 ,1,*
,1,*Decomposition of different root branch orders and its dominant controlling factors in four temperate tree species
GU Wei-Ping1, LIU Rui-Peng1, LI Xing-Huan1, SUN Tao2, ZHANG Zi-Jia2, ZAN Peng1, WEN Lu-Ning1, MA Peng-Yu1, MAO Zi-Jun,1,*通讯作者: E-mail:jshe@pku.edu.cn
编委: 王政权
责任编辑: 李 敏
收稿日期:2018-04-10修回日期:2018-09-10网络出版日期:2018-09-20
| 基金资助: |
Received:2018-04-10Revised:2018-09-10Online:2018-09-20
| Fund supported: |

摘要
细根分解是森林生态系统土壤碳和养分的主要输入途径, 但目前人们对于影响细根分解的主要因素和细根分解模式的了解仍然很少。该研究采用根序划分等级方法, 将红松(Pinus koraiensis)、落叶松(Larix gmelinii)、水曲柳(Fraxinus mandschurica)和白桦(Betula platyphylla)细根组分前四级根划分为两个等级: 一级和二级根混合为低级根, 三级和四级根混合为高级根。利用埋袋法对东北地区4个树种不同根序细根进行连续4年的分解实验, 并对其分解速率以及影响因素进行研究。结果显示, 红松低级根和高级根分解系数分别为0.342和0.461, 落叶松依次分别为0.304和0.436, 水曲柳分别为0.450和0.555, 白桦为0.441和0.579。4个树种均显示低级根分解速率较慢, 而高级根分解速率较快。实验表明, 根系分解系数与酸不溶性物质(AUF)和非结构性碳水化合物(TNC)均具有显著相关性。出现上述结果的主要原因是低级根含有较多的AUF, 很难被分解, 以及含有较少的TNC, 为分解者提供能量较少。
关键词:
Abstract
Keywords:
PDF (1009KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
顾伟平, 刘瑞鹏, 李兴欢, 孙涛, 张子嘉, 昝鹏, 温璐宁, 马鹏宇, 毛子军. 四个典型温带树种不同根序细根分解速率及其主要影响因素. 植物生态学报[J], 2018, 42(9): 955-962 DOI:10.17521/cjpe.2018.0080
GU Wei-Ping, LIU Rui-Peng, LI Xing-Huan, SUN Tao, ZHANG Zi-Jia, ZAN Peng, WEN Lu-Ning, MA Peng-Yu, MAO Zi-Jun.
在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(Wardle et al., 2004)。因此, 明确凋落物分解的一般机制对准确预测未来全球变化与陆地碳循环之间的相互作用具有重要意义(Aerts, 2006; Cornwell et al., 2008)。目前, 国内外对地上凋落物分解已进行了较为广泛的研究, 而有关地下凋落物的研究却很少, 如对细根分解的研究。通过细根分解输送给土壤的碳(C)是地上凋落物分解的4-5倍(宋森等, 2008), 细根分解释放的氮(N)也比地上部分高18%-45% (Vogt et al., 1986)。可见, 细根分解是C和养分归还土壤的主要途径(Dornbush et al., 2002)。已发表的文章表明, 细根分解的机理与以往研究有较大差异, Silver和Miya (2001)研究发现细根中的化学物质是与细根分解速率最密切相关的因子。其他研究表明, 根系初始N浓度、Ca浓度以及C:N对细根分解速率没有影响(Hobbie et al., 2010; Goebel et al., 2011; Sun et al., 2013a, 2013b)。可见, 对于细根分解的机理及其影响因子尚缺乏足够的认识。
近半个世纪以来, 大部分研究都把细根定义为一个均质系统, 即认为≤2 mm的根在结构和功能上基本相同。然而, 近年来越来越多的证据表明, 以这种单一径级定义细根的方法忽略了细根形态、化学以及生理方面的差异(Pregitzer et al., 2002; Guo et al., 2008a, 2008b; 刘莹等, 2010; McCormack et al., 2015; Xia et al., 2015)。专家研究发现, 树木根系具有复杂的分枝系统(即根序), 不同根序等级的个体根在形态结构、解剖构造和生理功能等方面存在显著差异(Pregitzer et al., 2002; Guo et al., 2008b)。根序等级的划分为: 根尖为1级根, 其母根为2级根, 以此类推(Pregitzer et al., 2002; 谷加存等, 2016)。因此, 不同根序等级的组织化学成分(如C:N、N、木质素、纤维素和酚类化合物的浓度)可能存在很大的差异。
现有文献资料表明, 迄今为止仅有少量已发表的文章量化了根序和分解速率之间的关系。有研究发现, 尽管低级根具有较小的直径, 高级根具有较大的直径并且木质化程度较高, 但低级根仍然比高级根分解速率低(Fan & Guo, 2010; Goebel et al., 2011; Xiong et al., 2012; Sun et al., 2016)。有研究表明, 非木质的、营养丰富的低级根, 尤其是根尖, 具有较高的酸不溶性化合物含量, 而高级根则相反。 这种难分解的酸不溶性化合物是导致低级根分解速率变慢的主要因素(Guo et al., 2004; Fan & Guo, 2010; Xiong et al., 2012; Sun et al., 2015, 2016)。为了解释这些现象的共性, 显然需要更多的实验研究来探索低级根分解缓慢的潜在机制。
为了探讨上述研究结果及其结论是否具有普遍意义, 本研究利用根序法划分细根等级(根尖为1级根, 其母根为2级根, 以此类推), 将1级根和2级根混合为低级根, 3级根和4级根混合为高级根。本研究采用分解袋方法, 对中国东北温带森林中4个常见的树种(红松(Pinus koraiensis)、落叶松(Larix gmelinii)、水曲柳(Fraxinus mandschurica)和白桦(Betula platyphylla))进行连续4年的细根分解实验, 探索4个树种的不同根序分解速率的规律, 以及根系当中初始化学成分对细根分解的影响。我们假设低级根分解慢, 而高级根分解快, 其主要原因是低级根含有较多难分解的酸不溶性物质。本研究旨在了解不同根序分解速率及其影响因素, 为探讨细根对森林生态系统C和养分循环提供理论依据。
1 研究地区和研究方法
1.1 研究区概况
分解实验样地设在黑龙江省东北林业大学帽儿山实验林场老山森林实验站(127.50°-127.57° E, 45.33°-45.42° N) 4个典型的温带森林群落中。当地气候条件受大陆性季风气候的影响, 季风性强, 春天风大, 夏天高温潮湿, 冬天干燥寒冷。年降水量730 mm, 主要集中在7、8月份。年平均气温2.8 ℃, 月平均最高气温20.9 ℃, 月平均最低气温-19.6 ℃。林下土壤为暗棕色森林土壤。在杨-桦林(优势种为山杨(Populus davidiana)和白桦)、落叶阔叶林(优势种为水曲柳、胡桃楸(Juglans mandshurica))、红松人工林(优势种为红松)以及落叶松人工林(优势种为落叶松) 4种温带森林内分别对白桦、水曲柳、落叶松、红松的细根分解进行了定量评估。本研究的4个森林类型的特征见表1。Table 1
表1
表14种森林类型的林分特征
Table 1
| 林型 Forest type | T10 (℃) | W10 (g·g-1) | 林龄 Age (a) | 密度 Density (trees·hm-2) | DBH (cm) | 主要树种 Dominant tree species |
|---|---|---|---|---|---|---|
| 落叶松人工林 Larix gmelinii plantation | 11.6 | 0.396 | 40 | 1 682 | 18.7 | 落叶松 Larix gmelinii |
| 红松人工林 Pinus koraiensis plantation | 10.8 | 0.458 | 46 | 2 904 | 16.1 | 红松 Pinus koraiensis |
| 杨-桦林 Aspen-birch forest | 12.9 | 0.552 | 58 | 2 381 | 12.9 | 山杨、白桦 Populus davidiana, Betula platyphylla |
| 阔叶林 Hardwood forest | 12.3 | 0.579 | 49 | 2 017 | 15.6 | 水曲柳、胡桃楸 Fraxinus mandschurica, Juglans mandshurica |
新窗口打开|下载CSV
1.2 实验设计
本实验采用分解袋法研究细根分解。于2009年, 在林地里选择5-8株目标树种挖取树根。在目标树种的树干基部找到主根, 然后顺着主根向外2 m左右范围内用铲子挖取枯枝落叶层和15 cm土壤深度之间的土块, 顺主根上的侧根寻找根序分支, 确保细根为目标树种的细根。然后小心地把根系从土块中分离出来, 尽量保持根系完整性, 确保根系末端最小的根尖完整并且着生在它的母根上。根系采集主要采用手工挖掘。清除死根(颜色发暗且无弹性)以及根表面土壤后, 尽快运回实验室, 清洗后低温储存, 以便后期处理。采用与Pregitzer等(2002)相同的根序划分方法, 识别并分离出前四级根。本实验将一级和二级细根混合为低级根, 三级和四级细根混合为高级根。并将每个等级的根序额外分出100根左右, 分别测量一级至四级根的平均直径(表2)。Table 2
表2
表24个温带树种一级至四级根平均直径(平均值±标准误差)
Table 2
| 树种 Tree species | 根直径 Root diameter (mm) | |||
|---|---|---|---|---|
| 一级 First-order | 二级 Second-order | 三级 Third-order | 四级 Fourth-order | |
| 红松 Pinus koraiensis | 0.29 ± 0.01 | 0.29 ± 0.02 | 0.53 ± 0.03 | 1.59 ± 0.12 |
| 落叶松 Larix gmelinii | 0.26 ± 0.02 | 0.28 ± 0.01 | 0.46 ± 0.05 | 0.85 ± 0.09 |
| 水曲柳 Fraxinus mandschurica | 0.34 ± 0.01 | 0.45 ± 0.01 | 0.55 ± 0.02 | 0.89 ± 0.07 |
| 白桦 Betula platyphylla | 0.21 ± 0.01 | 0.26 ± 0.03 | 0.27 ± 0.06 | 0.43 ± 0.09 |
新窗口打开|下载CSV
将所有细根样品置于65 ℃恒温烘箱中烘干至恒质量。称取每个根序等级的样品大约260 mg置于凋落物分解袋内(尼龙网, 10 cm × 10 cm, 网眼大小120 μm)。每个根序等级的细根装15袋, 2个等级(高级根和低级根)共装30袋, 4个树种总共120袋。在许多野外的凋落物分解实验中, 分解袋的网眼大小非常重要(Hobbie et al., 2006)。本实验选择的分解袋网眼大小能够阻止蚯蚓以及一些其他微小无脊椎动物的进入, 以使我们的研究主要针对微生物对细根分解的作用。
于2010年5月, 选取红松林、落叶松林、水曲柳林和白桦林样地。在每个样地内设置5个3 m × 3 m的样方, 每个样方内大概有5-8株目标树种, 样方距离15-20 m。每个样方内埋2个根序等级的根各3袋, 平埋于10 cm深的土层中, 表层覆盖林地凋落物, 尽量恢复原样, 以减少对分解环境的影响。做好标记, 以便取样。
1.3 数据采集及样品测定
本实验分别于2010年7月和10月、2011-2013年10月连续4年进行取样, 总共取5次样品, 分别在分解实验开始后的第69天、154天、513天、870天以及第1228天。每次取样的时候每个根序等级样品各取3袋, 小心去除附着的泥土杂物。在收集分解袋的过程中, 没有发现目标树种的根系生长到分解袋内部, 但偶尔会发现草根生长到了分解袋内部。基于它们明显的形态、结构以及颜色的不同, 利用镊子很容易将其去除。然后将根系样品置于65 ℃烘箱内烘干至恒质量并称量。在根系分解实验开始的时候, 将剩余的烘干初始样品磨碎并测定其初始化学物质。利用浓硫酸-重铬酸钾氧化法测定全C含量; 利用半微量凯氏定氮法测定全N含量; 用钼锑抗比色法测定全P含量; 采用火焰光度计法测定全K含量; 用有机质近似C组分分析法(Hendricks et al., 2002)测定酸不溶性物质(AUF)含量; 根据Seifter等(1950)的技术分析了根系样品的总非结构性碳水化合物(TNC)的浓度。
1.4 数据处理
采用根系的质量残留率来表征根系分解, 即各阶段的根系干质量占初始干质量的比例。根系分解率常数(k)采用负指数衰减模型(温达志等, 1998)来进行计算, 公式为:X = e-kt
式中: X为分解时间t (单位: a)时的质量残留率(%), k为年分解速率常数(kg·kg-1·a-1)。
采用Origin Pro 8.0软件对k和初始化学成分含量进行线性回归分析。利用Origin Pro 8.0软件画图。
2 结果和分析
2.1 细根分解速率
本研究的4个典型温带树种中, 根系分解质量残留率随时间的延长总体呈现低级根高于高级根的趋势(图1)。每个树种细根分解到513天的时候, 红松和落叶松低级根分解了接近36%, 而高级根则分解了45%左右; 水曲柳和白桦低级根分解了45%左右, 而高级根则分解了近54%, 每个树种的高级根均比低级根多分解9%左右。分解1228天后, 水曲柳和白桦低级根分解量相似, 分解了近76%, 高级根分解了82%左右, 两树种高级根均比低级根多分解6%; 红松低级根分解了64%, 比高级根少分解20%; 落叶松低级根分解了60%, 比高级根少分解13%。图1
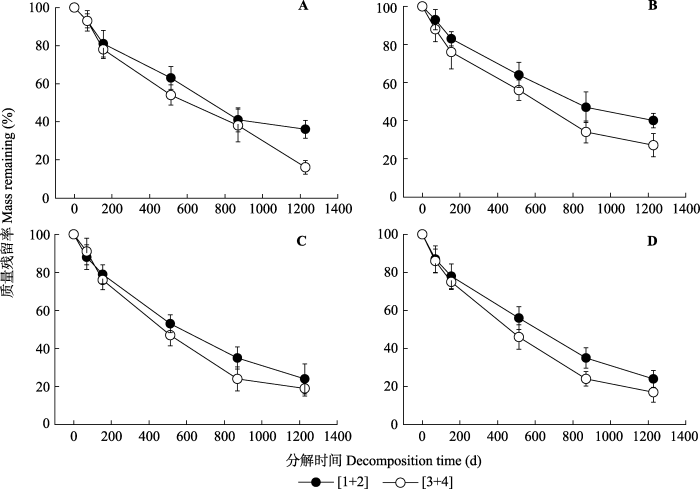 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图14个树种不同根序不同分解时间的质量残留率(平均值±标准误差)。[1+2]是指1-2级根的合并, [3+4]是指3-4级根的合并。A, 红松。B, 落叶松。C, 水曲柳。D, 白桦。
Fig. 1Mass remaining (%) of roots of different branch orders of the four tree species (mean ± SE). [1+2] refers to root orders 1 to 2, [3+4] refers to root orders 3 to 4. A, Pinus koraiensis. B, Larix gmelinii. C, Fraxinus mandschurica. D, Betula platyphylla.
采用负指数衰减模型计算的结果(表3)与实际分解结果(图1)相一致, 所有树种的低级根k值普遍小于高级根, 细根k值在根序间差异显著(p < 0.01)。说明4个树种低级根的分解率均低于高级根(图1; 表3)。
Table 3
表3
表3每个树种根系的分解常数(k)和相关系数(R2)
Table 3
| 根序 Root order | 红松 Pinus koraiensis | 落叶松 Larix gmelinii | 水曲柳 Fraxinus mandschurica | 白桦 Betula platyphylla | ||||
|---|---|---|---|---|---|---|---|---|
| k | R2 | k | R2 | k | R2 | k | R2 | |
| [1+2] | 0.342 | 0.983 | 0.304 | 0.985 | 0.450 | 0.993 | 0.441 | 0.987 |
| [3+4] | 0.461 | 0.987 | 0.436 | 0.980 | 0.555 | 0.995 | 0.579 | 0.994 |
新窗口打开|下载CSV
2.2 根系初始化学成分
根系中初始化学成分浓度常常被用来分析根系分解速率。本研究中4个温带树种根系的初始化学成分浓度在不同根序之间存在较大差异(表4)。从表中可以看出, 4个树种[1+2]级根中N、P、K初始浓度普遍高于[3+4]级根。例如[1+2]级根中初始N浓度几乎是[3+4]级根的两倍; 而C:N则呈现出相反的趋势; 然而, 值得注意的是4个树种的[1+2]级根中AUF的浓度始终高于[3+4]级根; 此外, [1+2]级根中的TNC的浓度普遍低于[3+4]级根。Table 4
表4
表4四个温带树种的细根分解袋实验的初始化学参数(平均值±标准误差)
Table 4
| 树种 Tree Species | 红松 Pinus koraiensis | 落叶松 Larix gmelinii | 水曲柳 Fraxinus mandschurica | 白桦 Betula platyphylla | ||||
|---|---|---|---|---|---|---|---|---|
| 根序 Root order | [1+2] | [3+4] | [1+2] | [3+4] | [1+2] | [3+4] | [1+2] | [3+4] |
| N (mg·g-1) | 21.6 ± 0.9 | 12.3 ± 0.5 | 25.5 ± 2.3 | 13.9 ± 1.2 | 31.9 ± 1.8 | 18.3 ± 1.3 | 28.1 ± 1.6 | 16.7 ± 1.1 |
| P (mg·g-1) | 1.9 ± 0.4 | 1.4 ± 0.4 | 2.8 ± 0.2 | 1.9 ± 0.2 | 2.5 ± 0.1 | 1.7 ± 0.2 | 2.2 ± 0.1 | 1.4 ± 0.2 |
| K (mg·g-1) | 4.4 ± 0.5 | 2.9 ± 0.1 | 5.8 ± 0.2 | 4.5 ± 0.3 | 4.0 ± 0.1 | 4.9 ± 0.3 | 5.3 ± 0.6 | 3.7 ± 0.3 |
| AUF (mg·g-1) | 528.5 ± 40.1 | 398.6 ± 32.9 | 513.4 ± 38.7 | 435.8 ± 37.5 | 401.7 ± 29.6 | 289.3 ± 26.4 | 397.5 ± 34.1 | 302.1 ± 28.2 |
| TNC (mg·g-1) | 101.7 ± 10.9 | 150.2 ± 18.4 | 98.3 ± 9.6 | 138.0 ± 14.1 | 195.8 ± 20.6 | 279.1 ± 23.5 | 164.9 ± 18.3 | 214.6 ± 21.4 |
| C:N | 29.6 ± 1.4 | 48.2 ± 1.9 | 23.6 ± 1.8 | 45.2 ± 3.1 | 14.4 ± 2.1 | 23.7 ± 2.4 | 18.7 ± 2.0 | 34.8 ± 3.1 |
新窗口打开|下载CSV
2.3 根系分解速率与初始化学成分的关系
4个树种[1+2]和[3+4]级根的k与所有树种的根系初始AUF浓度呈显著负相关关系(R2 = 0.945, p < 0.001, 图2)。表明AUF浓度越高, k越小, 即分解速率越低。k与初始TNC浓度呈正相关关系(R2 = 0.756, p < 0.001), 表明k随TNC浓度的增高而变大, 即分解速率变高。然而, 每个树种中[1+2]级根系初始AUF浓度普遍高于[3+4]级根(表4), 而TNC浓度呈相反的趋势。未观察到根系k值与N、P、K、C:N之间显著的相关性(数据未展示)。数据表明, 根系初始化学成分AUF浓度和TNC浓度是影响根系分解的主要因素, 其中AUF浓度具有更显著的相关性(图2)。图2
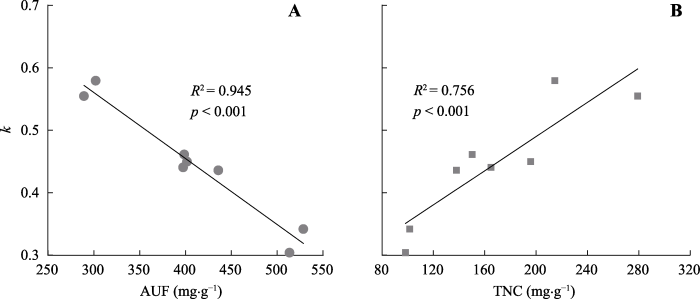 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2所有树种根系分解常数(k)与酸不溶性物质(AUF)(A)和非结构性碳水化合物(TNC)(B)之间的线性回归关系。
Fig. 2Regression relation between root decay constants (k) and root initial acid-unhydrolyzable fraction (AUF)(A) and total non-structural carbohydrat (TNC)(B) concentrations across all species.
3 讨论
在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率。而三四级根则部分已经木质化, 负责运输和贮藏等功能(Guo et al., 2008b; Xia et al., 2010; McCormack et al., 2015)。由于低级根和高级根功能上的不同, 导致根系化学成分产生差异, 并可能对细根分解速率产生影响(McCormack et al., 2015)。有研究表明, 非木质的、营养丰富的低级根, 尤其是根尖, 具有较多难分解的AUF。因此, 认为低级根分解较慢, 高级根分解较快(Guo et al., 2004; Fan & Guo, 2010; Xiong et al., 2012; Sun et al., 2015, 2016)。本研究的4个树种细根AUF浓度越高分解速率越低的结果与此相一致(图1; 表3)。其中Fan和Guo (2010)研究了水曲柳和落叶松不同根序的细根分解, 17个月后他们发现: 两个树种的一二级根的质量损失率普遍低于三四级根。而Goebel等(2011)对4个树种进行了36个月的研究发现, 三级和四级根的质量损失率为20%-40%, 而一级和二级根的质量损失率则不超过20%。Sun等(2016)对落叶松的实验研究显示, 三四级细根质量损失率比一二级细根高6%。可见不同根序的细根分解过程中, 普遍表现为低级根分解慢, 高级根分解快这一分解趋势。进一步分析表明, 直径越小的低级根根皮(皮层)组织所占比例越大, 根皮细胞壁中单宁等次生防御物质的含量越高(Chen et al., 2001; 张秀娟等, 2006)。AUF是难分解的有机高分子结构体(李晗等, 2015), 包括木质素、缩合单宁、角质、蜡质等物质。本研究结果表明导致低级根分解慢, 而高级根分解快的机制之一是低级根含有较多难分解的AUF。此外, 以直径划分等级的研究也发现, 直径越小, 分解速率越慢。谷利茶等(2017)研究油松细根分解发现, 分解到300天的时候, 直径为0-0.4、0.4-1、1-2 mm的细根各分解了19.8%、23.5%、30.5%。因此, 根序等级越低分解越慢似乎是目前细根分解研究的常见结论。另外, 根系分解受到微生物调控的影响(Magill & John, 1998)。在分解过程中, 微生物会利用根系中的能量物质, 例如还原糖、蔗糖和淀粉等TNC。Fan和Guo (2010)发现, 三级和四级根中的TNC浓度比一级和二级根高10%-30%, 分解两个月后, TNC浓度下降了50%-80%, 每一级根的TNC浓度基本相同。可能是因为这些容易降解的TNC已经为分解者提供了丰富的能量, 导致高级根持续更快的分解。本研究结果显示低级根[1+2]含有较低浓度的TNC, 而高级根[3+4]中含量较高(表4), 并且发现k与TNC浓度密切相关(p < 0.001, 图2)。因此, 含有较低TNC浓度的低级根[1+2]分解较慢, 相反高级根[3+4]分解较快。此外, 初始细根中N浓度和C:N均不是影响细根分解速率的因素。Xiong等(2012)研究表明, 细根的N浓度和C:N并没有影响细根分解速率, 本研究结果与之一致。他们的结果显示, AUF和N浓度呈负相关关系, 因此并不能检验AUF和N浓度是如何独立调节根系分解速率的。值得指出的是, 由于样品量较小, 本实验并未对分解过程中各时期根系内的AUF进行测定。因此细根内的AUF和TNC是否是一个恒定状态及其对根系分解速率的影响尚未可知, 有待于今后进一步深入研究。另外, 通过分析, P、K与k均不相关。气候因子也是影响根系分解的重要因素。在今后的研究中, 还应进一步进行气候因子的相关研究。
4 结论
本研究对4个典型温带树种的细根进行了研究, 通过不同根序法的实验进一步验证了低级根[1+2]分解慢, 而高级根[3+4]分解快这一普遍规律。并发现其主要原因是低级根[1+2]含有较多的AUF和较少的TNC, 即低级根[1+2]中含有较多难分解的高分子结构以及提供给分解者的能量物质较少。本实验测定的其他化学指标如N、P、K、C:N均和分解速率不相关。
致谢
感谢东北林业大学的王秀伟、程春香、任丹, 中国科学院沈阳应用生态研究所的董利利在文章撰写过程中提供的帮助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]

1 Decomposition of plant litter, a key component of the global carbon budget, is hierarchically controlled by the triad: climate > litter quality > soil organisms. Given the sensitivity of decomposition to temperature, especially in cold biomes, it has been hypothesized that global warming will lead to increased litter decomposition rates, both through direct temperature effects and through indirect effects on litter quality and soil organisms. 2 A meta-analysis of experimental warming studies in cold biomes (34 site-species combinations) showed that warming resulted in slightly increased decomposition rates. However, this response was strongly dependent on the method used: open top chambers reduced decomposition rates, whereas heating lamps stimulated decomposition rates. The low responsiveness was mainly due to moisture-limited decomposition rates in the warming treatments, especially at mesic and xeric sites. This control of litter decomposition by both temperature and moisture was corroborated by natural gradient studies. 3 Interspecific differences in litter quality and decomposability are substantially larger than warming-induced phenotypic responses. Thus, the changes in the species composition and structure of plant communities that have been observed in medium-term warming studies in cold biomes will have a considerably greater impact on ecosystem litter decomposition than phenotypic responses. 4 Soil fauna communities in cold biomes are responsive to climate warming. Moreover, temperature-driven migration of the, hitherto absent, large comminuters to high-latitude sites may significantly increase decomposition rates. However, we do not know how far-reaching the consequences of changes in the species composition and structure of the soil community are for litter decomposition, as there is a lack of data on functional species redundancy and the species' dispersal ability. 5 Global warming will lead to increased litter decomposition rates only if there is sufficient soil moisture. Hence, climate scenario and experimental studies should focus more on both factors and their interaction. As interspecific differences in potential decomposability and litter chemistry are substantially larger than phenotypic responses to warming, the focus of future research should be on the former. In addition, more light should be shed on the below-ground 'darkness' to evaluate the ecological significance of warming-induced soil fauna community changes for litter decomposition processes in cold biomes.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
Our understanding of fine-root decay processes is derived almost exclusively from litterbag studies. However, preparation of roots for litterbag studies and their subsequent decay within litterbags represent major departures from in situ conditions. We hypothesized that litterbag studies misrepresent fine-root decay and nutrient release rates during decomposition. To test these hypotheses we developed a new intact-core technique that requires no a priori root processing, retains natural rhizosphere associations, and maintains in situ decay conditions. Using both litterbags and intact cores, we measured annual decay rates and nitrogen release from newly senesced fine roots of silver maple, maize, and winter wheat. After one year, mass loss was 10-23% greater, and nitrogen release was 21-29% higher within intact cores. Differences appeared to result from litterbag-induced alterations to decomposer dynamics and from unavoidable changes to fine-root size-class composition within litterbags. Our results suggest that fine-root decay and nutrient turnover occur significantly faster than estimated from litterbag studies. By minimizing disturbances to roots, soil, and rhizosphere associates prior to root decay, the intact-core technique provides an improved alternative for measuring fine-root decomposition.
DOIURLPMID [本文引用: 5]
Among tree fine roots, the distal small-diameter lateral branches comprising first-and second-order roots lack secondary (wood) development. Therefore, these roots are expected to decompose more rapidly than higher order woody roots. But this prediction has not been tested and may not be correct. Current evidence suggests that lower order roots may decompose more slowly than higher order roots in tree species associated with ectomycorrhizal (EM) fungi because they are preferentially colonized by fungi and encased by a fungal sheath rich in chitin (a recalcitrant compound). In trees associated with arbuscular mycorrhizal (AM) fungi, lower order roots do not form fungal sheaths, but they may have poorer C quality, e. g. lower concentrations of soluble carbohydrates and higher concentrations of acid-insolubles than higher order roots, thus may decompose more slowly. In addition, litter with high concentrations of acid insolubles decomposes more slowly under higher N concentrations (such as lower order roots). Therefore, we propose that in both AM and EM trees, lower order roots decompose more slowly than higher order roots due to the combination of poor C quality and high N concentrations. To test this hypothesis, we examined decomposition of the first six root orders in Fraxinus mandshurica (an AM species) and Larix gmelinii (an EM species) using litterbag method in northeastern China. We found that lower order roots of both species decomposed more slowly than higher order roots, and this pattern appears to be associated mainly with initial C quality and N concentrations. Because these lower order roots have short life spans and thus dominate root mortality, their slow decomposition implies that a substantial fraction of the stable soil organic matter pool is derived from these lower order roots, at least in the two species we studied.
DOIURL [本文引用: 3]
Root turnover is fastest in the finest roots of the root system (first root order). Additionally, tissue chemistry varies among even the finest root orders and between white roots and older, pigmented roots. Yet the effects of pigmentation and order on root decomposition have rarely been examined. We separated the first four root orders (all <1 mm) of four temperate tree species into three classes: white first- and second-order roots; pigmented first- and second-order roots; and pigmented third- and fourth-order roots. Roots were enclosed in litterbags and buried under their own and under a common species canopy in a 34-year-old common garden in Poland. When comparing decomposition of different root orders over 36 months, pigmented third- and fourth-order roots with a higher C:N ratio decomposed more rapidly, losing 20—40% of their mass, than pigmented first- and second-order roots, which lost no more than 20%. When comparing decomposition of roots of different levels of pigmentation within the same root order over 14 months, pigmented (older) first- and second-order roots lost 6510% of their mass, while white (younger) first- and second-order roots lost 6530%. In contrast to root mass loss, root N content declined more rapidly in the first- and second-order roots than in third- and fourth-order roots. In higher-order roots, N increased in the first 10 months from 65110% to nearly 150% of initial N content, depending on species; by the end of the study N content had returned to initial levels. These findings suggest that, in plant communities where root mortality is primarily of pigmented first- and second-order roots, microbial decomposition may be slower than estimates derived from bulk fine-root litterbag experiments, which typically contain at least four root orders. Thus, a more mechanistic understanding of root decomposition and its contribution to ecosystem carbon and nutrient dynamics requires a fundamental shift in experimental methods that stratifies root samples for decomposition along more functionally based criteria such as root order and pigmentation, which parallel the markedly different longevities of these different root classes.
DOIURL [本文引用: 1]
树木细根具有高度的结构和功能的异质性。近20年来,人们逐渐认识到采用某一直径阈值来定义细根并开展研究的方法(即直径法)不能准确地描述相关的生理学和生态学过程。随着越来越多的研究证实根系功能与其分支等级密切相关,根序划分的方法(即根序法)在根系研究中得到更多的应用。但是,采用根序法测定相关功能属性需耗费大量的人力和时间。最近,有研究者在结合直径法和根序法优点的基础上提出了功能划分的方法。该方法将传统的细根区分为吸收根和运输根,在充分考虑根系结构和功能联系的同时,又能兼顾研究工作的效率和结果间的可比性,特别适用于根系生物量与周转的研究。采用功能划分方法来研究根系生物量(包括其他功能属性)是一个较新的观点,研究结果的规律性和存在的主要问题仍有待总结。该文作者通过查阅近年来的相关研究报告发现吸收根与运输根生物量在树种间存在较大的差异,目前对吸收根生物量在全球尺度上的变异格局并不清楚,吸收根与运输根对细根生物量周转的相对贡献有待探究,运输根在界定上存在着很大的挑战性。该文最后讨论了在根系研究中应用功能划分方法的优势与不足,并提出了建议。
DOIURL [本文引用: 1]
树木细根具有高度的结构和功能的异质性。近20年来,人们逐渐认识到采用某一直径阈值来定义细根并开展研究的方法(即直径法)不能准确地描述相关的生理学和生态学过程。随着越来越多的研究证实根系功能与其分支等级密切相关,根序划分的方法(即根序法)在根系研究中得到更多的应用。但是,采用根序法测定相关功能属性需耗费大量的人力和时间。最近,有研究者在结合直径法和根序法优点的基础上提出了功能划分的方法。该方法将传统的细根区分为吸收根和运输根,在充分考虑根系结构和功能联系的同时,又能兼顾研究工作的效率和结果间的可比性,特别适用于根系生物量与周转的研究。采用功能划分方法来研究根系生物量(包括其他功能属性)是一个较新的观点,研究结果的规律性和存在的主要问题仍有待总结。该文作者通过查阅近年来的相关研究报告发现吸收根与运输根生物量在树种间存在较大的差异,目前对吸收根生物量在全球尺度上的变异格局并不清楚,吸收根与运输根对细根生物量周转的相对贡献有待探究,运输根在界定上存在着很大的挑战性。该文最后讨论了在根系研究中应用功能划分方法的优势与不足,并提出了建议。
.
DOIURL [本文引用: 1]
采用4个梯度的林地氮处理(N0、N3、N6和N9依次为0、3、6和9 g N·m-2·a-1),利用分解袋试验,研究了N添加对油松不同径级细根分解及养分释放过程的影响.结果表明:细根分解过程分为快速分解(0~60 d)和慢速分解(60 ~ 300 d)两个阶段.0~0.4、0.4~1和1~2mm细根分解的质量百分数在第60天分别为7.6%、10.4%和11.4%,在第300天分别为19.8%、23.5%和30.5%,说明较细的根系分解较慢.N添加显著降低了0~0.4 mm细根的分解速率,但对0.4~1和1~~2 mm细根分解速率无显著影响,与对照(N0)相比,N3、N6和N9处理试验期间分解速率分别降低2.1%、4.5%和5.8%.N添加显著增加了0~0.4和0.4~1 mm细根C和N残留率,但对1~~2 mm细根C和N残留率无显著影响,且对3个径级细根P残留率无显著影响.与对照相比,N3、N6和N9处理分别增加了0~0.4 mm细根中8.1%、9.4%和4.5%的C残留率和5.3%、16.3%和16.7%的N残留率;同时增加了0.4~1 mm细根中2.5%、2.5%和0.9%的C残留率和0.9%、2.3%和3.9%的N残留率.0~0.4、0.4~1 mm细根C、N、P迁移模式总体表现为直接释放,而1~~2 mm细根N为富集-释放模式.氮沉降可能主要通过影响0~0.4mm细根(主要为1和2级细根)的分解过程,从而降低细根的分解速率.
DOIURL [本文引用: 1]
采用4个梯度的林地氮处理(N0、N3、N6和N9依次为0、3、6和9 g N·m-2·a-1),利用分解袋试验,研究了N添加对油松不同径级细根分解及养分释放过程的影响.结果表明:细根分解过程分为快速分解(0~60 d)和慢速分解(60 ~ 300 d)两个阶段.0~0.4、0.4~1和1~2mm细根分解的质量百分数在第60天分别为7.6%、10.4%和11.4%,在第300天分别为19.8%、23.5%和30.5%,说明较细的根系分解较慢.N添加显著降低了0~0.4 mm细根的分解速率,但对0.4~1和1~~2 mm细根分解速率无显著影响,与对照(N0)相比,N3、N6和N9处理试验期间分解速率分别降低2.1%、4.5%和5.8%.N添加显著增加了0~0.4和0.4~1 mm细根C和N残留率,但对1~~2 mm细根C和N残留率无显著影响,且对3个径级细根P残留率无显著影响.与对照相比,N3、N6和N9处理分别增加了0~0.4 mm细根中8.1%、9.4%和4.5%的C残留率和5.3%、16.3%和16.7%的N残留率;同时增加了0.4~1 mm细根中2.5%、2.5%和0.9%的C残留率和0.9%、2.3%和3.9%的N残留率.0~0.4、0.4~1 mm细根C、N、P迁移模式总体表现为直接释放,而1~~2 mm细根N为富集-释放模式.氮沉降可能主要通过影响0~0.4mm细根(主要为1和2级细根)的分解过程,从而降低细根的分解速率.
DOIURL [本文引用: 1]
1. Root life span regulates the quantity and quality of root-derived organic matter transferred to the soil organic matter pool. However, poor understanding of the rates and controls of root life span has hindered the prediction of carbon (C) flow and nutrient cycling dynamics at the ecosystem scale. 2. We examined the effects of root branch order, root diameter, mycorrhizal colonization, season of birth, depth in the soil, nitrogen (N) fertilization and foliage removal on root life span in a longleaf pine (Pinus palustris Mill.) forest from 2001 to 2004 using minirhizotron and soil monolith sampling. 3. Among all factors, root branch order had the strongest and most consistent effect on life span, with higher order roots having a 46% longer life span than roots one order lower. 4. Within first order roots, mycorrhizal colonization significantly increased root life span by > 45% in 2 of 3 years. 5. Roots born in winter and spring generally lived longer than roots born in summer and autumn. Root life span was positively correlated with depth in the soil and root diameter, but the correlations were weaker than with order, year and season. Neither N fertilization nor foliage removal had a significant impact on root life span. 6. When biomass mortality and associated N flux were estimated based on order-specific mean life span, N concentration and ecosystem-scale biomass estimates, first order roots constituted approximately 50% of the total biomass mortality and > 60% of the N flux for the first three root orders combined. 7. Synthesis. Our results show that (i) root branch order was the strongest predictor of life span among all covariates and can effectively partition the distal longleaf pine root systems into three or more populations with different turnover rates; (ii) only a fraction of fine roots turns over annually, whereas models of C cycles assume an annual turnover for the entire fine root system. We conclude that an order-based approach holds greater promise than the traditional diameter class approach for evaluating the role of different fine root populations in C flow and nutrient cycling.
DOIURLPMID [本文引用: 3]
090004 Different portions of tree root systems play distinct functional roles, yet precisely how to distinguish roots of different functions within the branching fine-root system is unclear. 090004 Here, anatomy and mycorrhizal colonization was examined by branch order in 23 Chinese temperate tree species of both angiosperms and gymnosperms forming ectomycorrhizal and arbuscular090009mycorrhizal associations. 090004 Different branch orders showed marked differences in anatomy. First-order roots exhibited primary development with an intact cortex, a high mycorrhizal colonization rate and a low stele proportion, thus serving absorptive functions. Second and third orders had both primary and secondary development. Fourth and higher orders showed mostly secondary development with no cortex or mycorrhizal colonization, and thus have limited role in absorption. Based on anatomical traits, it was estimated that c . 75% of the fine-root length was absorptive, and 68% was mycorrhizal, averaged across species. 090004 These results showed that: order predicted differences in root anatomy in a relatively consistent manner across species; anatomical traits associated with absorption and mycorrhizal colonization occurred mainly in the first three orders; the single diameter class approach may have overestimated absorptive root length by 25% in temperate forests.
DOIURLPMID [本文引用: 2]
Fine roots are a key component of carbon (C) flow and nitrogen (N) cycling in forest ecosystems. However, the complexity and heterogeneity of the fine root branching system have hampered the assessment and prediction of C and N dynamics at ecosystem scales. We examined how root morphology, biomass, and chemistry differed with root branch orders (1-5 with root tips classified as first order roots) and how different root orders responded to increased C sink strength (via N fertilization) and reduced carbon source strength (via canopy scorching) in a longleaf pine (Pinus palustris L.) ecosystem. With increasing root order, the diameter and length of individual roots increased, whereas the specific root length decreased. Total root biomass on an areal basis was similar among the first four orders but increased for the fifth order roots. Consequently, total root length and total root surface area decreased systematically with increasing root order. Fine root N and lignin concentrations decreased, while total non-structural carbohydrate (TNC) and cellulose concentrations increased with increasing root order. N addition and canopy disturbance did not alter root morphology, but they did influence root chemistry. N fertilization increased fine root N concentration and content per unit area in all five orders, while canopy scorching decreased root N concentration. Moreover, TNC concentration and content in fifth order roots were also reduced by canopy scorching. Our results indicate that the small, fragile, and more easily overlooked first and second order root may be disproportinately important in ecosystem scale C and N fluxes due to their large proportions of fine root biomass, high N concentrations, relatively short lifespans, and podentially high decomposition rates.
DOIURL [本文引用: 1]
Foliar litter position and decomposition were assessed in longleaf pine (Mill.) - wiregrass (Trin. & Rupr.) woodlands during a 3-year burn interval. Position assessments revealed 57.7 and 67.4% of foliar litter was elevated in wiregrass crowns 1 and 2 years, respectively, following burning. Decomposition assessments revealed soil-surface mass loss decay constants (range 0.097–0.282) similar to those measured in comparable pine forests. However, elevated longleaf pine and wiregrass litter exhibited decay constants (0.052 and 0.074, respectively) 50% lower than corresponding soil-surface rates and among the lowest values in the literature. With the exception of wiregrass, which did not exhibit an immobilization of the nutrients (N, P, Ca, K, and Mg) assessed, foliar litter exhibited either extensive P immobilization with minimal N immobilization or minimal, short-lived immobilization of N, P, or both N and P. The percentage of original N and P remaining after 3 years varied widely among the soil surface (N range 6.3–56.3%; P range 3.4–204.7%) and elevated (N range 76.8–94.9%; P range 52.0–99.2%) litter. These results suggest that fire regimes typically employed in longleaf pine – wiregrass woodlands may balance N losses via volatilization with P limitations via litter immobilization.La position et la décomposition de la litière de feuilles ont été évaluées dans des forêts de pin des marais (Mill.) et d'aristide bleutée (Trin. & Rupr.) au cours d'un intervalle de 3 ans entre deux feux. L'étude de la position révèle que respectivement 57,7 et 67,4% de la litière foliaire se retrouve dans la cime de l'aristide bleutée 1 et 2 ans après un feu. L'étude de la décomposition révèle que les constantes de décomposition qui traduisent la perte de masse à la surface du sol (0,097–0,282) sont semblables à celles qui ont été mesurées dans des forêts comparables de pin. Cependant, la litière surélevée de pin des marais et d'aristide bleutée ont des constantes de décomposition (respectivement 0,052 et 0,074) 50% plus faibles que les taux correspondants à la surface du sol et parmi les plus faibles rapportées dans la littérature. 08 l'exception de l'aristide bleutée qui n'immobilise pas les nutriments (N, P, Ca, K et Mg) mesurés, la litière de feuille immobilise soit beaucoup de P et peu de N, soit peu de N, de P ou de N et de P pendant de courtes périodes. Après 3 ans, les pourcentages initiaux de N et P encore présents varient beaucoup selon que la litière se trouve à la surface du sol (N varie de 6,3–56,3%; P varie de 3,4–204,7%) ou au-dessus du sol (N varie de 76,8–94,9%; P varie de 52,0–99,2%). Ces résultats indiquent que le régime de feux normalement utilisé dans les forêts de pin des marais et d'aristide bleutée peut contrebalancer les pertes de N par volatilisation mais peut causer des carences en P à cause de l'immobilisation dans la litière.[Traduit par la Rédaction]
DOIURLPMID [本文引用: 1]
Elucidating the function of and patterns among plant traits above ground has been a major research focus, while the patterns and functioning of belowground traits remain less well understood. Even less well known is whether species differences in leaf traits and their associated biogeochemical effects are mirrored by differences in root traits and their effects. We studied fine root decomposition and N dynamics in a common garden study of 11 temperate European and North American tree species (Abies alba, Acer platanoides, Acer pseudoplatanus, Carpinus betulus, Fagus sylvatica, Larix decidua, Picea abies, Pseudotsuga menziesii, Quercus robur, Quercus rubra and Tilia cordata) to determine whether leaf litter and fine root decomposition rates are correlated across species as well as which species traits influence microbial decomposition above versus below ground. Decomposition and N immobilization rates of fine roots were unrelated to those of leaf litter across species. The lack of correspondence of above-and belowground processes arose partly because the tissue traits that influenced decomposition and detritus N dynamics different for roots versus leaves, and partly because influential traits were unrelated between roots and leaves across species. For example, while high hemicellulose concentrations and thinner roots were associated with more rapid decomposition below ground, low lignin and high Ca concentrations were associated with rapid aboveground leaf decomposition. Our study suggests that among these temperate trees, species effects on and N dynamics in decomposing fine roots and leaf litter may not reinforce each other. Thus, species differences in rates of microbially mediated decomposition may not be as large as they would be if above-and belowground processes were working in similar directions (i.e., if faster decomposition above ground corresponded to faster decomposition below ground). Our results imply that studies that focus solely on aboveground traits may obscure some of the important mechanisms by which plant species influence ecosystem processes.
DOIURLPMID [本文引用: 1]
We studied the effects of tree species on leaf litter decomposition and forest floor dynamics in a common garden experiment of 14 tree species ( Abies alba, Acer platanoides, Acer pseudoplatanus, Betula pendula, Carpinus betulus, Fagus sylvatica, Larix decidua, Picea abies, Pinus nigra, Pinus sylvestris, Pseudotsuga menziesii, Quercus robur, Quercus rubra, and Tilia cordata) in southwestern Poland. We used three simultaneous litter bag experiments to tease apart species effects on decomposition via leaf litter chemistry vs. effects on the decomposition environment. Decomposition rates of litter in its plot of origin were negatively correlated with litter lignin and positively correlated with mean annual soil temperature (MAT(soil)) across species. Likewise, decomposition of a common litter type across all plots was positively associated with MATsoil, and decomposition of litter from all plots in a common plot was negatively related to litter lignin but positively related to litter Ca. Taken together, these results indicate that tree species influenced microbial decomposition primarily via differences in litter lignin ( and secondarily, via differences in litter Ca), with high-lignin ( and low-Ca) species decomposing most slowly, and by affecting MATsoil, with warmer plots exhibiting more rapid decomposition. In addition to litter bag experiments, we examined forest floor dynamics in each plot by mass balance, since earthworms were a known component of these forest stands and their access to litter in litter bags was limited. Forest floor removal rates estimated from mass balance were positively related to leaf litter Ca ( and unrelated to decay rates obtained using litter bags). Litter Ca, in turn, was positively related to the abundance of earthworms, particularly Lumbricus terrestris. Thus, while species influence microbially mediated decomposition primarily through differences in litter lignin, differences among species in litter Ca are most important in determining species effects on forest floor leaf litter dynamics among these 14 tree species, apparently because of the influence of litter Ca on earthworm activity. The overall influence of these tree species on leaf litter decomposition via effects on both microbial and faunal processing will only become clear when we can quantify the decay dynamics of litter that is translocated belowground by earthworms.
DOIURL [本文引用: 1]
高山森林冬季不同厚度雪被格局可能通过影响凋落物的分解过程中酸溶性和酸不溶性组分特征,改变凋落物分解过程,但缺乏必要关注。采用凋落物分解袋法,研究了高山森林林窗中央至林下形成的天然雪被厚度梯度(厚型雪被、中型雪被、薄型雪被和无雪被)覆盖下,6种典型物种岷江冷杉(Abies faxoniana)、红桦(Betula albo-sinensis)、四川红杉(Larix mastersiana)、方枝柏(Sabina sahuaria)、康定柳(Salixpara plesia)和高山杜鹃(Rhododendron lapponicum)凋落物在不同关键时期(雪被形成期、雪被覆盖期和雪被融化期)的酸溶性组分和酸不溶性组分变化特征。经历一个冬季的分解后,6种凋落物酸溶性组分绝对含量呈降低趋势,除红桦外5种凋落物酸不溶性组分绝对含量呈增加趋势。不同厚度雪被显著影响雪被覆盖期和融化期凋落物酸不溶性和酸溶性组分绝对变化量;其中方枝柏、红桦和康定柳凋落物酸不溶性组分增加量在厚型雪被下显著高于其它雪被覆盖;而相对于阔叶凋落物酸溶性组分变化量在薄型雪被和无雪被梯度达到最大值,针叶凋落物酸溶性组分在厚型雪被下具有最大的变化量。一个冬季分解结束后,表征6种凋落物酸溶性和酸不溶性组分含量相对比例的LCI指数(Lignocellulose index)总体升高,雪被对LCI指数的影响主要表现在雪被覆盖期和融化期,且方枝柏、岷江冷杉和康定柳凋落物LCI在冬季分解后均在厚型雪被达到最高值。同时统计分析结果表明,物种极显著影响冬季不同阶段凋落物酸溶性和酸不溶性组分的变化。这些结果意味着气候变暖情景下,高山森林冬季雪被和冻融格局的改变将显著影响凋落物分解过程中酸溶性、酸不溶性组分以及LCI指数代表的抵抗性组分结构的变化,且影响趋势受到凋落物质量的调控。
DOIURL [本文引用: 1]
高山森林冬季不同厚度雪被格局可能通过影响凋落物的分解过程中酸溶性和酸不溶性组分特征,改变凋落物分解过程,但缺乏必要关注。采用凋落物分解袋法,研究了高山森林林窗中央至林下形成的天然雪被厚度梯度(厚型雪被、中型雪被、薄型雪被和无雪被)覆盖下,6种典型物种岷江冷杉(Abies faxoniana)、红桦(Betula albo-sinensis)、四川红杉(Larix mastersiana)、方枝柏(Sabina sahuaria)、康定柳(Salixpara plesia)和高山杜鹃(Rhododendron lapponicum)凋落物在不同关键时期(雪被形成期、雪被覆盖期和雪被融化期)的酸溶性组分和酸不溶性组分变化特征。经历一个冬季的分解后,6种凋落物酸溶性组分绝对含量呈降低趋势,除红桦外5种凋落物酸不溶性组分绝对含量呈增加趋势。不同厚度雪被显著影响雪被覆盖期和融化期凋落物酸不溶性和酸溶性组分绝对变化量;其中方枝柏、红桦和康定柳凋落物酸不溶性组分增加量在厚型雪被下显著高于其它雪被覆盖;而相对于阔叶凋落物酸溶性组分变化量在薄型雪被和无雪被梯度达到最大值,针叶凋落物酸溶性组分在厚型雪被下具有最大的变化量。一个冬季分解结束后,表征6种凋落物酸溶性和酸不溶性组分含量相对比例的LCI指数(Lignocellulose index)总体升高,雪被对LCI指数的影响主要表现在雪被覆盖期和融化期,且方枝柏、岷江冷杉和康定柳凋落物LCI在冬季分解后均在厚型雪被达到最高值。同时统计分析结果表明,物种极显著影响冬季不同阶段凋落物酸溶性和酸不溶性组分的变化。这些结果意味着气候变暖情景下,高山森林冬季雪被和冻融格局的改变将显著影响凋落物分解过程中酸溶性、酸不溶性组分以及LCI指数代表的抵抗性组分结构的变化,且影响趋势受到凋落物质量的调控。
DOIURL [本文引用: 1]
植物根序和径级不仅反映细根的形态结构,而且能反映根系的一些生理特征,如细根寿命和周转等。该文以二年生油松(Pinus tabulaeformis)幼苗根系为研究对象,系统比较了根序分类方法和径级分类方法在描述根系特征上的优缺点,探索了两者之间的内在联系。结果表明:二年生油松幼苗最多可包括6级根序,直径的变化范围为0.169–3.877mm。按根序划分,I–VI级根序的总根长和总根表面积主要集中在前3级根序,这3级根序的根占总根长的78.77%和总根表面积的62.72%。前3级根序的比根长是后3级根序比根长的1.3–3.0倍,比根面积是后3级比根面积的1.0–1.5倍。按常用的径级(以0.5、1.0、1.5和2.0mm为阈值)划分方法,油松幼苗大部分根系直径≤1.5mm,此区间细根的根长和根表面积占总根长的93.76%和总根表面积的84.35%。直径≤1.5mm的细根平均比根长是〉1.5mm细根比根长的3–7倍,比根面积的1.5–3.0倍。由于油松根序和径级之间有显著的指数关系,依据径级最大程度反映根序的原则,提出了新的径级划分方法,即以0.4、0.8、1.3和2.0mm为阈值对油松幼苗根系径级重新进行划分。此时,上述区间可分别包括I级、II级、III级、IV级、V级根序中根尖数的93.22%、86.37%、75.96%、70.47%和76.67%。同时也可分别涵盖各径级根长的89.34%–70.83%、根面积的86.01%–76.12%以及体积的87.73%–76.12%。此时,根系不同径级与根序之间可以建立起良好的对应关系。这些结果表明,通过合理划分径级区间可以较好地反映根序特征。
DOIURL [本文引用: 1]
植物根序和径级不仅反映细根的形态结构,而且能反映根系的一些生理特征,如细根寿命和周转等。该文以二年生油松(Pinus tabulaeformis)幼苗根系为研究对象,系统比较了根序分类方法和径级分类方法在描述根系特征上的优缺点,探索了两者之间的内在联系。结果表明:二年生油松幼苗最多可包括6级根序,直径的变化范围为0.169–3.877mm。按根序划分,I–VI级根序的总根长和总根表面积主要集中在前3级根序,这3级根序的根占总根长的78.77%和总根表面积的62.72%。前3级根序的比根长是后3级根序比根长的1.3–3.0倍,比根面积是后3级比根面积的1.0–1.5倍。按常用的径级(以0.5、1.0、1.5和2.0mm为阈值)划分方法,油松幼苗大部分根系直径≤1.5mm,此区间细根的根长和根表面积占总根长的93.76%和总根表面积的84.35%。直径≤1.5mm的细根平均比根长是〉1.5mm细根比根长的3–7倍,比根面积的1.5–3.0倍。由于油松根序和径级之间有显著的指数关系,依据径级最大程度反映根序的原则,提出了新的径级划分方法,即以0.4、0.8、1.3和2.0mm为阈值对油松幼苗根系径级重新进行划分。此时,上述区间可分别包括I级、II级、III级、IV级、V级根序中根尖数的93.22%、86.37%、75.96%、70.47%和76.67%。同时也可分别涵盖各径级根长的89.34%–70.83%、根面积的86.01%–76.12%以及体积的87.73%–76.12%。此时,根系不同径级与根序之间可以建立起良好的对应关系。这些结果表明,通过合理划分径级区间可以较好地反映根序特征。
DOIURL [本文引用: 1]
Decomposition rates and N dynamics of foliar litter from 4 tree species were measured over a 72 month period on the Chronic Nitrogen Addition plots at the Harvard Forest, Petersham MA, beginning in November 1988. Plots received nitrogen additions of 0, 5 and 15 g N m -2 yr -1 in two different stand types: red pine and mixed hardwood. Bags were collected in August and November of each year and litter analysed for mass remaining, nitrogen, cellulose and lignin content. Mass remaining was significantly greater for litter in nitrogen treated plots than in control plots after 48 months. Lignin content of litter was significantly higher with nitrogen treatments but there was little effect of treatment on cellulose content. N concentration was similar between treatments, but greater mass remaining in treated plots resulted in a higher total amount of N in humus produced in the high N plot. This mechanism could be a sink for up to 1.5 g N m -2 yr -1 of the 1.5 g N m -2 yr -1 added annually to the high N plots. Reduced decomposition rates in conjunction with increased lignin accumulation could impact global carbon sequestration as well.
DOIURLPMID [本文引用: 3]
Abstract Fine roots acquire essential soil resources and mediate biogeochemical cycling in terrestrial ecosystems. Estimates of carbon and nutrient allocation to build and maintain these structures remain uncertain because of the challenges of consistently measuring and interpreting fine-root systems. Traditionally, fine roots have been defined as all roots 0909¤ 2 mm in diameter, yet it is now recognized that this approach fails to capture the diversity of form and function observed among fine-root orders. Here, we demonstrate how order-based and functional classification frameworks improve our understanding of dynamic root processes in ecosystems dominated by perennial plants. In these frameworks, fine roots are either separated into individual root orders or functionally defined into a shorter-lived absorptive pool and a longer-lived transport fine-root pool. Using these frameworks, we estimate that fine-root production and turnover represent 22% of terrestrial net primary production globally - a c. 30% reduction from previous estimates assuming a single fine-root pool. Future work developing tools to rapidly differentiate functional fine-root classes, explicit incorporation of mycorrhizal fungi into fine-root studies, and wider adoption of a two-pool approach to model fine roots provide opportunities to better understand below-ground processes in the terrestrial biosphere. 0008 2015 The Authors. New Phytologist 0008 2015 New Phytologist Trust.
DOIURL [本文引用: 4]
The fine roots of trees are concentrated on lateral branches that arise from perennial roots. They are important in the acquisition of water and essential nutrients, and at the ecosystem level, they make a significant contribution to biogeochemical cycling. Fine roots have often been studied according to arbitrary size classes, e.g., all roots less than 1 or 2 mm in diameter. Because of the size class approach, the position of an individual root on the complex lateral branching system has often been ignored, and relationships between the form of the branching root system and its function are poorly understood. The fine roots of both gymnosperms and angiosperms, which formed ectomycorrhizae (EM) and arbuscular mycorrhizae (AM) fungal associations, were sampled in 1998 and 1999. Study sites were chosen to encompass a wide variety of environments in four regions of North America. Intact lateral branches were collected from each species and 18561 individual roots were dissected by order, with distal roots numbered as first-order roots. This scheme is similar to the one commonly used to number the order of streams. Fine root diameter, length, specific root length (SRL; m/g), and nitrogen (N) concentration of nine North American tree species (Acer saccharum, Juniperus monosperma, Liriodendron tulipifera, Picea glauca, Pinus edulis, Pinus elliottii, Pinus resinosa, Populus balsamifera, and Quercus alba) were then compared and contrasted. Lateral roots <0.5 mm in diameter accounted for >75% of the total number and length of individual roots sampled in all species except Liriodendron tulipifera. Both SRL and N concentration decreased with increasing root order in all nine species, and this pattern appears to be universal in all temperate and boreal trees. Nitrogen concentrations ranged from 8.5 to 30.9 g/kg and were highest in the first-order "root tips." On a mass basis, first-order roots are expensive to maintain per unit time (high tissue N concentration). Tissue N appears to be a key factor in understanding the C cost of maintaining first- and second-order roots, which dominate the display of absorbing root length. There were many significant differences among species in diameter, length, SRL, and N concentration. For example, two different species can have similar SRL but very different tissue N concentrations. Our findings run contrary to the common idea that all roots of a given size class function the same way and that a common size class for fine roots works well for all species. Interestingly, fine root lateral branches are apparently deciduous, with a distinct lateral branch scar. The position of an individual root on the branching root system appears to be important in understanding the function of fine roots.
DOIURLPMID [本文引用: 1]
The colorimetric method for the estimation of carbohydrates was modified. Procedures are de scribed for use with tissues of high or low glycogen content; for the former, aliquots of an alkaline digest of the tissue are used and, for the latter, glycogen which has first been separated from an alkaline digest. The results obtained by both procedures were in good agreement with those given by Good...
DOIURLPMID [本文引用: 1]
Root decomposition represents a significant C flux in terrestrial ecosystems. Roots are exposed to a different decomposition environment than aboveground tissues, and few general principles exist regarding the factors controlling rates of root decay. We use a global dataset to explore the relative importance of climate, environmental variables, and litter quality in regulating rates of root decomposition. The parameters that explained the largest amount of variability in root decay were root Ca concentrations and C:N ratios, with a smaller proportion explained by latitude, mean annual temperature, mean annual precipitation, and actual evapotranspiration (AET). Root chemistry and decay rates varied by plant life form (conifer, broadleaf, or graminoid). Conifer roots had the lowest levels of Ca and N, the highest C:N and lignin:N ratios, and decomposed at the slowest rates. In a stepwise multiple linear regression, AET, root Ca, and C:N ratio accounted for approximately 90% of the variability in root decay rates. Root chemistry appeared to be the primary controller of root decomposition, while climate and environmental factors played secondary roles, in contrast to previously established leaf litter decomposition models.
DOIURL [本文引用: 1]
细根分解是陆地生态系统C和养分循环的重要环节.以往的细根分解研究以埋袋法的应用为主.然而,由于埋袋 法对分解材料的干扰以及对分解环境的改变使其很难揭示原位环境下根系的自然分解过程.该研究应用微根管(Minirhizotron)技术连续3年对水曲 柳(Fraxinus mandshurica)和兴安落叶松(Larix gmelinii)细根的分解过程进行原位监测,运用Kaplan-Meier方法估算细根分解的保存率及分解期中位值(即50%细根完全分解的时 间,Median root decomposition time),做分解曲线,用对数秩检验(Log-rank test)方法分析不同树种、直径、根序及土层对细根保存率的影响.结果表明,伴随时间延长,细根的保存率逐渐下降,兴安落叶松细根保存率的下降显著快于 水曲柳(p0.6 mm)细根的分解速率不同,两树种最长分解期中位值均出现在最细直径(≤0.3 mm)根中;高级根分解速率显著低于一级根(p<0.05);土壤上层分解速度快,随着土壤深度增加细根分解速率减小.微根管技术为了解细根自然分 解过程提供了有效途径.
DOIURL [本文引用: 1]
细根分解是陆地生态系统C和养分循环的重要环节.以往的细根分解研究以埋袋法的应用为主.然而,由于埋袋 法对分解材料的干扰以及对分解环境的改变使其很难揭示原位环境下根系的自然分解过程.该研究应用微根管(Minirhizotron)技术连续3年对水曲 柳(Fraxinus mandshurica)和兴安落叶松(Larix gmelinii)细根的分解过程进行原位监测,运用Kaplan-Meier方法估算细根分解的保存率及分解期中位值(即50%细根完全分解的时 间,Median root decomposition time),做分解曲线,用对数秩检验(Log-rank test)方法分析不同树种、直径、根序及土层对细根保存率的影响.结果表明,伴随时间延长,细根的保存率逐渐下降,兴安落叶松细根保存率的下降显著快于 水曲柳(p0.6 mm)细根的分解速率不同,两树种最长分解期中位值均出现在最细直径(≤0.3 mm)根中;高级根分解速率显著低于一级根(p<0.05);土壤上层分解速度快,随着土壤深度增加细根分解速率减小.微根管技术为了解细根自然分 解过程提供了有效途径.
DOIURL [本文引用: 2]
ABSTRACT Roots concentrated on the smallest distal branching orders have short life spans and thus dominate root mortality, and may contribute predominately to plant carbon and nutrient transfer into soil. Yet the effects of nitrogen (N) enrichment on decomposition of the finest root branching orders have not yet been examined. Resolving such N effects is critical for predicting the ecosystem consequences of increased anthropogenic N deposition. The first four root orders were separated into two classes: first- and second-order roots; third- and fourth-order roots. We studied the effects of N addition on decomposition of different root order classes in four temperate tree species over 4 years. Asymptotic decay models best fit the decomposition and allowed us to examine effects of N on initial versus later stages of decomposition separately. Very early in decomposition, N fertilization stimulated decomposition rates in higher-order roots, but had no effects on initial rates of decomposition in lower-order roots. In contrast, later in decomposition, N fertilization inhibited decomposition, ultimately resulting in a larger, slowly decomposing fraction in both lower-order and higher-order roots. Inhibitory effects of N addition on lignin-degrading enzyme activity might be an important mechanism explaining the negative effects of N on decomposition here. This study highlights the importance of long-term studies for understanding N effects on decomposition, and suggests that contrasting effects of N on different decomposition processes and carbon pools should be widely considered in biogeochemical models. Furthermore, the inhibitory effects of elevated atmospheric N deposition on decomposition of lower-order roots suggest that these roots may provide a critical mechanism of carbon and nutrient retention in soil because of their rapid input via root mortality.
[本文引用: 4]
DOIURL [本文引用: 1]
Root decomposition studies have rarely considered the heterogeneity within a fine-root system. Here, we investigated fine root (Both litterbag and intact-core techniques were used to examine decomposition dynamic and nutrient release of the two size class roots over a 498-day period. Moreover, we examined differences between the two approaches.The very fine roots (Our results suggest that the slow decomposition rate of very fine roots (< 0.5 mm) may be determined by their high concentration of acid-unhydrolyzable structural components. Additionally, the heterogeneity within a bulk fine-root system could lead to differences in their contribution to soil in terms of carbon and nitrogen dynamics.
DOIURL [本文引用: 1]
Despite our growing appreciation of the large contribution made by fine roots to soil organic carbon pool and nutrient cycling, their decomposition dynamics still remain poorly quantified.In a field experiment, we conducted a 4-year litterbag study on different root size classes (Very fine roots (We conclude that the quality of carbon in root detritus and not the concentration of initial nutrients control its decomposition dynamics in a relatively nutrient-rich soil in northeastern China. These patterns can be explained by lower easily accessible labile carbon and higher recalcitrant carbon fractions in very fine roots (< 0.5 mm) than in 0.5-2.0 mm roots, at least in the four species we studied.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURLPMID [本文引用: 1]
Most studies of forest litter dynamics examine the biochemical characteristics and decomposition of leaf litter, but fine roots are also a large source of litter in forests.We quantified the concentrations of eight biochemical fractions and nitrogen (N) in leaf litter and fine roots at four sugar maple (Acer saccharum)‐dominated hardwood forests in the north‐central United States. We combined these results with litter production data to estimate ecosystem biochemical fluxes to soil. We also compared how leaf litter and fine root biochemistry responded to long‐term simulated N deposition.Compared with leaf litter, fine roots contained 2.9‐fold higher acid‐insoluble fraction (AIF) and 2.3‐fold more condensed tannins; both are relatively difficult to decompose. Comparatively, leaf litter had greater quantities of more labile components: nonstructural carbohydrates, cellulose and soluble phenolics. At an ecosystem scale, fine roots contributed over two‐thirds of the fluxes ofAIFand condensed tannins to soil. Fine root biochemistry was also less responsive than leaf litter to long‐term simulated N deposition.Fine roots were the dominant source of difficult‐to‐decompose plant carbon fractions entering the soil at our four study sites. Based on our synthesis of the literature, this pattern appears to be widespread in boreal and temperate forests. Most studies of forest litter dynamics examine the biochemical characteristics and decomposition of leaf litter, but fine roots are also a large source of litter in forests. We quantified the concentrations of eight biochemical fractions and nitrogen (N) in leaf litter and fine roots at four sugar maple (Acer saccharum)‐dominated hardwood forests in the north‐central United States. We combined these results with litter production data to estimate ecosystem biochemical fluxes to soil. We also compared how leaf litter and fine root biochemistry responded to long‐term simulated N deposition. Compared with leaf litter, fine roots contained 2.9‐fold higher acid‐insoluble fraction (AIF) and 2.3‐fold more condensed tannins; both are relatively difficult to decompose. Comparatively, leaf litter had greater quantities of more labile components: nonstructural carbohydrates, cellulose and soluble phenolics. At an ecosystem scale, fine roots contributed over two‐thirds of the fluxes ofAIFand condensed tannins to soil. Fine root biochemistry was also less responsive than leaf litter to long‐term simulated N deposition. Fine roots were the dominant source of difficult‐to‐decompose plant carbon fractions entering the soil at our four study sites. Based on our synthesis of the literature, this pattern appears to be widespread in boreal and temperate forests.
[本文引用: 4]
URL [本文引用: 1]
采用埋袋法对水曲柳和落叶松粗根(5~10 mm)、中根(2~5 mm)、细根(<2 mm)的分解速率及其养分释放进行了为期2年的研究.结果表明,水曲柳粗根、中根和细根年分解系数分别为0.3649、0.4381和0.2720,落叶 松依次分别为0.1967、0.1955和0.2464.通过养分分析发现,根系分解过程释放大量C和养分.分解150 d后,两树种所有级别根系的可溶性糖释放均超过90%.水曲柳粗根和中根K的释放均在40%左右,细根K的释放为71%,落叶松所有级别的根系K的释放均 在95%以上.在根系分解第2年,两树种粗根和中根N的释放在50%左右波动,P在40%左右波动,两树种细根N和P的释放均达到60%.因此,根系分解 在C和养分循环中起重要作用,如果将其忽略,土壤有机质和养分元素的循环将会被严重低估.
URL [本文引用: 1]
采用埋袋法对水曲柳和落叶松粗根(5~10 mm)、中根(2~5 mm)、细根(<2 mm)的分解速率及其养分释放进行了为期2年的研究.结果表明,水曲柳粗根、中根和细根年分解系数分别为0.3649、0.4381和0.2720,落叶 松依次分别为0.1967、0.1955和0.2464.通过养分分析发现,根系分解过程释放大量C和养分.分解150 d后,两树种所有级别根系的可溶性糖释放均超过90%.水曲柳粗根和中根K的释放均在40%左右,细根K的释放为71%,落叶松所有级别的根系K的释放均 在95%以上.在根系分解第2年,两树种粗根和中根N的释放在50%左右波动,P在40%左右波动,两树种细根N和P的释放均达到60%.因此,根系分解 在C和养分循环中起重要作用,如果将其忽略,土壤有机质和养分元素的循环将会被严重低估.
The freezer defrosting: Global warming and litter decomposition rates in cold biomes
1
2006
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
Decomposition and nitrogen release from decomposition woody roots in coniferous forests of the Pacific Northwest
1
2001
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
Plant species traits are the predominant control on litter decomposition rates within biomes worldwide
1
2008
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
Quantifying fine root decomposition: An alternative to buried litterbags
1
2002
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
Slow decomposition of lower order roots: A key mechanism of root carbon and nutrient retention in the soil
5
2010
... 现有文献资料表明, 迄今为止仅有少量已发表的文章量化了根序和分解速率之间的关系.有研究发现, 尽管低级根具有较小的直径, 高级根具有较大的直径并且木质化程度较高, 但低级根仍然比高级根分解速率低(
... ;
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
... ).其中
... 另外, 根系分解受到微生物调控的影响(
Decomposition of the finest root branching orders linking belowground dynamics to fine-root function and structure
3
2011
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
... 现有文献资料表明, 迄今为止仅有少量已发表的文章量化了根序和分解速率之间的关系.有研究发现, 尽管低级根具有较小的直径, 高级根具有较大的直径并且木质化程度较高, 但低级根仍然比高级根分解速率低(
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
功能划分方法在树木细根生物量研究中的应用: 进展与评述
1
2016
... 近半个世纪以来, 大部分研究都把细根定义为一个均质系统, 即认为≤2 mm的根在结构和功能上基本相同.然而, 近年来越来越多的证据表明, 以这种单一径级定义细根的方法忽略了细根形态、化学以及生理方面的差异(
功能划分方法在树木细根生物量研究中的应用: 进展与评述
1
2016
... 近半个世纪以来, 大部分研究都把细根定义为一个均质系统, 即认为≤2 mm的根在结构和功能上基本相同.然而, 近年来越来越多的证据表明, 以这种单一径级定义细根的方法忽略了细根形态、化学以及生理方面的差异(
氮添加对油松不同径级细根分解及其养分释放的影响
1
2017
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
氮添加对油松不同径级细根分解及其养分释放的影响
1
2017
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
Endogenous and exogenous controls of root life span, mortality and nitrogen flux in a longleaf pine forest: Root branch order predominates
1
2008a
... 近半个世纪以来, 大部分研究都把细根定义为一个均质系统, 即认为≤2 mm的根在结构和功能上基本相同.然而, 近年来越来越多的证据表明, 以这种单一径级定义细根的方法忽略了细根形态、化学以及生理方面的差异(
Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species
3
2008b
... 近半个世纪以来, 大部分研究都把细根定义为一个均质系统, 即认为≤2 mm的根在结构和功能上基本相同.然而, 近年来越来越多的证据表明, 以这种单一径级定义细根的方法忽略了细根形态、化学以及生理方面的差异(
... ;
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest
2
2004
... 现有文献资料表明, 迄今为止仅有少量已发表的文章量化了根序和分解速率之间的关系.有研究发现, 尽管低级根具有较小的直径, 高级根具有较大的直径并且木质化程度较高, 但低级根仍然比高级根分解速率低(
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
Foliar litter position and decomposition in a fire-maintained longleaf pine-wiregrass ecosystem
1
2002
... 在根系分解实验开始的时候, 将剩余的烘干初始样品磨碎并测定其初始化学物质.利用浓硫酸-重铬酸钾氧化法测定全C含量; 利用半微量凯氏定氮法测定全N含量; 用钼锑抗比色法测定全P含量; 采用火焰光度计法测定全K含量; 用有机质近似C组分分析法(
Fine root decomposition rates do not mirror those of leaf litter among temperate tree species
1
2010
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
Tree species on decomposition and forest floor dynamics in a common garden
1
2006
... 将所有细根样品置于65 ℃恒温烘箱中烘干至恒质量.称取每个根序等级的样品大约260 mg置于凋落物分解袋内(尼龙网, 10 cm × 10 cm, 网眼大小120 μm).每个根序等级的细根装15袋, 2个等级(高级根和低级根)共装30袋, 4个树种总共120袋.在许多野外的凋落物分解实验中, 分解袋的网眼大小非常重要(
不同厚度雪被对高山森林6种凋落物分解过程中酸溶性和酸不溶性组分的影响
1
2015
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
不同厚度雪被对高山森林6种凋落物分解过程中酸溶性和酸不溶性组分的影响
1
2015
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
不同分类系统下油松幼苗根系特征的差异与联系
1
2010
... 近半个世纪以来, 大部分研究都把细根定义为一个均质系统, 即认为≤2 mm的根在结构和功能上基本相同.然而, 近年来越来越多的证据表明, 以这种单一径级定义细根的方法忽略了细根形态、化学以及生理方面的差异(
不同分类系统下油松幼苗根系特征的差异与联系
1
2010
... 近半个世纪以来, 大部分研究都把细根定义为一个均质系统, 即认为≤2 mm的根在结构和功能上基本相同.然而, 近年来越来越多的证据表明, 以这种单一径级定义细根的方法忽略了细根形态、化学以及生理方面的差异(
Long-term effects of experimental nitrogen additions on foliar litter decay and humus for mation in forest ecosystems
1
1998
... 另外, 根系分解受到微生物调控的影响(
Redefining fine roots improves understanding of belowground contributions to terrestrial biosphere processes
3
2015
... 近半个世纪以来, 大部分研究都把细根定义为一个均质系统, 即认为≤2 mm的根在结构和功能上基本相同.然而, 近年来越来越多的证据表明, 以这种单一径级定义细根的方法忽略了细根形态、化学以及生理方面的差异(
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
... ).由于低级根和高级根功能上的不同, 导致根系化学成分产生差异, 并可能对细根分解速率产生影响(
Fine root architecture of nine North American trees
4
2002
... 近半个世纪以来, 大部分研究都把细根定义为一个均质系统, 即认为≤2 mm的根在结构和功能上基本相同.然而, 近年来越来越多的证据表明, 以这种单一径级定义细根的方法忽略了细根形态、化学以及生理方面的差异(
... ).专家研究发现, 树木根系具有复杂的分枝系统(即根序), 不同根序等级的个体根在形态结构、解剖构造和生理功能等方面存在显著差异(
... ).根序等级的划分为: 根尖为1级根, 其母根为2级根, 以此类推(
... 本实验采用分解袋法研究细根分解.于2009年, 在林地里选择5-8株目标树种挖取树根.在目标树种的树干基部找到主根, 然后顺着主根向外2 m左右范围内用铲子挖取枯枝落叶层和15 cm土壤深度之间的土块, 顺主根上的侧根寻找根序分支, 确保细根为目标树种的细根.然后小心地把根系从土块中分离出来, 尽量保持根系完整性, 确保根系末端最小的根尖完整并且着生在它的母根上.根系采集主要采用手工挖掘.清除死根(颜色发暗且无弹性)以及根表面土壤后, 尽快运回实验室, 清洗后低温储存, 以便后期处理.采用与
The estimation of glycogen with the anthrone reagent
1
1950
... 在根系分解实验开始的时候, 将剩余的烘干初始样品磨碎并测定其初始化学物质.利用浓硫酸-重铬酸钾氧化法测定全C含量; 利用半微量凯氏定氮法测定全N含量; 用钼锑抗比色法测定全P含量; 采用火焰光度计法测定全K含量; 用有机质近似C组分分析法(
Global patterns in root decomposition: Comparisons of climate and litter quality effects
1
2001
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
水曲柳和兴安落叶松人工林细根分解研究
1
2008
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
水曲柳和兴安落叶松人工林细根分解研究
1
2008
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
Simulated atmospheric nitrogen deposition alters decomposition of ephemeral roots
2
2015
... 现有文献资料表明, 迄今为止仅有少量已发表的文章量化了根序和分解速率之间的关系.有研究发现, 尽管低级根具有较小的直径, 高级根具有较大的直径并且木质化程度较高, 但低级根仍然比高级根分解速率低(
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
Early stage fine-root decomposition and its relationship with root order and soil depth in a Larix gmelinii plantation
4
2016
... 现有文献资料表明, 迄今为止仅有少量已发表的文章量化了根序和分解速率之间的关系.有研究发现, 尽管低级根具有较小的直径, 高级根具有较大的直径并且木质化程度较高, 但低级根仍然比高级根分解速率低(
... ,
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
... 对4个树种进行了36个月的研究发现, 三级和四级根的质量损失率为20%-40%, 而一级和二级根的质量损失率则不超过20%.
Further evidence for slow decomposition of very fine roots using two methods: Litterbags and intact cores
1
2013a
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
Slow decomposition of very fine roots and some factors controlling the process: A 4-year experiment in four temperate tree species
1
2013b
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
Production, turnover, and nutrient dynamics of above- and belowground detritus of world forests
1
1986
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
Ecological linkages between aboveground and belowground biota
1
2004
... 在全球范围内, 超过50%的净初级生产量是通过植物组织分解(即凋落物的分解)的形式输送给土壤的(
鼎湖山南亚热带森林细根分解干物质损失和元素动态
1
1998
... 采用根系的质量残留率来表征根系分解, 即各阶段的根系干质量占初始干质量的比例.根系分解率常数(k)采用负指数衰减模型(
鼎湖山南亚热带森林细根分解干物质损失和元素动态
1
1998
... 采用根系的质量残留率来表征根系分解, 即各阶段的根系干质量占初始干质量的比例.根系分解率常数(k)采用负指数衰减模型(
Ephemeral root modules in Fraxinus mandschurica
1
2010
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
Fine roots are the dominant source of recalcitrant plant litter in sugar maple-?dominated northern hardwood forests
1
2015
... 近半个世纪以来, 大部分研究都把细根定义为一个均质系统, 即认为≤2 mm的根在结构和功能上基本相同.然而, 近年来越来越多的证据表明, 以这种单一径级定义细根的方法忽略了细根形态、化学以及生理方面的差异(
Slow decomposition and limited nitrogen release by lower order roots in eight Chinese temperate and subtropical trees
4
2012
... 现有文献资料表明, 迄今为止仅有少量已发表的文章量化了根序和分解速率之间的关系.有研究发现, 尽管低级根具有较小的直径, 高级根具有较大的直径并且木质化程度较高, 但低级根仍然比高级根分解速率低(
... ;
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
... 另外, 根系分解受到微生物调控的影响(
水曲柳和落叶松人工林细根分解与养分释放
1
2006
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(
水曲柳和落叶松人工林细根分解与养分释放
1
2006
... 在细根(≤2 mm)组分中, 树木的前2级根普遍非木质化, 具有吸收功能和较快的周转速率.而三四级根则部分已经木质化, 负责运输和贮藏等功能(

{kind=link}
{kind=link}
{kind=link}
{kind=link}