 ,, 王传宽,
,, 王传宽, ,*东北林业大学生态研究中心, 哈尔滨 150040
,*东北林业大学生态研究中心, 哈尔滨 150040Dynamics in foliar litter decomposition for Pinus koraiensis and Quercus mongolica in a snow-depth manipulation experiment
WU Qi-Qian,, WANG Chuan-Kuan,,*Center for Ecological Research, Northeast Forestry University, Harbin 150040, China通讯作者:
| 基金资助: |
Online:2018-04-19
| Supported by: | SupportedbytheNationalKeyTechnologyResearchandDevelopmentProgramoftheMinistryofScienceandTechnologyofChina( |

摘要
关键词:
Abstract
Keywords:
PDF (1144KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
武启骞, 王传宽. 控雪处理下红松和蒙古栎凋落叶分解动态. 植物生态学报, 2018, 42(2): 153-163 doi:10.17521/cjpe.2017.0184
WU Qi-Qian, WANG Chuan-Kuan.
森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(Berg & McClaugherty, 2014; Berger et al., 2015)。虽然低温限制了高寒地区森林凋落物的分解(Aerts, 2006), 但高寒地区季节性雪被的隔热和淋溶作用可促进凋落物分解(Christenson et al., 2010; Saccone et al., 2013), 从而使季节性雪被成为高寒地区森林凋落物分解的一个重要影响因素(Uchida et al., 2005; Baptist et al., 2010)。随着气候变化的加剧, 全球雪被格局已经发生了显著变化(IPCC, 2007), 并带有强烈的区域性和复杂性: 原本降水丰沛地区冬季变得更加潮湿多雪, 而某些地区的雪被可能会因全球增温而加速消融。例如, Beniston等(2003)报道阿尔卑斯山区冬季变暖使降水增加, 特别是在海拔1 700-2 000 m的地区雪被增厚; 而Ven?l?inen等(2001)报道芬兰经历暖冬, 其北部的雪被随着冬季降雪的减少而变薄。这些雪被变化格局对生物地球化学循环产生的显著影响已得到证实。然而, 目前有关森林凋落物的研究大多只关注气候变化引起的单一雪被格局的影响, 即单纯的增雪(如Aerts et al., 2012; Blok et al., 2016)或减雪(如Christenson et al., 2010; Bokhorst et al., 2013), 而雪被变化格局对森林凋落物分解影响的结果和机制尚不清楚。
虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异。例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素。但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(Mackelprang et al., 2011; Kreyling et al., 2013; Shibata et al., 2013), 从而提高分解者的数量和活性(Aanderud et al., 2013)。进入融化期, 融雪水的淋溶作用可提高凋落物内大分子有机物的降解速度(Li et al., 2016)。此外, 雪被的覆盖时间也会改变生长季长度, 进而影响生长季的凋落物分解(Christenson et al., 2010)。然而, 在少雪或无雪的地区, 严酷而多变的冬季环境一方面可限制森林凋落物的分解(Aerts, 2006), 另一方面其强烈的冻结作用又可改变凋落物的物理结构, 提高凋落物在无雪期中的可分解性(Lemma et al., 2007; Wu et al., 2010)。然而, 雪被对森林凋落物分解的综合影响尚不明确。
东北地区是中国气候变化最显著、植被系统对气候变化响应最为敏感的地区(杨金艳和王传宽, 2005), 季节性雪被期长达5个月。虽然已有大量研究关注该地区森林凋落物产量、分解动态和分解机制(如张新平等, 2008; 刘瑞鹏等, 2013; 吴鹏等, 2016), 但雪被变化格局对凋落物分解的影响尚不清楚。为此, 本研究以该区域的常绿针叶树种红松(Pinus koraiensis)和落叶阔叶树种蒙古栎(Quercus mongolica)的凋落叶为研究对象, 通过人工控雪模拟冬季雪被增加和减少, 研究雪被变化格局对不同类型和化学组分的凋落叶分解动态的影响, 为探索气候变化对森林生态系统地球生物化学循环影响提供理论依据。具体研究目标包括: (1)比较两种凋落叶在不同雪被格局下的分解动态; (2)探索凋落叶分解的调控因子; (3)量化雪被期和无雪期对凋落叶年分解总量的贡献; 以便验证3个假设: (1)凋落叶分解率受树种和雪深的显著影响, 分解率在增雪处理中最高, 除雪处理中最低; (2)分解率受分解阶段和环境因子的调控; (3)控雪将改变凋落叶在雪被期和无雪期的分解模式, 增雪处理提高雪被期对整个分解过程的贡献, 而除雪处理提高无雪期对整个分解过程的贡献。
1 研究方法
1.1 研究区域概况
研究地位于黑龙江帽儿山森林生态系统国家野外科学观测研究站(45.33°-45.42° N, 127.50°- 127.57° E)。平均海拔400 m, 平均坡度10°-15°, 土壤主要为暗棕色森林土。该地区属大陆性季风气候, 春季多风干燥, 夏季湿润多雨, 冬季干燥寒冷。年降水量629 mm, 年潜在蒸发量854 mm, 年平均气温3.1 ℃, 11月至次年4月地表有明显的季节性雪被覆盖(Wang et al., 2013)。植被是阔叶红松林经过不同程度的干扰(采伐、经营、火烧和开垦等)后形成的天然次生林和人工林。主要组成树种包括红松、蒙古栎、白桦(Betula platyphylla)等; 林下灌木主要包括丁香(Syringa spp.)、卫矛(Euonymus spp.)、绣线菊(Spiraea spp.)等; 草本植物主要包括薹草(Carex spp.)、山茄子(Brachybotrys paridiformis)、白花碎米荠(Cardamine leucantha)等(Zhang & Wang, 2010)。1.2 控雪试验
控雪试验设在研究区的红松人工林, 坡度12°, 坡向西北, 林龄约51 a, 密度约3 145株·hm-2, 平均胸径12.9 cm, 平均树高12.1 m。于2014年9月在该林分所在坡地上、中、下部各随机设置1个30 m × 20 m的标准样地, 样地总面积0.18 hm2。为保证各样地的相对独立性, 将样地间距保持在100 m以上。每个样地中随机设置3个5 m × 5 m的样方, 样方间距为3- 4 m。3个样方分别为: 对照(无处理)、增雪处理和除雪处理。11月至次年4月, 于各样地的除雪样方中设置除雪设施。除雪设施主体采用钢结构搭建(2.5 m × 2.5 m × 0.3 m), 上覆透明有机玻璃板(上海绅尔塑胶有限公司, 5 mm厚, 透光率> 92%), 同时采用遮阳网围挡除雪设施四周; 这样既可以保证设施内通风, 又可以避免风对雪的搬运作用的影响。每次降雪后, 小心将除雪设施上的积雪集中, 用大号网筛均匀地转移至增雪样方中心2.5 m × 2.5 m区域, 最大限度地模拟自然条件下的雪被增加。同时用直尺直接多点测量对照和增雪处理的雪深, 取得平均值(共18次降雪; 图1)。为避免融雪干扰, 在融化期到来之前(4月上旬), 于各样方周围挖一条20 cm深的阻水壕沟。次年5月至10月为无雪期, 将除雪设施移除, 整个阶段不做任何控制处理。此外, 在每个样方中设置纽扣式温度记录器(iButton DS1923-F5, Maxim/Dallas Semiconductor, Sunnyvale, USA), 自动记录试验期内凋落物层和大气每小时的温度变化(图2)。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同处理的雪深动态(平均值±标准偏差, n = 15)。*, p < 0.05。
Fig. 1Dynamics in snow-depth in different treatment plots (mean ± SD, n = 15). *, p < 0.05.
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2大气温度和不同处理的凋落物层温度动态(平均值, n = 3)。A, 大气温度和对照(CK)中凋落物层温度。B, 增雪处理(SA)和除雪处理(SR)与对照相比的凋落物层的温差。正值代表处理后温度升高, 负值代表处理后温度降低。
Fig. 2Dynamics in air temperature and the temperatures at the litter layer in different treatment plots (mean, n = 3). A, average air temperature and temperature at the litter layer under the control (CK). B, difference in the temperature between the snow-addition (SA) or snow-removal treatment (SR) and the control. Positive value indicates increased temperature after the treatment, while negative value indicates decreased temperature after the treatment.
1.3 凋落叶分解试验及室内分析
2014年9月初, 在研究区收集红松和蒙古栎新鲜凋落叶, 带回实验室自然风干。每个树种的凋落叶分别称取5份, 每份10.0 g, 置于70 ℃烘箱烘干至恒质量, 由此推算凋落叶样品含水率及初始干质量。烘干后的凋落叶研磨过60目筛, 测定其初始有机碳、全氮、全磷、木质素和纤维素含量(表1)。称取相当于烘干质量10.0 g的风干凋落叶放入分解袋中封口备用。分解袋为尼龙材质, 尺寸15 cm × 20 cm, 孔径1 mm。2014年10月25日将分解袋平铺于每个样方中, 相邻分解袋之间保持2 cm以上的间距, 以避免相互影响。试验按照12次取样设计, 并设置3个取样重复, 凋落物袋共为648个(3个样地× 3种控雪处理× 2个树种× 3个取样重复× 12次取样)。为排除干扰, 铺设凋落物袋前尽量去除地表已有的新鲜凋落叶。Table 1
表1
表1红松和蒙古栎凋落叶的初始质量(平均值±标准偏差, n = 5)
Table 1
| 树种 Tree species | 有机碳 Organic carbon (g·kg-1) | 全氮 Total nitrogen (g·kg-1) | 全磷 Total phosphorus (g·kg-1) | 碳/氮 C/N | 碳/磷 C/P | 氮/磷 N/P | 木质素 Lignin (%) | 纤维素 Cellulose (%) | 木质素/氮 Lignin/N |
|---|---|---|---|---|---|---|---|---|---|
| 红松 Pinus koraiensis | 489.6 ± 1.4a | 4.5 ± 0.1b | 0.55 ± 0.01b | 107.0 ± 1.2a | 893.5 ± 6.1a | 8.1 ± 0.5a | 29.8 ± 0.3a | 14.6 ± 0.4a | 66.8 ± 0.2a |
| 蒙古栎 Quercus mongolica | 458.8 ± 5.1b | 6.5 ± 0.1a | 1.30 ± 0.05a | 71.1 ± 0.6b | 351.7 ± 13.5b | 4.9 ± 0.2b | 16.1 ± 0.1b | 12.9 ± 0.1b | 25.0 ± 0.6b |
新窗口打开|下载CSV
为了解控雪对凋落叶分解的影响, 基于研究区历年气象观测数据, 根据Olsson等(2003)提出的划分原则, 将全年分为冻结初期、深冻期、融化期、无雪初期、无雪中期、无雪末期等6个阶段进行凋落叶取样分析测定(表2), 以探索其分解动态。
Table 2
表2
表2凋落叶分解过程中的取样阶段、日期及分解天数
Table 2
| 取样顺序 Sampling order | 取样阶段 Sampling stage | 取样日期 Sampling date | 分解天数 Decomposing days |
|---|---|---|---|
| 1 | 第一年冻结初期 1st year freezing onset stage | 2014-12-02 | 49 |
| 2 | 第一年深冻期 1st year deep freezing stage | 2015-03-18 | 145 |
| 3 | 第一年融化期 1st year thawing stage | 2015-04-18 | 176 |
| 4 | 第一年无雪初期 1st year early snow-free season | 2015-06-20 | 239 |
| 5 | 第一年无雪中期 1st year mid snow-free season | 2015-08-20 | 300 |
| 6 | 第一年无雪末期 1st year late snow-free season | 2015-10-20 | 366 |
| 7 | 第二年冻结初期 2nd year freezing onset stage | 2015-12-25 | 407 |
| 8 | 第二年深冻期 2nd year deep freezing stage | 2016-03-25 | 516 |
| 9 | 第二年融化期 2nd year thawing stage | 2016-04-22 | 544 |
| 10 | 第二年无雪初期 2nd year early snow-free season | 2016-06-20 | 603 |
| 11 | 第二年无雪中期 2nd year mid snow-free season | 2016-08-22 | 666 |
| 12 | 第二年无雪末期 2nd year late snow-free season | 2016-10-24 | 732 |
新窗口打开|下载CSV
每次取样时, 随机从各样方中采集红松和蒙古栎分解袋各3袋, 小心去掉新生根系及杂物后装入无菌聚乙烯袋中带回实验室, 70 ℃烘干至恒质量后测定凋落叶残留干质量。采集分解袋的同时, 采集袋下对应的土壤有机层样品(5 cm × 10 cm), 小心去除异物后风干, 供有机层有机碳、全氮和全磷含量的测定使用(表3)。在取样同时, 读取不同处理的温度, 据此计算凋落物层日平均温度、冻融循环次数和大气日平均温度(表3)。凋落物层温度高于或低于0 ℃ 3 h以上直到低于或高于0 ℃ 3 h以上计为1次冻融循环(Zhu et al., 2012)。凋落叶和有机层有机碳含量采取燃烧法, 用Multi N/C 3000分析仪和HT 1500 Solids Module固体模块(Analytik Jena AG, Thüringen, Germany)测定; 经H2SO4-H2O2消煮后, 在连续流动分析仪(Bran + Luebbe, Norderstedt, Germany)上测定凋落叶和有机层的全氮、全磷含量; 经酸性洗液充分洗涤后, 采用差重法测定凋落叶木质素和纤维素含量(Vanderbilt et al., 2008)。
Table 3
表3
表3不同处理下凋落叶分解过程中环境特征(平均值±标准偏差, n = 3)
Table 3
| 处理 Treatment | 凋落物层 平均温度 Average temperature in litter layer (°C) | 凋落物层 冻融循环 Freeze-thaw cycle in litter layer | 有机层有机碳 Organic carbon in organic layer (g·kg-1) | 有机层全氮 Total nitrogen in organic layer (g·kg-1) | 有机层全磷 Total phosphorus in organic layer (g·kg-1) | 有机层碳/氮 C/N in organic layer | 有机层碳/磷 C/P in organic layer | 有机层氮/磷 N/P in organic layer |
|---|---|---|---|---|---|---|---|---|
| 增雪 Snow-addition | 4.4 ± 0.3a | 58 ± 1c | 81.4 ± 2.2a | 7.4 ± 0.2a | 1.3 ± 0.1a | 10.9 ± 0.2b | 62.4 ± 0.3c | 5.5 ± 0.2c |
| 对照 Control | 3.8 ± 0.2b | 70 ± 1b | 75.4 ± 1.1b | 6.6 ± 0.3b | 0.9 ± 0.2b | 11.2 ± 0.2b | 83.9 ± 0.4b | 7.2 ± 0.1b |
| 除雪 Snow-removal | 2.7 ± 0.5c | 84 ± 2a | 72.6 ± 0.6c | 6.1 ± 0.1c | 0.7 ± 0.1c | 12.0 ± 0.3a | 104.3 ± 0.6a | 8.7 ± 0.3a |
新窗口打开|下载CSV
1.4 数据分析
凋落叶分解过程采用指数模型(Olson, 1963)模拟:y = ae-kt (1)
式中: y表示凋落叶月残留率(%); a表示拟合参数; e表示自然对数底; t表示分解时间(月); k表示分解系数。凋落叶半分解时间(t0.5)为:
t0.5 = ln (50/a)/(-k) (2)
凋落叶95%分解时间(t0.95)为:
t0. 95 = ln (5/a)/(-k) (3)
凋落叶分解率(Lt)为:
Lt (%) = (Mt+1 - Mt)/M0 × 100% (4)
式中: M0表示分解袋埋置前袋中凋落叶质量(g); (Mt+1 - Mt)表示相邻两阶段分解袋中凋落叶质量差(g)。凋落叶各季节分解量对分解总量的贡献率(P) (Zhu et al., 2012)为:
P(%) = (Mt+1 - Mt)/(M0 - MT) × 100% (5)
式中: MT表示最后一次取样时分解袋中凋落叶质量(g)。
采用独立样本t检验比较两个树种凋落叶初始质量的差异; 非线性回归分析拟合凋落叶分解曲线; 单因素方差分析和Bonferroni校正比较不同控雪处理中雪深、凋落叶分解率和环境因子的差异; 重复测量方差分析检验树种、雪深和分解阶段对分解率的影响; 逐步回归分析探索凋落叶的分解率与其初始质量、环境因子的关系。数据统计与分析采用SPSS 20.0 (SPSS, Chicago, USA)完成。
2 结果
2.1 不同处理下两个树种凋落叶分解动态
不同处理中两个树种的凋落叶分解动态格局基本一致, 但分解率的大小随分解阶段和处理而变(图3)。雪被期凋落叶分解率随雪被的增厚而升高, 即增雪处理最高, 除雪处理最低。相反, 无雪期的除雪处理的分解率最高, 增雪处理的分解率最低(图3B、3D)。红松凋落叶分解率在第一年无雪中期(第300天)和第二年深冻期(第516天)之外的所有阶段的处理之间差异均显著(p < 0.05; 图3A); 而蒙古栎凋落叶分解率在第一年无雪中期(第300天)和第二年无雪初期(第603天)之外的所有阶段的处理之间差异均显著(p < 0.05; 图3C)。综合两年分解过程, 与对照相比, 增雪处理使红松和蒙古栎的凋落叶分解率分别升高2.2%和2.6%; 而除雪处理使其分解率分别降低0.3%和2.7% (图3B、3D)。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同处理下两个树种的凋落叶分解率的比较及时间动态(平均值±标准偏差, n = 3)。*和不同字母代表处理间差异显著(p < 0.05)。
Fig. 3Comparisons and dynamics in the foliar litter decomposition rates of the two tree species under different treatments (mean ± SD, n = 3). * and different letters indicate significant differences among the treatments (p < 0.05).
两个树种凋落叶分解系数均以增雪处理最大, 除雪处理最小; 半分解时间和95%分解时间也按照增雪处理—对照—除雪处理的顺序依次延长(表4)。与对照相比, 增雪处理中红松凋落叶的半分解和95%分解时间分别缩短了1.38月和4.03月, 蒙古栎分别缩短了1.75月和7.24月; 而除雪处理中, 两个树种凋落叶半分解和95%分解时间分别延长了2.22月和8.10月(红松)及2.93月和9.49月(蒙古栎)。
Table 4
表4
表4不同处理下红松和蒙古栎凋落叶分解模型、分解系数(k)、决定系数(R2)、半分解(t0.5)和95%分解时间(t0.95)
Table 4
| 树种 Tree species | 处理 Treatment | 回归方程 Regression model | k | R2 | 半分解时间 t0.5 (month) | 95%分解时间 t0.95 (month) |
|---|---|---|---|---|---|---|
| 红松 Pinus koraiensis | 增雪 Snow-addition | y = 99.691e-0.030t | 0.030 | 0.982 | 23.00 | 99.75 |
| 对照 Control | y = 101.398e-0.029t | 0.029 | 0.979 | 24.38 | 103.78 | |
| 除雪 Snow-removal | y = 102.545e-0.027t | 0.027 | 0.965 | 26.60 | 111.88 | |
| 蒙古栎 Quercus mongolica | 增雪 Snow-addition | y = 100.026e-0.037t | 0.037 | 0.984 | 18.74 | 80.97 |
| 对照 Control | y = 100.342e-0.034t | 0.034 | 0.973 | 20.49 | 88.21 | |
| 除雪 Snow-removal | y = 103.359e-0.031t | 0.031 | 0.955 | 23.42 | 97.70 |
新窗口打开|下载CSV
2.2 凋落叶分解率的影响因素
重复测量方差分析表明, 树种、雪深和分解阶段及其交互作用均显著地影响凋落叶分解率(p < 0.05; 表5)。进一步的逐步回归分析表明, 虽然凋落叶初始质量和环境因子显著影响凋落叶分解率, 但各处理中有显著影响的因子不同(表6)。增雪处理中, 凋落叶分解率受凋落叶初始有机碳含量、凋落物层平均温度、冻融循环和有机层全磷含量的显著影响(R2 = 0.571); 对照中, 有显著影响的因子为凋落叶初始有机碳含量、初始C/N、凋落物层平均温度和有机层全磷含量(R2 = 0.697); 而除雪处理中则为凋落叶初始有机碳含量、初始木质素含量、凋落物层平均温度、有机层全氮和全磷含量(R2 = 0.404)。Table 5
表5
表5凋落叶分解率与树种、雪深和分解阶段的重复测量方差分析
Table 5
| 因子 Factor | df | F | p |
|---|---|---|---|
| 树种 Tree species | 1/2 | 85.9 | <0.001 |
| 雪深 Snow-depth | 2/12 | 52.8 | <0.001 |
| 分解阶段 Decomposition stage | 11/48 | 3371.1 | <0.001 |
| 树种×雪深 Tree species × Snow-depth | 2/12 | 4.33 | 0.014 |
| 树种×分解阶段 Tree species × Decomposition stage | 11/48 | 21.5 | <0.001 |
| 雪深×分解阶段 Snow-depth × Decomposition stage | 22/48 | 3.58 | 0.028 |
| 树种×雪深×分解阶段 Tree species × Snow-depth × Decomposition stage | 22/48 | 3.16 | 0.045 |
新窗口打开|下载CSV
表6
表6凋落叶分解率与初始质量和环境因子的逐步回归分析
| 处理 Treatment | 回归方程 Regression model | R2 | p |
|---|---|---|---|
| 增雪 Snow-addition | y = 107.012 - 0.139C + 0.237AT + 0.014FTC + 1.276TP | 0.571 | 0.02 |
| 对照 Control | y = 98.152 - 0.114C - 7.263C/N + 0.209AT + 1.482TP | 0.697 | < 0.01 |
| 除雪 Snow-removal | y = 102.834 - 0.107C - 7.074Lignin +0.384AT + 0.195TN + 1.425TP | 0.404 | 0.03 |
新窗口打开|下载CSV
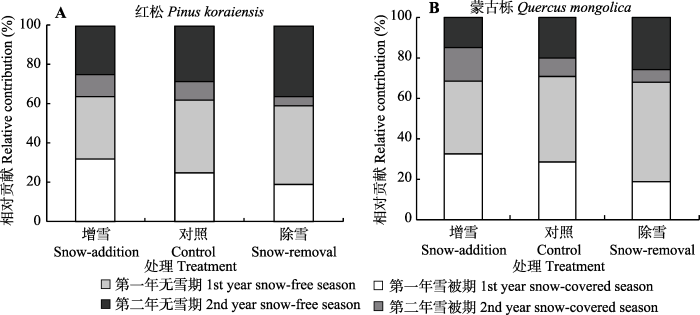
2.3 雪被期与无雪期凋落叶分解的贡献率
为期两年的试验表明, 雪被期是凋落叶分解的重要时期, 其中雪被期为红松凋落叶的分解总量贡献了24.1%以上, 为蒙古栎贡献了25.2%以上(图4)。控雪改变了两个树种凋落叶雪被期和无雪期的分解模式: 雪被期的贡献率随雪深增加而升高, 而无雪期的贡献率随雪深增加而降低。增雪处理提高了雪被期对凋落叶分解的贡献, 而降低了无雪期的贡献。相反, 除雪处理降低了雪被期对凋落叶分解的贡献, 提高了无雪期对分解的贡献。与对照相比, 增雪处理使红松和蒙古栎的雪被期分解贡献率分别提高了9.1%和10.4%; 除雪处理使无雪期贡献率分别提高了10.4%和12.7%。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同处理下雪被期和无雪期对两个树种凋落叶分解总量的相对贡献(平均值, n = 3)。
Fig. 4Relative contribution of litter loss during the snow-covered and snow-free seasons to the total annual litter loss for the two tree species under different treatments (mean, n = 3).
3 讨论
本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低。增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间。这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程。这可能有以下几方面解释: (1)控雪改变分解者动态。雪被良好的隔热能力可以提升雪下温度(表2)和湿度(Ayres et al., 2010), 使微环境相对稳定(图2)。参与分解的微生物和土壤动物在雪被下避免了频繁的冻融循环和动荡的环境变化, 其活性、种群结构和多样性得以改善(Brooks & Williams, 1999; Bokhorst et al., 2013)。而除雪处理导致凋落物层环境动荡(Comerford et al., 2013; Shibata et al., 2013), 耐受性较差的分解者会在严酷的环境下大量死亡(Templer et al., 2012)。这种分解者动态的变异是造成分解率差异的主要原因。(2)控雪改变酶活性。凋落叶的分解离不开酶系统的综合作用, 除雪处理后严酷的环境条件造成大量植物根系、土壤微生物和土壤动物死亡, 土壤中酶的输入来源减少, 阻碍了凋落叶分解进程。此外, 作为参与凋落叶分解的主要酶, 如纤维素酶、几丁质酶和磷酸酶等, 其活性都与环境温度或湿度呈正相关关系(Fioretto et al., 2000; Criquet et al., 2004)。因此, 增雪处理中适宜的温度和湿度提高了酶的活性, 促进了凋落叶分解; 相反, 除雪处理中凋落叶分解过程受到阻碍。(3)控雪改变凋落叶质量。与除雪处理相比, 增雪处理带来的自然粉碎作用(冻结、压迫等)可以直接破坏凋落叶的物理结构, 增加凋落叶表面积, 提高分解者对底物利用有效性, 同时加速了纤维素、半纤维素和木质素等组分的降解(Groffman et al., 2001), 使凋落叶中养分、酸溶性物质和未木质化的糖类等物质聚合形成酸不溶性物质, 促进凋落叶分解(Berg & McClaugherty, 2014)。本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子。与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(郭剑芬等, 2006)。与对照相比, 除雪处理中分解率还与初始木质素含量显著负相关, 这是由于除雪处理中严酷的自然条件抑制了木质素分解酶的活性, 从而限制了木质素降解, 降低凋落叶分解率(Berg & McClaugherty, 2014)。虽然气候条件、凋落物质量和分解者是影响分解的3个要素, 但越来越多的研究表明, 局域环境因子对凋落物分解的影响大于气候条件(Bradford et al., 2016), 本研究支持以上结论。温度作为影响生命活动的主导因子, 无论直接影响还是间接作用, 在一定范围内有机质的分解速率都随温度升高而加快(Hornsby et al., 1995)。增雪处理中凋落物层冻融循环与分解率显著正相关, 这是因为与对照和除雪处理相比, 增雪处理中冻融循环次数较少(表2), 冻融循环对凋落叶的破碎作用成为该处理中影响分解率的关键环境因子(Wu et al., 2010)。土壤养分对凋落叶分解的影响不可忽视, 凋落叶往往在养分有效性高的土壤中分解较快, 在养分有效性低的土壤中分解较慢。如Hobbie (1992)发现, 凋落物分解率与土壤氮有效性显著正相关。Kaspari等(2008)也报道, 添加P后凋落物分解率可以提升33%。在两年的增雪处理中, 有机层全氮、全磷含量显著提高; 相反, 除雪处理中两种元素含量显著降低(表3)。除雪处理中有机层全氮含量与分解率之间显著正相关, 表明N成为限制该处理中凋落叶分解的重要因素。这可能是因为除雪处理中相对更低的温度降低了微生物活性, 导致N的固定率下降(胡霞等, 2012), 限制了腐生真菌从土壤获取无机氮参与凋落叶分解的过程(郑俊强和韩士杰, 2016), 从而使凋落叶分解进程受阻。然而, N仅限制了除雪处理中的凋落叶分解, 可能是该区域冬季氮沉降累积在积雪中, 部分解除了N对微生物活动的限制(Madritch & Hunter, 2003)。值得注意的是, 本研究有机层全磷含量也与凋落叶分解率显著正相关。高寒地区所具有的强烈、频繁的冻结和融化作用使P元素经历了强烈的淋洗、降解过程(武启骞等, 2015), 可能造成P流失。因此与N相比, 微生物更难从外界获取P以满足分解过程的营养需求(郭剑芬等, 2006), 从而使P成为该区凋落叶分解更为重要的限制因子。随着全球氮沉降加剧, P对生态系统过程的限制可能逐步凸显。因此, P及氮磷耦合对凋落叶分解的限制作用需要在未来的研究中进一步关注。
经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(如Aerts, 2006; Bokhorst et al., 2013等)。本研究中雪被期对于两个树种凋落叶整个分解过程的贡献率达到24.1%以上, 体现了雪被期对凋落叶分解的重要作用, 与其他高寒生态系统凋落物分解的研究结果(Hobbie, 1996; 何伟等, 2013)一致。雪被期对凋落叶分解的贡献率随雪深的增加而增加; 但无雪期对凋落叶分解的贡献率随雪深的增加而减小。这主要与雪被带来的微环境变化有关: 一方面, 增雪处理可以为雪被期的凋落叶分解提供适宜的环境, 提高该时期凋落叶分解量和贡献率(胡霞等, 2012)。而除雪处理后, 地表凋落叶不得不面对严峻而频繁的温度变化, 凋落叶分解进程受到不同程度的阻碍, 分解量及贡献率随之降低; 另一方面, 增雪处理会在融化期和无雪初期中产生大量融雪水, 从而降低了环境中分解者的活性和数量, 使无雪期的凋落叶分解量和贡献率降低(Edwards et al., 2007; Blok et al., 2016)。另外, 增雪处理的凋落叶在雪被期快速分解, 消耗较多的易分解组分, 从而也会降低无雪期的分解量和贡献率(Tomaselli, 1991)。反之, 除雪处理后的凋落叶无需面临类似问题, 其中的分解者活性和数量一直保持在较稳定状态, 使得无雪期的分解量及贡献率维持在较高水平。这些结果表明, 雪被格局的改变不仅会对雪被期凋落叶分解产生即时的影响, 还会对后续分解过程产生不同程度的延续效应, 进一步影响凋落叶在无雪期的分解动态(Saccone et al., 2013; Carbognani et al., 2014)。
4 结论
全面理解雪被格局对森林凋落物分解的影响因子和驱动作用是森林生态学领域值得关注的重要命题, 需要采用多种情景模拟研究和长期观察相结合的方法。本研究首次在东北温带人工林中开展了为期2年的增雪和除雪人工控制对红松和蒙古栎凋落叶分解影响的研究探索, 虽然还有局限性(如重复性、长期性、多方案性等), 但我们发现: (1)控雪显著影响了凋落叶分解率。增雪处理使分解率升高, 半分解和95%分解时间缩短; 除雪处理使分解率降低, 半分解和95%分解时间延长。(2)雪深或分解阶段对凋落叶分解过程具有重要影响, 主要通过温度和环境中养分元素起调控作用。(3)控雪改变了凋落叶雪被期和无雪期的分解模式。增雪处理提高了雪被期贡献; 除雪处理提高了无雪期贡献。因此, 由气候变化带来的冬季雪被的变化不但会对温带森林凋落物分解产生显著的瞬时影响, 而且会对融雪之后无雪期的分解过程产生持续影响。致谢 感谢东北林业大学生态研究中心全先奎、窦佳和赵娟在野外取样和室内分析中提供的帮助; 感谢黑龙江帽儿山森林生态系统国家野外科学观测研究站供野外试验平台。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.soilbio.2012.07.022URL [本文引用: 1]

Winter respiration is a quantitatively important, yet variable flux of carbon dioxide (CO2) from soils to the atmosphere. Variability in winter soil respiration may be influenced by the effects of snowfall on microbial communities and their metabolic activities. In this study, we evaluated the importance of snowpack depth on soil respiration and microbial communities in a temperate deciduous forest. Snow removal created relatively dry, frequently frozen, and carbon substrate-poor soils, while snow additions led to wetter, warmer, and relatively carbon substrate-rich soils. Using time-series multiple regression, we observed enhanced sensitivity of respiration to moisture under ambient snow and snow removal; however, this effect was accompanied by a temporal lag suggesting that microorganisms had a delayed response to increases in free-water during soil thawing events. Conversely, soil respiration was only sensitive to temperature in the snow addition treatment when soil temperatures were consistently above 002°C. The snow-induced respiration dynamics were accompanied by shifts in the structure of wintertime fungal and bacterial communities. We detected an impact of altered snowpack on bacterial richness during the growing season, but our manipulation did not have legacy effects on other features of the soil microbial community at spring thaw. Our results suggest that microbial communities may be “reset” during seasonal transitions from winter to spring, and that soil microorganisms are likely adapted to annual fluctuations in snowpack depth. As snowpack becomes more variable in mid-latitude systems due to climate change, our findings suggest that soil moisture and temperature will co-regulate wintertime respiration through a non-linear relationship surrounding soil freeze–thaw cycles, with snow-mediated changes in microbial community structure likely influencing wintertime respiration dynamics.
DOI:10.1111/j.1365-2745.2006.01142.xURL [本文引用: 3]
Summary Top of page Summary Introduction Direct effects of temperature Indirect temperature effects on litter chemistry Indirect effects on detritivore and decomposer communities How to proceed? Acknowledgements References 1 Decomposition of plant litter, a key component of the global carbon budget, is hierarchically controlled by the triad: climate > litter quality > soil organisms. Given the sensitivity of decomposition to temperature, especially in cold biomes, it has been hypothesized that global warming will lead to increased litter decomposition rates, both through direct temperature effects and through indirect effects on litter quality and soil organisms. 2 A meta-analysis of experimental warming studies in cold biomes (34 site-species combinations) showed that warming resulted in slightly increased decomposition rates. However, this response was strongly dependent on the method used: open top chambers reduced decomposition rates, whereas heating lamps stimulated decomposition rates. The low responsiveness was mainly due to moisture-limited decomposition rates in the warming treatments, especially at mesic and xeric sites. This control of litter decomposition by both temperature and moisture was corroborated by natural gradient studies. 3 Interspecific differences in litter quality and decomposability are substantially larger than warming-induced phenotypic responses. Thus, the changes in the species composition and structure of plant communities that have been observed in medium-term warming studies in cold biomes will have a considerably greater impact on ecosystem litter decomposition than phenotypic responses. 4 Soil fauna communities in cold biomes are responsive to climate warming. Moreover, temperature-driven migration of the, hitherto absent, large comminuters to high-latitude sites may significantly increase decomposition rates. However, we do not know how far-reaching the consequences of changes in the species composition and structure of the soil community are for litter decomposition, as there is a lack of data on functional species redundancy and the species鈥 dispersal ability. 5 Global warming will lead to increased litter decomposition rates only if there is sufficient soil moisture. Hence, climate scenario and experimental studies should focus more on both factors and their interaction. As interspecific differences in potential decomposability and litter chemistry are substantially larger than phenotypic responses to warming, the focus of future research should be on the former. In addition, more light should be shed on the below-ground 鈥榙arkness鈥 to evaluate the ecological significance of warming-induced soil fauna community changes for litter decomposition processes in cold biomes.
DOI:10.1007/s00442-012-2330-zURLPMID:3470819 [本文引用: 1]
Litter decomposition and nutrient mineralization in high-latitude peatlands are constrained by low temperatures. So far, little is known about the effects of seasonal components of climate change (higher spring and summer temperatures, more snow which leads to higher winter soil temperatures) on these processes. In a 4-year field experiment, we manipulated these seasonal components in a sub-arctic bog and studied the effects on the decomposition and N and P dynamics of leaf litter ofCalamagrostis lapponica,Betula nana, andRubus chamaemorus, incubated both in a common ambient environment and in the treatment plots. Mass loss in the controls increased in the orderCalamagrostis<Betula<Rubus. After 4聽years, overall mass loss in the climate-treatment plots was 10聽% higher compared to the ambient incubation environment. Litter chemistry showed within each incubation environment only a few and species-specific responses. Compared to the interspecific differences, they resulted in only moderate climate treatment effects on mass loss and these differed among seasons and species. Neither N nor P mineralization in the litter were affected by the incubation environment. Remarkably, for all species, no net N mineralization had occurred in any of the treatments during 4聽years. Species differed in P-release patterns, and summer warming strongly stimulated P release for all species. Thus, moderate changes in summer temperatures and/or winter snow addition have limited effects on litter decomposition rates and N dynamics, but summer warming does stimulate litter P release. As a result, N-limitation of plant growth in this sub-arctic bog may be sustained or even further promoted.
DOI:10.1007/s00300-010-0766-3URL [本文引用: 1]
Snow accumulation can influence soil properties in arctic and alpine tundra, boreal and temperate forests, and temperate grasslands. However, snow may be even more influential in arid ecosystems, which by definition are water limited, such as the hyper-arid polar desert of the McMurdo Dry Valleys, Antarctica. Moreover, snow accumulation may be altered by climate change in the future. In order to investigate the impact of changes in snow accumulation on soils in the McMurdo Dry Valleys we experimentally manipulated the quantity of snow at two locations and monitored soil properties over 5聽years in relation to a snow depth gradient created by snow fences. We predicted that increased snow depth would be associated with increased soil moisture and a shift in soil animal community structure. While we did not observe changes in soil biochemistry or community structure along the snow depth gradient at either site, increased snow accumulation caused by the snow fence altered soil properties across the entire length of the transects at one site (Fryxell), which collected substantially more snow than the other site. At Fryxell, the presence of the snow fence increased gravimetric soil moisture from 1 to 5鈥9%. This was associated with a decline in abundance of the dominant animal, Scottnema lindsayae , a nematode typically found in dry soil, and an increase in Eudorylaimus sp. a nematode associated with moist soil. We also observed changes in soil pH, salinity, and concentrations of inorganic nitrogen and chlorophyll a over the course of the experiment, but it was difficult to determine if these were caused by snow accumulation or simply represented temporal variation related to other factors.
DOI:10.1007/s11104-009-0119-6URL [本文引用: 1]
DOI:10.1007/s00704-002-0709-1URL
09In many instances, snow cover and duration are a major controlling factor on a range of environmental systems in mountain regions. When assessing the impacts of climatic change on mountain ecosystems and river basins whose origin lie in the Alps, one of the key controls on such systems will reside in changes in snow amount and duration. At present, regional climate models or statistical downscaling techniques, which are the principal methods applied to the derivation of climatic variables in a future, changing climate, do not provide adequate information at the scales required for investigations in which snow is playing a major role. A study has thus been undertaken on the behavior of snow in the Swiss Alps, in particular the duration of the seasonal snow-pack, on the basis of observational data from a number of Swiss climatological stations. It is seen that there is a distinct link between snow-cover duration and height (i.e., temperature), and that this link has a specific “signature” according to the type of winter. Milder winters are associated with higher precipitation levels than colder winters, but with more solid precipitation at elevations exceeding 1,700–2,00065m above sea-level, and more liquid precipitation below. These results can be combined within a single diagram, linking winter minimum temperature, winter precipitation, and snow-cover duration. The resulting contour surfaces can then be used to assess the manner in which the length of the snow-season may change according to specified shifts in temperature and precipitation. While the technique is clearly empirical, it can be combined with regional climate model information to provide a useful estimate of the length of the snow season with snow cover, for various climate-impacts studies.
[本文引用: 3]
[本文引用: 1]
DOI:10.1007/s10021-015-9924-3URL [本文引用: 2]
The Arctic climate is projected to change during the coming century, with expected higher air temperatures and increased winter snowfall. These climatic changes might alter litter decomposition rates, which in turn could affect carbon (C) and nitrogen (N) cycling rates in tundra ecosystems. However, little is known of seasonal climate change effects on plant litter decomposition rates and N dynamics, hampering predictions of future arctic vegetation composition and the tundra C balance. We tested the effects of snow addition (snow fences), warming (open top chambers), and shrub removal (clipping), using a full-factorial experiment, on mass loss and N dynamics of two shrub tissue types with contrasting quality: deciduous shrub leaf litter (Salix glauca) and evergreen shrub shoots (Cassiope tetragona). We performed a 10.5-month decomposition experiment in a low-arctic shrub tundra heath in West-Greenland. Field incubations started in late fall, with harvests made after 249, 273, and 319聽days of field incubation during early spring, summer and fall of the next year, respectively. We observed a positive effect of deeper snow on winter mass loss which is considered a result of observed higher soil winter temperatures and corresponding increased winter microbial litter decomposition in deep-snow plots. In contrast, warming reduced litter mass loss during spring, possibly because the dry spring conditions might have dried out the litter layer and thereby limited microbial litter decomposition. Shrub removal had a small positive effect on litter mass loss for C. tetragona during summer, but not for S. glauca. Nitrogen dynamics in decomposing leaves and shoots were not affected by the treatments but did show differences in temporal patterns between tissue types: there was a net immobilization of N by C. tetragona shoots after the winter incubation, while S. glauca leaf N-pools were unaltered over time. Our results support the widely hypothesized positive linkage between winter snow depth and litter decomposition rates in tundra ecosystems, but our results do not reveal changes in N dynamics during initial decomposition stages. Our study also shows contrasting impacts of spring warming and snow addition on shrub decomposition rates that might have important consequences for plant community composition and vegetation-climate feedbacks in rapidly changing tundra ecosystems.
DOI:10.1016/j.soilbio.2013.03.016URL [本文引用: 3]
Decomposition of organic matter in high latitude biomes makes a significant contribution to global fluxes of nutrients and carbon and is expected to accelerate due to climate change. The majority of studies have focused on decomposition during the growing season, but winter climate is expected to change dramatically. Furthermore, knowledge of the drivers of organic matter decomposition, such as litter chemical composition, has primarily been tested across the growing season so it is unknown whether these drivers are also important during the winter. Given that the depth of snow cover insulates the sub-nivean climate from the much colder air, it is an important control on winter decomposition and is expected to be influenced by climate change, we experimentally manipulated snow cover to simulate impacts of different winter precipitation scenarios on soil processes. Our results show that despite snow reduction negatively affecting decomposer abundance (by 99%) and bulk soil respiration (by 47%), litter decomposition rates showed little to no response. Furthermore, variation in winter decomposition rates among litter types was unrelated to nutrient status, indicating that our current understanding of drivers of litter decomposition may not hold during winter months. Despite very large reductions in decomposer fauna due to snow removal, litter decomposition rates were not consistently responsive, indicative of decoupled responses of soil organisms and soil processes to winter climate change. (c) 2013 Elsevier Ltd. All rights reserved.
DOI:10.1111/1365-2745.12507URL [本文引用: 1]
Summary Litter decomposition is a biogeochemical process fundamental to element cycling within ecosystems, influencing plant productivity, species composition and carbon storage. Climate has long been considered the primary broad-scale control on litter decomposition rates, yet recent work suggests that plant litter traits may predominate. Both decomposition paradigms, however, rely on inferences from cross-biome litter decomposition studies that analyse site-level means. We re-analyse data from a classical cross-biome study to demonstrate that previous research may falsely inflate the regulatory role of climate on decomposition and mask the influence of unmeasured local-scale factors. Using the re-analysis as a platform, we advocate experimental designs of litter decomposition studies that involve high within-site replication, measurements of regulatory factors and processes at the same local spatial grain, analysis of individual observations and biome-scale gradients. Synthesis . We question the assumption that climate is the predominant regulator of decomposition rates at broad spatial scales. We propose a framework for a new generation of studies focused on factoring local-scale variation into the measurement and analysis of soil processes across broad scales. Such efforts may suggest a revised decomposition paradigm and ultimately improve confidence in the structure, parameter estimates and hence projections of biogeochemical models.
DOI:10.1002/(ISSN)1099-1085URL [本文引用: 1]
DOI:10.1007/s11104-013-1982-8URL [本文引用: 1]
Background and aims In cold biomes, litter decomposition, which controls the nutrient availability for plants and the ecosystem carbon budget, is strongly influenced by climatic conditions. In this...
DOI:10.1111/j.1365-2486.2009.02115.xURL [本文引用: 3]
Forests in northeastern North America are influenced by varying climatic and biotic factors; however, there is concern that rapid changes in these factors may lead to important changes in ecosystem processes such as decomposition. Climate change (especially warming) is predicted to increase rates of decomposition in northern latitudes. Warming in winter may result in complex effects including decreased levels of snow cover and an increased incidence of soil freezing that will effect decomposition. Along with these changes in climate, moose densities have also been increasing in this region, likely affecting nutrient dynamics. We measured decomposition and N release from 15N-labeled sugar maple leaf litter and moose feces over 20 months in reference and snow removal treatment (to induce soil freezing) plots in two separate experiments at the Hubbard Brook Experimental Forest in New Hampshire, USA. Snow removal/soil freezing decreased decomposition of maple litter, but stimulated N transfer to soil and microbial biomass. Feces decomposed more rapidly than maple litter, and feces N moved into the mineral soil more than N derived from litter, likely due to the lower C : N ratio of feces. Feces decomposition was not affected by the snow removal treatment. Total microbial biomass (measured as microbial N and C) was not significantly affected by the treatments in either the litter or feces plots. These results suggest that increases in soil freezing and/or large herbivore populations, increase the transfer rate of N from plant detritus or digested plants into the mineral soil. Such changes suggest that altering the spatial and temporal patterns of soil freezing and moose density have important implications for ecosystem N cycling.
DOI:10.1007/s00442-012-2393-xURL [本文引用: 1]
DOI:10.1016/j.soilbio.2004.02.021URL [本文引用: 1]
DOI:10.1016/j.quaint.2006.10.027URL [本文引用: 1]
At its maximum annual development, snow can cover more than half the Northern Hemisphere land area with one-third experiencing seasonal snow cover. The precise conditions that develop during the annual pattern of snowpack development formation have implications for: (i) soil microbiological activity and nutrient transformations; (ii) the capacity of the accumulating snowpack to retain atmospheric derived solutes; (iii) preferential elution and rapid runoff of solutes from the snowpack during periods of thaw; and (iv) leaching of solutes. Long-term records of annual snow accumulation suggest that substantial, regional scale shifts in snowpack characteristics have been occurring. The accompanying changes in the frequency and timing of freeze鈥搕haw episodes and the evidence of their disruptive and selective influence upon soil microbial processes, when human induced, suggests there are wider implications for nutrient cycling and functioning of mountain ecosystems. This review is focused on alpine landscapes.
[本文引用: 1]
DOI:10.1023/A:1013024603959URL [本文引用: 1]
Overwinter and snowmelt processes are thought to be critical to controllers of nitrogen (N) cycling and retention in northern forests. However, there have been few measurements of basic N cycle processes (e.g. mineralization, nitrification, denitrification) during winter and little analysis of the influence of winter climate on growing season N dynamics. In this study, we manipulated snow cover to assess the effects of soil freezing on in situ rates of N mineralization, nitrification and soil respiration, denitrification (intact core, C2H2 - based method), microbial biomass C and N content and potential net N mineralization and nitrification in two sugar maple and two yellow birch stands with reference and snow manipulation treatment plots over a two year period at the Hubbard Brook Experimental Forest, New Hampshire, U.S.A. The snow manipulation treatment, which simulated the late development of snowpack as may occur in a warmer climate, induced mild (temperatures > -5 °C) soil freezing that lasted until snowmelt. The treatment caused significant increases in soil nitrate (NO3-) concentrations in sugar maple stands, but did not affect mineralization, nitrification, denitrification or microbial biomass, and had no significant effects in yellow birch stands. Annual N mineralization and nitrification rates varied significantly from year to year. Net mineralization increased from 65 12.0 g N m-2 y-1 in 1998 to 65 22 g N m-2 y-1 in 1999 and nitrification increased from 65 8 g N m-2 y-1 in 1998 to 65 13 g N m-2 y-1 in 1999. Denitrification rates ranged from 0 to 0.65 g N m-2 y-1. Our results suggest that mild soil freezing must increase soil NO3- levels by physical disruption of the soil ecosystem and not by direct stimulation of mineralization and nitrification. Physical disruption can increase fine root mortality, reduce plant N uptake and reduce competition for inorganic N, allowing soil NO3- levels to increase even with no increase in net mineralization or nitrification.
DOI:10.3321/j.issn:1001-7488.2006.04.017URLMagsci [本文引用: 2]
<p>系统评述森林凋落物的分解过程、凋落物分解及养分释放的影响因素、分解研究的方法等。森林凋落物的分解既有物理过程,又有生物化学过程,一般由淋溶、自然粉碎、代谢作用等共同完成。凋落物分解过程先后出现分解速率较快和较慢2个阶段,元素迁移一般呈现淋溶-富集-释放的模式。凋落物分解主要受气候、凋落物性质、微生物和土壤动物的影响,气候是最基本的影响因素,常用实际蒸散(actual evapotranspiration 简称AET)作为指标。凋落物分解速率呈明显的气候地带性,与温度、湿度等紧密相关。从全球尺度来讲,凋落物质量对分解速率的影响处于次要地位,但在同一气候带内因AET变化较小,则起了主导作用。N、P和木质素浓度、C/N、C/P、木质素与养分比值是常见的凋落物质量指标,其中C/N和木质素/N最能反映凋落物分解速率。凋落物化学性质对其分解的影响作用又与分解阶段有关。凋落叶中N、P、K初始浓度高使得初期分解较快,而后期分解放慢。土壤理化性质及微生物区系也将不同程度地影响凋落物分解。尼龙网袋法(litter bag method)操作简单,是野外测定森林凋落物分解速率最常用的方法。除此之外,缩微试验也得到了广泛应用。目前普遍采用的衡量凋落物分解速率大小的指标主要有CO<sub>2</sub>释放速率、凋落物分解系数(<em>k</em>值)及质量损失率。在此基础上提出了指数衰减、线性回归等模型来模拟凋落物分解过程。尽管对凋落物分解在森林生态系统C、N、P循环、土壤肥力维持等方面已进行了较深入的研究,但未来研究应侧重以下方向:长期的定位观测;采用相对统一的研究方法,获得可比性强的数据进行综合;深化凋落物分解机理研究;探讨全球气候变化对森林凋落物分解的影响;评价营林措施(如林分皆伐、造林、施石灰和肥料等)对凋落物分解与养分释放的调节作用。</p>
DOI:10.3321/j.issn:1001-7488.2006.04.017URLMagsci [本文引用: 2]
<p>系统评述森林凋落物的分解过程、凋落物分解及养分释放的影响因素、分解研究的方法等。森林凋落物的分解既有物理过程,又有生物化学过程,一般由淋溶、自然粉碎、代谢作用等共同完成。凋落物分解过程先后出现分解速率较快和较慢2个阶段,元素迁移一般呈现淋溶-富集-释放的模式。凋落物分解主要受气候、凋落物性质、微生物和土壤动物的影响,气候是最基本的影响因素,常用实际蒸散(actual evapotranspiration 简称AET)作为指标。凋落物分解速率呈明显的气候地带性,与温度、湿度等紧密相关。从全球尺度来讲,凋落物质量对分解速率的影响处于次要地位,但在同一气候带内因AET变化较小,则起了主导作用。N、P和木质素浓度、C/N、C/P、木质素与养分比值是常见的凋落物质量指标,其中C/N和木质素/N最能反映凋落物分解速率。凋落物化学性质对其分解的影响作用又与分解阶段有关。凋落叶中N、P、K初始浓度高使得初期分解较快,而后期分解放慢。土壤理化性质及微生物区系也将不同程度地影响凋落物分解。尼龙网袋法(litter bag method)操作简单,是野外测定森林凋落物分解速率最常用的方法。除此之外,缩微试验也得到了广泛应用。目前普遍采用的衡量凋落物分解速率大小的指标主要有CO<sub>2</sub>释放速率、凋落物分解系数(<em>k</em>值)及质量损失率。在此基础上提出了指数衰减、线性回归等模型来模拟凋落物分解过程。尽管对凋落物分解在森林生态系统C、N、P循环、土壤肥力维持等方面已进行了较深入的研究,但未来研究应侧重以下方向:长期的定位观测;采用相对统一的研究方法,获得可比性强的数据进行综合;深化凋落物分解机理研究;探讨全球气候变化对森林凋落物分解的影响;评价营林措施(如林分皆伐、造林、施石灰和肥料等)对凋落物分解与养分释放的调节作用。</p>
DOI:10.5846/stxb201311292844URL
以针叶林代表性地被植物锦丝藓和高山冷蕨为研究对象,采用凋落物分解网袋法,研究了高山林线交错带(暗针叶林-林线-高山草甸)的锦丝藓植物残体、高山冷蕨凋落叶及混合凋落物经过一个雪被期分解后的质量损失与土壤动物群落结构特征.结果表明:雪被期林线交错带上两种地被物的质量损失率在高山草甸最大,锦丝藓表现更为显著,两种凋落物混合促进了分解过程且在林线上表现尤为显著.在交错带凋落物中共获取土壤动物968头,隶属于5纲10目35科,优势类群以弹尾目和蜱螨目为主.在林线上凋落物中获得的土壤动物个体数和类群数高于高山草甸和暗针叶林.典范对应分析(CCA)表明,土壤动物类群与雪被期平均温度关系最为密切,特有物种如等翅目和地蜈蚣目仅在暗针叶林出现,半翅目和啮目仅在高山草甸出现.地被物种类对土壤动物多样性的影响在暗针叶林和高山草甸大于林线.多元回归分析表明,日平均温度和雪被厚度能够解释凋落物质量损失率变异的30.8%,而土壤动物因子能解释质量损失率变异的8.3%,它们共同解释质量损失率变异的34.1%.雪被是影响高山两种地被物凋落物分解的最关键因子.
DOI:10.5846/stxb201311292844URL
以针叶林代表性地被植物锦丝藓和高山冷蕨为研究对象,采用凋落物分解网袋法,研究了高山林线交错带(暗针叶林-林线-高山草甸)的锦丝藓植物残体、高山冷蕨凋落叶及混合凋落物经过一个雪被期分解后的质量损失与土壤动物群落结构特征.结果表明:雪被期林线交错带上两种地被物的质量损失率在高山草甸最大,锦丝藓表现更为显著,两种凋落物混合促进了分解过程且在林线上表现尤为显著.在交错带凋落物中共获取土壤动物968头,隶属于5纲10目35科,优势类群以弹尾目和蜱螨目为主.在林线上凋落物中获得的土壤动物个体数和类群数高于高山草甸和暗针叶林.典范对应分析(CCA)表明,土壤动物类群与雪被期平均温度关系最为密切,特有物种如等翅目和地蜈蚣目仅在暗针叶林出现,半翅目和啮目仅在高山草甸出现.地被物种类对土壤动物多样性的影响在暗针叶林和高山草甸大于林线.多元回归分析表明,日平均温度和雪被厚度能够解释凋落物质量损失率变异的30.8%,而土壤动物因子能解释质量损失率变异的8.3%,它们共同解释质量损失率变异的34.1%.雪被是影响高山两种地被物凋落物分解的最关键因子.
DOI:10.3724/SP.J.1258.2013.00030URL [本文引用: 1]
高山/亚高山森林灌木层植物凋落物的分解对于系统物质循环等过程具有重要意义,并可能受到冬季不同厚度雪被斑块下冻融格局的影响。该文采用凋落物分解袋法,研究了高山森林典型灌层植物华西箭竹(Fargesia nitida)$1]康定柳(Satix paraplesia)凋落物在沿林窗一林下形成的冬季雪被厚度梯度(厚型雪被斑块、较厚型雪被斑块、中型雪被斑块、薄型雪被斑块、无雪被斑块)上在第一年不同关键时期(冻结初期、冻结期、融化期、生长季节初期和生长季节后期)的质量损失特征。在整个冻融季节,华西箭竹和康定柳凋落叶的平均质量损失分别占全年的(48.78±2.351%和(46.60±5.02)%。冻融季节雪被覆盖斑块下凋落叶的失重率表现出厚型雪被斑块大于薄型雪被斑块的趋势,而生长季节无雪被斑块的失重率明显较高。尽管如此,华西箭竹凋落物第一年分解表现出随冬季雪被厚度增加而增加的趋势,但康定柳凋落物第一年失重率以薄型雪被斑块最高,而无雪被斑块最低。同时,相关分析表明冻融季节凋落叶的失重率与平均温度和负积温呈极显著正相关,生长季节凋落叶的失重率与所调查的温度因子并无显著相关关系,但全年凋落物失重率与平均温度和正/负积温均显著相关。这些结果清晰地表明,未来冬季变暖情境下高山森林冬季雪被格局的改变将显著影响灌层植物凋落物分解,影响趋势随着物种的差异具有明显差异。
DOI:10.3724/SP.J.1258.2013.00030URL [本文引用: 1]
高山/亚高山森林灌木层植物凋落物的分解对于系统物质循环等过程具有重要意义,并可能受到冬季不同厚度雪被斑块下冻融格局的影响。该文采用凋落物分解袋法,研究了高山森林典型灌层植物华西箭竹(Fargesia nitida)$1]康定柳(Satix paraplesia)凋落物在沿林窗一林下形成的冬季雪被厚度梯度(厚型雪被斑块、较厚型雪被斑块、中型雪被斑块、薄型雪被斑块、无雪被斑块)上在第一年不同关键时期(冻结初期、冻结期、融化期、生长季节初期和生长季节后期)的质量损失特征。在整个冻融季节,华西箭竹和康定柳凋落叶的平均质量损失分别占全年的(48.78±2.351%和(46.60±5.02)%。冻融季节雪被覆盖斑块下凋落叶的失重率表现出厚型雪被斑块大于薄型雪被斑块的趋势,而生长季节无雪被斑块的失重率明显较高。尽管如此,华西箭竹凋落物第一年分解表现出随冬季雪被厚度增加而增加的趋势,但康定柳凋落物第一年失重率以薄型雪被斑块最高,而无雪被斑块最低。同时,相关分析表明冻融季节凋落叶的失重率与平均温度和负积温呈极显著正相关,生长季节凋落叶的失重率与所调查的温度因子并无显著相关关系,但全年凋落物失重率与平均温度和正/负积温均显著相关。这些结果清晰地表明,未来冬季变暖情境下高山森林冬季雪被格局的改变将显著影响灌层植物凋落物分解,影响趋势随着物种的差异具有明显差异。
DOI:10.1016/0169-5347(92)90126-VURLPMID:21236058
Plant species create positive feedbacks to patterns of nutrient cycling in natural ecosystems. For example, in nutrient-poor ecosystems, plants grow slowly, use nutrients efficiently and produce poor-quality litter that decomposes slowly and deters herbivores. /n contrast, plant species from nutrient-rich ecosystems grow rapidly, produce readily degradable litter and sustain high rates of herbivory, further enhancing rates of nutrient cycling. Plants may also create positive feedbacks to nutrient cycling because of species' differences in carbon deposition and competition with microbes for nutrients in the rhizosphere. New research is showing that species' effects can be as or more important than abiotic factors, such as climate, in controlling ecosystem fertility.
DOI:10.2307/2963492URL [本文引用: 1]
I compared effects of increased temperature and litter from different Alaskan tundra plant species on cycling of carbon and nitrogen through Litter and soil in microcosms. Warming between 4 degrees and 10 degrees C significantly increased rates of soil and litter respiration, litter decomposition, litter nitrogen release, and soil net nitrogen mineralization. Thus, future warming will directly increase rates of carbon and nitrogen cycling through litter and soil in tundra. In addition, differences among species' litter in rates of decomposition, N release, and effects on soil net nitrogen mineralization were sometimes larger than differences between the two temperature treatments within a species. Thus, changes in plant community structure and composition associated with future warming will have important consequences for how elements cycle through litter and soil in tundra. In general, species within a growth form (graminoids, evergreen shrubs, deciduous shrubs, and mosses) were more similar in their effects on decomposition than were species belonging to different growth forms, with graminoid litter having the fastest rate and litter of deciduous shrubs and mosses having the slowest rates. Differences in rates of litter decomposition were more related to carbon quality than to nitrogen concentration. Increased abundance of deciduous shrubs with future climate warming will promote carbon storage, because of their relatively large allocation to woody stems that decompose slowly. Changes in moss abundance will also have important consequences for future carbon and nitrogen cycling, since moss litter is extremely recalcitrant and has a low potential to immobilize nitrogen.
DOI:10.1139/x95-171URL [本文引用: 1]
URL [本文引用: 2]
2010年1-5月在川西高原采用人工雪厚度梯度试验(0、30和100 cm),应用网袋分解法对窄叶鲜卑花叶片凋落物进行分解试验,测定了凋落物的分解速率及其养分动态.结果表明:在无雪被覆盖的样地上分解5个月后的凋落物 质量损失率为29.9%,而中雪和深雪样地的凋落物质量损失率分别为33.8%和35.2%.分解过程中,凋落物氮存在一定的富集现象,磷处于波动的富集 状态,碳质量分数和碳氮比均呈现前期急剧下降后期逐渐上升的趋势.雪被覆盖显著增加了凋落物的质量损失率和氮含量,而对碳和磷含量无显著影响.在川西高原 地区,30 cm以上的持续雪被覆盖能够改变凋落物的分解过程,从而可能对土壤营养物质转化和植物群落构建产生实质性的影响.
URL [本文引用: 2]
2010年1-5月在川西高原采用人工雪厚度梯度试验(0、30和100 cm),应用网袋分解法对窄叶鲜卑花叶片凋落物进行分解试验,测定了凋落物的分解速率及其养分动态.结果表明:在无雪被覆盖的样地上分解5个月后的凋落物 质量损失率为29.9%,而中雪和深雪样地的凋落物质量损失率分别为33.8%和35.2%.分解过程中,凋落物氮存在一定的富集现象,磷处于波动的富集 状态,碳质量分数和碳氮比均呈现前期急剧下降后期逐渐上升的趋势.雪被覆盖显著增加了凋落物的质量损失率和氮含量,而对碳和磷含量无显著影响.在川西高原 地区,30 cm以上的持续雪被覆盖能够改变凋落物的分解过程,从而可能对土壤营养物质转化和植物群落构建产生实质性的影响.
[本文引用: 1]
DOI:10.1111/j.1461-0248.2007.01124.xURLPMID:18021246 [本文引用: 1]
To explore the importance of 12 elements in litter production and decomposition, we fertilized 36 1600 m(2)-plots with combinations of N, P, K, or micronutrients (i.e. B, Ca, Cu, Fe, Mg, Mn, Mo, S, Zn) for 6 years in a lowland Panamanian forest. The 90% of litter falling as leaves and twigs failed to increase with fertilization, but reproductive litter (fruits and flowers) increased by 43% with N. K enhanced cellulose decomposition; one or more micronutrients enhanced leaf-litter decomposition; P enhanced both. Our results suggest tropical forests are a non-Liebig world of multiple nutrient limitations, with at least four elements shaping rates of litterfall and decomposition. Multiple metallomic enzymes and cofactors likely create gradients in the break down of leaf litter. Selection favours individuals that make more propagules, and even in an N-rich forest, N is a non-substitutable resource for reproduction.
DOI:10.4141/CJSS2012-025URL [本文引用: 1]
Decomposition is a key process in carbon and nutrient cycling. However, little is known about its response to altered winter soil temperature regimes in boreal forests. Here, the impact of soil frost on cellulose decomposition over 1 yr and soil biotic activity (bait-lamina sticks) over winter, in spring, and in summer was investigated using a long-term (9-yr) snow-cover manipulation experiment in a boreal Picea abies forest. The experiment consisted of the treatments: snow removal, increased insulation, and ambient control. The snow removal treatment caused longer and deeper soil frost (minimum temperature - 8.6 degrees C versus - 1.4 degrees C) at 10 cm soil depth in comparison with control, while the increased insulation treatment resulted in nearly no soil frost during winter. Annual cellulose decomposition rates were reduced by 46% in the snow removal manipulation in comparison with control conditions. Increased insulation had no significant effect on decomposition. The decomposition was mainly driven by microorganisms, as no significant difference was observed for containers enclosed with a 44-mu m and a 1-mm mesh. Soil biotic activity was slightly increased by both the snow removal and the increased insulation treatment in comparison with control conditions over winter. However, this effect disappeared over spring and summer. We conclude that soil frost can have strong effects on decomposition in boreal ecosystems. Further studies should investigate to which degree the observed reduction in decomposition due to reduced snow cover in winter slows or even offsets the expected increase in decomposition rates with global warming.
DOI:10.1016/j.soilbio.2007.03.032URL [本文引用: 1]
Substrate quality and decomposition (measured as CO 2 release in laboratory microcosms) of fresh leaf litter and fine roots of Cupressus lusitanica, Pinus patula, Eucalyptus grandis and native forest trees were studied. Changes in litter chemistry in each forest stand were analysed by comparing fresh leaf litter (collected from trees) and decomposed litter from the forest floor. Elemental concentrations, proximate fractions including monomeric sugars, and cross polarisation magic-angle spinning (CPMAS) 13C NMR spectra were analysed in leaf litters, decomposed litter and fine roots. Leaf litters and fine roots varied in their initial substrate chemistry with Ca concentration in leaf litters being higher than that in fine roots. In each stand, fine roots had a higher acid unhydrolysable residue (AUR) (except for the Pinus stand), higher holocellulose concentration and lower concentration of water-soluble extractives (WSE) and dichloromethane extractives (NPE) than fresh leaf litter. Likewise, 13C NMR spectra of fine roots showed lower alkyl and carboxyl C, and higher phenolic (except P. patula), aromatic and O-alkyl C proportions than leaf litters. Compared with fresh leaf litter, decomposed litter had lower concentrations of potassium, holocellulose, WSE, NPE, arabinose and galactose, similar or higher concentrations of Mg, Ca, S and P, and higher concentrations of N and AUR. CPMAS 13C NMR spectra of decomposed litter showed a higher relative increase in signal intensity due to methoxyl C, aromatic C, phenolic C and carboxylic C compared with alkyl C. In a microcosm decomposition study, the proportion of initial C remaining in leaf litter and fine roots significantly fitted an exponential regression model. The decomposition constants ( k) ranged between 0.0013 and 0.0030 d 611 for leaf litters and 0.0010–0.0017 d 611 for fine roots. In leaf litters there was a positive correlation between the k value and the initial Ca concentration, and in fine roots there was an analogous positive correlation with initial WSE. Leaf litters decomposed in the order Cupressus>native forest> Eucalyptus65 Pinus, and fine roots in the order Pinus>native forest> Cupressus65 Eucalyptus. In each stand the fine root decomposition was significantly lower than the leaf litter decomposition, except for the P. patula stand where the order was reversed.
DOI:10.3390/f7020027URL [本文引用: 1]
To understand how forest gaps and the associated canopy control litter lignin and cellulose dynamics by redistributing the winter snow coverage and hydrothermal conditions in the growing season, a field litterbag trial was conducted in the alpine Minjiang fir (Abies faxoniana Rehder and E.H. Wilson) forest in a transitional area located in the upper reaches of the Yangtze River and the eastern Tibetan Plateau. Over the first year of litter decomposition, the litter exhibited absolute cellulose loss and absolute lignin accumulation except for the red birch litter. The changes in litter cellulose and lignin were significantly affected by the interactions among gap position, period and species. Litter cellulose exhibited a greater loss in the winter with the highest daily loss rate observed during the snow cover period. Both cellulose and lignin exhibited greater changes under the deep snow cover at the gap center in the winter, but the opposite pattern occurred under the closed canopy in the growing season. The results suggest that decreased snowpack seasonality due to winter warming may limit litter cellulose and lignin degradation in alpine forest ecosystems, which could further inhibit litter decomposition. As a result, the ongoing winter warming and gap vanishing would slow soil carbon sequestration from foliar litter in cold biomes.
DOI:10.5846/stxb201304140704URL [本文引用: 1]
采用凋落物分解袋法,研究了在土壤、水分相当的条件下模拟增温对红松(Pinus koraiensis)、蒙古栎(Quercus mongolica)及其混合凋落物分解的影响,以及在不同温度水平下,不同凋落物质量(两种单一凋落物和混合凋落物)的分解特性。利用碱式吸收法测量了凋落物分解累积释放CO2动态。将N浓度和C/N率作为凋落物质量参数,用呼吸产生CO2的积累值和凋落物质量损失率确定凋落物分解率。结果表明温度升高对单一凋落物和混合凋落物分解均有促进作用,在不同温度水平上,不同质量凋落物的分解特性有所差别,25℃和29℃条件下混合凋落物分解速率蒙古栎单一凋落物红松单一凋落物分解速率。然而,在31℃条件下混合凋落物与蒙古栎单一凋落物分解速率相差不大,二者均大于红松单一凋落物分解速率。
DOI:10.5846/stxb201304140704URL [本文引用: 1]
采用凋落物分解袋法,研究了在土壤、水分相当的条件下模拟增温对红松(Pinus koraiensis)、蒙古栎(Quercus mongolica)及其混合凋落物分解的影响,以及在不同温度水平下,不同凋落物质量(两种单一凋落物和混合凋落物)的分解特性。利用碱式吸收法测量了凋落物分解累积释放CO2动态。将N浓度和C/N率作为凋落物质量参数,用呼吸产生CO2的积累值和凋落物质量损失率确定凋落物分解率。结果表明温度升高对单一凋落物和混合凋落物分解均有促进作用,在不同温度水平上,不同质量凋落物的分解特性有所差别,25℃和29℃条件下混合凋落物分解速率蒙古栎单一凋落物红松单一凋落物分解速率。然而,在31℃条件下混合凋落物与蒙古栎单一凋落物分解速率相差不大,二者均大于红松单一凋落物分解速率。
DOI:10.1038/nature10576URLPMID:22056985 [本文引用: 1]
Abstract Permafrost contains an estimated 1672090009Pg carbon (C), an amount roughly equivalent to the total currently contained within land plants and the atmosphere. This reservoir of C is vulnerable to decomposition as rising global temperatures cause the permafrost to thaw. During thaw, trapped organic matter may become more accessible for microbial degradation and result in greenhouse gas emissions. Despite recent advances in the use of molecular tools to study permafrost microbial communities, their response to thaw remains unclear. Here we use deep metagenomic sequencing to determine the impact of thaw on microbial phylogenetic and functional genes, and relate these data to measurements of methane emissions. Metagenomics, the direct sequencing of DNA from the environment, allows the examination of whole biochemical pathways and associated processes, as opposed to individual pieces of the metabolic puzzle. Our metagenome analyses reveal that during transition from a frozen to a thawed state there are rapid shifts in many microbial, phylogenetic and functional gene abundances and pathways. After one week of incubation at 509000900°C, permafrost metagenomes converge to be more similar to each other than while they are frozen. We find that multiple genes involved in cycling of C and nitrogen shift rapidly during thaw. We also construct the first draft genome from a complex soil metagenome, which corresponds to a novel methanogen. Methane previously accumulated in permafrost is released during thaw and subsequently consumed by methanotrophic bacteria. Together these data point towards the importance of rapid cycling of methane and nitrogen in thawing permafrost.
DOI:10.1007/s00442-003-1253-0URLPMID:12684853 [本文引用: 1]
Anthropogenic forces are concurrently reducing biodiversity and altering terrestrial nutrient cycles. As natural populations decline, genetic diversity within single species also declines. The consequences of intraspecific genetic loss for ecosystem functions are poorly understood, and interactions among intraspecific diversity, nitrogen deposition, and nutrient cycling are unknown. We present results from an experiment that simulated both a decline in biodiversity and an increase in nitrogen deposition. In soil microcosms, we tested effects of variation in intraspecific litter diversity and nitrogen deposition on soil respiration and nitrogen leaching. Increases in intraspecific litter diversity increased soil respiration overall, with the greatest increases in respiration occurring under high nitrogen deposition. Nitrogen deposition increased the amount of inorganic nitrogen leached, while the amount of dissolved organic nitrogen leached was correlated with initial litter chemistry (lignin concentration) and remained independent of litter diversity and nitrogen deposition treatments. Our results demonstrate the potential for losses in genetic diversity to interact with other global environmental changes to influence terrestrial nutrient cycles.
DOI:10.2307/1932179URL
See full-text article at JSTOR
DOI:10.1657/1523-0430(2003)035[0074:FSOTAA]2.0.CO;2URL [本文引用: 3]
We divide the Alaskan Arctic cold season into five stages based on transitions in climatological and thermophysical conditions in the atmosphere, snowpack, and soil active layer. Each of these stages has distinct characteristics which drive ecosystem processes. During the two autumnal stages (and ) soils remain warm, unfrozen water is present, and the highest rates of cold-season soil respiration occur. The next two stages (and ) are characterized by a frozen active layer with decreasing temperature. is critical in determining the length of the growing season and the resumption of biological processes. and result from a radiation deficit, show little interannual variation, and will be resistant to change under almost any reasonable climate change scenario. These are also the stages with the least amount of biological activity and have the least impact on the ecosystem. However, and stages vary significantly from year to year, have more ecosystem implications, and are also the most likely to undergo significant change in timing and character as the arctic climate changes. This 5-fold subdivision is useful for framing discussions of biophysical interactions during the arctic winter and for focusing attention on critical cold-season periods.
DOI:10.1007/s11104-012-1307-3URL [本文引用: 2]
AbstractAimsClimate-induced changes in snow cover are likely to affect cold arctic and alpine ecosystems functioning and major processes such as wintertime plant litter decomposition. However, it remains poorly studied in subalpine systems where the snowpack may be irregular. In this paper we explored the dynamic of the winter plant litter decomposition process, its magnitude and its relationship with the snowpack properties.MethodsIn subalpine grasslands of the Central French Alps, we performed a litter bag experiment monitoring over a whole winter the litter decomposition from the exploitative ResultsLitter decomposition rates were stable during winter and 3-fold higher under deeper and permanent snowpack with higher thermal resistance. Litter quality appeared only significant under thinner snowpack with higher decomposition rates for the exploitative species. A snowpack with higher thermal resistance created an insulating layer promoting the decomposition process.ConclusionThese results suggest that the temporal (permanence vs. intermittency) and physical (depth and thermal resistance) characteristics of the snowpack should be considered when studying the response of winter ecosystems functioning to global changes.
DOI:10.1007/s10533-013-9882-9URL [本文引用: 1]
Winter climate change is an important environmental driver that alters the biogeochemical processes of forest soils. The decrease in snowpack amplifies soil freeze–thaw cycles and decreases the snowmelt water supply to soil. This study examined how snow decrease affects nitrogen (N) mineralization and nitrification in forest soil in northern Japan by conducting an in situ experimental snowpack manipulation experiment and a laboratory incubation of soil with different moisture, temperature and freeze–thaw magnitudes. For the incubation studies, surface mineral soil (0–1002cm) was collected from a cool-temperate natural mixed forest and incubated using the resin core method during the winter. In the field, there were two treatments: 50 and 10002% snow removal and control plots. The increase in the soil freeze–thaw cycle increased net N mineralization and marginally decreased the net nitrification in soil. The dissolved organic carbon (DOC) and DOC/DON ratio in soil increased with the decrease in snowpack especially during the snow melt period. These results suggested that the change in substrate quality by the increase in freeze–thaw cycles caused the significant enhancement of microbial ammonium production in soil. The lower soil moisture and higher gross immobilization of inorganic N by soil microbes may be maintaining the slow net nitrification and low nitrate leaching in freeze–thaw cycles with less snowpack. The results indicate that winter climate change would strongly impact N biogeochemistry through the increase in ammonium availability in soil for plants and microbes, whereas it would be unlikely that nitrate loss from surface soil would be enhanced.
DOI:10.1007/s00374-011-0636-3URL [本文引用: 1]
Abstract65=650.0001), Pseudoscorpionida (75%; 6565=650.0033), Collembola (24%; 65=650.0019), adult Coleoptera (23%; 65=650.0057), and larval Diptera (33%; 6565=650.032). Taxa that did not respond significantly to snow removal included Chilopoda (65=650.55), Acari (65=650.66), Diplopoda (65=650.66), adult Diptera (65=650.54), and larval Coleoptera (65=650.39). A delayed snowpack over two winters also resulted in decreased arthropod richness by 30% (6565=650.0002) during the two subsequent growing seasons. Results of this study demonstrate that predicted changes in the winter snowpack and depth and duration of soil frost may reduce the abundance and alter the community composition of arthropods living in the forest floor of northern hardwood forests, which could have implications for the structure and function of northern forest ecosystems.
DOI:10.1007/BF00032630URL [本文引用: 1]
The snow-bed vegetation in the Northern Apennines was studied by the Braun-Blanquet method. The phytosociological relev茅s were classified by numerical methods. Four main vegetation types were identified: Salicetum herbaceae, community of Carex foetida, Poo-Cerastietum cerastioidis and Oligotricho-Gnaphalietum supini. The latter was described as a new association. The vegetation types were ecologically characterized by an indirect gradient analysis based on principal component analysis. The results were interpreted by means of: i) geomorphological observations, ii) soil profiles, iii) measurements of the length of the snow-free period.
DOI:10.1016/j.agrformet.2005.11.003URL [本文引用: 1]
Winter CO 2 emissions from soil and snow-covered surfaces can represent a significant fraction of the annual carbon cycle in cool-temperate broad-leaved deciduous forests. The sources of this CO 2 production are, however, poorly understood. To clarify the role of saprophytic microorganisms in the carbon cycle during the winter (December–April), mass loss rates of organic substrates (leaf litter and cellulose sheets), and microbial activity and fungal biomass in the litter layer of a cool-temperate broad-leaved deciduous forest in Japan were investigated. The mass loss rate of leaves reached 13%, which accounted for 26% of the annual mass loss. Microbial respiration in the leaf litter, measured in the laboratory, was detected even at 612 °C. The ergosterol content (an indicator of fungal biomass) of the cellulose sheets increased significantly in the snow-covered season. Seven genera of fungi including Varicosporium elodeae, Epicoccum purpurascens and Cylindrocarpon destructans were isolated from leaf litter collected from under snow cover. Microbial respiration during the winter was estimated based on the relationships between the amount of leaf litter available, the temperature dependency of microbial respiration and the temperature in the litter layer. The microorganisms in the litter layer emitted 31 gCO 2-C m 612, which accounts for 38% of the total CO 2 efflux during the winter. These results suggest that through soil respiration, microorganisms in the litter layer play an important role in the carbon cycle during the winter.
DOI:10.1016/j.jaridenv.2007.10.010URL
The literature on aboveground litter decomposition in arid environments, which is dominated by short-term studies (one- to three-year duration), shows no consistent relationship between mass loss rates and factors controlling decomposition in mesic environments. For ten years, we repeated a decomposition study of five species in four arid to semi-arid biomes in central New Mexico. Annual precipitation during the study ranged from 60% to 150% of the long-term mean. Mass remaining after one year of decomposition varied widely between species: O. hymenoidesB. eriopodaJ. monospermaB. gracilis. Except for percent lignin, litter quality measures were not good predictors of relative mass loss rates among species. Although a few significant relationships between decomposition and precipitation and litter quality were found, our study results were overwhelmingly unsupportive of these factors having major impacts on litter decomposition processes in central New Mexico.
DOI:10.3354/cr017063URL
This study was aimed at assessing the potential impacts of climate change on the depth and duration of soil frost under snow cover in forests growing at different geographical locations in Finland. Frost simulations using a process-based forest ecosystem model (FinnFor) were made for Scots pine Pinus sylvestris L. stands (height 17 m, stand density 1100 stems ha(-1)) growing on a moraine sandy soil. The climate change forecast used in the computations was based on the global ocean-atmosphere general circulation model HadCM2 that was dynamically downscaled to the regional level. The simulated climate warming during the winter months was about 4 to 5 degreesC by the end of the 21st century. Frost simulations showed that the length of the soil frost period would lessen all over the country. Though winters will be warmer, the associated decrease in snow cover in southern Finland will increase the probability of frozen ground there in the middle of winter compar
DOI:10.2307/1937654URL [本文引用: 1]
Decomposition of grass leaf litter was studied on a shortgrass prairie using chemicals (HgCl"2 and CuSO"4) to prevent microbial activity (abiotic treatment), 53-@mm nylon mesh to exclude mesofauna (microbial treatment), and l-mm nylon mesh to allow the access of mesofauna. After 9 months, 15.2% of the blue grama grass litter was decomposed in the microbial treatment, and 29.4% was decomposed in the microbial plus mesofaunal treatment. After 7 months, 6.2% of the litter had disappeared from the abiotic treatment. There was a general decrease in C:N ratios with the microbial treatment lowest at the end of the experiment. Total available carbohydrates generally decreased with time. Certain mite families fluctuated with seasons. The tydeids were most active in winter and tetranychids were most active in summer. A correlation between abiotic factors and mite families was also observed.
DOI:10.1016/j.agrformet.2013.04.008URL [本文引用: 2]
Changes in characteristics of snowfall and spring freeze–thaw-cycle (FTC) events under the warming climate make it critical to understand biophysical controls on soil CO2 efflux (RS) in seasonally snow-covered ecosystems. We conducted a snow removal experiment and took year-round continuous automated measurements of RS, soil temperature (T5) and soil volumetric water content at the 5cm depth (W5) with a half-hour interval in a Chinese temperate forest in 2010–2011. Our objectives were to: (1) develop statistical models to describe the seasonality of RS in this forest; (2) quantify the contribution of seasonal RS to the annual budget; (3) examine biophysical effects of snowpack on RS; and (4) test the hypothesis that an FTC-induced enhancement of RS is jointly driven by biological and physical processes. Empirical RS–T5–W5 models explained 65.3–94.1% of the variability in the RS data, but the number of the regression terms and their coefficients varied with season. This indicates that the model should be fitted to the seasonal data sets separately to explicitly describe the seasonality of RS. The RS during the winter, spring FTC period, and growing season contributed 5.7%, 3.5%, and 91.1%, respectively, to the total annual RS. The relative enhancement of RS due to snowpack and FTCs averaged 3.4 and 2.5, respectively. The snowpack-induced enhancement of RS exponentially increased with T5 (R2=0.83) and snow depth (R2=0.16), while the FTC-induced enhancement of RS exponentially decreased with T5 (R2=0.45) and W5 (R2=0.67). These results suggest that the snowpack-induced enhancement mainly results from the snow-depth-dependent insulation of soil from low air temperatures, while the FTC-induced enhancement is dominantly driven by biological processes. Accumulatively, the snowpack and spring FTCs made a minor net contribution (2.3% and 1.2%, respectively) to the annual RS budget.
DOI:10.1016/j.actao.2009.11.002URL [本文引用: 1]
Mass loss and nutrient release of forest litter during the freeze–thaw season could play an essential role in C and nutrient cycling in cold regions, but few studies in some key ecosystems have been available. In order to characterize litter decomposition during the freeze–thaw season in a subalpine forest region of western China, a field experiment using the litterbag method was conducted on the decomposition of foliar litter of two dominant species, fir ( Abies faxoniana) and birch ( Betula platyphylla) under their respective forests. Over the freeze–thaw season following leaf-fall, about 18% and 20% of mass, 13% and 14% of lignin, 30% and 26% of cellulose, 14% and 21% of C, 30% and 27% of N, 17% and 15% of P, and 17% and 13% of K were lost from fir and birch litters, respectively. The lost mass and components accounted for more than 64% and 65% of mass, 72% and 69% of lignin, 75% and 60% of cellulose, 49% and 59% of C, 56% and 71% of N, 62% and 37% of P, and 38% and 37% of K in 1 year net loss rate of fir and birch litter, respectively. In addition, the loss of mass, lignin, cellulose and component bio-elements during the freeze–thaw season correlated closely with the initial substrate type and the levels of the individual bio-elements. The results demonstrated that litter decomposition during the freeze–thaw season contributes significantly to the first year decomposition in these subalpine forests.
DOI:10.5846/stxb201410101991URL [本文引用: 1]
在东北长白山、张广才岭、小兴 安岭、大兴安岭的主要森林类型中设置26块样地,进行为期3a(2004—2006年)凋落叶分解实验,以研究气候、林型、林冠透光率对凋落叶分解速率的 相对影响大小。结果表明,不同林型凋落叶分解速率依次为:落叶阔叶林针阔叶混交林落叶针叶林常绿针叶林岳桦林。对分解速率影响因素的分析表明,气候因子 (热量和水分)对分解速率有较强的解释力,分别解释了分解常数k和分解95%所需时间(t95%)的55.5%和65.0%的变异。但是,气候对分解速率 的影响在很大程度上是通过与林型、林冠透光率的协同作用而实现的,其独立解释力并不大(9%)。气候的变化导致林型(物种组成)的变化、进而影响分解速 率,这一因素解释了分解参数变异的46.8%(k)和56.8%(t95%)。与此同时,气候和林型的变化还导致林冠透光率的变化,随着热量水平的上升林 冠透光率下降、间接提高分解速率。这一因素分别解释了k值和t95%变异的23.9%和22.3%。研究结果表明,气候对凋落叶分解的影响主要是通过对物 种组成、林冠结构(影响透光率)等生物因素的间接作用实现的。忽视这些生物因素、简单研究气候和分解速率的关系可能难以正确预测未来气候变化对凋落物分解 的影响。
DOI:10.5846/stxb201410101991URL [本文引用: 1]
在东北长白山、张广才岭、小兴 安岭、大兴安岭的主要森林类型中设置26块样地,进行为期3a(2004—2006年)凋落叶分解实验,以研究气候、林型、林冠透光率对凋落叶分解速率的 相对影响大小。结果表明,不同林型凋落叶分解速率依次为:落叶阔叶林针阔叶混交林落叶针叶林常绿针叶林岳桦林。对分解速率影响因素的分析表明,气候因子 (热量和水分)对分解速率有较强的解释力,分别解释了分解常数k和分解95%所需时间(t95%)的55.5%和65.0%的变异。但是,气候对分解速率 的影响在很大程度上是通过与林型、林冠透光率的协同作用而实现的,其独立解释力并不大(9%)。气候的变化导致林型(物种组成)的变化、进而影响分解速 率,这一因素解释了分解参数变异的46.8%(k)和56.8%(t95%)。与此同时,气候和林型的变化还导致林冠透光率的变化,随着热量水平的上升林 冠透光率下降、间接提高分解速率。这一因素分别解释了k值和t95%变异的23.9%和22.3%。研究结果表明,气候对凋落叶分解的影响主要是通过对物 种组成、林冠结构(影响透光率)等生物因素的间接作用实现的。忽视这些生物因素、简单研究气候和分解速率的关系可能难以正确预测未来气候变化对凋落物分解 的影响。
[本文引用: 1]
[本文引用: 1]
DOI:10.3321/j.issn:1000-0933.2005.11.012URL [本文引用: 1]
土壤碳是高纬度地区森林生态系统最大的碳库,是森林生态系统碳循环的极其重要组分.研究了东北东部典型的6种次生林生态系统(天然蒙古栎林、杨桦林、杂木林、硬阔叶林、红松人工林和落叶松人工林)的土壤碳动态,包括(1)量化土壤有机碳(SOC)含量、碳密度及周转时间,(2)比较不同森林生态系统的土壤表面CO2通量(Rs)年通量差异,(3)建立Rs年通量及其分量与SOC的量化关系.研究结果表明:阔叶天然次生林和针叶人工林的SOC含量变化范围分别为52.63~66.29 g·kg-1和42.15~49.15g·kg-1;平均SOC密度分别为15.57和17.16 kg·m-2;平均SOC周转时间分别为32a和48a.各个生态系统的Rs依次为杂木林951 gC·m-2·a-1、硬阔叶林892 gC·m-2·a-1、杨桦林812 gC·m-2·a-1、蒙古栎林678gC·m-2·a-1、红松林596 gC·m-2·a-1和落叶松林451 gC·m-2·a-1.Rs年通量及其分量(土壤异养呼吸和自养呼吸)与SOC含量呈显著的正相关,但其相关程度因土层不同而异(R2=0.747~0.933).同一生态系统中,SOC含量随土深增加而降低,而SOC密度和SOC周转时间随深度增加而增大.采用统一规范的研究方法,获取大量有代表性的森林生态系统土壤碳贮量和Rs的实测数据,是减少区域尺度碳平衡研究中不确定性的不可缺少的研究内容.
DOI:10.3321/j.issn:1000-0933.2005.11.012URL [本文引用: 1]
土壤碳是高纬度地区森林生态系统最大的碳库,是森林生态系统碳循环的极其重要组分.研究了东北东部典型的6种次生林生态系统(天然蒙古栎林、杨桦林、杂木林、硬阔叶林、红松人工林和落叶松人工林)的土壤碳动态,包括(1)量化土壤有机碳(SOC)含量、碳密度及周转时间,(2)比较不同森林生态系统的土壤表面CO2通量(Rs)年通量差异,(3)建立Rs年通量及其分量与SOC的量化关系.研究结果表明:阔叶天然次生林和针叶人工林的SOC含量变化范围分别为52.63~66.29 g·kg-1和42.15~49.15g·kg-1;平均SOC密度分别为15.57和17.16 kg·m-2;平均SOC周转时间分别为32a和48a.各个生态系统的Rs依次为杂木林951 gC·m-2·a-1、硬阔叶林892 gC·m-2·a-1、杨桦林812 gC·m-2·a-1、蒙古栎林678gC·m-2·a-1、红松林596 gC·m-2·a-1和落叶松林451 gC·m-2·a-1.Rs年通量及其分量(土壤异养呼吸和自养呼吸)与SOC含量呈显著的正相关,但其相关程度因土层不同而异(R2=0.747~0.933).同一生态系统中,SOC含量随土深增加而降低,而SOC密度和SOC周转时间随深度增加而增大.采用统一规范的研究方法,获取大量有代表性的森林生态系统土壤碳贮量和Rs的实测数据,是减少区域尺度碳平衡研究中不确定性的不可缺少的研究内容.
DOI:10.1007/s11427-010-4026-0URLPMID:20697872
Quantifying forest carbon (C) storage and distribution is important for forest C cycling studies and terrestrial ecosystem modeling. Forest inventory and allometric approaches were used to measure C density and allocation in six representative temperate forests of similar stand age (42–59 years old) and growing under the same climate in northeastern China. The forests were an aspen-birch forest, a hardwood forest, a Korean pine plantation, a Dahurian larch plantation, a mixed deciduous forest, and a Mongolian oak forest. There were no significant differences in the C densities of ecosystem components (except for detritus) although the six forests had varying vegetation compositions and site conditions. However, the differences were significant when the C pools were normalized against stand basal area. The total ecosystem C density varied from 186.9 tC hm 612 to 349.2 tC hm 612 across the forests. The C densities of vegetation, detritus, and soil ranged from 86.3–122.7 tC hm 612 , 6.5–10.5 tC hm 612 , and 93.7–220.1 tC hm 612 , respectively, which accounted for 39.7% ± 7.1% (mean ± SD), 3.3% ± 1.1%, and 57.0% ± 7.9% of the total C densities, respectively. The overstory C pool accounted for > 99% of the total vegetation C pool. The foliage biomass, small root (diameter < 5mm) biomass, root-shoot ratio, and small root to foliage biomass ratio varied from 2.08–4.72 tC hm 612 , 0.95–3.24 tC hm 612 , 22.0%–28.3%, and 34.5%–122.2%, respectively. The Korean pine plantation had the lowest foliage production efficiency (total biomass/foliage biomass: 22.6 g g 611 ) among the six forests, while the Dahurian larch plantation had the highest small root production efficiency (total biomass/small root biomass: 124.7 g g 611 ). The small root C density decreased with soil depth for all forests except for the Mongolian oak forest, in which the small roots tended to be vertically distributed downwards. The C density of coarse woody debris was significantly less in the two plantations than in the four naturally regenerated forests. The variability of C allocation patterns in a specific forest is jointly influenced by vegetation type, management history, and local water and nutrient availability. The study provides important data for developing and validating C cycling models for temperate forests.
DOI:10.3773/j.issn.1005-264x.2008.05.008URL [本文引用: 2]
在中国东北长白山、帽儿山、凉水、根河的主要森林类型中设置27个样地,连续3年(2004~2006年)观测森林凋落物的生产量,以研究我国东北地区森林凋落物产量及其与环境因子的关系。结果表明,不同森林类型凋落物年产量存在显著差异,针阔叶混交林显著高于落叶针叶林和常绿针叶林,落叶针叶林、常绿针叶林、落叶阔叶林和针阔叶混交林的年平均产量分别为2337、2472、3130和4146kg·hm^-2;树叶、枝条、繁殖器官和其它组分占总凋落量的平均比例为71%、22%、6%和1%,不同森林类型凋落物组分的比例差异较大。森林凋落物产量主要受温度限制,降水、森林类型和群落结构无显著影响。不同组分凋落物量的影响因素不同;树叶凋落量主要受温度和森林类型的影响;枝条凋落量主要受降水和蓄积量的影响;而繁殖器官凋落量则与树种的繁殖特性以及年降水有关。各组分占总凋落量的比例主要受降水影响,树叶占凋落物比例随降水增加而下降,枝条所占比例很小,表现出与叶相反的变化趋势。
DOI:10.3773/j.issn.1005-264x.2008.05.008URL [本文引用: 2]
在中国东北长白山、帽儿山、凉水、根河的主要森林类型中设置27个样地,连续3年(2004~2006年)观测森林凋落物的生产量,以研究我国东北地区森林凋落物产量及其与环境因子的关系。结果表明,不同森林类型凋落物年产量存在显著差异,针阔叶混交林显著高于落叶针叶林和常绿针叶林,落叶针叶林、常绿针叶林、落叶阔叶林和针阔叶混交林的年平均产量分别为2337、2472、3130和4146kg·hm^-2;树叶、枝条、繁殖器官和其它组分占总凋落量的平均比例为71%、22%、6%和1%,不同森林类型凋落物组分的比例差异较大。森林凋落物产量主要受温度限制,降水、森林类型和群落结构无显著影响。不同组分凋落物量的影响因素不同;树叶凋落量主要受温度和森林类型的影响;枝条凋落量主要受降水和蓄积量的影响;而繁殖器官凋落量则与树种的繁殖特性以及年降水有关。各组分占总凋落量的比例主要受降水影响,树叶占凋落物比例随降水增加而下降,枝条所占比例很小,表现出与叶相反的变化趋势。
DOI:10.13332/j.1000-1522.20150438URL [本文引用: 2]
凋落物-土壤界面连续体是森林生态系统的最重要部分,也是氮素生物地球化学循环最活跃的场所。土壤氮元素的生物地球化学循环广义上可分为转运和转化2个环节,真菌和细菌分别在这2个环节上扮演重要角色。降雨、氮沉降和温度等变化能够改变森林生态系统的氮生物地球化学循环过程,在全球变化加剧背景下,深入了解凋落物-土壤界面连续体内氮的转运和转化过程和机制尤为重要。本文综述了凋落物-土壤界面连续体的研究现状,通过应用^(15)N示踪、分子生物学测序和15N-DNA-SIP分子探针技术,研究氮转运和转化的微生物群落及其过程的可行性,并提出今后森林生态系统凋落物-土壤界面连续体的氮循环模式,强调了真菌的转运和细菌的转化过程在氮固持中的贡献,有助于森林生态系统氮固持力和机制系统认知,为开展温带森林生态系统管理和氮排放控制提供了思路。
DOI:10.13332/j.1000-1522.20150438URL [本文引用: 2]
凋落物-土壤界面连续体是森林生态系统的最重要部分,也是氮素生物地球化学循环最活跃的场所。土壤氮元素的生物地球化学循环广义上可分为转运和转化2个环节,真菌和细菌分别在这2个环节上扮演重要角色。降雨、氮沉降和温度等变化能够改变森林生态系统的氮生物地球化学循环过程,在全球变化加剧背景下,深入了解凋落物-土壤界面连续体内氮的转运和转化过程和机制尤为重要。本文综述了凋落物-土壤界面连续体的研究现状,通过应用^(15)N示踪、分子生物学测序和15N-DNA-SIP分子探针技术,研究氮转运和转化的微生物群落及其过程的可行性,并提出今后森林生态系统凋落物-土壤界面连续体的氮循环模式,强调了真菌的转运和细菌的转化过程在氮固持中的贡献,有助于森林生态系统氮固持力和机制系统认知,为开展温带森林生态系统管理和氮排放控制提供了思路。
DOI:10.1080/02827581.2012.670726URL
Freeze–thaw events in winter may affect litter decomposition in cold biomes but few reports are available. We characterized the fir (Abies faxoniana) litter decomposition over a whole winter (November 2008 to April 2009) during the late autumn, deep winter, and early spring stages. The mass loss, nutrient release, and quality change of fir litter were determined using the litterbag method at 2700, 3000, 3300, and 3600 m altitude in southwest China. Over the winter an average of 18% mass, 27% C, 50% N, 40% P, 36% K, 30% cellulose, and 14% lignin were lost. Of these total losses, a majority loss of mass (70%), C (65%), N (50%), P (58%), K (42%), cellulose (70%), and lignin (68%) occurred during the deep winter stage. The highest loss rate of mass (19.2%) and lignin (16.4%) but the lowest N loss (47.9%) was at the highest 3600 m altitude. Soil freeze–thaw cycle resulted in significant losses of mass, while mass loss rate did not increase under the higher mean soil temperature during each stage. Our results confirmed that the physical process seemed to be the most important process for cold season decomposition in the cold biome.
Sensitivity of soil respiration and microbial communities to altered snowfall
1
2013
... id="C5">虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异.例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素.但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(
The freezer defrosting: Global warming and litter decomposition rates in cold biomes
3
2006
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
... id="C5">虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异.例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素.但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
Seasonal climate manipulations have only minor effects on litter decomposition rates and N dynamics but strong effects on litter P dynamics of sub-arctic bog species
1
2012
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
Experimentally increased snow accumulation alters soil moisture and animal community structure in a polar desert
1
2010
... id="C47">本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低.增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间.这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程.这可能有以下几方面解释: (1)控雪改变分解者动态.雪被良好的隔热能力可以提升雪下温度(
Direct and indirect control by snow cover over decomposition in alpine tundra along a snowmelt gradient
1
2010
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
Snow pack in the Swiss Alps under changing climatic conditions: An empirical approach for climate impacts studies
2003
Plant Litter-Decomposition, Humus Formation, Carbon Sequestration. 3rd edn. Springer
3
2014
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
... id="C47">本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低.增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间.这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程.这可能有以下几方面解释: (1)控雪改变分解者动态.雪被良好的隔热能力可以提升雪下温度(
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
Decomposition of beech (Fagus sylvatica) and pine (Pinus nigra) litter along an alpine elevation gradient: Decay and nutrient release.
1
2015
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
Initial stages of tundra shrub litter decomposition may be accelerated by deeper winter snow but slowed down by spring warming
2
2016
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
Reduction in snow depth negatively affects decomposers but impact on decomposition rates is substrate dependent
3
2013
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
... id="C47">本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低.增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间.这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程.这可能有以下几方面解释: (1)控雪改变分解者动态.雪被良好的隔热能力可以提升雪下温度(
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
Understanding the dominant controls on litter decomposition
1
2016
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
Snowpack controls on nitrogen cycling and export in seasonally snow-covered catchments
1
1999
... id="C47">本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低.增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间.这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程.这可能有以下几方面解释: (1)控雪改变分解者动态.雪被良好的隔热能力可以提升雪下温度(
Warming effects and plant trait control on the early-decomposition in alpine snowbeds
1
2014
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
Winter climate change implications for decomposition in northeastern forests, comparisons of sugar maple litter with herbivore fecal inputs
3
2010
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
... )或减雪(如
... id="C5">虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异.例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素.但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(
Influence of experimental snow removal on root and canopy physiology of sugar maple trees in a northern hardwood forest
1
2013
... id="C47">本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低.增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间.这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程.这可能有以下几方面解释: (1)控雪改变分解者动态.雪被良好的隔热能力可以提升雪下温度(
Annual dynamics of phosphatase activities in an evergreen oak litter: Influence of biotic and abiotic factors
1
2004
... id="C47">本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低.增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间.这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程.这可能有以下几方面解释: (1)控雪改变分解者动态.雪被良好的隔热能力可以提升雪下温度(
Changes in the seasonal snow cover of alpine regions and its effect on soil processes: A review
1
2007
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
Enzyme dynamics on decomposing leaf litter of Cistus incanus and Myrtus communis in a Mediterranean ecosystem.
1
2000
... id="C47">本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低.增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间.这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程.这可能有以下几方面解释: (1)控雪改变分解者动态.雪被良好的隔热能力可以提升雪下温度(
Effects of mild winter freezing on soil nitrogen and carbon dynamics in a northern hardwood forest
1
2001
... id="C47">本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低.增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间.这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程.这可能有以下几方面解释: (1)控雪改变分解者动态.雪被良好的隔热能力可以提升雪下温度(
森林凋落物分解研究进展
2
2006
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
... ), 可能造成P流失.因此与N相比, 微生物更难从外界获取P以满足分解过程的营养需求(
森林凋落物分解研究进展
2
2006
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
... ), 可能造成P流失.因此与N相比, 微生物更难从外界获取P以满足分解过程的营养需求(
雪被期川西高山林线交错带两种地被物凋落物分解与土壤动物多样性
2016
雪被期川西高山林线交错带两种地被物凋落物分解与土壤动物多样性
2016
雪被斑块对高山森林两种灌木凋落叶质量损失的影响
1
2013
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
雪被斑块对高山森林两种灌木凋落叶质量损失的影响
1
2013
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
Effects of plant species on nutrient cycling
1992
Temperature and plant species control over litter decomposition in Alaskan tundra
1
1996
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
Influence of microclimate on decomposition in loblolly pine stands: A field microcosm approach
1
1995
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
川西高原季节性雪被覆盖对窄叶鲜卑花凋落物分解和养分动态的影响
2
2012
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
川西高原季节性雪被覆盖对窄叶鲜卑花凋落物分解和养分动态的影响
2
2012
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
1
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
Multiple nutrients limit litterfall and decomposition in a tropical forest
1
2008
... id="C5">虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异.例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素.但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(
Snow removal reduces annual cellulose decomposition in a riparian boreal forest
1
2013
... id="C5">虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异.例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素.但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(
Decomposition and substrate quality of leaf litters and fine roots from three exotic plantations and a native forest in the southwestern highlands of Ethiopia
1
2007
... id="C5">虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异.例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素.但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(
Effects of forest gaps on litter lignin and cellulose dynamics vary seasonally in an alpine forest
1
2016
... id="C6">东北地区是中国气候变化最显著、植被系统对气候变化响应最为敏感的地区(
模拟增温和不同凋落物基质质量对凋落物分解速率的影响
1
2013
... id="C5">虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异.例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素.但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(
模拟增温和不同凋落物基质质量对凋落物分解速率的影响
1
2013
... id="C5">虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异.例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素.但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(
Metagenomic analysis of a permafrost microbial community reveals a rapid response to thaw
1
2011
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
Intraspecific litter diversity and nitrogen deposition affect nutrient dynamics and soil respiration
1
2003
... id="C23">凋落叶分解过程采用指数模型(
Energy storage and the balance of producers and decomposers in ecological systems
1963
Five stages of the Alaskan Arctic cold season with ecosystem implications
3
2003
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
... id="C15">为了解控雪对凋落叶分解的影响, 基于研究区历年气象观测数据,
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
The effects of snowpack properties and plant strategies on litter decomposition during winter in subalpine meadows
2
2013
... id="C5">虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异.例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素.但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(
... id="C47">本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低.增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间.这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程.这可能有以下几方面解释: (1)控雪改变分解者动态.雪被良好的隔热能力可以提升雪下温度(
Impact of snowpack decrease on net nitrogen mineralization and nitrification in forest soil of northern Japan
1
2013
... id="C47">本研究中两个性状各异的树种凋落叶分解率均在增雪处理中最高, 除雪处理中最低.增雪处理有效地缩短了凋落叶半分解和95%分解时间, 而除雪处理延长了相应的时间.这说明增雪处理促进了凋落叶整个分解过程, 而除雪处理抑制了该过程.这可能有以下几方面解释: (1)控雪改变分解者动态.雪被良好的隔热能力可以提升雪下温度(
Impact of a reduced winter snowpack on litter arthropod abundance and diversity in a northern hardwood forest ecosystem
1
2012
... id="C49">经典生态学中一直认为, 冬季是凋落物分解的“停滞”时期, 但越来越多的研究表明: 虽然不同树种凋落物分解机制不同, 但冬季与生长季节一样, 也是凋落物分解的重要时期(
The snow-bed vegetation in the Northern Apennines
1
1991
... id="C4">森林凋落物分解是生态系统物质循环的主要供给源, 是森林生态系统生产力、碳储量和土壤有机质形成的基础(
Microbial activity and litter decomposition under snow cover in a cool-temperate broad-leaved deciduous forest
1
2005
... id="C18">每次取样时, 随机从各样方中采集红松和蒙古栎分解袋各3袋, 小心去掉新生根系及杂物后装入无菌聚乙烯袋中带回实验室, 70 ℃烘干至恒质量后测定凋落叶残留干质量.采集分解袋的同时, 采集袋下对应的土壤有机层样品(5 cm × 10 cm), 小心去除异物后风干, 供有机层有机碳、全氮和全磷含量的测定使用(
Aboveground decomposition in arid environments: Results of a long-term study in central New Mexico
2008
Impact of climate change on soil frost under snow cover in a forested landscape
2001
Abiotic and biotic factors in litter decomposition in semiarid grassland
1
1979
... id="C7">研究地位于黑龙江帽儿山森林生态系统国家野外科学观测研究站(45.33°-45.42° N, 127.50°- 127.57° E).平均海拔400 m, 平均坡度10°-15°, 土壤主要为暗棕色森林土.该地区属大陆性季风气候, 春季多风干燥, 夏季湿润多雨, 冬季干燥寒冷.年降水量629 mm, 年潜在蒸发量854 mm, 年平均气温3.1 ℃, 11月至次年4月地表有明显的季节性雪被覆盖(
Seasonality of soil CO2 efflux in a temperate forest: Biophysical effects of snowpack and spring freeze-thaw cycles
2
2013
... id="C5">虽然环境因子、凋落物质量和分解者是调控凋落物分解的三大重要因素, 但各因素的作用时间和程度不同, 致使不同条件下影响凋落物分解的因子产生分异.例如, Vossbrinc等(1979)研究发现, 作为分解者的土壤动物, 对凋落物分解的贡献最大; 而和润莲等(2016)报道, 环境因子和分解者是影响两种地被植物凋落物分解的主要因素.但可以肯定的是, 雪被可通过改变以上因素影响凋落物分解过程, 并且这种影响在持续时间上还具有一定的阶段性: 在冻结初期和深冻期, 雪被控制了地表和土壤微环境, 可使雪下的温湿度保持适宜(
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
Litter decomposition in two subalpine forests during the freeze-thaw season
1
2010
... id="C6">东北地区是中国气候变化最显著、植被系统对气候变化响应最为敏感的地区(
东北地区森林凋落叶分解速率与气候, 林型, 林分光照的关系
1
2016
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
东北地区森林凋落叶分解速率与气候, 林型, 林分光照的关系
1
2016
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
冬季雪被对青藏高原东缘高海拔森林凋落叶P元素释放的影响
1
2015
... id="C6">东北地区是中国气候变化最显著、植被系统对气候变化响应最为敏感的地区(
冬季雪被对青藏高原东缘高海拔森林凋落叶P元素释放的影响
1
2015
... id="C6">东北地区是中国气候变化最显著、植被系统对气候变化响应最为敏感的地区(
东北东部森林生态系统土壤碳贮量和碳通量
1
2005
... id="C7">研究地位于黑龙江帽儿山森林生态系统国家野外科学观测研究站(45.33°-45.42° N, 127.50°- 127.57° E).平均海拔400 m, 平均坡度10°-15°, 土壤主要为暗棕色森林土.该地区属大陆性季风气候, 春季多风干燥, 夏季湿润多雨, 冬季干燥寒冷.年降水量629 mm, 年潜在蒸发量854 mm, 年平均气温3.1 ℃, 11月至次年4月地表有明显的季节性雪被覆盖(
东北东部森林生态系统土壤碳贮量和碳通量
1
2005
... id="C7">研究地位于黑龙江帽儿山森林生态系统国家野外科学观测研究站(45.33°-45.42° N, 127.50°- 127.57° E).平均海拔400 m, 平均坡度10°-15°, 土壤主要为暗棕色森林土.该地区属大陆性季风气候, 春季多风干燥, 夏季湿润多雨, 冬季干燥寒冷.年降水量629 mm, 年潜在蒸发量854 mm, 年平均气温3.1 ℃, 11月至次年4月地表有明显的季节性雪被覆盖(
Carbon density and distribution of six Chinese temperate forests
2010
我国东北主要森林类型的凋落物产量及其影响因素
2
2008
... id="C6">东北地区是中国气候变化最显著、植被系统对气候变化响应最为敏感的地区(
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
我国东北主要森林类型的凋落物产量及其影响因素
2
2008
... id="C6">东北地区是中国气候变化最显著、植被系统对气候变化响应最为敏感的地区(
... id="C48">本研究中, 有机碳含量、C/N、木质素含量等凋落叶初始质量指标是影响分解率的重要因子.与对照相比, 增雪处理中分解率仅受初始有机碳含量影响; 表明增雪处理中环境、分解者等适宜凋落叶分解, 因此两个树种凋落叶初始有机碳含量的差异是分解率变异的主要原因(
氮在凋落物-土壤界面连续体转移研究进展
2
2016
... id="C18">每次取样时, 随机从各样方中采集红松和蒙古栎分解袋各3袋, 小心去掉新生根系及杂物后装入无菌聚乙烯袋中带回实验室, 70 ℃烘干至恒质量后测定凋落叶残留干质量.采集分解袋的同时, 采集袋下对应的土壤有机层样品(5 cm × 10 cm), 小心去除异物后风干, 供有机层有机碳、全氮和全磷含量的测定使用(
... id="C31">式中: M0表示分解袋埋置前袋中凋落叶质量(g); (Mt+1 - Mt)表示相邻两阶段分解袋中凋落叶质量差(g).凋落叶各季节分解量对分解总量的贡献率(P) (
氮在凋落物-土壤界面连续体转移研究进展
2
2016
... id="C18">每次取样时, 随机从各样方中采集红松和蒙古栎分解袋各3袋, 小心去掉新生根系及杂物后装入无菌聚乙烯袋中带回实验室, 70 ℃烘干至恒质量后测定凋落叶残留干质量.采集分解袋的同时, 采集袋下对应的土壤有机层样品(5 cm × 10 cm), 小心去除异物后风干, 供有机层有机碳、全氮和全磷含量的测定使用(
... id="C31">式中: M0表示分解袋埋置前袋中凋落叶质量(g); (Mt+1 - Mt)表示相邻两阶段分解袋中凋落叶质量差(g).凋落叶各季节分解量对分解总量的贡献率(P) (
Decomposition of Abies faxoniana litter varies with freeze-thaw stages and altitudes in subalpine/alpine forests of southwest China
2012
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}