而功能多样性指数则是量化该性状的变异性对生产力的影响(
Tobner et al ., 2016 ).在草本植物生态系统中, 群落加权平均值能更好地解释生产力的变异(
Mokany et al ., 2008 ), 而在欧洲地中海森林(
Ratcliffe et al ., 2016 )和广西马尾松(
Pinus massoniana )林(
黄小荣, 2018 )则发现功能多样性指数更好地解释了生产力的变异, 本研究结果与之一致.这种不同群落类型的多样性-生产力关系结果的不一致, 可能是方法上的差异, 包括选择不同的样地监测周期和多样性指数(
Dănescu et al ., 2016 ); 也可能是由不同生态系统类型和演替阶段造成的.当森林的木本植物存在较大的功能性状差异时, 各物种具有不同获取和利用资源的策略, 减少了生态位重叠, 生态位互补效应会减弱物种间对资源的竞争, 最大化地利用光等有限资源, 从而提高生态系统生产力(
Díaz et al ., 2007 ).而在有生境过滤和扩散限制的群落中, 优势物种的影响可能更大, 生态系统过程主要取决于群落中优势物种的功能性状, 此时生物量比率效应更为重要(
Finegan et al ., 2015 ). ...
Applied Regression Analysis and Generalized Linear Models . 2nd edn
1
2008
... 多重共线性会影响解释变量对响应变量的解释和预测能力.利用方差膨胀因子(
VIF )检验变量间的多重共线性, 根据以往经验,
VIF < 10时认为变量间没有多重共线性(
Fox & Monette, 1992 ;
Fox, 2008 ).验证结果显示, 解释变量均匀度指数(
J )、叶片厚度的群落加权平均值(
CWM LT )和叶片厚度的功能分散指数(
FD LT )存在多重共线性, 故删去上述变量. ...
Generalized collinearity diagnostics
1
1992
... 多重共线性会影响解释变量对响应变量的解释和预测能力.利用方差膨胀因子(
VIF )检验变量间的多重共线性, 根据以往经验,
VIF < 10时认为变量间没有多重共线性(
Fox & Monette, 1992 ;
Fox, 2008 ).验证结果显示, 解释变量均匀度指数(
J )、叶片厚度的群落加权平均值(
CWM LT )和叶片厚度的功能分散指数(
FD LT )存在多重共线性, 故删去上述变量. ...
Functional diversity enhances silver fir growth resilience to an extreme drought
2
2016
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
...
Petchey等(2004) 首次证明功能多样性比物种多样性能更有效地预测生物量的变化,
Gazol和Camarero (2016) 指出功能多样性与生态系统生产力的关系非常密切, 功能多样性是解释生态系统功能变化的更有效的指标.相对于物种多样性, 功能多样性有两个优点: 首先, 各物种具有相异的功能特征, 物种多样性无法解释物种间功能特性的差异, 而功能多样性能够综合不同物种的属性(
Micheli & Halpern, 2010 ); 其次, 功能性状可以表征生物从环境中利用资源的能力(
McGill et al ., 2006 ).功能性状的多样性比物种多样性更好地反映不同物种的生态位, 因此能够更有效地揭示生产力的变异规律(
Petchey et al ., 2004 ). ...
Integrative modelling reveals mechanisms linking productivity and plant species richness
1
2016
... 多样性并非生态系统生产力的唯一驱动因素, 它同时还受非生物因素和其他生物因素的影响, 如初始生物量、土壤养分等环境因子(
Lin et al ., 2012 ;
Chen et al ., 2016 ).有研究认为, 林分的初始地上生物量是森林生产力的关键驱动因素, 是植被数量(即初始生物量)而不是植被质量(即多样性)对生产力变异起着主要作用(
Lohbeck et al ., 2015 ).研究发现单纯考虑多样性与生产力二元关系时, 物种丰富度显著影响生产力, 而整合考虑土壤肥力等非生物因子的作用后, 二者关系变得不明显(
Grace et al ., 2016 ); 与此相反, 当去除其他非生物(海拔、冠层光照)和生物因子(树干密度、森林演替阶段)后, 我国亚热带森林中多样性-生产力正相关关系变弱甚至不相关(
Ouyang et al ., 2016 ); 有研究表明, 土壤因素推动了生产力的变异, 而生态位互补和生物量比率机制在天然林中的重要性却微不足道(
Van et al ., 2017 ).综上来看, 生物因素(植物多样性、初始生物量等)与非生物因素(土壤、地形等)的相对重要性仍需进一步验证. ...
Benefits of plant diversity to ecosystems: Immediate, filter and founder effects
1
1998
... 植物多样性影响生态系统功能的机理存在两种假说.一是“生态位互补假说” (
Tilman, 1997 ), 认为随着多样性的增加, 群落内树种的功能特性差异增大, 如叶面积、冠层高度、生长速度等, 能够缓解竞争.通常用多样性指数(如物种、功能多样性)来表征该假说.二是“生物量比率假说” (
Grime, 1998 ), 认为某个时间点的生态系统功能主要由植物优势物种的功能性状值决定.当该假说占主导时, 可以检测到
CWM 是解释生态系统功能的最重要因子(
Tobner et al ., 2016 ;
Ali et al ., 2017 ).尽管目前研究普遍认为生态位互补与生物量比率假说并不相互排斥, 但二者的相对重要性还存在很大争论.
Cardinale和Gonzalez (2011) 整合分析多样性种植实验结果, 发现两种假说对生产力变异的解释基本持平;
Ali等(2017) 则发现生物量比率假说在解释中国东部亚热带次生林的生产力变异中占主导地位;
Ratcliffe等(2016) 发现在欧洲温带森林中生态位互补和生物量比率效应对树木生长均有促进作用, 但在水分受限制的地中海森林中生态位互补假说能够更好地解释生产力的变异. ...
Handbook of structural equation modeling
1
2013
... 根据变量筛选结果, 选择变量重要性最高、与生产力最为相关的多样性指数与潜在环境变量, 纳入到结构方程模型中.采用最大似然法对结构方程模型进行拟合, 利用比较拟合指数(
CFI )、标准均方根残差(
SRMR )、渐进残差均方和平方根(
RMSEA )、显著性概率值(
p )进行模型优度的评价(
Howard, 2013 ).拟合优度的临界值为
CFI > 0.9,
SRMR < 0.05,
RMSEA < 0.08,
p > 0.05.利用模型中各路径标准化系数, 定量表示不同因素对生产力的相对作用大小, 量化各个变量对森林地上生产力的直接、间接和总标准化效应(
Edwards & Lambert, 2007 ), 从而比较生态位互补假说与生物量比率假说对阔叶红松林的适用性.上述分析利用lavaan包完成(
Rosseel, 2012 ). ...
广西马尾松林植物功能多样性与生产力的关系
3
2018
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
... 基于所测定的8个植物功能性状, 计算每个小样方的功能均匀度(
FEve )、Rao二次熵指数(
FDQ )、功能分散指数(
FDis ), 计算基于8个功能性状组合的功能分散指数(
FD com )和分别基于单个功能性状的功能分散指数(
FD WD ,
FD MH ,
FD SLA ,
FD LA ,
FD LT ,
FD LDMC ,
FD LCC ), 同时计算了8个性状的群落加权平均值(
CWM ).各指数计算公式(
黄小荣, 2018 )如下: ...
... 群落加权平均值用于评估优势性状对生产力的影响, 而功能多样性指数则是量化该性状的变异性对生产力的影响(
Tobner et al ., 2016 ).在草本植物生态系统中, 群落加权平均值能更好地解释生产力的变异(
Mokany et al ., 2008 ), 而在欧洲地中海森林(
Ratcliffe et al ., 2016 )和广西马尾松(
Pinus massoniana )林(
黄小荣, 2018 )则发现功能多样性指数更好地解释了生产力的变异, 本研究结果与之一致.这种不同群落类型的多样性-生产力关系结果的不一致, 可能是方法上的差异, 包括选择不同的样地监测周期和多样性指数(
Dănescu et al ., 2016 ); 也可能是由不同生态系统类型和演替阶段造成的.当森林的木本植物存在较大的功能性状差异时, 各物种具有不同获取和利用资源的策略, 减少了生态位重叠, 生态位互补效应会减弱物种间对资源的竞争, 最大化地利用光等有限资源, 从而提高生态系统生产力(
Díaz et al ., 2007 ).而在有生境过滤和扩散限制的群落中, 优势物种的影响可能更大, 生态系统过程主要取决于群落中优势物种的功能性状, 此时生物量比率效应更为重要(
Finegan et al ., 2015 ). ...
广西马尾松林植物功能多样性与生产力的关系
3
2018
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
... 基于所测定的8个植物功能性状, 计算每个小样方的功能均匀度(
FEve )、Rao二次熵指数(
FDQ )、功能分散指数(
FDis ), 计算基于8个功能性状组合的功能分散指数(
FD com )和分别基于单个功能性状的功能分散指数(
FD WD ,
FD MH ,
FD SLA ,
FD LA ,
FD LT ,
FD LDMC ,
FD LCC ), 同时计算了8个性状的群落加权平均值(
CWM ).各指数计算公式(
黄小荣, 2018 )如下: ...
... 群落加权平均值用于评估优势性状对生产力的影响, 而功能多样性指数则是量化该性状的变异性对生产力的影响(
Tobner et al ., 2016 ).在草本植物生态系统中, 群落加权平均值能更好地解释生产力的变异(
Mokany et al ., 2008 ), 而在欧洲地中海森林(
Ratcliffe et al ., 2016 )和广西马尾松(
Pinus massoniana )林(
黄小荣, 2018 )则发现功能多样性指数更好地解释了生产力的变异, 本研究结果与之一致.这种不同群落类型的多样性-生产力关系结果的不一致, 可能是方法上的差异, 包括选择不同的样地监测周期和多样性指数(
Dănescu et al ., 2016 ); 也可能是由不同生态系统类型和演替阶段造成的.当森林的木本植物存在较大的功能性状差异时, 各物种具有不同获取和利用资源的策略, 减少了生态位重叠, 生态位互补效应会减弱物种间对资源的竞争, 最大化地利用光等有限资源, 从而提高生态系统生产力(
Díaz et al ., 2007 ).而在有生境过滤和扩散限制的群落中, 优势物种的影响可能更大, 生态系统过程主要取决于群落中优势物种的功能性状, 此时生物量比率效应更为重要(
Finegan et al ., 2015 ). ...
A distance-based framework for measuring functional diversity from multiple traits
2
2010
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
... 采用R 3.5.1 vegan包的diversity函数(
Oksanen et al ., 2009 ), 计算每个样方的物种多样性指标; FD包计算功能多样性指数与群落加权平均值(
Laliberté & Legendre, 2010 )(
表3 ). ...
Topographic variation in aboveground biomass in a subtropical evergreen broad-leaved forest in China
1
2012
... 多样性并非生态系统生产力的唯一驱动因素, 它同时还受非生物因素和其他生物因素的影响, 如初始生物量、土壤养分等环境因子(
Lin et al ., 2012 ;
Chen et al ., 2016 ).有研究认为, 林分的初始地上生物量是森林生产力的关键驱动因素, 是植被数量(即初始生物量)而不是植被质量(即多样性)对生产力变异起着主要作用(
Lohbeck et al ., 2015 ).研究发现单纯考虑多样性与生产力二元关系时, 物种丰富度显著影响生产力, 而整合考虑土壤肥力等非生物因子的作用后, 二者关系变得不明显(
Grace et al ., 2016 ); 与此相反, 当去除其他非生物(海拔、冠层光照)和生物因子(树干密度、森林演替阶段)后, 我国亚热带森林中多样性-生产力正相关关系变弱甚至不相关(
Ouyang et al ., 2016 ); 有研究表明, 土壤因素推动了生产力的变异, 而生态位互补和生物量比率机制在天然林中的重要性却微不足道(
Van et al ., 2017 ).综上来看, 生物因素(植物多样性、初始生物量等)与非生物因素(土壤、地形等)的相对重要性仍需进一步验证. ...
Biomass is the main driver of changes in ecosystem process rates during tropical forest succession
4
2015
... 多样性并非生态系统生产力的唯一驱动因素, 它同时还受非生物因素和其他生物因素的影响, 如初始生物量、土壤养分等环境因子(
Lin et al ., 2012 ;
Chen et al ., 2016 ).有研究认为, 林分的初始地上生物量是森林生产力的关键驱动因素, 是植被数量(即初始生物量)而不是植被质量(即多样性)对生产力变异起着主要作用(
Lohbeck et al ., 2015 ).研究发现单纯考虑多样性与生产力二元关系时, 物种丰富度显著影响生产力, 而整合考虑土壤肥力等非生物因子的作用后, 二者关系变得不明显(
Grace et al ., 2016 ); 与此相反, 当去除其他非生物(海拔、冠层光照)和生物因子(树干密度、森林演替阶段)后, 我国亚热带森林中多样性-生产力正相关关系变弱甚至不相关(
Ouyang et al ., 2016 ); 有研究表明, 土壤因素推动了生产力的变异, 而生态位互补和生物量比率机制在天然林中的重要性却微不足道(
Van et al ., 2017 ).综上来看, 生物因素(植物多样性、初始生物量等)与非生物因素(土壤、地形等)的相对重要性仍需进一步验证. ...
... 生物多样性与生产力之间的关系直接和间接地受多个因素的作用(
Lohbeck et al ., 2015 ).本研究运用结构方程模型评价了功能多样性、地上初始生物量、土壤养分和地形条件对生产力的相对贡献.结果显示, 地上初始生物量对生产力作用的总效应为0.449 (
表6 ), 在各因子中与生产力的相关路径系数最大, 表明地上初始生物量是森林生产力最重要的内在驱动因素, 即植被数量(即林分生物量)而非植被质量(即功能多样性和功能性状组成)是决定森林生产力的主要因素.地形因子对土壤因子的作用效应为-0.230, 并通过多样性、初始生物量对生产力产生间接影响, 其对生产力的总效应为-0.090, 这可能是由于海拔较高生境土壤水分和养分条件较差, 导致森林生产力下降. ...
... 在相对资源受限的环境中, 各物种采取不同的资源吸收策略才能共存, 此时互补效应为生产力的主要驱动力.在生物量较高的地区, 丰富的资源导致互补效应的相对重要性降低, 多样性对生产力的影响也将减弱(
Lohbeck et al ., 2015 ).当群落中出现非生物过滤(如环境过滤)时, 只有适应相应环境条件的物种才能够生存, 导致物种间的性状差异减少, 形成性状聚集(
Schellberg & Pontes, 2012 ), 致使局域范围内群落的优势物种表现出强烈的趋同现象.因此, 生物多样性并非生态系统生产力的唯一驱动因素, 而是受非生物因素和其他生物因素的共同影响(
Lohbeck et al ., 2015 ). ...
... ), 致使局域范围内群落的优势物种表现出强烈的趋同现象.因此, 生物多样性并非生态系统生产力的唯一驱动因素, 而是受非生物因素和其他生物因素的共同影响(
Lohbeck et al ., 2015 ). ...
草甸和沼泽植物群落功能多样性与生产力的关系
2
2014
... 本研究测定与植物生长和群落生产力密切相关的8种功能性状: 木材密度、最大树高、比叶面积、叶面积、叶片厚度、叶片干物质含量、叶片碳含量和叶片氮含量(
Finegan et al ., 2015 ;
Van et al ., 2017 ) (
表1 ).对41种木本植物的功能性状进行了测定, 这些物种的生物量总和占群落总生物量的85%以上(
吕亭亭等, 2014 ).每个物种随机选取5株生长良好、无病虫害的植株, 每株剪下20片健康无损、被日光照射无遮阴的叶片, 依据植物性状标准化测定方法(
Cornelissen et al ., 2003 ), 测定6种叶片性状; 在样地周围对每个物种随机选择3株健康植株, 在距离地面1.3 m处钻取一个生长芯, 把生长芯分成1 cm的片段, 用水置换法测定各节段鲜体积, 103 ℃烘干72 h, 用烘干的质量除以鲜体积来测量木材密度; 最大树高来源于《中国植物志》(
Wu & Raven, 1994- 2009 ). ...
... 越来越多的研究表明, 生态系统过程是由物种的功能特性、物种的丰富度和物种的分布共同决定的(
McGill et al ., 2006 ;
Mouchet et al ., 2010 ), 同时森林生产力随多样性的增加而增加(
Ruiz-Benito et al ., 2014 ).本研究得出相似的结果: 群落中物种丰富度越高、功能性状差异越大, 其生产力越高(
图1 ).我们的结果同样表明, 相较于物种多样性, 功能多样性与生产力的关系更加密切(
表4 ), 功能多样性比物种多样性更好地解释了该群落生产力的变异.这可能是因为物种数量和物种多样性所包含的关于物种的特征信息少(
Petchey et al ., 2004 ), 或者忽略了物种间功能性状的相似性和差异性(
吕亭亭等, 2014 ). ...
草甸和沼泽植物群落功能多样性与生产力的关系
2
2014
... 本研究测定与植物生长和群落生产力密切相关的8种功能性状: 木材密度、最大树高、比叶面积、叶面积、叶片厚度、叶片干物质含量、叶片碳含量和叶片氮含量(
Finegan et al ., 2015 ;
Van et al ., 2017 ) (
表1 ).对41种木本植物的功能性状进行了测定, 这些物种的生物量总和占群落总生物量的85%以上(
吕亭亭等, 2014 ).每个物种随机选取5株生长良好、无病虫害的植株, 每株剪下20片健康无损、被日光照射无遮阴的叶片, 依据植物性状标准化测定方法(
Cornelissen et al ., 2003 ), 测定6种叶片性状; 在样地周围对每个物种随机选择3株健康植株, 在距离地面1.3 m处钻取一个生长芯, 把生长芯分成1 cm的片段, 用水置换法测定各节段鲜体积, 103 ℃烘干72 h, 用烘干的质量除以鲜体积来测量木材密度; 最大树高来源于《中国植物志》(
Wu & Raven, 1994- 2009 ). ...
... 越来越多的研究表明, 生态系统过程是由物种的功能特性、物种的丰富度和物种的分布共同决定的(
McGill et al ., 2006 ;
Mouchet et al ., 2010 ), 同时森林生产力随多样性的增加而增加(
Ruiz-Benito et al ., 2014 ).本研究得出相似的结果: 群落中物种丰富度越高、功能性状差异越大, 其生产力越高(
图1 ).我们的结果同样表明, 相较于物种多样性, 功能多样性与生产力的关系更加密切(
表4 ), 功能多样性比物种多样性更好地解释了该群落生产力的变异.这可能是因为物种数量和物种多样性所包含的关于物种的特征信息少(
Petchey et al ., 2004 ), 或者忽略了物种间功能性状的相似性和差异性(
吕亭亭等, 2014 ). ...
Rebuilding community ecology from functional traits
2
2006
... 越来越多的研究表明, 生态系统过程是由物种的功能特性、物种的丰富度和物种的分布共同决定的(
McGill et al ., 2006 ;
Mouchet et al ., 2010 ), 同时森林生产力随多样性的增加而增加(
Ruiz-Benito et al ., 2014 ).本研究得出相似的结果: 群落中物种丰富度越高、功能性状差异越大, 其生产力越高(
图1 ).我们的结果同样表明, 相较于物种多样性, 功能多样性与生产力的关系更加密切(
表4 ), 功能多样性比物种多样性更好地解释了该群落生产力的变异.这可能是因为物种数量和物种多样性所包含的关于物种的特征信息少(
Petchey et al ., 2004 ), 或者忽略了物种间功能性状的相似性和差异性(
吕亭亭等, 2014 ). ...
...
Petchey等(2004) 首次证明功能多样性比物种多样性能更有效地预测生物量的变化,
Gazol和Camarero (2016) 指出功能多样性与生态系统生产力的关系非常密切, 功能多样性是解释生态系统功能变化的更有效的指标.相对于物种多样性, 功能多样性有两个优点: 首先, 各物种具有相异的功能特征, 物种多样性无法解释物种间功能特性的差异, 而功能多样性能够综合不同物种的属性(
Micheli & Halpern, 2010 ); 其次, 功能性状可以表征生物从环境中利用资源的能力(
McGill et al ., 2006 ).功能性状的多样性比物种多样性更好地反映不同物种的生态位, 因此能够更有效地揭示生产力的变异规律(
Petchey et al ., 2004 ). ...
Tree species diversity promotes aboveground carbon storage through functional diversity and functional dominance
1
2016
... 众多研究表明, 有关多样性对木本植物生物量及其生产力的作用机理, 既有生物量比率效应也有生态位互补效应(
Ruiz-Benito et al ., 2014 ;
Mensah et al ., 2016 ), 但两种机制在生态系统过程中的作用不相等(
Mokany et al ., 2008 ).本研究中Rao二次熵指数、功能分散指数在解释该群落生产力变异的能力上明显优于群落加权平均值(
图2 ;
表4 ); 在解释多样性-生产力关系的最优结构方程模型中, 两者均与生产力显著相关, 功能多样性指数的总效应(0.383)高于群落加权平均值的总效应(0.242)(
表6 ).说明生态位互补效应和生物量比率效应共存, 前者更适于解释阔叶红松林生产力的变异. ...
Low functional redundancy in coastal marine assemblages
1
2010
...
Petchey等(2004) 首次证明功能多样性比物种多样性能更有效地预测生物量的变化,
Gazol和Camarero (2016) 指出功能多样性与生态系统生产力的关系非常密切, 功能多样性是解释生态系统功能变化的更有效的指标.相对于物种多样性, 功能多样性有两个优点: 首先, 各物种具有相异的功能特征, 物种多样性无法解释物种间功能特性的差异, 而功能多样性能够综合不同物种的属性(
Micheli & Halpern, 2010 ); 其次, 功能性状可以表征生物从环境中利用资源的能力(
McGill et al ., 2006 ).功能性状的多样性比物种多样性更好地反映不同物种的生态位, 因此能够更有效地揭示生产力的变异规律(
Petchey et al ., 2004 ). ...
Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland
3
2008
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
... 众多研究表明, 有关多样性对木本植物生物量及其生产力的作用机理, 既有生物量比率效应也有生态位互补效应(
Ruiz-Benito et al ., 2014 ;
Mensah et al ., 2016 ), 但两种机制在生态系统过程中的作用不相等(
Mokany et al ., 2008 ).本研究中Rao二次熵指数、功能分散指数在解释该群落生产力变异的能力上明显优于群落加权平均值(
图2 ;
表4 ); 在解释多样性-生产力关系的最优结构方程模型中, 两者均与生产力显著相关, 功能多样性指数的总效应(0.383)高于群落加权平均值的总效应(0.242)(
表6 ).说明生态位互补效应和生物量比率效应共存, 前者更适于解释阔叶红松林生产力的变异. ...
... 群落加权平均值用于评估优势性状对生产力的影响, 而功能多样性指数则是量化该性状的变异性对生产力的影响(
Tobner et al ., 2016 ).在草本植物生态系统中, 群落加权平均值能更好地解释生产力的变异(
Mokany et al ., 2008 ), 而在欧洲地中海森林(
Ratcliffe et al ., 2016 )和广西马尾松(
Pinus massoniana )林(
黄小荣, 2018 )则发现功能多样性指数更好地解释了生产力的变异, 本研究结果与之一致.这种不同群落类型的多样性-生产力关系结果的不一致, 可能是方法上的差异, 包括选择不同的样地监测周期和多样性指数(
Dănescu et al ., 2016 ); 也可能是由不同生态系统类型和演替阶段造成的.当森林的木本植物存在较大的功能性状差异时, 各物种具有不同获取和利用资源的策略, 减少了生态位重叠, 生态位互补效应会减弱物种间对资源的竞争, 最大化地利用光等有限资源, 从而提高生态系统生产力(
Díaz et al ., 2007 ).而在有生境过滤和扩散限制的群落中, 优势物种的影响可能更大, 生态系统过程主要取决于群落中优势物种的功能性状, 此时生物量比率效应更为重要(
Finegan et al ., 2015 ). ...
Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules
1
2010
... 越来越多的研究表明, 生态系统过程是由物种的功能特性、物种的丰富度和物种的分布共同决定的(
McGill et al ., 2006 ;
Mouchet et al ., 2010 ), 同时森林生产力随多样性的增加而增加(
Ruiz-Benito et al ., 2014 ).本研究得出相似的结果: 群落中物种丰富度越高、功能性状差异越大, 其生产力越高(
图1 ).我们的结果同样表明, 相较于物种多样性, 功能多样性与生产力的关系更加密切(
表4 ), 功能多样性比物种多样性更好地解释了该群落生产力的变异.这可能是因为物种数量和物种多样性所包含的关于物种的特征信息少(
Petchey et al ., 2004 ), 或者忽略了物种间功能性状的相似性和差异性(
吕亭亭等, 2014 ). ...
3
2009
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
... 多样性并非生态系统生产力的唯一驱动因素, 它同时还受非生物因素和其他生物因素的影响, 如初始生物量、土壤养分等环境因子(
Lin et al ., 2012 ;
Chen et al ., 2016 ).有研究认为, 林分的初始地上生物量是森林生产力的关键驱动因素, 是植被数量(即初始生物量)而不是植被质量(即多样性)对生产力变异起着主要作用(
Lohbeck et al ., 2015 ).研究发现单纯考虑多样性与生产力二元关系时, 物种丰富度显著影响生产力, 而整合考虑土壤肥力等非生物因子的作用后, 二者关系变得不明显(
Grace et al ., 2016 ); 与此相反, 当去除其他非生物(海拔、冠层光照)和生物因子(树干密度、森林演替阶段)后, 我国亚热带森林中多样性-生产力正相关关系变弱甚至不相关(
Ouyang et al ., 2016 ); 有研究表明, 土壤因素推动了生产力的变异, 而生态位互补和生物量比率机制在天然林中的重要性却微不足道(
Van et al ., 2017 ).综上来看, 生物因素(植物多样性、初始生物量等)与非生物因素(土壤、地形等)的相对重要性仍需进一步验证. ...
... 采用R 3.5.1 vegan包的diversity函数(
Oksanen et al ., 2009 ), 计算每个样方的物种多样性指标; FD包计算功能多样性指数与群落加权平均值(
Laliberté & Legendre, 2010 )(
表3 ). ...
Significant effects of biodiversity on forest biomass during the succession of subtropical forest in south China
2
2016
... 越来越多的研究表明, 生态系统过程是由物种的功能特性、物种的丰富度和物种的分布共同决定的(
McGill et al ., 2006 ;
Mouchet et al ., 2010 ), 同时森林生产力随多样性的增加而增加(
Ruiz-Benito et al ., 2014 ).本研究得出相似的结果: 群落中物种丰富度越高、功能性状差异越大, 其生产力越高(
图1 ).我们的结果同样表明, 相较于物种多样性, 功能多样性与生产力的关系更加密切(
表4 ), 功能多样性比物种多样性更好地解释了该群落生产力的变异.这可能是因为物种数量和物种多样性所包含的关于物种的特征信息少(
Petchey et al ., 2004 ), 或者忽略了物种间功能性状的相似性和差异性(
吕亭亭等, 2014 ). ...
...
Petchey等(2004) 首次证明功能多样性比物种多样性能更有效地预测生物量的变化,
Gazol和Camarero (2016) 指出功能多样性与生态系统生产力的关系非常密切, 功能多样性是解释生态系统功能变化的更有效的指标.相对于物种多样性, 功能多样性有两个优点: 首先, 各物种具有相异的功能特征, 物种多样性无法解释物种间功能特性的差异, 而功能多样性能够综合不同物种的属性(
Micheli & Halpern, 2010 ); 其次, 功能性状可以表征生物从环境中利用资源的能力(
McGill et al ., 2006 ).功能性状的多样性比物种多样性更好地反映不同物种的生态位, 因此能够更有效地揭示生产力的变异规律(
Petchey et al ., 2004 ). ...
How do different measures of functional diversity perform?
2
2004
...
Petchey等(2004) 首次证明功能多样性比物种多样性能更有效地预测生物量的变化,
Gazol和Camarero (2016) 指出功能多样性与生态系统生产力的关系非常密切, 功能多样性是解释生态系统功能变化的更有效的指标.相对于物种多样性, 功能多样性有两个优点: 首先, 各物种具有相异的功能特征, 物种多样性无法解释物种间功能特性的差异, 而功能多样性能够综合不同物种的属性(
Micheli & Halpern, 2010 ); 其次, 功能性状可以表征生物从环境中利用资源的能力(
McGill et al ., 2006 ).功能性状的多样性比物种多样性更好地反映不同物种的生态位, 因此能够更有效地揭示生产力的变异规律(
Petchey et al ., 2004 ). ...
... 群落加权平均值用于评估优势性状对生产力的影响, 而功能多样性指数则是量化该性状的变异性对生产力的影响(
Tobner et al ., 2016 ).在草本植物生态系统中, 群落加权平均值能更好地解释生产力的变异(
Mokany et al ., 2008 ), 而在欧洲地中海森林(
Ratcliffe et al ., 2016 )和广西马尾松(
Pinus massoniana )林(
黄小荣, 2018 )则发现功能多样性指数更好地解释了生产力的变异, 本研究结果与之一致.这种不同群落类型的多样性-生产力关系结果的不一致, 可能是方法上的差异, 包括选择不同的样地监测周期和多样性指数(
Dănescu et al ., 2016 ); 也可能是由不同生态系统类型和演替阶段造成的.当森林的木本植物存在较大的功能性状差异时, 各物种具有不同获取和利用资源的策略, 减少了生态位重叠, 生态位互补效应会减弱物种间对资源的竞争, 最大化地利用光等有限资源, 从而提高生态系统生产力(
Díaz et al ., 2007 ).而在有生境过滤和扩散限制的群落中, 优势物种的影响可能更大, 生态系统过程主要取决于群落中优势物种的功能性状, 此时生物量比率效应更为重要(
Finegan et al ., 2015 ). ...
Modes of functional biodiversity control on tree productivity across the European continent
2
2016
... 植物多样性影响生态系统功能的机理存在两种假说.一是“生态位互补假说” (
Tilman, 1997 ), 认为随着多样性的增加, 群落内树种的功能特性差异增大, 如叶面积、冠层高度、生长速度等, 能够缓解竞争.通常用多样性指数(如物种、功能多样性)来表征该假说.二是“生物量比率假说” (
Grime, 1998 ), 认为某个时间点的生态系统功能主要由植物优势物种的功能性状值决定.当该假说占主导时, 可以检测到
CWM 是解释生态系统功能的最重要因子(
Tobner et al ., 2016 ;
Ali et al ., 2017 ).尽管目前研究普遍认为生态位互补与生物量比率假说并不相互排斥, 但二者的相对重要性还存在很大争论.
Cardinale和Gonzalez (2011) 整合分析多样性种植实验结果, 发现两种假说对生产力变异的解释基本持平;
Ali等(2017) 则发现生物量比率假说在解释中国东部亚热带次生林的生产力变异中占主导地位;
Ratcliffe等(2016) 发现在欧洲温带森林中生态位互补和生物量比率效应对树木生长均有促进作用, 但在水分受限制的地中海森林中生态位互补假说能够更好地解释生产力的变异. ...
... 根据变量筛选结果, 选择变量重要性最高、与生产力最为相关的多样性指数与潜在环境变量, 纳入到结构方程模型中.采用最大似然法对结构方程模型进行拟合, 利用比较拟合指数(
CFI )、标准均方根残差(
SRMR )、渐进残差均方和平方根(
RMSEA )、显著性概率值(
p )进行模型优度的评价(
Howard, 2013 ).拟合优度的临界值为
CFI > 0.9,
SRMR < 0.05,
RMSEA < 0.08,
p > 0.05.利用模型中各路径标准化系数, 定量表示不同因素对生产力的相对作用大小, 量化各个变量对森林地上生产力的直接、间接和总标准化效应(
Edwards & Lambert, 2007 ), 从而比较生态位互补假说与生物量比率假说对阔叶红松林的适用性.上述分析利用lavaan包完成(
Rosseel, 2012 ). ...
lavaan: An R package for structural equation modeling
3
2012
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
... 越来越多的研究表明, 生态系统过程是由物种的功能特性、物种的丰富度和物种的分布共同决定的(
McGill et al ., 2006 ;
Mouchet et al ., 2010 ), 同时森林生产力随多样性的增加而增加(
Ruiz-Benito et al ., 2014 ).本研究得出相似的结果: 群落中物种丰富度越高、功能性状差异越大, 其生产力越高(
图1 ).我们的结果同样表明, 相较于物种多样性, 功能多样性与生产力的关系更加密切(
表4 ), 功能多样性比物种多样性更好地解释了该群落生产力的变异.这可能是因为物种数量和物种多样性所包含的关于物种的特征信息少(
Petchey et al ., 2004 ), 或者忽略了物种间功能性状的相似性和差异性(
吕亭亭等, 2014 ). ...
... 众多研究表明, 有关多样性对木本植物生物量及其生产力的作用机理, 既有生物量比率效应也有生态位互补效应(
Ruiz-Benito et al ., 2014 ;
Mensah et al ., 2016 ), 但两种机制在生态系统过程中的作用不相等(
Mokany et al ., 2008 ).本研究中Rao二次熵指数、功能分散指数在解释该群落生产力变异的能力上明显优于群落加权平均值(
图2 ;
表4 ); 在解释多样性-生产力关系的最优结构方程模型中, 两者均与生产力显著相关, 功能多样性指数的总效应(0.383)高于群落加权平均值的总效应(0.242)(
表6 ).说明生态位互补效应和生物量比率效应共存, 前者更适于解释阔叶红松林生产力的变异. ...
Diversity increases carbon storage and tree productivity in Spanish forests
1
2014
... 在相对资源受限的环境中, 各物种采取不同的资源吸收策略才能共存, 此时互补效应为生产力的主要驱动力.在生物量较高的地区, 丰富的资源导致互补效应的相对重要性降低, 多样性对生产力的影响也将减弱(
Lohbeck et al ., 2015 ).当群落中出现非生物过滤(如环境过滤)时, 只有适应相应环境条件的物种才能够生存, 导致物种间的性状差异减少, 形成性状聚集(
Schellberg & Pontes, 2012 ), 致使局域范围内群落的优势物种表现出强烈的趋同现象.因此, 生物多样性并非生态系统生产力的唯一驱动因素, 而是受非生物因素和其他生物因素的共同影响(
Lohbeck et al ., 2015 ). ...
Plant functional traits and nutrient gradients on grassland
2012
2
2002
... 植物多样性影响生态系统功能的机理存在两种假说.一是“生态位互补假说” (
Tilman, 1997 ), 认为随着多样性的增加, 群落内树种的功能特性差异增大, 如叶面积、冠层高度、生长速度等, 能够缓解竞争.通常用多样性指数(如物种、功能多样性)来表征该假说.二是“生物量比率假说” (
Grime, 1998 ), 认为某个时间点的生态系统功能主要由植物优势物种的功能性状值决定.当该假说占主导时, 可以检测到
CWM 是解释生态系统功能的最重要因子(
Tobner et al ., 2016 ;
Ali et al ., 2017 ).尽管目前研究普遍认为生态位互补与生物量比率假说并不相互排斥, 但二者的相对重要性还存在很大争论.
Cardinale和Gonzalez (2011) 整合分析多样性种植实验结果, 发现两种假说对生产力变异的解释基本持平;
Ali等(2017) 则发现生物量比率假说在解释中国东部亚热带次生林的生产力变异中占主导地位;
Ratcliffe等(2016) 发现在欧洲温带森林中生态位互补和生物量比率效应对树木生长均有促进作用, 但在水分受限制的地中海森林中生态位互补假说能够更好地解释生产力的变异. ...
... 计算每个样方的物种多样性, 包括物种丰富度(
C )、辛普森指数(
D )、香农指数(
H´ )、均匀度指数(
J ), 各指数计算公式(
孙儒泳等, 2002 )如下: ...
2
2002
... 植物多样性影响生态系统功能的机理存在两种假说.一是“生态位互补假说” (
Tilman, 1997 ), 认为随着多样性的增加, 群落内树种的功能特性差异增大, 如叶面积、冠层高度、生长速度等, 能够缓解竞争.通常用多样性指数(如物种、功能多样性)来表征该假说.二是“生物量比率假说” (
Grime, 1998 ), 认为某个时间点的生态系统功能主要由植物优势物种的功能性状值决定.当该假说占主导时, 可以检测到
CWM 是解释生态系统功能的最重要因子(
Tobner et al ., 2016 ;
Ali et al ., 2017 ).尽管目前研究普遍认为生态位互补与生物量比率假说并不相互排斥, 但二者的相对重要性还存在很大争论.
Cardinale和Gonzalez (2011) 整合分析多样性种植实验结果, 发现两种假说对生产力变异的解释基本持平;
Ali等(2017) 则发现生物量比率假说在解释中国东部亚热带次生林的生产力变异中占主导地位;
Ratcliffe等(2016) 发现在欧洲温带森林中生态位互补和生物量比率效应对树木生长均有促进作用, 但在水分受限制的地中海森林中生态位互补假说能够更好地解释生产力的变异. ...
... 计算每个样方的物种多样性, 包括物种丰富度(
C )、辛普森指数(
D )、香农指数(
H´ )、均匀度指数(
J ), 各指数计算公式(
孙儒泳等, 2002 )如下: ...
Distinguishing between the effects of species diversity and species composition
2
1997
... 植物多样性影响生态系统功能的机理存在两种假说.一是“生态位互补假说” (
Tilman, 1997 ), 认为随着多样性的增加, 群落内树种的功能特性差异增大, 如叶面积、冠层高度、生长速度等, 能够缓解竞争.通常用多样性指数(如物种、功能多样性)来表征该假说.二是“生物量比率假说” (
Grime, 1998 ), 认为某个时间点的生态系统功能主要由植物优势物种的功能性状值决定.当该假说占主导时, 可以检测到
CWM 是解释生态系统功能的最重要因子(
Tobner et al ., 2016 ;
Ali et al ., 2017 ).尽管目前研究普遍认为生态位互补与生物量比率假说并不相互排斥, 但二者的相对重要性还存在很大争论.
Cardinale和Gonzalez (2011) 整合分析多样性种植实验结果, 发现两种假说对生产力变异的解释基本持平;
Ali等(2017) 则发现生物量比率假说在解释中国东部亚热带次生林的生产力变异中占主导地位;
Ratcliffe等(2016) 发现在欧洲温带森林中生态位互补和生物量比率效应对树木生长均有促进作用, 但在水分受限制的地中海森林中生态位互补假说能够更好地解释生产力的变异. ...
... 群落加权平均值用于评估优势性状对生产力的影响, 而功能多样性指数则是量化该性状的变异性对生产力的影响(
Tobner et al ., 2016 ).在草本植物生态系统中, 群落加权平均值能更好地解释生产力的变异(
Mokany et al ., 2008 ), 而在欧洲地中海森林(
Ratcliffe et al ., 2016 )和广西马尾松(
Pinus massoniana )林(
黄小荣, 2018 )则发现功能多样性指数更好地解释了生产力的变异, 本研究结果与之一致.这种不同群落类型的多样性-生产力关系结果的不一致, 可能是方法上的差异, 包括选择不同的样地监测周期和多样性指数(
Dănescu et al ., 2016 ); 也可能是由不同生态系统类型和演替阶段造成的.当森林的木本植物存在较大的功能性状差异时, 各物种具有不同获取和利用资源的策略, 减少了生态位重叠, 生态位互补效应会减弱物种间对资源的竞争, 最大化地利用光等有限资源, 从而提高生态系统生产力(
Díaz et al ., 2007 ).而在有生境过滤和扩散限制的群落中, 优势物种的影响可能更大, 生态系统过程主要取决于群落中优势物种的功能性状, 此时生物量比率效应更为重要(
Finegan et al ., 2015 ). ...
Functional identity is the main driver of diversity effects in young tree communities
2
2016
... 多样性并非生态系统生产力的唯一驱动因素, 它同时还受非生物因素和其他生物因素的影响, 如初始生物量、土壤养分等环境因子(
Lin et al ., 2012 ;
Chen et al ., 2016 ).有研究认为, 林分的初始地上生物量是森林生产力的关键驱动因素, 是植被数量(即初始生物量)而不是植被质量(即多样性)对生产力变异起着主要作用(
Lohbeck et al ., 2015 ).研究发现单纯考虑多样性与生产力二元关系时, 物种丰富度显著影响生产力, 而整合考虑土壤肥力等非生物因子的作用后, 二者关系变得不明显(
Grace et al ., 2016 ); 与此相反, 当去除其他非生物(海拔、冠层光照)和生物因子(树干密度、森林演替阶段)后, 我国亚热带森林中多样性-生产力正相关关系变弱甚至不相关(
Ouyang et al ., 2016 ); 有研究表明, 土壤因素推动了生产力的变异, 而生态位互补和生物量比率机制在天然林中的重要性却微不足道(
Van et al ., 2017 ).综上来看, 生物因素(植物多样性、初始生物量等)与非生物因素(土壤、地形等)的相对重要性仍需进一步验证. ...
... 本研究测定与植物生长和群落生产力密切相关的8种功能性状: 木材密度、最大树高、比叶面积、叶面积、叶片厚度、叶片干物质含量、叶片碳含量和叶片氮含量(
Finegan et al ., 2015 ;
Van et al ., 2017 ) (
表1 ).对41种木本植物的功能性状进行了测定, 这些物种的生物量总和占群落总生物量的85%以上(
吕亭亭等, 2014 ).每个物种随机选取5株生长良好、无病虫害的植株, 每株剪下20片健康无损、被日光照射无遮阴的叶片, 依据植物性状标准化测定方法(
Cornelissen et al ., 2003 ), 测定6种叶片性状; 在样地周围对每个物种随机选择3株健康植株, 在距离地面1.3 m处钻取一个生长芯, 把生长芯分成1 cm的片段, 用水置换法测定各节段鲜体积, 103 ℃烘干72 h, 用烘干的质量除以鲜体积来测量木材密度; 最大树高来源于《中国植物志》(
Wu & Raven, 1994- 2009 ). ...
Abiotic and biotic drivers of biomass change in a neotropical forest
2017
2
1994-2009
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
... 本研究测定与植物生长和群落生产力密切相关的8种功能性状: 木材密度、最大树高、比叶面积、叶面积、叶片厚度、叶片干物质含量、叶片碳含量和叶片氮含量(
Finegan et al ., 2015 ;
Van et al ., 2017 ) (
表1 ).对41种木本植物的功能性状进行了测定, 这些物种的生物量总和占群落总生物量的85%以上(
吕亭亭等, 2014 ).每个物种随机选取5株生长良好、无病虫害的植株, 每株剪下20片健康无损、被日光照射无遮阴的叶片, 依据植物性状标准化测定方法(
Cornelissen et al ., 2003 ), 测定6种叶片性状; 在样地周围对每个物种随机选择3株健康植株, 在距离地面1.3 m处钻取一个生长芯, 把生长芯分成1 cm的片段, 用水置换法测定各节段鲜体积, 103 ℃烘干72 h, 用烘干的质量除以鲜体积来测量木材密度; 最大树高来源于《中国植物志》(
Wu & Raven, 1994- 2009 ). ...
Individual size inequality links forest diversity and above-ground biomass
1
2015
... 探索生物多样性与生态系统功能之间的关系已经成为生态学领域的一个核心问题, 尤其是更好地了解天然林地上生产力的驱动因素, 对于森林的可持续经营十分重要(
Cardinale et al ., 2012 ;
Zhang & Chen, 2015 ).众多的研究表明, 生物多样性对生态系统功能的影响, 不仅归因于物种数量, 还依赖于物种种类及物种所具有的功能特性(
Mokany et al ., 2008 ).关于哪些多样性指标与地上生物量和生产力有关, 以及应如何量化, 一直存在争议(
Díaz et al ., 2007 ;
Ouyang et al ., 2016 ).以物种丰富度为代表的物种多样性是最简单的多样性衡量标准, 通常被用作地上生物量和生产力的生物预测指标, 但研究表明其只能解释生产力的较小一部分变异(
Zhang et al ., 2012 ;
Ruiz-Benito et al ., 2014 ).功能多样性是指植物群落中功能性状的值、范围、分布或分散程度(
Laliberté & Legendre, 2010 ), 主要包括功能多样性指数与群落加权平均值(
CWM )两部分(
黄小荣, 2018 ).研究表明, 相较于物种多样性, 功能性状多样性是解释生态系统功能变化更有效的指标, 其与生态系统生产力的关系更为密切(
Gazol & Camarero, 2016 ). ...
Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis
2012
 ,
, ,1,2,*
,1,2,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT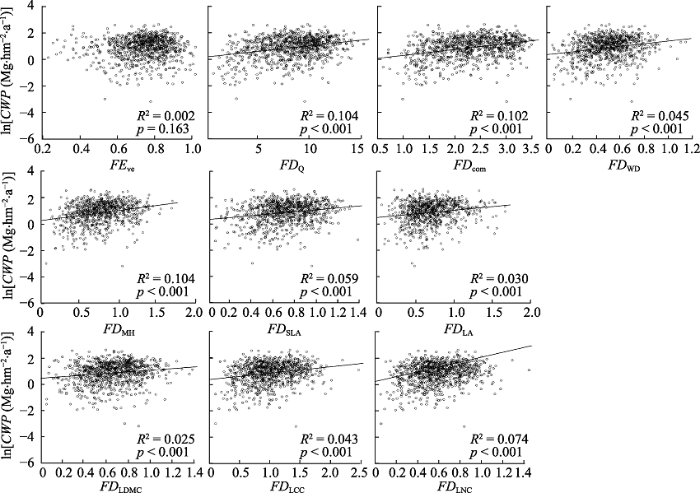 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT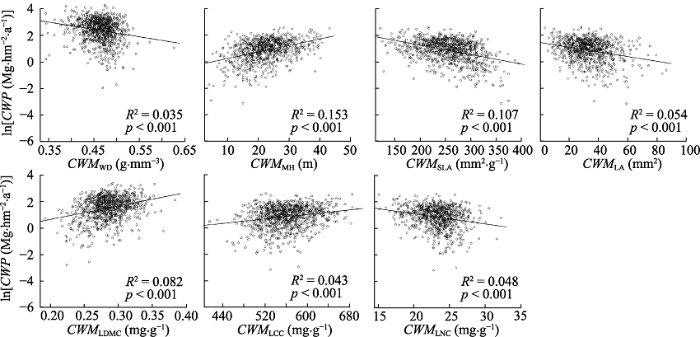 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT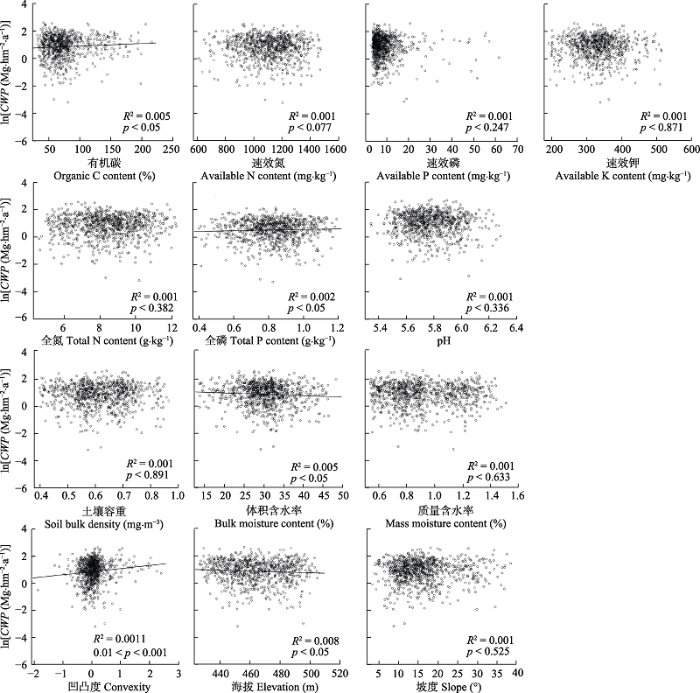 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT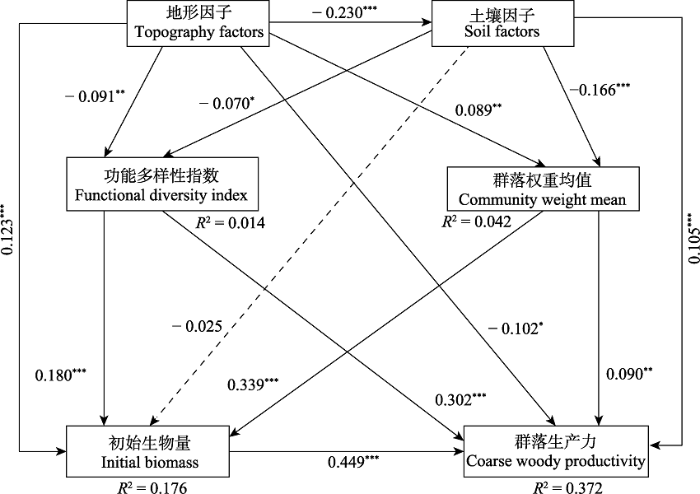 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}